
Abstract
Neuromodulation with electromagnetic stimulation is widely used for the control of abnormal neural activity, and has been proven to be a valuable alternative to pharmacological tools for the treatment of many neurological diseases. Tremendous efforts have been focused on the design of the stimulation apparatus (i.e., electrodes and magnetic coils) that delivers the electric current to the neural tissue, and the optimization of the stimulation parameters. Less attention has been given to the complicated, dynamic properties of the neurons, and their context-dependent impact on the stimulation effects. This review focuses on the neuronal factors that influence the outcomes of electromagnetic stimulation in neuromodulation. Evidence from multiple levels (tissue, cellular, and single ion channel) are reviewed. Properties of the neural elements and their dynamic changes play a significant role in the outcome of electromagnetic stimulation. This angle of understanding yields a comprehensive perspective of neural activity during electrical neuromodulation, and provides insights in the design and development of novel stimulation technology.
Similar content being viewed by others


Introduction
As electrically excitable cells, neurons transfer information among each other using synapses or electric connections. Fluctuations of the transmembrane potential and dynamic changes in the voltage-dependent ion channels produce action potentials and induce synaptic connections with other neurons. These unique features of the neurons, therefore, allow the exogenous electrical signals to control or modify their excitability, behavior, and functions. Numerous electrical stimulation devices have been invented to deliver electric currents that target the brain and spinal cord for neuromodulation. Among these many clinical successes, deep brain stimulation (DBS) uses implanted electrode leads. DBS is an established symptomatic surgical therapy for Parkinson’s disease, essential tremor, and several other movement and neuropsychiatric disorders [1]. Transcranial magnetic stimulation (TMS), an electromagnetic technology, induces electric currents to control neural activities inside the brain [2]. Transcranial direct-current stimulation (tDCS) uses weak, direct currents to shift the resting potential of cortical neurons in a non-invasive manner. Spinal cord stimulation (SCS) uses minimally invasive stimulation for the treatment of chronic neuropathic pain [3]. From the start of these clinical practices with electrical neuromodulation, researchers began to ask the questions, “What determines the outcome of electrical stimulation? How can we improve it?”.
From the early years of its practice, researchers have realized that optimal control of neural activity with the electric field seemed largely dependent on the properties of the electric currents that were applied to the target neuronal tissue. Many key parameters that define the electric current have been exclusively studied, such as intensity, duration, frequency, and orientation [4,5,6]. For example, neurons in the motor cortex displayed different sensitivities to transcranial magnetic fields with differing coil orientations, shapes of the induced current pulse, and intensities [7,8,9,10,11,12]. Development of various technologies has allowed researchers to link various stimulation parameters to fundamental cellular physiology, such as the excitation of individual neurons [13], changes in ion channel dynamics [14], and alternation of synaptic transmission [15, 16], etc. Cellular analysis reveals that the orientation of the magnetic field determines the geometric pattern of membrane polarization on the neuron [13, 17]. To achieve optimal outcomes, clinical research witnesses the continuous optimization of the stimulating parameters [18,19,20]. In TMS, with careful coil design and placement around the head, the magnetic field can eventually achieve improved clinical outcomes [10, 11].
Even though substantial variance has been observed in patients and normal subjects, at the individual neuron level, the impacts of the targeted tissue and neurons on the outcome of the electrical stimulation has drawn far less attention. For example, repetitive transcranial magnetic stimulation (rTMS) over the occipital cortex led to a significantly increased visual cortex excitability in subjects affected by migraine with aura, but a decreased visual cortex excitability in normal subjects [21]. At the cellular level, the same electrical stimulation protocol can cause significant difference in neural activity. Identical stimulation parameters can result in neuronal activation, suppression, or both, depending on the brain region [22]. Considering the complicated interactions between the applied electric field and the biological tissue (reviewed in [23]), it is essential to understand the outcome of electrical stimulation and its dependence on both the properties of the stimuli and its neural targets.
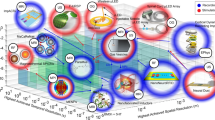
This paper will review the impact of biological complexity on the outcomes of neuromodulation with electromagnetic stimulation. We organize our evidence in a multidimensional format—at the tissue level, cell level, and molecular/ion channel level, respectively (Fig. 1). It is our hope that reviewing these biological complexities during electrical stimulation will provoke awareness in the scientific community during the investigation of the underlying mechanisms of stimulation, and will facilitate the design and development of novel protocols and technologies for neural control with electrical stimulation.

Conceptual figure that summarizes multiple different electromagnetic stimulation modalities across different biological levels. The text of the paper is organized to provide evidence for how the different factors at each of these levels could impact the outcome of the stimulation. Abbreviations are provided separately in the “abbreviations list” of the text
Review
Dynamic changes in tissue properties impact neural stimulation with electromagnetic fields
Biophysical complexity of the neural tissue that surrounds the stimulating electrode alters the electric current distribution and the stimulation outcome. This includes, but is not limited to, inhomogeneity and anisotropy of the tissue, compactness of the cell, and impedance change due to electrode-tissue interaction.
Impact of tissue inhomogeneity and anisotropy on electrical stimulation
Under electrical stimulation, distribution of the electric current inside the neural tissue is largely dependent on the electrical properties of the tissue, including homogeneity and isotropy. A homogeneous material has the same properties at every point. An isotropic material has the same physical properties in all directions. Most neural tissues are inhomogeneous, compositing different types of cells, tissues, fat, irregular vascularization, pH variation, and heterogeneous oxygen concentration, etc. Inhomogeneous resistance is present in the extracellular space in many brain regions, such as the hippocampus and the cerebellum [24]. For example, the tissue conductivity can vary in different layers of the hippocampus [25], with the resistance in the stratum pyramidale two times higher than other hippocampal layers. Additionally, tissue swelling has been observed after intense neural activity, which could lead to an increase in extracellular resistance [26]. Similarly, directional variance in electrophysical properties (anisotropy) is frequently observed in biological tissue. This material property allows tissues to change or assume different properties in different directions. For example, the specific direction of a fiber track generates anisotropy in the brain [27].
Under electrical stimulation, the electric field could be significantly dispersed by non-homogenous tissue conductivity and anisotropy. A recent numerical study [28] suggested that a decrease in tissue conductivity resulted in a smaller volume of neuronal tissue being activated by an electrode. Anisotropic properties of the head play significant roles in the distribution of the electric fields and the effects of transcranial direct-current stimulation (tDCS) [29, 30], since the electric current tends to flow in directions more parallel to the white matter fiber tracts.
In magnetic stimulation, electric currents inside the tissue are generated by electromagnetic induction, which can also be significantly dispersed by the inhomogeneity and anisotropy of the tissue. During TMS, the electric field and current density distribution induced in the brain were significantly altered by the tissue [27, 31, 32]. Specifically, it was found that tissue types and fiber orientation affect the induced electric field [32] and the outcome of the treatment [27].
Impact of cells and cell density on electrical stimulation
The cell membrane is largely non-conductive. Consequently, the presence of the cells will inevitability perturb the externally-applied electric field.
At the single cell level, a spherical cell disturbs the electric potential generated by a microelectrode [33], or a magnetic field [13]. This altered distribution of the microscopic electric field around a single cell could cause secondary effects to the neighboring cells. For example, it will change the membrane potentials in the neighboring cell without direct physical contact between the two cells [34]. Normally, a single cell is surrounded by many other cells. Therefore, although it is possible to predict the polarization and excitability of a single neuron under a specific stimulation electrode arrangement [33], it is nearly impossible to describe the polarization patterns of a single neuron embedded in a large cluster of cells. Indeed, polarization in individual cells depends on the overall density and spatial arrangement of the cells in the three-dimensional cell cluster [35].
Impact of impedance change on electrical stimulation at the electrode-tissue interface
When electric currents are delivered by the implanted electrode for neural stimulation, the impedance between the electrode and the contacting tissue determines the electric current flow inside the tissue [36]. However, impedance changes during chronic implantation. The devastated path left by an intracortical-inserted electrode can lead to vascular damage and tearing of the extracellular matrix, damaging both neurons and glial cells [37, 38]. Consequently, increased electrode-tissue impedance is observed in the weeks following electrode implantation in the brain, stabilizing at approximately 3 to 6 months. Postmortem studies from patients confirmed the presence of a fibrous sheath, astrocytosis, neuronal loss, and neuroinflammation in the immediate vicinity of the electrode [39]. Effects of glial scar formation around the electrode also prevent the readout of nerve activity [40]. In a study that investigated the relationship between impedance change and neural recording in a set of chronically implanted animals, the authors found a positive correlation between glial cells and tissue impedance, and a negative correlation between glial cell density and neural signal quality [41]. Establishing how electrical stimulation influences the electrical and histological properties of the surrounding tissue is essential for the development of future neuromodulation systems for more efficient stimulation.
Cell properties impact neural stimulation with electromagnetic fields
Different cells respond differently to the same electrical stimulation, due to the variabilities in the morphology, biophysical properties, and physiology among different neuronal populations.
