
Abstract
Background
Amelioration of biofuel feedstock of microalgae using sustainable means through synthetic ecology is a promising strategy. The co-cultivation model (Tetraselmis striata and Pelagibaca bermudensis) was evaluated for the robust biofuel production under varying stressors as well as with the selected two-stage cultivation modes. In addition, the role of metabolic exudates including the quorum-sensing precursors was assessed.
Results
The co-cultivation model innovated in this study supported the biomass production of T. striata in a saline/marine medium at a broad range of pH, salinity, and temperature/light conditions, as well as nutrient limitation with a growth promotion of 1.2–3.6-fold. Hence, this developed model could contribute to abiotic stress mitigation of T. striata. The quorum-sensing precursor dynamics of the growth promoting bacteria P. bermudensis exhibited unique pattern under varying stressors as revealed through targeted metabolomics (using liquid chromatography–mass spectrometry, LC–MS). P. bermudensis and its metabolic exudates mutually promoted the growth of T. striata, which elevated the lipid productivity. Interestingly, hydroxy alkyl quinolones independently showed growth inhibition of T. striata on elevated concentration. Among two-stage cultivation modes (low pH, elevated salinity, and nitrate limitation), specifically, nitrate limitation induced a 1.5 times higher lipid content (30–31%) than control in both axenic and co-cultivated conditions.
Conclusion
Pelagibaca bermudensis is established as a potential growth promoting native phycospheric bacteria for robust biomass generation of T. striata in varying environment, and two-stage cultivation using nitrate limitation strategically maximized the biofuel precursors for both axenic and co-cultivation conditions (T and T-PB, respectively). Optimum metabolic exudate of P. bermudensis which act as a growth substrate to T. striata surpasses the antagonistic effect of excessive hydroxy alkyl quinolones [HHQ, 4-hydroxy-2-alkylquinolines and PQS (pseudomonas quorum signal), 2-heptyl-3-hydroxy-4(1H)-quinolone].
Similar content being viewed by others

Background
Microalgae are an emerging potential source of sustainable biofuel in addition to efficient carbon sequestration and bioremediation [1,2,3,4]. Innovative marine microalgal cultivation has been suggested for biofuel production because of its extremely low freshwater and cultivable land crop footprint [5]. The cost-effective cultivation and determination of optimum environmental conditions for achieving the desired improvement in biomass and biofuel precursors is an essential step for the technological advancement [6, 7]. Most of the research in microalgal technologies has focused on elevation and amelioration of desired biological yields of the axenic or monoalgal strains without considering the potential of phycospheric bacteria [3, 4, 8, 9].
However, limited research has been conducted on growth promoting sustainable cultivation techniques to ascertain the biomass productivity through biotic interactions despite the enormous potential of synthetic/natural co-cultivation. The metabolite and nutrients exuded by the phycospheric growth promoting bacteria could offer impressive biomass yields with suitable fatty acid properties to make biodiesel [10,11,12]. Furthermore, the mutualistic phycospheric bacteria can grow in similar physicochemical environments whenever co-cultivated, while it could not grow alone in the same media, and remarkable growth co-operation for the nutrients was shown which could reduce the cost of the cultivation [12].
Most of the co-cultured mutual consortia may release volatile or non-volatile growth promoting metabolites (such as vitamins, hormones, amino acids, and fatty acids), macronutrients (K, N, and P), and micronutrients such as trace metals (including B, Mn, Ni, Cu, Fe, and Co), and these metabolites (including infochemicals) support to the respective partner during the growth [13,14,15,16,17]. Some bacteria exhibit potential for environmental applications in wastewater remediation and biofuel production and a consortium (natural or synthetically engineered) could promote the growth of desired microalgae as well as exhibit algicidal effects on undesired microalgae [12, 14, 16, 17].
Some of the different microalgal seawater strains are accorded a relatively higher significance for open/semi-enclosed cultivation in coastal waters. Tetraselmis is well known for its commercial potential for biomass and biofuel production in coastal areas [18, 19]. Tetraselmis spp. are highly influenced by the associated bacteria; there is an opportunity to boost the biomass productivity via synthetic phycosphere engineering upon establishing the mechanism and optimal processes [12, 20,21,22]. The previous study on Tetraselmis striata and its native phycospheric bacterium Pelagibaca bermudensis under axenic and co-cultivation conditions (T and T-PB, respectively) elucidated the mutual co-operative effects of the phosphate and organic/inorganic carbons. However, the study was conducted under sole unvarying light, temperature, and salinity conditions [12]. There is a growing consensus on the importance of understanding the growth promoting effects caused by bacteria under various environmental stressors. Furthermore, determining whether P. bermudensis has growth promoting effects on microalgae under varying abiotic conditions is imperative, because the coastal aquatic environment and effluent load are extremely variable where it needs to be applicable.
Pelagibaca bermudensis belongs to the well-known Roseobacter clade of the Alphaproteobacteria family, which is one of the predominant seawater bacterial communities and is also found in a wide range of habitats [23, 24]. Some members of this family have known quorum-sensing abilities for biofilm formation on biotic/abiotic surfaces and can use a variety of organic carbon sources, which they degrade effectively [25,26,27]. Roseobacter P. bermudensis may degrade aliphatic and aromatic hydrocarbons due to its hydrocarbonoclastic properties [28]. P. bermudensis HTCC2601 isolated from surface water has genes that encode all C3 cycle enzymes [29]. It is also established as a d-carbamoylase-producing bacterium and synthesizes antimicrobial agents such as tropodithietic acid [23, 30]. The marine bacterium P. bermudensis encodes an uncharacterized member of the phosphopyruvate hydratase (enolase) superfamily [31]. It predominantly has fatty acid 18 : 1ω7c (79.7 % of total fatty acids) [24].
Quorum sensing is the ability of the organisms to regulate gene expression of the varied function using the precursors and to act as a multicellular system for collective decision making which is often correlated to their cell populations [27, 32]. Most members of the Roseobacter clade also contain the RuBISCO enzyme, which is involved in CO2 fixation and has an oligophototrophic/chemoheterotrophic/photoheterotrophic nature [33, 34].
The growth dynamics of P. bermudensis in the presence of T. striata as a mutual partner, as well as the extent of this mutual partnership and role of quorum-sensing precursors (acyl homoserine lactones and alkyl quinolones) concentrations in this consortium are currently unexplored. The role of acyl homoserine lactones and alkyl quinolones in the intrapopulation and interpopulation communication during co-cultivation may be a significant factor for the possible relationships, colonization, quorum sensing, biofilm formation, cellular recognition, and cell density-dependent regulation of target genes as well as in growth dynamics [32, 35, 36]. The primary objective of this research was to study the mutual growth, biomass, and lipid productivity of the T. striata under co-cultivated (T-PB) and axenic (T) conditions in varying environmental stressors; second, to monitor the quorum-sensing precursors of P. bermudensis during co-cultivation, and their effect on growth of the T. striata; furthermore, to assess the amelioration of the biomass and lipid of the T. striata for feasible biofuel production under varying crude exo-metabolic feeding and with selected two-stage cultivation modes.
Methods
Culture development and maintenance
P. bermudensis
The P. bermudensis (KCTC13073BP) used in study was isolated from the phycosphere of T. striata. P. bermudensis maintained and cultured in the axenic form in cell culture flasks (containing marine broth) and marine nutrient agar plates. Periodical maintenance and axenic culture monitoring was performed. The details of the isolation of P. bermudensis from phycosphere of Tetraselmis, axenic (T) culture preparation, and molecular identification of the strain used in this study are available elsewhere [12].
