
Abstract
Many activities from livestock husbandry contribute to emission and concentration of carbon dioxide (CO2), nitrous oxide (N2O) and methane (CH4) gases to the atmosphere; activities such as grazing, manure and urine deposited or stored on land as well as crop farming practices such as tilling, burning of biomass or crop residues. A better understanding of the extent of emission sources and carbon sequestration potential for Eastern Africa rangelands is vital for developing mitigation strategies. In this article, we review the sources of emission with a focus on land conversion for crop farming, livestock husbandry, wildfire/burning and biotic processes such as soil biota activity in the ecosystem. The trade-offs of using rangeland with an emphasis on enhancing carbon sequestration potential are also addressed. This review revealed that many practices that enhance carbon capture process show promising benefits with sink capacity of −0.004 to 13 Mg C ha−1 year−1. However, given multiple land-use and environmental dynamics in Eastern African rangelands, it is imperative to generate more data across various land management and climatic zones in order to ascertain varied sink capacity. Improving carbon sequestration in rangelands through appropriate land management is a promising cost-effective strategy to mitigate climate change. Through improved farming or grazing management practice and restoring of degraded areas, there are significant benefits to enhance carbon sequestration. As rangeland resources are multi-faceted, engaging trans-disciplinary approaches is necessary to allow analyses of co-benefits of improved management or trade-offs degrading.
Similar content being viewed by others


Introduction
Atmospheric concentrations of greenhouse gases
An increase in the concentration of atmospheric greenhouse gases (GHGs) methane (CH4), carbon dioxide (CO2) and nitrous oxide (N2O) has led to widespread concern about human activities that contribute heavily to global environmental change (IPCC 2013; Aneja et al. 2019). Between 1959 and 2018, the global mean annual concentration of CO2 rose from 315.97 parts per million (ppm) to 408.52 ppm (NOAA/ESRL 2018), while CH4 rose from 719.01 parts per billion (ppb) in 1750 to 1857.62 ppb in 2018 (EEA and NOAA 2019), and N2O increased from 270.00 ppb in 1750 to 328.51 ppb in 2016 (EEA 2019). Studies show that human activities such as deforestation, crop farming, livestock husbandry, land-use change and industrial developments have contributed to the observed increases in GHG concentrations (IPCC 2007, 2013; MacCarthy and Zougmoré 2018), which, over the last century, have resulted in increasing global mean surface air temperature and climate change. These changes have a range of potential impacts including extreme weather events, sea level rise and changes in biogeochemical cycles and net primary productivity (Ritchie and Roser 2019). While climate change is projected to aggravate more (McDermot and Elavarthi 2014), implementing practices that can help to improve sinks and intensify their sequestration from the atmosphere are necessary in order to mitigate climate change.
Terrestrial carbon sinks
From a land management perspective, greenhouse gas mitigation should focus on reducing emissions from livestock husbandry and land-use change, such as farming expansion or deforestation (Niles et al. 2002; Ringius 2002; Scharlemann et al. 2014). Additionally, mitigation may take advantage of the fact that, as plants and trees grow, they take CO2 from the atmosphere through the photosynthesis process and store it in woody trees and soils. Therefore, enhancing the capacity of natural sinks such as forests (Pan et al. 2011; FAO 2020) and rangelands, is crucial (McDermot and Elavarthi 2014).
Forests form the most significant global carbon sink, because trees store carbon in woody tissues that delay decomposition and reduce the release of CO2 through respiration (Keenan and Williams 2018). The proportion and spatial distribution of forests include tropical 45%, boreal 27%, temperate 16% and sub-tropical 11% (FAO and UNEP 2020). Given its coverage of 4.06 billion hectares (ha), about 31% of the total land area (FAO 2020), if well managed, forests have huge potential to sequester carbon in woody biomass and soils (Pugh et al. 2019). The current estimates based on FRA 2020 data show that forests acted globally as a net carbon sink (−0.6 Gt CO2 year−1) in the period 2011–2015 (FAO 2020). Forests contribute a large fraction of the global terrestrial CO2 sink as they experience enhanced growth from improved environmental and resource conditions (Keenan and Williams 2018).
Rangelands are also an important carbon sink (McDermot and Elavarthi 2014; Derner et al. 2017). Globally, 41.3% of the world’s land surface is classified as rangelands (MEA 2005; Nosetto et al. 2006; Lal 2011; UN 2011; Laban et al. 2018). The rangeland vegetation consists of grasses, grass-like plants, forbs, shrubs or trees that are grazed or have the potential to be grazed or browsed and which are used as a natural ecosystem for raising grazing livestock and conserving wildlife (Allen et al. 2011; Liniger and Studer 2019). The Millennium Ecosystem Assessment (MEA 2005) includes rangelands with drylands, because the occurrence of vegetation subtypes is influenced by the quantity of precipitation. Based on aridity, rangelands may be classified into hyper-arid, arid, semi-arid, dry sub-humid and humid, with rainfall ranging from less than 200 millimetres (mm) in hyper-arid to above 1500 mm in humid areas. These ecosystems support the livelihoods of millions of people worldwide and are important providers of other services (Godde et al. 2020). Because of their global extent, if properly managed, rangelands have the potential to sequester carbon in woody plants and store organic and inorganic carbon in soils (WRI 2000; IPCC 2007; Derner et al. 2017). Some studies show that rangelands can store up to 10–30% of global soil organic carbon (SOC) (Lal 2004; Derner and Schuman 2007; Laban et al. 2018) and sequester up to 179.623 Mg of CO2 from the atmosphere per year (Lal 2011; McDermot and Elavarthi 2014).
Africa’s rangelands
Rangelands occupy 43–45% of Africa’s land surface, with woody and shrub trees comprising the dominant vegetation types (23.8%) and grassland with bush trees making up 19.5% (WRI 2003; UNEP 2009). The remaining African land area is made up of dense forest (7.9%), mosaic forest (4.3%), cultivated land (10.1%), wetland (0.9%) and bare soil (33.5%) (Hoffman and Vogel 2008). Given their extent and heterogeneity, rangelands provide suitable habitat for wild and domesticated herbivores (as well as many other animals), ecosystem services for communities and support many different socio-economic activities (Mortimore et al. 2009). It is estimated that 40–55% of the continent’s population (268 to 384 million people) live in rangelands and are engaged in pastoralism, rainfed cultivation (in humid areas) and extraction of non-timber forest products (UNEP 2009; IIED and SOS 2010; Derner et al. 2017; Liniger and Studer 2019).
Land-use dynamics
African rangelands have multiple uses, with pastoralists, crop farmers, commercial investors and conservation competing for land and related resources (Kideghesho et al. 2013; Ykhanbai et al. 2014). This competition continues to occur under the influences of changing climate, land-use and national policy, such as the conversion of communal lands into private ranches (Neely et al. 2009; Reid et al. 2014). These changing trends affect the traditional land-use systems, vegetation cover and carbon balances of the ecosystem. Climate variability reduces the resilience of the ecosystem through deviations in precipitation, rising temperature and changes of net primary productivity. In Eastern Africa rangelands, increased intensity of livestock grazing around watering points, and growing of new permanent to semi-permanent farming settlements are also replacing nomadic pastoral systems (FAO 2019). In the contemporary discourse on land use, greenhouse gas emissions, climate change and strategies for poverty reduction, it has been suggested that development carbon finance schemes are a win-win strategy for African rangelands (Perez et al. 2007; Wilkes and Tennigkeit 2010; Dean et al. 2015). The schemes are perceived as ways to exploit the synergy between climate change mitigation and the enhancement of pastoral livelihoods for the benefit of the larger ecosystem, with co-benefits for livestock production and carbon sequestration. However, the current United Nations (UN’s) Programme on Reducing Emissions from Deforestation and Forest Degradation (REDD+) carbon-based payment for ecosystem services’ programmes (UN 2018) has put little emphasis on arid and semi-arid rangelands (Wilkes and Tennigkeit 2010). This is attributed to inadequate monitoring of the changes on carbon stocks in the rangelands (cf. Lipper et al. 2010), as well as methodological complexities (Ciais et al. 2011; Stringer et al. 2012). A limited understanding of carbon market opportunities by the stakeholders, unclear land rights in most communal rangelands and uncertainty of the capacity of rangelands to compete for limited carbon markets with forest biomes are other uncertainties (Tennigkeit and Wilkies 2008; UNEP 2008; Wilkes and Tennigkeit 2010). Few studies have analysed the effects of these dynamics across land management practices (Houghton et al. 2012; MacCarthy and Zougmoré 2018). A recent study by Tessema et al. (2019) assessed the potential of grasslands to sequester soil organic carbon in six East African countries (Burundi, Ethiopia, Kenya, Rwanda, Tanzania and Uganda). However, these estimates are not sufficient to inform country-specific policy recommendations and intervention practices by local resource users; hence, detailed quantification of the processes exacerbating GHG emissions and carbon dynamics is required (Stringer et al. 2012; Tessema et al. 2019). Given the modifications in rangelands, quantifying carbon stock in established allotments can help to predict the impacts of change in land management on carbon sequestration (Lal 2004; Lipper et al. 2010; Stringer et al. 2012). The objectives of this article are to review the major sources of GHG emissions related to land conversion for crop farming, livestock husbandry and soil biota processes and to identify and synthesize the knowledge gaps and uncertainties regarding the potential of rangeland ecosystems to sequester carbon. The specific focus is on the Eastern Africa experience, but also some examples are drawn from a broader perspective. The review provides specific recommendations for enhancing carbon sink capacity.
Methods
Data sources and compilation
The peer-reviewed articles, international reports, books, proceeding documents and web data were searched using the keywords “greenhouse gases”, “rangelands”, “land-use”, “crop farming/husbandry”, “livestock husbandry”, “wildfires/burning”, “soil biota”, “termites” and “carbon sequestration”. Seven hundred publications were identified from the search (Google Scholar, InWorldInData.org, Scopus.com, ResearchGate.net, Springer.com and ScienceDirect.com), of which 4 publications on atmospheric GHG data obtained from InWorldInData.org, 28 international reports (FAO/org, WRI/org, IPPC, IIED, MEA, UN, UNEP, IUCN, ICRAF/org, TerrAfrica, FIAN International, HPG, and ILCA), 1 proceeding report, 9 books and 171 peer-reviewed articles published from 1970 to 2020 were included during the preparation of this article. After screening the documents using the specific keywords, 213 relevant publications with information on land use and management, biotic activities, GHG emission sources and carbon sequestration potential and rangelands were reviewed, analysed and synthesized. The approach is semi-systematic and integrative where we begin from broad to specific (Snyder 2019). Abiotic processes are important as they influence biotic activity and GHG emissions and therefore were considered in this review. Similarly, ammonia (NH3), which is predominantly from manure storage and land application, is not a GHG but has significant environmental implications and therefore was considered in the review. Available data on carbon sequestration capacity of the forest biome is referred for comparative purposes. This review is a continuation of the CarbonQuest project: Livelihood diversifying potential of livestock based carbon sequestration options in pastoral and agro-pastoral systems in Africa funded by the Federal Ministry of Economic Cooperation and Development (BMZ) (Germany) 2011 to 2014. The review, therefore, focuses on the biophysical potential of rangelands to sequester carbon given the different land-use options such as livestock and crop husbandry. It aims to address the knowledge gap on how much carbon could potentially be stored and what emissions could be avoided in African rangelands, when effectively controlling factors influencing the carbon balance under conditions of pastoral land use, such as grazing, wildfire/burning, extraction of woody plants and land conversion to crop farming, are understood.
Results
Sources of greenhouse gas emissions in rangeland ecosystems
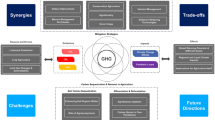
The major sources of GHG emissions related to land-use management are livestock or crop husbandry and plant biomass extraction (Fig. 1). Biotic activities which contribute to emissions are largely influenced by the quantity of precipitation and temperature or land use. Figure 1 illustrates the sources, activities and processes contributing to carbon dynamics and emission of other non-carbon greenhouse gases in rangelands via different pathways. The emission of CH4 is generated in anaerobic manure storage and eructation from ruminants, and N2O occurs from livestock manure. In high temperatures, volatilization of animal urine as urea-nitrogen occurs primarily in the form of NH3 and NOx. Carbon dioxide (CO2) is released during grazing and farming practice while carbon monoxide (CO) during burning of plant biomass. An increase in the atmospheric concentration of GHGs requires scientific strategies and policy responses for offsetting emissions through enhancing carbon sink. This is possible if there is enough data to support the strategies at various spatial and temporal scales.

