
Abstract
Purpose
Temporomandibular joint osteoarthritis (TMJOA) is a common disease that negatively affects the life quality of human beings. Circadian rhythm acts an important role in life activities. However, whether the clock genes are rhythmic expressed in mandibular condylar chondrocytes, or the clock genes have an effect on the progression of TMJOA remains unknown. In this study, we aim to explore expression of clock genes and regulatory mechanism of TMJOA in rat mandibular condylar chondrocytes.
Methods
After synchronized by dexamethasone, the expression of core clock genes Per1, Per2, Clock, Cry1, Cry2 and Bmal1 and cartilage matrix degrading factor gene Mmp13 were analyzed in mandibular condylar chondrocytes every 4 h with RT-qPCR. The mandibular condylar chondrocytes were stimulated with IL-1β, and expression of Per1, Mmp13, P65 and p-P65 was assessed by RT-qPCR and Western blot. Sh-Per1 lentivirus was used to assess the effect of clock gene Per1 in IL-1β-induced chondrocytes, and expression of Mmp13, P65 and p-P65 was measured. After establishing a rat TMJOA model using unilateral anterior crossbite (UAC), micro-CT, H & E, Alcian Blue & Nuclear Fast Red and Safranin O & Fast Green, cartilage thickness was utilized to assess the damage of cartilage and subchondral bone. Immunohistochemistry of PER1, MMP13 and P65 was performed in condylar sections.
Results
All core clock genes and Mmp13 were rhythmically expressed. And Mmp13 expression curve was closed in phase and amplitude with Per1. After stimulation with IL-1β, the expression of MMP13, PER1 and P65 and ratio of p-P65/P65 increased in condylar chondrocytes. After Per1 was down-regulated in condylar chondrocytes, the expression of MMP13 and P65 and ratio of p-P65/P65 decreased. Compared with the condyles of Sham group, the bony parameters of UAC group were significantly worse. The thickness of cartilage in UAC group significantly reduced. The modified Mankin scores and the expression of PER1, MMP13 and P65 in cartilage of UAC group significantly increased compared with Sham group.
Conclusion
Core clock genes and Mmp13 are rhythmic expressed in rat mandibular condylar chondrocytes. PER1 can regulate the expression of MMP13 through NF-κB pathway in IL-1β-induced mandibular condylar chondrocytes.
Similar content being viewed by others


Introduction
Temporomandibular joint osteoarthrosis (TMJOA) is a common manifestation of temporomandibular joint disorder that often presents with bone resorption, condylar cysts or osteophyte formation [1, 2]. Severe cases can lead to joint dyskinesia, mouth restriction, etc. Condylar resorption can even lead to mandibular retraction and other facial deformities, seriously affecting the living quality and psychological health of patients [3]. Its etiology includes local factors such as injury, parafunction, occlusion and joint overload [4], as well as systemic factors such as age, sex, autoimmune diseases and hormonal disorders [5]. Mandibular condylar chondrocytes are the only cells in condylar cartilage, so the factors affecting the proliferation, differentiation and mineralization of condylar chondrocytes will affect the growth and reconstruction of condyles, which can eventually be manifested as condylar resorption [6,7,8]. Early cartilage degeneration is caused by metabolic or local mechanical factors, which trigger an immune response. And then, immune cells release cytokines such as IL-1β, TNF-α, etc., and other inflammatory factors to initiate the inflammatory response, which increases the synthesis of cartilage matrix degrading factors, including matrix metalloproteinases (MMPs) and prostaglandin E (PGE). These factors further aggravate the decomposition of cartilage and eventually lead to the degeneration of articular cartilage, which, in turn, leads to the resorption and degeneration of subchondral bone [2, 9, 10].
Circadian rhythm is a continuously biological rhythm regulated by the clock genes, with a cycle of about 24 h. It is a periodic oscillation of biochemical activities in the organism, regulating human body temperature, hormone, sleep, blood pressure and other life activities [11, 12]. There is also a circadian clock in the peripheral tissues, which is regulated by the central clock. After the central clock is destroyed, the peripheral clock will gradually disappear [13]. The circadian rhythm is regulated by multiple clock genes. At present, more than ten core clock genes have been found, including Bma1, Clock, Cry1, Cry2, Per1, Per2, Rev-erb α, ROR α, etc. Bmal1 and Clock are positive regulatory clock genes, and Cry1, Cry2, Per1, Per2, Rev-erb α and ROR α are negative regulatory genes [14, 15]. There are rhythmic changes in cartilage metabolism, such as the proliferation of chondrocytes mainly occurring in the early morning and mineralization mainly occurring at night. The expression of various matrix proteins in cartilage, such as hyaluronic acid (HA) and collagen II, also is rhythmic expressed [16,17,18]. Studies have shown that compared with the knee cartilage of normal individuals, the expression of NR1D1 and BMAL1 in the knee cartilage of osteoarthritis (OA) patients is relatively low, and the expression of Bmal1 is also lower in the OA rat model, which is negatively correlated with the severity of OA [19, 20]. In the OA mice model, it was found that the expression of clock gene Per2 reduced [21]. The clock gene Cry1 may be involved in anti-COL2 antibody-induced mouse OA model and exacerbate the progression of osteoarthritis [22].