T. striata
Tetraselmis striata was isolated from seawater samples from the Incheon coast mass cultivation site of Inha University. Culture of T. striata was deposited at the Korean Culture Repository after molecular identification by Inha Univeristy and was assigned Accession No. KCTC1432BP. The microalgal culture was also maintained in O3 medium in the algal culture laboratory of the Department of Life Science, Hanyang University. The O3 media was containing: 20 g L−1 of NaCI, 5.0 g L−1 of MgSO4·7H2O, 4.25 g L−1 of MgCl2·6H2O, 1.13 g L−1 of CaCl2·2H2O, 0.76 g L−1 of KNO3, 0.05 g L−1 of KH2PO4, 0.03 g L−1 of NaHCO3, and 0.76 mL of stock solution A (18.6 g L−1 of Na2EDTA and 2.4 g L−1 of FeCl3·6H2O L−1 dissolved in distilled H2O) and 0.76 mL of stock solution B [40 mg L−1 of ZnCl2, 600 mg L−1 of H3BO3, 15 mg L−1 of CoCl2·6H2O, 40 mg L−1 of CuCl2·2H2O, 488 mg L−1 of MgCl2·4H2O, and 37 mg L−1 of (NH4)6MoO24·4H2O dissolved in distilled H2O and 0.38 mL of vitamin B12 stock solution (0.1 mg of vitamin B12 in 100 mL of distilled water). 1 M Tris–HCl and 1 M NaOH were used for adjusting the pH [37]. The modification of the media according to the experiments (salinity, pH, and nutrient limitation) is mentioned in the separate section.
The axenic T. striata culture cells (T) were maintained in cell culture flasks containing O3 media (pH 8.0) incubated at 20 °C with exposure of 50 µM m−2 s−1 by cool fluorescent lamps (12:12 h photoperiod). Axenic culture and experimental culture were routinely maintained by repetitive agar plating techniques and microscopic observations.
Furthermore, prior to initiate the experiment, axenic status of the culture was also monitored using 4′,6-diamidino-2-phenylindole (DAPI, Sigma-Aldrich, St. Louis, MO, USA) staining for bacterial presence under a microscope (BX51 Olympus, Japan). The inoculum volume to attain the final cell abundance of T. striata and P. bermudensis in experimental culture flask on the initial hours of the experiment was determined by actual cell abundance of mother culture. Cell abundance was measured according to the “Cell abundance of microalgae and bacteria, and growth promotion” section. The cells of the determined inoculum volume were harvested using the centrifugation; complying with standard sterile conditions and resulting axenic washed pellet were re-suspended in axenic and co-cultivated conditions.
T-PB and T cultivation experiments under varying stress conditions
To investigate the effect of pH, the O3 media were initially adjusted for the different pH of 4, 6, 8, and 10 followed by the autoclave. After centrifuging the T. striata and P. bermudensis culture and repeatedly washing the pellet, the starved cells were inoculated in 100-mL O3 medium culture flask (150 mL) under sterile laminar flow conditions. T. striata and P. bermudensis were co-inoculated and adjusted to the initial cell densities up to 104 and 1.72 × 108 cells mL−1, respectively, for each experiment of co-cultivation (T-PB). The control culture under T conditions was only inoculated with axenic T. striata. All the conditions were investigated in triplicates.
Similarly, the O3 media containing different salinity with sodium chloride (NaCl) at 20, 25, 30, and 35 gL−1 and an initial pH of 8.0 were used for both the T and T-PB experiments. To examine the effect of varying light and temperature on T. striata under T-PB and T conditions, low-light (30 µM m−2 s−1 with 20 °C, 12:12 h), and low-temperature (15 °C with 45 µM m−2 s−1, 12:12 h) conditions were compared with comparatively higher light (147 µM m−2 s−1, 20 °C, 24:00 and 12:12 h.) and normal light and temperature (45 µM m−2 s−1, 20 °C, 12:12 h) conditions (Table 1). All the experiments related to light/temp were run in triplicates using the O3 media at an initial pH of 8.0 and salinity of 20 g L−1.
Cell abundance of microalgae and bacteria, and growth promotion
To determine the cell abundance (cells mL−1), the cells were counted every third day. Samples (100 µL) from each experimental flask were ejected out under sterile laminar flow conditions for the T. striata and P. bermudensis, and immediately fixed with Lugol’s solution (final concentration 1%) and glutaraldehyde (2.5% glutaraldehyde), respectively. The microalgae growth was determined by enumerating the cells (fixed with 1% Lugol’s solution) observed using a hemocytometer under a light microscope at 200× magnification (BX51 Olympus, Japan). Similarly, the bacterial samples (fixed with glutaraldehyde) were counted using the DAPI epifluorescence technique. Before microscopy and enumeration of bacteria, each sample was filtered through a 0.2-μm GTTP Millipore filter membrane (Millipore Filter Corporation, Ireland) under 178 mmHg and then stained with 4′,6-diamidino-2-phenylindole (DAPI) for 8–12 min under dark conditions. The filter membrane containing fixed and stained bacteria was taken on the glass slide and covered by cover slip containing Fluor-mount (Sigma, USA). The slides were kept in the cool and dark place until analysis. During the observation under microscope, UV transparent immersion oil was used. The cell abundance of P. bermudensis was then enumerated on the basis of cell counts at 1000× under an epifluorescence microscope (Olympus, Japan equipped with wavelength filter set) from a microscope slide (Marienfeld, Germany) as per Eq. 1. The cell abundance was analyzed in ten randomly selected fields of view across each of the slides based on the method of Joo et al. [38]:
The growth promotion (fold) was determined as the ratio of the cell abundance of co-cultivated T. striata (T-PB) and axenic T. striata (T) in the stationary phase of the same cultivation time.
Biomass productivity and lipid content measurement
A known amount of the experimental culture was harvested, while the T. striata cells reached to stationary phase (day 18). The cultured cells were harvested by centrifugation (3700×g for 10 min), the cell pellet was washed twice with distilled water (DW) at the same centrifugal force, and then, the wet biomass was dried at 70 °C in a drying oven for 24 h. The biomass productivity of both the axenic T. striata (T) and co-cultivated T. striata and P. bermudensis (T-PB) was determined based on the cell dry weight using a gravimetric method [8] by the following equation:
The total lipids were extracted from a known weight of dried biomass using a solvent mixture of chloroform and methanol (2:1, v/v) according to the Folch method [39] and were finally determined using the following equation:
The total chlorophyll concentration error in folch extracted lipid fraction was 0.031–0.042% of dry weight of biomass, which was less than the error of the total lipid (%) found in the experimental triplicates.
Determination of Acyl homoserine lactones and alkyl quinolones concentration in cultures
The cultures grown for different days under the varying physicochemical conditions described in “T-PB and T cultivation experiments under varying stress conditions” section were harvested by centrifugation at 3700×g for 10 min; the supernatant was frozen immediately and then transferred to a − 80 °C deep freezer till the analysis. The supernatants were thawed and filtered through a 0.22-µm sterile Whatman filter paper.
An equal amount of sodium acetate was used to extract the metabolites. After ensuring maximum metabolite extraction by vortex and mixing, the extracted layers were separated and then freeze-dried under vacuum conditions. The dried extracts were treated with acidic methanol for derivatization. Acyl homoserine lactone (AHL) and alkyl quinolone (AQ) standards, namely N-(3-oxo decanoyl)-l-homoserine lactone, N-[-(RS)-3-hydroxybutyryl] l-homoserine lactone, N-butyryl-dl-homoserine lactone, 4-hydroxy-2-heptylquinoline (HHQ), and 2 heptyl-3 hydroxy-4 (1H) quinolone (PQS) (Sigma-Aldrich), were used as the standards. The derivatized products from the samples were analyzed using an ultra-performance-liquid chromatography (UPLC) system (Ultimate 3000 RS UHPLC, Thermo Fisher Scientific) coupled with mass spectrometry (MS, Quantiva, Thermo Fisher Scientific, USA).
