
Abstract
Epithelial-mesenchymal transition (EMT) is a fundamental process for embryonic development during which epithelial cells acquire mesenchymal characteristics, and the underlying mechanisms confer malignant features to carcinoma cells such as dissemination throughout the organism and resistance to anticancer treatments. During the past decades, an entire class of molecules, called non-coding RNA (ncRNA), has been characterized as a key regulator of almost every cellular process, including EMT. Like protein-coding genes, ncRNAs can be deregulated in cancer, acting as oncogenes or tumor suppressors. The various forms of ncRNAs, including microRNAs, PIWI-interacting RNAs, small nucleolar RNAs, transfer RNA-derived RNA fragments, long non-coding RNAs, and circular RNAs can orchestrate the complex regulatory networks of EMT at multiple levels. Understanding the molecular mechanism underlying ncRNAs in EMT can provide fundamental insights into cancer metastasis and may lead to novel therapeutic approaches. In this review, we describe recent advances in the understanding of ncRNAs in EMT and provide an overview of recent ncRNA applications in the clinic.
Similar content being viewed by others

Background
Epithelial-mesenchymal transition (EMT) is a highly dynamic cellular process that transiently converts epithelial into mesenchymal characteristics. Although, EMT and the reverse process, mesenchymal-epithelial transition (MET), play crucial roles in specific steps of embryogenesis, the underlying molecular mechanisms are reactivated during tumor progression [1]. Upon activation of EMT, epithelial cells deconstruct their junctions, reprogram gene expression signatures and signaling pathways, modulate their cytoskeletal organization, destabilize apical–basal polarity and concomitantly acquire front–rear polarity [2]. These changes confer attributes of ‘high-grade malignancy’ on carcinoma cells, including motility, invasive behavior, cancer stem cell (CSC)-properties, resistance to conventional therapies and immunoevasive and immunosuppressive features [2]. Extracellular stimuli derived from the tumor microenvironment can trigger activation of several signaling pathways such as TGFβ/SMAD, WNT/β-catenin, Notch, and receptor tyrosine kinase signaling pathways [1]. These signaling pathways initiate EMT programs in tumor cells through induction of a core set of EMT transcription factors including three different protein families, namely the basic helix–loop–helix factors TWIST1 and TWIST2, the zinc-finger E-box-binding homeobox factors ZEB1 and ZEB2 and the SNAIL family of zinc-finger factors SNAI1 (also known as SNAIL) and SNAI2 (also known as SLUG) [3]. These transcription factors, in different combinations, repress the expression of epithelial-related genes such as CDH1 (E-cadherin) and concomitantly induce the expression of mesenchymal-related genes such as CDH2 (N-cadherin) and vimentin. Moreover, cellular motility and cytoskeletal changes are regulated by Rho GTPase family such as RhoA, RAC1 and CDC42 [1].
Preliminary investigation on the molecular mechanisms underlying EMT has widely focused on protein-coding genes, however, the discovery of non-coding RNAs (ncRNAs) has revolutionized our perception of the molecular mechanisms behind EMT. ncRNAs comprise a heterogeneous class of RNA transcripts (Table 1) with different biogenesis, length, and functions, which account for almost 60% of the human transcriptome [4,5,6]. Recently, functional studies have uncovered that ncRNAs, like protein-coding genes, can participate in diverse cellular processes such as EMT.
In this review, we discuss the roles of various forms of ncRNAs in regulation of EMT in cancer and mention RNA modifications, which may result in the loss or gain of binding sites on ncRNAs with relation to EMT. Lastly, we focus on the potential clinical relevance of EMT-related ncRNAs in oncology.
General functions of ncRNAs
microRNAs
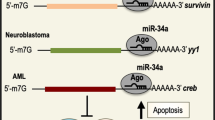
microRNAs (miRNAs) constitute a highly conserved class of small ncRNAs (19–24 nucleotides) that lead to translational repression or degradation of target mRNAs through binding between the seed sequence of the miRNA and complementary sequences in the 3′ untranslated region (3′UTR) of the mRNA [7]. miRNAs can interact with various key players of EMT leading to the formation of highly complex gene-regulatory networks during tumor progression and metastasis (Fig. 1).

miRNAs regulate the EMT process in cancer. miRNAs post-transcriptionally suppress the expression of key players of the EMT program at multiple levels. Initially, EMT is triggered upon activation of several pathways such as TGF-β and WNT/β-catenin signaling pathways. The multiple components of these signaling pathways are targeted by various miRNAs. Activation of these signaling pathways promotes the expression of EMT-inducing transcription factors (ZEB, SNAIL and TWIST) that function pleiotropically to induce the acquisition of the mesenchymal properties. These transcription factors bind to the promoter regions of specific miRNAs and regulate their expression. On the other hand, miRNAs can target the 3′UTRs of the mRNAs that encode these transcription factors. Some of these miRNAs and transcription factors form a double negative feedback loop. The TGF-β signaling pathway regulates cytoskeletal dynamics through regulating RhoA and CDC42, which are targeted by miRNAs. mRNAs encoding adhesion molecules such as E-cadherin and N-cadherin are also targeted by miRNAs. Green boxes represent an EMT inhibitory role by the indicated miRNAs, whereas red boxes represent the induction of the EMT process by the indicated miRNAs.
PIWI-interacting RNAs
P-element Induced WImpy testis (PIWI)-interacting RNAs (piRNAs) are a class of small ncRNAs (21–35 nucleotides in length) that are loaded onto members of the PIWI subfamily of Argonaute proteins to repress transposons in germline cells [8]. However, recent findings have indicated that aberrantly expressed and malfunctioning piRNAs can lead to the development and progression of human malignancies [9, 10].
Small nucleolar RNAs
Small nucleolar RNAs (snoRNAs) are generally classified as small ncRNAs of 60–300 nucleotides in length. snoRNAs act as guide RNAs for the post-transcriptional modification of ribosomal and small nuclear RNAs [11]. Based on structural characteristics, snoRNAs can be categorized into two families: C/D box snoRNAs (SNORDs) and H/ACA box snoRNAs (SNORAs) [12].
Transfer RNA-derived RNA fragments
Transfer RNA-derived RNA fragments (tRFs) are single-stranded ncRNA transcripts (14–30 nucleotides) that are cleaved from mature transfer RNAs (tRNAs). Like miRNAs, tRFs interact with Argonaute proteins to impair the translation of mRNAs through binding to target 3′UTRs [13].
Long non-coding RNAs
Long non-coding RNAs (lncRNAs) constitute a heterogeneous class of ncRNAs that are at least 200 nucleotides [13]. Contrary to small ncRNAs, lncRNAs employ diverse mechanisms of action to perform their functional roles [14], and can be classified as scaffolds, decoys, or guides (Fig. 2). Scaffold lncRNAs can facilitate the interactions between various components of a complex, decoy lncRNAs can interact with either proteins or RNAs and titrate them away from their natural targets and guide lncRNAs can interact with proteins to localize them at specific genomic loci [12, 15] (Fig. 2). However, the miRNA decoy mechanism is somewhat controversial [16], in particular when the proposed decoy only contributes a tiny fraction of the total pool of miRNA targets in a cell [17]. Nevertheless, recent studies suggest that individual lncRNAs may function at substoichiometric ratios through seeding of concentration gradients in specific spatial territories or phase-separated compartments [18, 19] or by mediating target-directed miRNA degradation [20, 21].

lncRNAs and circRNAs have versatile modes of action: A Guide lncRNAs can interact with regulatory proteins (such as epigenetic activators, epigenetic repressors or transcription factors) and direct them to their target regions. B Decoy lncRNAs and circRNAs can bind and sequester miRNAs or proteins. C Scaffold lncRNAs can function as a central platform to allow the assembly of various molecular components to facilitate their intermolecular interactions. D Enhancer RNAs are a class of ncRNAs that are transcribed from enhancer regions and act in regulating mRNA transcription. E mRNA-binding lncRNAs can bind to mRNAs and enhance or reduce its stability
Circular RNAs
Circular RNAs (circRNAs) belong to a family of large ncRNAs that are characterized by covalently closed circular structures that are generated by a process called backsplicing of linear precursor RNAs [22, 23]. Like lncRNAs, circRNAs can act as decoys to sequester specific miRNAs [24, 25] or, conversely, may act to stabilize specific miRNAs [20, 22, 26]. Besides, circRNAs can interact with proteins by serving as scaffolds to assemble multiple components or as decoys that sequester the target proteins [13, 27,28,29,30,31]. In addition, some circRNAs have been proposed to encode unique proteins [32], however, the vast majority are believed to be non-coding [33]. As for the lncRNAs, controversies are associated with many circRNAs being proposed to function as miRNA decoys [34, 35]. Despite, hundreds of circRNAs have been claimed to have miRNA sponging properties, only few harbors more miRNA binding sites than expected by change [36] and most are very lowly expressed and probably non-functional [37]. Intriguingly, one of the most studied circRNAs in cancer, ciRS-7, was recently shown to be absent in cancer cells within solid tumors [38], even though numerous previous studies suggested that it functions as a miR-7 sponge in cancer cells. Rather it is expressed in the stromal cells and positive correlations with miR-7 target genes can be explained by stromal co-expression and were also observed for other stromal-enriched circRNAs that do not contain miR-7 binding sites [38].
Small non-coding RNAs in EMT
miRNA
miRNAs and EMT-transcription factors
Single miRNAs can target several genes and a single gene can be targeted by multiple miRNAs. Thus, in combination with downstream transcriptional changes arising from miRNA-targeted transcription factors highly complex gene-regulatory networks result, which might buffer or enforce signaling pathways [39]. For example, members of the miR-200 family and miR-205 cooperate to strongly inhibit EMT through suppressing ZEB1/2 expression [40]; conversely, ZEB1/2 directly suppress the transcription of the miR-200 family [41]. A similar mutually inhibitory loop exists between miR-1199-5p and Zeb1, coordinating EMT and tumor metastasis [42]. Such double-negative feedback loops might act as epithelial/mesenchymal switches that confer a high degree of plasticity required during the complex route from the primary tumor to metastasis [42, 43]. Similarly, double-negative feedback loops occur between SLUG and miR-1 and miR-200 [44], as well as between SLUG and miR-203. Interestingly, SLUG also inhibits miR-200 family members and thus might act in concert with the mutually inhibitory loop between miR-200 and ZEB1/2, to promote the EMT process [45]. Additionally, double-negative feedback loops also occurs between SNAIL and the miR-34 family (miR-34a/b/c) [46] as well as between SNAIL and miR-203 [47]. The miR-203/SNAIL regulatory complex in concert with the miR200/ZEB feedback loop might construct an EMT core network that could function as a robust switch regulating cell plasticity [47]. In addition, TWIST1 binds to the promoter regions of miR-200 and miR-205 and probably inhibits their expression to promote EMT in bladder cancer [48]. The detailed interplay between miRNAs and TWIST1 has been reviewed elsewhere [49]. Finally, the tumor-suppressor miRNA let-7 inhibits EMT through binding to multiple target sites in the 3′UTR of high mobility group A2 (HMGA2), a chromatin-binding protein that could directly bind to the promoter regions of SNAIL and TWIST1 and promote their expression [50]. Together, several intertwined mutually inhibitory loops, in which miRNAs and EMT master transcription factors repress the expression of each other, robustly reinforce the epithelial or mesenchymal states and may orchestrate a quick and strong response to minimal stimuli [43].
miRNAs regulate cell adhesion and cytoskeletal components
In addition to targeting EMT master transcription factors, miRNAs also modulate the expression of cytoskeletal and cell adhesion components. For example, miR-9, which is directly activated by MYC and MYCN, induces EMT and metastasis by repressing the translation of E-cadherin [51]. Similarly, miR-495 represses E-cadherin expression and promotes EMT and tumorigenicity of breast cancer cells [52]. Alternatively, miR-194, whose expression is reduced in liver mesenchymal-like cancer cells, inhibits invasion and migration, partly through targeting of N-cadherin [53] and miR-30a, whose expression is induced by RUNX3, directly targets vimentin and represses its expression [54]. Finally, miR-375 suppresses the expression of CLAUDIN-1, thereby contributing to the dissolution of tight junctions during EMT in lung cancer [55].
miRNAs regulate EMT signaling pathways
The TGF-β signaling pathway, which plays a central role in promoting EMT in various tissue types, modulates miRNA expression at both transcriptional and post-transcriptional levels through Smads [56]. Upon treatment with TGF-β or bone morphogenetic protein (BMP), the Smad proteins directly bind to the RNA-Smad binding element (R-SBE) found in the stem region of primary transcripts of a set of miRNAs (such as miR-21and miR-181) and facilitate the recruitment of Drosha to the R-SBE-containing pri-miRNAs, promoting the processing of these primary transcripts [57, 58]. Moreover, TGF-β control the transcription of various miRNAs through R-SBEs in the miRNA promoter. For example, TGF-β released from M2 macrophages promotes binding of Smad2/3 to the miR-362-3p promoter, resulting in upregulation of miR-362-3p in hepatocellular carcinoma (HCC) cells [59]. miR-362-3p directly targets CD82, a key player in the tetraspanin network, thereby maintaining EMT in HCC cells [59]. In addition, TGF-β-activated Smads indirectly regulate miRNA expression through activation of transcription factors that bind to their promoters [56]. On the other hand, miRNAs may regulate the TGF-β signaling pathway at multiple levels through targeting of ligands and receptors as well as Smad and non-Smad pathway components. For instance, members of the miR-200 and/or miR-30 families inhibit the invasive ability and promote MET in anaplastic thyroid carcinoma cells through targeting TGFBR-1 and Smad2 [60]. Similarly, miR-190, which is directly suppressed by ZEB1, inhibits TGF-β-induced EMT and metastasis through targeting Smad2 in breast cancer both in vitro and in vivo [61].
