
Abstract
Background
In utero exposure to diabetes has been shown to contribute to preterm birth, though the underlying biological mechanisms are yet to be fully elucidated. Fetal epigenetic variations established in utero may be a possible pathway. This study aimed to investigate whether in utero exposure to diabetes was associated with a change in newborn DNA methylation, and whether the identified CpG sites mediate the association between diabetes and preterm birth in a racially diverse birth cohort population.
Methods
This study included 954 mother–newborn pairs. Methylation levels in the cord blood were determined using the Illumina Infinium MethylationEPIC BeadChip 850 K array platform. In utero exposure to diabetes was defined by the presence of maternal pregestational or gestational diabetes. Preterm birth was defined as gestational age at birth less than 37 weeks. Linear regression analysis was employed to identify differentially methylated CpG sites. Differentially methylated regions were identified using the DMRcate Package.
Results
126 (13%) newborns were born to mothers with diabetes in pregnancy and 173 (18%) newborns were born preterm, while 41 newborns were born both preterm and to mothers with diabetes in pregnancy. Genomic-wide CpG analysis found that eighteen CpG sites in cord blood were differentially methylated by maternal diabetes status at an FDR threshold of 5%. These significant CpG sites were mapped to 12 known genes, one of which was annotated to gene Major Histocompatibility Complex, Class II, DM Beta (HLA-DMB). Consistently, one of the two identified significant methylated regions overlapped with HLA-DMB. The identified differentially methylated CpG sites mediated the association between diabetes in pregnancy and preterm birth by 61%.
Conclusions
In this US birth cohort, we found that maternal diabetes was associated with altered fetal DNA methylation patterns, which substantially explained the link between diabetes and preterm birth.
Similar content being viewed by others

Background
With the increasing prevalence of obesity, the rates of type 2 diabetes and gestational diabetes (GDM) have been rising globally [1]. The consequences of these rising rates are that in utero exposure to diabetes has become a common scenario that is linked to a wide range of detrimental health outcomes [2,3,4]. A growing number of studies have demonstrated that in utero exposure to diabetes has contributed to adverse birth outcomes, such as preterm birth and macrosomia [3, 4]. Along the same lines, even maternal hyperglycemia which did not met the criteria of diabetes diagnosis was also significantly associated with preterm birth [5]. However, the underlying biological mechanisms driving these outcomes are yet to be fully elucidated.
Preterm birth, a birth occurring at less than 37 completed gestational weeks, has been identified as the most important cause of perinatal mortality and infant morbidity and has a lifelong impact [6]. Despite limited understanding about the root cause and biological mechanisms underlying preterm birth, it is increasingly recognized that genetic and environmental interaction plays a critical role [7, 8], while epigenetic mechanisms are likely the interface that coordinates environmental factors with genes to drive such changes [9]. Studies have provided evidence that fetal epigenetic variations [10], which are largely established in utero [11], may contribute to preterm birth [12]. Several studies have documented that maternal diabetes in pregnancy was associated with altered methylation levels in cord blood DNA [13,14,15]. However, little is known about whether and to what degree the influence of diabetes has on preterm birth through variation in DNA methylation.
Using the Boston Birth Cohort (BBC), a well-established racially and ethnically diverse birth cohort, we sought to identify the associations between exposure to diabetes in utero (assessed as maternal diabetes in pregnancy) and DNA methylation changes in cord blood. Next, we further explored whether these differentially methylated positions (DMPs) mediated the association between diabetes and preterm birth.
Results
Study characteristics
This study included 954 maternal-infant dyads. Mean (standard deviation (SD)) maternal age at delivery was 28.3 (6.6) years. Among the mothers, 679 (71.2%) self-identified as non-Hispanic Black and 126 (13.2%) experienced diabetes (including GDM (n = 83), type 2 diabetes (n = 39), and type 1 diabetes (n = 4)). Compared to mothers with GDM, mothers who had pregestational diabetes had higher blood glucose concentrations and a higher proportion used insulin as treatment (Additional file 1: Table S1). Among the newborns, 504 (52.8%) were male and 173 (18.1%) were born prematurely, of which 41 newborns were born to mothers with diabetes in pregnancy. Maternal and neonatal characteristics are shown in Table 1. As expected, mothers with diabetes were more likely to be older, had higher prepregnancy body mass index (BMI), and were more likely to be with overweight or obesity. Newborns of mothers with diabetes were more likely to have shorter gestation and to be born prematurely, to be large for gestational age (LGA), and to have lower proportions of granulocytes and nature killer cells and higher proportion of nucleated red blood cells in their cord blood. An overall predominance of non-Hispanic Black race and ethnicity was observed among participating mothers both with and without diabetes.