The analysis was carried out in the positive polarity mode using an electrospray ionization (ESI) source (3500 V). The ion transfer tube and vaporizer temperatures were 342 and 350 °C, respectively. The analytes were separated using a Thermo Hypersil gold C18 column (2.1 × 100 mm, 1.9 µm) at 35 °C with a 0.4 mL/min flow rate. The mobile phase consisted of 0.1% formic acid in water (A) and 0.1% formic acid in acetonitrile (B). The gradient used was 10% B in A for 1.0 min, 50% B in A for 1.5 min, up to 99% B in A over 5.5 min and 7.5 min, and finally, the process was held at 90% B in A for 3 min. The standards of the AHL (acyl homoserine lactones) and AHQ (alkyl hydroxy quinolones) were run at different serially diluted concentrations, and a standard curve was constructed. Injection volume for each sample was 2 µL. Blanks were also similarly run for the quality control. The peaks were analyzed using relevant software and the concentrations were determined for the respective analyte molecules [40]
Effects of crude metabolic exudates, HHQ, and PQS on microalgal cells
The P. bermudensis cultures were harvested after 2.5 days of the growth incubation in the marine broth media. The control (marine broth) without any bacterial inoculation was also incubated for the same time. The supernatants of the P. bermudensis culture (t) and media alone (MB) were collected, filtered through a 0.22-µm sterile syringe filter, and added to autoclaved samples of the O3 media in cell culture flasks under sterile conditions. The ratio of culture filtrates (MB with the P. bermudensis exudates) to the final experimental O3 media volume was 0:100 (0% t, control), 1:99 (1% t), 5:95 (5% t), 10:90 (10% t), and 100:00 (100% t). A similar ratio was used for the control (MB, the supernatant of the MB of the same incubation time) instead of using the P. bermudensis culture supernatant. The microalgal cells at the late log phase were inoculated to adjust the initial cell density to 104 cells mL−1 in the cell culture flask on day 0 of the experiment. All the experiments were performed in triplicates (n = 3). Similarly, the cell abundance, biomass productivity, and total lipid content were assessed using the methods described in this section.
To investigate an independent effect of HHQ and PQS on T. striata, HHQ and PQS were dissolved in acetonitrile and added in different concentration in the O3 media, and initial cell density of the T. striata (104 cells mL−1) was adjusted by inoculating the starved cells and growth was observed during the experiment.
Investigation of nutrient limitation conditions
T. striata was grown under T and T-PB mode under limited nitrate, sulfate, and trace metal (iron and other trace metals) levels in the O3 medium and compared with the nutrient replete condition (O3 control). The nitrate limited conditions were devoid of nitrate in the O3 media, since KNO3 was not added. However, nitrogen in the form of ammonia was present as it was added as constituent of stock A of O3 media. The trace metal-limited conditions were devoid of Iron and stock B. The sulfate-limited media were containing ~ 4–5 µM of sulfate after washing the cell pellet with sterile distilled water. However, the sulfate in the media was not added as a component of O3 for the sulfate-limited media. The inoculation conditions and measurement of cell abundance, lipid content, and biomass productivity were carried out using similar methods to those of the other experiments.
Comparative two-stage cultivation
The two-stage cultivation mode including nutrient replete and nutrient limiting were employed for both T and T-PB. The microalgal cultures containing T. striata (axenic, T) and T. striata with the P. bermudensis (co-cultivated, T-PB) were cultivated in the O3 medium for 15 days (n = 12) under NLT conditions, as shown in Table 1. All experimental cultures were harvested using centrifugation and the cell pellet was washed with distilled water (DW). The starved cell pellets were re-suspended in fresh O3 medium of three different conditions for the second-stage cultivation of 3 days. Herein, three-distinct two-stage cultivation performed for elevated salinity (containing modified O3 with 35 g L−1 NaCl; n = 3), nitrate limitation (modified O3 without nitrate; n = 3), and low pH (O3 media with pH 6.0; n = 3) along with a parallel control (20 g L−1 NaCl, pH 8.0 and nutrient replete condition) for both T and T-PB conditions. After 3 days, cultures were harvested for the biomass productivity and total lipid content assessment.
Statistical analysis
All experiments were run in triplicates (except quorum-sensing precursors monitoring, n = 2), while the error bars are represented as mean ± SD. Data were analyzed using the one-way analysis of variance (ANOVA) followed by post hoc test to determine the significance of differences (LSD) at p ≤ 0.05 between the treatments using SPSS.
Results and discussion
Effect of varying pH
The total cell abundance of the microalgae under the T-PB condition with P. bermudensis and T conditions at different growth interval was the highest at pH 8.0 followed by pH 10.0, 6.0, and 4.0. T. striata showed a higher cell abundance in the presence of P. bermudensis than in its absence. The optimum pH conditions for the biomass productivity in both T and T-PB conditions were pH 8.0 and 10.0 (Figs. 1a and 2a). It was interesting to note that although the cell division rate of T. striata was restricted at pH 4.0 and 6.0, growth promotion was observed (Fig. 1a). This clearly indicates that the P. bermudensis provided a growth promoting environment for the T. striata under almost all the various pH conditions. The growth promotion rates (expressed as fold increases) on the cell harvesting day were 2.8-, 1.3-, 1.2-, and 1.3-fold at pH 4.0, 6.0, 8.0, and 10.0, respectively.

Cell abundance of Tetraselmis striata under axenic (T) and co-culture (T-PB) conditions at different (a) pH, b salinity (g L−1 NaCl), and c light/temperature in O3 medium. The values are presented in mean ± SD (n = 3). Error bars are showing SD. Details of the condition are exhibited in Table 1

Biomass productivity and total lipid (%) of Tetraselmis striata under axenic (T) and co-cultivated (T-PB) conditions at different a pH, b salinity (g L−1 NaCl), and c light/temperature in O3 medium. The values are presented in mean ± SD (n = 3). Error bars are showing SD. Post hoc analysis is shown in the Additional file 2: Table S1 (A–F) for comparing the means according to their significant difference (LSD)
The maximum mean cell abundance on the harvesting day was obtained at the pH 8.0 and 10.0 (1.83 × 106 and 1.31 × 106 cells mL−1, respectively) under T-PB conditions (Fig. 1a). The cell density of P. bermudensis showed a sudden decrease at the initial T-PB phase but later improved under the optimal and suboptimal pH conditions (Additional file 1: Figure S1). Although both the bacterial and microalgal growth dynamics were affected by the H+ concentrations, the P. bermudensis remarkably survived under the acidic pH (4.0) and a multiple-fold enhancement of the growth in T. striata (6.41- and 2.81-fold on days 15 and 18, respectively; Fig. 1a and Additional file 1: Figure S1) was observed.
Furthermore, the inoculated cell density of P. bermudensis (108 cells) was higher than that used previously (106 cells mL−1) by Park et al. [12]. The higher cell density of P. bermudensis required higher levels of microalgal metabolites to support the growth in the initial phase and to initiate the mutualistic interactions. The lipid content of the biomass obtained under T conditions was approximately 1–3% higher than the that of the T-PB conditions at pH 8 and 10; however, the mean biomass productivity of T-PB increased by up to 19.30 to 22.67% at the same pH. The highest total lipid content was 23.8 ± 0.21% at pH 6.0, while the highest biomass productivity was 43.52 ± 3.2 mg L−1 day−1 at pH 8.0 under T-PB conditions (Fig. 2a). The overall results exhibited that the T-PB conditions induced higher lipid productivity than the T condition at each investigated pH.
Effect of varying salinity
The maximum cell abundance of the T. striata was observed in 20 g L−1 NaCl on day 15/18 under T-PB conditions. There was a decline in the cell abundance at concentrations of 25, 30, and 35 g L−1 NaCl (Fig. 1b). The results suggest that the cells could cope with the wide range of salinity levels (20–35 g L−1). The elevated salinity levels in the T condition could lead to impeded microalgal cell abundance and lower growth than that observed under the respective T-PB conditions. The growth promotion was 1.18-, 2.51-, 2.53-, and 3.58-fold in 20, 25, 30, and 35 g L−1 NaCl, respectively, for the microalgae on day 18. T. striata (under T-PB) grew in 20 g L−1 NaCl with a higher growth promotion rate during the earlier lag phase (1.97- and 1.89-fold on days 3 and 6, respectively).