The causes and sources of greenhouse gas emissions and carbon dynamics in ecosystems (authors’ conceptualization). The (+) and (−) signs indicate a positive and negative contribution of an activity or process to GHG emissions. Direct arrows show emission sources or processes and dash arrow indicates direct or indirect emission sources
Activities contributing to greenhouse gas emissions
Land conversion for crop farming
Conversion of rangelands to croplands is one of the largest sources of CO2 emissions and the leading cause of loss of soil organic carbon (Conant and Paustian 2002; Farage et al. 2007; Luo et al. 2011; Valentini et al. 2014). Expansion of crop cultivation depletes soil organic matter (SOM) stored in the soil (Smith et al. 2020), thereby decreasing the carbon storage potential in rangelands (Mauda et al. 2017). The availability of SOM is very important in supporting microbes, nutrient cycling and water storage in the soil (Cano et al. 2018). However, in wet and humid areas of the Eastern Africa rangelands, people are increasingly intensifying small-scale crop farming to supplement pastoralism (e.g. Egeru et al. 2010; Reda 2016; Mekuyie et al. 2018). In particular, low-lying wet areas of the rangelands are converted into cultivated croplands (Abate and Angassa 2016). Distinct from livestock grazing, crop farming expansion into rangelands accelerates losses of soil organic carbon (SOC) by erosion and decomposition due to constant disturbance of the land (Franzluebbers et al. 2012). While grassland soils are the major stock of SOC (Batjes 1996), with a sequestration capacity of 90 to 160 Mg C ha−1 (e.g. Lal 2000b), conversion of these landscapes into croplands through permanent or shifting cultivation will decrease the SOC (Lal 2000c, 2003). In the western grasslands of the USA, Lal (2002) reported losses of SOC by 30 to 50% (25–40 Mg C ha−1) in the past century due to the expansion of cultivated lands. In semi-arid rangelands of northern Tanzania, Solomon et al. (2000) revealed a marked decline of the SOC after clearing native vegetation for crop cultivation. According to FAO (2004), grassland soils can lose their SOC at a rate of 0.17 Mg C ha−1 year−1 over the first 25 years to 0.04 Mg C ha−1 year−1 over the next 25 years following land conversion.
Table 1 shows that from 1980 to 2009, the emission of carbon from permanent croplands was higher than that from land under shifting cultivation. The carbon fluxes of land under shifting cultivation are low due to the fallow system that allows recovery of vegetation, and therefore improve sequestration of carbon. Conversely, under permanent cropping, other than the cultivated crop, limited or no vegetation cover is left on the land, resulting in carbon flux than storage. As crop farming is needed to improve food security, implementing recommended management practices in African rangelands is crucial to enhance carbon storage in the soil. Agronomic practices such as conservation tillage, restoration of degraded areas and planting of cover crops have long-term benefits for maintaining soil moisture, vegetation biomass and carbon storage and sequestration in rangeland soils (Olsson and Ardö 2002; Lal 2003). In addition, no-till farming, addition of livestock manure, water harvesting and the construction of water breaks can enhance the storage of organic matter (Lal 2004, 2011; Branca et al. 2013). Table 2 presents various management practices and their contribution to carbon sequestration in rangeland soils.
As shown in Table 2, there are differences in the capacity to sequester carbon for different management practices. This is due to the heterogeneity of agro-ecosystems such soil, climate (see Batjes 2004a), and period of managing. Nevertheless, it is clear that the transitions from croplands to grassland or croplands to fallow have great potential to enhance carbon sequestration (cf. Table 2). Given the spatial variations of soil and land management practices across Eastern Africa rangelands, specific strategies that enhance the uptake of carbon and sequestration potential must be designed based on environments of specific locations. To design appropriate strategies to enhance carbon sequestration potential, there is a need to generate detailed data and comparative studies across regions to capture the variabilities (Tessema et al. 2019). This will help to provide information to rangeland planners and managers on land-use practices that can offset greenhouse gas emissions by enhancing carbon sequestration. Different literature suggests that adopting recommended practices can offer opportunities to store soil organic carbon (e.g. Batjes 2004b). However, in different farming communities, small-scale farmers may be inactive to adopt these practices mainly because of unclear land tenure, poverty and lack of economic incentives. A study by Elias et al. (2015) in agro-pastoral communities of Borana rangeland in Southern Ethiopia revealed that the majority of farmers were uncertain about implementing recommended soil management practices because of unclear land rights. Such conditions challenge efforts aimed at offsetting GHG emissions related to crop farming in the rangelands. Therefore, studies that would consider these constraints and land users are needed because they are the custodians of the land in these rangelands (Reid et al. 2004). As productive wet areas of rangelands are increasingly converted to crop farm for food, higher resolution data from remote sensing technology can help to map the extent and trends (Brink et al. 2014), and fieldwork inventories can be useful for identifying and monitoring carbon dynamics after converting rangelands into croplands.
Livestock husbandry
Livestock farming is the most widespread livelihood activity and the dominant land use in rangeland ecosystems (Alkemade et al. 2013). Yet, the sector is an important source of global GHG emissions. The largest source of emissions in ruminant production is CH4 from enteric fermentation (Valentini et al. 2014) and N2O from feed production (IPCC 2007; Herrero et al. 2013a) while CO2 is associated with the expansion of grazing into grasslands, burning of pastures or land use change (Gerber et al. 2013). Globally, livestock account about 9 to 18% of anthropogenic GHG emissions (IPCC 2007; Caro et al. 2014; Herrero et al. 2015). On a per capita basis in Africa, livestock-related GHG emissions are highest in East and Southern Africa which are about 0.6 t CO2-equivalent/year and in West and Central Africa are 0.4 and 0.3 t CO2-equivalent/year in 2010 (Otte et al. 2019). It is better to understand the sources of livestock husbandry greenhouse gas emissions and mitigation options.
Animal nutrition and feeding
Livestock contribute to CH4 emissions during microbial fermentation of the carbohydrates, proteins, free amino-N and secondary plant components of their feed or forage (McAllister et al. 1996; Kebreab et al. 2006). Johnson and Johnson (1995) found that 4–12% of the gross energy ingested by livestock is converted to CH4 by microbial fermentation in the gastrointestinal tract, though this is largely dependent on feed type. Table 3 shows the estimates of methane gas emissions from enteric fermentation from three livestock systems in rangelands of East and Horn of Africa in 2000 (Herrero et al. 2008). Reported estimates of methane gas emissions in millions of kg CH4 were converted to carbon dioxide equivalent (CO2e) (Table 3).
In addition to the estimates reported by Herrero et al. (2008), a recent study by Otte et al. (2019) reported about 8,6484,000 tonne CO2 eq in 1990 and 128,250,000 tonne CO2 eq 2010 emissions from livestock production systems within the East African region. However, these studies are not sufficient for policy recommendations; more data is required across African regions to guide mitigation strategies. To mitigate enteric methane emission per unit of product is possible through the provision of quality and more digestible feeds (Herrero et al. 2015). Boadi et al. (2004) reported a decline of about 50% in CH4 production from steers grazing on high-quality pastures compared to steers grazing on lower quality pastures. McCaughey et al. (1999) revealed that feeding livestock on alfalfa (Medicago sativa) pastures could reduce CH4 production by 7.1% of gross energy ingested, compared to 9.5% on grass-only pastures. Other feeds such as hay and maize silage can reduce CH4 emissions while also increasing animal productivity (Herrero et al. 2013b; Hristov et al. 2013). Thornton and Herrero (2010) suggested that improved diet, feed digestibility, diet intensification and grain supplements could reduce CH4 emissions from livestock. Depending on the socio-economic and environmental situation, providing improved feeds could be constraints in low input production systems (Herrero et al. 2013a, b, 2015). Although some of the mitigation options reported may have positive results, they may not be feasible for extensive livestock production in Eastern Africa rangelands. This is because grazing depends entirely on mobile systems and pastoralists have different economic and production goals (HPG 2009). In this context, there are few options for improving livestock feeds except in cases where crop residues such as sorghum, teff and wheat straw are available, or under agro-silvo-pastoral systems.
Other challenges facing pastoralism in Africa rangelands are issues of land tenure and privatization, affecting the choices to improve feeds (FIAN 2010; Flintan 2011; Ykhanbai et al. 2014). In communal grazing systems, drought, insecurity and conflict over resources hinder the selection or planting of forage (grass and shrubs). Considering these constraints, the options of reducing CH4 emission through improved feeds are uncertain. It is therefore important to quantify livestock-derived emissions in the Eastern Africa rangelands and explore alternative avenues to minimize GHG emissions.
Manure management
Most livestock in semi-arid and arid rangelands of Africa roam freely for grazing on native pasture during the day and are kept in enclosures during the night. Faeces and urine deposited by animals on the rangeland or in enclosures can be an important source of CH4 and N2O emissions (Zhu et al. 2020). Methane gas is produced through microbial breakdown processes similar to those in the gastrointestinal system of ruminants. Methane from manure is generated in anaerobic conditions through a decomposition process of organic matter in faecal or during handling of livestock excreta (Steinfeld et al. 2006; Dijkstra et al. 2013; Herrero et al. 2015; Otte et al. 2019). Much of the carbon in faeces, when allowed to decompose anaerobically, is converted to CH4 (Forabosco et al. 2017), although temperature, moisture and type of manure (dry or wet matter) also play a role. The emission of nitrous oxide occurs from stored manure and in soil after the addition of manure in farm fields. The emission of nitrogen depends on the level of dietary nitrogen intake as well as the animal’s ability to metabolize nitrogen feeds into protein such as milk and meat (Gay and Knowlton 2009). In situations where livestock consume low to medium quality feeds, only 7–33% of the ingested nitrogen is metabolized while the rest is excreted as dung or urine (Gourley et al. 2012; Silva et al. 2019). This contributes to a significant concentration of nitrogen in excreta. Most inorganic nitrogen (N) in fresh solid manure is in the form of ammonium (Forabosco et al. 2017). Much of the excreta deposited on rangelands can stay without use except in very few areas where farmers use manure on their fields. The environmental conditions such as temperature can influence volatilization from excreta. Under high temperatures, volatilization of animal urine as urea-nitrogen occurs primarily in the form of NH3 and NOx (Caro et al. 2014), hence contributing to emissions. Other biological processes producing N2O and NO are nitrification and denitrification (Dijkstra et al. 2013): N2O gas is produced during the denitrification of NO3− of manure under anaerobic condition (Swamy and Bhattacharya 2006) and occurs in intensive and extensive livestock production systems. The nitrification of surface-dropped faeces by free-range grazing animals in Eastern Africa rangelands produces aerobic NOx (Mosier et al. 1998; Swamy and Bhattacharya 2006), although temperature, soil moisture, soil pH levels and wind speed influence the rate of NOx production (Barton et al. 2008). In developing countries such as the Eastern Africa livestock production systems, the larger emission of methane comes from enteric fermentation of ruminants (cattle, sheep and goats) while for manure, all domestic ruminants contribute to the production of CH4 and N2O gases (Gerber et al. 2013). However, with proper manure management strategies, it is a key element in the mitigation of CH4, N2O and NH3 emissions from livestock (Herrero et al. 2015). Another important element is the improvement of diet to reduce the nitrogen in livestock excreta. For example, dietary tannins affect nitrogen partitioning between faeces and urine, thus channelling a higher proportion of nitrogen to faeces (Somda et al. 1995; Powell et al. 1999). Therefore, tannin reduces nitrogen loss from urine through volatilization because nitrogen in faeces is less volatile than in urine. Finally, proper management of manure remains important not only for Eastern Africa livestock systems but also for continental Africa, because manure stored or abandoned in enclosures remains an active source of N2O (Butterbach-Bahl et al. 2020). A viable option is to use dung and urine in crop fields to replenish C and N in soils that are often weathered or depleted. The practice also shapes the source-sink relationships between different land or vegetation units (Schlecht et al. 2007). Principally, reducing CH4, N2O or NH3 emissions requires substantial data and strategies that support low input livestock production systems and overcome socio-economic and environmental constraints (Derner et al. 2017).
Grazing management
Grazing by livestock is one of the most widespread uses of rangelands and a means of modifying habitats (Brooks et al. 2002; Eldridge et al. 2016). However, grazing can affect soil properties, with direct effects on soil organic carbon (SOC) and soil organic nitrogen (SON) (Baisden and Amundson 2003; Yusuf et al. 2015), but this is influenced by the frequency, intensity and duration of livestock (Liebig et al. 2014). The changes in C and N in grazing lands arise mainly from changes in soil organic matter decomposition and mineralization rates (Piñeiro et al. 2006), as well as animal movement, grazing intensity and associated erosion (Wang et al. 2006). Studies show that the number of livestock and herding mode determines the degree of animal grazing impact (Turner et al. 2005; Schlecht et al. 2006, 2009). Therefore, improving grazing practice and resting the land are important practices for sustainable rangeland management (Garnett 2009).
