
Abstract
Background
Endophytic microorganisms are a rich source of bioactive natural products. They are considered as promising biofertilizers and biocontrol agents due to their growth-promoting interactions with the host plants and their bioactive secondary metabolites that can help manage plant pathogens. Identification of new endophytes may lead to the discovery of novel molecules or provide new strains for production of valuable compounds.
Results
In this study, we isolated an endophytic bacterium from the leaves of Taxus chinensis, which was identified as Pseudomonas sp. 102515 based on the 16S rRNA gene sequence and physiological characteristics. Analysis of its secondary metabolites revealed that this endophytic strain produces a major product zeaxanthin diglucoside, a promising antioxidant natural product that belongs to the family of carotenoids. A carotenoid (Pscrt) biosynthetic gene cluster was amplified from this strain, and the functions of PsCrtI and PsCrtY in the biosynthesis of zeaxanthin diglucoside were characterized in Escherichia coli BL21(DE3). The entire Pscrt biosynthetic gene cluster was successfully reconstituted in E. coli BL21(DE3) and Pseudomonas putida KT2440. The production of zeaxanthin diglucoside in Pseudomonas sp. 102515 was improved through the optimization of fermentation conditions such as medium, cultivation temperature and culture time. The highest yield under the optimized conditions reached 206 mg/L. The engineered strain of P. putida KT2440 produced zeaxanthin diglucoside at 121 mg/L in SOC medium supplemented with 0.5% glycerol at 18 °C, while the yield of zeaxanthin diglucoside in E. coli BL21(DE3) was only 2 mg/L. To further enhance the production, we introduced an expression plasmid harboring the Pscrt biosynthetic gene cluster into Pseudomonas sp. 102515. The yield in this engineered strain reached 380 mg/L, 85% higher than the wild type. Through PCR, we also discovered the presence of a turnerbactin biosynthetic gene cluster in Pseudomonas sp. 102515. Because turnerbactin is involved in nitrogen fixation, this endophytic strain might have a role in promoting growth of the host plant.
Conclusions
We isolated and identified an endophytic strain of Pseudomonas from T. chinensis. A zeaxanthin diglucoside biosynthetic gene cluster was discovered and characterized in this bacterium. Through fermentation and genetic engineering, the engineered strain produced zeaxanthin diglucoside at 380 ± 12 mg/L, representing a promising strain for the production of this antioxidant natural product. Additionally, Pseudomonas sp. 102515 might also be utilized as a plant-promoting strain for agricultural applications.
Similar content being viewed by others

Background
Nature provides a huge repertoire of structurally and functionally diverse bioactive molecules. As such, natural products are a major source of new drugs. Microorganisms are known to produce many pharmaceuticals such as lovastatin (anti-cholesterol), penicillin (antibacterial) and vancomycin (antibacterial). There is an estimated 99.999% of microbial species left undiscovered in the world. Therefore, microorganisms remain an underexplored source of bioactive natural products. In particular, endophytic microorganisms hold huge potential for the discovery of natural products with pharmaceutical importance [1]. Endophytes, microorganisms (bacteria and fungi) that live in the tissues of living plants without causing any apparent disease symptoms in the host, are potential sources of novel natural products with applications in medicine, agriculture, and industry [2, 3]. They have been isolated from medicinal plants, weeds, as well as ornamental and fruit trees [1]. Endophytes enter plants through the roots and the aerial portions of plants, such as leaves, flowers, stems and cotyledons [4]. Upon entering the host, they reside within cells, the intercellular spaces or the vascular system [5]. Some endophytes exhibit plant growth-promoting effects and protect plants from biotic and abiotic stresses under different environmental conditions. Thus, they are considered as endosymbiotic microorganisms with potential agricultural applications as biofertilizers and biocontrol agents [5,6,7,8].
Another important aspect is that the endophytes are known to produce a wide variety of natural products. For example, ecomycins produced by Pseudomonas viridiflava are a family of novel lipopeptides and consist of some unusual amino acids including homoserine and β-hydroxy aspartic acid [9]. Another example is pseudomycins, which represent a group of antifungal peptide compounds isolated from Pseudomonas syringae, a plant-associated bacterium [10]. The new species can sometimes lead to the rediscovery of known natural products as happened in the example of paclitaxel (Taxol®). Taxol, the world’s first billion-dollar anticancer drug, was isolated from the Pacific yew tree, Taxus wallachiana. Interestingly, endophytic fungus (Pestalotiopsis microspora) isolated from this plant was reported to produce paclitaxel [11], representing a potential microbial host for the production of this pharmaceutically important molecule.
Carotenoids are isoprenoid pigments that are widely seen in nature. They are produced by all known phototrophic organisms and some non-phototrophic fungi, bacteria and archaea [12]. Carotenoids exhibit diverse biological functions in different organisms that either produce or consume carotenoids. They serve as accessory pigments in the light harvesting center of phototrophic organisms [13]. The primary roles of carotenoids in non-phototrophic organisms are membrane stability and relief of oxidative stress [14]. Additionally, carotenoids have been used in various applications, ranging from food colorants and feed supplements to nutritional and cosmetics purposes [15]. This family of natural products has shown a variety of biological activities, such as antioxidant, anticancer, and anti-inflammatory properties. They are also used as precursors for the synthesis of vitamin A and as nutritional factors for the prevention of chronic diseases [16, 17]. Due to the negative impacts of synthetic coloring agents on human health (e.g., toxicity, hyperallergenicity, and carcinogenicity) and the increase in the consumer demand for natural and health-promoting food ingredients, the utilization of carotenoids as colorants and supplements is rising in food, cosmetics, nutraceutical and pharmaceutical industries, with an expected global market value of $1.8 billion in 2019 [18, 19].
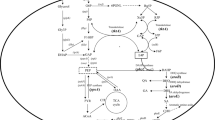
More than 750 different natural carotenoids have been identified. These molecules share a linear, conjugated chromophore backbone and a common biosynthetic pathway (Fig. 1). Carotenoid biosynthesis typically begins with the isomerization of isopenthyl diphosphate (IPP) from the mevalonate pathway to yield dimethylallyl diphosphate (DMAPP), catalyzed by the IPP isomerase (Idi). Geranyl diphosphate (GPP) is synthesized through the head-to-tail condensation of IPP and DMAPP. Addition of IPP to GPP generates farnesyl pyrophosphate (FPP) formation and a further IPP molecule yields geranylgeranyl diphosphate (GGPP) by geranylgeranyl diphosphate synthase (CrtE). Phytoene synthase (CrtB) catalyzes the head-to-head condensation of two GGPP to phytoene. Phytoene desaturase (CrtI) extends the double bond conjugation of phytoene to generate lycopene. Terminal β-cyclization catalyzed by lycopene β-cyclase (CrtY) results in the formation of β-carotene, which is subsequently hydroxylated by β-carotene hydroxylase (CrtZ), yielding zeaxanthin. Zeaxanthin can be further modified by different tailoring enzymes to yield a variety of carotenoids. For example, it can be glycosylated by the glycosyltransferase (CrtX) to generate zeaxanthin diglucoside [20,21,22,23].

Proposed biosynthetic pathway of zeaxanthin diglucoside in Pseudomonas sp. 102515. The involved enzymes include isopentenyl-diphosphate delta-isomerase (PsIdi), geranylgeranyl pyrophosphate synthetase (PsCrtE), phytoene synthase (PsCrtB), lycopene synthase (PSCrtI), lycopene cyclase (PsCrtY), β-carotene hydroxylase (PsCrtZ), and zeaxanthin glucosyltransferase (PsCrtX)
In this study, our primary purpose was to isolate endophytic bacteria with the purpose of discovering new natural products and/or new effective means to produce carotenoids. We isolated an endophytic Pseudomonas strain, named Pseudomonas sp. 102515, from the leaves of the yew tree, which was found to produce zeaxanthin diglucoside as the major product. Based on the genome of P. psychrotolerans PRS08–11306 (the closest relative to the endophytic strain) as reference, we were able to amplify a carotenoid (Pscrt) biosynthetic gene cluster from Pseudomonas sp. 102515. Two genes in this gene cluster were functionally characterized. The entire Pscrt gene cluster was successfully expressed in two heterologous hosts, Escherichia coli BL21(DE3) and Pseudomonas putida KT2440. Additionally, the production of zeaxanthin diglucoside in Pseudomonas sp. 102515 was significantly improved by optimizing the fermentation conditions, with a yield of 206 ± 6 mg/L. This endophyte was further engineered by introducing an expression plasmid harboring the Pscrt gene cluster. The yield of zeaxanthin diglucoside by the engineered strain reached 380 ± 12 mg/L, 85% higher than the wild type strain. This strain represents a promising host for the production of zeaxanthin diglucoside.