From day 6 onwards, T. striata (under T conditions) cultured with elevated salinity did not grow effectively compare to cells grown under T-PB (Fig. 1b). The presence of P. bermudensis in the cultivation medium of the T. striata not only enhanced the microalgae growth but also showed a pronounced multifold growth promotion. This clearly indicates that P. bermudensis likely exuded some specific metabolites or nutrients under higher salinity levels, which mitigated the intracellular stress levels of the microalgal cells. The biomass productivity of the microalgae (under T conditions) was 36.48 ± 1.39, 15.90 ± 1.12, 12.35 ± 1.20, and 9.31 ± 1.81 mg L−1 day−1 with lipid contents of 22.27 ± 0.38, 23.1 ± 0.80, 23.81 ± 1.0, 21 ± 0.7% at 20, 25, 30, and 35 g L−1 NaCl, respectively. While the biomass productivities of the T-PB were 43.52 ± 3.2, 26.57 ± 1.56, 23.47 ± 0.52, 20.47 ± 2.48 mg L−1 day−1 with the lipid contents of 21.23 ± 0.56, 23.48 ± 0.30, 19.67 ± 0.52, 18.63 ± 0.82% at 20, 25, 30, and 35 g L−1 NaCl, respectively (Fig. 2b).
These results suggest that the salinity endurance and wide salinity tolerance of T. striata (T-PB) enhance its ability to produce consistent biomass. Furthermore, this effect could only be due to the phycospheric native growth promoting bacteria, since T. striata alone (T) did not exhibit a consistent biomass productivity under the wide salinity range used in this study. The growth dynamics of P. bermudensis was also found to be unique in each case owing to the varied salinity (Additional file 1: Figure S2). In fact, the cells of P. bermudensis that are lysed may also provide metabolic nutrients to the microalgae. Many of these metabolites are osmolytes and phytohormones, which potentially regulate the membrane integrity of the microalgal cells as well as alter or mitigate the intracellular stress levels [41, 42].
Effect of varying light and temperature
Tetraselmis striata did not exhibit better growth under low-light conditions than it did under high-light intensity in the presence or absence of P. bermudensis. The continuous exposure of T. striata to high-light conditions increased its growth by several degrees under both conditions compared to that under the other experimental settings. The growth promotion (1.85-fold) was also pronounced in the presence of P. bermudensis under high-light intensity with a maximum cell abundance (4.6 × 106 cells mL−1). The light intensity and photoperiods were highly dominating factors to affect the T. striata and, growth promotion enhanced under high-light intensity as well as photoperiod (Fig. 1c and Additional file 1: Figure S3). Tetraselmis can grow at a relatively wide temperature range with a low rate of variation, so the effect of temperature on the cell abundance of T. striata were minimum [43]. Irrespective of the temperature/light conditions, T-PB conditions remained superior to T conditions with a 1.26–1.96-fold growth promotion in the microalgal cell abundance on day 18 (Fig. 1c and Additional file 1: Figure S3).
Unlike the oscillated bacterial growth dynamics observed under the varying temperature, salinity, and pH conditions, the P. bermudensis showed consistent growth when continuously exposed to high-light intensity. Furthermore, a high cell abundance (3.189 × 109 cells mL−1 on day 18) was observed during the experimental period, which might have been due to luxuriant microalgal growth and continuous photosynthesis (Additional file 1: Figure S4). The continuous high-light exposure induced a maximum biomass productivity of 79.85 and 56.00 mg L−1 day−1 with 18.38 and 20.29% lipid content under T-PB and T conditions, respectively. In contrast, the values were the lowest under low-light conditions (24.89 and 18.51 mg L−1 day−1 with 19.57 and 22.46% total lipids under T-PB and T conditions, respectively) under varying light/temperature conditions. Low-temperature conditions induced maximum lipid contents of 25.67 and 28.26% under T-PB and T conditions, respectively (Fig. 2c).
Dynamics of quorum-sensing precursors under T-PB condition
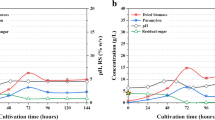
The 2-heptyl-4-quinolone (HHQ) concentration in the lag phase was similar (2.02–2.14 µM) for all of the varying experimental T-PB conditions (Fig. 3). The accumulated HHQ was exuded under most conditions because of the decline in cell growth of P. bermudensis during the initial phase to the log phase except the high-light exposure (Fig. 3 and Additional file 1: Figure S4). The high growth rate of T. striata (high cell abundance) could have produced a high organic carbon content in the form of metabolites, which likely led to the growth of P. bermudensis and improved cell density in the T-PB condition. The HHQ was higher at the log or stationary phase of the microalgae growth cycle, especially under the optimum conditions of biomass and lipid, at a range of 2.28–17.19 µM (Fig. 3). The highest HHQ (17.19 µM) was observed with the continuous exposure to high-light intensities where the P. bermudensis constantly grow without any decline.

2-Heptyl-3-hydroxy-4(1H)-quinolone (PQS) and 2-heptyl-4-quinolone (HHQ) precursor concentrations under varying pH, salinity, light, and temperature in co-culture (T-PB) conditions at lag phase (day 3), log phase (day 9), and stationary phase (day 15). The values are presented in mean ± SD (n = 2). Error bars are showing SD. HHQ and PQS were below detectable range at 00 h of the experiment
However, this observation was not correlated with the lag, log, and stationary phases of the P. bermudensis under different experimental conditions, since the cell abundance measured every third day, which is a long growth cycle for the P. bermudensis. This phenomenon could not be investigated with axenic P. bermudensis (without T. striata) in O3 media, since P. bermudensis alone did not grow without the T. striata metabolites (data not shown). However, there was an oscillation in the cell density of P. bermudensis (day 3–18, Additional file 1: Figure S4). The growth cycle of the P. bermudensis remained at a low range of the growth intervals.
Apart from the quorum-sensing function for the bacterial biofilm formation, HHQ is also known to exhibit algicidal activity. Previously, Roseobacter clade have been reported to show algicidal activity against Emiliania huxleyi strains with IC50 of 88–115 ng m L−1. In contrast, marine chlorophytes were not affected with elevated HHQ (IC50 = 105ng mL−1) [44]. The HHQ and PQS concentration mediate the mutualistic interactions by indirect promotion of the algal growth through increasing cell abundance of P. bermudensis to form biofilm on the biotic surface and releases concentrated metabolic stock for their algal partner. However, while HHQ and PQS reached higher concentration, they can arrest the growth of T. striata. This example could be similar as a role of other algicidal metabolite roseobacticide. Although Roseobacter have symbiotic association for the phytoplanktons under mutualistic conditions, they switch to parasitic relationship by sensing the senescence signal (p-coumaric acid) of their host algae and releases roseobacticides to control the growth of algae [45].
In the current study, the release of higher HHQ concentrations in the log/stationary phase could not be detrimental for the T. striata and did not likely hamper the growth promoting effect on the algal cells under T-PB conditions (Fig. 3). In fact, the dead cells could be degraded by the HHQ and the degradation complex molecules were probably simpler growth metabolites that could be used by the active cells of T. striata and P. bermudensis. HHQ is also known to degrade dimethylsulfoniopropionate (DMSP) [44]. Therefore, the degradation of DMSP may provide sulfur to the T. striata and P. bermudensis. 2-Heptyl-3-hydroxy-4(1H) quinolone (PQS) was found at a high concentration (188–544 µM) except at the low pH conditions where the concentration was 20.84 µM.
