
Abstract
CircRNAs are a novel type of RNAs. With the newly developed technology of next-generation sequencing (NGS), especially RNA-seq technology, over 30,000 circRNAs have already been found. Owing to their unique structure, they are more stable than linear RNAs. CircRNAs play important roles in the carcinogenesis of cancer. The expression of circRNAs is correlated with patients’ clinical characteristics, and circRNAs play a vital role in many aspects of malignant phenotypes, including cell cycle, apoptosis, vascularization, and invasion; metastasis as a RNA sponge, binding to RBP; or translation. Therefore, it is meaningful to further study the mechanism of interactions between circRNAs and tumors. The role of circRNAs as molecular markers or potential targets will provide promising application perspectives, such as early tumor diagnosis, therapeutic evaluation, prognosis prediction, and even gene therapy for tumors.
Similar content being viewed by others


Background
Circular RNAs (circRNAs) are a large group of RNAs that form covalently closed continuous loops. The first circRNA was found in a RNA virus in 1976 [1, 2]. Not long after that, Hsu MT et al. discovered the existence of a circular form of RNA in the cytoplasm of monkey renal CV-1 cells using electron microscopy [3]. In 1996, circRNAs were found to consist of exons from RNA transcripts in human cells [4]. In recent years, an increasing number of circRNAs have been found owing to the development of sequencing technologies, with the RNA-seq technology of next-generation sequencing being the most notable. To date, the total number of known circRNAs has reached up to 30,000.
CircRNAs have long been regarded as an accidental byproduct, resulting from errors during post-transcriptional processing, and recent studies have found important functions of circRNAs in multiple biological processes [5]. CircRNAs can act in a broad range of roles, including as miRNA sponges [6] and regulators of RNA binding protein (RBP) in modulating the expression of protein-encoding genes [7]. Some circRNAs even encode peptides [8]. Thus, circRNA disorders should influence the development of a variety of human diseases, including cancer [9]. Furthermore, the unique structures of circRNAs lead to their insensitivity to ribonucleases. Thus, unlike linear RNAs, circRNA molecules can exist in tissues, serum, and urine. Owing to this characteristic, the discovery of circRNAs as biomarkers for human cancer has been shown to be a tremendous possibility.
Biogenesis of circRNAs
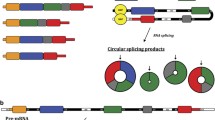
CircRNAs can be sorted into three types, including exonic circRNAs, intronic circRNAs, and retained-intron circRNAs, which are produced from different cycling mechanisms. During the formation of circRNAs, specific sequences are conserved and sequences from upstream and downstream regions are linked together in a reverse direction, all of which contribute to mature circRNAs by back splicing [10]. Although the efficiency of back splicing is much lower than in linear RNAs [5], there is still a large intracellular abundance of circRNAs due to their stability and relatively long half-life period [11]. The earliest back splicing mechanism is dependent on complementary intron matches [12], which widely exist in the exonic circRNA formation process. CircRNAs structured from this mechanism have special repeated sequences, such as Alu and other short sequences, on their two sides [13, 14]. After transcription into RNA precursors, the exon sequences on the two sides of the cycling region will complementarily match with each other under the promotion of repetitive sequences, such as Alu, to form a circle shape. Following this, the spliceosome binds to the cycled RNA precursor under the effect of U6 and U2. Then, it selectively cuts out the exon in the cycling region by cooperating with the protein complex and the U5 core [15, 16]. At the same time, exon sequences connection are reversely cut, forming mature circRNA.
The mechanism of intronic circRNA formation is different from that of exonic circRNA. The formation of intronic circRNAs relies on GU-rich sequences near the 5′ splice site and C-rich sequences near the branch point. During the back splicing process, the two segments bind into a circle first. Then exonic sequences and intronic sequences in the binding part are cut out by the spliceosome. The remaining introns are eventually pieced together to form mature circRNA [17, 18]. Recently, Li et al. found another novel type of circRNA consisting of both exons and introns that overexpress themselves according to their complementary sequences [7]; however, the exact splicing mechanism has yet to be determined (Fig. 1a).