Variability among neuron types
The nervous system is composed of various types of glia cells and neurons that have separate identities compared to other cells in the system. The outcome of electrical stimulation is dependent on these different types of neurons. For example, the excitability of somatostatin-positive interneurons is higher than that of regular spiking pyramidal neurons in response to various activating stimuli, including extracellular current, low-Mg2+/Ca2+ artificial cerebrospinal fluid, metabotropic glutamate receptor agonists, and cholinergic agonists [42]. In TMS practice, it was found that there exists select groups of neurons with higher sensitivity to the magnetic field [43]. Indeed, the same magnetic stimulation caused different levels of excitation in layer 5 (L5) pyramidal neurons and interneurons in cortical slices of rats [44].
Cell orientation to the electric field
Orientation of the cell to the electric current determines the pattern and intensity of the field-induced membrane potential at the single cell level. This concept was established mostly due to the biophysics modeling of the neuron as a simple geometric shape, such as a spherical cell or spheroidal cell [33, 45, 46]. Numerical modeling with a multi-compartmental method revealed a detailed activation pattern of the targeted neurons with variable orientation. For example, it was reported that clinical DBS electrodes have different effects in axon activation, depending on the axonal orientation. The activation threshold of the axons that were parallel to the electrode was the lowest. As the axons increased their angle toward the perpendicular placement, the activation threshold increased [47].
The biophysics of such orientation-dependent axonal activation has been extensively investigated. Location of activation is largely determined by the gradients of the electric field along the neural tissue, or the activating function [48,49,50]. Orientations that favor the establishment of large field gradients are essential for electrical activation of the neuron. This theoretical understanding of the impact of orientation on neural activation led to the delicate control of retina ganglion cells by the electric field [51].
Under magnetic stimulation, orientation of the magnetic coil is also a major concern in effective TMS of the motor cortex [7, 11, 12]. At the cellular level, a greater magnetic stimulation is needed when a cell is placed perpendicularly to the magnetically-induced electric field [52, 53]. In an in vitro study, when axons from multiple neurons were forced to form a ring in the culture, the neurons could be magnetically stimulated. In contrast, two-dimensional cultures of comparable size did not respond to magnetic excitation [43]. This happens because the one-dimensional pattern allowed axons in the ring structure to follow the magnetically induced electric field to generate a large field gradient [43].
Neural morphology and electrical properties
A typical neuron consists of three major components: the soma, axon, and dendrite. Under electrical stimulation, the basic morphologies of the neuronal elements, including the size and shape of the neural structure, all play a significant role in somatic depolarization by altering the current sources or sinks created by the applied field [54, 55]. Consequently, the geometric and electrical properties of a single neuron affect electrical stimulation.
Computational work revealed that larger cell bodies are associated with greater transmembrane potentials under electrical [56] and magnetic [13] stimulation. The stimulation effects within different brain regions were found to be dependent on the axon fiber size in DBS [57]. A larger axon diameter is associated with a smaller axonal resistance, leading to a lower threshold for excitation.
Similarly, electroporation studies have demonstrated that the threshold for membrane permeabilization is associated with the size of the target cells [58, 59]. The larger cell requires lower external fields to create permeable cell membranes [60]. Computational work also confirms the importance of cell shape to electrical stimulation. Distribution of the induced transmembrane potential under a homogeneous electric field is affected by the shape of the cell. This distribution is more complicated in a spheroidal cell than in a spherical cell [45, 61].
In a typical neuron, the soma leads to the axon through the axon hillock. Placement of electrical stimulation to the neuron, specifically at the initial axonal segment, has resulted in a lower threshold in mouse and rat cortical brain slices. This is due to the great number of voltage gated sodium channels in this area [44, 62]. Axons are further divided regarding the presence or absence of myelin. Myelin sheaths act as insulators, where the thickness, length, and amount of axon coverage greatly affects the stimulation needed. Factors such as the degree of axonal myelination and the presence of large bending axons are known to have an important influence on electrical stimulation [2]. For example, dorsal root ganglion stimulation (DRGS) with electric current was developed in the mid-2010s to treat chronic pain [63]. The stimulation drives the activity of large, myelinated Aβ-fibers, but not small, unmyelinated C-fibers [64]. The last prominent property of the neuron is the dendrite. Dendritic complexity influences electrophysiological characteristics through unusual firing frequency and patterns [65]. The threshold for electrical activation is lower in neurons with symmetrical dendrites than in those with asymmetrical dendrites, due to less axial resistance in the symmetrical dendrites [54].
In addition to the geometric properties of the cell, electrical properties of the cellular environment affect outcomes of electrical stimulation. These biophysical properties include the conductance of the medium, the membrane, and the cytoplasm [66]. Transmembrane potential is dependent on the electric conductance of the media and cytoplasm, and the membrane conductivity under electrical [46] and magnetic stimulation [13, 53, 67]. Medium conductivity also plays a significant role in causing membrane permeability under electroporation [45, 68, 69].
Physiological properties of the neurons
Physiological properties of the neurons play compounded roles in electrical stimulation. These include, but are not limited to, the threshold of activation, refractory period, and capability of rebounding after hyperpolarization.
The threshold of neural activation is defined as the minimal current needed to excite a neuron [70]. Neurons are more excitable when their membrane potential is just below threshold but not discharging [2]. Similarly, neurons with a low current threshold are more susceptible to magnetic stimulation [44, 52]. The refractory period of the neuron puts constraints on the frequency of electrical stimuli for neural activation. There is a limit to the highest frequency of stimulation at which neurons can still respond to individual stimulation pulses.
Post inhibitory rebound (PIR) is a phenomenon where a cell will spontaneously fire after it has been intensively inhibited [71, 72]. For example, some inhibitory interneurons in goldfish exhibited a remarkable excitation triggered by a brief hyperpolarization [73]. In clinical practice, this cellular property converts the outcome of electrical stimulation from inhibition to excitation. For example, transcranial alternating current stimulation (tACS) causes a rebound of neural activity, rather than entrenchment [74]. In TMS of the cat primary visual cortex, strong stimuli could lead to an early suppression of activity during the first 100-200 ms, followed by stronger (rebound) facilitation [75]. In DBS, the globus pallidus internus (GPi) and thalamic neuronal activity were initially inhibited, but immediately increased after the stimulation [76]. In DBS on human thalamic neurons, a transient rebound of bursting activity was found, followed by prolonged inhibition after the cessation of the high frequency extracellular stimulation [77]. PIR observed in DBS raised questions about the role of inhibition in the current concepts of basal ganglia physiology [76, 77].
Dynamic changes in the neurons impact the outcome of electrical stimulation
Neurons are dynamic entities. They grow, mature, and age. Neurons also die in pathological conditions. Neuronal properties could also be altered by electrical stimulation. Activation state of the neuron consistently changes over time. All these dynamic changes will impact the outcome of electrical stimulation.
Changes in the biophysical properties and functions of the neurons during development
Neuronal growth can lead to profound effects on the biophysical properties, excitation levels, and function changes of the cells. For example, the electrotonic structure and synaptic integration of the lateral giant neurons in crayfish grow in size during development, leading to decreased input resistance and increased cell membrane constants [78]. These structural and biophysical changes can impact the electrophysiological profile of the brain tissue. For example, certain electric potentials can be generated from the fusiform gyrus in adult brains, but not in infants [79,80,81]. Neuron excitation also changes during aging. Intracranial electroencephalography (EEG) and spontaneous field potential recordings demonstrate that the overall network activity in the hippocampus increases during aging [82].
Neurons change their functional roles during development. For example, by releasing the neurotransmitter gamma-aminobutyric acid (GABA), GABAergic interneurons are traditionally regarded as inhibitory to network activity. However, GABAergic interneurons can excite and inhibit postsynaptic neurons, depending on the GABA reversal potential in the postsynaptic cells [83, 84]. During early mammalian embryonic development, the level of sodium–potassium-chloride cotransporter 1 (NKCC1) is high, and the level of potassium chloride cotransporter 2 (KCC2) expression is low [85]. This causes a high concentration of intracellular Cl− and depolarization of the reversal potential for GABA (EGABA). As a result, GABA transmission to these postsynaptic neurons is depolarizing but not hyperpolarizing [86,87,88,89,90,91,92].
Dynamic changes of the neurons during pathological conditions
Pathological conditions lead to profound changes at the cellular level. In animal models of epilepsy, abnormalities were found in the inhibitory GABAergic neurons [93, 94]. Furthermore, in human temporal lobe epilepsy, the loss of hippocampal interneurons was observed (TLE) [95]. Many neurological diseases cause systemic changes in the axons, such as reduction in internodal lengths [96], segmental demyelination [97], and changes in innervation [98].
Pathophysiological changes in the neurons could alter their excitation states. For example, hemisection of the spinal cord in rats led to changes in synaptic transmission. It caused a reduction in the amplitude and frequency of spontaneous inhibitory postsynaptic currents (sIPSC) in substantia gelatinosa neurons. This led to a change in the overall hyperexcitation state in these neurons [99]. In another example, dendritic sprouting occurred during chronic levodopa (L-DOPA) treatments in parkinsonian rats, leading to a reduction of intrinsic excitability [100].
Ionic gradients can also be altered in pathological conditions. For example, potassium ions are responsible for neuronal excitation and network excitability. During seizure, the ion concentration fluctuated in the neuronal tissue [101].