Furthermore, the concentration trend varied under different environmental conditions (Fig. 3). PQS is known to entrap iron by chelation and indirectly promotes siderophores production during the limiting nutrient conditions. PQS is known to act through at least three-distinct signaling pathways and separately has an iron scavenging mechanism, while its precursor, HHQ, cannot form an iron complex. In the previous studies, the exposure of P. aeruginosa mutant to PQS prevented the cells from synthesizing pyoverdine or pyochelin. PQS was found to be involved in cell envelope formation and inhibited bacterial growth. The secondary functions/multi-functions of PQS include iron entrapment, which facilitates siderophore-mediated iron delivery [46]. Thus, based on the current results, the higher PQS concentration may be involved in the entrapment and release of iron under iron-starved conditions wherein T. striata could have benefited.
Effect of nutrient limitation and replete conditions
Tetraselmis striata under T-PB conditions (with P. bermudensis) and nitrogen, sulfur, and trace metal limitations showed a higher growth rate than it did under T conditions. However, all the nutrient limitation conditions induced restricted growth compare to the nutrient replete conditions (Additional file 1: Figure S6). The trace metal and nitrate limitation exhibited higher growth inhibition. In comparison, the nitrate limitation results revealed as the initial increase in the cell abundance of the microalgae with a slower growth rate. This observation indicates that an organic nitrogen source was potentially provided by the partner bacteria (P. bermudensis) or was due to strategical utilization of the organic nitrogen released by the cells of the same population (Additional file 1: Figure S6).
The total lipid contents of T-PB under limited sulfur, metal, and nitrate conditions were 7.65 ± 0.08, 15.34 ± 0.62, and 32.47 ± 0.24% with biomass productivities of 18.93 ± 1.15, 10.97 ± 0.77, and 16.48 ± 1.18 mg L−1 day−1, respectively, while the total lipid contents under T conditions with limited sulfate, metal, and nitrate were 7.34 ± 0.48, 20.17 ± 0.82, and 31.47 ± 0.2% with biomass productivities of 10.60 ± 1.15, 7.45 ± 1.88, and 2.08 ± 0.72, mg L−1 day−1, respectively (Fig. 4). The sulfate limitation reduced the lipid content, but it maximized the biomass productivity most among the nutrient limitation conditions. Sulfur limitation in Tetraselmis has been reported to cause higher accumulation of starch than lipid [47]. The higher biomass productivity under co-cultivated conditions compared to axenic conditions indicated that the P. bermudensis involved in multi-functional co-operation to the T. striata. Furthermore, of the nutrient limitation conditions, nitrate limitation enhanced the lipid content, while the sulfur and trace metal limitations reduced the lipid content. Moreover, only nitrate limitation induced lipid accumulation, which showed a 1.46–1.55-fold higher lipid content than that under the nutrient replete condition (O3-control) and, therefore, was selected for the two-stage cultivation in the current study (Fig. 4).

Biomass productivity and total lipid (%) content of Tetraselmis striata in axenic conditions (T) with different treatments of marine broth (MB) and Pelagibaca bermudensis exudates (t) in O3 medium. The values are presented in mean ± SD (n = 3). Error bars are showing SD. Post hoc analysis is shown in the Additional file 2: Table S1 (G, H) for comparing the means according to their significant difference (LSD)
However, all nutrient limited conditions restricted the growth of T. striata in the presence and absence of P. bermudensis. T. striata cells exhibited a higher survival rate in the presence of P. bermudensis than under T conditions (Additional file 1: Figure S5). The trace metal might have been regenerated by the re-mineralization process of the decaying cells or the metalloproteins/metallopeptides, or both [48]. The hypothesis proposed in an independent study on DMSP degradation by the Roseobacter clade bacteria using HHQ [44] could be one of the strategies to address the partial requirement of sulfur for T. striata. Therefore, the microalgal cell abundance and biomass productivity were higher under limited sulfur levels than under axenic T. striata cultivation with limited sulfur levels (Fig. 4 and Additional file 1: Figure S5).
Effect of exo-metabolites and mixotrophy
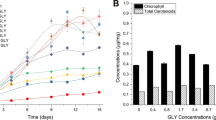
The crude exudates containing metabolites released by P. bermudensis were shown to elevate the biomass productivity by up to 41.63 and 91.73% at 1 and 5% of exudates, respectively (Fig. 5). Furthermore, 1 and 5% exo-metabolite treatment increased the biomass of the T. striata successively, while the 10% treatment did not enhance the growth as reflected in the reduced cell abundance in the stationary phase compared to the MB-treated cultures (Fig. 6). This indicated that the T. striata could not cope with the higher metabolite concentration of P. bermudensis. The results indicate that the physical presence of P. bermudensis was not essential for the growth promoting effect, and the metabolites released by P. bermudensis were closely involved in promoting the microalgal growth. The microalgal cells did not grow in 100% exudate of P. bermudensis and MB (t), and the 100% MB-containing medium (without exudates of P. bermudensis) showed that T. striata did not grow mixotrophically or heterotrophically under these conditions.

Biomass productivity, lipid productivity, and total lipid content (%) of Tetraselmis striata in axenic (T) and co-culture (T-PB) conditions under nutrient limitation (sulfate, nitrate, and trace metal), nutrient replete, and two-stage cultivation mode (salinity increase, nitrate limitation, and decreased pH). The values are presented in mean ± SD (n = 3). Error bars are showing SD

Cell abundance of T. striata in varying P. bermudensis exudates (t) and marine broth exudates (MB) in O3 media. The values are presented in mean ± SD (n = 3). Error bars are showing SD
The HHQ (10 µM) and PQS (1000 µM) individually exhibited algicidal effect on the axenic T. striata (Additional file 1: Figure S6). These precursors are beneficial in formation of biofilm of P. bermudensis on the cell wall of T. striata for mutualistic growth but increased concentration control the growth of T. striata. In addition, the exudates released by P. bermudensis containing the cocktail of growth promoting metabolites as well as algicidal compounds, so the net effect of the metabolites was growth promoting (Fig. 5).
However, the culture, which was exposed to only MB in the O3 medium (without the exudates of P. bermudensis), also exhibited enhanced growth under successive concentration (5 and 10% MB in O3) due to the mixotrophic capability of the T. striata; however, the rate was less than that of the exudate-treated cultures (t). MB also contains organic substrates and nutrients. A higher organic substrate concentration (100% MB and 100% t) elevates the intracellular CO2 level and, hence, causes toxicity to microalgae [9]. Furthermore, 5% was the optimum condition with a 68.28 ± 2.12 mg L−1 day−1 biomass productivity and 21.43 ± 1.41% lipid content. Therefore, this condition is lucrative for the scaleup in wastewater/saline effluent as well as in seawater (Fig. 5). Mixotrophic cultivation is an impressive strategy for wastewater remediation where high organic load containing various organic substrates with inorganic nutrients would be available to synergize the maximum biomass production [49,50,51]. Furthermore, biomass productivity would be even higher under sunlight conditions, since the exudates enhanced mixotrophic growth employed in the current experiment had only 45 µM m−2 s−1. Furthermore, to scale up the cultivation of T. striata, extensive investigation on mechanisms of growth stimulating and growth inhibiting allelopathic compounds is essential, at lab, pilot, and commercial production scales [52].
Amelioration of biomass productivity and lipid content using two-stage cultivation
The two-stage cultivation (15 day, first stage; 3 day, second stage) technique was performed in an attempt to ameliorate the lipid content. The biochemical composition and precursors were modulated by the abrupt changes in the medium and physical environment, which decreased the microalgal cell abundance. The salinity increase in the two-stage cultivation mode exhibited a biomass productivity of 14.81 ± 1.52 and 28.52 ± 2.39 mg L−1 day−1 with 23.04 ± 0.71 and 19.04 ± 0.58% lipid content under T and T-PB conditions, respectively. Furthermore, the pH decrease (low pH, 6.0) in the stage II resulted in a biomass productivity of 15.23 ± 1.16 and 21.62 ± 3.44 mg L−1 day−1 with a lipid content of 22.78 ± 0.39 and 19.20 ± 0.17%, respectively (Fig. 4). The nitrate limitation in the two-stage cultivation resulted in a biomass productivity of 30.28 ± 2.18 and 38.95 ± 2.45 mg L−1 day−1 with an increased lipid content of 29.75 ± 0.97 and 31.37 ± 1.46% under T and T-PB conditions, respectively (Fig. 4). Salinity and pH in the two-stage mode did not result in higher lipid content with a more impressive biomass productivity than the control (nutrient replete condition).