Similarly, WNT/β-catenin signaling orchestrates a reciprocal crosstalk with miRNAs to regulate EMT in tumor cells. The β-catenin/LEF1 complex transcribes expression of miR-150, which in turn directly targets CREB1 and EP300 to facilitate EMT in colorectal cancer (CRC) cells [62]. On the other hand, the miR-34 family, inhibits EMT through targeting key components of the canonical WNT signaling pathway such as β-catenin, WNT1, WNT3, LRP6, SNAIL, and AXIN2 [63, 64]. Moreover, miR-374a, whose ectopic expression induces EMT and metastasis both in vitro and in vivo, promotes the WNT/β-catenin pathway by targeting negative regulators of the WNT/β-catenin signaling cascade, such as WIF1, PTEN, and WNT5A [65].
miRNAs regulate cell motility
miRNAs can modulate actin cytoskeletal dynamics during EMT. A study using high-throughput sequencing of RNA isolated by HITS-CLIP technology uncovered hundreds of miR-200a and miR-200b targets that are predominantly enriched for genes associated with the cytoskeletal remodeling [66]. Similarly, miR-23b, a metastatic suppressor miRNA, could regulate cytoskeletal remodeling in breast cancer through directly suppressing a set of genes, including PAK2, LIMK2, ARHGEF6, CFL2, PIK3R3, PLAU, and ANXA2 [67]. Inhibition of miR-23b, using a miR-23b sponge construct, induces spontaneous metastasis in an orthotopic mouse model of breast cancer [67]. In contrast, miR-155 induces EMT, tight junction dissolution, migration and invasion through targeting RhoA in breast cancer [68].
Other small non-coding RNAs and EMT
In addition to miRNAs, other small non-coding RNAs, such as snoRNAs, piRNAs and tRFs can regulate EMT in cancer; however, their contribution to EMT, is only beginning to be uncovered. For instance, SNORD78, which is upregulated in non-small cell lung cancer (NSCLC) and associated with poor prognosis, may contribute to the invasion and EMT in NCSLC probably through aberrant methylation of CDH1 [69] and SNORD113–1, which is downregulated in HCC, inhibits tumor cell growth via inhibiting the phosphorylation of ERK1/2 and Smad2/3 in the MAPK-ERK and TGF-β pathways [70].
piRNAs may also regulate EMT. piR-932 is highly expressed in breast cancer cells with EMT and CSC phenotypes [9]. Functionally, the combination of piR-932 and PIWIL2 serve as a positive regulator of EMT in breast CSCs through mediating the methylation of LXN [9]. Similarly, piR-1037, which is upregulated in oral squamous cell carcinoma (OSCC) cells and xenografts, promotes EMT and motility of OSCC cells likely through interacting with XIAP [10].
Besides, tRF may participate in the regulation of EMT. For example, tRF/miR-1280, a small fragment (17-bp) that is derived from both tRNALeu and pre-miRNA inhibits CSC and EMT phenotypes and metastasis in CRC [71]. Mechanistically, tRF/miR-1280 interacts with 3′UTR of JAG2, a Notch ligand, and inhibits its expression, leading to suppression of Notch/Gata and miR-200b pathways [71].
Long non-coding RNAs in EMT
lncRNAs
lncRNAs and EMT-transcription factors
Recently, an increasing number of lncRNAs have been implicated in EMT and tumor metastasis through regulating EMT-transcription factors via various mechanisms. For instance, the lncRNA antisense to ZEB1 (ZEB1-AS1) can interact with MLL1 and recruit it to the promoter region of ZEB1 to epigenetically induce ZEB1 transcription, promoting EMT and tumor metastasis [72, 73]. In addition, lncRNA-BX111887 directly interacts with transcriptional factor Y-box protein (YB1) and recruits it to the ZEB1 promoter region and subsequently transactivates ZEB1 expression, promoting tumor growth and metastasis of pancreatic cancer in a xenograft mouse model [74]. Besides, a set of lncRNAs has been identified that can post-transcriptionally regulate ZEB expression by sequestering miRNAs (Table 2). lncRNA-activated by TGF-β (lncRNA-ATB) functions as a competitive endogenous RNA (ceRNA) to sequester members of the miR-200 family and sustain ZEB1/2 expression in HCC [75]. Interestingly, SNHG14, through acting as a miRNA sponge for miR-5590-3p, derepresses the expression of ZEB1 which in turn induces SNHG14 expression, forming a positive feedback loop [76]. Moreover, lncRNA RP11–138 J23.1 (RP11) binds hnRNPA2B1 protein to facilitate the interaction between hnRNPA2B1 and the mRNA of SIAH1 and FBXO45, ubiquitin E3 ligases that stimulate ZEB1 degradation. This complex facilitates the mRNA degradation of Siah1 and Fbxo45, preventing ZEB1 degradation [77]. Another lncRNA, lncRNA-HIT, interacts with ZEB1 and enhances its protein stability [78].
Similarly, the expression of SNAIL and SLUG is regulated by lncRNAs through multiple mechanisms. The lncRNA PVT1 transcriptionally promotes SLUG expression through directly interacting with the transcription factor STAT3 and recruiting it to the SNAI2 promoter [90]. Interestingly, the PVT1 locus produces multiple linear and circular transcripts, and it is important to consider that not all molecular assays can distinguish the linear and circular forms [141]. Besides, multiple lncRNAs have been identified that could post-transcriptionally regulate SNAIL or SLUG expression (Table 2). For instance, lncRNA GCMA functions as a ceRNA for miR-34a and miR-124 to derepress SNAIL and SLUG, respectively, thereby promoting EMT in vitro and metastasis in vivo [92]. Interestingly, TGF-β-induced lincRNAs, AC026904.1 and UCA1, cooperatively promote SLUG expression at transcriptional and post-transcriptional levels, respectively; UCA1 functions as a ceRNA to sequester miR-1 and miR-203 and derepress SLUG expression, while, AC026904.1 behaves as an enhancer RNA and directly binds the promoter region of SNAI2 to facilitate its transcription [91]. Besides, lncRNAs, such as HOTAIR and NEAT1, can mediate interaction between SNAIL and epigenetic machineries to specifically localize this complex at SNAIL binding sites on epithelial gene promoters thereby inducing EMT [142, 143]. Moreover, SNHG15 interacts with SLUG and prevents BTRC-induced SLUG ubiquitination and degradation, promoting EMT in CRC [94].
Finally, lncRNA JPX derepresses TWIST1 expression through sponging miR-33a-5p and this regulatory axis activates EMT and metastasis in vitro and in vivo through inducing WNT/β-catenin signaling [98]. Additionally, TWIST1 transcriptionally induces lncRNA LINC01296, which sequesters miR-598 and in turn derepresses its direct target TWIST1, thereby forming a positive feedback loop [99].
lncRNAs regulate cell adhesion and cytoskeletal components
Several lncRNAs (listed in Table 2) have been demonstrated to interact with epigenetic regulators to recruit them to the regulatory regions of CDH1. For example, TRERNA1 behaves as a scaffold to facilitate the recruitment of EZH2 to the CDH1 promoter and silence its expression in gastric cancer [100]. Additionally, MEG3 induces E-cadherin expression via functioning as a ceRNA for miR-21 and miR-421 [107, 108]. Besides, NEAT1 facilitates the interaction between FOXN3 and SIN3A, a transcriptional repressor complex that repress the expression of GATA3 and ZO1 (an epithelial marker), resulting in EMT promotion [109]. In addition to epithelial markers, mesenchymal markers such as vimentin are regulated by lncRNAs. As previously indicated, LINC00460 directly interacts with PRDX1 and recruits it to the promoter region of vimentin and other mesenchymal markers, and consequently induces their transcription [144]. In addition, MALAT1 and H19 upregulate vimentin expression through acting as a sponge for miR-30a-5p and miR-138, respectively [79, 110]. lncRNAs may also post-translationally regulate vimentin stability [111,112,113]. For instance, amine oxidase, copper containing 4, pseudogene (AOC4P) interacts with vimentin to promote its ubiquitin-dependent degradation, impairing EMT, tumor growth and metastasis in animal models [111] and LINC00675 interacts with vimentin to facilitate its phosphorylation thereby destabilizing vimentin filaments and inhibiting metastasis in gastric cancer [112].
lncRNAs regulate EMT signaling pathways
lncRNAs can regulate key components of TGF-β pathway through various mechanisms. TGFB2-antisense RNA1 (TGFB2-AS1), which is induced by canonical TGF-β signaling, physically associates with the PRC2 adaptor protein EED, and recruits it to the TGF-β-target genes to facilitate their methylation and suppression, thus forming a feedback loop [119]. Moreover, lncRNA ELIT-1, which its upregulation positively correlates with poor outcome in patients with lung adenocarcinoma and gastric cancer, interacts with Smad3 and recruits it to the promoters of multiple genes including SNAIL, vimentin, N-cadherin, API-1, and ELIT-1 itself to accelerate their transcription, forming a positive feedback loop which promotes EMT [145]. Additionally, lncRNA-ATB and lncRNA-XIST could promote TGFβ2 expression and tumor progression through acting as a sponge for miR-141-3p [114, 115]. Furthermore, MIR22HG lncRNA inhibits TGF-β signaling and EMT via competitively binding to Smad2 and perturbing the interaction between Smad2 and Smad4 in CRC [118]. Moreover, MALAT1 cooperates with SETD2 to form a scaffold, which facilitates binding of phosphorylated Smad2/3 to their specific phosphatase PPM1A, thus promoting the dephosphorylation of Smads and subsequently inhibiting TGF-β signaling [120].