Results
Identification and phylogenetic analysis of an endophytic bacterium from Taxus chinensis
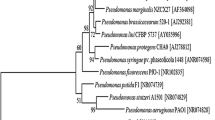
We isolated an endophytic bacterium from the leaves of Taxus chinensis, whose colonies showed a bright yellow color. To identify this strain, we amplified the 16S ribosomal RNA (rRNA) gene fragment by PCR using a set of universal primers. The gene sequence of this fragment was subjected to BLAST analysis, with the 16S rRNA sequences (bacteria and archaea) as the reference database. The BLASTn analysis revealed that there are three Pseudomonas strains with 99% identity and coverage. Some other Pseudomonas strains (Pseudomonas stutzeri, Pseudomonas indoloxydans, Pseudomonas luteola, etc.) also had 99% coverage but are 96% identical to our endophytic isolate. Using an online platform (https://itol.embl.de/), we constructed a phylogenetic tree by the neighbor-joining method (Fig. 2a). The phylogenetic tree indicated that P. psychrotolerans is the closest relative to our isolate. P. oryzihabitans and P. oleovorans are also closely related to this endophytic strain. We also looked into the physiology of our isolate under a scanning electron microscope (SEM) (Fig. 2b) and confirmed that it is a rod-shaped bacterium. Additionally, it is a non-spore-forming and a yellow-pigment-producing bacterium. The closest relative, P. psychrotolerans, was also reported as a yellow-pigmented bacterium [24]. All these physiological characteristics and genetic analysis indicated that this endophyte is a Pseudomonas strain, which was named as Pseudomonas sp. 102515.

Identification of Pseudomonas sp. 102515. (a) A phylogenetic tree constructed based on the 16S rRNA sequences of Pseudomonas strains from BLASTn analysis. It estimates the relationship between strain 102515 and other Pseudomonas strains that shared the highest 16S rRNA gene sequence similarities. Numbers at the nodes indicate the normalized percentages of occurrence in 100 bootstrapped trees, and only values greater than 0.5 are shown. Sequences of reference species were obtained from GenBank, including P. luteola (NR_114215), P. asuensis (NR_136445), P. duriflava (NR_044390), P. benzenivorans (NR_116904), P.stutzeri (NR_041715), P. songnenensis (NR_148295), P. alcaligenes (NR_113646), P. otitidis (NR_043289), P. aeruginosa (NR_117678), P. alcaliphila (NR_114072), P. toyotomiensis (NR_112808), P. chengduensis (NR_125523), P.oleovorans (NR_043423), P. oryzihabitans (NR_114041), and P. psychrotolerans (NR_042191.1). (b) SEM micrograph of Pseudomonas sp. 102515. The bacterium was fixed with 2% glutaraldehyde in 0.1% HEPES buffer overnight. The samples were subjected to alcohol series dehydration (50–100% ethanol) and then chemically dried using hexamethyldisilazane
Amplification and analysis of a zeaxanthin diglucoside biosynthetic gene cluster from Pseudomonas sp. 102515
As described above, P. psychrotolerans is the closest relative strain to Pseudomonas sp. 102515. The genome of this strain is available in GenBank under the accession number, NZ_CP018758. We analyzed the genome of P. psychrotolerans with the help of the AntiSMASH online genome analysis platform. Four major natural product biosynthetic gene clusters were identified. The first one is a nonribosomal peptide synthetase (NRPS)-type gene cluster with 50% similarity to the known taiwachelin biosynthetic gene cluster. A turnerbactin (tnb) biosynthetic gene cluster was also found to be located in this NRPS gene cluster. Turnerbactin was reported to have an important role in the plant-microbe interactions [25]. In order to find out whether a tnb gene cluster is present in Pseudomonas sp. 102515, we amplified the tnbA gene from its genomic DNA using a pair of primers designed based on the genome of P. psychrotolerans. The sequence of tnbA was provided in the Supporting Information (Sequence S1). The second gene cluster is an arylpolyene (APE)-type biosynthetic gene cluster from which 40% of genes show a similarity to the genes from the characterized APE biosynthetic gene cluster. APE gene clusters are widely distributed in gram-negative bacteria, including Pseudomonas strains [26]. In addition to another siderophore biosynthetic gene cluster, genome analysis revealed that P. psychrotolerans has a carotenoid (crt) biosynthetic gene cluster as expected from its carotenoid production capability. The genes from this crt gene cluster is similar to those found in the crt gene cluster from Pantoea agglomerans, which contains crtE, idi, crtX, crtY, crtI, crtB and crtZ [27].
To find out what yellow compound Pseudomonas sp. 102515 produced, we extracted the cells with methanol/chloroform (2:1, v/v) and analyzed the products by LC-MS. The extract of Pseudomonas sp. 102515 showed a major peak at 18.2 min (Fig. 3a). Based on its UV and MS spectra (Fig. 3b and c), this compound was characterized as zeaxanthin diglucoside. Furthermore, this peak has the same retention time and UV spectrum as zeaxanthin diglucoside produced by E. coli BL21(DE3)/pAC-EHER [28]. Therefore, we confirmed that the yellow pigment from this endophyte is zeaxanthin diglucoside.

Production of zeaxanthin diglucoside by Pseudomonas sp. 102515. (a) HPLC analysis (460 nm) of zeaxanthin diglucoside production by Pseudomonas sp. 102515. (i) Pseudomonas sp. 102515, (ii) E. coli BL21(DE3)/pAC-EHER. (b) Comparison of the UV spectra of the major product of Pseudomonas sp. 102515 and E. coli BL21(DE3)/pAC-EHER. (c) ESI-MS(−) spectrum of the major product of Pseudomonas sp. 102515
Based on the sequence of the crt gene cluster in P. psychrotolerans, we designed two sets of primers and successfully amplified a ~ 9.4-kb carotenoid (Pscrt) biosynthetic gene cluster from Pseudomonas sp. 102515. The open reading frames (orfs) in this gene cluster were analyzed and annotated (Table 1), including PscrtZ, PscrtB, PscrtI, PscrtY, PscrtX, Psidi, PscrtE, and two orfs. Based on the predicted functions of these genes, we propose that this gene cluster is responsible for the biosynthesis of zeaxanthin diglucoside (Fig. 1).
Functional characterization of two carotenoid biosynthetic genes from the endophyte
To characterize the functions of the carotenoid biosynthetic genes from Pseudomonas sp. 102515, it is necessary to develop an effective expression system for carotenoid biosynthetic enzymes in E. coli. We first ligated the crtX (zeaxanthin glucosyltransferase) gene from pAC-EHER into pET28a(+) to yield pOKF63. However, co-expression of pOKF63 and zeaxanthin-producing pAC-ZEAXipi in E. coli BL21(DE3) did not produce zeaxanthin diglucoside upon induction with 200 μM of IPTG. We next replaced the T7 promoter in pET28a(+) with a strong constitutive promoter J23119 and the B0034 ribosome binding site (pOKF72). Co-expression of pOKF72 and pAC-ZEAXipi in E. coli BL21(DE3) did yield zeaxanthin diglucoside, which was confirmed by HPLC through a comparison of the retention time and UV spectrum of the authentic sample obtained from E. coli BL21(DE3) harboring pAC-EHER (data not shown). We next used this system to test the functions of selected biosynthetic genes in the Pscrt gene cluster. PsCrtI and PsCrtY were chosen as their functions can be easily observed by the color change in the products. Co-expression of phytoene-producing plasmid pAC-PHYTipi with PsCrtI in E. coli BL21(DE3) led to the production of lycopene. We observed that the color of cell pellets changed from colorless (E. coli BL21(DE3)/pAC-PHYTipi) to red, as seen in Fig. 4a, indicating that a red compound was produced. For further confirmation, we compared the extract of this engineered strain with those of the negative (pAC-PHYTipi) and positive (pAC-LYCipi) controls by HPLC (Fig. 4a), which clearly revealed that lycopene was formed. This was supported by the UV spectra (Fig. 4b). This result confirmed that PsCrtI is a phytoene desaturase that converts phytoene to lycopene (Fig. 1). Similarly, we also confirmed the function of PsCrtY as a lycopene cyclase in E. coli BL21(DE3) by co-expressing pAC-LYCipi with PsCrtY. We observed the color change from red to yellow due to the conversion of lycopene into β-carotene by PsCrtY (Fig. 5a). Production of β-carotene was further confirmed by its retention time (Fig. 5)a and UV spectrum (Fig. 5b).