The strategic stress induced for the 3 days could not be optimum, but the results of this study support the usefulness of the two-stage cultivation model for T. striata under both T and T-PB conditions using nitrate limitation. The optimum stress induced by the elevated salinity/temperature/nutrient limitation mediated biochemical macromolecular synthesis, which would modulate metabolism to induce the levels of carbohydrate or lipid accumulation in Tetraselmis; however, the response varies within genera/species [10, 18, 19, 47]. Optimum stress conditions in two-stage cultivation for the amelioration of lipid productivity have been previously demonstrated in Tetraselmis, Scenedesmus, Acutodesmus, Nannochloropsis, Chlorella, Synechococcus, and Monoraphidium and Chlamydomonas [9, 19, 53,54,55,56,57] where the type, time, and intensity of the stress matter. Two-stage cultivation could enhance potential of overall biofuel production process by increasing the desirable biochemical feedstock. Furthermore, it creates an opportunity to use the enormous potential of varied effluents strategically, while providing nutrients and a suitable physical environment for attaining the maximum lipid productivity [50, 51, 57]. The growth promotion effect shown in some of the optimized conditions for higher biomass productivity and lipid productivity are in agreement with the previous studies performed in other co-cultivation strategies. However, the increment folds in lipid yields compared to the axenic condition were higher in the immobilized conditions in Chlorella spp. (Table 2) [58,59,60,61,62,63,64,65,66].
The results obtained in the current study, indicating that the microalgal biofuel co-cultivation model would be applicable for the range of environmental variables. However, the open pond cultivation would have several climatic factors as well as biotic factors. The dominant biotic stressor and interaction of P. bermudensis with the native seawater bacterial communities needed to be investigated in more details. Optimization of the inoculum density of P. bermudensis as well as the search for the other co-operating bacteria would be essential to overcome with the competition of seawater native bacterial communities.
The previous studies done by our group [12] to evaluate the feasibility of this model with native seawater bacteria indicated that the cell abundance of P. bermudensis (105) was incapable to outcompete the seawater bacterial communities, but growth promotion was also seen under the adverse conditions. This was probably due to metabolic products released by the growth promoting probiotic bacteria. The effect shown by the cell culture filtrate treatment of P. bermudensis (t) on T. striata confirmed this hypothesis that metabolic exudates of the P. bermudensis could show growth promoting effect. The large-scale open pond cultivation should be investigated to unravel the exo-metabolomic dynamics (growth hormones as well as algicidal compounds released by the native seawater or saline water communities) to exactly pinpoint the mechanisms and realize the application.
It would be an interesting methodological question that which growth phase of T. striata and P. bermudensis should be employed to see the mutualistic interaction in addition to varying stressors (as variables) in different culture mode. Furthermore, the interactions studies in varying co-cultivation in light of multi omics approaches will certainly unfold the vivid transcriptomic and metabolic changes, real-time growth promoting effects, and metabolic nutrients trade-off between the organisms [67, 68].
Conclusion
Co-cultivation of T. striata with P. bermudensis could be a potentially robust and sustainable technique for biomass generation using a wide spectrum of abiotic parameters with ameliorated stress tolerance. P. bermudensis also demonstrated a growth promoting effect on T. striata. Two-stage cultivation using nitrate limitation and continuous high-light exposure with optimum salinity (20 g L−1) at pH 8.0, as well as metabolic feed of P. bermudensis is suggested as a useful strategy for attaining higher lipid productivity of T. striata. Exo-metabolomics would certainly unfold the vivid interactions between microalgae and bacteria, and would elucidate the specific role of growth promoting metabolites and their implications for ensuring lipid productivity.
Change history
03 July 2018
After publication of the original article [1], it was brought to our attention that Figs. 4, 5, and 6 and their captions were incorrect. The correct figures and captions are presented below:
Abbreviations
- HHQ:
-
4-hydroxy-2-alkylquinolines
- PQS (Pseudomonas quorum signal):
-
2-heptyl-3-hydroxy-4(1H)-quinolone
- T:
-
axenic condition
- T-PB:
-
co-cultivation condition
References
González-González LM, Correa DF, Ryan S, Jensen PD, Pratt S, Schenk PM. Integrated biodiesel and biogas production from microalgae: towards a sustainable closed loop through nutrient recycling. Renew Sustain Energy Rev. 2018;82:1137–48.
Guldhe A, Bhola V, Rawat I, Bux F. Carbon dioxide sequestration by microalgae: biorefinery approach for clean energy and environment. In: Singh B, Bauddh K, Bux F, editors. Algae and environmental sustainability. New Delhi: Springer India; 2015. p. 147–54.
Patidar SK, Mishra SK, Bhattacharya S, Ghosh T, Paliwal C, Goel S, Mishra S. Naturally floating microalgal mat for in situ bioremediation and potential for biofuel production. Algal Res. 2015;9:275–82.
Patidar SK, Mishra S. Carbon sequestration by microalgae: a green approach for climate change mitigation. In: Abraham M, editor. Reference module in Earth systems and environmental sciences from encyclopedia of sustainable technologies. Amsterdam: Elsevier; 2017. p. 477–83.
Amaro HM, Guedes AC, Malcata FX. Advances and perspectives in using microalgae to produce biodiesel. Appl Energy. 2011;88:3402–10.
Borowitzka MA, Moheimani NR. Sustainable biofuels from algae. Mitig Adapt Strateg Glob Change. 2013;18:13–25.
Patidar SK, Mitra M, Goel S, Mishra S. Effect of carbon supply mode on biomass and lipid in CSMCRI’s Chlorella variabilis (ATCC 12198). Biomass Bioenergy. 2016;86:1–10.
Mitra M, Patidar SK, Mishra S. Integrated process of two stage cultivation of Nannochloropsis sp for nutraceutically valuable eicosapentaenoic acid along with biodiesel. Bioresour Technol. 2015;193:363–9.
Patidar SK, Mitra M, George B, Soundarya R, Mishra S. Potential of Monoraphidium minutum for carbon sequestration and lipid production in response to varying growth mode. Bioresour Technol. 2014;172:32–40.
Ishika T, Moheimani NR, Bahri PA. Sustainable saline microalgae co-cultivation for biofuel production: a critical review. Renew Sustain Energy Rev. 2017;78:356–68.
Wang H, Hill RT, Zheng T, Hu X, Wang B. Effects of bacterial communities on biofuel-producing microalgae: stimulation, inhibition and harvesting. Crit Rev Biotechnol. 2016;36:341–52.
Park J, Park BS, Wang P, Patidar SK, Kim JH, Kim SH, Han MS. Phycospheric native bacteria Pelagibaca bermudensis and Stappia sp. ameliorate biomass productivity of Tetraselmis striata (KCTC1432BP) in co-cultivation system through mutualistic interaction. Front. Plant Sci. 2017;8:289.
Amavizca E, Bashan Y, Ryu CM, Farag MA, Bebout BM. de-Bashan LE, Enhanced performance of the microalga Chlorella sorokiniana remotely induced by the plant growth-promoting bacteria Azospirillum brasilense and Bacillus pumilus. Sci Rep. 2017;7:41310.
de-Bashan LE, Mayali X, Bebout BM, Weber PK, Detweiler AM, Hernandez JP, Bashan Y. Establishment of stable synthetic mutualism without co-evolution between microalgae and bacteria demonstrated by mutual transfer of metabolites (NanoSIMS isotopic imaging) and persistent physical association (fluorescent in situ hybridization). Algal Res. 2016;15:179–86.