Epigenome-wide analysis of differentially methylated positions (DMPs)
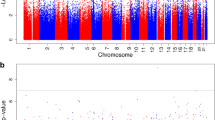
Quantile–quantile (Q-Q) plots and lambda (λ) values (1.14) are provided in Additional file 1: Fig. S1. Epigenome-wide associations between diabetes in pregnancy and cord blood DNA methylation are visualized in the Manhattan plot (Additional file 1: Fig. S2). As shown in Table 2, 18 CpG probes were identified as significantly differentially methylated at a false discovery rate (FDR) of P < 0.05 and annotated to 12 known genes. Among these DMPs, the top DMP was located within the 5 prime untranslated region (5’UTR) of the Leucine Rich Repeat Containing 8 VRAC Subunit B (LRRC8B) gene, which is involved in anion transmembrane transport; two top DMPs were located in genes Dynamin 3 (DNM3) and Family With Sequence Similarity 163 Member A (FAM163A), which are involved in components and functions of membranes; and another top DMP was mapped to the body of gene Major Histocompatibility Complex (MHC), Class II, DM Beta (HLA-DMB), which is associated with autoimmune diseases. Of these DMPs, 17 (94.4%) showed hypermethylation in newborns whose mothers had diabetes in pregnancy, whereas only 1 DMP (cg21363811) located within the CpG island of the FAM163A gene showed hypomethylation (Table 2). When analyses were conducted separately by GDM and pregestational diabetes, the association patterns were similar (Additional file 1: Table S2). Furthermore, no significantly differential methylation levels were found in the identified DMPs between GDM and pregestational diabetes (Additional file 1: Table S3).
Identification of differentially methylated regions (DMRs)
DMRs were investigated using ‘DMRcate’ by windows 500- and 1000-bp. Of the 4 DMRs identified by window 500-bp, 2 were significant using an FDR threshold of 0.10, including chromosome 17: 19631464–19631485, which contained 2 probes and lacked overlapping known genes and chromosome 6: 32908466–32908718, which contained 5 probes located within the 5’UTR and first exon as well as the body of the HLA-DMB gene, which is associated with autoimmune diseases [16,17,18] (Table 3). All probes covered in the two significant DMRs had higher methylation levels in the cord blood of mothers with diabetes (Additional file 1: Table S4). Although three DMRs were identified by window 1000-bp, none of them survived after correction for multiple testing (Additional file 1: Table S5).
Lookups in the literature
Of 10 studies [13,14,15, 19,20,21,22,23,24,25] measured DNA methylation using the 450 K or 850 K microarray and investigated the associations between maternal pregestational diabetes/GDM and cord blood DNA methylation, 4 studies [13, 14, 22, 23] reported DMPs on EWAS. Two study was removed due to small sample size of GDM cases (n < 50) [22, 23]. Across the 2 studies [13, 14], 307 unique CpGs were associated with diabetes exposure after adjustment for multiple comparisons. Of these CpGs, 274 were evaluated in the current study. Of 274 CpGs examined, 120 were nominally associated with diabetes in pregnancy and 115 with the same direction of effect, of which, 87 CpGs were significant with an FDR-corrected P value less than 0.05 based on 274 tests conducted (Additional file 1: Table S6).
Gene set enrichment analysis
We did not find statistically significant enriched pathways using ontologies defined in the KEGG (Kyoto Encyclopedia of Genes and Genomes) and GO (Gene Ontology) databases at an FDR p < 0.05. However, as shown by the enrichment results at both the DMP-level and DMR-level, the top 10 KEGG pathways showed enrichment of inflammatory and immune-related pathways (Additional file 1: Tables S7 and S9). Notably, the top 10 KEGG pathways in DMRs identified by windows of both 500-bp and 1000-bp were highly concordant. Along this same functional theme, GO terms for DMRs showed enrichment of immune response (Additional file 1: Tables S10), while GO terms for DMPs included multiple arginine metabolism pathways, which are involved in cellular metabolism and function [26] (Additional file 1: Table S8).