Functional identification of PsCrtI as a phytoene desaturase. (a) HPLC analysis of lycopene production through co-expression of PsCrtI with the phytoene biosynthetic enzymes. (i) E. coli BL21(DE3)/pAC-PHYTipi (negative control) producing phytoene (retention time: 17.5 min, 280 nm), (ii) E. coli BL21(DE3)/pAC-PHYTipi+pOKF89 (pAC-PHYTipi+PsCrtI) producing lycopene (retention time: 14.8 min, 460 nm), (iii) E. coli BL21(DE3)/pAC-LYCipi (positive control) producing lycopene (retention time: 14.8 min, 460 nm). Color change of harvested cells due to the co-expression of PsCrtI with pAC-PHYTipi in E. coli BL21(DE3) is shown on the right. (b) A comparison of the UV spectra of lycopene produced by E. coli BL21(DE3)/pAC-PHYTipi+pOKF89 with lycopene produced by the positive control and phytoene produced by the negative control

Functional identification of PsCrtY as a lycopene cyclase. (a) HPLC analysis (460 nm) of β-carotene production through co-expression of PsCrtY with the lycopene biosynthetic enzymes. (i) E. coli BL21(DE3)/pAC-LYCipi (negative control) producing lycopene (retention time: 14.8 min), (ii) E. coli BL21(DE3)/pAC-LYCipi+pOKF91 (pAC-LYCipi+PsCrtY) producing β-carotene (retention time: 13.3 min), (iii) E. coli BL21(DE3)/pAC-BETAipi (positive control) producing β-carotene (retention time: 13.3 min). Color change of harvested cells due to the co-expression of PsCrtY with pAC-LYCipi in E. coli BL21(DE3) is shown on the right. (b) A comparison of the UV spectra of β-carotene produced by E. coli BL21(DE3)/pAC-LYCipi+pOKF91 with β-carotene produced by the positive control and lycopene produced by the negative control
Heterologous expression of the Pscrt gene cluster in E. coli BL21(DE3) and Pseudomonas putida KT2440
Identification of PsCrtI and PsCrtY further suggested that this gene cluster is responsible for the biosynthesis of zeaxanthin diglucoside in Pseudomonas sp. 102515. Instead of characterizing each of the remaining genes in this gene cluster, we attempted to express the entire gene cluster in a heterologous host. We first transferred the Pscrt gene cluster into pET28a(+). Heterologous expression of the Pscrt gene cluster through the pET28a(+) expression system did not produce detectable amounts of zeaxanthin diglucoside. Then, we cloned the Pscrt gene cluster into a pACYC184-based expression system that has been previously used for the heterologous production of carotenoids in E. coli [28]. Expression of the Pscrt gene cluster in E. coli BL21(DE3) led to the production of zeaxanthin diglucoside (Fig. 6), which confirmed that this gene cluster is indeed responsible for the biosynthesis of this glycosylated carotenoid. However, the yield of zeaxanthin diglucoside was very low in E. coli BL21(DE3) as the cells only showed a slight yellow color. We figured that P. putida KT2440 might be a better host since it belongs to the genus of Pseudomonas. To this end, we cloned the Pscrt biosynthetic gene cluster into the pMIS1-mva vector, and expressed the resulting plasmid (pOKF192) in P. putida KT2440. Expression of this plasmid in P. putida KT2440 was deemed successful by observing the bright yellow color of the cells, and the formation of zeaxanthin diglucoside was verified by HPLC analysis (Fig. 6).

Heterologous reconstitution of the zeaxanthin diglucoside biosynthetic pathway in E. coli BL21(DE3) and P. putida KT2440. Shown are HPLC traces (460 nm) of the extracts of Pseudomonas sp. 102515 (i), E. coli BL21(DE3)/pOKF184 (ii), and P. putida KT2440/pOKF192 (iii)
Enhanced production of zeaxanthin diglucoside in Pseudomonas sp. 102515
To improve the production of zeaxanthin diglucoside in Pseudomonas sp. 102515, we next attempted to optimize the culture conditions. We used a UV spectrophotometer to measure the amount of zeaxanthin diglucoside in the extracts at 456 nm, which is the wavelength of maximum absorbance for zeaxanthin diglucoside as reported in the literature [29]. Using the standard curve prepared with purified zeaxanthin diglucoside (Additional file 1: Figure S1), we calculated the yields from the crude extracts of each sample. Seven different media with 3 replicates were tested for the production of zeaxanthin diglucoside by Pseudomonas sp. 102515 in culture tubes. LB, SOC and superbroth media showed better yields compared to the other media (Additional file 1: Figure S2). We then tested LB, SOC and superbroth media in 50-mL flasks, which showed even better yields of zeaxanthin diglucoside than the tubes. As shown in Fig. 7a, SOC exhibited the best yield (98 ± 7 mg/L) among these three media.

Optimization of zeaxanthin diglucoside production in Pseudomonas sp. 102515. (a) The effects of culture media on the production of zeaxanthin diglucoside. (b) The effects of supplementation of glycerol on the production of zeaxanthin diglucoside in the flasks. (c) The effects of cultivation temperature and time on the production of zeaxanthin diglucoside by Pseudomonas sp. 102515 and Pseudomonas sp. 102515/pOKFF192. Cells were grown in 50 mL of SOC medium supplemented with 0.5% glycerol. Black unfilled circle: 102515 at 18 °C; orange square: 102515 at 23 °C; purple unfilled triangle: 102515 at 28 °C; red filled circle: 102515/pOKF192 at 18 °C; blue filled square: 102515/pOKF192 at 23 °C; green filled triangle: 102515/pOKF192 at 28 °C. (d) A comparison of the production yield of zeaxanthin diglucoside by Pseudomonas sp. 102515 and three engineered strains
Glycerol was previously reported to enhance the production of carotenoid [30]. Therefore, we tested the effects of glycerol on the production of zeaxanthin diglucoside by this endophyte in flasks at different concentrations (0.5, 1 and 2% final concentration). Supplementation of 0.5% glycerol (final concentration) into SOC medium increased the yield of zeaxanthin diglucoside to 127 ± 2 mg/ L after 3 days at 28 °C, representing a 30% increase in the yield (Fig. 7b). We next examined how different cultivation times and temperatures (18 °C, 23 °C, 28 °C and 37 °C) affect the yield of zeaxanthin diglucoside in SOC medium with 0.5% glycerol. The cultivation temperature of 37 °C resulted in extremely low yields (data not shown). Among the tested temperatures, 18 °C worked best and the yield reached 206 ± 6 mg/L in SOC medium after 5 days of cultivation (Fig. 7c). Under these conditions, we also tested the yields of zeaxanthin diglucoside in the engineered strains of E. coli BL21(DE3) and P. putida KT2440. Consistent with the cell colors, E. coli BL21(DE3) only produced 2 mg/L zeaxanthin diglucoside, while P. putida KT2440/pOKF192 generated the product in a much higher yield (121 ± 6 mg/L). However, since this yield is still lower than the wild type (206 mg/L), the latter is a better starting strain for further improvements. We successfully engineered Pseudomonas sp. 102515 by introducing pOKF192 into this strain, which allows this endophyte to have another copy of the Pscrt gene cluster. Expression of this plasmid further improved the yield to 380 ± 12 mg/L after 5 days of incubation at 18 °C (Fig. 7c and d).
Discussion
Carotenoids have a variety of health-benefiting activities. They are also an important dietary source of vitamin A [31], thus representing a group of industrially important natural products. Carotenoids are considered to protect cells from the damaging effects of reactive oxygen species (ROS), which are formed by normal metabolic activities and lifestyle factors such as diet, smoking and exercise. Carotenoids might potentially diminish the destructive effects of ROS such as superoxide (O2−), hydrogen peroxide (H2O2), singlet oxygen (1O2) and hydroxyl radical (OH) [32, 33]. In specific, 1O2 is a product in both biochemical and photochemical systems that is responsible for the cell destruction caused by light and certain photosensitizers. The number of conjugated double bands in the carotenoids is a significant factor for the 1O2 quenching activity [34, 35]. Addition of the glucose moiety was found to enhance the 1O2 quenching activity of carotenoids. For example, the 1O2 quenching activity of a series of carotenoids were tested in E. coli. The kq value of zeaxanthin diglucoside was 3.5 times higher than that of zeaxanthin [33]. Similarly, the viability (38.7%) of zeaxanthin-diglucoside-producing strain of E. coli was higher than that (25.7%) of zeaxanthin-producing strain in the 1O2 generation medium [33], further indicating that zeaxanthin diglucoside has better protecting effects than its aglycone.