In open-access systems (such as different parts of African rangelands), where mobile and sedentary forms of livestock coexist, pastures are exploited by multi-species herds. As a result, grazing causes defoliation of plants, over-concentration of livestock excreta on one area and increased ammonia emissions (Arsenault and Owen-Smith 2002). Defoliation of vegetation is another impact of grazing and affects below- and above-ground biomass production in a given vegetation period (Hiernaux and Turner 1996; Quiroga et al. 2018). In perennial and annual herbaceous species, moderate defoliation may also result in over-compensatory growth. In this context, livestock grazing can affect nutrient matter fluxes, particularly in heterogeneous areas (Wezel and Schlecht 2004; Herrero et al. 2013b; Waters et al. 2017). Therefore, varying stock densities, grazing itineraries, herd mobility, enclosures, rest periods and fencing offer the potential to redirect nutrient redistribution processes (Schlecht et al. 1998; Wilkinson et al. 2013).
The practices can offer sustainability in land use where non-equilibrium systems are present (Vetter 2005). Sustainability relates to both the productivity of the grazed pastures and animals. However, implementing such measures depends on production goals, access to grazing lands or water points, property and user rights for grazing lands and the level of economic development (Turner 1995; Selemani 2014). In well-organized livestock production systems, enforcement of customary laws can regulate resource use. However, in the absence of effective traditional resource management practices, proper grazing practice is undermined. Over the past decade, for example, competition and land grabbing for the establishment of biofuel plantations in productive rangeland areas directly interfered with the traditional use of pasture and resulted in the reduction of grazing lands in East Africa (Reid et al. 2014). Intensive grazing by livestock results in trampling which can accelerate the deterioration of vegetation, transforming standing materials into litter and eventually incorporating litter into soil (Hiernaux et al. 1999; Hiernaux 2001). On different soil types (e.g. clays), trampling breaks surface crust, compacts soil and reduces infiltration; consequently, it influences nutrient fluxes and grassland productivity and hence reduces carbon sink potential (Waters et al. 2017). Excessive trampling reduces the ability of plants to access nutrients and water and hence limits plant growth. In turn, the process exhausts carbon reserves and the capacity of grasslands to store carbon (Hiernaux and Turner 1996; Thornton and Herrero 2010; Herrero et al. 2015). Despite the fact that intensive grazing influences soil C and N dynamics and storage in rangelands, few studies have quantified these effects in Eastern Africa (Mcsherry and Ritchie 2013; Angassa 2014; Yusuf et al. 2015). Recently, Oduor et al. (2018a, b) analysed soil organic carbon and microbial biomass in the semi-arid rangeland of Kenya and showed that total SOC and N have increased due to the establishment of pasture enclosures in degraded lands. Similarly, Tessema et al. (2019) show that soil organic carbon stocks in the grasslands of Burundi, Ethiopia, Kenya, Rwanda, Tanzania and Uganda range from 0.1 to 93 Mg C ha−1 in areas under differently managed practices such as grassland regeneration and restoration of degraded lands.
We can deduce that by promoting sustainable grazing through restoring degraded grasslands, moderate grazing and enclosures, there is potential to enhance carbon sequestration in rangelands. This can be achieved if there is enough data on soil carbon stock changes caused by land-use changes, grazing practice or farming expansion across East African rangelands (Bikila et al. 2016). Such assessments will provide a better understanding of the relationship between change in land management and carbon sequestration in rangelands.
Biomass burning
Burning grass, bushes, wood trees and crop residues has been a practice in many pastoral and agro-pastoral traditions (Mammino 2020). In Africa, the practice is more extensive and frequent in rangelands. The demand for wood or charcoal for domestic cooking and heating, expansion of pastureland or farming are the major causes for burning. Fire is extensively used as a tool for managing domestic livestock and indigenous wildlife (Trollope 2011; Keywood et al. 2013). Its primary purpose is to control the encroachment of undesirable plants, stimulate germination of certain seed species and reduce vegetation patchiness and dead plant biomass. Burning replaces nutrients in the soil, paves the way for lush vegetation and manipulates vegetation to optimize forage and pest control (Andrew 1986; Paton and Rickert 1989; Orr et al. 1991; Higgins et al. 2000; Turner et al. 2007; Bowman et al. 2011; Lohmann et al. 2014). Some studies show an increase in the number of recurring burning in African rangelands (Mbow et al. 2000; Van Wilgen et al. 2004).
Besides its positive role, burning deteriorates forage and surface litter materials, changes species composition and may increase wood weeds or undesirable herbs (Trollope 1999; Higgins et al. 2007; Lohmann et al. 2014). Many studies describe the negative effects of burning on soil organisms, the oxidation of humus and the deterioration of organic matter in the soil ecosystem (Ojima et al. 1990, 1994; Crutzen and Andreae 1990; Synmann 2002; Bucini and Lambin 2002; Fynn et al. 2003; Perez et al. 2007; Fynn 2008; Santín and Doerr 2016).
Other effects of burning are the release of gases and aerosols to the atmosphere (Turner et al. 2007). Incomplete combustion contributes to CH4 emissions and NH3 gas is released from the volatilization of nitrogen compounds during biomass burning (Radojevic 2003; Andersson et al. 2004; Edwards et al. 2006; Bell and Adams 2009; Castaldi et al. 2010). Other gases released are CO2, CO, N2O and NOx. Alongside gas emissions, burning intensifies nutrient loss through run-off during rainfall and decreases soil carbon storage in the rangelands (Snyman 2015). In sub-humid Great Plains rangelands, the rate of CO2 flux was higher while soil carbon flux increases with periodic burning on non-grazed areas (Limb et al. 2016). However, there are relatively few studies that have quantified the effects of burning in African rangelands (e.g. Mapiye et al. 2008; Stavi 2019), and even fewer in Eastern Africa. Studies that have quantified the effect of burning in African rangelands have used remotely sensed data without measuring the quantity of gas emissions (van der Werf et al. 2006; Govender et al. 2006; Valentini et al. 2014). Table 4 summarizes the estimates of emission sources from African rangelands.
The reported estimates show not only scanty, but also aggregated data on emissions from four sources. It is possible that emissions attributed to land-use change for pasture or farming expansion could be higher than those associated with wildfire. Since extensive burning on the rangelands is continuing, quantifying their emissions and carbon sequestration potential especially in Eastern Africa is important.
Soil biota activity
The microbes and macro-decomposers play a critical role in the flow of energy through an ecosystem (Dutta and Dutta 2016). Soil microbes break plant litter and mix organic components in the soil (De Deyn et al. 2008) while macro-decomposers such as termites, darkling beetles (Tenebrionidae) and invertebrates (many of which are soil dwellers) prepare litter for soil microbes, mix organic and mineral fractions and carry out nutrient recycling in arid rangelands (MEA 2005).
Although soil organisms play a crucial role in all ecosystem processes, they are rarely the focus of greenhouse gas emissions. Soil organisms significantly contribute to the emissions of CO2 (Hashimoto et al. 2015), CH4 and N2O gases in different situations. However, not many studies have quantified these emissions especially in Eastern Africa rangelands, except a recent one by Oduor et al. (2018a, b) in the semi-arid rangelands of Kenya as they analysed the soil organic carbon and microbial biomass. In different environments, soil microbes (denitrifier) use nitrogen oxides as alternative electron acceptors under anaerobic condition and in the process they release NO, N2O and N2 gases (Sugihara et al. 2012; Butterbach-Bahl et al. 2013). Aerobic emission predominantly originates from nitrifying microbes which convert NH4+ to NO3− to obtain energy and results in N2O production as a by-product of N-transformations (Barton et al. 2008). The microbial and root respiration (autotrophic and heterotrophic) and decomposition also result in CO2 emissions from soil (Austin and Vivanco 2006; Ferréa et al. 2012; Stringer et al. 2012; Hashimoto et al. 2015). Although the emissions occur naturally in ecosystems, high temperatures and dry conditions in arid and semi-arid rangelands can intensify these processes (Jenkinson et al. 1991; Davidson and Janssens 2006; Perez et al. 2007; Bardgett et al. 2008). Without quantifying these emission sources, it is difficult to mitigate (Stringer et al. 2012).
Extensive data about soil organisms are needed to reliably predict how microbial functions and processes contribute to GHG emissions (yet only very few such datasets exist—cf. Table 5). Furthermore, termites in arid and semi-arid rangelands are significant source of CH4, CO2 (Brümmer et al. 2009; Khan et al. 2018; Govorushko 2019) and N2O gas emissions (Brauman et al. 2015). Naturally, methanogenesis in the symbiotic metabolic cascade of lignocellulose in termite hindguts produces the gas (Brune 2014). CH4 and CO2 gases originate from microbial degradation of the ingested feeds like soil or plant materials (Sanderson 1996). However, few studies have quantified the GHG emissions from termites. For example, Brümmer et al. (2009) quantify N2O released from mounds and Nauer et al. (2018) estimate the role of termite mounds to sink CH4. Other studies that examined the activity of termites have focused on the environmental impacts and control measures (Wood 1991), ethno-ecological measures for their management (Sileshi et al. 2009) or their role in vegetation heterogeneity in Africa (Sileshi et al. 2010).
Elsewhere in the world, studies that quantify CH4 and CO2 fluxes from mounds in tropical savannas of northern Australia showed that termites are a significant source of CO2 as compared to CH4 on an annual CO2-e basis (Jamali et al. 2013). An experiment testing the extent of N2O emissions in relation to feed ingested (rich in N and low in N) also showed that N2O production was higher in termites feeding on substrates with higher N content such as soil and fungi compared to those feeding on N-deficient wood (Brauman et al. 2015). This highlights the need to generate long-term data on GHG emissions from soil organisms. There is also evidence that the rates of emissions depend on species, feeding habits, availability of feeds and components, and human disturbances (Jamali et al. 2013; Khan et al. 2018). Earlier studies reported that flagellate protozoa in lower and in higher termite groups emit CO2 and CH4 gases during digestion of feed components (Zimmerman et al. 1982; Sapunov 2008; Velu et al. 2011). The lower and higher termite species could emit up to 0.425 μg CH4 and 0.397 μg CH4 respectively per termite per day (e.g. Zimmerman et al. 1982). Majeed et al. (2012) showed that xylophagous termites could constitute a potential atmospheric sink of N2O from three species studied: Hodotermes mossambicus (grass-harvesting), Nasutitermes voeltzkowi (dry wood feeding), Hodotermopsis sjoestedti (damp wood feeding), in dry savannas and wet forest. Indeed, soil biota play a crucial entity in the emission and removal of greenhouse gas; unfortunately, only a few studies have quantified their role (Khan et al. 2018; Govorushko 2019). It is necessary to consider their roles in different analyses especially for Eastern Africa rangelands. Table 5 compiles the sources of emissions from termites consuming diverse feeds in rangeland ecosystems of the world.
Discussion
Land management and carbon sequestration in rangelands
Terrestrial carbon sequestration is the process through which carbon dioxide from the atmosphere is absorbed by vegetation through photosynthesis and stored as carbon in biomass (tree trunks, branches, grasses, foliage and roots) and soils (Lal 2004, 2008, 2018; Olson et al. 2014). The ability of rangelands to sequester carbon depends on plant species, soil type, regional climate and management practices (Ciais et al. 2011). Basically, the per hectare capacity of rangelands to sequester carbon may be less than that of forestlands; the size of land that rangeland occupies shows a great potential to sequester carbon in below-ground biomass and soils (Derner and Schuman 2007). It is critical that rangelands globally are managed appropriately to develop their potential to sequester carbon. Rangeland management practices that contribute to increase carbon storage in the soil contribute to better soil quality (Tessema et al. 2019). Such management practices must avoid land-use change for farming expansion or deforestation, burning of vegetation and over-grazing. The practices that enhance carbon sequestration potential are moderate grazing, restoring degraded lands, using proper grazing management such as enclosures, implementing non-till farming and sowing drought-tolerant legumes or grass species (Smith et al. 2020). As synthesized in the schematic model (see Fig. 1), it shows that livestock or crop husbandry and plant biomass harvest are the land uses contributing to change the carbon balance in rangelands. They alter carbon and nitrogen cycles, biotic and abiotic processes, and climate system. As a result, they reduce the potential of rangelands to sequester carbon. The potential options to enhance rangelands’ capacity to sequester carbon are regeneration of degraded areas, strategic conservation and reduction in total grazing pressure on sensitive or degraded areas to allow natural revegetation to occur. Using different case studies in the world, Gerber et al. (2013) stated a number of practices that can mitigate GHG emissions with the potential to sequester carbon.