Mediation effect of DMPs
The associations of diabetes in pregnancy and 18 identified DMPs with preterm birth and subtypes are presented in Additional file 1: Table S11. Diabetes in pregnancy and 17 DMPs were significantly associated with an increased risk of preterm birth, especially medically indicated preterm birth. One CpG (cg21363811) mapped to gene FAM163A was associated with a reduced risk of preterm birth. Among the 18 DMPs, 5 were selected as mediators in the association between diabetes in pregnancy and preterm birth at a p level of < 0.10 (Additional file 1: Table S12). Taken as a whole, the 5 DMPs mediated 61% of the association (Table 4). In sensitivity analyses, we evaluated the mediation effect of these DMPs on medically indicated preterm. About 66% of the associations of diabetes with indicated preterm birth were explained by these DMPs (Additional file 1: Table S13).
Discussion
In this study, we examined the associations between maternal diabetes in pregnancy and cord blood DNA methylation patterns at both individual and regional CpG levels. Eighteen individual CpG sites and two regions were identified to differentially methylate by diabetes in pregnancy. In our data, the directions of the effect for DMPs were mostly positive, indicating a hypermethylation effect. Furthermore, our data showed that diabetes in pregnancy was associated with an increased risk of preterm birth, and about 61% of the association was explained by the DMPs.
LRRC8B, encoding leucine-rich repeat containing 8 family, member B, is one of the LRRC8 gene family members and LRRC8B protein acting as a leak channel in the endoplasmic reticulum plays an important role in intracellular Ca2+ homeostasis [27]. Although altered methylation in gene LRRC8B has not been previously reported in existing literature as differentially methylated in cord blood in regard to maternal diabetes in pregnancy, cell-type-specific methylation changes in gene of LRRC8B have been associated with Alzheimer's disease [28]. Previous studies have also implicated the role of prenatal exposure to diabetes in autism and neuropsychiatric disease [29, 30]. Another hit was on cg25953130, located in the body of ARID5B gene which is involved in the regulation of the transcription of target genes involved in adipogenesis and liver development and plays a role in cell growth and differentiation of B-lymphocyte progenitors [31]. In agreement with our findings, Antoun et al. also found that a CpG site which was annotated to the ARID5B gene was positively associated with GDM [13].
One DMP (cg22324029) was annotated to the HLA-DMB gene, which was previously linked to several autoimmune diseases, such as multiple sclerosis [16], rheumatoid arthritis [17], and type 1 diabetes [18]. Consistently, one significant DMR (Chr 6, 32908239–32909282) also overlapped with the HLA-DMB gene, suggesting that the association between diabetes in pregnancy and the CpG site was robust. More importantly, this DMR covered 4 CpG sites in the 5’UTR and exon 1 of HLA-DMB, indicating the regulatory functions of the DMR. HLA-DMB belongs to the HLA class II beta chain paralogs, which play a critical role in the peptide loading of major histocompatibility complex (MHC) class II molecules. The abnormal DNA methylation of HLA-DMB modified the gene expression and was associated with an increased risk of asthma [32], suggesting HLA-DMB is involved in human immune response. In support of our finding, a previous study reported an association between GDM and a CpG site located in gene MHC, class II, DR beta (HLA-DRB) [13], an important paralog of HLA-DMB; and alterations in DNAm of the MHC region in placentas from mothers with GDM [25, 33], as well as in cord blood of offspring from mothers with GDM [24]. In addition, strong enrichment of inflammation and immune response pathways of DNAm was also found in human amniocytes exposed to GDM in utero [34]. Gene set enrichment analysis also showed that these identified DMPs and DMRs were involved in the pathways that are integral to autoimmune diseases. Previous studies showed that maternal hyperglycemia can exacerbate inflammatory and immune responses during pregnancy, leading to adverse fetal outcomes [35]. Epidemiological study also provides evidence that GDM increases the risk of asthma in the offspring [36].
In agreement with previous studies which reported a positive association between preterm birth and type 1 diabetes [3], type2 diabetes [4], and GDM [37], our data showed a strong association between diabetes in pregnancy and preterm birth and its subtype (medically indicated preterm birth). We explored beyond previous studies and demonstrated that the DMPs induced by diabetes mediated the association by more than two-thirds. Since diabetes-induced DMPs were enriched in the immune response and inflammatory pathway, our data support the hypothesis implicating the involvement of inflammatory and immune response in the etiology of preterm birth [38]. In further support of our findings, one study using two dependent cohorts also showed that the preterm birth-associated CpGs were mapped to genes enriched for inflammation and immune pathways [12].