Neuron’s properties change during electrical stimulation
Electric pulses applied to the neuron can change its physiological properties. It is well known that high frequency stimulation protocols lead to long-term potentiation in the synaptic strength [102]. When stimulated with high frequency pulses, axons in the spinal cord demonstrate some “fatigue” responses, manifested as a reduced size of compound action potentials (CAPs) [103]. High frequency electrical stimulation can also change the refractory period of a neuron. Typically, the refractory period lasts about 2 ms, but with high frequency stimulation, the refractory period is extended to 5–10 ms in rat brains. This can be explained by changes in ionic concentrations and channel dynamics [104].
The microscopic ionic environment surrounding a cell changes during electrical stimulation. For example, an increase in potassium concentration was observed in hemiparkinsonian rats that were undergoing DBS [105]. Similarly, excessive potassium ion release happens in the brain slices during electrical stimulation, which leads to the depolarization block of epileptic neurons [106, 107]. Calcium ion concentrations also change during electrical stimulation. Monai et al. [108] demonstrated that tDCS caused synchronous, large-amplitude Ca2+ surges across the cortex of transgenic mice.
Changes in the activation state of the neurons impacts electrical stimulation
The brain has dynamic internal states [109, 110], which may play a significant role in the outcome of electrical and magnetic stimulation (termed “state-dependent”). For example, electrical stimulation of the ventral tegmental area in rhesus monkeys produced different responses, depending on if the animal was awake or under anesthesia [111]. Similarly, TMS produces different perceptual or behavioral outcomes that may depend on the excitability levels of specific neuronal populations [112]. Recording of extracellular spikes and local field potentials from the cat visual cortex has demonstrated that the outcome of stimulation depends on the pre-TMS state of network activity [113]. The neurobiological mechanisms underlying state-dependent electrical stimulation are largely unknown. Some works have highlighted the importance of neural connectivity underlying state-dependent neural stimulation. Considerable changes have been observed in functional connectivity and correlated activity between the awake state and anesthesia both in monkeys [114] and rodents [115], and animals under these different states have distinct responses to neural stimulation [111].
These observations suggest that stimulation effects could be dependent on the active state of individual neurons. Li et al. found that the neuronal response to electromagnetic stimulation highly depends on the intrinsic cellular dynamics in each state [116]. Yang et al. found that neurons at three different levels of excitation (spiking, bursting, and bistable) demonstrate different dynamic responses under magnetic stimulation. Magnetic stimulation has a minimized impact on the temporal firing sequences of bursting neurons in comparison to the spiking and bi-stable neurons [117]. Clinically, the state of cortical neurons can affect the neuronal response to TMS [118, 119]. Using neurons from the model system Aplysia californica, we have found state-dependent neural inhibition by the magnetic field, with neurons firing at a low frequency being more susceptible to inhibition than those firing at a higher frequency [120]. However, as of today, there is a lack of systematic understanding in how stimulation interacts with endogenous neural activity.
Ephaptic effects between neurons affect electrical stimulation
Neurons use field potentials to communicate, namely by ephaptic effects. By altering the field potential, the exogenously applied electric field could modify neuron behavior by interfering with the ephaptic effects. On the other hand, the ephaptic effects between neurons reshape the efficacy of electrical stimulation.
Ephaptic effects refer to the electrical interactions mediated across the extracellular space. Studies of the goldfish Mauthner cell provided the first clear evidence for such interactions [121]. A single impulse in the M-cell leads to the nearly simultaneous firing of 40 to 80 interneurons nearby [73]. The local electric fields generated by the cooperative action of brain cells can influence the timing of neural activity in the brain [122], and facilitate synchronized, even epileptic-like, neuronal bursting [123] and neuron-glia communication [124]. It is proposed that bioelectric and biomagnetic fields of the astroglial network could equalize extracellular local field potentials (LFPs) and associated local magnetic field potentials (LMFPs) in the cortical layers of the brain. This could contribute to the adequate and coherent integration of external and internal signals in the processing of information [125].
Since ephaptic coupling and electrical stimulation share the same biophysical nature, it is appealing to speculate that the externally applied electric field could interact with the ephaptic coupling between neurons. For example, ephaptic interactions between neurons are known to be a key neuronal mechanism underlying seizure. External electrical stimulation via the interruption of the ephaptic effect is used to suppress seizure [126]. In another example, it was found that nonlinear neuronal oscillators could be coupled indirectly via electromagnetic induction with magnetic flux, through which neurons can communicate without physical connections [127].
On the other hand, ephaptic coupling between neurons could alter the outcome of electrical stimulation. In a computational model of a peripheral nerve trunk, the interstitial space between the fibers and the tissues was modeled using a resistor network, enabling distance-dependent ephaptic coupling between myelinated axons, and between fascicles. Under electrical stimulation, this ephaptic coupling can increase the number of fibers that are activated, reducing the artificial currents required for axonal recruitment. Ephaptic coupling also reduces and shifts the range of optimal stimulation amplitudes for maximum inter-fascicular selectivity [128].
Molecular changes that affect electrical stimulation
Ion channels are the fundamental units that support neural excitation. Properties of the ion channels shape the output of electrical stimulation. Ion channels change during development, under pathological conditions, and during electrical stimulation. Therefore, the outcome of electrical stimulation may be interpreted in the context of dynamic channel expression and functions.
Distribution and properties of ion channels impact electrical stimulation
The mechanisms of neuronal activation by electrical stimulation are largely governed by voltage-gated ion channels. An electrical stimulus leads to membrane potential depolarization and ion flow across the membrane via voltage-gated channels.
Location of voltage sensitive ion channels affects the stimulation. Normally, dendrites (which have lower sodium channel density) have higher activation thresholds than axons of the same diameter in electrical stimulation [129]. A portion of retinal ganglion cell axons, about 40 μm from the soma, demonstrates a specific biophysical property of highly dense sodium channels. Electrode locations close to this band of highly dense sodium channels show the lowest thresholds during electrical stimulation [130].
Variability in different ion channel kinetics affects electrical stimulation. The expression and kinetics of several different voltage-dependent potassium (KV) channel subtypes in retina cells play determinant roles in the sensitivity of neurons to electrical stimulation. Consequently, when retina cells are electrically stimulated, the non-spiking amacrine neurons of the retina exhibit a large variety of responses [131].
State of the ion channel affects electrical stimulation. Blockage of voltage-gated potassium (Kv) channels may either increase or decrease cellular excitability, depending on the state of the ion channels. Blocking Kv when the channel is at a closed-state increased the excitability, while a selective block of the open Kv channels decreased the rate of repetitive and consequent Ca influx [132].
Ion channels change during development, in pathological conditions, and during electrical stimulation
Mutation of ion channels causes dysfunction in neurons. For example, the action potential initiation mechanism was impaired in GABAergic interneurons in a mouse model that expresses a mutated human Na (V)1.1 gene, resulting in a hyperexcitable network [133]. When the functions of voltage-dependent sodium channels were impaired in the GABAergic interneurons, it led to a reduced threshold and accelerated propagation in febrile seizures, and reduced threshold in flurothyl-induced seizures [134].
Ion channels change during neural injury. For example, after spinal cord injury (SCI), dispersed distributions of potassium channel subunit Kv1.1 and Kv1.2 along the injured axons were observed, in contrast to the tight, clustered locations of these channels to the juxtaparanodes of non-injured axons [135].
Electrical stimulation can regulate the expression and dynamics of the ion channels involved in neural excitability. For example, retinal ganglion cells (RGCs) can fire spikes at frequencies greater than 200 Hz when driven by light. RGC spiking rates decreased when electrically stimulated at 50–200 Hz. This depression was caused by a stimulus-frequency-dependent decline of RGC voltage-gated sodium current, mediated by the sodium channels [136].
Electrical stimulation can also regulate the expression and dynamics of the ion channels involved in synaptic transmission. Long-term potentiation (LTP) refers to persistent strengthening of neuronal synapses. It is normally induced by a strong tetanic (high frequency) stimulation. Long-term depression (LTD) refers to the persistent decrease in synaptic strength following weak, low frequency stimulation. AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) and NMDA (N-methyl-d-aspartate) receptors are permeable to Na+ and K+ (and Ca2+ in NMDARs). The activity of those channels, and the consequent strength of the synapse, depends greatly on the strength and length of stimulation of both presynaptic and postsynaptic neurons [137]. In LTP, more AMPA receptors are inserted into the postsynaptic membrane to potentiate the synapse; in LTD, AMPA receptors are absorbed and removed from the postsynaptic membrane, which weakens the synapse [137]. These neuronal changes, at the ion channel level, result in enhanced or attenuated synaptic transmission triggered by electrical stimulation.
Lessons learned—neurons have dynamic identities during neuromodulation with electromagnetic stimulations
Converging pieces of evidence have suggested a broader frame of reference in which the interactions between the cell and the electric field define the outcome of electrical stimulation [23]. Specifically, this paper reviews the impact of the dynamic aspects of neurons on the efficacy of stimulation. There are several lessons learned from this study.