The biogenesis of circRNAs. a Alternative splicing of an RNA precursor. A linear RNA precursor can be cis-spliced into linear RNA, including mRNA and lncRNA. Special sequences, such as Alu, in the RNA precursor can combine to form a cycle and lead the spliceosome to produce different types of circRNAs through back splicing. b Different circRNAs have different back splicing mechanisms. Exonic circRNAs have cycling sequences, such as the Alu sequence, on the two sides. The two sequences bind to each other complementarily and are then spliced by the spliceosome, which consists of U2 and U6. Intronic circRNAs form a cycle through the combination of upstream introns of the GU rich sequence and the downstream C rich sequence, and they are spliced by spliceosome
Similar to linear RNA, the biogenesis of circRNAs is regulated by various types of proteins, such as the RNA spliceosome and RNA binding protein (RBP). The RNA spliceosome and spicing process are vital research frontiers in biomedicine. The RNA spliceosome is a RNA-based enzyme with a U5 core consisting of 5 snRNAs and a number of proteins. Under the effect the U6 and U2 promoters, many proteins cooperatively process pre-RNAs [16]. Similar to mRNA formation, circRNAs can be spliced from linear RNA precursors by the spliceosome. Furthermore, one linear RNA molecule can be processed into different types of RNAs, including mRNA, circRNA, and lncRNA, through different splicing events. For example, HIPK3 pre-mRNA can be spliced into HIPK3 mRNA and its second exon can also form circRNA hsa_circ_0018082 [13]. There is also a competition between inverted splicing of circRNA, mRNA, and lncRNA during splicing [19], which may be a regulation mechanism of circRNAs. Another regulated protein is the Quaking (QKI) protein, a representative molecule. It can bind to specific downstream sites on the linear pre mRNA sequence and promote cycling and inverted splicing of circRNAs [20] (Fig. 1b).
Databases of circRNAs
With the application of next-generation sequencing and bioinformatics, many circRNAs are being discovered and integrated into the circRNA database [9] (Table 1). The Cirbase and Deepbase 2.0 databases contain over 150,000 circRNA genomes and mature RNA sequences of different species, including human, fruit fly, and nematode [21–23]. The Cir2Traits database contains circRNAs relating to various diseases [24]. Circnet and circRNAbase contain alternatively spliced variants and their expression patterns in different tissues and possible miRNA regulated networks [21, 25]. The CircInteractome database contains corresponding genome sequences, primers for qRT-PCR, and siRNAs of mature circRNAs. It can also be used to predict RBP binding sites [26]. The CIRCpedia and TSCD databases contain RNA-seq data of several human cell lines and samples [27, 28].
CircRNAs can be a novel biomarker in cancer
Unlike linear RNA, 3′ and 5′ terminals are not exposed in circRNAs; thus, they are not sensitive to ribonucleases, such as exonuclease or RNase R and have a longer half-life period with a more prominent specificity in histology and pathology [29]. Meanwhile, circRNAs are easier to extract and detect with higher specificity compared with proteins; thus, circRNAs are an ideal molecular marker for cancer. With the application of RNA-seq [12, 30], changes in the expression of circRNAs are found to be prevalent in many cancers, including esophageal cancer [31], glioma [32], and hepatocellular carcinoma [33]. Further experiments and analysis on clinical files confirm that the expression levels of some circRNAs vary according to TNM stage, metastasis, and size the of tumor in esophageal cancer [34], hepatocellular carcinoma [35, 36], and colorectal cancer [37]. Free circRNA molecules are degraded in the extracellular environment, but studies show that circRNAs that are produced intracellularly can be secreted to the exterior of an exosome and be restored after an extended period of time. Li et al. found high expression of circ_0001649 in patients with liver cancer through RNA sequencing and qRT-PCR, and the expression level in the exosome is closely related to clinical characteristics, such as tumor size [33] (Table 2).
Because of their unique structure, circRNAs are more stable in tissues, serum, savory, or urine compared with circulating tumor DNAs (ctDNAs) or linear RNAs. There is also a greater possibility to screen for sensitive and specific molecular biomarkers from thousands of circRNAs with regard to the relatively small number of miRNAs. Testing circRNAs by RT-PCR and in-situ hybridization is also more specific and sensitive than detecting proteins by an antigen-antibody reaction. In conclusion, the use of circRNAs outweighs the use of current tumor biomarkers in many fields, and they could be promising molecular biomarkers and new targets for future drugs.