Besides, lncRNAs can regulate various key players of the WNT pathway. For example, GATA6 antisense RNA 1 (GATA6-AS1) interacts with EZH2 and increases its occupation at FZD4 promoter to repress its expression, leading to inactivation of WNT/β-catenin pathway [121]. Similarly, oncogenic lncRNAs such as HERES and NEAT1 can epigenetically activate the WNT/β-catenin pathway via binding to EZH2 and recruiting it to the promoter regions of multiple negative regulators of the WNT/β-catenin pathway [122, 123]. Furthermore, RBM5 antisense (RBM5-AS1) directly associates with β-catenin and facilitates the recruitment of β-catenin and TCF4 complex to the WNT target genes SGK1, YAP1, and MYC, activating the WNT pathway in CRC [146]. Moreover, lincRNA-p21 physically associates with mRNAs encoding β-catenin (CTNNB1) and JunB (JUNB) and induces their translational repression via translational repressor Rck, leading to inhibition of WNT/β-catenin pathway [124]. Besides, lncRNA cytoskeleton regulator RNA (CYTOR) interacts with cytoplasmic β-catenin to prevent casein kinase 1 (CK1)-mediated phosphorylation of β-catenin, which leads to accumulation of β-catenin in the nucleus and induces transcriptional activity of the β-catenin/TCF complex [126]. Interestingly, lncRNA-MUF and LncCCAT1 facilitate the interaction between ANXA2 and GSK3β, which prevents β-catenin phosphorylation and degradation, through disrupting the formation of the GSK3β/β-catenin complex [95, 127]. In addition, NEAT1 directly interacts with DDX1, a protein that binds to β-catenin and enhances its protein stability and transcriptional activity, thus activating the WNT/β-catenin pathway [147].
lncRNAs regulate cell motility
lncRNAs also regulate EMT through modulating cytoskeletal dynamics. For instance, lncRNA-UCA1 promotes EMT in bladder cancer through sequestering tumor suppressive miR-145 that targets ZEB1/2 and fascin homologue 1 (FSCN1), an actin-binding protein that regulate actin-based cellular protrusions and cellular motility [130]. Besides, lncMER52A directly interacts with and stabilizes p120-catenin by preventing β-TrCP1-mediated ubiquitination and degradation [131]. Moreover, ABHD11 antisense RNA1 (ABHD11-AS1) and actin filament-associated protein 1-antisense RNA1 (AFAP1-AS1) directly interact with RhoC and facilitate EMT in ovarian cancer and osteosarcoma, respectively [148, 149]. Moreover, multiple, lncRNAs have been shown to upregulate the expression of Cdc42, RAC1 or RhoA through functioning as decoys for various miRNAs (Table 2), including H19 and MALAT1 that upregulate Cdc42 by sponging miR-15b and miR-1, respectively [132, 133].
circRNAs
circRNAs and EMT-transcription factors
In the first study to indicate a role for circRNAs in EMT a high-throughput RNA sequencing analysis demonstrated that the expression of hundreds of circRNAs are modulated during EMT, most of them upregulated [150]. The functions of circRNAs in EMT remain largely unknown, although more recently, several circRNAs have been proposed to regulate EMT transcription factors. For example, circPRMT5 may induce EMT in UCB cells and promotes an aggressive phenotype in a xenograft mouse model via sponging miR-30c, a tumor suppressor that directly inhibits SNAIL expression [151]. Similarly, circRNA_0084043 and circ-ZNF652 was suggested to derepress SNAIL expression through sponging of miRNAs [152, 153] and has-circ-000684, which is upregulated in gastric cancer cell lines and tumor tissues, derepresses ZEB1 by sponging miR-186 [154]. Moreover, circPCNXL2, which is highly expressed in clear cell renal cell carcinoma (ccRCC) and positively correlates with poor prognosis, induces invasion and proliferation in vitro and promotes tumor growth in a xenograft mouse model by acting as a miRNA sponge to inhibit the miR-153-mediated suppression of ZEB2 [155]. In addition, EMT transcription factors may transcriptionally regulate circRNA expression. For example, TWIST1 binds to the promoter region of the CUL2 gene to selectively induce transcription of a circRNA (circRNA-10,720), which exerts its oncogenic function in HCC by sponging multiple miRNAs that target vimentin [156].
circRNAs regulate cell junction and cytoskeletal components
Several circRNAs have been identified as EMT regulators through modulating key components of cell junctions and the cytoskeleton. Circ-AKT3 may sponge miR-296-3p to promote CDH1 expression, thereby suppressing migration and invasion in vitro as well as tumor cell dissemination in vivo [157]. Similarly, circPTPRA regulates E-cadherin and EMT in NSCLC by sponging of miR-96-5p, permitting the expression of the downstream tumor suppressor RASSF8, which interacts with E-cadherin and stabilizes adhesive junctions [158, 159]. In addition, circAMOTL1L upregulates the expression of protocadherin-α (Pcdha), a member of cadherin superfamily which mediates cell-cell adhesion, through behaving as a miRNA sponge for miR-193a-5p, thus, its loss in prostate cancer (PCa) promotes cell migration, invasion and EMT in vitro and tumor growth in vivo [160]. Interestingly, circPTK2 can bind to Ser38, Ser55, and Ser82 sites of vimentin and induce EMT in vitro and in vivo and targeting of circPTK2 using shRNA significantly suppressed tumor metastasis in a xenograft mouse model of CRC [161].
circRNAs regulate EMT signaling pathways
Deregulated circRNAs in cancer may also regulate EMT-related signaling pathways such as the TGF-β and WNT pathways. For example, circPTK2 was suggested to sponge miR-429 and miR-200b-3p to derepress TIF1γ, a protein that negatively regulates TGF-β/Smad signaling through Smad4 ubiquitination and competing with Smad4 to bind Smad2/3 complex [162,163,164]. Additionally, circANKS1B, whose upregulation is associated with lymph node metastasis and poor prognosis, induces EMT through sponging multiple miRNAs, including miR-148a and miR-152-3p that target USF1, a transcription factor that induces expression of TGF-β and ESPR1, forming a feedback loop [165]. Similarly, circUHRF1 promotes EMT in vitro and tumor growth in vivo via sponging miR-526b-5p to derepress the expression of c-Myc, which in turn promotes expression of TGF-β1 and ESRP1 [166]. Additionally, hsa_circ_0009361, which is downregulated in CRC tissues and cells, inhibits Wnt/β-catenin pathway and EMT through sponging miR-582 to upregulate APC2, a Wnt/β-catenin pathway inhibitor [167].
Another interesting study suggests that circGSK3β, directly interacts with GSK3β and inhibits GSK3β activity, thereby protecting β-catenin from phosphorylation and degradation which subsequently results in promoting β-catenin pathway and EMT in esophageal squamous cell carcinoma (ESCC) [168]. GSK3β-mediated β-catenin phosphorylation and degradation can be also antagonized via direct interaction of GSK3β with a novel 370-amino acid β-catenin isoform that is derived from circβ-catenin [169]. Moreover, circ-CTNNB1 directly interacts with DEAD-box polypeptide 3 (DDX3) to promote its physical association with transcription factor Yin Yang 1 (YY1), leading to transactivation of YY1 and transcriptional regulation of β-catenin pathway key components such as WNT1, WNT3, AXIN2, FZD10 and BMP4 [170]. Besides, circRNA-MYLK, whose expression is upregulated in bladder cancer and positively associated with advanced clinical stage, behaves as a miR-29a sponge to upregulate VEGF expression inducing VEGFA/VEGFR2 signaling pathway which consequently promotes EMT and angiogenesis in bladder cancer [171].
circRNAs regulate cell motility
circRNAs may also regulate key players of cytoskeletal dynamics. Interestingly, circHIAT1 was suggested to behave as a ‘miRNA reservoir’ to stabilize multiple tumor suppressive miRNAs such as miR-195-5p/29a-3p/29c-3p that target CDC42 in ccRCC [172]. Additionally, FLI1 exonic circRNAs (FECR) including FECR1 and FECR2 participate in the activation of ROCK1, a key effector of RhoA GTPase that promotes actin polymerization, through sequestering miR-584-3p from binding to ROCK1 [173]. Furthermore, circ-133, which is enriched in the exosomes derived from hypoxic CRC cells promotes tumor metastasis through regulating miR-133a/GEF-H1/RhoA axis [174]. These data indicate that the deregulated circRNA could regulate the expression of major regulators of cytoskeletal dynamics.
RNA modifications in EMT
Adenosine-to-inosine (A-to-I) RNA editing modifies targeting by ncRNAs
RNA editing is a post-transcriptional modification which remodels the transcriptional landscape and consequently governs cell fate decisions. To date, more than 100 distinct modifications of RNA have been identified, emphasizing the critical role of these modifications on genome output [175]. Of the RNA modifications, the conversion of adenosine to inosine, termed A-to-I editing, is the most abundant form of RNA editing in Metazoans which is mediated by the adenosine deaminase acting on RNA (ADAR) family of enzymes [175]. Upon A-to-I editing inosines are recognized as guanosines and base-pair with cytosines, suggesting that A-to-I editing can modify the transcripts secondary structures [176]. A-to-I editing can occur in protein-coding sequences, however most of these events occur in non-coding regions [177, 178]. RNA editing in miRNAs may have a profound effect on miRNA regulation, because alteration of a single nucleotide in the seed sequence of an individual miRNA can alter the base pairing properties, potentially leading to creation or disruption of miRNA binding sites [6]. For example, miR-200b has been found to be overedited in various tumors, and its edited form correlates with worse prognosis [179, 180]. Unlike wild-type miR-200b, the edited miR-200b loses its capacity to suppress ZEB1 and ZEB2 and concomitantly acquires the capability to suppress novel targets such as LIFR, a well-known anti-metastatic gene (Fig. 3A) [179]. This editing is mediated by ADAR1; thus, its inhibition impairs miR-200b editing which in turn inhibits tumor cell invasion and migration and impairs tumor growth in a xenograft mouse model of thyroid cancer [179, 180]. ADAR1 can also interact with Dicer and facilitates maturation of a set of oncogenic miRNAs that promotes migration, invasion and EMT in OSCC [181], and has recently been shown to regulate a large number of cancer-relevant circRNAs via both editing-dependent and independent mechanisms [182].

RNA modification alters ncRNA targeting. A Adenosine-to-inosine (A-to-I) editing of the seed sequence of a miRNA can alter the base pairing properties of the miRNA. The double-stranded RNA-specific adenosine deaminases (ADARs) can interact with target site (here the target site is the seed sequence of miR-200b) and change adenosine bases to inosine, thereby changing the sequence of the target site. In this example, ADARs change the seed sequence of the miR-200b. The edited miR-200b loses its ability to interact with 3′UTR of ZEB1 and ZEB2; while it concomitantly acquires the capability to interact with novel targets such as LIFR, a well-known anti-metastatic gene. Therefore, this process can change the tumor-suppressive miR-200b to an oncogenic miRNA. B Alternative polyadenylation (APA) in the 3′UTR can generate multiple mRNA transcripts with different 3′ UTRs. As shown here, the 3′ UTR of the candidate gene includes two APA sites which can give rise to two isoforms with short and long 3′ UTRs. The short isoform might produce more proteins due to escaping from repression by various components such as miRNAs, lncRNAs, and RNA-binding proteins
Alternative polyadenylation modifies targeting by ncRNAs
High-throughput sequencing technologies, genome-wide experimental and bioinformatic tools have uncovered that more than 70% of mammalian genes have alternative polyadenylation regions in the 3ʹUTR, which can give rise to multiple mRNA transcripts with different 3′ UTRs [183]. Tumor cells mainly express mRNA isoforms with shortened 3′UTRs which results from alternative polyadenylation [184]. Shortening of 3′UTRs can remarkably generate more protein relative to their full-length 3′ UTR counterparts probably by escaping from miRNA-mediated translational repression (Fig. 3B) [184]. Tumor cells expressing mRNA isoforms with shorter 3′UTR are considered to be more aggressive and result in a poorer prognosis [185]. In pancreatic ductal adenocarcinoma (PDAC), treatment with genotoxic agents induces shortening of ZEB1 3′UTR which increases ZEB1 protein production through escaping from repression by miRNAs such as miR-200 [186]. ZEB1 protein production directly associates with alternative polyadenylation of ZEB1 3′ UTR [186]. Additionally, during EMT and tumor progression, the TWIST1 3′UTR, containing several regulatory elements, is shortened which leads to increased TWIST1 protein production [187]. Similarly, the 3′ UTR of RAC1 and Fibronectin type III domain containing 3B (FNDC3B) is shortened which results in enhanced protein production during cancer progression [188, 189]. FNDC3B directly associates with and stabilize myosin heavy chain 9 (MYH9) to facilitate Wnt/β-catenin pathway activation [189].