Carotenoids such as lycopene, lutein, β-carotene, astaxanthin, and zeaxanthin are quite common in nature. However, glycosylated carotenoids such as dihydroxylycopene diglucoside, adonixanthin diglucoside and zeaxanthin diglucoside are relatively rare [36,37,38,39]. Glycosylated carotenoids can be potent antioxidant agents with protective properties against photooxidative damages from ROS and visible light [40, 41]. For instance, a novel glycosylated carotenoid, caloxanthin 3′-β-D-glucoside, was reported to have a potent 1O2 quenching activity with an IC50 of 19 μM [42]. Moreover, Tatsuzawa et al. reported that the viability of zeaxanthin diglucoside producing E. coli cells is higher in an 1O2 generation mixture compared to the cells that produce other carotenoids such as zeaxanthin, astaxanthin, β-carotene, and canthaxanthin [33]. Glycosylated carotenoids are also reported to stabilize the membrane through integrating within the lipid membrane due to their polar functional groups [40, 43]. Synthetic coloring agents have the negative effects on human health and thus there is an increasing consumer demand for natural and health-promoting food ingredients. In addition to their promising biological activities, such as antioxidant, anticancer, and anti-inflammatory properties, glycosylated carotenoids can be used in a wide variety of applications, ranging from food colorants and feed supplements to nutritional and cosmetics purposes [15]. Typically, carotenoids are poorly soluble in water. By contrast, glycosylated carotenoids were found to have a much higher water solubility than the corresponding carotenoid aglycones while maintaining similar biological activities. For instance, the water solubility of zeaxanthin diglucoside is more than 60-fold higher than that of zeaxanthin, which makes the use of this carotenoid in the water environment much easier [44]. Therefore, engineered production of glycosylated carotenoids including zeaxanthin diglucoside that are rare in nature is of great importance.
In this study, we isolated an endophytic bacterium Pseudomonas sp. 102515 from the leaves of T. chinensis, which was found to produce zeaxanthin diglucoside as a major metabolite. BLAST and phylogenetic analyses (Fig. 2) revealed that our endophytic isolate is a Pseudomonas strain. Pseudomonas species have been isolated from diverse sources including marine, freshwater, animals and plants [45,46,47,48]. Certain Pseudomonas strains, such as fluorescent Pseudomonads, are predominantly found in the rhizosphere and have been reported to move from rhizosphere to aerial plant tissue as in the case of P. aeurofaciens [49]. Pseudomonas species were also reported as endophytes in the literature [50, 51]. For instance, P. stutzeri A15 is an endophytic nitrogen-fixing bacterium isolated from paddy rice [52]. P. fluorescens was also reported to be an endophyte with beneficial interaction with plants [53]. The plant-endophyte interactions have not been fully understood. Many endophytes not only have beneficial effects on their hosts, but also plays a significant role in plant physiology. For example, some endophytic bacteria were reported to provide phytohormones, low-molecular compounds or enzymes to the host plants that led to the enhanced plant growth [54,55,56]. Endophytic bacteria also provide an alternative way to manage plant pathogens as a promising biocontrol agent through various ways, such as releasing antimicrobial substances, producing siderophores, and inducing the systemic resistance to pathogens [57,58,59]. For instance, biological control of P. syringae pv. actinidiae, the causal agent of bacterial canker of kiwifruit, was achieved by using an endophytic bacterium isolated from a medicinal plant [60].
To better understand Pseudomonas sp. 102515, we looked into the closely related strains, P. psychrotolerans and P. oryzihabitans. Both strains were reported as yellow-pigmented gram-negative bacteria and isolated from different sources including clinical samples, copper coins, diseased rice and rice seeds [61,62,63,64,65]. P. oryzihabitans was previously isolated from Hibiscus rosasinensis as an endophyte [66]. P. psychrotolerans was found to be an endosymbiotic bacterium from diseased rice and rice seeds in two different studies [61, 65]. The later endophytic bacterium was reported to enhance the plant growth due to potential nitrogen fixing characteristics of the turnerbactin (tnb) biosynthetic gene cluster, which is responsible for the biosynthesis of turnerbactin, a tricatecholate siderophore. Plants infected with P. psychrotolerans PRS08–11306 showed enhanced growth [25, 65]. We were able to amplify one of the key genes, tnbA, suggesting the existence of a tnb gene cluster in Pseudomonas sp. 102515. Thus, this endophyte may have potential as plant growth-promoting bacterium for agricultural applications.
In addition to the tnb gene cluster, we found a complete carotenoid (Pscrt) biosynthetic gene cluster in Pseudomonas sp. 102515, which contains a series of carotenoid biosynthetic genes (Table 1). However, there are some differences in the organization of the genes between the Pscrt gene cluster and other reported ones (Fig. 8). One obvious difference is the additional non-carotenoid gene (orf2) encoding for gluconate 2-dehydrogenase, which has not been reported in a carotenoid biosynthetic gene cluster to our knowledge. The role of this gene remains to be characterized. Typically, all genes in a crt biosynthetic gene cluster except crtZ have the same direction and are controlled by the same promoter in many γ-proteobacteria [27]. However, PscrtE also has a different direction compared to most of the genes in the Pscrt gene cluster and is controlled by its own promoter (Fig. 8). The similar organization of crtE in the gene cluster was also reported in Pseudomonas sp. strain Akiakane isolated from the excrement of autumn darker, yet its crtZ gene in that gene cluster is located differently than ours and overlapped with the sequences of crtX and crtY as shown in Fig. 8 [48]. We analyzed the functions of two Pscrt genes amplified from Pseudomonas sp. 102515 to confirm their functions through co-expression with heterologous carotenoid-producing genes in E. coli BL21(DE3) [67, 68]. PsCrtI was confirmed to be a phytoene desaturase and PsCrtY is a lycopene cyclase. The function of the Pscrt gene cluster was characterized by expression of the entire biosynthetic gene cluster in two heterologous hosts, E. coli BL21(DE3) and P. putida KT2440, both yielding zeaxanthin diglucoside (Fig. 6). This heterologous expression strategy not only allows the functional confirmation of this gene cluster, but also provides an alternative way to produce zeaxanthin diglucoside. However, the yield of zeaxanthin diglucosde in E. coli was very low compared to Pseudomonas sp. 102515, likely due to the low efficiency of the native promoter in this host. Sometimes a native promoter might not work well in a heterologous host. For instance, expression of the two phaC genes from P. putida were tested in E. coli under the control of a native promoter or an external promoter. They were only expressed in E. coli with the external promoter [69]. In another study, the levansucrase genes from P. syringae pv. glycinea PG4180 and P. syringae pv. phaseolicola NCPPB 1321 were cloned with the native promoters, yet the expression of levansucrase was only achieved upon the use of a lac promoter [70]. Therefore, we decided to use external promoters, in particular constitutive promoters, for heterologous expression of the Pscrt biosynthetic gene cluster from Pseudomonas sp. 102515 in E. coli. This approach allowed us to successfully reconstitute the production of zeaxanthin diglucoside in E. coli. Another approach we used was to choose another host microorganism. Since our isolate is a Pseudomonas strain, we proposed that the Pscrt gene cluster might be expressed better in P. putida KT2440 than E. coli. P. putida KT2440 was previously used as a heterologous host for the production of natural products including zeaxanthin [71, 72]. Our results confirmed that expression of the Pscrt gene cluster in P. putida KT2440 resulted in a much higher yield of zeaxanthin diglucoside than in E. coli (Fig. 7d).

Organization of several carotenoid biosynthetic gene clusters in γ-proteobacteria. GenBank accession numbers: M87280 for Pantoa agglomerans, CP001875 for Pantoa ananatis, and LC317091 and LC317092 for Pseudomonas sp. strain Akiakane [48]
A common strategy to improve the yield is the optimization of culture conditions [73]. We used LB medium for the isolation of Pseudomonas sp. 102515 and the initial product analysis of this endophyte. However, the nutrients in different media can affect the production of zeaxanthin diglucoside. A scanning of 7 different media (Additional file 1: Table S1) in culture tubes showed that LB, superbroth and SOC media had better yields ranging from 13 to 15 mg/L (Additional file 1: Figure S2). We chose SOC medium for further optimization studies due to its higher yield in flasks (Fig. 7a) and found that 0.5% glycerol supplementation further increased the yield to 127 ± 2 mg/L (Fig. 7b), which is consistent with a previous work on the effect of glycerol on the production of carotenoids [30]. It was found that supplementation of higher concentrations of glycerol decreased the titer of zeaxanthin diglucoside (Fig. 7B). This might be due to the inhibitory effect of 3-hydroxypropionaldehyde, a metabolite of glycerol. This compound was previously reported to inhibit the growth of Enterobacter agglomerans, which, similar to our strain, is a gram-negative bacterium with the carotenoid biosynthesis capability [74]. The effects of cultivation temperature and time were also examined. We originally used 28 °C as cultivation temperature as reported in the literature for the production of carotenoids [28, 67, 68, 71]. However, a comparison of four different temperatures revealed that 18 °C was the best cultivation temperature for the production of zeaxanthin diglucoside in Pseudomonas sp. 102515. The yield reached 145 ± 5 mg/L in 50 mL of SOC medium. We then cultivated this endophyte in SOC medium supplemented with 0.5% glycerol (final concentration) at 18 °C for a week to find out the best fermentation time. Based on the time course analysis, the yield reached the highest (206 ± 6 mg/L) after 5 days of cultivation. Low cultivation temperature is an economic burden for industrial production although it often favors the expression of enzymes [75]. Therefore, we cultivated Pseudomonas sp. 102515 in 50 mL of SOC medium supplemented with 0.5% glycerol at 23 °C for a week (Fig. 7c). After 5 days of cultivation, we obtained the yield of 197 ± 8 mg/L that is slightly lower than but comparable to the yield at 18 °C. Thus, 23 °C can be used for industrial production of zeaxanthin diglucoside with Pseudomonas sp. 102515. To further improve the yield, we introduced pOKF192 carrying the Pscrt gene cluster into Pseudomonas sp. 102515. With this additional copy of the Pscrt gene cluster, Pseudomonas sp. 102515/pOKF192 produced zeaxanthin diglucoside at 380 ± 12 mg/L, which is 85% higher than the parent strain and represents the highest yield of this promising antioxidant (Fig. 7c and d). Similar to Pseudomonas sp. 102515, this engineered strain at 23 °C led to a slightly lower but comparable yield than 18 °C (Fig. 7c).