This study has several strengths. First, to the best of our knowledge, this study is the first to explore the mediation effect of DNA methylation patterns induced by diabetes in pregnancy on the associations between diabetes and preterm birth. Second, our study examined the DMRs in addition to DMPs.
Several limitations should also be acknowledged. First, the number of newborns born to mothers with diabetes in pregnancy included in this study was relatively small, especially for that coupled with preterm birth. Second, a lambda of 1.14 suggests that not all possible test-statistic inflation has been ruled out. Third, due to the combination of GDM with pregestational diabetes, we cannot differentiate the effects of the two diabetic conditions on the cord blood methylation. Unavailability of blood glucose data for mothers without diabetes limited our ability to investigate the effect of glycemia on the methylation. Fourth, although the look-up analysis showed that some DMPs identified in our study were consistent with previous studies, future studies are needed to replicate our results in larger samples. Fifth, because of a substantial missing in the genotyping data, we could not perform mQTL analysis. Finally, the study population is a predominantly urban and primarily Black cohort from low-resourced communities and households; thus, caution is needed when assessing the generalizability of the findings to other populations with different characteristics.
Conclusion
This study found that exposure to diabetes in utero was associated with DMPs and DMRs. Furthermore, these identified DNA methylation alterations substantially mediated the association between diabetes in pregnancy and preterm births.
Materials and methods
Study design and population
The study population was a subset of the BBC, an ongoing prospective birth cohort, recruited at 2–3 days after delivery at Boston Medical Center, MA, USA, starting in 1998. A detailed description of the BBC has been reported previously [39]. In brief, at enrollment, each mother completed a questionnaire interview that gathered demographic information. Cord blood was collected immediately after birth. As of December 2018, 8623 mother-infant pairs had been enrolled. Of these pairs, 963 cord blood samples were analyzed by DNA methylation array. Of these, 9 samples were removed due to not passing the quality control process. Finally, this study included 954 mother-infant pairs. Maternal demographic characteristics and birth outcomes were comparable between the study sample included and the total BBC, except that more Black participants and preterm births among the sample were included in this study (Additional file 1: Table S14). The study protocol was approved by the institutional review boards of Boston University Medical Center and Johns Hopkins Bloomberg School of Public Health. Written informed consent was obtained from all the study mothers.
Definition of maternal diabetes status in pregnancy
Diabetes in pregnancy included pregestational diabetes (type 1 and type 2 diabetes) and GDM, which was abstracted from electronic medical records (EMRs) and verified by blood glucose profiles extracted from EMRs. Pregestational diabetes was defined as fasting glucose concentration ≥ 7 mmol/L, 2 h plasma glucose of 75 g oral glucose tolerance test (OGTT) or random plasma glucose ≥ 11.1 mmol/L, HbA1c ≥ 6.5%, or receiving treatment for diabetes prior to pregnancy or in the first trimester [40]. GDM was diagnosed if women were free from pregestational diabetes and at least two of the following plasma glucose values were met: fasting glucose ≥ 5.3 mmol/l, 1 h ≥ 10.0 mmol/l, 2 h ≥ 8.6 mmol/l, and 3 h ≥ 7.8 mmol/l in response to a 100 g oral glucose load around 20–32 weeks of the index pregnancy according to American Diabetes Association criteria [40].
Definition of preterm birth
Gestational age was abstracted from EMRs. Preterm birth was defined as gestational age < 37 weeks and then further classified into two subtypes: spontaneous preterm birth (spontaneous preterm labor, preterm prelabor rupture of membranes, or cervical insufficiency) and medically indicated preterm birth (indicated by a specific maternal or fetal complication) according to the definition of the American College of Obstetricians and Gynecologists [41].