There are few studies on whether there is a peripheral clock in the mandibular condylar chondrocytes, and whether the clock gene is involved in the progression of TMJOA. This study is aimed to explore whether clock genes are rhythmic expressed in condylar chondrocytes in vitro, investigate whether clock genes are abnormally expressed through TMJOA model in vitro and vivo and preliminarily explore possible mechanism.
Materials and methods
Isolation and culture of mandibular condylar chondrocytes
The temporomandibular joint cartilage was isolated from 15 male Sprague Dawley rats (3–5 d) in the present study. After digested with 0.25% trypsin and 0.1 collagenase type I for 2 h at 37 °C, the chondrocytes were collected by centrifugation at 1000 rpm/min and cultured with DMEM (Gibco, USA) containing 20% fetal bovine serum (FBS, Gibco, USA), 100-IU penicillin and 100-μg/mL streptomycin (Gibco, USA) in incubator at 37 °C with 5% CO2. Cells in passage 2 were used for further experiments.
Circadian rhyme synchronization of mandibular condylar chondrocytes
After passaging the cells to passage 2, the chondrocytes were treated with a known synchronizer of peripheral clock (100-nM dexamethasone) for 1 h [21, 23]. And then, the total RNA was extracted from cultured chondrocytes using TRIzol (Invitrogen, USA) every 4 h and reverse transcribed to cDNA using PrimeScript™ RT Master Mix (Takara, Japan). All the cDNA samples were subsequently conducted to polymerase chain reaction with Hieff UNICON® Universal Blue qPCR SYBR Green Master Mix (Yeasen, China). The expression of clock genes (Per1, Per2, Clock, Cry1, Cry2 and Bmal1) and Mmp13 was calculated by using cycle threshold method (2−△△Ct). The primer sequence was attached in Additional file 1. And the internal control gene was Gapdh. All experiments were repeated three times. The relative expression of genes at a given time was normalized as a percent value of mean level across all the time points. The results were fitted with trigonometric functions by Origin Pro 10.0 (OriginLab, USA).
Cell treatment and gene inhibition
After synchronized with 100-nM dexamethasone for 1 h, the mandibular chondrocytes were treated with 10-ng/mL IL-1β for 24 h [24, 25] to simulate temporomandibular joint osteoarthritis in vitro. Total RNA and protein were extracted from IL-1β-treated cells. In order to inhibit the expression of Per1, the chondrocytes were transfected with sh-Per1 and sh-control vectors via lentiviruses (Genechem, China) for 24 h. And the cells were stimulated with IL-1β following inhibition of Per1 by lentiviruses for next 24 h. After that, total RNA and protein were extracted at the same time point during a day. Western blot and RT-qPCR were performed to detect the expression of Mmp13 and Per1.
Western blot
The collected protein samples were quantified using BCA method (Yeasen, China) and separated by 10% SDS-PAGE. Then, the protein was transferred onto the PVDF membrane (Millipore, Germany). After blocking with 5% non-fat milk at room temperature for 1 h, the membrane was incubated with primary antibodies including rabbit anti-PER1 (Affinity, USA, 1:800), rabbit anti-MMP13 (Proteintech, China, 1:1000), rabbit anti-NF-kB p65 (Affinity, USA, 1:500), rabbit anti-Phospho-NF-kB p65 (Ser536) (Affinity, USA, 1:500) and rabbit anti-GAPDH (Affinity, USA, 1:2000) at 4 °C for 16 h. Following the washing with 0.1% TBST, the secondary antibody (Beyotime, China) was incubated with the membrane. After washing with 0.1% TBST, a chemiluminescence ECL system (Millipore, Germany) was used to visualize the immunoreactive proteins.
Unilateral anterior crossbite (UAC)-induced TMJOA
All operations are carried out in accordance with animal ethics (approved by Ethics Committee of Sun Yat-sen University, China). Fourteen male Sprague Dawley rats (6 w, 150–160 g) were randomly divided into experimental group and control group, with seven rats in each group. Prior to the start of the experiment, the rats were acclimated for 1 week under a 12-h light/12-h dark environment (8:00 AM–8:00 PM light and 8:00 PM–8:00 AM dark), and the conditions for the formal experiment are consistent with the conditions stated above. The animals were housed with two in a cage in a room maintained at 21–24 °C. All animals were fed a regular laboratory diet with daily changing soft feed and sterile water. After intraperitoneal injection of 1% pentobarbital sodium (0.3 mL/100-g body weight) as anesthesia, we used 25# cow lacteal needle to make the upper anterior tooth restoration and 20# needle to make the lower anterior tooth restoration. The lower restoration made the angle of 135° with the upper anterior tooth restoration, forming unilateral anterior crossbite according to Wang’s method [26]. All the rats ate soft feed after surgery, periodically check whether the restoration has fallen off and bond in time if it falls off. After 8 weeks, the rats were sacrificed, and the condyles were isolated at the same time point during a day for subsequent experiments.