Conclusions
In conclusion, we isolated a carotenoid-producing endophytic Pseudomonas strain from the yew tree, which can potentially offer an alternative way to produce zeaxanthin diglucoside. A complete Pscrt gene cluster was discovered in this endophyte, from which the functions of two Pscrt genes were characterized in E. coli BL21(DE3). We cloned the whole Pscrt gene cluster and successfully expressed it in the heterologous hosts, E. coli BL21(DE3) and P. putida KT2440. PCR analysis also showed that a turnerbactin biosynthetic gene cluster exists in the endophytic strain, which renders our isolate a potential plant-growth-promoting bacterium for agricultural applications. Through fermentation and genetic engineering, we increased the yield of zeaxanthin diglucoside to 380 ± 12 mg/L. This engineered strain represents a great host for industrial production of this promising glycosylated carotenoid.
Methods
Bacterial strains, media and culture conditions
E. coli XL1-Blue, which was routinely grown in LB (Luria-Bertani) medium at 37 °C, was used for general genetic manipulations. Pseudomonas sp. 102515 was grown in LB overnight for genomic DNA extraction. LB, SOB, SOC, 2 × YT, TB, 2 × TB and superbroth media (Recipes for media are in Additional file 1: Table S1) were tested for the production of zeaxanthin diglucoside by Pseudomonas sp. 102515. E. coli BL21(DE3) and P. putida KT2440 were utilized for heterologous expression and co-expression studies. All wild type and engineered bacterial strains are listed in Table 2. Chloramphenicol (25 μg/mL), ampicillin (50 μg/mL) and kanamycin (50 and 30 μg/mL) were supplemented when appropriate. The concentrations of kanamycin for engineered E. coli and Pseudomonas strains are 50 and 30 μg/mL, respectively.
Isolation of endophytic bacteria from T. chinensis
The leaves were collected from a cultivated shrub of T. chinensis on the Logan campus of Utah State University (Latitude: 41.7424, Longitude: − 111.8080). A voucher specimen is deposited in the Intermountain herbarium, accession number UTC00282046. The collected leaves were thoroughly washed in running water. The disinfection and isolation were performed according to de Oliveira Costa et al. with minor modifications [77]. Briefly, the leaves were disinfected superficially through dipping into 70% ethanol for 2 min 3 times, followed by rinsing with 70% ethanol 3 times. Then, the leaves were dipped into sterile distilled water for a few minutes and rinsed with sterile distilled water. This process was repeated 3 times. To confirm the disinfection protocol, aliquots of the sterile water used in the final rinse were plated in LB plates at 28 °C for 7 days and the plates are examined for the presence or absence of any microbial colonies.
The disinfected leaves were grounded with 6 mL of an aqueous NaCl solution (0.85%) using a sterile mortar and pestle. The tissue extract was subsequently incubated at 28 °C for 3 h to allow the complete release of endophytic microorganisms from the host tissue. For the isolation of endophytic bacteria, the tissue extract was diluted with the NaCl solution and plated on LB plates with different dilutions. The plates were incubated for around a week at 28 °C. Promising colonies were selected and streaked on fresh LB plates for the isolation of promising bacteria. In particular, we picked the pigment-producing colonies in an attempt to isolate a carotenoid-producing endophyte.
PCR and general genetic manipulations
Standard molecular biology protocols were performed as previously described [78]. Genomic DNA of Pseudomonas sp. 102515 was extracted with a ZR Fungal/Bacterial DNA Miniprep Kit. Plasmid DNA extraction from E. coli cells was performed using a Thermo Scientific GeneJET Plasmid Miniprep Kit. PCR reactions were performed with an Arktik™ Thermal Cycler using Phusion DNA polymerase. Primers were ordered from Thermo Fisher Scientific and dissolved in TE buffer to the final concentration of 100 ng/mL.
Identification and phylogenetic analysis of the endophytic bacterium
The partial 16S rRNA gene fragment was PCR amplified from the genomic DNA of Pseudomonas sp. 102515 using the 16S universal primers of GGCTACCTTGTTACGACTTC and AGTTTGATCCTGGCTCAG [79]. The PCR product was sequenced by Sanger’s method. The 16S rDNA sequence was deposited into GenBank under the accession number of MK610450. BLAST analysis of this 16S rDNA sequence was performed using the 16S ribosomal RNA sequences database as the reference. Using the 16S rRNA sequences of the related bacteria from BLAST analysis, we created a phylogenetic tree by the neighbor-joining method using an online platform (https://itol.embl.de/) [80]. For physiological characteristics, scanning electron microscopy images were taken after performing the fixation of the sample [81].
Amplification and annotation of the Pscrt biosynthetic gene cluster
The colonies of Pseudomonas sp. 102515 on LB agar plates showed a yellow color. LC-MS analysis of the chloroform extract of the cells confirmed that zeaxanthin diglucoside was produced. To discover the carotenoid (Pscrt) biosynthetic gene cluster in Pseudomonas sp. 102515, we first analyzed the genome of P. psychrotolerans (GenBank accession number: NZ_CP018758), which is the closest relative to our strain based on the phylogenetic analysis. We used an online genome analysis platform, AntiSMASH, and found a complete crt biosynthetic gene cluster responsible for the biosynthesis of zeaxanthin diglucoside. We designed two sets of primers (primers 3–6, Additional file 1: Table S2) based on the DNA sequence of this crt gene cluster. Briefly, the whole Pscrt gene cluster was divided into two fragments, which were PCR amplified from the genomic DNA of Pseudomonas sp. 102515 using the primers (primers 3 and 4 for fragment A, and primers 5 and 6 for fragment B) listed in Additional file 1: Table S2. Fragments A and B were first ligated into pJET1.2, yielding pOKF163 and pOKF166 (Table 3), respectively. These two plasmids were sent out for DNA sequencing to obtain the whole DNA sequence of the Pscrt gene cluster using the walking primers (primes 7–16) listed in Additional file 1: Table S2. The sequence of the Pscrt biosynthetic gene cluster from Pseudomonas sp. 102515 was deposited into GenBank under the accession number of MK613929.
Construction of plasmids for functional characterization of PsCrtI and PsCrtY
We first constructed a pET28a(+)-based expression plasmid for the carotenoid biosynthetic genes in E. coli. crtX (glycosyltransferase) was PCR amplified using primers 17 and 18 (Additional file 1: Table S2) from pAC-EHER, which was a gift from Francis X Cunningham Jr. (Addgene plasmid # 53262). PCR product was directly ligated into the cloning vector, pJET1.2, yielding pOKF55 (Table 3), which was sequenced using the Sanger method. crtX was excised with NcoI and HindIII from pOKF55 and ligated into pET28a(+) between the same sites to yield pOKF63 (Table 3). pOKF63 and pAC-ZEAXipi (Addgene plasmid # 53287) were introduced into E. coli BL21(DE3) for co-expression. The engineered strain was grown in LB medium with 50 μg/mL kanamycin and induced with 200 μM of IPTG for product analysis.
To replace the T7 promoter in pET28a (+) with a strong constitutive promoter, J23119, we redesigned the forward primer for crtX by including the J23119 promoter and B0034 ribosome binding site before the start codon (Additional file 1: Table S2). We re-amplified crtX with J23119 and B0034 from pAC-EHER using this new forward primer and previous reverse primer (primers 19 and 20). The new PCR product was ligated into pJET1.2 (pOKF69, Table 3) and the sequence was confirmed by sequencing. Subsequently, crtX with J23119 and B0034 was excised from pOKF69 using BglII and HindIII, and then ligated into pET28a(+) between the same sites to yield pOKF72 (Table 3). pOKF72 was co-expressed with pAC-ZEAXipi (Addgene plasmid # 53287) in E. coli BL21(DE3), and the products of the engineered strain was analyzed by HPLC.