As for the case of Eastern Africa land-use systems, if rangelands are used for livestock husbandry, improving grazing practices, such as herd mobility over diverse landscapes (Dabasso et al. 2014), and revegetation of eroded lands are crucial for enhancing soil carbon storage (Tessema et al. 2019). Proper grazing on grasslands and fire management can increase vegetation cover and build soil carbon pool (Lal 2018). Other practices such as forage improvement, improved animal health and breeding, and manure management are also important. Moreover, since crop farming activity is inevitable in humid areas of rangelands, improving agronomic practices is very important for enhancing soil organic carbon storage, sequestration and food production. Sustainable farming practice that ensures the increased nitrogen availability and water use efficiency accelerates carbon input in the soil ecosystem. The adoption of less tillage (Lal 2013), planting leguminous crops, crop rotation, inter-cropping and adequate fallow periods for cultivated fields increases nitrogen availability and helps to sequester organic carbon in rangelands. Such practices must involve the use of manure and crop residue into cropland fields to increase carbon stocks.
However, before introducing any intervention, location-specific research and more data are needed to recommend practices that enhance carbon sequestration and are accepted in Eastern Africa rangelands. In the absence of scientific research data, it is impossible to implement management strategies that enhance carbon sequestration in rangelands. Additional data on the socio-economic and ecological conditions are needs before introducing soil management practice. At present, few studies have examined the management practices and their contribution on carbon sequestration in African rangelands (Tables 2 and 6). Given the spatial and temporal heterogeneity, a management practice at one area may contribute to carbon sequestration and not on another. In the same way, duration of management practice and land-use history can influence the capacity to sink carbon (Soussana et al. 2010; Ciais et al. 2011). This variability explains one of the weaknesses of relying on continental or regional estimates to guide policy recommendations to land users at the local level. Studies such as Dabasso et al. (2014), Maestre et al. (2016) and Tessema et al. (2019) insisted on the necessity of producing more data in order to capture the heterogeneity across African rangelands.
Conclusion and further research
Globally, enhancement of carbon sequestration in rangelands not only is an essential strategy to mitigate climate change, but forms part of adaptation, through improved land productivity and livelihoods. Building up carbon stocks in rangeland soils through proper land management is a promising cost-effective, natural process to aid in mitigating climate change impacts with numerous benefits. However, given information inconsistency and the knowledge gaps revealed in this review, there is a need to generate more data on land-use and management practices across African climatic zones, to capture the variations in GHG emissions associated with land use or natural process. This information would be useful for supporting climate change policies and adaptation plans. It is necessary to adopt trans-disciplinary methods for promoting social acceptance to land users, to improve methods for estimating carbon flux and to effect technical and policy change. Lastly, this review emphasizes the importance and a need to link national and regional policies to global strategies to reduce emissions of greenhouse gases and enhance carbon sequestration potential in rangelands.
Availability of data and materials
Not applicable.
Abbreviations
- C:
-
Carbon
- CH4 :
-
Methane
- CO:
-
Carbon monoxide
- CO2 :
-
Carbon dioxide
- g:
-
Gramme
- GEI:
-
Gross energy ingested
- GHGs:
-
Greenhouse gases
- h−1 :
-
Per hour per day
- MgC:
-
Megagramme of carbon
- N:
-
Nitrogen
- N2 :
-
Nitrogen gas
- N2O:
-
Nitrous oxide
- NH3 :
-
Ammonia gas
- NO:
-
Nitric oxide gas
- NPP:
-
Net primary productivity
- ppb:
-
Parts per billion
- ppm:
-
Parts per million
- SD:
-
Standard deviation
- SOC:
-
Soil organic carbon
- SON:
-
Soil organic nitrogen
- TgC:
-
Tetragramme of carbon
- UN’s REDD:
-
The United Nations Programme on Reducing Emissions from Deforestation and Forest Degradation
- μg:
-
Microgramme
References
Abate, T., and A. Angassa. 2016. Conversion of savanna rangelands to bush dominated landscape in Borana, Southern Ethiopia. Ecological Processes 5 (6): 6 https://doi.org/10.1186/s13717-016-0049-1.
Alkemade, R., R.S. Reid, M. van den Berg, J. de Leeuw, and M. Jeuken. 2013. Assessing the impacts of livestock production on biodiversity in rangeland ecosystems. Proceedings of the National Academy of Sciences (PNAS) 110 (52): 20900–20905 https://doi.org/10.1073/pnas.1011013108.
Allen, V.G., C. Batello, E.J. Berretta, J. Hodgson, M. Kothmann, X. Li, J. McIvor, J. Milne, C. Morris, A. Peeters, and M. Sanderson. 2011. An international terminology for grazing lands and grazing animals. Grass and Forage Science 66 (1): 2–28 https://doi.org/10.1111/j.1365-2494.2010.00780.x.
Andersson, M., A. Michelsen, M. Jensen, and A. Kjoller. 2004. Tropical savannah woodland: Effects of experimental fire on soil microorganisms and soil emissions of carbon dioxide. Soil Biology & Biochemistry 36 (5): 849–858 https://doi.org/10.1016/j.soilbio.2004.01.015.
Andrew, M.H. 1986. The use of fire for spelling monsoon tallgrass pasture grazed by cattle. Tropical Grasslands 20: 69–78.
Aneja, V.P., W.H. Schlesinger, Q. Li, A. Nahas, and W.H. Battye. 2019. Characterization of atmospheric nitrous oxide emissions from global agricultural soils. SN Applied Sciences 1 (12): 1662 https://doi.org/10.1007/s42452-019-1688-5.
Angassa, A. 2014. Effects of grazing intensity and bush encroachment on herbaceous species and rangeland condition in Southern Ethiopia. Land Degradation and Development 25 (5): 438–451 https://doi.org/10.1002/ldr.2160.
Arsenault, R., and N. Owen-Smith. 2002. Facilitation versus competition in grazing herbivore assemblages. Oikos 97 (3): 313–318 https://doi.org/10.1034/j.1600-0706.2002.970301.x.
Austin, A.T., and L. Vivanco. 2006. Plant litter decomposition in a semi-arid ecosystem controlled by photo-degradation. Nature 442 (7102): 555–558 https://doi.org/10.1038/nature05038.
Baisden, W.T., and R. Amundson. 2003. An analytical approach to ecosystem biogeochemistry modeling. Ecological Applications 13 (3): 649–663 https://doi.org/10.1890/1051-0761(2003)013[0649:AAATEB]2.0.CO;2.
Bardgett, R., C. Freeman, and N.J. Ostle. 2008. Microbial contributions to climate change through carbon cycle feedbacks. The International Society for Microbial Ecology (ISME) Journal 2: 805–814.
Barton, L., R. Kiese, D. Gatter, K. Butterbach-Bahl, R. Buck, C. Hinz, and D.N. Muphy. 2008. Nitrous oxide emissions from a cropped soil in a semi-arid climate. Global Change Biology 14 (1): 177–192 https://doi.org/10.1111/j.1365-2486.2007.01474.x.
Batjes, N.H. 1996. Total carbon and nitrogen in the soils of the world. European Journal of Soil Science 47 (1): 151–163 https://doi.org/10.1111/ejss.12115.
Batjes, N.H. 2004a. Carbon stocks and projected changes according to land use and management: A case study for Kenya. Soil Use and Management 20 (3): 350–356 https://doi.org/10.1079/SUM2004269.
Batjes, N.H. 2004b. Estimation of soil carbon gains upon improved management within croplands and grasslands of Africa. Environment, Development and Sustainability 6 (1/2): 133–143 https://doi.org/10.1023/B:ENVI.0000003633.14591.fd.
Bell, T., and M. Adams. 2009. Smoke from wildfires prescribed burning in Australia: Effects on human health and ecosystems. Development in Environmental Sciences 8: 289–308.
Bikila, N.G., Z.K. Tessema, and E.G. Abule. 2016. Carbon sequestration potentials of semi-arid rangelands under traditional management practices in Borana, Southern Ethiopia. Agriculture, Ecosystems and Environment 223: 108–114 https://doi.org/10.1016/j.agee.2016.02.028.
Boadi, D.A., K.M. Wittenberg, S.L. Scott, D. Burton, K. Buckley, J.A. Small, and K.H. Ominski. 2004. Effect of low and high forage diet on enteric and manure pack greenhouse gas emissions from a feedlot. Canadian Journal of Animal Science 84 (3): 445–453 https://doi.org/10.4141/A03-079.
Bowman, D.M.J.S., J. Balch, P. Artaxo, W.J. Bond, M.A. Cochrane, C.M. D’Antonio, R. DeFries, F.H. Johnston, J.E. Keeley, M.A. Krawchuk, C.A. Kull, M. Mack, M.A. Moritz, S. Pyne, C.I. Roos, A.C. Scott, N.S. Sodhi, and T.W. Swetnam. 2011. The human dimension of fire regimes on Earth. Journal of Biogeography 38 (12): 2223–2236 https://doi.org/10.1111/j.1365-2699.2011.02595.x.
Branca, G., L. Lipper, N. McCarthy, and M.C. Jolejole. 2013. Food security, climate change, and sustainable land management. Agronomy for Sustainable Development 33 (4): 635–650 https://doi.org/10.1007/s13593-013-0133-1.
Brauman, A., M.D. Kane, M. Labat, and J.A. Breznak. 1992. Genesis of acetate and methane by gut bacteria of nutritionally diverse termites. Science 257 (5075): 1384–1387 https://doi.org/10.1126/science.257.5075.1384.
Brauman, A., M.Z. Majeed, B. Buatois, A. Robert, A.-L. Pablo, and E. Miambi. 2015. Nitrous oxide (N2O) emissions by termites: Does the feeding guild matter? PLoS One 10 (12): e0144340 https://doi.org/10.1371/journal.pone.0144340.
Brink, A.B., C. Bodart, L. Brodsky, P. Defourney, C. Ernst, F. Donney, A. Lupi, and K. Tuckova. 2014. Anthropogenic pressure in East Africa—Monitoring 20 years of land cover changes by means of medium resolution satellite data. International Journal of Applied Earth Observation and Geoinformation 28: 60–69 https://doi.org/10.1016/j.jag.2013.11.006.
Brooks, T.M., R.A. Mittermeier, C.G. Mittermeier, G.A.B. Da Fonseca, A.B. Rylands, W.R. Konstant, P. Flick, J. Pilgrim, S. Oldfield, G. Magin, and C. Hilton-Taylor. 2002. Habitat loss and extinction in the hotspots of biodiversity. Conservation Biology 16 (4): 909–923 https://doi.org/10.1046/j.1523-1739.2002.00530.x.
Brümmer, C., H. Papen, R. Wassmann, and N. Brüggemann. 2009. Termite mounds as hot spots of nitrous oxide emissions in South-Sudanian savanna of Burkina Faso (West Africa). Geophysical Research Letters 36 (L09814): 1–4 https://doi.org/10.1029/2009GL037351.
Brune, A. 2014. Symbiotic digestion of lignocellulose in termite guts. Nature Reviews Microbiology 12 (3): 168–180 https://doi.org/10.1038/nrmicro3182.
Bucini, G., and E.F. Lambin. 2002. Fire impacts on vegetation in Central Africa: A remote-sensing-based statistical analysis. Applied Geography 22 (1): 27–48 https://doi.org/10.1016/S0143-6228(01)00020-0.
Butterbach-Bahl, K., E.M. Baggs, M. Dannenmann, R. Kiese, and S. Zechmeister-Boltenstern. 2013. Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Philosophical Transactions of The Royal Society B Biological Sciences 368 (1621): 20130122 https://doi.org/10.1098/rstb.2013.0122.
Butterbach-Bahl, K., G. Gettel, R. Kiese, K. Fuchs, C. Werner, J. Rahimi, M. Barthel, and L. Merbold. 2020. Livestock enclosures in drylands of Sub-Saharan Africa are overlooked hotspots of N2O emissions. Nature Communications 11 (1): 4644 https://doi.org/10.1038/s41467-020-18359-y.
Cano, A., A. Núñez, V. Acosta-Martinez, M. Schipanski, R. Ghimire, C. Rice, and C. West. 2018. Current knowledge and future research directions of soil health and water conservation in the Ogallala aquifer region. Geoderma 328: 109–118 https://doi.org/10.1016/j.geoderma.2018.04.027.
Caro, D., S.J. Davis, S. Bastianoni, and K. Caldeira. 2014. Global and regional trends in greenhouse gas emissions from livestock. Climatic Change 126 (1-2): 203–216 https://doi.org/10.1007/s10584-014-1197-x.
Castaldi, S., A. de Grandcourt, A. Rasile, U. Skiba, and R. Valentini. 2010. CO2, CH4 and N2O fluxes from soil of burned grassland in central Africa. Biogeoscience 7 (11): 3459–3471 https://doi.org/10.5194/bg-7-3459-2010.