Micro-CT
Condyles of rats in each groups were collected and fixed with 4% paraformaldehyde solution for 24 h and then scanned by micro-CT (70 kV, 114 mA, 6.8 μm, Scanco.μCT 50). The parameters including bone surface/bone volume (BS/BV), bone volume fraction (BV/TV), trabecular thickness (Tb.Th), trabecular separation (Tb.Sp) and trabecular number (Tb.N) were analyzed. Three-dimensional images were reconstructed by μCT evaluation Program V6.6 for morphological assessment.
Histological analysis
Condyles of rats were demineralized in 10% ethylenediaminetetraacetic acid for 8 weeks and then embedded with paraffin after dehydration. About 5-μm mid-sagittal sections parallel to condyle were cut and dewaxed in xylene, hydrated in gradient alcohol. H & E, Alcian Blue & Nuclear Fast Red and Safranin O & Fast Green staining were applied according to protocol. Modified Mankin score and cartilage thickness [27, 28] were calculated according to the histological sections to assess severity of TMJOA.
Immunohistochemistry
Dewaxing and hydration were performed as we described previously. We used 3% hydrogen peroxide to block endogenous peroxidase activity for 15 min. And the sections were antigen-retrieved in citrate solution by microwave. Following by serum blocking for 1 h and primary antibody including rabbit anti-PER1 (Affinity, USA, 1:100), rabbit anti-MMP13 (Proteintech, China, 1:300) and rabbit anti-NF-κB p65 (Affinity, USA, 1:100) at 4 °C were incubated overnight individually. Secondary antibody was incubated for 1 h after PBS washing. Finally, we used DAB method to visualize the immunoreactive cells and hematoxylin to stain the nucleus. We used ImageJ 1.52i (National Institutes of Health, Germany) to measure the average optical density in anterior, middle and posterior fields of each section.
Statistical analysis
Data were performed by Prism 8.0 (GraphPad, USA). Student’s t-test (two-tailed) was used to compare means of two groups, and one-way ANOVA was used to compare means of three or more groups. Test level was α = 0.05. All the quantitative data were presented as mean ± SEM in figures.
Results
Expression of core clock genes and Mmp13 is rhythmic
In order to explore the existence of peripheral clock in mandibular condylar chondrocytes, we used the dexamethasone to synchronize the circadian rhythm and measured the mRNA expression of core clock genes and Mmp13. All core clock genes and Mmp13 (Fig. 1) were rhythmically expressed. We calculated the functions of all the genes using Origin software and found that the function of Per1 (Fig. 1B) is closest to the function of Mmp13 (Fig. 1A) in terms of phase and amplitude, which indicates a possible relationship between clock gene Per1 and Mmp13. Therefore, we chose the clock gene Per1 as the target gene for further study.

Mmp13 and core clock genes are rhythmically expressed in rat mandibular condylar chondrocytes after synchronization of dexamethasone. A Expression of Mmp13 within 24 h. B Expression of Per1 within 24 h. C Expression of Per2 within 24 h. D Expression of Clock within 24 h. E Expression of Cry1 within 24 h. F Expression of Cry2 within 24 h. G Expression of Bmal1 within 24 h
IL-1β increases the expression of Per1 and Mmp13 and activates NF-κB pathway in mandibular condylar chondrocytes
To simulate TMJOA in vitro, we added IL-1β to the rat mandibular chondrocytes. After stimulation with IL-1β, the mRNA and protein expression levels of Mmp13 and Per1 and protein expression level of P65 and p-P65 were measured using RT-qPCR and Western blot. As shown in Fig. 2A–C, mRNA and protein levels of Mmp13 and Per1 in chondrocytes significantly increased compared to control group. It indicates that clock gene Per1 may have a potential positive correlation with Mmp13. And we examined the NF-κB pathway-related proteins in the IL-1β-induced chondrocytes. As shown in Fig. 2C and D, the protein expression level of P65 (P < 0.01) and ratio of p-P65/P65 (P < 0.05) significantly increased after stimulation of IL-1β.

IL-1β increases the expression of Per1 and Mmp13 and activates the NF-κB pathway in mandibular condylar chondrocytes. A The mRNA levels of Mmp13 and Per1 after stimulation of IL-1β in chondrocytes. B, C Protein level and analysis of MMP13, PER1, P65 and p-P65 after stimulation of IL-1β in chondrocytes. D The ratio of p-P65/P65 in two groups. Values are displayed as the mean ± SEM. *P < 0.05 and **P < 0.01
Down-regulation of Per1 inhibits the expression of Mmp13 and NF-κB pathway induced by IL-1β
To further explore the correlation between clock gene Per1 and Mmp13, and potential regulatory mechanism, we examined the expression of Mmp13 and NF-κB pathway-related proteins with Per1 down-regulation. After down-regulation of Per1 by lentiviruses, the mRNA and protein levels of Per1 in IL-1β-induced mandibular condylar chondrocytes were significantly reduced compared with sh-control+IL-1β group (Fig. 3A-B, mRNA level P < 0.01. Protein level P < 0.05). The efficacy of sh-Per1 was verified. Compared with the sh-control group, sh-Per1 lentivirus significantly reduced the mRNA (P < 0.001) and protein (P < 0.01) levels of Mmp13. And down-regulation of Per1 also significantly inhibited the expression of P65 (Fig. 3B-C) and ratio of p-P65/P65 (Fig. 3D). Take together, these results indicate that clock gene Per1 can activate the expression of Mmp13 through NF-κB pathway in IL-1β-induced mandibular condylar chondrocytes.