PscrtI and PscrtY were PCR amplified from the genomic DNA of Pseudomonas sp. 102515 using the primers (primers 21 and 22 for PscrtI, and primers 23 and 24 for PscrtY) listed in Additional file 1: Table S2 and were subsequently ligated into pJET1.2 to yield pOKF85 and pOKF88 (Table 3), respectively. After sequencing, PscrtI and PscrtY were transferred into pET28a(+) with J23119 and B0034 using the NdeI and HindIII sites to yield pOKF89 and pOKF91 (Table 3), respectively. PsCrtI (pOKF89) and PsCrtY (pOKF91) were separately co-expressed with pAC-PHYTipi (Addgene plasmid #: 53283) and pAC-LYCipi (Addgene plasmid #: 53279) in E. coli BL21(DE3), respectively.
Heterologous expression of the Pscrt biosynthetic gene cluster from Pseudomonas sp. 102515 in E. coli BL21(DE3) and P. putida KT2440
For the heterologous expression of the Pscrt gene cluster from the endophytic strain, we first excised fragment A from pOKF163 using NheI and MfeI and ligated it into pOKF166 that contains fragment B between the same sites to yield pOKF169 (Table 3), which harbors the entire Pscrt gene cluster in pJET1.2. We then ligated the Pscrt gene cluster excised from pOKF169 using SpeI and HindIII into pET28a(+) digested with the NheI and HindIII sites (pOKF173, Table 3). Since pET28a(+) has a relatively higher copy number, we decided to also ligate the Pscrt gene cluster from pOKF169 into pACYC184 between the XbaI and HindIII sites to yield pOKF184 (Table 3). The corresponding expression plasmids, including pOKF173 and pOKF184, were transferred into E. coli BL21(DE3) for heterologous expression of the Pscrt gene cluster.
We also constructed an expression plasmid for expression of the Pscrt gene cluster in P. putida KT2440. The pMIS1-mva-ges plasmid was generously provided by Jens Schrader and Josef Altenbuchner at Dechema-Forschungsinstitut and University of Stuttgart, respectively [71, 72]. The whole Pscrt gene cluster was excised from pOKF169 using SpeI and PmeI and ligated into pMIS1-mva-ges digested with AvrII and PmeI to yield pOKF192 (Table 3). pOKF192 was introduced into P. putida KT2440 by electroporation as described in the literature [82]. Briefly, P. putida KT2440 was grown until the OD600 reached 0.4 at 28 °C. The cells were immediately placed on ice and harvested by centrifugation. Then, the cells were washed three times with 300 mM sterile sucrose solution and re-suspended in 100 μL of 300 mM sucrose solution. The cells were mixed with pOKF192 and the mixture was transferred into a pre-chilled electroporation cuvette. After 10 min of incubation on ice, the following settings were used for electroporation: set voltage - 2.5 kV (12.5 kV/cm); capacitor - 25 μF. After electroporation, 900 μL of LB broth was immediately added to the cuvette. The mixture was then transferred into a culture tube, which was incubated at 28 °C with shaking (250 rpm) for 2 h. Finally, the cells were plated on LB agar plates with 30 μg/mL kanamycin and incubated at 28 °C. The transformants were picked and grown in LB with kanamycin for product analysis. The same electroporation protocol was followed to introduce pOKF192 into Pseudomonas sp. 102515.
Discovery of a turnerbactin biosynthetic gene cluster in Pseudomonas sp. 102515 by PCR
To find out whether Pseudomonas sp. 102515 contains the turnerbaction biosynthetic genes, the tnbA gene was amplified from the genomic DNA of Pseudomonas sp. 102515 using the primers 25 and 26 listed in Additional file 1: Table S2. The PCR product was ligated into the pJET1.2 cloning vector to yield pOKF187 (Table 3), which was subsequently sent out for sequencing (Sequence S1).
Optimization of carotenoid production, extraction and HPLC-MS analysis
For Pseudomonas sp. 102515, different culture media were tested to increase the yield of zeaxanthin diglucoside. Upon the determination of a suitable medium, we investigated the effect of glycerol supplementation as additional carbon source. Cultivation time and temperature were also tested for an improved production of carotenoids. We also analyzed the yield of zeaxanthin diglucoside in heterologous expression systems.
The wild type and engineered strains were typically cultured in 250-mL flasks containing 50 mL of medium. The cultures were centrifuged at 3500 rpm for 8 min to harvest the cells and the cell pellets were suspended in 50 mL of methanol. The cell suspension in methanol was sonicated for 5 min to extract carotenoids. After centrifugation at 3500 rpm for 8 min, the resulting extract was dried in vacuo, and the residues were redissolved in a dimethyl sulfoxide (DMSO)-methanol mixture (10%, v/v) for HPLC-MS analysis with a gradient mobile phase of acetonitrile-water from 50 to 90% over 45 min at 1 mL/min. Low-resolution ESI-MS spectra were obtained on an Agilent 6130 LC-MS to confirm the molecular weights of carotenoids. For co-expression experiments, we used a different HPLC condition in order for the better separation of phytoene, lycopene and beta-carotene. For this HPLC method, we used an isocratic elution system of methanol/tetrahydrofuran (6/4, v/v) at 1 mL/min over 45 min.
Zeaxanthin diglucoside was subsequently purified by HPLC with an Agilent Eclipse XDB-C18 column (5 μm, 250 mm × 4.6 mm) for the standard curve preparation. Purified zeaxanthin diglucoside powder was measured and dissolved in a specific volume of methanol to make a zeaxanthin diglucoside solution. Next, this zeaxanthin diglucoside solution was serially diluted. The absorbance value of each dilution was measured at 456 nm on a UV-Vis spectrophotometer, with methanol as the blank control. Based on the correlation between the absorbance value and concentration, a standard curve of zeaxanthin diglucoside was prepared to calculate the yields in the optimization experiments. All crude extracts were dissolved in methanol and their absorbance values at 456 nm were measured on the UV-Vis spectrophotometer. The yields were calculated based on the measured data and standard curve. All samples were done in three replicates.
Availability of data and materials
The partial 16S rRNA gene sequence of Pseudomonas sp. 102515 is available to the public in NCBI under accession number MK610450. The sequence of the Pscrt biosynthetic gene cluster is available to the public in NCBI under accession number MK613929.
Abbreviations
- DMAPP:
-
Dimethylallyl diphosphate
- DMSO:
-
Dimethyl sulfoxide
- FPP:
-
Farnesyl pyrophosphate
- GGPP:
-
Geranylgeranyl diphosphate
- GPP:
-
Geranyl diphosphate
- IPP:
-
Isopenthyl diphosphate
- IPTG:
-
Isopropyl β-D-1-thiogalactopyranoside
- LC-MS:
-
Liquid chromatography-mass spectrometry
- ROS:
-
Reactive oxygen species
- rRNA:
-
ribosomal RNA
References
Christina A, Christapher V, Bhore SJ. Endophytic bacteria as a source of novel antibiotics: an overview. Pharmacogn Rev. 2013;7(13):11.
Strobel G, Daisy B, Castillo U, Harper J. Natural products from endophytic microorganisms. J Nat Prod. 2004;67(2):257–68.
Staniek A, Woerdenbag HJ, Kayser O. Endophytes: exploiting biodiversity for the improvement of natural product-based drug discovery. J Plant Interact. 2008;3(2):75–93.
Palumbo J, Kobayashi D. Bacterial endophytes and their effects on plants and uses in agriculture. In: Bacon CW, editor. White J, editors. Microbial Endophytes: CRC Press; 2000. p. 213–50.
Mitter B, Brader G, Afzal M, Compant S, Naveed M, Trognitz F, et al. Advances in elucidating beneficial interactions between plants, soil, and bacteria. Advances in Agronomy. 121: Elsevier; 2013. p. 381–445.
Glassner H, Zchori-Fein E, Compant S, Sessitsch A, Katzir N, Portnoy V, et al. Characterization of endophytic bacteria from cucurbit fruits with potential benefits to agriculture in melons (Cucumis melo L.). FEMS Microbiol Ecol. 2015;91(7):fiv074.
Puri A, Padda KP, Chanway CP. Plant growth promotion by endophytic bacteria in nonnative crop hosts. In: Dinesh KM, Annapurma K, editors. . Springer: Endophytes: Crop Productivity and Protection; 2017. p. 11–45.
Ting A, Mah S, Tee C. Prevalence of endophytes antagonistic towards Fusarium oxysporum f. sp. cubense race 4 in various plants. Am Eurasian J Agric Environ Sci. 2009;3(3):399–406.
Miller CM, Miller RV, Garton-Kenny D, Redgrave B, Sears J, Condron MM, et al. Ecomycins, unique antimycotics from Pseudomonas viridiflava. J Appl Microbiol. 1998;84(6):937–44.