Maternal and neonatal characteristics
Maternal socio-demographic information was collected using a questionnaire interview. Educational attainment was classified into high school and below vs college and above; maternal smoking during pregnancy was classified into never smoker vs smoker; and maternal race and ethnicity were classified as non-Hispanic Black vs. non-Black (which included non-Hispanic White, Hispanic, Asian, Pacific Islander, and mixed race) based on self-reported race and ethnicity. Prepregnancy BMI was calculated as prepregnancy weight (kg) divided by height (m) squared. Maternal overweight and obesity were defined as BMI ≥ 25 kg/m2 [42]. Neonatal sex and birth weight were abstracted from EMRs. Fetal growth was assessed by birthweight for gestational age and categorized into 3 groups: small for gestational age (SGA, < 10th percentile), LGA (> 90th percentile), and appropriate for gestational age (AGA, 10th-90th percentile) according to an established local sex and race-specific reference population [43].
Genome-wide DNA methylation assessment in cord blood
Cord blood samples were collected at birth for genomic DNA extraction. DNA methylation profiling was analyzed using the Illumina Infinium MethylationEPIC BeadChip 850 K array platform (Illumina, California, USA) at the University of Minnesota Genomics Center, Minneapolis, MN, USA. Systematic quality control steps were reported in a previous publication [44]. In brief, profiling was conducted using existing analytic pipelines with R/Bioconductor package minfi [45]. At the sample level, we removed samples that appeared to be outliers (n = 1) or had a high missing rate (n = 1) or mismatch between data abstracted from EMRs and predicted sex (n = 7). At the probe level, we excluded probes with detection P values > 0.01 (n = 4,193), probes with known single nucleotide polymorphisms within the CpG motif or that have been shown to cross-hybridize (n = 140,271), probes with missing values (n = 44,900), and probes located on the X and Y chromosomes (n = 14,499). After these exclusions, a total of 661,996 CpG sites were interrogated in this study.
Statistical analysis
Socio-demographic and clinical variables are presented as mean ± SD for continuous variables and frequencies with proportions for categorical variables across maternal diabetes status in pregnancy. Raw methylation (beta) values were converted to M-values to better approximate a normal distribution.
Linear regression analysis was performed using the ‘limma’ package [46] to examine the association of maternal diabetes in pregnancy with each DNA CpG site in cord blood. Maternal diabetes in pregnancy was modeled as the exposure of interest, and cord blood DNA methylation levels (M-values) at each CpG site were modeled as the response variable, adjusted for maternal age, educational attainment, race and ethnicity, smoking during pregnancy, prepregnancy overweight or obesity, infant’s sex, fetal growth, and cell proportions (B cells, CD8 T cells, CD4 T cells, granulocytes, natural killer cells, monocytes, and nucleated red blood cells) which were estimated based on cord blood reference data [47] using ‘minfi’ package, as reported previously [44]. Genomic inflation was evaluated by constructing Q-Q plots and calculating λ for the epigenome-wide analyses. We corrected for multiple testing using the Benjamini–Hochberg FDR procedure. DMRs that were associated with maternal diabetes were identified using the Bioconductor package DMRcate [48] with the default settings using the same model and covariates as the abovementioned DMPs analysis.
A logistic regression model was applied to examine the association between maternal diabetes in pregnancy and preterm birth. To examine whether the identified DMPs mediated the association between diabetes and preterm birth, we performed mediation analysis for multiple mediators simultaneously using mma package [49], which allows for correlations among mediators. To evaluate the potential biological pathways in which diabetes-associated DMPs or DMRs were implicated, we performed probe-wise and region-based gene set enrichment analysis using GOmeth and GOregion functions in missMethyl R package [50], respectively. All statistical analyses were performed using the R statistical package, version 4.1.2 (R Foundation for Statistical Computing, Vienna).
Availability of data and materials
The datasets supporting these findings are not publicly available. The datasets used and/or analyzed for the current study are available from the corresponding author on reasonable request and after Institutional Review Board review and approval.
Abbreviations
- AGA:
-
Appropriate for gestational age
- BBC:
-
Boston Birth Cohort
- BMI:
-
Body mass index
- DE:
-
Direct effect
- DMP:
-
Differentially methylated position
- DMR:
-
Differentially methylated region
- EMR:
-
Electronic medical record
- GDM:
-
Gestational diabetes mellitus
- GO:
-
Gene Ontology
- IE:
-
Indirect effect
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- LGA:
-
Large for gestational age
- OGTT:
-
Oral glucose tolerance test
- RE:
-
Relative effect
- SGA:
-
Small for gestational age
- TE:
-
Total effect
References
Chen L, Magliano DJ, Zimmet PZ. The worldwide epidemiology of type 2 diabetes mellitus--present and future perspectives. Nat Rev Endocrinol. 2011;8:228–36.