EMT-related ncRNAs as cancer biomarkers
Accumulating evidence have demonstrated that ncRNAs are dysregulated across diverse cancers. Given the tissue and stage-specific expression patterns of many ncRNAs they are promising biomarkers for cancer detection and prognosis (Table 3). Owing to the covalently closed structure, circRNAs are highly resistant to exonucleases, making them a promising biomarker compared to other sub-classes of ncRNAs as they may be more readily detected in serum, plasma or urine, where the obtaining procedures are relatively convenient and less invasive compared to obtaining a tumor biopsy [190,191,192]. For example, the plasma levels of circGSK3β, a circRNA that promotes EMT and cancer progression, were shown to be significantly elevated in patients with ESCC [168]. Additionally, circPTK2 which promotes EMT and tumor metastasis, is significantly elevated in the serum of patients with CRC and the expression levels of circPTK2 is a metastatic indicator in patients with CRC [161]. Intriguingly, ncRNAs can be selectively packaged into exosomes or other extracellular vesicles, which protect them from RNase-mediated degradation in the extracellular space and in body fluids. The serum exosomal FECR1 (FLI1 exonic circular RNA) was shown to be markedly higher in patients with SCLC compared with that in heathy individuals and higher levels of exosomal FECR1 could serve as a biomarker of an unfavorable outcome in patients with SCLC [173]. Moreover, circPRMT5 is enriched in the serum and urinary exosomes of patients with UCB and upregulation of circPRMT5 in the serum and urinary exosomes is associated with lymph node metastasis [151]. These findings suggest that EMT-related ncRNAs, particularly circRNAs, can serve as diagnostic or prognostic biomarkers in cancer.
EMT-related ncRNAs as therapeutic targets
The EMT program leads to increased invasion and migration of tumor cells. Thus, targeting of EMT-related ncRNAs may hold therapeutic potential. To date, miRNAs are the most widely studied class of ncRNAs in cancer. One of the features of miRNAs that makes them attractive tools and targets for novel therapeutic strategies is their capacity to regulate several targets within a specific pathway or a set of targets across various pathways. Thus, certain miRNAs may lead to a stronger therapeutic effect if their multiple targets are enriched within a specific pathway [39]. The miR-200 family that regulates EMT process at multiple levels through targeting a set of mRNAs involved in Rho signaling pathway, invadopodia formation, focal adhesions, and EMT-related transcription factors, is a prominent example [39, 66].
The current strategies for miRNA-based therapeutics include reintroduction of tumor suppressive miRNAs using synthetic double-stranded oligoribonucleotides (also known as miRNA mimics) and/or inhibition of the oncogenic miRNAs via single-stranded antisense oligoribonucleotides (also known as antimiRs). These oligoribonucleotides are chemically modified to prevent RNase-mediated degradation, enhance binding affinity, and improve their pharmacokinetic characteristics in vivo [193, 194]. In addition, various delivery vehicles, including lipid-based nanoparticles and peptide and polymer-based systems are developed to encapsulate oligoribonucleotides to protect them from degradation and facilitate endosomal escape [194].
To date, several miRNAs have reached clinical development (Table 4). For example, miR-16 mimics has recently completed a phase I clinical trial (NCT02369198) with encouraging results in patients with malignant pleural mesothelioma [195]. Additionally, a locked nucleic acid (LNA)-modified antimir-155 has advanced to a phase II clinical trial (NCT03837457) in patients with certain lymphomas and leukemias.
The development of lncRNA-based therapeutics is only in its infancy, however, their appealing properties such as their dysregulation in malignancies, tissue-specific expression, and tight-transcriptional control make them promising candidates for cancer therapeutic targeting. In parallel, targeting a specific lncRNA using antisense oligonucleotides (ASOs) which triggers RNases H-mediated degradation has made lncRNA-based therapeutics feasible [211]. These lncRNA-targeting ASOs can be chemically modified using 2′-O-Me or LNAs to improve their stability in vivo [211]. Taken together, growing knowledge of ncRNAs in EMT and metastasis and further improvements of nucleotide modifications and in vivo delivery systems may ultimately enable the translation of this novel knowledge into clinical practice.
Concluding remarks and future perspectives
EMT is a highly dynamic cellular program that enables epithelial cells to transiently acquire a mesenchymal phenotype. Instead of oscillating between a complete epithelial and a complete mesenchymal state, cells can reside at an intermediate state, displaying both epithelial and mesenchymal features [43, 212]. Partial EMT in a cancerous context has been observed in experimental models and clinical settings [213, 214]. These studies demonstrated that the high plasticity of partial EMT enables tumor cells to adapt to a stressful environment during circulation, colonizing and formation of metastases, emphasizing the pivotal function of partial EMT in metastasis [213]. Although the detailed molecular mechanisms underlying partial EMT still remain elusive, mathematic modeling and experimental approaches demonstrated that double-negative feedback loops such as the miR-34/SNAIL and the miR-200/ZEB loops function as the ‘motor of cellular plasticity’ to control epithelial-hybrid-mesenchymal transitions [215,216,217]. Furthermore, cancer cells at the invasive front of solid tumors may undergo EMT, dependent on the link between miR-205 and ZEB1/2 [218]. These observations suggest that miRNAs are highly involved in regulating partial EMT and tumor metastasis, making them attractive candidates for diagnostic and therapeutic approaches. However, further investigation of the link between partial EMT and various forms of ncRNAs will be required to employ them in the development of novel therapeutic and diagnostic tools.
Moreover, tumor cells with an intermediate EMT phenotype, tend to develop CSC-like characteristics [43, 212, 219]. CSCs represent a minor subpopulation of tumor cells that can evade the immune system and are more resistant to most conventional therapeutics. Besides, CSCs may harbor tumor-propagating and metastatic capabilities [3]. Thus, given the crucial role of EMT and CSCs in tumor progression and the frequent development of resistance to various therapeutics, yielding novel therapeutic approaches designed to target the EMT process is promising. Anti-EMT therapeutic approaches can be achieved via inhibiting EMT initiation, targeting cancer cells that have undergone EMT as well as induction of an EMT reversal program in cancer cells [219]. Application of these strategies, using small-molecule inhibitors and biological agents, such as monoclonal antibodies, have been reviewed elsewhere and promisingly several of them has reached clinical trials or even received FDA approval [219]. ncRNA-based therapeutics can also be used to target EMT and may have distinct advantages over other compounds [13], for instance by targeting several RNAs simultaneously, which may lead to less acquired therapy resistance and better clinical outcomes. Additionally, as the ncRNA-based therapeutics are mainly based on base-pairing, the potential therapeutics can be easily designed and synthesized to be tested in preclinical and clinical models, while high-throughput screening and structure-based strategies should be employed for potential small molecule inhibitors and biological agents [13]. Despite these advantages, the major challenges in developing ncRNA-based therapeutics include successful delivery to the target cells while preventing nuclease-mediated degradation, avoiding immune system activation, and reducing off-target effects [13].
As summarized in this review, ncRNAs regulate EMT at multiple levels. Although the role of miRNAs in regulating EMT has been intensively investigated, the impact of other ncRNAs is still far from being fully understood and several controversies are associated with the lncRNA and circRNA research fields. In particular, concerning studies suggesting that lowly expressed lncRNAs or circRNAs containing only one or few miRNA binding sites function as efficient miRNA sponges [16, 34, 220]. More research is needed to gain a deeper understanding of the roles of ncRNAs as EMT-regulators in cancer, which may facilitate the development of novel diagnostic and therapeutic tools to impair metastasis and resistance to therapy.
Availability of data and materials
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Abbreviations
- 3′UTR:
-
3′ untranslated region
- A-to-I:
-
Adenosine-to-inosine
- ASO:
-
Antisense oligonucleotide
- ccRCC:
-
Clear cell renal cell carcinoma
- ceRNA:
-
Competitive endogenous RNA
- circRNA:
-
Circular RNA
- CRC:
-
Colorectal cancer
- CSC:
-
Cancer stem cell
- EMT:
-
Epithelial-mesenchymal transition
- ESCC:
-
Esophageal squamous cell carcinoma
- HCC:
-
Hepatocellular carcinoma
- LNA:
-
Locked nucleic acid
- lncRNA:
-
Long non-coding RNAs
- MET:
-
Mesenchymal-epithelial transition
- miRNAs:
-
microRNAs
- ncRNA:
-
Non-coding RNA
- NSCLC:
-
Non-small cell lung cancer
- OSCC:
-
Oral squamous cell carcinoma
- piRNA:
-
P-element Induced WImpy testis (PIWI)-interacting RNA
- R-SBE:
-
RNA-Smad binding element
- snoRNA:
-
Small nucleolar RNA
- SNORA:
-
H/ACA box snoRNA
- SNORD:
-
C/D box snoRNA
- tRF:
-
Transfer RNA-derived RNA fragment
- tRNA:
-
Transfer RNA
References
Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–96.
Yang J, Antin P, Berx G, Blanpain C, Brabletz T, Bronner M, et al. Guidelines and definitions for research on epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2020;21(6):341–52.
Dongre A, Weinberg RA. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat Rev Mol Cell Biol. 2019;20(2):69–84.
Djebali S, Davis CA, Merkel A, Dobin A, Lassmann T, Mortazavi A, et al. Landscape of transcription in human cells. Nature. 2012;489(7414):101–8.
Luo Y, Hitz BC, Gabdank I, Hilton JA, Kagda MS, Lam B, et al. New developments on the Encyclopedia of DNA elements (ENCODE) data portal. Nucleic Acids Res. 2020;48(D1):D882–d9.
Anastasiadou E, Jacob LS, Slack FJ. Non-coding RNA networks in cancer. Nat Rev Cancer. 2018;18(1):5–18.
Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol. 2014;15(8):509–24.
Ozata DM, Gainetdinov I, Zoch A, O’Carroll D, Zamore PD. PIWI-interacting RNAs: small RNAs with big functions. Nat Rev Genet. 2019;20(2):89–108.
Zhang H, Ren Y, Xu H, Pang D, Duan C, Liu C. The expression of stem cell protein Piwil2 and piR-932 in breast cancer. Surg Oncol. 2013;22(4):217–23.
Li G, Wang X, Li C, Hu S, Niu Z, Sun Q, et al. Piwi-interacting RNA1037 enhances chemoresistance and motility in human oral squamous cell carcinoma cells. Onco Targets Ther. 2019;12:10615–27.
Williams GT, Farzaneh F. Are snoRNAs and snoRNA host genes new players in cancer? Nat Rev Cancer. 2012;12(2):84–8.
Wong CM, Tsang FH, Ng IO. Non-coding RNAs in hepatocellular carcinoma: molecular functions and pathological implications. Nat Rev Gastroenterol Hepatol. 2018;15(3):137–51.
Slack FJ, Chinnaiyan AM. The role of non-coding RNAs in oncology. Cell. 2019;179(5):1033–55.
Fico A, Fiorenzano A, Pascale E, Patriarca EJ, Minchiotti G. Long non-coding RNA in stem cell pluripotency and lineage commitment: functions and evolutionary conservation. Cell Mol Life Sci. 2019;76(8):1459–71.
Wang KC, Chang HY. Molecular mechanisms of long noncoding RNAs. Mol Cell. 2011;43(6):904–14.
Thomson DW, Dinger ME. Endogenous microRNA sponges: evidence and controversy. Nat Rev Genet. 2016;17(5):272–83.
Bosson AD, Zamudio JR, Sharp PA. Endogenous miRNA and target concentrations determine susceptibility to potential ceRNA competition. Mol Cell. 2014;56(3):347–59.
Unfried JP, Ulitsky I. Substoichiometric action of long noncoding RNAs. Nat Cell Biol. 2022;24(5):608–15.
Elguindy MM, Mendell JT. NORAD-induced Pumilio phase separation is required for genome stability. Nature. 2021;595(7866):303–8.
Kleaveland B, Shi CY, Stefano J, Bartel DP. A network of noncoding regulatory RNAs acts in the mammalian brain. Cell. 2018;174(2):350–62.e17.
Ghini F, Rubolino C, Climent M, Simeone I, Marzi MJ, Nicassio F. Endogenous transcripts control miRNA levels and activity in mammalian cells by target-directed miRNA degradation. Nat Commun. 2018;9(1):3119.
Chen LL. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat Rev Mol Cell Biol. 2020;21(8):475–90.
Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019;20(11):675–91.
Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495(7441):333–8.
Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495(7441):384–8.