Importance of improving the biocompatibility of the implants for consistent electrical stimulation
To eliminate inconsistency in neural recording and stimulation due to foreign body reactions, it is essential to improve the biocompatibility of the implants. Next-generation intracortical microelectrodes are being developed with an increased emphasis on reducing the neuro-inflammatory response. The field is actively progressing from traditional, inorganic materials towards approaches that either minimize the microelectrode footprint, or incorporate compliant materials, bioactive molecules, conducting polymers, or nanomaterials with the implants [40]. Biocompatibility can also be improved with drugs. For example, administration of a cell cycle inhibitor (flavopiridol) at the time of surgery would reduce reactive gliosis surrounding neural prostheses. Purcell et al. reported that flavopiridol reduced the expression of a cell cycle protein (cyclin D1) in microglia surrounding the probes, decreased the impedance at 1 kHz, and increased the neural signal [41].
The recent development of micromagnetic stimulation (µMS) technology significantly improves the biocompatibility of the coil implantation. These coils can be manufactured to match the size of the targeted neurons for focal stimulation. They can also be encapsulated under the cover of biocompatible, soft material to avoid the inflammatory and immune responses that occur due to direct contact with the tissue [138, 139]. Evidence has shown that the miniature coil could be used to control retinal ganglion neuron activity with high amplitude pulses [140]. Recently, we reported that µMS could block axonal conductance in unmyelinated axons [141] and in ganglion cells [142]. This technology could also block hyperactive circuit activity in the hippocampus [143].
Importance of understanding neural anatomy for efficient and selective stimulation
Since neural morphology plays a significant role in the neuron’s response to electrical stimulation, an in depth understanding of neural anatomy at the microscopic level is essential in the design of efficient protocols for selective stimulation. The importance of neural anatomy includes elements of orientational and directional selectivity. Studies have suggested that these two factors are related in natural firing rates under electrical stimulation [129]. For example, the µMS technology has been used to activate vertically oriented pyramidal neurons (PNs) without activating horizontally oriented passing axons [144]. Achieving such improved selectivity depends on the understanding of brain anatomy, and the theory that the field parallel to the axon is more effective in neural activation than the field that is perpendicular to the axon [145].
Importance of monitoring neural activity during electrical stimulation
To ensure electrical stimulation takes effect in the most ideal way, it is essential to use imaging or electrophysiology tools to monitor neural activity changes during the process. This practice is necessary in the execution, evaluation and optimization of electrical stimulation. There are several advantages for such an approach:
-
a.
Ensure precise location for the stimulation electrode The same stimulation electrode could be used for neural recording from the implanted area, for it is possible to use the shape of neural activity to deduce the location of the extracellular electrodes [146]. This is a reliable strategy to guide the precise positioning of the extracellular electrode, especially when it is challenging to directly monitor the electrode with imaging technology [147]. Rapid predictability based on the waveform shape of the neural activity provides researchers and clinicians with valuable information right from the implantation of an electrode. For example, characteristics of evoked compound action potentials (ECAPs) and local field potentials (LFPs) recorded during DBS provide valuable information between the contact of the electrode and the tissue [148].
-
b.
Reveal any pathological changes in the targeted area Monitoring neural activity also provides insights to the potential pathological changes in the targeted neural population. Any discrepancies or unexpected shape changes in the neural recording can, therefore, be used to reveal such damage, as seen with strokes [149] and post-concussive syndrome [150]. In another example, acute edema caused a reduction in the single neuron and population ECAP signal energy, as well as LFP magnitude. This information can be used to optimize electrode parameters in a closed-loop DBS system for the treatment of movement disorders [151, 152].
-
c.
Provide predictions of the stimulation outcomes Since pre-stimulation neural activity is highly related to the post-stimulation outcome [113], it is important to monitor the neural activity prior to stimulation. This technique can generate valuable predictions and a comprehensive understanding of the stimulation outcome.
-
d.
Guide the adjustment of the stimulation protocol Changes in neural activity can empirically guide the effective use of TMS in both clinical and experimental settings [153]. For example, the instantaneous brain state can be used to promote efficacious plasticity induction by TMS [154]. Combining TMS with functional magnetic resonance imaging (fMRI) is powerful in revealing if different TMS intensities are needed to induce neural activation [155].
Importance of utilizing the dynamic state of the neurons for efficient neural stimulation
Because the dynamic changes at the cellular level impact the outcome of electrical stimulation, it is essential to apply state-dependent brain stimulation to match and compensate for the state changes.
-
a.
Preconditioning the brain state for efficient stimulation It is essential to develop technology that can precondition the state of the neural network to enhance the stimulation outcome [156]. For example, tDCS was used to precondition the motor cortex for a better outcome in low-frequency rTMS [157, 158]. In another example, transcranial random noise stimulation (tRNS) was used to precondition the primary motor cortex. This led to a lower response threshold and increased responsiveness when probed with TMS [159].
-
b.
Adjusting the brain state with pharmacology for efficient stimulation It is possible to consider using a pharmacological approach to alter the excitability of the nervous system and maximize the clinical outcome of rTMS. As a proof, when anticonvulsant phenytoin was administrated, the magnetic field was more effective in decreasing audiogenic seizure severity in mice [160, 161]. Similarly, bursts of high frequency rTMS, together with lorazepam, suppressed seizures in a rat kainate status epilepticus model [162], with the combined methods more effective than rTMS alone. Future research should explore the possibility of improving complementary therapies by adjusting the excitability state of the nervous system with drugs.
Importance of developing closed-loop stimuli to adapt to the quick changes in neural activity
To apply fast, purpose-driven stimulation to compensate for the dynamic changes in the brain, new technology should explore real-time, closed-loop measurement and stimulation paradigms. Real-time, multi-channel EEG data can be used to monitor the brain state online and modify stimulation parameters [163] to apply state-dependent brain stimulation. EEG can also be used to design closed-loop, purpose-driven stimuli to provide brain-state guided stimulation [164, 165]. For example, Ganzer et al. [166] used closed-loop vagus nerve stimulation (CLVS) to enhance recovery after SCI. To generate the best control, the authors developed a real-time, closed-loop neuromodulation paradigm based on the synaptic eligibility trace, to deliver CLVS immediately after the most successful forelimb movements during motor rehabilitation.
Importance of including neural dynamics in computer simulations of neuromodulation
To appreciate the complexity and reveal the underlying mechanisms of electrical stimulation, researchers use computer simulation to address large sets of parameters and variabilities. The typical stimulation protocol includes a three-step simulation approach [70, 167,168,169]. First, the electric current distribution generated by an electrode/coil is computed in the 3D space. Second, a multi-compartment neural model is built to represent the fine, geometric structure of the neuron or neural networks, with channel mechanisms incorporated into each component. Finally, the electric field obtained from the first step is used to activate the neurons. This classic framework of computation assumes, first, that the extracellular electric field is always treated equally around each model compartment. Second, the extracellular electric field is not affected by the presence of the tissue. Third, the extracellular voltage generated by the membrane current is neglected. Under these assumptions, the extracellular electric field is computed without considering the existence of the tissue, or its counter effect to the externally-applied electric fields. Although such an approach is typical in the field [23, 167], ignorance of the presence of the neuron and its dynamics could potentially cause underestimation of the applied electric fields, and introduce potential inaccuracies in the modeling [33, 170].
Evidence provided in this review paper further suggests that all three aspects of this modeling approach should be improved. Since tissue inhomogeneities play significant roles in the distribution of the externally applied electrical stimulation, a precise understanding of the electric field distribution in the context of the complicated, dynamic tissue properties is essential in studying neural stimulation with the electric field. Computational tools, such as finite element modeling (FEM), which can delineate the tissue complexity and represent the resistive and capacitive properties of the tissue at the micro scale [171], will be appealing. A successful example is the model study on parkinsonian symptoms with electrical stimulation of the subthalamic nucleus (STN). Sotiropoulos and Steinmetz [57] included tissue inhomogeneity and anisotropy in the simulation. The model successfully validated that the tissue’s properties (i.e., degree of inhomogeneity and anisotropy) have a direct impact on stimulation effects, and that increasing STN conductivity could cause neuronal activation outside the STN.
We must consider the neuron as a dynamic entity that consistently changes in anatomy and variable excitation states. Some recent developments of computational technology have begun to address these shortcomings. As the first step, the impact of the presence of the neuron must be better represented in the model. For example, a “whole finite element” approach allows the simultaneous calculation of the extracellular and intracellular potentials by representing the neuronal membrane with a thin-film approximation [167]. This approach illustrates the difference of the neuronal response between the sides of the soma membrane either facing or opposite to the stimulating electrode. By simultaneous calculation of the stimulation potential and the neuron response, this novel approach provides a mechanism to better represent the neuron and its dynamic behavior during electrical stimulation. These improvements result in more accurate results that more closely match the biological reality.
Conclusions
Neurons are dynamic elements in the practice of neuromodulation with electromagnetic stimulation. Ultimately, the outcome of neural stimulation depends on the complicated interactions between the dynamic neurons and the externally applied electric field. Therefore, the neuron matters when we consider the outcomes of electrical stimulation in both laboratory research and in clinical practice. Further research should consider the dynamic properties of neurons in interpreting experimental data and incorporate cell-field interactions when setting up computer simulation.