Casagrande SS, Linder B, Cowie CC. Prevalence of gestational diabetes and subsequent Type 2 diabetes among U.S. women. Diabetes Res Clin Pract. 2018;141:200–8.
Lin SF, Kuo CF, Chiou MJ, Chang SH. Maternal and fetal outcomes of pregnant women with type 1 diabetes, a national population study. Oncotarget. 2017;8(46):80679–87.
Kong L, Nilsson IAK, Gissler M, Lavebratt C. Associations of maternal diabetes and body mass index with offspring birth weight and prematurity. JAMA Pediatr. 2019;173(4):371–8.
Group HSCR, Metzger BE, Lowe LP, Dyer AR, Trimble ER, Chaovarindr U, et al. Hyperglycemia and adverse pregnancy outcomes. N Engl J Med. 2008;358(19):1991–2002.
Berkowitz GS, Papiernik E. Epidemiology of preterm birth. Epidemiol Rev. 1993;15(2):414–43.
Hong X, Hao K, Ji H, Peng S, Sherwood B, Di Narzo A, et al. Genome-wide approach identifies a novel gene-maternal pre-pregnancy BMI interaction on preterm birth. Nat Commun. 2017;8:15608.
Tsai HJ, Liu X, Mestan K, Yu Y, Zhang S, Fang Y, et al. Maternal cigarette smoking, metabolic gene polymorphisms, and preterm delivery: new insights on GxE interactions and pathogenic pathways. Hum Genet. 2008;123(4):359–69.
Feinberg AP. Epigenetics at the epicenter of modern medicine. JAMA. 2008;299(11):1345–50.
Barker D. Fetal and infant origins of adult disease. London: BMJ Publishing; 1992.
Wang D, Liu X, Zhou Y, Xie H, Hong X, Tsai HJ, et al. Individual variation and longitudinal pattern of genome-wide DNA methylation from birth to the first two years of life. Epigenetics. 2012;7(6):594–605.
York TP, Latendresse SJ, Jackson-Cook C, Lapato DM, Moyer S, Wolen AR, et al. Replicated umbilical cord blood DNA methylation loci associated with gestational age at birth. Epigenetics. 2020;15(11):1243–58.
Antoun E, Kitaba NT, Titcombe P, Dalrymple KV, Garratt ES, Barton SJ, et al. Maternal dysglycaemia, changes in the infant’s epigenome modified with a diet and physical activity intervention in pregnancy: Secondary analysis of a randomised control trial. PLoS Med. 2020;17(11):e1003229.
Haertle L, El Hajj N, Dittrich M, Muller T, Nanda I, Lehnen H, et al. Epigenetic signatures of gestational diabetes mellitus on cord blood methylation. Clin Epigenet. 2017;9:28.
Finer S, Mathews C, Lowe R, Smart M, Hillman S, Foo L, et al. Maternal gestational diabetes is associated with genome-wide DNA methylation variation in placenta and cord blood of exposed offspring. Hum Mol Genet. 2015;24(11):3021–9.
Bennetts BH, Teutsch SM, Buhler MM, Heard RN, Stewart GJ. HLA-DMB gene and HLA-DRA promoter region polymorphisms in Australian multiple sclerosis patients. Hum Immunol. 1999;60(9):886–93.
Pinet V, Combe B, Avinens O, Caillat-Zucman S, Sany J, Clot J, et al. Polymorphism of the HLA-DMA and DMB genes in rheumatoid arthritis. Arthritis Rheum. 1997;40(5):854–8.
Siegmund T, Donner H, Braun J, Usadel KH, Badenhoop K. HLA-DMA and HLA-DMB alleles in German patients with type 1 diabetes mellitus. Tissue Antigens. 1999;54(3):291–4.
Yang IV, Zhang W, Davidson EJ, Fingerlin TE, Kechris K, Dabelea D. Epigenetic marks of in utero exposure to gestational diabetes and childhood adiposity outcomes: the EPOCH study. Diabet Med. 2018;35(5):612–20.
Ruchat SM, Houde AA, Voisin G, St-Pierre J, Perron P, Baillargeon JP, et al. Gestational diabetes mellitus epigenetically affects genes predominantly involved in metabolic diseases. Epigenetics. 2013;8(9):935–43.