Piwecka M, Glazar P, Hernandez-Miranda LR, Memczak S, Wolf SA, Rybak-Wolf A, et al. Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science. 2017;357(6357):eaam8526.
Holdt LM, Stahringer A, Sass K, Pichler G, Kulak NA, Wilfert W, et al. Circular non-coding RNA ANRIL modulates ribosomal RNA maturation and atherosclerosis in humans. Nat Commun. 2016;7:12429.
Ashwal-Fluss R, Meyer M, Pamudurti NR, Ivanov A, Bartok O, Hanan M, et al. circRNA biogenesis competes with pre-mRNA splicing. Mol Cell. 2014;56(1):55–66.
Pandey PR, Yang JH, Tsitsipatis D, Panda AC, Noh JH, Kim KM, et al. circSamd4 represses myogenic transcriptional activity of PUR proteins. Nucleic Acids Res. 2020;48(7):3789–805.
Abdelmohsen K, Panda AC, Munk R, Grammatikakis I, Dudekula DB, De S, et al. Identification of HuR target circular RNAs uncovers suppression of PABPN1 translation by CircPABPN1. RNA Biol. 2017;14(3):361–9.
Du WW, Yang W, Chen Y, Wu ZK, Foster FS, Yang Z, et al. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur Heart J. 2017;38(18):1402–12.
Pamudurti NR, Bartok O, Jens M, Ashwal-Fluss R, Stottmeister C, Ruhe L, et al. Translation of CircRNAs. Mol Cell. 2017;66(1):9–21.e7.
Stagsted LV, Nielsen KM, Daugaard I, Hansen TB. Noncoding AUG circRNAs constitute an abundant and conserved subclass of circles. Life Sci Alliance. 2019;2(3):e201900398.
Jarlstad Olesen MT, Kristensen LS. Circular RNAs as microRNA sponges: evidence and controversies. Essays Biochem. 2021;65(4):685–96.
Kristensen LS, Jakobsen T, Hager H, Kjems J. The emerging roles of circRNAs in cancer and oncology. Nat Rev Clin Oncol. 2022;19(3):188–206.
Guo JU, Agarwal V, Guo H, Bartel DP. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014;15(7):409.
Xu C, Zhang J. Mammalian circular RNAs result largely from splicing errors. Cell Rep. 2021;36(4):109439.
Kristensen LS, Ebbesen KK, Sokol M, Jakobsen T, Korsgaard U, Eriksen AC, et al. Spatial expression analyses of the putative oncogene ciRS-7 in cancer reshape the microRNA sponge theory. Nat Commun. 2020;11(1):4551.
Bracken CP, Scott HS, Goodall GJ. A network-biology perspective of microRNA function and dysfunction in cancer. Nat Rev Genet. 2016;17(12):719–32.
Gregory PA, Bert AG, Paterson EL, Barry SC, Tsykin A, Farshid G, et al. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat Cell Biol. 2008;10(5):593–601.
Bracken CP, Gregory PA, Kolesnikoff N, Bert AG, Wang J, Shannon MF, et al. A double-negative feedback loop between ZEB1-SIP1 and the microRNA-200 family regulates epithelial-mesenchymal transition. Cancer Res. 2008;68(19):7846–54.
Diepenbruck M, Tiede S, Saxena M, Ivanek R, Kalathur RKR, Lüönd F, et al. miR-1199-5p and Zeb1 function in a double-negative feedback loop potentially coordinating EMT and tumour metastasis. Nat Commun. 2017;8(1):1168.
Lu W, Kang Y. Epithelial-mesenchymal plasticity in cancer progression and metastasis. Dev Cell. 2019;49(3):361–74.
Liu YN, Yin JJ, Abou-Kheir W, Hynes PG, Casey OM, Fang L, et al. MiR-1 and miR-200 inhibit EMT via Slug-dependent and tumorigenesis via Slug-independent mechanisms. Oncogene. 2013;32(3):296–306.
Ding X, Park SI, McCauley LK, Wang C-Y. Signaling between transforming growth factor β (TGF-β) and transcription factor SNAI2 represses expression of microRNA miR-203 to promote epithelial-mesenchymal transition and tumor metastasis. J Biol Chem. 2013;288(15):10241–53.
Siemens H, Jackstadt R, Hünten S, Kaller M, Menssen A, Götz U, et al. miR-34 and SNAIL form a double-negative feedback loop to regulate epithelial-mesenchymal transitions. Cell Cycle (Georgetown, Tex). 2011;10(24):4256–71.
Moes M, Le Béchec A, Crespo I, Laurini C, Halavatyi A, Vetter G, et al. A novel network integrating a miRNA-203/SNAI1 feedback loop which regulates epithelial to mesenchymal transition. PLoS One. 2012;7(4):e35440.
Wiklund ED, Bramsen JB, Hulf T, Dyrskjot L, Ramanathan R, Hansen TB, et al. Coordinated epigenetic repression of the miR-200 family and miR-205 in invasive bladder cancer. Int J Cancer. 2011;128(6):1327–34.
Khanbabaei H, Teimoori A, Mohammadi M. The interplay between microRNAs and Twist1 transcription factor: a systematic review. Tumour Biol. 2016;37(6):7007–19.
Lee YS, Dutta A. The tumor suppressor microRNA let-7 represses the HMGA2 oncogene. Genes Dev. 2007;21(9):1025–30.
Ma L, Young J, Prabhala H, Pan E, Mestdagh P, Muth D, et al. miR-9, a MYC/MYCN-activated microRNA, regulates E-cadherin and cancer metastasis. Nat Cell Biol. 2010;12(3):247–56.
Hwang-Verslues WW, Chang PH, Wei PC, Yang CY, Huang CK, Kuo WH, et al. miR-495 is upregulated by E12/E47 in breast cancer stem cells, and promotes oncogenesis and hypoxia resistance via downregulation of E-cadherin and REDD1. Oncogene. 2011;30(21):2463–74.
Meng Z, Fu X, Chen X, Zeng S, Tian Y, Jove R, et al. miR-194 is a marker of hepatic epithelial cells and suppresses metastasis of liver cancer cells in mice. Hepatology (Baltimore, Md). 2010;52(6):2148–57.
Liu Z, Chen L, Zhang X, Xu X, Xing H, Zhang Y, et al. RUNX3 regulates vimentin expression via miR-30a during epithelial-mesenchymal transition in gastric cancer cells. J Cell Mol Med. 2014;18(4):610–23.
Yoda S, Soejima K, Hamamoto J, Yasuda H, Nakayama S, Satomi R, et al. Claudin-1 is a novel target of miR-375 in non-small-cell lung cancer. Lung Cancer (Amsterdam, Netherlands). 2014;85(3):366–72.
Blahna MT, Hata A. Smad-mediated regulation of microRNA biosynthesis. FEBS Lett. 2012;586(14):1906–12.
Davis BN, Hilyard AC, Lagna G, Hata A. SMAD proteins control DROSHA-mediated microRNA maturation. Nature. 2008;454(7200):56–61.
Davis BN, Hilyard AC, Nguyen PH, Lagna G, Hata A. Smad proteins bind a conserved RNA sequence to promote microRNA maturation by Drosha. Mol Cell. 2010;39(3):373–84.
Zhang Q, Huang F, Yao Y, Wang J, Wei J, Wu Q, et al. Interaction of transforming growth factor-β-Smads/microRNA-362-3p/CD82 mediated by M2 macrophages promotes the process of epithelial-mesenchymal transition in hepatocellular carcinoma cells. Cancer Sci. 2019;110(8):2507–19.
Braun J, Hoang-Vu C, Dralle H, Hüttelmaier S. Downregulation of microRNAs directs the EMT and invasive potential of anaplastic thyroid carcinomas. Oncogene. 2010;29(29):4237–44.
Yu Y, Luo W, Yang ZJ, Chi JR, Li YR, Ding Y, et al. miR-190 suppresses breast cancer metastasis by regulation of TGF-β-induced epithelial-mesenchymal transition. Mol Cancer. 2018;17(1):70.
Guo YH, Wang LQ, Li B, Xu H, Yang JH, Zheng LS, et al. Wnt/β-catenin pathway transactivates microRNA-150 that promotes EMT of colorectal cancer cells by suppressing CREB signaling. Oncotarget. 2016;7(27):42513–26.
Kim NH, Kim HS, Li X-Y, Lee I, Choi H-S, Kang SE, et al. A p53/miRNA-34 axis regulates Snail1-dependent cancer cell epithelial–mesenchymal transition. J Cell Biol. 2011;195(3):417–33.
Kim NH, Kim HS, Kim NG, Lee I, Choi HS, Li XY, et al. p53 and microRNA-34 are suppressors of canonical Wnt signaling. Sci Signal. 2011;4(197):ra71.
Cai J, Guan H, Fang L, Yang Y, Zhu X, Yuan J, et al. MicroRNA-374a activates Wnt/β-catenin signaling to promote breast cancer metastasis. J Clin Invest. 2013;123(2):566–79.
Bracken CP, Li X, Wright JA, Lawrence DM, Pillman KA, Salmanidis M, et al. Genome-wide identification of miR-200 targets reveals a regulatory network controlling cell invasion. EMBO J. 2014;33(18):2040–56.
Pellegrino L, Stebbing J, Braga VM, Frampton AE, Jacob J, Buluwela L, et al. miR-23b regulates cytoskeletal remodeling, motility and metastasis by directly targeting multiple transcripts. Nucleic Acids Res. 2013;41(10):5400–12.
Kong W, Yang H, He L, Zhao J-j, Coppola D, Dalton WS, et al. MicroRNA-155 is regulated by the transforming growth factor β/Smad pathway and contributes to epithelial cell plasticity by targeting RhoA. Mol Cell Biol. 2008;28(22):6773–84.
Zheng D, Zhang J, Ni J, Luo J, Wang J, Tang L, et al. Small nucleolar RNA 78 promotes the tumorigenesis in non-small cell lung cancer. J Exp Clin Cancer Res. 2015;34(1):49.
Xu G, Yang F, Ding CL, Zhao LJ, Ren H, Zhao P, et al. Small nucleolar RNA 113-1 suppresses tumorigenesis in hepatocellular carcinoma. Mol Cancer. 2014;13:216.
Huang B, Yang H, Cheng X, Wang D, Fu S, Shen W, et al. tRF/miR-1280 suppresses stem cell-like cells and metastasis in colorectal cancer. Cancer Res. 2017;77(12):3194–206.
Su W, Xu M, Chen X, Chen N, Gong J, Nie L, et al. Long noncoding RNA ZEB1-AS1 epigenetically regulates the expressions of ZEB1 and downstream molecules in prostate cancer. Mol Cancer. 2017;16(1):142.
Li T, Xie J, Shen C, Cheng D, Shi Y, Wu Z, et al. Upregulation of long noncoding RNA ZEB1-AS1 promotes tumor metastasis and predicts poor prognosis in hepatocellular carcinoma. Oncogene. 2016;35(12):1575–84.
Deng SJ, Chen HY, Ye Z, Deng SC, Zhu S, Zeng Z, et al. Hypoxia-induced LncRNA-BX111 promotes metastasis and progression of pancreatic cancer through regulating ZEB1 transcription. Oncogene. 2018;37(44):5811–28.
Yuan JH, Yang F, Wang F, Ma JZ, Guo YJ, Tao QF, et al. A long noncoding RNA activated by TGF-β promotes the invasion-metastasis cascade in hepatocellular carcinoma. Cancer Cell. 2014;25(5):666–81.
Zhao L, Liu Y, Zhang J, Liu Y, Qi Q. LncRNA SNHG14/miR-5590-3p/ZEB1 positive feedback loop promoted diffuse large B cell lymphoma progression and immune evasion through regulating PD-1/PD-L1 checkpoint. Cell Death Dis. 2019;10(10):731.
Wu Y, Yang X, Chen Z, Tian L, Jiang G, Chen F, et al. m(6)A-induced lncRNA RP11 triggers the dissemination of colorectal cancer cells via upregulation of Zeb1. Mol Cancer. 2019;18(1):87.
Jia X, Wang Z, Qiu L, Yang Y, Wang Y, Chen Z, et al. Upregulation of LncRNA-HIT promotes migration and invasion of non-small cell lung cancer cells by association with ZEB1. Cancer Med. 2016;5(12):3555–63.