Availability of data and materials
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Abbreviations
- AMPA:
-
α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid
- CAPs:
-
Compound action potentials
- CLVS:
-
Closed-loop vagus nerve stimulation
- DBS:
-
Deep brain stimulation
- DRGS:
-
Dorsal root ganglion stimulation
- ECAPs:
-
Evoked compound action potentials
- EEG:
-
Electroencephalography
- E GABA :
-
Reversal potential of GABA
- FEM:
-
Finite element modeling
- fMRI:
-
Functional magnetic resonance imaging
- GABA:
-
Gamma-aminobutyric acid
- KCC2:
-
Potassium chloride cotransporter 2
- L-DOPA:
-
Levodopa
- LFPs:
-
Local field potentials
- LMFPs:
-
Local magnetic field potentials
- LTD:
-
Long-term depression
- LTP:
-
Long-term potentiation
- µMS:
-
Micromagnetic stimulation
- NKCC1:
-
Sodium–potassium-chloride cotransporter 1
- NMDA:
-
N-Methyl-d-aspartate
- PIR:
-
Post inhibitory rebound
- PNs:
-
Pyramidal neurons
- RGCs:
-
Retinal ganglion cells
- rTMS:
-
Repetitive transcranial magnetic stimulation
- SCI:
-
Spinal cord injury
- SCS:
-
Spinal cord stimulation
- sIPSC:
-
Spontaneous inhibitory postsynaptic currents
- STN:
-
Subthalamic nucleus
- tACS:
-
Transcranial alternating current stimulation
- tDCS:
-
Transcranial direct-current stimulation
- TLE:
-
Temporal lobe epilepsy
- TMS:
-
Transcranial magnetic stimulation
- tRNS:
-
Transcranial random noise stimulation
References
Lozano AM, Lipsman N, Bergman H, Brown P, Chabardes S, Chang JW, et al. Deep brain stimulation: current challenges and future directions. Nat Rev Neurol. 2019;15(3):148–60.
Siebner HR, Hartwigsen G, Kassuba T, Rothwell JC. How does transcranial magnetic stimulation modify neuronal activity in the brain? Implications for studies of cognition. Cortex. 2009;45(9):1035–42.
Sdrulla AD, Guan Y, Raja SN. Spinal cord stimulation: clinical efficacy and potential mechanisms. Pain Pract. 2018;18(8):1048–67.
Paffi A, Camera F, Apollonio F, d’Inzeo G, Liberti M. Numerical characterization of intraoperative and chronic electrodes in deep brain stimulation. Front Comput Neurosci. 2015;9:2.
Agnesi F, Muralidharan A, Baker KB, Vitek JL, Johnson MD. Fidelity of frequency and phase entrainment of circuit-level spike activity during DBS. J Neurophysiol. 2015;114:825–34.
Stratton J. Electromagnetic theory. New York: McGraw-Hill; 1941.
Di Lazzaro V, Oliviero A, Pilato F, Saturno E, Dileone M, Mazzone P, et al. The physiological basis of transcranial motor cortex stimulation in conscious humans. Clin Neurophysiol. 2004;115(2):255–66.
Mills KR, Boniface SJ, Schubert M. Magnetic brain stimulation with a double coil: the importance of coil orientation. Electroencephalogr Clin Neurophysiol. 1992;85(1):17–21.
Bashir S, Perez JM, Horvath JC, Pascual-Leone A. Differentiation of motor cortical representation of hand muscles by navigated mapping of optimal TMS current directions in healthy subjects. J Clin Neurophysiol. 2013;30(4):390–5.
Laakso I, Hirata A, Ugawa Y. Effects of coil orientation on the electric field induced by TMS over the hand motor area. Phys Med Biol. 2014;59(1):203–18.
Salvador R, Silva S, Basser PJ, Miranda PC. Determining which mechanisms lead to activation in the motor cortex: a modeling study of transcranial magnetic stimulation using realistic stimulus waveforms and sulcal geometry. Clin Neurophysiol. 2011;122(4):748–58.
Janssen AM, Oostendorp TF, Stegeman DF. The coil orientation dependency of the electric field induced by TMS for M1 and other brain areas. J Neuroeng Rehabil. 2015;12:47.
Ye H, Cotic M, Carlen PL. Transmembrane potential induced in a spherical cell model under low-frequency magnetic stimulation. J Neural Eng. 2007;4(3):283–93.
Tan T, Xie J, Tong Z, Liu T, Chen X, Tian X. Repetitive transcranial magnetic stimulation increases excitability of hippocampal CA1 pyramidal neurons. Brain Res. 2013;1520:23–35.
Zheng Y, Ma W, Dong L, Dou JR, Gao Y, Xue J. Influence of the on-line ELF-EMF stimulation on the electrophysiological properties of the rat hippocampal CA1 neurons in vitro. Rev Sci Instrum. 2017;88(10): 105106.
Tokay T, Holl N, Kirschstein T, Zschorlich V, Kohling R. High-frequency magnetic stimulation induces long-term potentiation in rat hippocampal slices. Neurosci Lett. 2009;461(2):150–4.
Durand DM. Electric field effects in hyperexcitable neural tissue: a review. Radiat Prot Dosim. 2003;106(4):325–31.
Pascual-Leone A, Tormos JM, Keenan J, Tarazona F, Canete C, Catala MD. Study and modulation of human cortical excitability with transcranial magnetic stimulation. J Clin Neurophysiol. 1998;15(4):333–43.
Dayan E, Censor N, Buch ER, Sandrini M, Cohen LG. Noninvasive brain stimulation: from physiology to network dynamics and back. Nat Neurosci. 2013;16(7):838–44.
Parkin BL, Ekhtiari H, Walsh VF. Non-invasive human brain stimulation in cognitive neuroscience: a primer. Neuron. 2015;87(5):932–45.
Brighina F, Piazza A, Daniele O, Fierro B. Modulation of visual cortical excitability in migraine with aura: effects of 1 Hz repetitive transcranial magnetic stimulation. Exp Brain Res. 2002;145(2):177–81.
Paus T. Inferring causality in brain images: a perturbation approach. Philos Trans R Soc Lond B Biol Sci. 2005;360(1457):1109–14.
Ye H, Steiger A. Neuron matters: electric activation of neuronal tissue is dependent on the interaction between the neuron and the electric field. J Neuroeng Rehabil. 2015;12:65.
Okada YC, Huang JC, Rice ME, Tranchina D, Nicholson C. Origin of the apparent tissue conductivity in the molecular and granular layers of the in vitro turtle cerebellum and the interpretation of current source-density analysis. J Neurophysiol. 1994;72(2):742–53.
Holsheimer J. Electrical conductivity of the hippocampal CA1 layers and application to current-source-density analysis. Exp Brain Res. 1987;67(2):402–10.
Autere AM, Lamsa K, Kaila K, Taira T. Synaptic activation of GABAA receptors induces neuronal uptake of Ca2+ in adult rat hippocampal slices. J Neurophysiol. 1999;81(2):811–6.
De Lucia M, Parker GJ, Embleton K, Newton JM, Walsh V. Diffusion tensor MRI-based estimation of the influence of brain tissue anisotropy on the effects of transcranial magnetic stimulation. Neuroimage. 2007;36(4):1159–70.
Yousif N, Bayford R, Wang S, Liu X. Quantifying the effects of the electrode-brain interface on the crossing electric currents in deep brain recording and stimulation. Neuroscience. 2008;152(3):683–91.
Rampersad SM, Janssen AM, Lucka F, Aydin U, Lanfer B, Lew S, et al. Simulating transcranial direct current stimulation with a detailed anisotropic human head model. IEEE Trans Neural Syst Rehabil Eng. 2014;22(3):441–52.
Wagner S, Rampersad SM, Aydin U, Vorwerk J, Oostendorp TF, Neuling T, et al. Investigation of tDCS volume conduction effects in a highly realistic head model. J Neural Eng. 2014;11(1): 016002.
Miranda PC, Hallett M, Basser PJ. The electric field induced in the brain by magnetic stimulation: a 3-D finite-element analysis of the effect of tissue heterogeneity and anisotropy. IEEE Trans Biomed Eng. 2003;50(9):1074–85.
Opitz A, Windhoff M, Heidemann RM, Turner R, Thielscher A. How the brain tissue shapes the electric field induced by transcranial magnetic stimulation. Neuroimage. 2011;58(3):849–59.
Lee DC, Grill WM. Polarization of a spherical cell in a nonuniform extracellular electric field. Ann Biomed Eng. 2005;33(5):603–15.
Pucihar G, Kotnik T, Valic B, Miklavcic D. Numerical determination of transmembrane voltage induced on irregularly shaped cells. Ann Biomed Eng. 2006;34(4):642–52.
Pavlin M, Pavselj N, Miklavcic D. Dependence of induced transmembrane potential on cell density, arrangement, and cell position inside a cell system. IEEE Trans Biomed Eng. 2002;49(6):605–12.
Newbold C, Richardson R, Millard R, Seligman P, Cowan R, Shepherd R. Electrical stimulation causes rapid changes in electrode impedance of cell-covered electrodes. J Neural Eng. 2011;8(3): 036029.
Campbell A, Wu C. Chronically implanted intracranial electrodes: tissue reaction and electrical changes. Micromachines. 2018;9(9):430.
Patil AC, Thakor NV. Implantable neurotechnologies: a review of micro- and nanoelectrodes for neural recording. Med Biol Eng Comput. 2016;54(1):23–44.
Evers J, Lowery M. The active electrode in the living brain: the response of the brain parenchyma to chronically implanted deep brain stimulation electrodes. Oper Neurosurg. 2021;20(2):131–40.