The function of circRNAs in human cancer
CircRNAs are widely involved in physiological and pathological processes, and studies have found that circRNAs can be involved in biological activities by acting as a miRNA sponge [6], binding RBPs [7], and translating peptides [8, 38]. Of these, the role of circRNAs as a miRNA sponge is the main mechanism of circRNAs in tumor cells. There are many miRNA binding sites on circRNAs. Thus, circRNAs can bind to miRNA as a RNA sponge and can regulate miRNA and downstream gene expression through a ceRNA mechanism and contribute to tumor progression. The following examples present current studies on circRNAs and their functions in human cancer.
ciRS-7
ciRS-7, also called CDT1as, is one of the earliest known circRNAs and regulates miRNA in tumor cells [39]. ciRS-7 is spliced from the antisense transcript of the CDR1 gene. It was first found to be upregulated in malignant tumor cell lines, such as HeLa cells, and further studies found more than seventy miRNA binding sites in ciRS-7 [40]. Later, it was discovered that ciRS-7 can bind to miR7 as a miRNA sponge to downregulate miR-7 expression [41]. miR-671 was found to be another binding site, and the combination of the two molecules mediates the Ago2-mediated cleavage of ciRS-7 to release the absorbed miR-7 [42, 43]. This suggests that miR-671 diminishes the inhibition of miR-7 expression through ciRS-7; improves the miR7 level in tumor cells; contributes to the increase of downstream target oncogenes, such as EGFR and XIAP; and promotes vascularization, metastasis and reproduction of tumor cells [41, 44].
circ-Foxo3
Despite encoding for Foxo3, the Foxo3 gene can encode for two noncoding RNAs, the pseudogene FOXO3P and circ-Foxo3. Yang et al. found that the two RNAs can both act as miRNA sponges for miR-22, miR-136*, miR-138, miR-149*, miR-433, miR-762, miR-3614-5p, and miR-3622b-5p, influencing their function by binding to them. Thus, they are involved in the progression of tumors [45]. Studies have shown that FOXO3P, circ-Foxo3, and Foxo3 mRNA expressed elsewhere can inhibit the growth of a tumor. Further studies have found high levels of circ-Foxo3 in normal cells and that it binds to p21 and CDK2 to form a circ-Foxo3-p21-CDK2 tripolymer. Thus, in this way, the function of CDK2 is restricted, leading to a decrease in cell proliferation, and the silencing of circ-Foxp3 will contribute to much faster growth in malignant tumor cells [46].
Hsa_circ_001569
Hsa_circ_001569 is a circRNA of 1776 bp that is spliced by linear a RNA precursor from the ABCC1 gene. The official name of hsa_circ_001569 is hsa_circ_0000677. Liang et al. found that the hsa_circ_001569 had significantly high expression in colorectal cancer tissue and that it correlated positively with the degree of clinical features, such as the TNM stage [37]. This means hsa_circ_001569 may be closely related to colorectal cancer progression. Another study later confirmed this, and hsa_circ_001569 can act as a sponge to bind to miR-145 and inhibit the transcriptional activity of miR-145 without influencing its expression level. Thus, has_circ_001569 leads to the upregulation of target genes of miR-145, including E2F5, BAG4, and FMNL2, and acts as a positive regulator in cell proliferation and invasion of colorectal cancer.
circHIPK3
circHIPK3 is produced by the second exon of the HIPK3 gene. Huang found higher circRNA levels, including circHIPK3, by RNA sequencing seven types of tumor samples and six types of normal tissues [13]. Silencing circHIPK3 relieves the proliferation speed of tumor cells. Further studies found that circHIPK3 binds to eighteen sites of nine miRNAs. It was found that circHIPK3 inhibits miR-124 activity by working as the sponge of miR-124; thus, it can influence the proliferation of tumor cells through genes, including iASPP [47].
Fusion-circRNAs
The genomes of tumor cells are unstable; thus, many chromosome translocations exist, especially in lymphoma. Gene fusions may lead to incorrectly spliced mRNAs, during which circRNA (fusion-circRNA,f-circRNA) comes into being. For example, the PML/RARa and MLL genes fused and produced f-circM9 and f-circPR and knockouts of f-circM9 and f-circPR lead to apoptosis of a large amount of tumor cells as well as increases their sensitivity to drugs, such as arsenic [48]. Together, these results suggest that f-circM9 and f-circPR play important roles in hematological malignancy.