Ciais, P., A. Bombelli, M. Williams, S.L. Piao, J. Chave, C.M. Ryan, M. Henry, P. Brender, and R. Valentini. 2011. The carbon balance of Africa: Synthesis of recent research studies. Philosophical Transactions of the Royal Society A 369: 1–20.
Conant, R.T., and K. Paustian. 2002. Potential soil carbon sequestration in overgrazed grassland ecosystems. Global Biogeochemical Cycles 16 (4): 11–43 https://doi.org/10.1029/2001GB001661.
Conant, R.T., K. Paustian, and E.T. Elliott. 2001. Grassland management and conversion into grassland: Effects on soil carbon. Ecological Applications 11 (2): 343–355 https://doi.org/10.1890/1051-0761(2001)011[0343:GMACIG]2.0.CO;2.
Crutzen, P.J., and M.O. Andreae. 1990. Biomass burning in the tropics: Impact on atmospheric chemistry and biogeochemical cycles. Science 250 (4988): 1669–1678 https://doi.org/10.1126/science.250.4988.1669.
Dabasso, B.H., Z. Taddese, and D. Hoag. 2014. Carbon stocks in semi-arid pastoral ecosystems of northern Kenya. Pastoralism: Research, Policy and Practice 4 (5): 1–8 http://www.pastoralismjournal.com/content/4/1/5.
Davidson, E.A., and I.A. Janssens. 2006. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440 (7081): 165–173 https://doi.org/10.1038/nature04514.
De Deyn, G.B., J.H.C. Cornelissen, and R.D. Bardgett. 2008. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecology Letters 11 (5): 516–531 https://doi.org/10.1111/j.1461-0248.2008.01164.x.
Dean, C., J.B. Kirkpatrick, R.J. Harper, and D.J. Eldridge. 2015. Optimising carbon sequestration in arid and semiarid rangelands. Ecological Engineering 74: 148–163 https://doi.org/10.1016/j.ecoleng.2014.09.125.
Derner, J.D., L. Hunt, K.E. Filho, J. Ritten, J. Capper, and G. Han. 2017. Livestock production systems. In Rangeland systems processes, management and challenges, ed. D.D. Briske, 347–372. Chapter 10. Springer Series on Environmental Management https://doi.org/10.1007/978-3-319-46709-2_10.
Derner, J.D., and G.E. Schuman. 2007. Carbon sequestration and rangelands: A synthesis of land management effects. Journal of Soil and Water Conservation 62: 77–85.
Dijkstra, J., O. Oenema, J.W. van Groenigen, J.W. Spek, A.M. van Vuuren, and A. Bannink. 2013. Diet effects on urine composition of cattle and N2O emissions. Animal 7 (s2): 292–302 https://doi.org/10.1017/S1751731113000578.
Don, A., J. Schumacher, and A. Freibauer. 2011. Impact of tropical land-use change on soil organic carbon stocks - A meta-analysis. Global Change Biology 17 (4): 1658–1670 https://doi.org/10.1111/j.1365-2486.2010.02336.x.
Dutta, H., and A. Dutta. 2016. The microbial aspect of climate change. Energy, Ecology and Environment 1 (4): 209–232 https://doi.org/10.1007/s40974-016-0034-7.
Edwards, D.P., L.K. Emmons, J.C. Gille, A. Chu, J.L. Attie, L. Giglio, S.W. Wood, J. Haywood, M.N. Deeter, S.T. Massie, D.C. Ziskin, and J.R. Drummond. 2006. Satellite-observed pollution from Southern Hemisphere biomass burning. Journal of Geophysical Research 111 (D14312) https://doi.org/10.1029/2005JD006655.
EEA (European Environment Agency). 2019. Global annual averaged atmospheric concentration of nitrous oxide (N2O), from 1750–2016, measured in parts per billion (ppb). https://www.eea.europa.eu/data-and-maps/indicators/atmospheric-greenhouse-gas-concentrations-6/assessment. In Ritchie H., and Roser M. (2020) – N2O. OurWorldInData.org. https://ourworldindata.org/N2O. Accessed in June 2020.
EEA (European Environment Agency) and NOAA (National Oceanic and Atmospheric Administration). 2019. Global annual averaged atmospheric concentration of methane (CH4), from 1750–2018 measured in parts per billion (ppb). In Ritchie H., and Roser M. (2020) - CH4. OurWorldInData.org. https://ourworldindata.org/CH4. Accessed in June 2020.
Egeru, A., P. Okello, M.G.J. Majaliwa, P. Mukwaya, and P. Isubikalu. 2010. The effects of land cover change on biomass stock in dryland areas of Eastern Uganda. A case study of Olio Sub-county in Soroti District. Journal of Applied Sciences and Environmental Management 14 (4): 101–106.
Eldridge, D.J., A.G.B. Poore, M. Ruiz-Colmenero, M. Letnic, and S. Soliveres. 2016. Ecosystem structure, function, and composition in rangelands are negatively affected by livestock grazing. Ecological Applications 26 (4): 1273–1283 https://doi.org/10.1890/15-1234.
Elias, M., O. Hensel, U. Richter, C. Hülsebusch, B. Kaufmann, and O. Wasonga. 2015. Land conversion dynamics in the Borana rangelands of Southern Ethiopia: An integrated assessment using remote sensing techniques and field survey data. Environments 2 (4): 1–31 https://doi.org/10.3390/environments2010001.
FAO. 2004. Carbon sequestration in dryland soils. In World soils resources reports 102. Rome: Food and Agriculture Organization of the United Nations.
FAO. 2019. Trees, forests and land use in drylands: The first global assessment – Full report. Rome: FAO Forestry Paper No. 184.
FAO. 2020. Global Forest Resources Assessment 2020 – Key findings. Rome: https://doi.org/10.4060/ca8753en.
FAO and UNEP. 2020. The State of the World’s Forests 2020. Forests, biodiversity and people. Rome: https://doi.org/10.4060/ca8642en.
Farage, P.K., J. Ardö, L. Olsson, E.A. Rienzi, A.S. Ball, and J.N. Pretty. 2007. The potential for soil carbon sequestration in three tropical dryland farming systems of Africa and Latin America: A modelling approach. Soil & Tillage Research 94 (2): 457–472 https://doi.org/10.1016/j.still.2006.09.006.
Ferréa, C., T. Zenoneb, R. Comolli, and G. Seufert. 2012. Estimating heterotrophic and autotrophic soil respiration in a semi-natural forest of Lombardy, Italy. Pedobiologia 55: 285–294 https://doi.org/10.1016/j.pedobi.2012.05.001.
FIAN. 2010. In Land Grabbing in Kenya and Mozambique. A report on two research missions and human rights analysis of land grabbing, ed. E. Odeny, R. Leonhard, B. Saturnino Jr., and M. Rocha, 1–43. Germany: FIAN.
Flintan, F. 2011. The causes, process and impacts of land fragmentation in the rangelands of Ethiopia, Kenya and Uganda. Regional Learning and Advocacy programme for Vulnerable Dryland Communities, 1–12.
Forabosco, F., Zh. Chitchyan, and R. Mantovani. 2017. Methane, nitrous oxide emissions and mitigation strategies for livestock in developing countries: A review. South African Journal of Animal Science 47 (3): 268–270 https://doi.org/10.4314/sajas.v47i3.3.
Franzluebbers, A.J., L.K. Paine, J.R. Winsten, M. Krome, M.A. Sanderson, K. Ogles, and D. Thompson. 2012. Well-managed grazing systems: A forgotten hero of conservation. Journal of Soil and Water Conservation 67 (4): 100A–104A https://doi.org/10.2489/jswc.67.4.100A.
Fynn, R. 2008. Savory insights – Is rangeland science due for a paradigm shift? Rangeland management, Grassroots. Newsletter of the Grassland Society of Southern Africa 8 (3): 25–38.
Fynn, R.W.S., R.J. Haynes, and T.G. O’Connor. 2003. Burning causes long-term changes in soil organic matter content of South African grassland. Soil Biology & Biochemistry 35 (5): 677–687 https://doi.org/10.1016/S0038-0717(03)00054-3.
Garnett, T. 2009. Livestock-related greenhouse gas emissions: Impacts and options for policy makers. Environmental Science & Policy 12 (4): 491–503 https://doi.org/10.1016/j.envsci.2009.01.006.
Gay, S.W., and K.F. Knowlton. 2009. Ammonia emissions and animal agriculture. Virginia Cooperative Extension. Publication 442–110: 1–5.
Gerber, P.J., H. Steinfeld, B. Henderson, A. Mottet, C. Opio, J. Dijkman, A. Falcucci, and G. Tempio. 2013. Tackling climate change through livestock – A global assessment of emissions and mitigation opportunities. Rome: Food and Agriculture Organization of the United Nations.
Godde, C.M., R.B. Boone, A.J. Ash, K. Waha, L.L. Sloat, P.K. Thornton, and M. Herrero. 2020. Global rangeland production systems and livelihoods at threat under climate change and variability. Environmental Research Letters 15 (044021): 1–15 https://doi.org/10.1088/1748-9326/ab7395.
Gourley, C.J.P., S.R. Aarons, and J.M. Powell. 2012. Nitrogen use efficiency and manure management practices in contrasting dairy production systems. Agriculture, Ecosystems and Environment 147: 73–81 https://doi.org/10.1016/j.agee.2011.05.011.
Govender, N., W.S.W. Trollope, and B.W. Van Wilgen. 2006. The effect of fire season, fire frequency, rainfall and management on fire intensity in savanna vegetation in South Africa. Journal of Applied Ecology 43 (4): 748–758 https://doi.org/10.1111/j.1365-2664.2006.01184.x.
Govorushko, S. 2019. Economic and ecological importance of termites: A global review. Entomological Science 22 (1): 21–35 https://doi.org/10.1111/ens.12328.
Grace, J., J. San José, P. Meir, H.S. Miranda, and R.A. Montes. 2006. Productivity and carbon fluxes of tropical savannas. Journal of Biogeography 33 (3): 387–400 https://doi.org/10.1111/j.1365-2699.0148x.
Hashimoto, S., N. Carvalhais, A. Ito, M. Migliavacca, K. Nishina, and M. Reichstein. 2015. Global spatiotemporal distribution of soil respiration modeled using a global database. Biogeosciences 12 (13): 4121–4132 https://doi.org/10.5194/bg-12-4121-2015.
Herrero, M., D. Grace, J. Njuki, N. Johnson, D. Enahoro, S. Silvestri, and M.C. Rufino. 2013a. The roles of livestock in developing countries. Animal 7 (1): 3–18 https://doi.org/10.1017/S1751731112001954.
Herrero, M., P. Havlík, H. Valin, A. Notenbaert, M.C. Rufino, P.K. Thornton, M. Blümmel, F. Weiss, D. Grace, and M. Obersteiner. 2013b. Biomass use, production, feed efficiencies and greenhouse gas emissions from global livestock systems. PNAS 110 (52): 20888–20893 www.pnas.org/cgi/doi/10.1073/pnas.1308149110.
Herrero, M., P.K. Thornton, R. Kruska, and R.S. Reid. 2008. Systems dynamics and the spatial distribution of methane emissions from African domestic ruminants to 2030. Agriculture, Ecosystems and Environment 126 (1-2): 122–137 https://doi.org/10.1016/j.agee.2008.01.017.
Herrero, M., S. Wirsenius, B. Henderson, C. Rigolot, P. Thornton, P. Havlík, I. de Boer, and P.J. Gerber. 2015. Livestock and the environment: What have we learned in the past decade? Annual Review of Environment and Resources 40 (1): 177–202. https://doi.org/10.1146/annurev-environ-031113-093503.
Hiernaux, P. 2001. Fondements écologiques de la gestion des parcours au Sahel. In Elevage ET gestion de parcours au Sahel, implications pour le développement, ed. E. Tielkes, E. Schlecht, and P. Hiernaux, 65–77. Beuren-Stuttgart: Verlag E. Grauer.
Hiernaux, P., C.L. Bielders, C. Valentin, A. Bationo, and S. Fernández-Rivera. 1999. Effects of livestock grazing on physical and chemical properties of sandy soils in Sahelian rangelands. Journal of Arid Environments 41 (3): 231–245 https://doi.org/10.1006/jare.1998.0475.
Hiernaux, P., and M.D. Turner. 1996. The effect of clipping on growth and nutrient uptake of Sahelian annual rangelands. Journal of Applied Ecology 33 (2): 387–399 https://doi.org/10.2307/2404760.
Higgins, S.I., W.J. Bond, E.C. February, A. Bronn, and D.I.W. Euston-Brown. 2007. Effects of four decades of fire manipulation on woody vegetation structure in savanna. Journal of Ecology 88 (5): 1119–1125 https://doi.org/10.1890/06-1664.