Jorfi M, Skousen JL, Weder C, Capadona JR. Progress towards biocompatible intracortical microelectrodes for neural interfacing applications. J Neural Eng. 2015;12(1): 011001.
Purcell EK, Thompson DE, Ludwig KA, Kipke DR. Flavopiridol reduces the impedance of neural prostheses in vivo without affecting recording quality. J Neurosci Methods. 2009;183(2):149–57.
Fanselow EE, Richardson KA, Connors BW. Selective, state-dependent activation of somatostatin-expressing inhibitory interneurons in mouse neocortex. J Neurophysiol. 2008;100(5):2640–52.
Rotem A, Moses E. Magnetic stimulation of one-dimensional neuronal cultures. Biophys J. 2008;94(12):5065–78.
Pashut T, Magidov D, Ben-Porat H, Wolfus S, Friedman A, Perel E, et al. Patch-clamp recordings of rat neurons from acute brain slices of the somatosensory cortex during magnetic stimulation. Front Cell Neurosci. 2014;8:145.
Kotnik T, Bobanovic F, Miklavcic D. Sensitivity of transmembrane voltage induced by applied electric fields—a theoretical analysis. Bioelectrochem Bioenergy. 1997;43(2):285–91.
Kotnik T, Miklavcic D. Analytical description of transmembrane voltage induced by electric fields on spheroidal cells. Biophys J. 2000;79(2):670–9.
Slopsema JP, Pena E, Patriat R, Lehto LJ, Grohn O, Mangia S, et al. Clinical deep brain stimulation strategies for orientation-selective pathway activation. J Neural Eng. 2018;15(5): 056029.
Rattay F. Analysis of models for extracellular fiber stimulation. IEEE Trans Biomed Eng. 1989;36(7):676–82.
Lee SW, Fried SI. Enhanced control of cortical pyramidal neurons with micromagnetic stimulation. IEEE Trans Neural Syst Rehabilit Eng. 2017;25(9):1375–86.
Rattay F. Analysis of models for external stimulation of axons. IEEE Trans Biomed Eng. 1986;33(10):974–7.
Grumet AE, Wyatt JL Jr, Rizzo JF 3rd. Multi-electrode stimulation and recording in the isolated retina. J Neurosci Methods. 2000;101(1):31–42.
Pashut T, Wolfus S, Friedman A, Lavidor M, Bar-Gad I, Yeshurun Y, et al. Mechanisms of magnetic stimulation of central nervous system neurons. PLoS Comput Biol. 2011;7(3): e1002022.
Ye H, Cotic M, Fehlings MG, Carlen PL. Transmembrane potential generated by a magnetically induced transverse electric field in a cylindrical axonal model. Med Biol Eng Comput. 2011;49(1):107–19.
Yi GS, Wang J, Deng B, Wei XL. Morphology controls how hippocampal CA1 pyramidal neuron responds to uniform electric fields: a biophysical modeling study. Sci Rep. 2017;7(1):3210.
Lin C, Huang Y, Quan T, Zhang Y. Modelling brain-wide neuronal morphology via rooted Cayley trees. Sci Rep. 2018;8(1):15666.
Kotnik T, Miklavcic D. Theoretical evaluation of voltage inducement on internal membranes of biological cells exposed to electric fields. Biophys J. 2006;90(2):480–91.
Sotiropoulos SN, Steinmetz PN. Assessing the direct effects of deep brain stimulation using embedded axon models. J Neural Eng. 2007;4(2):107–19.
Rols MP, Delteil C, Serin G, Teissie J. Temperature effects on electrotransfection of mammalian cells. Nucleic Acids Res. 1994;22(3):540.
Calvin NM, Hanawalt PC. High-efficiency transformation of bacterial cells by electroporation. J Bacteriol. 1988;170(6):2796–801.
Gehl J. Electroporation: theory and methods, perspectives for drug delivery, gene therapy and research. Acta Physiol Scand. 2003;177(4):437–47.
Gimsa J, Wachner D. Analytical description of the transmembrane voltage induced on arbitrarily oriented ellipsoidal and cylindrical cells. Biophys J. 2001;81(4):1888–96.
Werginz P, Raghuram V, Fried SI. Tailoring of the axon initial segment shapes the conversion of synaptic inputs into spiking output in OFF-alpha T retinal ganglion cells. Sci Adv. 2020;6(37): eabb6642.
Huygen F, Kallewaard JW, Nijhuis H, Liem L, Vesper J, Fahey ME, et al. Effectiveness and safety of dorsal root ganglion stimulation for the treatment of chronic pain: a pooled analysis. Neuromodulation. 2020;23(2):213–21.
Graham RD, Bruns TM, Duan B, Lempka SF. Dorsal root ganglion stimulation for chronic pain modulates Abeta-fiber activity but not C-fiber activity: a computational modeling study. Clin Neurophysiol. 2019;130(6):941–51.
Zhu G, Du L, Jin L, Offenhausser A. Effects of morphology constraint on electrophysiological properties of cortical neurons. Sci Rep. 2016;6:23086.
Ye H, Cotic M, Fehlings MG, Carlen PL. Influence of cellular properties on the electric field distribution around a single cell. Progr Electromagn Res B. 2012;39:141–61.
Ye H, Cotic M, Kang EE, Fehlings MG, Carlen PL. Transmembrane potential induced on the internal organelle by a time-varying magnetic field: a model study. J Neuroeng Rehabilit. 2010;7:12.
Mossop BJ, Barr RC, Henshaw JW, Zaharoff DA, Yuan F. Electric fields in tumors exposed to external voltage sources: implication for electric field-mediated drug and gene delivery. Ann Biomed Eng. 2006;34(10):1564–72.
Mossop BJ, Barr RC, Zaharoff DA, Yuan F. Electric fields within cells as a function of membrane resistivity—a model study. IEEE Trans Nanobiosci. 2004;3(3):225–31.
Lu H, Chestek CA, Shaw KM, Chiel HJ. Selective extracellular stimulation of individual neurons in ganglia. J Neural Eng. 2008;5(3):287–309.
Ferrante M, Shay CF, Tsuno Y, William Chapman G, Hasselmo ME. Post-inhibitory rebound spikes in rat medial entorhinal layer II/III principal cells: in vivo, in vitro, and computational modeling characterization. Cereb Cortex. 2017;27(3):2111–25.
Angstadt JD, Grassmann JL, Theriault KM, Levasseur SM. Mechanisms of postinhibitory rebound and its modulation by serotonin in excitatory swim motor neurons of the medicinal leech. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2005;191(8):715–32.
Faber DS, Korn H. Field effects trigger post-anodal rebound excitation in vertebrate CNS. Nature. 1983;305(5937):802–4.
Haberbosch L, Schmidt S, Jooss A, Kohn A, Kozarzewski L, Ronnefarth M, et al. Rebound or entrainment? The influence of alternating current stimulation on individual alpha. Front Hum Neurosci. 2019;13:43.
Moliadze V, Zhao Y, Eysel U, Funke K. Effect of transcranial magnetic stimulation on single-unit activity in the cat primary visual cortex. J Physiol. 2003;553(Pt 2):665–79.
Montgomery EB Jr. Effects of GPi stimulation on human thalamic neuronal activity. Clin Neurophysiol. 2006;117(12):2691–702.
Birdno MJ, Tang W, Dostrovsky JO, Hutchison WD, Grill WM. Response of human thalamic neurons to high-frequency stimulation. PLoS ONE. 2014;9(5): e96026.
Edwards DH, Yeh SR, Barnett LD, Nagappan PR. Changes in synaptic integration during the growth of the lateral giant neuron of crayfish. J Neurophysiol. 1994;72(2):899–908.
Deffke I, Sander T, Heidenreich J, Sommer W, Curio G, Trahms L, et al. MEG/EEG sources of the 170-ms response to faces are co-localized in the fusiform gyrus. Neuroimage. 2007;35(4):1495–501.
Gao C, Conte S, Richards JE, Xie W, Hanayik T. The neural sources of N170: understanding timing of activation in face-selective areas. Psychophysiology. 2019;56(6): e13336.
Guy MW, Richards JE, Tonnsen BL, Roberts JE. Neural correlates of face processing in etiologically-distinct 12-month-old infants at high-risk of autism spectrum disorder. Dev Cogn Neurosci. 2018;29:61–71.
El-Hayek YH, Wu C, Ye H, Wang J, Carlen PL, Zhang L. Hippocampal excitability is increased in aged mice. Exp Neurol. 2013;247:710–9.
Staley KJ, Soldo BL, Proctor WR. Ionic mechanisms of neuronal excitation by inhibitory GABAA receptors. Science. 1995;269(5226):977–81.
Bernard C, Cossart R, Hirsch JC, Esclapez M, Ben-Ari Y. What is GABAergic inhibition? How is it modified in epilepsy? Epilepsia. 2000;41(Suppl 6):S90–5.
Stil A, Liabeuf S, Jean-Xavier C, Brocard C, Viemari JC, Vinay L. Developmental up-regulation of the potassium-chloride cotransporter type 2 in the rat lumbar spinal cord. Neuroscience. 2009;164(2):809–21.
Lopantsev V, Avoli M. Participation of GABAA-mediated inhibition in ictallike discharges in the rat entorhinal cortex. J Neurophysiol. 1998;79(1):352–60.