Down-regulation of Per1 inhibits the expression of Mmp13 and NF-κB pathway induced by IL-1β. A RT-qPCR analysis of Per1 and Mmp13 in chondrocytes of NC, IL-1β, sh-control+IL-1β and sh-Per1+IL-1β groups. B Western blot analysis of PER1, MMP13, P65 and p-P65 in four groups. C The relative protein expression level of PER1, MMP13, P65 and p-P65 in four groups. D The ratio of p-P65/P65 in four groups. Values are displayed as the mean ± SEM. *P < 0.05, **P < 0.01 and ***P < 0.001
Degenerative lesions were observed in unilateral anterior crossbite TMJOA rat model
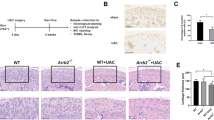
To explore the relationship between PER1 and MMP13 in vivo, we established a TMJOA rat model through unilateral anterior crossbite. As shown in micro-CT analysis, there were obvious bone lesions in 3D reconstruction images of UAC group. Both the thickness of the cartilage layer and the density of the subchondral trabeculae are significantly reduced in UAC group compared with Sham group (Fig. 4A). Similarly, quantitative evaluation results (Fig. 4B) including BS/BV, BV/TV, Tb.SP and Tb.N in UAC group significantly decreased compared with Sham group. The Tb.Th in UAC group was significantly higher than the Sham group. As Fig. 4D shown, the thickness of cartilage was significantly thinner (P < 0.05) in UAC group than Sham group. The modified Mankin score (Fig. 4E) in UAC group was also significantly higher than the Sham group (P < 0.01).

Degenerative lesions are observed in unilateral anterior crossbite TMJOA rat model. A Three-dimensional reconstruction of condyles in UAC group and Sham group. B Quantitative evaluation including BS/BV, BV/TV, Tb.Th, Tb.Sp and Tb.N of two groups (n = 3). C H & E, Alcian Blue & Nuclear Fast Red and Safranin O & Fast Green staining of two groups (n = 4). D The thickness of cartilage in two groups (n = 4). E The modified Mankin scores of two groups (n = 4). Values are displayed as the mean ± SEM. *P < 0.05, **P < 0.01 and ***P < 0.001
The expression of PER1, MMP13 and P65 increased in the mandibular condylar cartilage of UAC rats
As Fig. 5B shown, compared with Sham group, the matrix degrading factor MMP13 significantly increased in mandibular condylar cartilage of UAC rats (P < 0.05). And clock protein PER1 also significantly increased in UAC group (P < 0.01). P65, the core protein of NF-κB pathway, significantly increased in cartilage of UAC group (P < 0.05), which is similar to the results of experiments in vitro.

Expression of PER1, MMP13 and P65 increases in UAC-induced TMJOA rats. A Immunohistochemical staining of PER1, MMP13 and P65 in condyles of UAC group and Sham group. B The average optical density of PER1, MMP13 and P65 of temporomandibular cartilage in UAC and Sham groups (n = 4). Values are displayed as the mean ± SEM. *P < 0.05 and **P < 0.01
Discussion
After three-dimensional cultivation of mouse chondrocytes, a study found through gene microarray analysis that the clock genes Per1, Per2 and Clock were expressed in chondrocytes and exhibited rhythmic expression [29]. Several studies have shown that circadian rhythm also exists in cartilage metabolism. The proliferation of chondrocytes and subchondral mineralization activities are rhythmic [16,17,18]. Mice with specific Bmal1-knockout chondrocytes show the disappearance of circadian rhythm, progressive injury of joints and decrease in bone mass of the whole body [19, 30].
Matrix metalloproteinase-13 (MMP13), mainly secreted by chondrocytes, is the most powerful COL2-degrading enzyme known in matrix metalloproteinases [31]. As mentioned above, during the decompensation phase of TMJOA, the synthesis of matrix metalloproteinases increases, a large amount of cartilage matrix is decomposed and the activity of MMP13 in chondrocytes during the progression of OA is significantly increased [32]. And MMP13 is also an important marker of terminal hypertrophy differentiation of chondrocytes [33].
This study found that Mmp13 also showed rhythmic oscillation in mandibular condylar chondrocytes, suggesting that clock genes may affect the expression of Mmp13. Compared with several core clock genes, the phase of the expression curve of Mmp13 is the closest to that of clock gene Per1. Therefore, we speculate that the expression of Per1 may potentially affect the expression of Mmp13, and we chose Per1 among the core clock genes for subsequent research.
Among various cytokines, IL-1β plays a crucial role in the pathogenesis of TMJOA. It can initiate an increase in the expression of matrix degrading factors including MMP3 and MMP13 in chondrocytes, making it an ideal cytokine for modeling TMJOA in vitro [34]. After stimulation with IL-1β concentration of 10 ng/mL for 24 h [24, 25], the expression of Mmp13 significantly increased in mRNA and protein levels.