cZNF292
Silencing cZNF292 in human endothelium cells will inhibit the tube formation process these cells undergo [49]. cZNF292 also exists in glioma cells [50]. Downregulation of cZNF292 inhibits the proliferation and vascularization of glioma cells and reduces the levels of cellular Cyclin A, CDK2, p-CDK2, β-catenin, p-STAT3 (Tyr705) and p-STAT5 (Tyr694). Thus, the cells are arrested in the G2/M phase due to modifications in the functioning of the Wnt/β-catenin pathway. Meanwhile, it was found that downregulation of cZNF292 contributes to decreased transcription of E2F1, NF-κB, Sp1, HIF-1, AP-1, STAT3, and STAT5, thus inhibiting tube formation of tumor cells. However, studies show that cZNF292 does not function as a RNA sponge, and the mechanism requires further study [49].
circTCF25
In 467 samples of carcinomas of the urinary bladder, Zhang et al. detected circRNAs with prominent differential expression through microarray analysis. It was confirmed by qRT-PCR that six circRNAs, including circTCF25 (hsa_circ_0041103), were highly expressed in tumor tissues. Another study demonstrated that circTCF25 can upregulate the levels of CDK6 by working as a RNA sponge for miR-103a-3p and miR-107. Thus, it promotes the proliferation and metastasis of urinary bladder carcinoma [51].
Prospect
CircRNAs are novel molecules that have been discovered in recent years. They are no longer recognized as products of transcription errors. On the contrary, they have become a popular research subject, after miRNAs and lncRNAs, owing to their important functions [52–56]. The formation of a mature circRNA molecule has at least two mechanisms, which are different from selective splicing in linear RNAs; however, the detailed molecular mechanism requires further studies. There may be a third mechanism involving the formation of retained-intron circRNAs that contain both exons and introns, and this has also yet to be explored. Moreover, different types of proteins and miRNAs have been found to take part in the processing of pre-RNAs to circRNAs. The mechanism of circRNAs formation has yet to be discovered, and it remains an attractive mystery.
A tremendous amount of data indicate that circRNAs regulate other genes at the transcriptional and post-transcriptional levels. Some can even encode proteins directly and are thus involved in physiological and pathological processes, including influencing malignant tumors. Until now, the miRNA sponge has been found to be a possible mechanism for circRNA function. They can specifically bind to miRNAs and regulate gene expression by competing with competing endogenous RNA (ceRNA), including lncRNAs, mRNAs, and pseudogenes. The participation of circRNAs in the ceRNA network renders it more complete and complex. CircRNAs compete with mRNAs from the same preRNAs during the splicing process, and circRNAs were recently found to encode for proteins. This demonstrates their involvement in biological procedures, provides a more complete understanding of the RNA language, and enriches the central dogma [57–59]. Knowledge of circRNAs still remains at a superficial level, and more functions remain to be discovered among over 30,000 molecules, as only a limited number of reports describe their functions. Exploration of their functions will bring humans closer to the essence of life.
CircRNAs are much more stable compared to linear RNAs owing to their unique structure. With regard to biological markers of cancer, circRNAs have demonstrated their wide perspective. Studies have already found differential expression of circRNAs in tissues and blood, and their expression differs from those of miRNA and lncRNA [60–62]. It is possible to build a model for prognosis by combining them with various other biomarkers and thus improve the accuracy and specificity of diagnosis. The clinical applications of circRNAs continues to expand, with their use in samples, such as saliva and urine, and research on circRNAs is indeed a very promising aspect of the study of life sciences.
Previous studies have reported that circRNAs are one type of molecular marker in tumors and that they can influence apoptosis and infiltration. CircRNAs are potential targets for new drugs. For example, the fusion-circRNA found in leukemia is associated with chemotherapy resistance, and knocking out expression of f-circM9 can lead to apoptosis of leukemia cells and decrease its drug resistance to Ara-C [48]. On the other hand, circRNAs perform their functions by mediating miRNAs through a miRNA sponge. This process can also act as a treatment target for malignant tumors. It was also revealed that circRNAs can encode multiple peptides, which can also be used to treat tumors. However, more progress needs to be made between circ-RNA and tumor treatment since the process to effectively and efficiently transform circRNAs into targeted cells is still a major problem that needs to be solved. Although nanoparticles have been applied as transforming vehicles in gene therapy and the Synergistic Photodynamic/Photothermal/Chemotherapy of Cancer drug releasing system has also made some advancement, neither have been utilized in clinical practice. Further studies are still needed to reach a definite resolution.
Conclusions
In conclusion, current achievements in the study of circRNAs and their functions are still very limited. To our excitement, this mystery will eventually be solved through scientists’ endeavors and applications of new methods and techniques. CircRNAs function in an increasingly significant way in the diagnosis and therapy of cancers and will provide precise strategies for affecting human cancers.