Kohling R, Vreugdenhil M, Bracci E, Jefferys JG. Ictal epileptiform activity is facilitated by hippocampal GABAA receptor-mediated oscillations. J Neurosci. 2000;20(18):6820–9.
Dzhala VI, Staley KJ. Transition from interictal to ictal activity in limbic networks in vitro. J Neurosci. 2003;23(21):7873–80.
Perez Velazquez JL. Bicarbonate-dependent depolarizing potentials in pyramidal cells and interneurons during epileptiform activity. Eur J Neurosci. 2003;18(5):1337–42.
Khalilov I, Le Van QM, Gozlan H, Ben-Ari Y. Epileptogenic actions of GABA and fast oscillations in the developing hippocampus. Neuron. 2005;48(5):787–96.
Fujiwara-Tsukamoto Y, Isomura Y, Imanishi M, Fukai T, Takada M. Distinct types of ionic modulation of GABA actions in pyramidal cells and interneurons during electrical induction of hippocampal seizure-like network activity. Eur J Neurosci. 2007;25(9):2713–25.
Lasztoczi B, Nyitrai G, Heja L, Kardos J. Synchronization of GABAergic inputs to CA3 pyramidal cells precedes seizure-like event onset in juvenile rat hippocampal slices. J Neurophysiol. 2009;102(4):2538–53.
Olsen RW, Avoli M. GABA and epileptogenesis. Epilepsia. 1997;38(4):399–407.
DeLorey TM, Olsen RW. GABA and epileptogenesis: comparing gabrb3 gene-deficient mice with Angelman syndrome in man. Epilepsy Res. 1999;36(2–3):123–32.
de Lanerolle NC, Kim JH, Robbins RJ, Spencer DD. Hippocampal interneuron loss and plasticity in human temporal lobe epilepsy. Brain Res. 1989;495(2):387–95.
Low PA, Dyck PJ. Splanchnic preganglionic neurons in man: II. Morphometry of myelinated fibers of T7 ventral spinal root. Acta Neuropathol. 1977;40(3):219–25.
Appenzeller O, Ogin G. Myelinated fibres in the human paravertebral sympathetic chain; quantitative studies on white rami communicantes. J Neurol Neurosurg Psychiatry. 1973;36(5):777–85.
Phillips RJ, Walter GC, Powley TL. Age-related changes in vagal afferents innervating the gastrointestinal tract. Auton Neurosci. 2010;153(1–2):90–8.
Kozuka Y, Kawamata M, Furue H, Ishida T, Tanaka S, Namiki A, et al. Changes in synaptic transmission of substantia gelatinosa neurons after spinal cord hemisection revealed by analysis using in vivo patch-clamp recording. Mol Pain. 2016;12:1744806916665827.
Fieblinger T, Zanetti L, Sebastianutto I, Breger LS, Quintino L, Lockowandt M, et al. Striatonigral neurons divide into two distinct morphological-physiological phenotypes after chronic L-DOPA treatment in parkinsonian rats. Sci Rep. 2018;8(1):10068.
Shin DS, Carlen PL. Enhanced Ih depresses rat entopeduncular nucleus neuronal activity from high-frequency stimulation or raised Ke+. J Neurophysiol. 2008;99(5):2203–19.
Bramham CR, Srebro B. Induction of long-term depression and potentiation by low- and high-frequency stimulation in the dentate area of the anesthetized rat: magnitude, time course and EEG. Brain Res. 1987;405(1):100–7.
Ye H, Buttigieg J, Wan Y, Wang J, Figley S, Fehlings MG. Expression and functional role of BK channels in chronically injured spinal cord white matter. Neurobiol Dis. 2012;47(2):225–36.
Feng Z, Yu Y, Guo Z, Cao J, Durand DM. High frequency stimulation extends the refractory period and generates axonal block in the rat hippocampus. Brain Stimul. 2014;7(5):680–9.
Sutton AC, Yu W, Calos ME, Mueller LE, Berk M, Shim J, et al. Elevated potassium provides an ionic mechanism for deep brain stimulation in the hemiparkinsonian rat. Eur J Neurosci. 2013;37(2):231–41.
Bikson M, Lian J, Hahn PJ, Stacey WC, Sciortino C, Durand DM. Suppression of epileptiform activity by high frequency sinusoidal fields in rat hippocampal slices. J Physiol. 2001;531(Pt 1):181–91.
Lian J, Bikson M, Sciortino C, Stacey WC, Durand DM. Local suppression of epileptiform activity by electrical stimulation in rat hippocampus in vitro. J Physiol. 2003;547(Pt 2):427–34.
Monai H, Ohkura M, Tanaka M, Oe Y, Konno A, Hirai H, et al. Calcium imaging reveals glial involvement in transcranial direct current stimulation-induced plasticity in mouse brain. Nat Commun. 2016;7:11100.
Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van Essen DC, Raichle ME. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. P Natl Acad Sci USA. 2005;102(27):9673–8.
Marques JC, Li M, Schaak D, Robson DN, Li JM. Internal state dynamics shape brainwide activity and foraging behaviour. Nature. 2020;577(7789):239–43.
Murris SR, Arsenault JT, Vanduffel W. Frequency- and state-dependent network effects of electrical stimulation targeting the ventral tegmental area in macaques. Cereb Cortex. 2020;30(8):4281–96.
Silvanto J, Muggleton NG. New light through old windows: moving beyond the “virtual lesion” approach to transcranial magnetic stimulation. Neuroimage. 2008;39(2):549–52.
Pasley BN, Allen EA, Freeman RD. State-dependent variability of neuronal responses to transcranial magnetic stimulation of the visual cortex. Neuron. 2009;62(2):291–303.
Uhrig L, Sitt JD, Jacob A, Tasserie J, Barttfeld P, Dupont M, et al. Resting-state dynamics as a cortical signature of anesthesia in monkeys. Anesthesiology. 2018;129(5):942–58.
Paasonen J, Stenroos P, Salo RA, Kiviniemi V, Grohn O. Functional connectivity under six anesthesia protocols and the awake condition in rat brain. Neuroimage. 2018;172:9–20.
Li G, Henriquez CS, Frohlich F. Rhythmic modulation of thalamic oscillations depends on intrinsic cellular dynamics. J Neural Eng. 2019;16(1): 016013.
Yang H, Wang H, Guo L, Xu G. Dynamic responses of neurons in different states under magnetic field stimulation. J Comput Neurosci. 2022;50(1):109–20.
Keil J, Timm J, Sanmiguel I, Schulz H, Obleser J, Schonwiesner M. Cortical brain states and corticospinal synchronization influence TMS-evoked motor potentials. J Neurophysiol. 2014;111(3):513–9.
Ruddy K, Balsters J, Mantini D, Liu Q, Kassraian-Fard P, Enz N, et al. Neural activity related to volitional regulation of cortical excitability. Elife. 2018;7: e40843.
Ye H, Chen V, Hendee J. Cellular mechanisms underlying state-dependent neural inhibition with magnetic stimulation. Sci Rep. 2022;12(1):12131.
Furukawa T, Furshpan EJ. Two inhibitory mechanisms in the Mauthner neurons of goldfish. J Neurophysiol. 1963;26:140–76.
Anastassiou CA, Montgomery SM, Barahona M, Buzsaki G, Koch C. The effect of spatially inhomogeneous extracellular electric fields on neurons. J Neurosci. 2010;30(5):1925–36.
Dudek FE, Yasumura T, Rash JE. ‘Non-synaptic’ mechanisms in seizures and epileptogenesis. Cell Biol Int. 1998;22(11–12):793–805.
Amzica F, Steriade M. Neuronal and glial membrane potentials during sleep and paroxysmal oscillations in the neocortex. J Neurosci. 2000;20(17):6648–65.
Martinez-Banaclocha M. Astroglial isopotentiality and calcium-associated biomagnetic field effects on cortical neuronal coupling. Cells. 2020;9(2):439.
Ghai RS, Bikson M, Durand DM. Effects of applied electric fields on low-calcium epileptiform activity in the CA1 region of rat hippocampal slices. J Neurophysiol. 2000;84(1):274–80.
Majhi S, Ghosh D. Alternating chimeras in networks of ephaptically coupled bursting neurons. Chaos. 2018;28(8): 083113.
Capllonch-Juan M, Sepulveda F. Modelling the effects of ephaptic coupling on selectivity and response patterns during artificial stimulation of peripheral nerves. PLoS Comput Biol. 2020;16(6): e1007826.
Nowak LG, Bullier J. Axons, but not cell bodies, are activated by electrical stimulation in cortical gray matter. II. Evidence from selective inactivation of cell bodies and axon initial segments. Exp Brain Res. 1998;118(4):489–500.
Werginz P, Fried SI, Rattay F. Influence of the sodium channel band on retinal ganglion cell excitation during electric stimulation—a modeling study. Neuroscience. 2014;266:162–77.
Cameron MA, Al Abed A, Buskila Y, Dokos S, Lovell NH, Morley JW. Differential effect of brief electrical stimulation on voltage-gated potassium channels. J Neurophysiol. 2017;117(5):2014–24.
Agren R, Nilsson J, Arhem P. Closed and open state dependent block of potassium channels cause opposing effects on excitability—a computational approach. Sci Rep. 2019;9(1):8175.