Higgins, S.I., W.J. Bond, and W.S.W. Trollope. 2000. Fire, resprouting and variability: A recipe for grass-tree coexistence in savanna. Journal of Ecology 88 (2): 213–229 https://doi.org/10.1046/j.1365-2745.2000.00435.x.
Hoffman, T.M., and C. Vogel. 2008. Climate change impacts on African rangelands. Rangelands 30 (3): 12–17 https://doi.org/10.2111/1551-501X(2008)30[12:CCIOAR]2.0.CO;2.
Houghton, R.A., J.I. House, J. Pongratz, G.R. van der Werf, R.S. DeFries, M.C. Hansen, C. Le Quéré, and N. Ramankutty. 2012. Carbon emissions from land use and land-cover change. Biogeosciences 9 (12): 5125–5142 https://doi.org/10.5194/bg-9-5125-2012.
HPG (Humanitarian Policy Group). 2009. Pastoralism, policies and practice in the Horn and East Africa. A review of current trends. Synthesis Paper, 1–4.
Hristov, A.N., J. Oh, C. Lee, R. Meinen, F. Montes, T. Ott, J. Firkins, A. Rotz, C. Dell, A. Adesogan, W. Yang, J. Tricarico, E. Kebreab, G. Waghorn, J. Dijkstra, and S. Oosting. 2013. Mitigation of greenhouse gas emissions in livestock production – A review of technical options for non-CO2 emissions. In Animal Production and Health Paper No. 177, ed. P.J. Gerber, B. Henderson, and H.P.S. Makkar. Rome: FAO.
IIED and SOS. 2010. In Modern and mobile: The future of livestock production in Africa’s drylands, ed. H. De Jode, 11–87. International Institute for Environment & Development (IIED) and SOS Sahel International UK.
IPCC. 2007. Climate change 2007: Mitigation. In Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Summary for Policymakers, ed. B. Metz, O.R. Davidson, P.R. Bosch, R. Dave, and L.A. Meyer. Cambridge and New York: Cambridge University Press.
IPCC. 2013. Climate change 2013: The physical science basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, ed. T.F. Stocker, D. Qin, G.-K. Plattner, M. Tignor, S.K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex, and P.M. Midgley, 4–29. Cambridge and New York: Cambridge University Press.
Jamali, H., S.J. Livesley, L.B. Hutley, B. Fest, and S.K. Arndt. 2013. The relationships between termite mound CH4/CO2 emissions and internal concentration ratios are species specific. Biogeosciences 10 (4): 2229–2240 https://doi.org/10.5194/bg-10-2229-2013.
Jenkinson, D.S., D.E. Adams, and A. Wild. 1991. Model estimates of CO2 emissions from soil in response to global warming. Nature 351 (6324): 304–306 https://doi.org/10.1038/351304a0.
Johnson, K.A., and D.E. Johnson. 1995. Methane emissions from cattle. Journal of Animal Science 73 (8): 2483–2492 https://doi.org/10.2527/1995.7382483x.
Kebreab, E., K. Clark, R. Wagner, and J. France. 2006. Methane and nitrous oxide emissions from Canadian animal agriculture: A review. Canadian Journal of Animal Science 86: 135–158.
Keenan, T.F., and C.A. Williams. 2018. The terrestrial carbon sink. Annual Review of Environment and Resources 43 (1): 219–243 https://doi.org/10.1146/annurev-environ-102017-030204.
Keywood, M., M. Kanakidou, A. Stohl, F. Dentener, G. Grassi, C.P. Meyer, K. Torseth, D. Edwards, A.M. Thompson, U. Lohmann, and J. Burrows. 2013. Fire in the air: Biomass burning impacts in a changing climate. Critical Reviews in Environmental Science and Technology 43 (1): 40–83 https://doi.org/10.1080/10643389.2011.604248.
Khan, Md, A.W. Ahmad, and B. Paul. 2018. Ecological impacts of termites. In Termites and sustainable management, Sustainability in plant and crop protection, ed. M.A. Khan and W. Ahmad. Springer International Publishing AG 2018 https://doi.org/10.1007/978-3-319-72110-1_10.Chapter10.
Kideghesho, J.R., A.A. Rija, K.A. Mwamende, and I.S. Selemani. 2013. Emerging issues and challenges in conservation of biodiversity in the rangelands of Tanzania. Nature Conservation 6: 1–29 https://doi.org/10.3897/natureconservation.6.5407.
Laban, P., G. Metternicht, and J. Davies. 2018. Soil biodiversity and soil organic carbon: Keeping drylands alive, viii + 24. Gland: IUCN.
Lal, R. 2000b. Carbon sequestration in drylands. Annals of Arid Zone 39 (1): 1–10.
Lal, R. 2000c. World cropland soils as a source or sink for atmospheric carbon. Advances in Agronomy 71: 145–191.
Lal, R. 2002. Soil carbon dynamics in cropland and rangeland. Environmental Pollution 116 (3): 353–362 https://doi.org/10.1016/S0269-7491(01)00211-1.
Lal, R. 2003. Carbon sequestration in dryland ecosystems. Environmental Management 33 (4): 528–544 https://doi.org/10.1007/s00267-003-9110-9.
Lal, R. 2004. Soil carbon sequestration to mitigate climate change. Geoderma 123 (1-2): 1–22 https://doi.org/10.1016/j.geoderma.2004.01.032.
Lal, R. 2008. Carbon sequestration. Philosophical Transactions of The Royal Society B 363 (1492): 815–830 https://doi.org/10.1098/rstb.2007.2185.
Lal, R. 2011. Sequestering carbon in soils of agro-ecosystems. Food Policy 36: 33–39 https://doi.org/10.1016/j.foodpol.2010.12.001.
Lal, R. 2013. Soil carbon management and climate change. Carbon Management 4 (4): 439–462 https://doi.org/10.4155/cmt.13.31.
Lal, R. 2018. Digging deeper: A holistic perspective of factors affecting soil organic carbon sequestration in agroecosystems. Global Change Biology 24 (8): 3285–3301 https://doi.org/10.1111/gcb.14054.
Lehsten, V., K. Tansey, H. Balzter, K. Thonicke, A. Spessa, U. Weber, B. Smith, and A. Arneth. 2009. Estimating carbon emissions from African wildfires. Biogeosciences 6 (3): 349–360 www.biogeosciences.net/6/349/2009/. https://doi.org/10.5194/bg-6-349-2009.
Liebig, M.A., S.L. Kronberg, J.R. Hendrickson, and J.R. Gross. 2014. Grazing management, season, and drought contributions to near-surface soil property dynamics in semiarid rangelands. Rangeland Ecology & Management 67 (3): 266–274 https://doi.org/10.2111/REM-D-13-00145.1.
Limb, R.F., S.D. Fuhlendorf, D.M. Engle, and R.F. Miller. 2016. Synthesis paper: Assessment of research on rangeland fire as a management practice. Rangeland Ecology & Management 69 (6): 415–422 https://doi.org/10.1016/j.rama.2016.07.013.
Liniger, H.P., and R.M. Studer. 2019. Sustainable rangeland management in Sub-Saharan Africa – Guidelines to good practice. TerrAfrica; World Bank, Washington D.C.; World Overview of Conservation Approaches and Technologies (WOCAT); World Bank Group (WBG), Washington DC, USA and Centre for Development and Environment (CDE), University of Bern, Switzerland.
Lipper, L., C. Dutilly-Diane, and N. McCarthy. 2010. Supplying carbon sequestration from West African rangelands: Opportunities and barriers. Rangeland Ecology & Management 63 (1): 155–166 https://doi.org/10.2111/REM-D-09-00009.1.
Lohmann, D., B. Tietjen, N. Blaum, D.F. Joubert, and F. Jeltsch. 2014. Prescribed fire as a tool for managing shrub encroachment in semi-arid savanna rangelands. Journal of Arid Environments 107: 49–56 https://doi.org/10.1016/j.jaridenv.2014.04.003.
Luo, Z., E. Wang, O.J. Sun, C.J. Smith, and M.E. Probert. 2011. Modeling long-term soil carbon dynamics and sequestration potential in semi-arid agro-ecosystems. Agricultural and Forest Meteorology 151 (12): 1529–1544 https://doi.org/10.1016/j.agrformet.2011.06.011.
MacCarthy, D.S., and R.B. Zougmoré. 2018. Assessment of greenhouse gas emissions from different land-use systems: A case study of CO2 in the Southern Zone of Ghana. Applied and Environmental Soil Science Article ID 1057242: 1–12 https://doi.org/10.1155/2018/1057242.
Maestre, F.T., D.J. Eldridge, S. Soliveres, S. Kéfi, M. Delgado-Baquerizo, M.A. Bowker, P. García-Palacios, J. Gaitán, A. Gallardo, R. Lázaro, and M. Berdugo. 2016. Structure and functioning of dryland ecosystems in a changing world. Annual Review of Ecology, Evolution, and Systematics 47 (1): 215–237 https://doi.org/10.1146/annurev-ecolsys-121415-032311.
Majeed, M.Z., E. Miambi, A. Robert, M. Bernoux, and A. Brauman. 2012. Xylophagous termites: A potential sink for atmospheric nitrous oxide. European Journal of Soil Biology 53: 121–125 https://doi.org/10.1016/j.ejsobi.2012.10.002.
Mammino, L. 2020. Biomass burning in sub-Saharan Africa: Chemical issues and action outreach. Dordrecht: Springer Nature B.V https://doi.org/10.1007/978-94-007-0808-2.
Mapiye, C., M. Mwale, N. Chikumba, and M. Chimonyo. 2008. Fire as a rangeland management tool in the savannas of southern Africa: A review. Tropical and Subtropical Agroecosystems 8 (2): 115–124.
Mauda, E.V., G.S. Joseph, C.L. Seymour, T.C. Munyai, and S.H. Foord. 2017. Changes in land use alter ant diversity, assemblage composition and dominant functional groups in African savannas. Biodiversity and Conservation 27 (4): 947–965 https://doi.org/10.1007/s10531-017-1474-x.
Mbow, C., T.T. Nielson, and K. Rausmussen. 2000. Savanna fires in east central Senegal: Distribution patterns, resource management perceptions. Human Ecology 28 (4): 561–583 https://doi.org/10.1023/A:1026487730947.
McAllister, T.A., E.K. Okine, G.W. Mathison, and K.J. Cheng. 1996. Dietary, environmental and microbiological aspects of methane production in ruminants. Canadian Journal of Animal Science 76 (2): 231–243 https://doi.org/10.4141/cjas96-035.
McCaughey, W.P., K.M. Wittenberg, and D. Corrigan. 1999. Impact of pasture type on methane production by lactating beef cows. Canadian Journal of Animal Science 79 (2): 221–226 https://doi.org/10.4141/A98-107.
McDermot, C., and S. Elavarthi. 2014. Rangelands as carbon sinks to mitigate climate change: A review. Journal of Earth Science and Climate Change 5 (08): 221 https://doi.org/10.4172/2157-7617.1000221.
Mcsherry, M.E., and M.E. Ritchie. 2013. Effects of grazing on grassland soil carbon: A global review. Global Change Biology 19 (5): 1347–1357 https://doi.org/10.1111/gcb.12144.
MEA. 2005. Millennium ecosystem assessment and human well-being: Desertification synthesis: By Safriel, U., Z. Adeel, D. Niemeijer, J. Puigdefabregas, R. White, R. Lal, M. Wilson, J. Ziedler, S. Prince, E. Archer, C. King, B. Shapiro, K. Wessels, T. Nielsen, B. Portnov, I. Reshef, J. Thonell, E. Lachman, and D. McNab. In Ecosystems and human well-being: Current state and trends, ed. R. Hassan, R. Scholes, and N. Ash, 625–653. Washington, DC: World Resource Institute.Chapter 22 “Dryland Systems”. Volume 1.
Mekuyie, M., A. Jordaan, and Y. Melka. 2018. Land-use and land-cover changes and their drivers in rangeland-dependent pastoral communities in the southern Afar Region of Ethiopia. African Journal of Range and Forage Science 35 (1): 33–43 https://doi.org/10.2989/10220119.2018.1442366.
Mortimore, M., S. Anderson, L. Cotula, J. Davies, K. Faccer, C. Hesse, J. Morton, W. Nyangena, J. Skinner, and C. Wolfangel. 2009. Dryland opportunities: A new paradigm for people, ecosystems and development, 86. Gland, London, Nairobi: IUCN, IIED, UNDP/DDC.