Liang WC, Fu WM, Wong CW, Wang Y, Wang WM, Hu GX, et al. The lncRNA H19 promotes epithelial to mesenchymal transition by functioning as miRNA sponges in colorectal cancer. Oncotarget. 2015;6(26):22513–25.
Gao H, Li X, Zhan G, Zhu Y, Yu J, Wang J, et al. Long noncoding RNA MAGI1-IT1 promoted invasion and metastasis of epithelial ovarian cancer via the miR-200a/ZEB axis. Cell Cycle (Georgetown, Tex). 2019;18(12):1393–406.
Chen DL, Chen LZ, Lu YX, Zhang DS, Zeng ZL, Pan ZZ, et al. Long noncoding RNA XIST expedites metastasis and modulates epithelial-mesenchymal transition in colorectal cancer. Cell Death Dis. 2017;8(8):e3011.
Tang J, Yu B, Li Y, Zhang W, Alvarez AA, Hu B, et al. TGF-β-activated lncRNA LINC00115 is a critical regulator of glioma stem-like cell tumorigenicity. EMBO Rep. 2019;20(12):e48170.
Grelet S, Link LA, Howley B, Obellianne C, Palanisamy V, Gangaraju VK, et al. A regulated PNUTS mRNA to lncRNA splice switch mediates EMT and tumour progression. Nat Cell Biol. 2017;19(9):1105–15.
Liang H, Yu T, Han Y, Jiang H, Wang C, You T, et al. LncRNA PTAR promotes EMT and invasion-metastasis in serous ovarian cancer by competitively binding miR-101-3p to regulate ZEB1 expression. Mol Cancer. 2018;17(1):119.
Cheng Y, Chang Q, Zheng B, Xu J, Li H, Wang R. LncRNA XIST promotes the epithelial to mesenchymal transition of retinoblastoma via sponging miR-101. Eur J Pharmacol. 2019;843:210–6.
Li T, Xie J, Shen C, Cheng D, Shi Y, Wu Z, et al. Amplification of long noncoding RNA ZFAS1 promotes metastasis in hepatocellular carcinoma. Cancer Res. 2015;75(15):3181–91.
Zhang Z, Liu F, Yang F, Liu Y. Kockdown of OIP5-AS1 expression inhibits proliferation, metastasis and EMT progress in hepatoblastoma cells through up-regulating miR-186a-5p and down-regulating ZEB1. Biomed Pharmacother. 2018;101:14–23.
Zhu C, Cheng D, Qiu X, Zhuang M, Liu Z. Long noncoding RNA SNHG16 promotes cell proliferation by sponging microRNA-205 and upregulating ZEB1 expression in osteosarcoma. Cell Physiol Biochem. 2018;51(1):429–40.
Liu C, Lin J. Long noncoding RNA ZEB1-AS1 acts AS an oncogene in osteosarcoma by epigenetically activating ZEB1. Am J Transl Res. 2016;8(10):4095–105.
Zhao J, Wu J, Qin Y, Zhang W, Huang G, Qin L. LncRNA PVT1 induces aggressive vasculogenic mimicry formation through activating the STAT3/Slug axis and epithelial-to-mesenchymal transition in gastric cancer. Cell Oncol. 2020;43(5):863–76.
Li GY, Wang W, Sun JY, Xin B, Zhang X, Wang T, et al. Long non-coding RNAs AC026904.1 and UCA1: a “one-two punch” for TGF-β-induced SNAI2 activation and epithelial-mesenchymal transition in breast cancer. Theranostics. 2018;8(10):2846–61.
Tian Y, Ma R, Sun Y, Liu H, Zhang H, Sun Y, et al. SP1-activated long noncoding RNA lncRNA GCMA functions as a competing endogenous RNA to promote tumor metastasis by sponging miR-124 and miR-34a in gastric cancer. Oncogene. 2020;39(25):4854–68.
Dong H, Hu J, Zou K, Ye M, Chen Y, Wu C, et al. Activation of LncRNA TINCR by H3K27 acetylation promotes Trastuzumab resistance and epithelial-mesenchymal transition by targeting MicroRNA-125b in breast cancer. Mol Cancer. 2019;18(1):3.
Jiang H, Li T, Qu Y, Wang X, Li B, Song J, et al. Long non-coding RNA SNHG15 interacts with and stabilizes transcription factor Slug and promotes colon cancer progression. Cancer Lett. 2018;425:78–87.
Yan X, Zhang D, Wu W, Wu S, Qian J, Hao Y, et al. Mesenchymal stem cells promote hepatocarcinogenesis via lncRNA-MUF interaction with ANXA2 and miR-34a. Cancer Res. 2017;77(23):6704–16.
Zhang Y, Yuan Y, Zhang Y, Cheng L, Zhou X, Chen K. SNHG7 accelerates cell migration and invasion through regulating miR-34a-Snail-EMT axis in gastric cancer. Cell Cycle (Georgetown, Tex). 2020;19(1):142–52.
Wang YQ, Jiang DM, Hu SS, Zhao L, Wang L, Yang MH, et al. SATB2-AS1 suppresses colorectal carcinoma aggressiveness by inhibiting SATB2-dependent snail transcription and epithelial-mesenchymal transition. Cancer Res. 2019;79(14):3542–56.
Pan J, Fang S, Tian H, Zhou C, Zhao X, Tian H, et al. lncRNA JPX/miR-33a-5p/Twist1 axis regulates tumorigenesis and metastasis of lung cancer by activating Wnt/β-catenin signaling. Mol Cancer. 2020;19(1):9.
Xu L, Wei B, Hui H, Sun Y, Liu Y, Yu X, et al. Positive feedback loop of lncRNA LINC01296/miR-598/Twist1 promotes non-small cell lung cancer tumorigenesis. J Cell Physiol. 2019;234(4):4563–71.
Wu H, Hu Y, Liu X, Song W, Gong P, Zhang K, et al. LncRNA TRERNA1 function as an enhancer of SNAI1 promotes gastric cancer metastasis by regulating epithelial-mesenchymal transition. Mol Ther Nucleic Acids. 2017;8:291–9.
Li P, Zhang X, Wang H, Wang L, Liu T, Du L, et al. MALAT1 is associated with poor response to Oxaliplatin-based chemotherapy in colorectal cancer patients and promotes chemoresistance through EZH2. Mol Cancer Ther. 2017;16(4):739–51.
Cai Q, Jin L, Wang S, Zhou D, Wang J, Tang Z, et al. Long non-coding RNA UCA1 promotes gallbladder cancer progression by epigenetically repressing p21 and E-cadherin expression. Oncotarget. 2017;8(29):47957–68.
Terashima M, Tange S, Ishimura A, Suzuki T. MEG3 long noncoding RNA contributes to the epigenetic regulation of epithelial-mesenchymal transition in lung cancer cell lines. J Biol Chem. 2017;292(1):82–99.
Dong Z, Li S, Wu X, Niu Y, Liang X, Yang L, et al. Aberrant hypermethylation-mediated downregulation of antisense lncRNA ZNF667-AS1 and its sense gene ZNF667 correlate with progression and prognosis of esophageal squamous cell carcinoma. Cell Death Dis. 2019;10(12):930.
Wang B, Zhao L, Chi W, Cao H, Cui W, Meng W. Aberrant methylation-mediated downregulation of lncRNA SSTR5-AS1 promotes progression and metastasis of laryngeal squamous cell carcinoma. Epigenetics Chromatin. 2019;12(1):35.
Liu J, Liu L, Wan JX, Song Y. Long noncoding RNA SNHG20 promotes gastric cancer progression by inhibiting p21 expression and regulating the GSK-3β/ β-catenin signaling pathway. Oncotarget. 2017;8(46):80700–8.
Wu L, Zhu L, Li Y, Zheng Z, Lin X, Yang C. LncRNA MEG3 promotes melanoma growth, metastasis and formation through modulating miR-21/E-cadherin axis. Cancer Cell Int. 2020;20:12.
Zhang W, Shi S, Jiang J, Li X, Lu H, Ren F. LncRNA MEG3 inhibits cell epithelial-mesenchymal transition by sponging miR-421 targeting E-cadherin in breast cancer. Biomed Pharmacother. 2017;91:312–9.
Li W, Zhang Z, Liu X, Cheng X, Zhang Y, Han X, et al. The FOXN3-NEAT1-SIN3A repressor complex promotes progression of hormonally responsive breast cancer. J Clin Invest. 2017;127(9):3421–40.
Pan Y, Tong S, Cui R, Fan J, Liu C, Lin Y, et al. Long non-coding MALAT1 functions as a competing endogenous RNA to regulate vimentin expression by sponging miR-30a-5p in hepatocellular carcinoma. Cell Physiol Biochem. 2018;50(1):108–20.
Wang TH, Lin YS, Chen Y, Yeh CT, Huang YL, Hsieh TH, et al. Long non-coding RNA AOC4P suppresses hepatocellular carcinoma metastasis by enhancing vimentin degradation and inhibiting epithelial-mesenchymal transition. Oncotarget. 2015;6(27):23342–57.
Zeng S, Xie X, Xiao YF, Tang B, Hu CJ, Wang SM, et al. Long noncoding RNA LINC00675 enhances phosphorylation of vimentin on Ser83 to suppress gastric cancer progression. Cancer Lett. 2018;412:179–87.
Yang Y, Zhang J, Chen X, Xu X, Cao G, Li H, et al. LncRNA FTX sponges miR-215 and inhibits phosphorylation of vimentin for promoting colorectal cancer progression. Gene Ther. 2018;25(5):321–30.
Lei K, Liang X, Gao Y, Xu B, Xu Y, Li Y, et al. Lnc-ATB contributes to gastric cancer growth through a MiR-141-3p/TGFβ2 feedback loop. Biochem Biophys Res Commun. 2017;484(3):514–21.
Sun J, Zhang Y. LncRNA XIST enhanced TGF-β2 expression by targeting miR-141-3p to promote pancreatic cancer cells invasion. Biosci Rep. 2019;39(7):BSR20190332.
Huang WJ, Tian XP, Bi SX, Zhang SR, He TS, Song LY, et al. The β-catenin/TCF-4-LINC01278-miR-1258-Smad2/3 axis promotes hepatocellular carcinoma metastasis. Oncogene. 2020;39(23):4538–50.
Chen X, Xiong D, Yang H, Ye L, Mei S, Wu J, et al. Long noncoding RNA OPA-interacting protein 5 antisense transcript 1 upregulated SMAD3 expression to contribute to metastasis of cervical cancer by sponging miR-143-3p. J Cell Physiol. 2019;234(4):5264–75.
Xu J, Shao T, Song M, Xie Y, Zhou J, Yin J, et al. MIR22HG acts as a tumor suppressor via TGFβ/SMAD signaling and facilitates immunotherapy in colorectal cancer. Mol Cancer. 2020;19(1):51.
Papoutsoglou P, Tsubakihara Y, Caja L, Morén A, Pallis P, Ameur A, et al. The TGFB2-AS1 lncRNA regulates TGF-β signaling by modulating corepressor activity. Cell Rep. 2019;28(12):3182–98.e11.
Zhang J, Han C, Song K, Chen W, Ungerleider N, Yao L, et al. The long-noncoding RNA MALAT1 regulates TGF-β/Smad signaling through formation of a lncRNA-protein complex with Smads, SETD2 and PPM1A in hepatic cells. PLoS One. 2020;15(1):e0228160.
Li ZT, Zhang X, Wang DW, Xu J, Kou KJ, Wang ZW, et al. Overexpressed lncRNA GATA6-AS1 inhibits LNM and EMT via FZD4 through the Wnt/β-catenin signaling pathway in GC. Mol Ther Nucleic Acids. 2020;19:827–40.
You BH, Yoon JH, Kang H, Lee EK, Lee SK, Nam JW. HERES, a lncRNA that regulates canonical and noncanonical Wnt signaling pathways via interaction with EZH2. Proc Natl Acad Sci U S A. 2019;116(49):24620–9.