Hedrich UB, Liautard C, Kirschenbaum D, Pofahl M, Lavigne J, Liu Y, et al. Impaired action potential initiation in GABAergic interneurons causes hyperexcitable networks in an epileptic mouse model carrying a human Na(V)1.1 mutation. J Neurosci. 2014;34(45):14874–89.
Martin MS, Dutt K, Papale LA, Dube CM, Dutton SB, de Haan G, et al. Altered function of the SCN1A voltage-gated sodium channel leads to gamma-aminobutyric acid-ergic (GABAergic) interneuron abnormalities. J Biol Chem. 2010;285(13):9823–34.
Karimi-Abdolrezaee S, Eftekharpour E, Fehlings MG. Temporal and spatial patterns of Kv1.1 and Kv1.2 protein and gene expression in spinal cord white matter after acute and chronic spinal cord injury in rats: implications for axonal pathophysiology after neurotrauma. Eur J Neurosci. 2004;19(3):577–89.
Tsai D, Morley JW, Suaning GJ, Lovell NH. Frequency-dependent reduction of voltage-gated sodium current modulates retinal ganglion cell response rate to electrical stimulation. J Neural Eng. 2011;8(6): 066007.
Luscher C, Malenka RC. NMDA receptor-dependent long-term potentiation and long-term depression (LTP/LTD). Cold Spring Harb Perspect Biol. 2012;4(6): a005710.
Kim YT, Hitchcock RW, Bridge MJ, Tresco PA. Chronic response of adult rat brain tissue to implants anchored to the skull. Biomaterials. 2004;25(12):2229–37.
Liu B, Kim E, Meggo A, Gandhi S, Luo H, Kallakuri S, et al. Enhanced biocompatibility of neural probes by integrating microstructures and delivering anti-inflammatory agents via microfluidic channels. J Neural Eng. 2017;14(2): 026008.
Bonmassar G, Lee SW, Freeman DK, Polasek M, Fried SI, Gale JT. Microscopic magnetic stimulation of neural tissue. Nat Commun. 2012;3:921.
Skach J, Conway C, Barrett L, Ye H. Axonal blockage with microscopic magnetic stimulation. Sci Rep. 2020;10(1):18030.
Ye H, Barrett L. Somatic inhibition by microscopic magnetic stimulation. Sci Rep. 2021;11(1):13591.
Ye H, Chen VC, Helon J, Apostolopoulos N. Focal suppression of epileptiform activity in the hippocampus by a high-frequency magnetic field. Neuroscience. 2020;432:1–14.
Lee SW, Fallegger F, Casse BD, Fried SI. Implantable microcoils for intracortical magnetic stimulation. Sci Adv. 2016;2(12): e1600889.
Golestanirad L, Gale JT, Manzoor NF, Park HJ, Glait L, Haer F, et al. Solenoidal micromagnetic stimulation enables activation of axons with specific orientation. Front Physiol. 2018;9:724.
Domacena J, Ruan J, Ye H. Improving suction technology for nerve activity recording. J Neurosci Methods. 2022;365: 109401.
Buzsaki G, Anastassiou CA, Koch C. The origin of extracellular fields and currents–EEG, ECoG, LFP and spikes. Nat Rev Neurosci. 2012;13(6):407–20.
Kent AR, Grill WM. Neural origin of evoked potentials during thalamic deep brain stimulation. J Neurophysiol. 2013;110(4):826–43.
Wilkinson CM, Burrell JI, Kuziek JWP, Thirunavukkarasu S, Buck BH, Mathewson KE. Predicting stroke severity with a 3-min recording from the Muse portable EEG system for rapid diagnosis of stroke. Sci Rep. 2020;10(1):18465.
Suleiman A, Lithgow B, Dastgheib Z, Mansouri B, Moussavi Z. Quantitative measurement of post-concussion syndrome using electrovestibulography. Sci Rep. 2017;7(1):16371.
Kent AR, Grill WM. Analysis of deep brain stimulation electrode characteristics for neural recording. J Neural Eng. 2014;11(4): 046010.
Priori A, Foffani G, Rossi L, Marceglia S. Adaptive deep brain stimulation (aDBS) controlled by local field potential oscillations. Exp Neurol. 2013;245:77–86.
Allen EA, Pasley BN, Duong T, Freeman RD. Transcranial magnetic stimulation elicits coupled neural and hemodynamic consequences. Science. 2007;317(5846):1918–21.
Stefanou MI, Baur D, Belardinelli P, Bergmann TO, Blum C, Gordon PC, et al. Brain state-dependent brain stimulation with real-time electroencephalography-triggered transcranial magnetic stimulation. J Vis Exp. 2019;150: e59711.
Bohning DE, Shastri A, McConnell KA, Nahas Z, Lorberbaum JP, Roberts DR, et al. A combined TMS/fMRI study of intensity-dependent TMS over motor cortex. Biol Psychiatry. 1999;45(4):385–94.
Ridding MC, Ziemann U. Determinants of the induction of cortical plasticity by non-invasive brain stimulation in healthy subjects. J Physiol. 2010;588(Pt 13):2291–304.
Siebner HR, Lang N, Rizzo V, Nitsche MA, Paulus W, Lemon RN, et al. Preconditioning of low-frequency repetitive transcranial magnetic stimulation with transcranial direct current stimulation: evidence for homeostatic plasticity in the human motor cortex. J Neurosci. 2004;24(13):3379–85.
Hurley R, Machado L. Using tDCS priming to improve brain function: can metaplasticity provide the key to boosting outcomes? Neurosci Biobehav Rev. 2017;83:155–9.
Potok W, Bachinger M, van der Groen O, Cretu AL, Wenderoth N. Transcranial random noise stimulation acutely lowers the response threshold of human motor circuits. J Neurosci. 2021;41(17):3842–53.
McLean MJ, Engstrom S, Holcomb RR, Sanchez D. A static magnetic field modulates severity of audiogenic seizures and anticonvulsant effects of phenytoin in DBA/2 mice. Epilepsy Res. 2003;55(1–2):105–16.
McLean MJ, Engstrom S, Qinkun Z, Spankovich C, Polley DB. Effects of a static magnetic field on audiogenic seizures in black Swiss mice. Epilepsy Res. 2008;80(2–3):119–31.
Gersner R, Dhamne SC, Zangen A, Pascual-Leone A, Rotenberg A. Bursts of high-frequency repetitive transcranial magnetic stimulation (rTMS), together with lorazepam, suppress seizures in a rat kainate status epilepticus model. Epilepsy Behav. 2016;62:136–9.
HabibollahiSaatlou F, Rogasch NC, McNair NA, Biabani M, Pillen SD, Marshall TR, et al. MAGIC: an open-source MATLAB toolbox for external control of transcranial magnetic stimulation devices. Brain Stimul. 2018;11(5):1189–91.
Tanskanen JMA, Ahtiainen A, Hyttinen JAK. Toward closed-loop electrical stimulation of neuronal systems: a review. Bioelectricity. 2020;2(4):328–47.
Leite J, Morales-Quezada L, Carvalho S, Thibaut A, Doruk D, Chen CF, et al. Surface EEG-transcranial direct current stimulation (tDCS) closed-loop system. Int J Neural Syst. 2017;27(6):1750026.
Ganzer PD, Darrow MJ, Meyers EC, Solorzano BR, Ruiz AD, Robertson NM, et al. Closed-loop neuromodulation restores network connectivity and motor control after spinal cord injury. Elife. 2018;7: e32058.
Joucla S, Gliere A, Yvert B. Current approaches to model extracellular electrical neural microstimulation. Front Comput Neurosci. 2014;8:13.
Tai C, de Groat WC, Roppolo JR. Simulation of nerve block by high-frequency sinusoidal electrical current based on the Hodgkin-Huxley model. IEEE Trans Neural Syst Rehabilit Eng. 2005;13(3):415–22.
Tai C, Roppolo JR, de Groat WC. Analysis of nerve conduction block induced by direct current. J Comput Neurosci. 2009;27(2):201–10.
McIntyre CC, Grill WM, Sherman DL, Thakor NV. Cellular effects of deep brain stimulation: model-based analysis of activation and inhibition. J Neurophysiol. 2004;91(4):1457–69.
Vermaas M, Piastra MC, Oostendorp TF, Ramsey NF, Tiesinga PHE. FEMfuns: a volume conduction modeling pipeline that includes resistive, capacitive or dispersive tissue and electrodes. Neuroinformatics. 2020;18(4):569–80.
Acknowledgements
We thank the two anonymous reviewers for their many insightful comments and suggestions.
Funding
This work was supported by the National Institute of Neurological Disorders and Stroke of the National Institutes of Health (R03NS130387), and a research support grant (1100) from Loyola University Chicago. Joyce Ruan was supported by a LUROP research fellowship award from Loyola University Chicago.
Author information
Authors and Affiliations
Contributions
HY conceptualized the project. HY, JH, JR, AZ, JY, and MD worked together to draft and revise the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ye, H., Hendee, J., Ruan, J. et al. Neuron matters: neuromodulation with electromagnetic stimulation must consider neurons as dynamic identities. J NeuroEngineering Rehabil 19, 116 (2022). https://doi.org/10.1186/s12984-022-01094-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12984-022-01094-4