Fuentes JL, Garbayo L, Cuaresma M, Montero Z, González-del-Valle M, Vílchez C. Impact of microalgae-bacteria interactions on the production of algal biomass and associated compounds. Mar Drugs. 2016;14:100.
Amin SA, Hmelo LR, van Tol HM, Durham BP, Carlson LT, Heal KR, et al. Interaction and signaling between a cosmopolitan phytoplankton and associated bacteria. Nature. 2015;522:98–101.
Park BS, Joo JH, Baek KD, Han MS. A mutualistic interaction between the bacterium Pseudomonas asplenii and the harmful algal species Chattonella marina (Raphidophyceae). Harmful Algae. 2016;56:29–36.
Kim ZH, Park H, Lee CG. Seasonal assessment of biomass and fatty acid productivity by Tetraselmis sp in the ocean using semi-permeable membrane photobioreactors. J Microbiol Biotechnol. 2016;26:1098–102.
Pereira H, Gangadhar KN, Schulze PS, Santos T, de Sousa CB, Schueler LM, Malcata FX, Gouveia L, Varela JCS, Barreira L. Isolation of a euryhaline microalgal strain, Tetraselmis sp. CTP4, as a robust feedstock for biodiesel production. Sci Rep. 2016;6:35663.
Biondi N, Cheloni G, Rodolfi L, Viti C, Giovannetti L, Tredici MR. Tetraselmis suecica F&M-M33 growth is influenced by its associated bacteria. Microbial Biotechnol. 2017. https://doi.org/10.1111/1751-7915.12865.
Meseck SL, Smith BC, Wikfors GH, Alix JH, Kapareiko D. Nutrient interactions between phytoplankton and bacterioplankton under different carbon dioxide regimes. J Appl Phycol. 2007;19:229–37.
Arora M, Chandrashekar A, Delany J, Rajarajan N, Emami K, Mesbahi E. Carbohydrate degrading bacteria closely associated with Tetraselmis indica: influence on algal growth. Aquat Biol. 2012;15:61–71.
Brinkhoff T, Giebel HA, Simon M. Diversity, ecology, and genomics of the Roseobacter clade: a short overview. Arch Microbiol. 2008;189:531–9.
Cho JC, Giovannoni SJ. Pelagibaca bermudensis gen. nov., sp. nov., a novel marine bacterium within the Roseobacter clade in the order Rhodobacterales. Int J Syst Evol Microbiol. 2006;56(4):855–9.
Cude WN, Buchan A. Acyl-homoserine lactone-based quorum sensing in the Roseobacter clade: complex cell-to-cell communication controls multiple physiologies. Front Microbiol. 2013;4:336.
Michael V, Frank O, Bartling P, Scheuner C, Göker M, Brinkmann H, Petersen J. Biofilm plasmids with a rhamnose operon are widely distributed determinants of the ‘swim-or-stick’ lifestyle in Roseobacters. ISME J. 2016;10:2498–513.
Miller MB, Bassler BL. Quorum sensing in bacteria. Annu Rev Microbiol. 2001;55(1):165–99.
Al-Mailem D, Kansour M, Radwan S. Bacterial communities associated with biofouling materials used in bench-scale hydrocarbon bioremediation. Environ Sci Pollut R. 2015;22(5):3570–85.
Thrash JC, Cho JC, Ferriera S, Johnson J, Vergin KL, Giovannoni SJ. Genome sequences of Pelagibaca bermudensis HTCC2601T and Maritimibacter alkaliphilus HTCC2654T, the type strains of two marine Roseobacter genera. J Bacteriol. 2010;192:5552–3.
Liu Y, Xu G, Han R, Dong J, Ni Y. Identification of d-carbamoylase for biocatalytic cascade synthesis of d-tryptophan featuring high enantioselectivity. Bioresour Technol. 2018;249:720–8.
Zhao S, et al. Discovery of new enzymes and metabolic pathways by using structure and genome context. Nature. 2013;502(7473):698.
Choudhary S, Claudia S-D. Applications of quorum sensing in biotechnology. Appl Microbiol Biot. 2010;86:1267–79.
Petersen J, Brinkmann H, Bunk B, Michael V, Päuker O, Pradella S. Think pink: photosynthesis, plasmids and the Roseobacter clade. Environ Microbiol. 2012;14:2661–72.
Wagner-Döbler I, Biebl H. Environmental biology of the marine Roseobacter lineage. Annu Rev Microbiol. 2006;60:255–80.
Natrah FMI, Kenmegne MM, Wiyoto W, Sorgeloos P, Bossier P, Defoirdt T. Effects of micro-algae commonly used in aquaculture on acyl-homoserine lactone quorum sensing. Aquaculture. 2011;317:53–7.
Bruhn JB, Gram L, Belas R. Production of antibacterial compounds and biofilm formation by Roseobacter species are influenced by culture conditions. Appl Environ Microbiol. 2007;73:442–50.
McIntosh L, Cattolico RA. Preservation of algal and higher plant ribosomal RNA during extraction and electrophoretic quantitation. Anal Biochem. 1978;91:600–12.
Joo JH, Kuang Z, Wang P, Park BS, Patidar SK, Han MS. Ecological assessment of an algaecidal naphthoquinone derivate for the mitigation of Stephanodiscus within a mesocosm. Environ Pollut. 2017;229:735–45.
Folch J, Lees M, Sloane-Stanley GH. A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem. 1957;226:497–509.
Ortori CA, Halliday N, Cámara M, Williams P, Barrett DA. LC–MS/MS quantitative analysis of quorum sensing signal molecules. Pseudomonas Methods Protoc. 2014;1149:255–70.
Bohnert HJ, Nelson DE, Jensen RG. Adaptations to environmental stresses. Plant Cell. 1995;7:1099.
Kleist S, Ulbrich M, Bill N, Schmidt-Hohagen K, Geffers R, Schomburg D. Dealing with salinity extremes and nitrogen limitation–an unexpected strategy of the marine bacterium Dinoroseobacter shibae. Environ Microbiol. 2017;19:894–908.
Shin H, Hong SJ, Yoo C, Han MA, Lee H, Choi HK, Cho BK. Genome-wide transcriptome analysis revealed organelle specific responses to temperature variations in algae. Sci Rep. 2016;6:37770.
Harvey EL, Deering RW, Rowley DC, Gamal AE, Schorn M, Moore BS, Johnson MD, Mincer TJ, Whalen KE. A bacterial quorum-sensing precursor induces mortality in the marine coccolithophore. Emiliania huxleyi. Front Microbiol. 2016;7:59.
Seyedsayamdost MR, Wang R, Kolter R, Clardy J. Hybrid biosynthesis of roseobacticides from algal and bacterial precursor molecules. J Am Chem Soc. 2014;136(43):15150–3.
Diggle SP, Matthijs S, Wright VJ, Fletcher MP, Chhabra SR, Lamont IL, Kong X, Hider RC, Cornelis P, Cámara M, Williams P. The Pseudomonas aeruginosa 4-quinolone signal molecules HHQ and PQS play multifunctional roles in quorum sensing and iron entrapment. Chem Biol. 2007;14:87–96.
Yao C, Ai J, Cao X, Xue S, Zhang W. Enhancing starch production of a marine green microalga Tetraselmis subcordiformis through nutrient limitation. Bioresour Technol. 2012;118:438–44.
Maranger R, Bird DF, Price NM. Iron acquisition by photosynthetic marine phytoplankton from ingested bacteria. Nature. 1998;396:248–51.
Bhatnagar A, Chinnasamy S, Singh M, Das KC. Renewable biomass production by mixotrophic algae in the presence of various carbon sources and wastewaters. Appl Energy. 2011;88:3425–31.