Mosier, A., C. Kroeze, C. Nevison, O. Oenema, S. Seitzinger, and O. van Cleemput. 1998. Closing the global N2O budget: nitrous oxide emissions through the agricultural nitrogen cycle. Nutrient Cycling in Agroecosystems 52 (2/3): 225–248 https://doi.org/10.1023/A:1009740530221.
Nauer, P.A., L.B. Hutley, and S.K. Arndt. 2018. Termite mounds mitigate half of termite methane emissions. PNAS 115 (52): 13306–13311 www.pnas.org/cgi/doi/10.1073/pnas.1809790115.
Neely, C., S. Bunning, and A. Wilkes, eds. 2009. Review of evidence on drylands pastoral systems and climate change: Implications and opportunities for mitigation and adaptation. Land and Water Discussion Paper 8, 1–38. Rome: FAO.
Niles, J.O., S. Brown, J. Pretty, A.S. Ball, and J. Fay. 2002. Potential carbon mitigation and income in developing countries from changes in use and management of agricultural and forest lands. Philosophical Transactions of the Royal Society of London. Series A, Mathematical, Physical, and Engineering Sciences 360 (1797): 1621–1639 https://doi.org/10.1098/rsta.2002.1023.
NOAA/ESRL (National Oceanic and Atmospheric Administration). 2018. Global mean annual concentration of carbon dioxide (CO2) from 1959 to 2018, measured in parts per million (ppm). https://www.esrl.noaa.gov/gmd/ccgg/trends/data.html. In Hannah Ritchie and Max Roser (2020) - “CO2 and greenhouse gas emissions”. OurWorldInData.org. https://ourworldindata.org/co2-and-other-greenhouse-gas-emissions. Accessed in June 2020.
Nosetto, M.D., E.G. Jobbágy, and J.M. Paruelo. 2006. Carbon sequestration in semi-arid rangelands: Comparison of Pinus ponderosa plantations and grazing exclusion in NW Patagonia. Journal of Arid Environments 67 (1): 142–156 https://doi.org/10.1016/j.jaridenv.2005.12.008.
Oduor, C.O., N. Karanja, R. Onwong’a, S. Mureithi, D. Pelster, and G. Nyberg. 2018a. Pasture enclosures increase soil carbon dioxide flux rate in Semiarid Rangeland, Kenya. Carbon Balance and Management 13 (24): 1–12 https://doi.org/10.1186/s13021-018-0114-4.
Oduor, C.O., N.K. Karanja, R.N. Onwonga, S.M. Mureithi, D. Pelster, and G. Nyberg. 2018b. Enhancing soil organic carbon, particulate organic carbon and microbial biomass in semi-arid rangeland using pasture enclosures. BMC Ecology 18 (45): 1–9 https://doi.org/10.1186/s12898-018-0202-z.
Ojima, D.S., W.J. Parton, D.S. Schimel, and C.E. Owensby. 1990. Simulated impacts of annual burning on prairie ecosystems. In Fire in North American tall grass prairie, ed. S.L. Collins and L.L. Wallace, 118–132. Norman: University of Oklahoma Press.
Ojima, D.S., D.S. Schimel, W.J. Parton, and C.E. Owensby. 1994. Long and short-term effects of fire on nitrogen cycling in tallgrass prairie. Biogeochemistry 24 (2): 67–84 https://doi.org/10.1007/BF02390180.
Olson, K.R., M. Al-Kaisi, R. Lal, and B. Lowery. 2014. Experimental considerations, treatments and methods in determining soil organic carbon sequestration rates. Soil Science Society of America Journal 78 (2): 348–360 https://doi.org/10.2136/sssaj2013.09.0412.
Olsson, L., and J. Ardö. 2002. Soil carbon sequestration in degraded semiarid agro-ecosystems—Perils and potentials. A Journal of the Human Environment 31 (6): 471–477 https://doi.org/10.1579/0044-7447-31.6.471.
Orr, D.M., G.M. McKeon, and K.A. Day. 1991. Burning and exlosure can rehabilitate degraded black speargrass (Heteropogon contortus) pasture. Tropical Grasslands 25: 333–336.
Otte, J., U. Pica-Ciamarra, and S. Morzaria. 2019. A comparative overview of the livestock-environment interactions in Asia and Sub-Saharan Africa. Frontiers in Vetenary Science 6 (37): 1–13 https://doi.org/10.3389/fvets.2019.00037.
Pan, Y., R.A. Birdsey, J. Fang, R. Houghton, P.E. Kauppi, W.A. Kurz, O.L. Phillips, A. Shvidenko, S.L. Lewis, J.G. Canadell, P. Ciais, R.B. Jackson, S.W. Pacala, A.D. McGuire, S. Piao, A. Rautiainen, S. Sitch, and D. Hayes. 2011. A large and persistent carbon sink in the world’s forests. Science 333 (6045): 988–993 https://doi.org/10.1126/science.1201609.
Paton, C.J., and K.G. Rickert. 1989. Burning, then resting, reduces wiregrass (Aristida ssp.) in black speargrass pastures. Tropical Grasslands 23: 211–216.
Perez, C., C. Roncoli, C. Neely, and J.L. Steiner. 2007. Can carbon sequestration markets benefit low-income producers in semi-arid Africa? Potentials and challenges. Agricultural Systems 94 (1): 2–12 https://doi.org/10.1016/j.agsy.2005.09.009.
Piñeiro, G., J.M. Paruelo, and M. Oesterheld. 2006. Potential long-term impacts of livestock introduction on carbon and nitrogen cycling in grasslands of Southern South America. Global Change Biology 12 (7): 1267–1284 https://doi.org/10.1111/j.1365-2486.2006.01173.x.
Powell, J.M., F.N. Ikpe, and Z.C. Somda. 1999. Crop yield and the fate of nitrogen and phosphorus following application of plant material and faeces to soil. Nutrient Cycling in Agroecosystems 54 (3): 215–226 https://doi.org/10.1023/A:1009762900258.
Pugh, T.A.M., M. Lindeskog, B. Smith, B. Poultere, A. Arneth, V. Haverd, and L. Callee. 2019. Role of forest regrowth in global carbon sink dynamics. PNAS 116 (10): 4382–4387 https://doi.org/10.1073/pnas.1810512116.
Quiroga, R.E., L.J. Blanco, and P.R. Namur. 2018. Defoliation intensity and simulated grazing strategy effects on three-C4 rangeland bunchgrasses. Rangeland Ecology & Management 71 (1): 58–66 https://doi.org/10.1016/j.rama.2017.09.002.
Radojevic, M. 2003. Chemistry of forestry fires and regional haze emphasis on southern Asia. Pure and Applied Geophysics 160 (1-2): 157–187 https://doi.org/10.1007/s00024-003-8771-x.
Reda, T. 2016. Dynamics in pastoral resource management and conflict in the Borana rangelands of southern Ethiopia. African Security Review 25 (1): 31–43 https://doi.org/10.1080/10246029.2015.1126526.
Reid, R.S., M.E. Fernández-Giménez, and K.A. Galvin. 2014. Dynamics and resilience of rangelands and pastoral peoples around the globe. Annual Review of Environment and Resources 39 (1): 217–242 https://doi.org/10.1146/annurev-environ-020713-163329.
Reid, R.S., P.K. Thornton, G.J. McCrabb, R.L. Kruska, F. Atieno, and P.G. Jones. 2004. Is it possible to mitigate greenhouse gas emissions in pastoral ecosystems of the tropics? Environment. Development and Sustainability 6 (1/2): 91–109 https://doi.org/10.1023/B:ENVI.0000003631.43271.6b.
Ringius, L. 2002. Soil carbon sequestration and the CDM: Opportunities and challenges for Africa. Climatic Change 54 (4): 471–495 https://doi.org/10.1023/A:1016108215242.
Ritchie, H., and M. Roser. 2019. “CO2 and greenhouse gas emissions”. Published online at OurWorldInData.org. https://ourworldindata.org/co2. Accessed in June 2020.
Rouland, C., A. Brauman, M. Labat, and M. Lepage. 1993. Nutritional factors affecting methane emission from termites. Chemosphere 26 (1932): 617–622 https://doi.org/10.1016/0045-6535(93)90447-D.
Sanderson, M.G. 1996. Biomass of termites and their emissions of methane and carbon dioxide: A global database. Global Biogeochemical Cycles 10 (4): 543–557 https://doi.org/10.1029/96GB01893.
Santín, C., and S.H. Doerr. 2016. Fire effects on soils: The human dimension. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 371 (20150171) https://doi.org/10.1098/rstb.2015.0171.
Sapunov, V.B. 2008. Global dynamics of termite population: Modeling, control and role in greenhouse effect. In Proceeding of the Sixth International Conference on Urban Pest, ed. W.H. Robinson and D. Bajomi, 390–393.
Scharlemann, J.P.W., E.V.J. Tanner, R. Hiederer, and V. Kapos. 2014. Global soil carbon: Understanding and managing the largest terrestrial carbon pool. Carbon Management 5 (1): 81–91 https://doi.org/10.4155/cmt.13.77.
Schlecht, E., U. Dickhoefer, E. Gumpertsberger, and A. Buerkert. 2009. Grazing itineraries and forage selection of goats in the Jabal al Akhdar Mountain range of northern Oman. Journal of Arid Environments 73 (3): 355–363 https://doi.org/10.1016/j.jaridenv.2008.10.013.
Schlecht, E., S. Fernández-Rivera, and P. Hiernaux. 1998. Timing, size and N-concentration of faecal and urinary excretions in cattle, sheep and goats - Can they be used for better manuring of cropland? In Soil fertility management in West African land use systems, ed. G. Renard, A. Neef, K. Becker, and M. von Oppen, 361–368. Weikersheim: Margraf Verlag.
Schlecht, E., P. Hiernaux, I. Kadaouré, C. Hülsebusch, and F. Mahler. 2006. A spatio-temporal analysis of forage availability, grazing and excretion behaviour of cattle, sheep and goats in Western Niger. Agriculture, Ecosystems and Environment 113 (1-4): 226–242 https://doi.org/10.1016/j.agee.2005.09.008.
Schlecht, E., H. Richter, S. Fernández-Rivera, and K. Becker. 2007. Gastrointestinal passage of Sahelian roughages in cattle, sheep and goats, and implications for livestock-mediated nutrient transfers. Animal Feed Science and Technology 137 (1-2): 93–114 https://doi.org/10.1016/j.anifeedsci.2006.11.001.
Scholes, R.J., S. Archibald, and G. von Maltitz. 2011. Emissions from fire in Sub-Saharan Africa: The magnitude of sources, their variability and uncertainty. Global Environmental Research 15: 53–63.
Selemani, I.S. 2014. Communal rangelands management and challenges underpinning pastoral mobility in Tanzania: A review. Livestock Research for Rural Development 26 (78) http://www.lrrd.org/lrrd26/5/sele26078.html. Accessed 30 Sept 2021.
Sharma, P., V. Abrol, S. Abrol, and R. Kumar. 2012. Climate change and carbon sequestration in dryland soils. Resource Management for Sustainable Agriculture Chapter 6: 140–164 https://doi.org/10.5772/52103.
Sileshi, G.W., M.A. Arshad, S. Konaté, and P.O.Y. Nkunika. 2010. Termite-induced heterogeneity in African savanna vegetation: mechanisms and patterns. Journal of Vegetation Science 21 (5): 923–937 https://doi.org/10.1111/j.1654-1103.2010.01197.x.
Sileshi, G.W., P. Nyeko, O.Y. Nkunika, B.M. Sekematte, F.K. Akinnifesi, and O.C. Ajayi. 2009. Integrating ethno-ecological and scientific knowledge of termites for sustainable termite management and human welfare in Africa. Ecology and Society 14 (1): 48 http://www.ecologyandsociety.org/vol14/iss1/art48/.
Silva, L.F.P., R.M. Dixon, and D.F.A. Costa. 2019. Nitrogen recycling and feed efficiency of cattle fed protein-restricted diets. Animal Production Science 59 (11): 2093–2107 https://doi.org/10.1071/AN19234.
Smith, P., J. Soussana, D. Angers, L. Schipper, C. Chenu, D.P. Rasse, N.H. Batjes, F. van Egmond, S. McNeill, M. Kuhnert, C. Arias-Navarro, J.E. Olesen, N. Chirinda, D. Fornara, E. Wollenberg, J. Álvaro-Fuentes, A. Sanz-Cobena, and K. Klumpp. 2020. How to measure, report and verify soil carbon change to realize the potential of soil carbon sequestration for atmospheric greenhouse gas removal. Global Change Biology 26: 219–241 https://doi.org/10.1111/gcb.14815.
Snyder, H. 2019. Literature review as a research methodology: An overview and guidelines. Journal of Business Research 104: 333–339 https://doi.org/10.1016/j.jbusres.2019.07.039.