Abbreviations
- ABCC1:
-
ATP binding cassette subfamily C member 1
- Ago:
-
Argonaute
- BAG2:
-
BCL2 associated athanogene 4
- CDK2:
-
Cyclin dependent kinase 2
- CDK6:
-
Cyclin dependent kinase 5
- CDR1:
-
Cerebellar degeneration related protein 1
- circRNAs:
-
Circular RNAs
- ctDNAs:
-
Circulating tumor DNAs
- E2F1:
-
E2F transcription factor 1
- E2F5:
-
E2F transcription factor 5
- EGFR:
-
Epidermal growth factor receptor
- FMNL2:
-
Formin like 2
- FOXO3:
-
Forkhead box O3
- HIPK3:
-
Homeodomain interacting protein kinase 3
- iASPP:
-
Inhibitor of apoptosis-stimulating protein of p53
- miRNAs:
-
microRNAs
- NGS:
-
Next generation sequencing
- PML:
-
Promyelocytic leukemia
- RBP:
-
RNA binding protein
- START3:
-
Signal transducer and activator of transcription 3
- TCF25:
-
Transcription factor 25
- XIAP:
-
X-linked inhibitor of apoptosis
- ZNF292:
-
Zinc finger protein 292
References
Kolakofsky D. Isolation and characterization of Sendai virus DI-RNAs. Cell. 1976;8:547–55.
Sanger HL, Klotz G, Riesner D, Gross HJ, Kleinschmidt AK. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc Natl Acad Sci U S A. 1976;73:3852–6.
Hsu MT, Coca-Prados M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature. 1979;280:339–40.
Pasman Z, Been MD, Garcia-Blanco MA. Exon circularization in mammalian nuclear extracts. RNA. 1996;2:603–10.
Cocquerelle C, Mascrez B, Hetuin D, Bailleul B. Mis-splicing yields circular RNA molecules. FASEB J. 1993;7:155–60.
Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK, Kjems J. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495:384–8.
Li Z, Huang C, Bao C, Chen L, Lin M, Wang X, Zhong G, Yu B, Hu W, Dai L, et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat Struct Mol Biol. 2015;22:256–64.
Granados-Riveron JT, Aquino-Jarquin G. The complexity of the translation ability of circRNAs. Biochim Biophys Acta. 2016;1859:1245–51.
Szabo L, Salzman J. Detecting circular RNAs: bioinformatic and experimental challenges. Nat Rev Genet. 2016;17:679–92.
Vicens Q, Westhof E. Biogenesis of Circular RNAs. Cell. 2014;159:13–4.
Qu S, Yang X, Li X, Wang J, Gao Y, Shang R, Sun W, Dou K, Li H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015;365:141–8.
Zhang XO, Wang HB, Zhang Y, Lu X, Chen LL, Yang L. Complementary sequence-mediated exon circularization. Cell. 2014;159:134–47.
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen B, Luo Y, Lyu D, Li Y, Shi G, et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat Commun. 2016;7:11215.
Jeck WR, Sorrentino JA, Wang K, Slevin MK, Burd CE, Liu J, Marzluff WF, Sharpless NE. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA. 2013;19:141–57.
Hang J, Wan R, Yan C, Shi Y. Structural basis of pre-mRNA splicing. Science. 2015;349:1191–8.
Yan C, Wan R, Bai R, Huang G, Shi Y. Structure of a yeast activated spliceosome at 3.5 A resolution. Science. 2016;353:904–11.
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin QF, Xing YH, Zhu S, Yang L, Chen LL. Circular intronic long noncoding RNAs. Mol Cell. 2013;51:792–806.
Talhouarne GJ, Gall JG. Lariat intronic RNAs in the cytoplasm of Xenopus tropicalis oocytes. RNA. 2014;20:1476–87.
Chen LL, Yang L. Regulation of circRNA biogenesis. RNA Biol. 2015;12:381–8.
Conn SJ, Pillman KA, Toubia J, Conn VM, Salmanidis M, Phillips CA, Roslan S, Schreiber AW, Gregory PA, Goodall GJ. The RNA binding protein quaking regulates formation of circRNAs. Cell. 2015;160:1125–34.
Li JH, Liu S, Zhou H, Qu LH, Yang JH. starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014;42:D92–7.