Mohan SV, Rohit MV, Chiranjeevi P, Hariprasad R, Arora S. Integrating microalgae cultivation with wastewater treatment for biodiesel production. In: Das D, editor. Algal biorefinery an integrated approach. Cham: Springer International Publishing; 2015. p. 321–37.
Mitra M, Shah F, Bharadwaj SV, Patidar SK, Mishra S. Cultivation of Nannohloropsis oceanica biomass rich in eicosapentaenoic acid utilizing wastewater as nutrient resource. Bioresour Technol. 2016;218:1178–86.
Mendes LBB, Vermelho AB. Allelopathy as a potential strategy to improve microalgae cultivation. Biotechnol Biofuels. 2013;6(1):152.
Li X, Wang M, Liao X, Chen H, Dai Y, Chen B. Two stages of N-deficient cultivation enhance the lipid content of Microalga Scenedesmus sp. J Am Oil Chem Soc. 2015;92:503–12.
Gruber-Brunhumer M, Nussbaumer M, Jerney J, Ludwig I, Zohar E, Lang I, Bochmann G, Schagerl M, et al. Two-stage cultivation of N-rich and N-deprived biomass: influence of cultivation and dewatering methods on microalgal biomass used in anaerobic digestion. Algal Res. 2016;17:105–12.
Wensel P, Helms G, Hiscox B, Davis WC, Kirchhoff H, Bule M, Yu L, Chen S. Isolation, characterization, and validation of oleaginous, multi-trophic, and haloalkaline-tolerant microalgae for two-stage cultivation. Algal Res. 2014;4:2–11.
Ho SH, Nakanishi A, Ye X, Chang JS, Hara K, Hasunuma T, Kondo A. Optimizing biodiesel production in marine Chlamydomonas sp. JSC4 through metabolic profiling and an innovative salinity-gradient strategy. Biotechnol Biofuels. 2014;7(1):97.
Schulze PS, Carvalho CF, Pereira H, Gangadhar KN, Schüler LM, Santos TF, Varela JCS, Barreira L. Urban wastewater treatment by Tetraselmis sp. CTP4 (Chlorophyta). Bioresour Technol. 2017;223:175–83.
Kim BH, Ramanan R, Cho DH, Oh HM, Kim HS. Role of Rhizobium, a plant growth promoting bacterium, in enhancing algal biomass through mutualistic interaction. Biomass Bioenerg. 2014;69:95–105.
Cho DH, Ramanan R, Heo J, Lee J, Kim BH, Oh HM, Kim HS. Enhancing microalgal biomass productivity by engineering a microalgal–bacterial community. Bioresour Technol. 2015;175:578–85.
Ryu BG, Kim EJ, Kim HS, Kim J, Choi YE, Yang JW. Simultaneous treatment of municipal wastewater and biodiesel production by cultivation of Chlorella vulgaris with indigenous wastewater bacteria. Biotechnol Bioprocess Eng. 2014;19(2):201–10.
Do Nascimento M, de los Angeles Dublan M, Ortiz-Marquez JCF, Curatti L. High lipid productivity of an Ankistrodesmus–Rhizobium artificial consortium. Bioresour Technol. 2013;146:400–7.
Tanabe Y, Okazaki Y, Yoshida M, Matsuura H, Kai A, Shiratori T, Ishida K, Nakano S, Watanabe MM. A novel alphaproteobacterial ectosymbiont promotes the growth of the hydrocarbon-rich green alga Botryococcus braunii. Sci Rep. 2015;5:10467.
Kim HJ, Choi YK, Jeon HJ, Bhatia SK, Kim YH, Kim YG, Choi KY, Kim HJ, Lee SH, Lee YK, Yang YH. Growth promotion of Chlorella vulgaris by modification of nitrogen source composition with symbiotic bacteria, Microbacterium sp. HJ1. Biomass Bioenergy. 2015;74:213–9.
Wang R, Xue S, Zhang D, Zhang Q, Wen S, Kong D, Yan C, Cong W. Construction and characteristics of artificial consortia of Scenedesmus obliquus-bacteria for S. obliquus growth and lipid production. Algal Res. 2015;12:436–45.
de-Bashan LE, Bashan Y, Moreno M, Lebsky VK, Bustillos JJ. Increased pigment and lipid content, lipid variety, and cell and population size of the microalgae Chlorella spp. when co-immobilized in alginate beads with the microalgae-growth-promoting bacterium Azospirillum brasilense. Can J Microbiol. 2002;48(6):514–21.
Gonzalez LE, Bashan Y. Increased growth of the microalga Chlorella vulgaris when coimmobilized and cocultured in alginate beads with the plant-growth-promoting bacterium Azospirillum brasilense. Appl Environ Microbiol. 2000;66(4):1527–31.
Hom EF, Aiyar P, Schaeme D, Mittag M, Sasso S. A chemical perspective on microalgal–microbial interactions. Trends Plant Sci. 2015;20(11):689–93.
Padmaperuma G, Kapoore RV, Gilmour DJ, Vaidyanathan S. Microbial consortia: a critical look at microalgae co-cultures for enhanced biomanufacturing. Crit Rev Biotechnol. 2017. https://doi.org/10.1080/07388551.2017.1390728.
Authors’ contributions
SKP, M-SH, BSP, and JHK contributed for the research design and data analyses. All authors have read the final version of the manuscript and gave feedback on the content. The experimental work was done by SKP. JP and SHK assisted in axenic culture preparation and maintenance of cultures for the experiments. Manuscript is written and edited by SKP and approved by M-SH. All authors read and approved the final manuscript.
Acknowledgements
SKP is grateful for the BK21plus postdoctoral fellowship, Marine Biotechnology Program (PJT200255) and Industry-Academic Co-operation Foundation. Authors are also acknowledging the anonymous reviewers for the constructive suggestions.
Competing interests
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential competing interests.
Availability of data and materials
The data sets used and/or analyzed during the current study are available from the corresponding author/lead author on reasonable request.
Consent for publication
All the authors agree for their authorship, declaration of the results, and submission of the manuscript for peer review.
Ethics approval and consent to participate
No conflicts, informed consent, human or animal rights applicable for this research.
Funding
This research was supported by a grant from Marine Biotechnology Program (PJT 200255) and a project entitled “Development of marine microalgal biofuel production technology” funded by the Ministry of Oceans and Fisheries, South Korea.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding authors
Additional files
Additional file 1: Figure S1.
Cell abundance of Pelagibaca bermudensis in co-cultivation (Tetraselmis striata–P. bermudensis; T-PB) culture during the different interval of the growth (at different pH). Figure S2. Cell abundance of Pelagibaca bermudensis in co-cultivation (Tetraselmis striata–P. bermudensis; T-PB) culture during the different interval of the growth (at different salinity). Figure S3. Cell abundance of Tetraselmis striata in axenic (T) and co-cultivated (T-PB) growth mode at different photoperiods (12:12 and 24:00 hrs with 147 µM m−2 s−1). Figure S4. Cell abundance of Pelagibaca bermudensis in co-cultivation (Tetraselmis striata–P. bermudensis; T-PB) culture during the different interval of the growth (at different temperature and light conditions). Figure S5. Cell abundance of Tetraselmis striata (T) exposed to varied HHQ and PQS concentration and controls (without HHQ and PQS; with equal nontoxic acetonitrile). Figure S6. Cell abundance of Tetraselmis striata in axenic (T) and co-cultivated (T-PB) growth mode at different nutrient limited and replete conditions in O3 media.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Patidar, S.K., Kim, SH., Kim, J.H. et al. Pelagibaca bermudensis promotes biofuel competence of Tetraselmis striata in a broad range of abiotic stressors: dynamics of quorum-sensing precursors and strategic improvement in lipid productivity. Biotechnol Biofuels 11, 102 (2018). https://doi.org/10.1186/s13068-018-1097-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13068-018-1097-9