UAC rat model is a classic TMJOA modeling method, and multiple studies have confirmed that it can successfully lead to TMJOA [26, 35, 36]. In our study, the three-dimensional morphology of the condyles in the UAC group was worse than that in Sham group, with a decrease in trabecular density and bone mass of the condyles. In terms of histopathology, the modified Mankin score and thickness of cartilage of UAC group also significantly decreased compared to the Sham group.
The previous studies have shown that compared with normal individuals, the expression of clock gene BMAL1 in the knee cartilage of OA patients is lower [19], and the expression of PER2 increased in OA cartilage [37]. Clock genes play a crucial role in the occurrence and development of OA. Per1 can inhibit the differentiation of pre-chondrocytes into chondrocytes by suppressing the expression of Sox6 [38]. Silencing Bmal1 in mouse articular chondrocytes resulted in decreased expression levels of p-SMAD2/3 and NFATC2, as well as decreased expression of chondroprotective factors and matrix-related genes such as Sox9, Acan and Col2a1, but increased levels of p-SMAD1/5, indicating that the clock gene Bmal1 is involved in the maintenance and repair of cartilage. Disrupted circadian rhythms are a risk factor for OA and other joint diseases [19]. Specific knockout of Bmal1 in cartilage promotes cartilage degradation and chondrocyte apoptosis in OA mice, inhibits chondrocyte synthetic metabolism and accelerates cartilage degeneration and osteophyte formation during OA progression [39]. Silencing NR1D1 or BMAL1 in human knee joint chondrocytes affects the TGFβ signaling pathway, thereby affecting cartilage homeostasis [20]. In human OA cartilage, the expression of CRY2 increases, and it plays a positive role in maintaining the extracellular matrix homeostasis of chondrocytes [40].
We also found the expression of clock gene Per1 increased significantly in TMJOA rat model in vitro and in vivo. After down-regulation of Per1 in chondrocytes using lentivirus in vitro, the expression of Mmp13 also reduced, suggesting that Per1 may participate in the occurrence and development of TMJOA by regulating the expression of Mmp13. In a study, chronic sleep deprivation was found to disrupt the normal rhythm of Wistar rats, leading to TMJOA-like lesions such as osteophyte formation. Increased expression of IL-6 and p-ERK was observed in the mandibular condylar cartilage, while Bmal1 expression was decreased. Overexpression of Bmal1 in condylar chondrocytes suppressed the ERK pathway, indicating that Bmal1 in chondrocytes can regulate the synthesis and secretion of IL-6 through the MAPK/ERK pathway, thereby inducing cartilage matrix degradation and participating in the pathological process of TMJOA [41]. During low-intensity pulsed ultrasound (LIPUS) treatment in rats with TMJOA, an increase in Per2 expression was detected through RNA-seq [42]. Loss of Bmal1 down-regulated the expression of cartilage-related genes Prg4, Sox9 and Col7a1 in bone marrow mesenchymal stem cells (BMSCs), while Bmal1 enhanced the migration capability of BMSCs and attenuated age-related temporomandibular joint osteophyte degeneration, indicating the crucial role of circadian rhythm in maintaining condylar osteochondral homeostasis [43].
The IHC results showed that both PER1 and P65 increased in the cartilage of condyles in UAC rats. And after down-regulation of Per1 in chondrocytes in vitro, the expression of P65 and p-P65/P65 also reduced. Take these results together, as Fig. 6 shown, clock gene Per1 can regulate the expression of MMP13 through NF-κB pathway and further to regulate the progression of TMJOA. NF-κB family plays an important role in inflammation, cell differentiation and proliferation. Human NF-κB sub-family includes five types of NF-κB transcription factors: RelA/p65, NF-κB1/p105, NF-κB2/p100, c-Rel and RelB. P65 is the core protein of NF-κB pathway. P65 and P50 can form a protein dimer. When P65 is phosphorylated, the dimer will enter the nucleus to initiate the transcription and expression of downstream genes [44, 45]. NF-κB pathway is also one of the most important signaling pathways that regulate cartilage metabolism. Activating NF-κB pathway can promote inflammation of cartilage, trigger chondrocyte apoptosis and enhance the production of matrix-degrading proteinases such as MMP1, MMP3, MMP13, ADAMTS4 and ADAMTS5, which lead to the erosion of cartilage [46,47,48].

Schematic diagram of PER1-mediated increasing expression of MMP13 by modulating NF-κB pathway in rat mandibular condylar chondrocytes
PER1 is a core molecular regulator of circadian rhythm in organisms. CLOCK and BMAL1 form a positive regulatory complex that activates a series of downstream genes, including PER1 itself. As a negative regulator of circadian rhythm, PER1, in turn, inhibits the expression of CLOCK and BMAL1, forming a feedback loop that maintains circadian clock [49]. PER1 itself is a transcriptional regulator that controls the transcription of downstream genes [50]. This study reveals that PER1 can regulate the expression of MMP13 by controlling the expression of P65 and p-P65. The mechanism involved may be PER1 regulating the transcription of a downstream molecule, which can affect the activation of the NF-κB pathway. Another possibility is that PER1 interacts with a protein in the NF-κB pathway, thereby influencing its activation. The underlying mechanisms need further in-depth research, which is also a limitation of this study.