Chen Q, Cai J, Wang Q, Wang Y, Liu M, Yang J, et al. Long noncoding RNA NEAT1, regulated by the EGFR pathway, contributes to glioblastoma progression through the WNT/β-catenin pathway by scaffolding EZH2. Clin Cancer Res. 2018;24(3):684–95.
Yoon JH, Abdelmohsen K, Srikantan S, Yang X, Martindale JL, De S, et al. LincRNA-p21 suppresses target mRNA translation. Mol Cell. 2012;47(4):648–55.
Lu Y, Zhao X, Liu Q, Li C, Graves-Deal R, Cao Z, et al. lncRNA MIR100HG-derived miR-100 and miR-125b mediate cetuximab resistance via Wnt/β-catenin signaling. Nat Med. 2017;23(11):1331–41.
Yue B, Liu C, Sun H, Liu M, Song C, Cui R, et al. A positive feed-forward loop between LncRNA-CYTOR and Wnt/β-catenin signaling promotes metastasis of colon cancer. Mol Ther. 2018;26(5):1287–98.
Tang T, Guo C, Xia T, Zhang R, Zen K, Pan Y, et al. LncCCAT1 promotes breast cancer stem cell function through activating WNT/β-catenin signaling. Theranostics. 2019;9(24):7384–402.
Li Y, Guo D, Zhao Y, Ren M, Lu G, Wang Y, et al. Long non-coding RNA SNHG5 promotes human hepatocellular carcinoma progression by regulating miR-26a-5p/GSK3β signal pathway. Cell Death Dis. 2018;9(9):888.
Shao Q, Xu J, Deng R, Wei W, Zhou B, Yue C, et al. SNHG 6 promotes the progression of colon and rectal adenocarcinoma via miR-101-3p and Wnt/β-catenin signaling pathway. BMC Gastroenterol. 2019;19(1):163.
Xue M, Pang H, Li X, Li H, Pan J, Chen W. Long non-coding RNA urothelial cancer-associated 1 promotes bladder cancer cell migration and invasion by way of the hsa-miR-145-ZEB1/2-FSCN1 pathway. Cancer Sci. 2016;107(1):18–27.
Wu Y, Zhao Y, Huan L, Zhao J, Zhou Y, Xu L, et al. An LTR retrotransposon-derived long noncoding RNA lncMER52A promotes hepatocellular carcinoma progression by binding p120-catenin. Cancer Res. 2020;80(5):976–87.
Zhou Y, Fan RG, Qin CL, Jia J, Wu XD, Zha WZ. LncRNA-H19 activates CDC42/PAK1 pathway to promote cell proliferation, migration and invasion by targeting miR-15b in hepatocellular carcinoma. Genomics. 2019;111(6):1862–72.
Chou J, Wang B, Zheng T, Li X, Zheng L, Hu J, et al. MALAT1 induced migration and invasion of human breast cancer cells by competitively binding miR-1 with cdc42. Biochem Biophys Res Commun. 2016;472(1):262–9.
Yang J, Qiu Q, Qian X, Yi J, Jiao Y, Yu M, et al. Long noncoding RNA LCAT1 functions as a ceRNA to regulate RAC1 function by sponging miR-4715-5p in lung cancer. Mol Cancer. 2019;18(1):171.
Li H, Wang X, Wen C, Huo Z, Wang W, Zhan Q, et al. Long noncoding RNA NORAD, a novel competing endogenous RNA, enhances the hypoxia-induced epithelial-mesenchymal transition to promote metastasis in pancreatic cancer. Mol Cancer. 2017;16(1):169.
Yu X, Wang D, Wang X, Sun S, Zhang Y, Wang S, et al. CXCL12/CXCR4 promotes inflammation-driven colorectal cancer progression through activation of RhoA signaling by sponging miR-133a-3p. J Exp Clin Cancer Res. 2019;38(1):32.
Zong W, Feng W, Jiang Y, Cao Y, Ke Y, Shi X, et al. LncRNA CTC-497E21.4 promotes the progression of gastric cancer via modulating miR-22/NET1 axis through RhoA signaling pathway. Gastric Cancer. 2020;23(2):228–40.
Yang G, Song R, Wang L, Wu X. Knockdown of long non-coding RNA TP73-AS1 inhibits osteosarcoma cell proliferation and invasion through sponging miR-142. Biomed Pharmacother. 2018;103:1238–45.
Li J, Guo W, Xue W, Xu P, Deng Z, Zhang D, et al. Long noncoding RNA AURKAPS1 potentiates malignant hepatocellular carcinoma progression by regulating miR-142, miR-155 and miR-182. Sci Rep. 2019;9(1):19645.
Zheng X, Tang H, Zhao X, Sun Y, Jiang Y, Liu Y. Long non-coding RNA FTH1P3 facilitates uveal melanoma cell growth and invasion through miR-224-5p. PLoS One. 2017;12(11):e0184746.
Traversa D, Simonetti G, Tolomeo D, Visci G, Macchia G, Ghetti M, et al. Unravelling similarities and differences in the role of circular and linear PVT1 in cancer and human disease. Br J Cancer. 2022;126(6):835–50.
Battistelli C, Cicchini C, Santangelo L, Tramontano A, Grassi L, Gonzalez FJ, et al. The Snail repressor recruits EZH2 to specific genomic sites through the enrollment of the lncRNA HOTAIR in epithelial-to-mesenchymal transition. Oncogene. 2017;36(7):942–55.
Li Y, Cheng C. Long noncoding RNA NEAT1 promotes the metastasis of osteosarcoma via interaction with the G9a-DNMT1-Snail complex. Am J Cancer Res. 2018;8(1):81–90.
Jiang Y, Cao W, Wu K, Qin X, Wang X, Li Y, et al. LncRNA LINC00460 promotes EMT in head and neck squamous cell carcinoma by facilitating peroxiredoxin-1 into the nucleus. J Exp Clin Cancer Res. 2019;38(1):365.
Sakai S, Ohhata T, Kitagawa K, Uchida C, Aoshima T, Niida H, et al. Long noncoding RNA ELIT-1 acts as a Smad3 cofactor to facilitate TGFβ/Smad signaling and promote epithelial-mesenchymal transition. Cancer Res. 2019;79(11):2821–38.
Di Cecilia S, Zhang F, Sancho A, Li S, Aguiló F, Sun Y, et al. RBM5-AS1 is critical for self-renewal of colon cancer stem-like cells. Cancer Res. 2016;76(19):5615–27.
Zhang M, Weng W, Zhang Q, Wu Y, Ni S, Tan C, et al. The lncRNA NEAT1 activates Wnt/β-catenin signaling and promotes colorectal cancer progression via interacting with DDX5. J Hematol Oncol. 2018;11(1):113.
Wu DD, Chen X, Sun KX, Wang LL, Chen S, Zhao Y. Role of the lncRNA ABHD11-AS(1) in the tumorigenesis and progression of epithelial ovarian cancer through targeted regulation of RhoC. Mol Cancer. 2017;16(1):138.
Shi D, Wu F, Mu S, Hu B, Zhong B, Gao F, et al. LncRNA AFAP1-AS1 promotes tumorigenesis and epithelial-mesenchymal transition of osteosarcoma through RhoC/ROCK1/p38MAPK/Twist1 signaling pathway. J Exp Clin Cancer Res. 2019;38(1):375.
Conn SJ, Pillman KA, Toubia J, Conn VM, Salmanidis M, Phillips CA, et al. The RNA binding protein quaking regulates formation of circRNAs. Cell. 2015;160(6):1125–34.
Chen X, Chen RX, Wei WS, Li YH, Feng ZH, Tan L, et al. PRMT5 circular RNA promotes metastasis of urothelial carcinoma of the bladder through sponging miR-30c to induce epithelial-mesenchymal transition. Clin Cancer Res. 2018;24(24):6319–30.
Luan W, Shi Y, Zhou Z, Xia Y, Wang J. circRNA_0084043 promote malignant melanoma progression via miR-153-3p/Snail axis. Biochem Biophys Res Commun. 2018;502(1):22–9.
Guo J, Duan H, Li Y, Yang L, Yuan L. A novel circular RNA circ-ZNF652 promotes hepatocellular carcinoma metastasis through inducing snail-mediated epithelial-mesenchymal transition by sponging miR-203/miR-502-5p. Biochem Biophys Res Commun. 2019;513(4):812–9.
Lin S, Song S, Sun R, Zhang M, Du Y, Zhang D, et al. Oncogenic circular RNA Hsa-circ-000684 interacts with microRNA-186 to upregulate ZEB1 in gastric cancer. FASEB J. 2020;34(6):8187–203.
Zhou B, Zheng P, Li Z, Li H, Wang X, Shi Z, et al. CircPCNXL2 sponges miR-153 to promote the proliferation and invasion of renal cancer cells through upregulating ZEB2. Cell Cycle (Georgetown, Tex). 2018;17(23):2644–54.
Meng J, Chen S, Han JX, Qian B, Wang XR, Zhong WL, et al. Twist1 regulates vimentin through Cul2 circular RNA to promote EMT in hepatocellular carcinoma. Cancer Res. 2018;78(15):4150–62.
Xue D, Wang H, Chen Y, Shen D, Lu J, Wang M, et al. Circ-AKT3 inhibits clear cell renal cell carcinoma metastasis via altering miR-296-3p/E-cadherin signals. Mol Cancer. 2019;18(1):151.
Wei S, Zheng Y, Jiang Y, Li X, Geng J, Shen Y, et al. The circRNA circPTPRA suppresses epithelial-mesenchymal transitioning and metastasis of NSCLC cells by sponging miR-96-5p. EBioMedicine. 2019;44:182–93.
Lock FE, Underhill-Day N, Dunwell T, Matallanas D, Cooper W, Hesson L, et al. The RASSF8 candidate tumor suppressor inhibits cell growth and regulates the Wnt and NF-kappaB signaling pathways. Oncogene. 2010;29(30):4307–16.
Yang Z, Qu CB, Zhang Y, Zhang WF, Wang DD, Gao CC, et al. Dysregulation of p53-RBM25-mediated circAMOTL1L biogenesis contributes to prostate cancer progression through the circAMOTL1L-miR-193a-5p-Pcdha pathway. Oncogene. 2019;38(14):2516–32.
Yang H, Li X, Meng Q, Sun H, Wu S, Hu W, et al. CircPTK2 (hsa_circ_0005273) as a novel therapeutic target for metastatic colorectal cancer. Mol Cancer. 2020;19(1):13.
Wang L, Tong X, Zhou Z, Wang S, Lei Z, Zhang T, et al. Circular RNA hsa_circ_0008305 (circPTK2) inhibits TGF-β-induced epithelial-mesenchymal transition and metastasis by controlling TIF1γ in non-small cell lung cancer. Mol Cancer. 2018;17(1):140.
He W, Dorn DC, Erdjument-Bromage H, Tempst P, Moore MA, Massagué J. Hematopoiesis controlled by distinct TIF1gamma and Smad4 branches of the TGFbeta pathway. Cell. 2006;125(5):929–41.
Dupont S, Zacchigna L, Cordenonsi M, Soligo S, Adorno M, Rugge M, et al. Germ-layer specification and control of cell growth by Ectodermin, a Smad4 ubiquitin ligase. Cell. 2005;121(1):87–99.
Zeng K, He B, Yang BB, Xu T, Chen X, Xu M, et al. The pro-metastasis effect of circANKS1B in breast cancer. Mol Cancer. 2018;17(1):160.
Zhao W, Cui Y, Liu L, Qi X, Liu J, Ma S, et al. Splicing factor derived circular RNA circUHRF1 accelerates oral squamous cell carcinoma tumorigenesis via feedback loop. Cell Death Differ. 2020;27(3):919–33.
Geng Y, Zheng X, Hu W, Wang Q, Xu Y, He W, et al. Hsa_circ_0009361 acts as the sponge of miR-582 to suppress colorectal cancer progression by regulating APC2 expression. Clin Sci (London, England: 1979). 2019;133(10):1197–213.
Hu X, Wu D, He X, Zhao H, He Z, Lin J, et al. circGSK3β promotes metastasis in esophageal squamous cell carcinoma by augmenting β-catenin signaling. Mol Cancer. 2019;18(1):160.