Glazar P, Papavasileiou P, Rajewsky N. circBase: a database for circular RNAs. RNA. 2014;20:1666–70.
Zheng LL, Li JH, Wu J, Sun WJ, Liu S, Wang ZL, Zhou H, Yang JH, Qu LH. deepBase v2.0: identification, expression, evolution and function of small RNAs, LncRNAs and circular RNAs from deep-sequencing data. Nucleic Acids Res. 2016;44:D196–202.
Ghosal S, Das S, Sen R, Basak P, Chakrabarti J. Circ2Traits: a comprehensive database for circular RNA potentially associated with disease and traits. Front Genet. 2013;4:283.
Liu YC, Li JR, Sun CH, Andrews E, Chao RF, Lin FM, Weng SL, Hsu SD, Huang CC, Cheng C, et al. CircNet: a database of circular RNAs derived from transcriptome sequencing data. Nucleic Acids Res. 2016;44:D209–15.
Dudekula DB, Panda AC, Grammatikakis I, De S, Abdelmohsen K, Gorospe M. CircInteractome: A web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016;13:34–42.
Zhang XO, Dong R, Zhang Y, Zhang JL, Luo Z, Zhang J, Chen LL, Yang L. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016;26:1277–87.
Xia S, Feng J, Lei L, Hu J, Xia L, Wang J, Xiang Y, Liu L, Zhong S, Han L, He C. Comprehensive characterization of tissue-specific circular RNAs in the human and mouse genomes. Brief Bioinform. 2016. [Epub ahead of print]. doi:10.1093/bib/bbw081.
Suzuki H, Zuo Y, Wang J, Zhang MQ, Malhotra A, Mayeda A. Characterization of RNase R-digested cellular RNA source that consists of lariat and circular RNAs from pre-mRNA splicing. Nucleic Acids Res. 2006;34:e63.
Gao Y, Wang J, Zhao F. CIRI: an efficient and unbiased algorithm for de novo circular RNA identification. Genome Biol. 2015;16:4.
Su H, Lin F, Deng X, Shen L, Fang Y, Fei Z, Zhao L, Zhang X, Pan H, Xie D, et al. Profiling and bioinformatics analyses reveal differential circular RNA expression in radioresistant esophageal cancer cells. J Transl Med. 2016;14:225.
Song X, Zhang N, Han P, Moon BS, Lai RK, Wang K, Lu W. Circular RNA profile in gliomas revealed by identification tool UROBORUS. Nucleic Acids Res. 2016;44:e87.
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J, Chen D, Gu J, He X, Huang S. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res. 2015;25:981–4.
Xuan L, Qu L, Zhou H, Wang P, Yu H, Wu T, Wang X, Li Q, Tian L, Liu M, Sun Y. Circular RNA: a novel biomarker for progressive laryngeal cancer. Am J Transl Res. 2016;8:932–9.
Qin M, Liu G, Huo X, Tao X, Sun X, Ge Z, Yang J, Fan J, Liu L, Qin W. Hsa_circ_0001649: A circular RNA and potential novel biomarker for hepatocellular carcinoma. Cancer Biomark. 2016;16:161–9.
Shang X, Li G, Liu H, Li T, Liu J, Zhao Q, Wang C. Comprehensive Circular RNA Profiling Reveals That hsa_circ_0005075, a New Circular RNA Biomarker, Is Involved in Hepatocellular Crcinoma Development. Medicine (Baltimore). 2016;95:e3811.
Xie H, Ren X, Xin S, Lan X, Lu G, Lin Y, Yang S, Zeng Z, Liao W, Ding YQ, Liang L. Emerging roles of circRNA_001569 targeting miR-145 in the proliferation and invasion of colorectal cancer. Oncotarget. 2016;7:26680–91.
Wang Y, Wang Z. Efficient backsplicing produces translatable circular mRNAs. RNA. 2015;21:172–9.
Peng L, Yuan XQ, Li GC. The emerging landscape of circular RNA ciRS-7 in cancer (Review). Oncol Rep. 2015;33:2669–74.
Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer M, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495:333–8.
Xu H, Guo S, Li W, Yu P. The circular RNA Cdr1as, via miR-7 and its targets, regulates insulin transcription and secretion in islet cells. Sci Rep. 2015;5:12453.