Investigating the mechanisms by which clock genes affect TMJOA can facilitate the future development of related therapeutic or preventive drugs. Currently, there is no cure for TMJOA, only methods to alleviate symptoms. Studying the role of clock genes in TMJOA can help us develop new approaches for prevention and treatment. At the cellular level, novel drugs could directly target clock molecules in chondrocytes, influencing their molecular clock and thereby affecting chondrocyte metabolism to treat TMJOA [51]. At the systemic level, manipulating molecules that regulate peripheral circadian rhythm can regulate chondrocyte metabolism. For example, the melatonin, which regulates peripheral circadian rhythm, could be used to impact chondrocyte metabolism [52], or optimizing light–dark cycles could alleviate OA pathology caused by disruption of circadian rhythm [53]. By studying the circadian rhythm of cartilage tissue, we could optimize drug administration timing, as the target molecules for OA treatment may also exhibit circadian rhythms. Aligning the peak concentration of a drug with the peak expression of target molecules after administration could maximize the efficacy of the drug [54]. PER1 could serve as a potential target for the prevention and treatment of TMJOA or OA.
Conclusion
Core clock genes and Mmp13 are rhythmic expressed in rat mandibular condylar chondrocytes. PER1 can regulate the expression of MMP13 in mandibular condylar chondrocytes through NF-κB pathway. Clock gene Per1 may be a potential target gene for TMJOA treatment.
Availability of data and materials
The data are available upon reasonable request.
References
Lim M-J, Lee J-Y. Computed tomographic study of the patterns of oesteoarthritic change which occur on the mandibular condyle. J Craniomaxillofac Surg. 2014;42:1897–902.
Tanaka E, Detamore MS, Mercuri LG. Degenerative disorders of the temporomandibular joint: etiology, diagnosis, and treatment. J Dent Res. 2008;87:296–307.
Allori AC, Chang CC, Fariña R, Grayson BH, Warren SM, McCarthy JG. Current concepts in pediatric temporomandibular joint disorders: part 1 Etiology, epidemiology, and classification. Plast Reconstr Surg. 2010;126:1263–75.
Derwich M, Mitus-Kenig M, Pawlowska E. Interdisciplinary approach to the temporomandibular joint osteoarthritis-review of the literature. Medicina (Kaunas). 2020;56:225.
Cevidanes LHS, Walker D, Schilling J, Sugai J, Giannobile W, Paniagua B, et al. 3D osteoarthritic changes in TMJ condylar morphology correlates with specific systemic and local biomarkers of disease. Osteoarthritis Cartilage. 2014;22:1657–67.
Cardoneanu A, Macovei LA, Burlui AM, Mihai IR, Bratoiu I, Rezus II, et al. Temporomandibular joint osteoarthritis: pathogenic mechanisms involving the cartilage and subchondral bone, and potential therapeutic strategies for joint regeneration. Int J Mol Sci. 2022;24:171.
Li B, Guan G, Mei L, Jiao K, Li H. Pathological mechanism of chondrocytes and the surrounding environment during osteoarthritis of temporomandibular joint. J Cell Mol Med. 2021;25:4902–11.
Utreja A, Dyment NA, Yadav S, Villa MM, Li Y, Jiang X, et al. Cell and matrix response of temporomandibular cartilage to mechanical loading. Osteoarthritis Cartilage. 2016;24:335–44.
Kalladka M, Quek S, Heir G, Eliav E, Mupparapu M, Viswanath A. Temporomandibular joint osteoarthritis: diagnosis and long-term conservative management: a topic review. J Indian Prosthodont Soc. 2014;14:6–15.
Wang XD, Zhang JN, Gan YH, Zhou YH. Current understanding of pathogenesis and treatment of TMJ osteoarthritis. J Dent Res. 2015;94:666–73.
Gamble KL, Berry R, Frank SJ, Young ME. Circadian clock control of endocrine factors. Nat Rev Endocrinol. 2014;10:466–75.
Serin Y, Acar TN. Effect of circadian rhythm on metabolic processes and the regulation of energy balance. Ann Nutr Metab. 2019;74:322–30.
Patke A, Young MW, Axelrod S. Molecular mechanisms and physiological importance of circadian rhythms. Nat Rev Mol Cell Biol. 2020;21:67–84.
Rath MF, Rohde K, Fahrenkrug J, Møller M. Circadian clock components in the rat neocortex: daily dynamics, localization and regulation. Brain Struct Funct. 2013;218:551–62.
Yamajuku D, Shibata Y, Kitazawa M, Katakura T, Urata H, Kojima T, et al. Cellular DBP and E4BP4 proteins are critical for determining the period length of the circadian oscillator. FEBS Lett. 2011;585:2217–22.
Russell JE, Grazman B, Simmons DJ. Mineralization in rat metaphyseal bone exhibits a circadian stage dependency. Proc Soc Exp Biol Med. 1984;176:342–5.