Liang WC, Wong CW, Liang PP, Shi M, Cao Y, Rao ST, et al. Translation of the circular RNA circβ-catenin promotes liver cancer cell growth through activation of the Wnt pathway. Genome Biol. 2019;20(1):84.
Yang F, Fang E, Mei H, Chen Y, Li H, Li D, et al. Cis-acting circ-CTNNB1 promotes β-catenin signaling and cancer progression via DDX3-mediated transactivation of YY1. Cancer Res. 2019;79(3):557–71.
Zhong Z, Huang M, Lv M, He Y, Duan C, Zhang L, et al. Circular RNA MYLK as a competing endogenous RNA promotes bladder cancer progression through modulating VEGFA/VEGFR2 signaling pathway. Cancer Lett. 2017;403:305–17.
Wang K, Sun Y, Tao W, Fei X, Chang C. Androgen receptor (AR) promotes clear cell renal cell carcinoma (ccRCC) migration and invasion via altering the circHIAT1/miR-195-5p/29a-3p/29c-3p/CDC42 signals. Cancer Lett. 2017;394:1–12.
Li L, Li W, Chen N, Zhao H, Xu G, Zhao Y, et al. FLI1 exonic circular RNAs as a novel oncogenic driver to promote tumor metastasis in small cell lung cancer. Clin Cancer Res. 2019;25(4):1302–17.
Yang H, Zhang H, Yang Y, Wang X, Deng T, Liu R, et al. Hypoxia induced exosomal circRNA promotes metastasis of colorectal cancer via targeting GEF-H1/RhoA axis. Theranostics. 2020;10(18):8211–26.
Walkley CR, Li JB. Rewriting the transcriptome: adenosine-to-inosine RNA editing by ADARs. Genome Biol. 2017;18(1):205.
Nishikura K. A-to-I editing of coding and non-coding RNAs by ADARs. Nat Rev Mol Cell Biol. 2016;17(2):83–96.
Kim DD, Kim TT, Walsh T, Kobayashi Y, Matise TC, Buyske S, et al. Widespread RNA editing of embedded alu elements in the human transcriptome. Genome Res. 2004;14(9):1719–25.
Daniel C, Lagergren J, Öhman M. RNA editing of non-coding RNA and its role in gene regulation. Biochimie. 2015;117:22–7.
Wang Y, Xu X, Yu S, Jeong KJ, Zhou Z, Han L, et al. Systematic characterization of A-to-I RNA editing hotspots in microRNAs across human cancers. Genome Res. 2017;27(7):1112–25.
Ramírez-Moya J, Baker AR, Slack FJ, Santisteban P. ADAR1-mediated RNA editing is a novel oncogenic process in thyroid cancer and regulates miR-200 activity. Oncogene. 2020;39(18):3738–53.
Liu X, Fu Y, Huang J, Wu M, Zhang Z, Xu R, et al. ADAR1 promotes the epithelial-to-mesenchymal transition and stem-like cell phenotype of oral cancer by facilitating oncogenic microRNA maturation. J Exp Clin Cancer Res. 2019;38(1):315.
Shen H, An O, Ren X, Song Y, Tang SJ, Ke XY, et al. ADARs act as potent regulators of circular transcriptome in cancer. Nat Commun. 2022;13(1):1508.
Elkon R, Ugalde AP, Agami R. Alternative cleavage and polyadenylation: extent, regulation and function. Nat Rev Genet. 2013;14(7):496–506.
Mayr C, Bartel DP. Widespread shortening of 3’UTRs by alternative cleavage and polyadenylation activates oncogenes in cancer cells. Cell. 2009;138(4):673–84.
Lembo A, Di Cunto F, Provero P. Shortening of 3’UTRs correlates with poor prognosis in breast and lung cancer. PLoS One. 2012;7(2):e31129.
Passacantilli I, Panzeri V, Bielli P, Farini D, Pilozzi E, Fave GD, et al. Alternative polyadenylation of ZEB1 promotes its translation during genotoxic stress in pancreatic cancer cells. Cell Death Dis. 2017;8(11):e3168.
Nairismägi ML, Vislovukh A, Meng Q, Kratassiouk G, Beldiman C, Petretich M, et al. Translational control of TWIST1 expression in MCF-10A cell lines recapitulating breast cancer progression. Oncogene. 2012;31(47):4960–6.
Chen X, Zhang JX, Luo JH, Wu S, Yuan GJ, Ma NF, et al. CSTF2-induced shortening of the RAC1 3’UTR promotes the pathogenesis of urothelial carcinoma of the bladder. Cancer Res. 2018;78(20):5848–62.
Li YQ, Chen Y, Xu YF, He QM, Yang XJ, Li YQ, et al. FNDC3B 3′-UTR shortening escapes from microRNA-mediated gene repression and promotes nasopharyngeal carcinoma progression. Cancer Sci. 2020;111(6):1991–2003.
Hansen EB, Fredsøe J, Okholm TLH, Ulhøi BP, Klingenberg S, Jensen JB, et al. The transcriptional landscape and biomarker potential of circular RNAs in prostate cancer. Genome Med. 2022;14(1):8.
Li P, Xu Z, Liu T, Liu Q, Zhou H, Meng S, et al. Circular RNA sequencing reveals serum exosome circular RNA panel for high-grade astrocytoma diagnosis. Clin Chem. 2022;68(2):332–43.
Vo JN, Cieslik M, Zhang Y, Shukla S, Xiao L, Zhang Y, et al. The landscape of circular RNA in cancer. Cell. 2019;176(4):869–81.e13.
Rezaeian AH, Khanbabaei H, Calin GA. Therapeutic potential of the miRNA-ATM axis in the management of tumor radioresistance. Cancer Res. 2020;80(2):139–50.
Rupaimoole R, Slack FJ. MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat Rev Drug Discov. 2017;16(3):203–22.
van Zandwijk N, Pavlakis N, Kao SC, Linton A, Boyer MJ, Clarke S, et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: a first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol. 2017;18(10):1386–96.
Seto AG, Beatty X, Lynch JM, Hermreck M, Tetzlaff M, Duvic M, et al. Cobomarsen, an oligonucleotide inhibitor of miR-155, co-ordinately regulates multiple survival pathways to reduce cellular proliferation and survival in cutaneous T-cell lymphoma. Br J Haematol. 2018;183(3):428–44.
Hong DS, Kang YK, Borad M, Sachdev J, Ejadi S, Lim HY, et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br J Cancer. 2020;122(11):1630–7.
Kasinski AL, Kelnar K, Stahlhut C, Orellana E, Zhao J, Shimer E, et al. A combinatorial microRNA therapeutics approach to suppressing non-small cell lung cancer. Oncogene. 2015;34(27):3547–55.
Shao S, Hu Q, Wu W, Wang M, Huang J, Zhao X, et al. Tumor-triggered personalized microRNA cocktail therapy for hepatocellular carcinoma. Biomater Sci. 2020;8(23):6579–91.
Mai D, Ding P, Tan L, Zhang J, Pan Z, Bai R, et al. PIWI-interacting RNA-54265 is oncogenic and a potential therapeutic target in colorectal adenocarcinoma. Theranostics. 2018;8(19):5213–30.
Cui L, Nakano K, Obchoei S, Setoguchi K, Matsumoto M, Yamamoto T, et al. Small nucleolar noncoding RNA SNORA23, up-regulated in human pancreatic ductal adenocarcinoma, regulates expression of spectrin repeat-containing nuclear envelope 2 to promote growth and metastasis of xenograft tumors in mice. Gastroenterology. 2017;153(1):292–306.e2.
Kim HK, Fuchs G, Wang S, Wei W, Zhang Y, Park H, et al. A transfer-RNA-derived small RNA regulates ribosome biogenesis. Nature. 2017;552(7683):57–62.
Zhang Y, Pitchiaya S, Cieślik M, Niknafs YS, Tien JC, Hosono Y, et al. Analysis of the androgen receptor-regulated lncRNA landscape identifies a role for ARLNC1 in prostate cancer progression. Nat Genet. 2018;50(6):814–24.
Hu Q, Ye Y, Chan LC, Li Y, Liang K, Lin A, et al. Oncogenic lncRNA downregulates cancer cell antigen presentation and intrinsic tumor suppression. Nat Immunol. 2019;20(7):835–51.
Leucci E, Vendramin R, Spinazzi M, Laurette P, Fiers M, Wouters J, et al. Melanoma addiction to the long non-coding RNA SAMMSON. Nature. 2016;531(7595):518–22.
Kim J, Piao HL, Kim BJ, Yao F, Han Z, Wang Y, et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat Genet. 2018;50(12):1705–15.
Gutschner T, Hämmerle M, Eissmann M, Hsu J, Kim Y, Hung G, et al. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 2013;73(3):1180–9.
Arun G, Diermeier S, Akerman M, Chang KC, Wilkinson JE, Hearn S, et al. Differentiation of mammary tumors and reduction in metastasis upon Malat1 lncRNA loss. Genes Dev. 2016;30(1):34–51.
Chen Y, Yang F, Fang E, Xiao W, Mei H, Li H, et al. Circular RNA circAGO2 drives cancer progression through facilitating HuR-repressed functions of AGO2-miRNA complexes. Cell Death Differ. 2019;26(7):1346–64.
Han K, Wang FW, Cao CH, Ling H, Chen JW, Chen RX, et al. CircLONP2 enhances colorectal carcinoma invasion and metastasis through modulating the maturation and exosomal dissemination of microRNA-17. Mol Cancer. 2020;19(1):60.
Adams BD, Parsons C, Walker L, Zhang WC, Slack FJ. Targeting noncoding RNAs in disease. J Clin Invest. 2017;127(3):761–71.
Nieto MA, Huang RY, Jackson RA, Thiery JP. EMT: 2016. Cell. 2016;166(1):21–45.
Pastushenko I, Brisebarre A, Sifrim A, Fioramonti M, Revenco T, Boumahdi S, et al. Identification of the tumour transition states occurring during EMT. Nature. 2018;556(7702):463–8.
Yu M, Bardia A, Wittner BS, Stott SL, Smas ME, Ting DT, et al. Circulating breast tumor cells exhibit dynamic changes in epithelial and mesenchymal composition. Science (New York, NY). 2013;339(6119):580–4.
Lu M, Jolly MK, Levine H, Onuchic JN, Ben-Jacob E. MicroRNA-based regulation of epithelial-hybrid-mesenchymal fate determination. Proc Natl Acad Sci U S A. 2013;110(45):18144–9.
Tian XJ, Zhang H, Xing J. Coupled reversible and irreversible bistable switches underlying TGFβ-induced epithelial to mesenchymal transition. Biophys J. 2013;105(4):1079–89.
Zhang J, Tian XJ, Zhang H, Teng Y, Li R, Bai F, et al. TGF-β-induced epithelial-to-mesenchymal transition proceeds through stepwise activation of multiple feedback loops. Sci Signal. 2014;7(345):ra91.
Dang TT, Esparza MA, Maine EA, Westcott JM, Pearson GW. ΔNp63α promotes breast cancer cell motility through the selective activation of components of the epithelial-to-mesenchymal transition program. Cancer Res. 2015;75(18):3925–35.
Shibue T, Weinberg RA. EMT, CSCs, and drug resistance: the mechanistic link and clinical implications. Nat Rev Clin Oncol. 2017;14(10):611–29.
Li HM, Ma XL, Li HG. Intriguing circles: conflicts and controversies in circular RNA research. Wiley Interdiscip Rev RNA. 2019;10(5):e1538.
Acknowledgements
We apologize to the many investigators whose important studies could not be cited here because of space limitations.
Funding
Financial support was granted by The Lundbeck Foundation to LSK (R307–2018-3433).
Author information
Authors and Affiliations
Contributions
HK and LSK contributed to the direction and guidance of this review. SE and ZG drafted the manuscript and prepared the figures. JLGR, HP, MM, LSK and HK made the tables and revised the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Khanbabaei, H., Ebrahimi, S., García-Rodríguez, J.L. et al. Non-coding RNAs and epithelial mesenchymal transition in cancer: molecular mechanisms and clinical implications. J Exp Clin Cancer Res 41, 278 (2022). https://doi.org/10.1186/s13046-022-02488-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13046-022-02488-x