Howe CG, Cox B, Fore R, Jungius J, Kvist T, Lent S, et al. Maternal gestational diabetes mellitus and newborn DNA Methylation: findings from the pregnancy and childhood epigenetics consortium. Diabetes Care. 2020;43(1):98–105.
Kang J, Lee CN, Li HY, Hsu KH, Lin SY. Genome-wide DNA methylation variation in maternal and cord blood of gestational diabetes population. Diabetes Res Clin Pract. 2017;132:127–36.
Rizzo HE, Escaname EN, Alana NB, Lavender E, Gelfond J, Fernandez R, et al. Maternal diabetes and obesity influence the fetal epigenome in a largely Hispanic population. Clin Epigenet. 2020;12(1):34.
Weng X, Liu F, Zhang H, Kan M, Wang T, Dong M, et al. Genome-wide DNA methylation profiling in infants born to gestational diabetes mellitus. Diabetes Res Clin Pract. 2018;142:10–8.
Awamleh Z, Butcher DT, Hanley A, Retnakaran R, Haertle L, Haaf T, et al. Exposure to gestational diabetes mellitus (GDM) alters DNA methylation in placenta and fetal cord blood. Diabetes Res Clin Pract. 2021;174:108690.
Morris SM Jr. Arginine metabolism revisited. J Nutr. 2016;146(12):2579S-S2586.
Ghosh A, Khandelwal N, Kumar A, Bera AK. Leucine-rich repeat-containing 8B protein is associated with the endoplasmic reticulum Ca(2+) leak in HEK293 cells. J Cell Sci. 2017;130(22):3818–28.
Gasparoni G, Bultmann S, Lutsik P, Kraus TFJ, Sordon S, Vlcek J, et al. DNA methylation analysis on purified neurons and glia dissects age and Alzheimer’s disease-specific changes in the human cortex. Epigenet Chromatin. 2018;11(1):41.
Li M, Fallin MD, Riley A, Landa R, Walker SO, Silverstein M, et al. The association of maternal obesity and diabetes with autism and other developmental disabilities. Pediatrics. 2016;137(2):e20152206.
Nahum Sacks K, Friger M, Shoham-Vardi I, Abokaf H, Spiegel E, Sergienko R, et al. Prenatal exposure to gestational diabetes mellitus as an independent risk factor for long-term neuropsychiatric morbidity of the offspring. Am J Obstet Gynecol. 2016;215(3):380 e1-7.
GwnwCards: ARID5B Gene - AT-Rich Interaction Domain 5B. https://www.genecards.org/cgi-bin/carddisp.pl?gene=ARID5B Accessed 9 Oct 2022
Yang H, Na FY, Guo L, Liang X, Zhang RF. The landscape of DNA methylation in asthma: a data mining and validation. Bioengineered. 2021;12(2):10063–72.
Binder AM, LaRocca J, Lesseur C, Marsit CJ, Michels KB. Epigenome-wide and transcriptome-wide analyses reveal gestational diabetes is associated with alterations in the human leukocyte antigen complex. Clin Epigenet. 2015;7(1):79.
Pinney SE, Joshi A, Yin V, Min SW, Rashid C, Condon DE, et al. Exposure to gestational diabetes enriches immune-related pathways in the transcriptome and methylome of human amniocytes. J Clin Endocrinol Metab. 2020;105(10):3250–64.
Pantham P, Aye IL, Powell TL. Inflammation in maternal obesity and gestational diabetes mellitus. Placenta. 2015;36(7):709–15.
Nasreen S, Wilk P, Mullowney T, Karp I. The effect of gestational diabetes mellitus on the risk of asthma in offspring. Ann Epidemiol. 2021;57:7–13.
Hedderson MM, Ferrara A, Sacks DA. Gestational diabetes mellitus and lesser degrees of pregnancy hyperglycemia: association with increased risk of spontaneous preterm birth. Obstet Gynecol. 2003;102(4):850–6.
Romero R, Espinoza J, Goncalves LF, Kusanovic JP, Friel LA, Nien JK. Inflammation in preterm and term labour and delivery. Semin Fetal Neonatal Med. 2006;11(5):317–26.
Wang G, Divall S, Radovick S, Paige D, Ning Y, Chen Z, et al. Preterm birth and random plasma insulin levels at birth and in early childhood. JAMA. 2014;311(6):587–96.