Snyman, H.A. 2015. Short-term responses of Southern African semi-arid rangelands to fire: A review of impact on soils. Arid Land Research and Management 29 (2): 222–236 https://doi.org/10.1080/15324982.2014.944244.
Solomon, D., J. Lehmann, and W. Zech. 2000. Land-use effects on soil organic matter properties of chromic luvisols in semi-arid northern Tanzania: Carbon, nitrogen, lignin and carbohydrates. Agriculture, Ecosystems and Environment 78 (3): 203–213 https://doi.org/10.1016/S0167-8809(99)00126-7.
Somda, Z.C., J.M. Powell, S. Fernández-Rivera, and J. Reed. 1995. Feed factors affecting nutrient excretion by ruminants and the fate of nutrients when applied to soil. In Livestock and sustainable nutrient cycling in mixed farming systems of sub-Saharan Africa, ed. J.M. Powell, S. Fernández-Rivera, T.O. Williams, and C. Renard, 227–243. Addis Ababa: International Livestock Center for Africa 2.
Soussana, J.F., T. Tallec, and V. Blanfort. 2010. Mitigating the greenhouse gas balance of ruminant production systems through carbon sequestration in grasslands. Animal 4 (3): 334–350 https://doi.org/10.1017/S1751731109990784.
Stavi, I. 2019. Wildfires in Grasslands and Shrublands: A Review of Impacts on Vegetation, Soil, Hydrology, and Geomorphology. Water 11: 1042 https://doi.org/10.3390/w11051042.
Steinfeld, H., P. Gerber, T. Wassenaar, V. Castel, M. Rosales, and C. de Haan. 2006. Livestock’s long shadow-environmental issues and options, Chapter 3 & 4, 79–176. Rome: FAO.
Stringer, L.C., A.J. Dougill, A.D. Thomas, D.V. Spracklen, S. Chesterman, C. IfejikaSperanza, H. Rueff, M. Riddell, M. Williams, T. Beedy, D.J. Abson, P. Klintenberg, S. Syampungani, P. Powell, A.R. Palmer, M.K. Seely, D.D. Mkwambisi, M. Falcao, A. Sitoe, S. Ross, and G. Kopolo. 2012. Challenges and opportunities in linking carbon sequestration, livelihoods and ecosystem service provision in drylands. Environmental Science and Policy 19 (20): 121–135 https://doi.org/10.1016/j.envsci.2012.02.004.
Sugihara, S., S. Funakawa, M. Kilasara, and T. Kosaki. 2012. Effects of land management on microbial N supply to crop N uptake in dry tropical cropland in Tanzania. Agriculture, Ecosystems and Environment 146 (1): 209–219 https://doi.org/10.1016/j.agee.2011.11.008.
Swamy, M., and S. Bhattacharya. 2006. Budgeting anthropogenic greenhouse gas emission from Indian livestock using country specific emission coefficients. Current Science 91: 1340–1353.
Synmann, H.A. 2002. Fire and the dynamics of semi-arid grassland: Influence on soil characteristics. African Journal of Range and Forage Science 19 (3): 137–145 https://doi.org/10.2989/10220110209485786.
Tennigkeit, T., and A. Wilkies. 2008. An assessment of the potential for carbon finance in rangelands. ICRAF Working Paper 68: 1–31.
Tessema, B., R. Sommer, K. Piikki, M. Söderström, S. Namirembe, A. Notenbaert, L. Tamene, S. Nyawira, and B. Paul. 2019. Potential for soil organic carbon sequestration in grasslands in East African countries: A review. Grassland Science 2020 (00): 1–10 https://doi.org/10.1111/grs.12267.
Thornton, P.K., and M. Herrero. 2010. Potential for reduced methane and carbon dioxide emissions from livestock and pasture management in the tropics. PNAS 107 (46): 19667–19672 www.pnas.org/cgi/doi/10.1073/pnas.0912890107.
Trollope, W.S.W. 1999. Veld burning. In Veld and pasture management in South Africa, ed. N.M. Tainton, 217–243. Pieternmaritzburg: Shutter and Shooter.
Trollope, W.S.W. 2011. Personal perspectives on commercial versus communal African fire paradigms when using fire to manage rangelands for domestic livestock and wildlife in southern and east African ecosystems. Fire Ecology 7 (1): 57–73 https://doi.org/10.4996/fireecology.0701057.
Turner, M.D. 1995. The sustainability of rangeland to cropland nutrient transfer in semi-arid West Africa: Ecological and social dimensions neglected in the debate. In Livestock and sustainable nutrient cycling in mixed farming systems of sub-Saharan Africa, eds, J. M. Powell, S. Fernández-Rivera, T.O. Williams and C. Renard, 1993(II): 435–452, Proceedings of a conference at ILCA. Addis Ababa, Ethiopia, Nov. 22-26.
Turner, M.D., P. Hiernaux, and E. Schlecht. 2005. The distribution of grazing pressure in relation to vegetation resources in semi-arid West Africa: The role of herding. Ecosystems 8 (6): 668–281 https://doi.org/10.1007/s10021-003-0099-y.
Turner, M.G., E.A.H. Smithwick, K.L. Metzger, D.B. Tinker, and W.H. Romme. 2007. Inorganic nitrogen availability after severe stand-replacing fire in the Greater Yellowstone ecosystem. PNAS 104 (12): 124782–124789 www.pnas.org/cgi/doi/10.1073/pnas.0700180104.
UN. 2011. Global drylands: A UN system-wide response. Prepared by the Environment Management Group. Chapter 1–4, 14–129.
UN. 2018. Forests: A natural solution to climate change, crucial for a sustainable future. 10th Consolidated Annual Progress Report of the UN-REDD Programme Fund, 11–13. Geneva: Chemin des Anémones, CH-1219 Châtelaine www.un-redd.org. Accessed 4 June 2020.
UNEP. 2008. Carbon in drylands: Desertification, climate change and carbon finance: A UNEP-UNDP-UNCCD Technical Note for Discussion at CRIC 7: 2–12, Istanbul, Turkey-03-14 November, 2008.
UNEP. 2009. Climate change in the African drylands: Options and opportunities for adaptation and mitigation. UNEP, UNCCD and UNDP, United Nations Development Programme, 17–44. New York: One UN Plaza.
Vagen, T.G., R. Lal, and B.R. Singh. 2005. Soil carbon sequestration in Sub-Saharan Africa: A review. Land Degradation and Development 16 (1): 53–71 https://doi.org/10.1002/ldr.644.
Valentini, R., A. Arneth, A. Bombelli, S. Castaldi, C.R. Gatti, F. Chevallier, P. Ciais, E. Grieco, J. Hartmann, M. Henry, R.A. Houghton, M. Jung, W.L. Kutsch, Y. Malhi, E. Mayorga, L. Merbold, G. Murray-Tortarolo, D. Papale, P. Peylin, B. Poulter, P.A. Raymond, M. Santini, S. Sitch, G.V. Laurin, G.R. van der Werf, C.A. Williams, and R.J. Scholes. 2014. A full greenhouse gases budget of Africa: Synthesis, uncertainties, and vulnerabilities. Biogeosciences 11 (2): 381–407 https://doi.org/10.5194/bg-11-381-2014.
van der Werf, G.R., J.T. Randerson, L. Giglio, G.J. Collatz, P.S. Kasibhatla, and A.F. Arellano Jr. 2006. Inter-annual variability in global biomass burning emissions from 1997 to 2004. Atmospheric Chemistry and Physics 6 (11): 3423–3441 https://doi.org/10.5194/acp-6-3423-2006.
Van Wilgen, B.W., N. Govender, H.C. Biggs, D. Ntsala, and X.N. Funda. 2004. Response of savanna fire regimes to changing fire-management policies in a large African national park. Conservation Biology 18 (6): 1533–1540 https://www.jstor.org/stable/3589034. https://doi.org/10.1111/j.1523-1739.2004.00362.x.
Velu, G., K. Ramasamy, K. Kumar, N. Sivaramaiah, and V.R.M. Ramanjaneya. 2011. Green house gas emission from termite’s ecosystem. African Journal of Environmental Science and Technology 5 (2): 56–64.
Vetter, S. 2005. Rangelands at equilibrium and non-equilibrium: Recent developments in the debate. Journal of Arid Environments 62 (2): 321–241 https://doi.org/10.1016/j.jaridenv.2004.11.015.
Wang, C., S. Wan, X. Xing, L. Zhang, and X. Han. 2006. Temperature and soil moisture interactively affected soil net N mineralization in temperate grassland in Northern China. Soil Biology and Biochemistry 38 (5): 1101–1110 https://doi.org/10.1016/j.soilbio.2005.09.009.
Waters, C.M., S.E. Orgill, G.J. Melville, I.D. Toole, and W.J. Smith. 2017. Management of grazing intensity in the semi-arid rangelands of Southern Australia: Effects on soil and biodiversity. Land Degradation and Development 28 (4): 1363–1375 https://doi.org/10.1002/ldr.2602.
Wezel, A., and E. Schlecht. 2004. Inter-annual variation of species composition in fallow plant communities of semi-arid Niger. Journal of Arid Environments 56 (2): 265–282 https://doi.org/10.1016/S0140-1963(03)00049-1.
Wilkes, A., and T. Tennigkeit. 2010. Carbon finance in extensively managed rangelands: Issues in project, programmatic and sectoral approached. In Grassland carbon sequestration: Management, policy and economics; CHAPTER XII. Vol. 11, 211–233.
Wilkinson, S.N., G.J. Hancocka, R. Bartley, A.A. Hawdon, and R.J. Keen. 2013. Using sediment tracing to assess processes and spatial patterns of erosion in grazed rangelands, Burdekin River basin, Australia. Agriculture, Ecosystems and Environment 180: 90–102 https://doi.org/10.1016/j.agee.2012.02.002.
Wood, T.G. 1991. Termites in Ethiopia: The environmental impact of their damage and resultant control measures. Ambio 20 (3/4): 136–138 http://www.jstor.org/stable/4313800.
WRI (World Resources Institute). 2000. In Pilot analysis of global ecosystems grassland ecosystems, ed. R.P. White, S. Murray, and M. Rohweder, 1–70 http://www.wri.org/wr2000.
WRI (World Resources Institute). 2003. In Drylands, people, and ecosystem goods and services: A web-based geospatial analysis, ed. R.P. White and J. Nackoney, 1–58.
Ykhanbai, H., R. Garg, A. Singh, S. Moiko, C.E. Beyene, D. Roe, F. Nelson, T. Blomley, and F. Flintan. 2014. Conservation and “land grabbing” in rangelands: Part of the problem or part of the solution? International Land Coalition, +42.
Yusuf, H.M., A.C. Treydte, and J. Sauerborn. 2015. Managing semi-arid rangelands for carbon storage: Grazing and woody encroachment effects on soil carbon and nitrogen. PLoS One 10 (10): 1–18. e0109063 https://doi.org/10.1371/journal.pone.0109063.
Zhu, Y., L. Merbold, S. Leitner, L. Xia, D.E. Pelster, E. Diaz-Pines, S. Abwanda, P.M. Mutuo, and K. Butterbach-Bahl. 2020. Influence of soil properties on N2O and CO2 emissions from excreta deposited on tropical pastures in Kenya. Soil Biology and Biochemistry 140 (107636) https://doi.org/10.1016/j.soilbio.2019.107636.
Zimmerman, P.R., J.P. Greenberg, S.O. Wandiga, and P. Crutzen. 1982. Termites: A potentially large source of atmospheric methane, carbon dioxide, and molecular hydrogen. Science 218 (4572): 563–565 https://doi.org/10.1126/science.218.4572.563.
Acknowledgements
The authors acknowledge the German Academic Exchange Service (DAAD), the Ministry of Education and Vocational Training (MoEVT) of Tanzania, the Federal Ministry for Economic Cooperation and Development (BMZ) of Germany and the German Institute for Tropical and Subtropical Agriculture (DITSL) in Witzenhausen, Germany, for supporting this work.
Funding
The research was funded by the German Academic Exchange Service (DAAD) and the Ministry of Education and Vocational Training (MoEVT) of Tanzania through the scholarship programme received by Michael Mgalula.
Author information
Authors and Affiliations
Contributions
All authors contributed to the conceptualization of the idea; MEM, OVW, CH, UR and OH: collection of secondary data; MEM, OVW, CH and UR: data analysis and preparation of the article. All authors read and approved the final manuscript and take responsibility for its content.
Authors’ information
Not applicable.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mgalula, M.E., Wasonga, O.V., Hülsebusch, C. et al. Greenhouse gas emissions and carbon sink potential in Eastern Africa rangeland ecosystems: A review. Pastoralism 11, 19 (2021). https://doi.org/10.1186/s13570-021-00201-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13570-021-00201-9