Harrison L, Teplow DB, Rinaldi M, Strobel G. Pseudomycins, a family of novel peptides from Pseudomonas syringae possessing broad-spectrum antifungal activity. Microbiology. 1991;137(12):2857–65.
Strobel G, Yang X, Sears J, Kramer R, Sidhu RS, Hess W. Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology. 1996;142(2):435–40.
Frank H, Cogdell R. The photochemistry and function of carotenoids in photosynthesis. In: Yound AJ, Britton G, editors. . Springer: Carotenoids in Photosynthesis; 1993. p. 252–326.
Griffiths M, Sistrom W, Cohen-Bazire G, Stanier R. Function of carotenoids in photosynthesis. Nature. 1955;176(4495):1211.
Britton G. Structure and properties of carotenoids in relation to function. FASEB J. 1995;9(15):1551–8.
Chemler JA, Koffas MA. Metabolic engineering for plant natural product biosynthesis in microbes. Curr Opin Biotechnol. 2008;19(6):597–605.
Olson JA, Krinsky NI. Introduction: the colorful, fascinating world of the carotenoids: important physiologic modulators. FASEB J. 1995;9(15):1547–50.
Fidan O, Zhan J. Reconstitution of medicinally important plant natural products in microorganisms. In: Kermode AR, Jiang L, editors. Molecular Pharming: Applications, Challenges, and Emerging Areas: John Wiley & Sons, Inc; 2018. p. 383–415.
Asker D. High throughput screening and profiling of high-value carotenoids from a wide diversity of bacteria in surface seawater. Food Chem. 2018;261:103–11.
Marz U. The global market for carotenoids FOD025E. BBC Research. 2015:2–5.
Fraser PD, Bramley PM. The biosynthesis and nutritional uses of carotenoids. Prog Lipid Res. 2004;43(3):228–65.
Furubayashi M, Ikezumi M, Takaichi S, Maoka T, Hemmi H, Ogawa T, et al. A highly selective biosynthetic pathway to non-natural C50 carotenoids assembled from moderately selective enzymes. Nat Commun. 2015;6:7534.
Kim SH, Kim MS, Lee BY, Lee PC. Generation of structurally novel short carotenoids and study of their biological activity. Sci Rep. 2016;6:21987.
Verdoes JC, Misawa N, van Ooyen AJ. Cloning and characterization of the astaxanthin biosynthetic gene encoding phytoene desaturase of Xanthophyllomyces dendrorhous. Biotechnol Bioeng. 1999;63(6):750–5.
Hauser E, Kämpfer P, Busse H-J. Pseudomonas psychrotolerans sp. nov. Int J Syst Evol Microbiol. 2004;54(5):1633–7.
Liu R, Zhang Y, Chen P, Lin H, Ye G, Wang Z, et al. Genomic and phenotypic analyses of Pseudomonas psychrotolerans PRS08-11306 reveal a turnerbactin biosynthesis gene cluster that contributes to nitrogen fixation. J Biotechnol. 2017;253:10–3.
Cimermancic P, Medema MH, Claesen J, Kurita K, Brown LCW, Mavrommatis K, et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell. 2014;158(2):412–21.
Sedkova N, Tao L, Rouvière PE, Cheng Q. Diversity of carotenoid synthesis gene clusters from environmental Enterobacteriaceae strains. Appl Environ Microbiol. 2005;71(12):8141–6.
Cunningham FX, Sun Z, Chamovitz D, Hirschberg J, Gantt E. Molecular structure and enzymatic function of lycopene cyclase from the cyanobacterium Synechococcus sp strain PCC7942. Plant Cell. 1994;6(8):1107–21.
Choudhari SM, Ananthanarayan L, Singhal RS. Use of metabolic stimulators and inhibitors for enhanced production of β-carotene and lycopene by Blakeslea trispora NRRL 2895 and 2896. Bioresour Technol. 2008;99(8):3166–73.
Yoon S-H, Lee S-H, Das A, Ryu H-K, Jang H-J, Kim J-Y, et al. Combinatorial expression of bacterial whole mevalonate pathway for the production of β-carotene in E. coli. J Biotechnol. 2009;140(3):218–26.
Johnson EJ. The role of carotenoids in human health. Nutr Clin Care. 2002;5(2):56–65.
Rao AV, Rao LG. Carotenoids and human health. Pharmacol Res. 2007;55(3):207–16.
Tatsuzawa H, Maruyama T, Misawa N, Fujimori K, Nakano M. Quenching of singlet oxygen by carotenoids produced in Escherichia coli-attenuation of singlet oxygen-mediated bacterial killing by carotenoids. FEBS Lett. 2000;484(3):280–4.
Ito T. Cellular and subcellular mechanisms of photodynamic action: the 1O2 hypothesis as a driving force in recent research. Photochem Photobiol. 1978;28(4–5):493–506.
Weishaupt KR, Gomer CJ, Dougherty TJ. Identification of singlet oxygen as the cytotoxic agent in photo-inactivation of a murine tumor. Cancer Res. 1976;36(7 Part 1):2326–9.
Cazzonelli CI. Carotenoids in nature: insights from plants and beyond. Funct Plant Biol. 2011;38(11):833–47.
Misawa N, Nakagawa M, Kobayashi K, Yamano S, Izawa Y, Nakamura K, et al. Elucidation of the Erwinia uredovora carotenoid biosynthetic pathway by functional analysis of gene products expressed in Escherichia coli. J Bacteriol. 1990;172(12):6704–12.
Takaichi S, Maoka T, Akimoto N, Khan ST, Harayama S. Major carotenoid isolated from Paracoccus schoinia NBRC 100637T is adonixanthin diglucoside. J Nat Prod. 2006;69(12):1823–5.
Takaichi S, Maoka T, Hanada S, Imhoff JF. Dihydroxylycopene diglucoside diesters: a novel class of carotenoids from the phototrophic purple sulfur bacteria Halorhodospira abdelmalekii and Halorhodospira halochloris. Arch Microbiol. 2001;175(3):161–7.
Seo YB, Choi SS, Nam SW, Lee JH, Kim YT. Cloning and characterization of the zeaxanthin glucosyltransferase gene (crtX) from the astaxanthin-producing marine bacterium. Paracoccus haeundaensis J Microbiol Biotechnol. 2009;19(12):1542–6.
Tuveson R, Larson R, Kagan J. Role of cloned carotenoid genes expressed in Escherichia coli in protecting against inactivation by near-UV light and specific phototoxic molecules. J Bacteriol. 1988;170(10):4675–80.
Osawa A, Harada H, Choi S-K, Misawa N, Shindo K. Production of caloxanthin 3′-β-D-glucoside, zeaxanthin 3,3′-β-D-diglucoside, and nostoxanthin in a recombinant Escherichia coli expressing system harboring seven carotenoid biosynthesis genes, including crtX and crtG. Phytochemistry. 2011;72(8):711–6.
Richter TK, Hughes CC, Moore BS. Sioxanthin, a novel glycosylated carotenoid, reveals an unusual subclustered biosynthetic pathway. Environ Microbiol. 2015;17(6):2158–71.
Hundle BS, O'Brien DA, Alberti M, Beyer P, Hearst JE. Functional expression of zeaxanthin glucosyltransferase from Erwinia herbicola and a proposed uridine diphosphate binding site. Proc Natl Acad Sci U S A. 1992;89(19):9321–5.
Achouak W, Sutra L, Heulin T, Meyer JM, Fromin N, Degraeve S, et al. Pseudomonas brassicacearum sp. nov. and Pseudomonas thivervalensis sp. nov., two root-associated bacteria isolated from Brassica napus and Arabidopsis thaliana. Int J Syst Evol Microbiol. 2000;50(1):9–18.
Asif H, Studholme DJ, Khan A, Aurongzeb M, Khan IA, Azim MK. Comparative genomics of an endophytic Pseudomonas putida isolated from mango orchard. J Genet Mol Biol. 2016;39(3):465–73.
Eljounaidi K, Lee SK, Bae H. Bacterial endophytes as potential biocontrol agents of vascular wilt diseases-review and future prospects. Biol Control. 2016;103:62–8.
Fukaya Y, Takemura M, Koyanagi T, Maoka T, Shindo K, Misawa N. Structural and functional analysis of the carotenoid biosynthesis genes of a Pseudomonas strain isolated from the excrement of autumn darter. Biosci Biotechnol Biochem. 2018;82(6):1043–52.
Lamb TG, Tonkyn DW, Kluepfel DA. Movement of Pseudomonas aureofaciens from the rhizosphere to aerial plant tissue. Can J Microbiol. 1996;42(11):1112–20.
Miliute I, Buzaite O, Baniulis D, Stanys V. Bacterial endophytes in agricultural crops and their role in stress tolerance: a review. Zemdirbyste. 2015;102(4):465–78.