Stevenson S, Hunziker EB, Herrmann W, Schenk RK. Is longitudinal bone growth influenced by diurnal variation in the mitotic activity of chondrocytes of the growth plate? J Orthop Res. 1990;8:132–5.
Kong SY, Stabler TV, Criscione LG, Elliott AL, Jordan JM, Kraus VB. Diurnal variation of serum and urine biomarkers in patients with radiographic knee osteoarthritis. Arthritis Rheum. 2006;54:2496–504.
Dudek M, Gossan N, Yang N, Im H-J, Ruckshanthi JPD, Yoshitane H, et al. The chondrocyte clock gene Bmal1 controls cartilage homeostasis and integrity. J Clin Invest. 2016;126:365–76.
Akagi R, Akatsu Y, Fisch KM, Alvarez-Garcia O, Teramura T, Muramatsu Y, et al. Dysregulated circadian rhythm pathway in human osteoarthritis: NR1D1 and BMAL1 suppression alters TGF-β signaling in chondrocytes. Osteoarthritis Cartilage. 2017;25:943–51.
Gossan N, Zeef L, Hensman J, Hughes A, Bateman JF, Rowley L, et al. The circadian clock in murine chondrocytes regulates genes controlling key aspects of cartilage homeostasis. Arthritis Rheum. 2013;65:2334–45.
Bang J, Chang HW, Jung H-R, Cho C-H, Hur J-A, Lee S-I, et al. Melatonin attenuates clock gene cryptochrome1, which may aggravate mouse anti-type II collagen antibody-induced arthritis. Rheumatol Int. 2012;32:379–85.
Balsalobre A, Brown SA, Marcacci L, Tronche F, Kellendonk C, Reichardt HM, et al. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science. 2000;289:2344–7.
Ying W, Yuan F, He P, Ji P. Inhibition of Notch1 protects against IL-1β-induced inflammation and cartilage destruction in temporomandibular chondrocytes. Mol Med Rep. 2017;15:4391–7.
Li Y, Sun H, Liu X, Hu Z, Jiang H, Guo H, et al. Transglutaminase 2 inhibitors attenuate osteoarthritic degeneration of TMJ-osteoarthritis by suppressing NF-κB activation. Int Immunopharmacol. 2023;114: 109486.
Wang YL, Zhang J, Zhang M, Lu L, Wang X, Guo M, et al. Cartilage degradation in temporomandibular joint induced by unilateral anterior crossbite prosthesis. Oral Dis. 2014;20:301–6.
Ootake T, Ishii T, Sueishi K, Watanabe A, Ishizuka Y, Amano K, et al. Effects of mechanical stress and deficiency of dihydrotestosterone or 17β-estradiol on Temporomandibular Joint Osteoarthritis in mice. Osteoarthritis Cartilage. 2021;29:1575–89.
Xu L, Polur I, Lim C, Servais JM, Dobeck J, Li Y, et al. Early-onset osteoarthritis of mouse temporomandibular joint induced by partial discectomy. Osteoarthritis Cartilage. 2009;17:917–22.
McDearmon EL, Patel KN, Ko CH, Walisser JA, Schook AC, Chong JL, et al. Dissecting the functions of the mammalian clock protein BMAL1 by tissue-specific rescue in mice. Science. 2006;314:1304–8.
Samsa WE, Vasanji A, Midura RJ, Kondratov RV. Deficiency of circadian clock protein BMAL1 in mice results in a low bone mass phenotype. Bone. 2016;84:194–203.
Wang J, Ma J, Gu J-H, Wang F-Y, Shang X-S, Tao H-R, et al. Regulation of type II collagen, matrix metalloproteinase-13 and cell proliferation by interleukin-1β is mediated by curcumin via inhibition of NF-κB signaling in rat chondrocytes. Mol Med Rep. 2017;16:1837–45.
Hirata M, Kugimiya F, Fukai A, Saito T, Yano F, Ikeda T, et al. C/EBPβ and RUNX2 cooperate to degrade cartilage with MMP-13 as the target and HIF-2α as the inducer in chondrocytes. Hum Mol Genet. 2012;21:1111–23.
Tchetina EV, Kobayashi M, Yasuda T, Meijers T, Pidoux I, Poole AR. Chondrocyte hypertrophy can be induced by a cryptic sequence of type II collagen and is accompanied by the induction of MMP-13 and collagenase activity: implications for development and arthritis. Matrix Biol. 2007;26:247–58.
Fei J, Liang B, Jiang C, Ni H, Wang L. Luteolin inhibits IL-1β-induced inflammation in rat chondrocytes and attenuates osteoarthritis progression in a rat model. Biomed Pharmacother. 2019;109:1586–92.
Wang H, Shi Y, He F, Ye T, Yu S, Miao H, et al. GDF11 inhibits abnormal adipogenesis of condylar chondrocytes in temporomandibular joint osteoarthritis. Bone Joint Res. 2022;11:453–64.
Ye T, He F, Lu L, Miao H, Sun D, Zhang M, et al. The effect of oestrogen on mandibular condylar cartilage via hypoxia-inducible factor-2α during osteoarthritis development. Bone. 2020;130: 115123.