American Diabetes Association. 2 classification and diagnosis of diabetes: standards of medical care in diabetes-2020. Diabetes Care. 2020;43:S14–31.
American College of O, Gynecologists’ Committee on Practice B-O. Prediction and prevention of spontaneous preterm birth: ACOG practice bulletin, Number 234. Obstet Gynecol. 2021;138(2):e65–90.
Centers for Disease Control and Prevention. Overweight & Obesity, Defining Adult Overweight and Obesity. 2020. https://www.cdc.gov/obesity/adult/defining.html. Accessed 23 Nov 2020.
Wang L, Wang X, Laird N, Zuckerman B, Stubblefield P, Xu X. Polymorphism in maternal LRP8 gene is associated with fetal growth. Am J Hum Genet. 2006;78(5):770–7.
Xu R, Hong X, Zhang B, Huang W, Hou W, Wang G, et al. DNA methylation mediates the effect of maternal smoking on offspring birthweight: a birth cohort study of multi-ethnic US mother-newborn pairs. Clin Epigenet. 2021;13(1):47.
Hansen K, Ayree M, Irizarry RA, Jaffe A, Maksimovic J, Houseman A, et al. Analyze Illumina Infinium DNA methylation arrays. Package ‘minfi’ version 1.38.0. https://github.com/hansenlab/minfi.
Smyth G, Hu Y, Ritchie M, Silver j, Wettenhall j, McCarthy d, et al. Linear Models for Microarray Data. http://bioinf.wehi.edu.au/limma.
Bakulski KM, Feinberg JI, Andrews SV, Yang J, Brown S, McKenney S, et al. DNA methylation of cord blood cell types: applications for mixed cell birth studies. Epigenetics. 2016;11(5):354–62.
Peters TJ, Buckley MJ, Statham AL, Pidsley R, Samaras K, V Lord R, et al. De novo identification of differentially methylated regions in the human genome. Epigenet Chromatin. 2015;8:6.
Yu Q, Scribner RA, Leonardi C, Zhang L, Park C, Chen L, et al. Exploring racial disparity in obesity: a mediation analysis considering geo-coded environmental factors. Spat Spatiotemporal Epidemiol. 2017;21:13–23.
Phipson B MJ, missMethyl: Analysing Illumina HumanMethylation BeadChip Data. https://www.bioconductor.org/packages/devel/bioc/vignettes/missMethyl/inst/doc/missMethyl.html.
Acknowledgements
We would like to thank the study participants, the nursing staff at Labor and Delivery of the Boston Medical Center and the field team for their contributions to the Boston Birth Cohort. Linda Rosen, MSEE, and the Clinical Data Warehouse assisted in obtaining relevant clinical information; she was compensated for her time. The Clinical Data Warehouse service is supported by Boston University’s Clinical and Translational Institute and the National Institutes of Health Clinical and Translational Science Award (grant U54-TR001012).
Funding
The Boston Birth Cohort (the parent study) is supported in part by the National Institutes of Health (NIH) (2R01HD041702, R01HD086013, R01HD098232, R01ES031272, R01ES031521, and U01ES034983); and the Health Resources and Services Administration (HRSA) of the U.S. Department of Health and Human Services (HHS) (UT7MC45949). The contents of the manuscript are solely the responsibility of the authors and do not necessarily represent the official views of the supporting agencies.
Author information
Authors and Affiliations
Contributions
GW was responsible for the study concept and design, acquisition and interpretation of data, statistical analysis, and drafting of the manuscript. TRB, CP, LL, and XW were responsible for the critical revision of the manuscript. RX, BZ and XH were responsible for statistical analysis and critical revision of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study has been approved by the Institutional Review Boards of Boston Medical Center and Johns Hopkins Bloomberg School of Public Health. Written informed consent was obtained from each participant.
Consent for publication
Not applicable.
Competing interests
The authors declare no potential conflicts of interest with respect to the research, authorship, and publication of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
. Supplementary figures and tables.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, G., Xu, R., Zhang, B. et al. Impact of intrauterine exposure to maternal diabetes on preterm birth: fetal DNA methylation alteration is an important mediator. Clin Epigenet 15, 59 (2023). https://doi.org/10.1186/s13148-023-01473-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13148-023-01473-1