Ulrich K, Ulrich A, Ewald D. Diversity of endophytic bacterial communities in poplar grown under field conditions. FEMS Microbiol Ecol. 2008;63(2):169–80.
Rediers H, Bonnecarrere V, Rainey PB, Hamonts K, Vanderleyden J, De Mot R. Development and application of a dapB-based in vivo expression technology system to study colonization of rice by the endophytic nitrogen-fixing bacterium Pseudomonas stutzeri A15. Appl Environ Microbiol. 2003;69(11):6864–74.
Lally RD, Galbally P, Moreira AS, Spink J, Ryan D, Germaine KJ, et al. Application of endophytic Pseudomonas fluorescens and a bacterial consortium to Brassica napus can increase plant height and biomass under greenhouse and field conditions. Front Plant Sci. 2017;8:2193.
Frommel MI, Nowak J, Lazarovits G. Growth enhancement and developmental modifications of in vitro grown potato (Solanum tuberosum spp. tuberosum) as affected by a nonfluorescent Pseudomonas sp. Plant Physiol. 1991;96(3):928–36.
Glick BR, Penrose DM, Li J. A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J Theor Biol. 1998;190(1):63–8.
Hallmann J, Quadt-Hallmann A, Mahaffee W, Kloepper J. Bacterial endophytes in agricultural crops. Can J Microbiol. 1997;43(10):895–914.
Bangera MG, Thomashow LS. Characterization of a genomic locus required for synthesis of the antibiotic 2, 4-diacetylphloroglucinol by the biological control agent Pseudomonas fluorescens Q2-87. Mol Plant-Microbe Interact. 1996;9(2):83–90.
Madhaiyan M, Poonguzhali S, Senthilkumar M, Seshadri S, Chung H, Jinchul Y, et al. Growth promotion and induction of systemic resistance in rice cultivar Co-47 (Oryza sativa L.) by Methylobacterium spp. Bot Bull Acad Sinica. 2004:45.
O'sullivan DJ, O'Gara F. Traits of fluorescent Pseudomonas spp. involved in suppression of plant root pathogens. Microbiol Rev. 1992;56(4):662–76.
Wicaksono WA, Jones EE, Casonato S, Monk J, Ridgway HJ. Biological control of Pseudomonas syringae pv. actinidiae (Psa), the causal agent of bacterial canker of kiwifruit, using endophytic bacteria recovered from a medicinal plant. Biol Control. 2018;116:103–12.
Adorada DL, Stodart BJ, Tpoi RP, Costa SS, Ash GJ. Bacteria associated with sheath browning and grain discoloration of rice in East Timor and implications for Australia’s biosecurity. Australas Plant Dis Notes. 2013;8(1):43–7.
Kodama K, Kimura N, Komagata K. Two new species of Pseudomonas: P. oryzihabitans isolated from rice paddy and clinical specimens and P. luteola isolated from clinical specimens. Int J Syst Evol Microbiol. 1985;35(4):467–74.
Santo CE, Lin Y, Hao X, Wei G, Rensing C, Grass G. Draft genome sequence of Pseudomonas psychrotolerans L19, isolated from copper alloy coins. J Bacteriol. 2012;194(6):1623–4.
Simmon KE, Croft AC, Petti CA. Application of SmartGene IDNS software to partial 16S rRNA gene sequences for a diverse group of bacteria in a clinical laboratory. J Clin Microbiol. 2006;44(12):4400–6.
Xie G-l, Soad A, Swings J, Mew T. Diversity of gram negative bacteria antagonistic against major pathogens of rice from rice seed in the tropic environment. J Zhejiang Univ Sci A. 2003;4(4):463–8.
Bhagat J, Kaur A, Chadha BS. Single step purification of asparaginase from endophytic bacteria Pseudomonas oryzihabitans exhibiting high potential to reduce acrylamide in processed potato chips. Food Bioprod Process. 2016;99:222–30.
Cunningham FX, Lee H, Gantt E. Carotenoid biosynthesis in the primitive red alga Cyanidioschyzon merolae. Eukaryot Cell. 2007;6(3):533–45.
Cunningham FX Jr, Gantt E. A study in scarlet: enzymes of ketocarotenoid biosynthesis in the flowers of Adonis aestivalis. Plant J. 2005;41(3):478–92.
Ren Q, Van Beilen JB, Sierro N, Zinn M, Kessler B, Witholt B. Expression of PHA polymerase genes of Pseudomonas putida in Escherichia coli and its effect on PHA formation. Antonie Van Leeuwenhoek. 2005;87(2):91.
Hettwer U, Jaeckel FR, Boch J, Meyer M, Rudolph K, Ullrich MS. Cloning, nucleotide sequence, and expression in Escherichia coli of levansucrase genes from the plant pathogens Pseudomonas syringae pv. glycinea and P. syringae pv. phaseolicola. Appl Environ Microbiol. 1998;64(9):3180–7.
Beuttler H, Hoffmann J, Jeske M, Hauer B, Schmid RD, Altenbuchner J, et al. Biosynthesis of zeaxanthin in recombinant Pseudomonas putida. Appl Microbiol Biotechnol. 2011;89(4):1137–47.
Mi J, Becher D, Lubuta P, Dany S, Tusch K, Schewe H, et al. De novo production of the monoterpenoid geranic acid by metabolically engineered Pseudomonas putida. Microb Cell Factories. 2014;13(1):1.
Ser H-L, Law JW-F, Chaiyakunapruk N, Jacob SA, Palanisamy UD, Chan K-G, et al. Fermentation conditions that affect clavulanic acid production in Streptomyces clavuligerus: a systematic review. Front Microbiol. 2016;7:522.
Barbirato F, Grivet JP, Soucaille P, Bories A. 3-Hydroxypropionaldehyde, an inhibitory metabolite of glycerol fermentation to 1,3-propanediol by enterobacterial species. Appl Environ Microbiol. 1996;62(4):1448–51.
Zhang Y, Taiming L, Liu J. Low temperature and glucose enhanced T7 RNA polymerase-based plasmid stability for increasing expression of glucagon-like peptide-2 in Escherichia coli. Protein Expr Purif. 2003;29(1):132–9.
Miller C, Pettee B, Zhang C, Pabst M, McLean J, Anderson A. Copper and cadmium: responses in Pseudomonas putida KT2440. Lett Appl Microbiol. 2009;49(6):775–83.
de Oliveira Costa LE, de Queiroz MV, Borges AC, de Moraes CA, de Araújo EF. Isolation and characterization of endophytic bacteria isolated from the leaves of the common bean (Phaseolus vulgaris). Braz J Microbiol. 2012;43(4):1562–75.
Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual: cold Spring Harbor laboratory press; 1989.
Barghouthi SA. A universal method for the identification of bacteria based on general PCR primers. Indian J Microbiol. 2011;51(4):430–44.
Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4(4):406–25.
Rice MC, Norton JM, Valois F, Bollmann A, Bottomley PJ, Klotz MG, et al. Complete genome of Nitrosospira briensis C-128, an ammonia-oxidizing bacterium from agricultural soil. Stand Genomic Sci. 2016;11(1):46.
Iwasaki K, Uchiyama H, Yagi O, Kurabayashi T, Ishizuka K, Takamura Y. Transformation of Pseudomonas putida by electroporation. Biosci Biotechnol Biochem. 1994;58(5):851–4.
Acknowledgements
The authors are thankful to Dr. Paul Wolf and Michael B. Piep at Utah State University Intermountain Herbarium for their help to identify and deposit the plant specimen. We thank Dr. Jens Schrader and Dr. Josef Altenbuchner at Dechema-Forschungsinstitut and University of Stuttgart, respectively, for providing the pMIS1-mva-ges expression plasmid. We are also grateful to Dr. Charles Miller at Utah State University for generously providing Pseudomonas putida KT2440 and Dr. Francis X Cunningham Jr. at the University of Maryland for sharing the carotenoid producing plasmids through Addgene.
Funding
This work was financially supported by a Grant-In-Aid (16GRNT26430067) from the American Heart Association.
Author information
Authors and Affiliations
Contributions
JZ supervised this study. OF performed the experiments. Both authors analyzed the data and wrote the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Table S1. Recipes of the media used in this study. Table S2. Oligonucleotides used in this study. Figure S1. Standard curve for zeaxanthin diglucoside using UV spectrophotometer at 456 nm. Figure S2. The effects of different media on the yield of zeaxanthin diglucoside in the culture tubes (5 mL). (DOCX 114 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Fidan, O., Zhan, J. Discovery and engineering of an endophytic Pseudomonas strain from Taxus chinensis for efficient production of zeaxanthin diglucoside. J Biol Eng 13, 66 (2019). https://doi.org/10.1186/s13036-019-0196-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13036-019-0196-x