Snelling SJB, Forster A, Mukherjee S, Price AJ, Poulsen RC. The chondrocyte-intrinsic circadian clock is disrupted in human osteoarthritis. Chronobiol Int. 2016;33:574–9.
Le NQ, Binh NT, Takarada T, Takarada-Iemata M, Hinoi E, Yoneda Y. Negative correlation between Per1 and Sox6 expression during chondrogenic differentiation in pre-chondrocytic ATDC5 cells. J Pharmacol Sci. 2013;122:318–25.
Qian Z, Gao X, Jin X, Kang X, Wu S. Cartilage-specific deficiency of clock gene Bmal1 accelerated articular cartilage degeneration in osteoarthritis by up-regulation of mTORC1 signaling. Int Immunopharmacol. 2023;115: 109692.
Bekki H, Duffy T, Okubo N, Olmer M, Alvarez-Garcia O, Lamia K, et al. Suppression of circadian clock protein cryptochrome 2 promotes osteoarthritis. Osteoarthritis Cartilage. 2020;28:966–76.
Chen G, Zhao H, Ma S, Chen L, Wu G, Zhu Y, et al. Circadian rhythm protein bmal1 modulates cartilage gene expression in temporomandibular joint osteoarthritis via the MAPK/ERK pathway. Front Pharmacol. 2020;11: 527744.
He D, An Y, Li Y, Wang J, Wu G, Chen L, et al. RNA sequencing reveals target genes of temporomandibular joint osteoarthritis in rats after the treatment of low-intensity pulsed ultrasound. Gene. 2018;672:126–36.
Cha S, Lee S-M, Wang J, Zhao Q, Bai D. Enhanced circadian clock in MSCs-based cytotherapy ameliorates age-related temporomandibular joint condyle degeneration. Int J Mol Sci. 2021;22:10632.
Oeckinghaus A, Ghosh S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb Perspect Biol. 2009;1: a000034.
Ge X-P, Gan Y-H, Zhang C-G, Zhou C-Y, Ma K-T, Meng J-H, et al. Requirement of the NF-κB pathway for induction of Wnt-5A by interleukin-1β in condylar chondrocytes of the temporomandibular joint: functional crosstalk between the Wnt-5A and NF-κB signaling pathways. Osteoarthritis Cartilage. 2011;19:111–7.
Liao S, Zhou K, Li D, Xie X, Jun F, Wang J. Schisantherin A suppresses interleukin-1β-induced inflammation in human chondrocytes via inhibition of NF-κB and MAPKs activation. Eur J Pharmacol. 2016;780:65–70.
Li Y, Jiang W, Wang H, Deng Z, Zeng C, Tu M, et al. Osteopontin promotes expression of matrix metalloproteinase 13 through NF-κB signaling in osteoarthritis. Biomed Res Int. 2016;2016:6345656.
Lepetsos P, Papavassiliou KA, Papavassiliou AG. Redox and NF-κB signaling in osteoarthritis. Free Radic Biol Med. 2019;132:90–100.
Huang W, Ramsey KM, Marcheva B, Bass J. Circadian rhythms, sleep, and metabolism. J Clin Invest. 2011;121:2133–41.
Koike N, Yoo S-H, Huang H-C, Kumar V, Lee C, Kim T-K, et al. Transcriptional architecture and chromatin landscape of the core circadian clock in mammals. Science. 2012;338:349–54.
He T, Pang S, Wang H, Yun H, Hao X, Jia L, et al. Drugging the circadian clock feedback cycle to ameliorate cartilage degeneration. FEBS J. 2022;289:6643–58.
Lu K-H, Lu PW-A, Lu EW-H, Tang C-H, Su S-C, Lin C-W, et al. The potential remedy of melatonin on osteoarthritis. J Pineal Res. 2021;71:12762.
Kc R, Li X, Voigt RM, Ellman MB, Summa KC, Vitaterna MH, et al. Environmental disruption of circadian rhythm predisposes mice to osteoarthritis-like changes in knee joint. J Cell Physiol. 2015;230:2174–83.
Cutolo M. Circadian rhythms and rheumatoid arthritis. Joint Bone Spine. 2019;86:327–33.
Acknowledgements
Not applicable.
Funding
This work is supported by National Natural Science Foundation of China (No. 82271021), the Guangdong Basic and Applied Basic Research Foundation, China (No. 2021A1515010460), Fundamental Research Funds for the Central Universities, Sun Yat-sen University (23qnpy143) and the Science and Technology Projects in Guangzhou, China (No. 202102080157).
Author information
Authors and Affiliations
Contributions
JMW, HA and ZC designed the research. JMW and SQT performed the research. YXW and SZ analyzed data. YF contributed to the data interpretation. JMW wrote the paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All operations are carried out in accordance with animal ethics (approved by Ethics Committee of Sun Yat-sen University, China).
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Primer sequence of core clock genes and Mmp13.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wei, Jm., Tu, Sq., Wang, Yx. et al. Clock gene Per1 regulates rat temporomandibular osteoarthritis through NF-κB pathway: an in vitro and in vivo study. J Orthop Surg Res 18, 817 (2023). https://doi.org/10.1186/s13018-023-04301-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13018-023-04301-7