Chou YT, Lin HH, Lien YC, Wang YH, Hong CF, Kao YR, Lin SC, Chang YC, Lin SY, Chen SJ, et al. EGFR promotes lung tumorigenesis by activating miR-7 through a Ras/ERK/Myc pathway that targets the Ets2 transcriptional repressor ERF. Cancer Res. 2010;70:8822–31.
Hansen TB, Wiklund ED, Bramsen JB, Villadsen SB, Statham AL, Clark SJ, Kjems J. miRNA-dependent gene silencing involving Ago2-mediated cleavage of a circular antisense RNA. EMBO J. 2011;30:4414–22.
Hansen TB, Kjems J, Damgaard CK. Circular RNA and miR-7 in cancer. Cancer Res. 2013;73:5609–12.
Yang W, Du WW, Li X, Yee AJ, Yang BB. Foxo3 activity promoted by non-coding effects of circular RNA and Foxo3 pseudogene in the inhibition of tumor growth and angiogenesis. Oncogene. 2016;35(30):3919-31.
Du WW, Yang W, Liu E, Yang Z, Dhaliwal P, Yang BB. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016;44:2846-58.
Chen J, Xiao H, Huang Z, Hu Z, Qi T, Zhang B, Tao X, Liu SH. MicroRNA124 regulate cell growth of prostate cancer cells by targeting iASPP. Int J Clin Exp Pathol. 2014;7:2283–90.
Guarnerio J, Bezzi M, Jeong JC, Paffenholz SV, Berry K, Naldini MM, Lo-Coco F, Tay Y, Beck AH, Pandolfi PP. Oncogenic Role of Fusion-circRNAs Derived from Cancer-Associated Chromosomal Translocations. Cell. 2016;165:289–302.
Boeckel JN, Jae N, Heumuller AW, Chen W, Boon RA, Stellos K, Zeiher AM, John D, Uchida S, Dimmeler S. Identification and Characterization of Hypoxia-Regulated Endothelial Circular RNA. Circ Res. 2015;117:884–90.
Yang P, Qiu Z, Jiang Y, Dong L, Yang W, Gu C, Li G, Zhu Y. Silencing of cZNF292 circular RNA suppresses human glioma tube formation via the Wnt/beta-catenin signaling pathway. Oncotarget. 2016;7:63449-55.
Zhong Z, Lv M, Chen J. Screening differential circular RNA expression profiles reveals the regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in bladder carcinoma. Sci Rep. 2016;6:30919.
Bo H, Gong Z, Zhang W, Li X, Zeng Y, Liao Q, Chen P, Shi L, Lian Y, Jing Y, et al. Upregulated long non-coding RNA AFAP1-AS1 expression is associated with progression and poor prognosis of nasopharyngeal carcinoma. Oncotarget. 2015;6:20404–18.
Gong Z, Yang Q, Zeng Z, Zhang W, Li X, Zu X, Deng H, Chen P, Liao Q, Xiang B, et al. An integrative transcriptomic analysis reveals p53 regulated miRNA, mRNA, and lncRNA networks in nasopharyngeal carcinoma. Tumour Biol. 2016;37:3683–95.
He B, Li W, Wu Y, Wei F, Gong Z, Bo H, Wang Y, Li X, Xiang B, Guo C, et al. Epstein-Barr virus-encoded miR-BART6-3p inhibits cancer cell metastasis and invasion by targeting long non-coding RNA LOC553103. Cell Death Dis. 2016;7:e2353.
Li Q, Chen P, Zeng Z, Liang F, Song Y, Xiong F, Li X, Gong Z, Zhou M, Xiang B, et al. Yeast two-hybrid screening identified WDR77 as a novel interacting partner of TSC22D2. Tumour Biol. 2016;37:12503–12.
Liang F, Li Q, Li X, Li Z, Gong Z, Deng H, Xiang B, Zhou M, Li X, Li G, et al. TSC22D2 interacts with PKM2 and inhibits cell growth in colorectal cancer. Int J Oncol. 2016;49:1046–56.
Wang Y, Xue D, Li Y, Pan X, Zhang X, Kuang B, Zhou M, Li X, Xiong W, Li G, et al. The Long Noncoding RNA MALAT-1 is A Novel Biomarker in Various Cancers: A Meta-analysis Based on the GEO Database and Literature. J Cancer. 2016;7:991–1001.
Zeng Z, Bo H, Gong Z, Lian Y, Li X, Li X, Zhang W, Deng H, Zhou M, Peng S, et al. AFAP1-AS1, a long noncoding RNA upregulated in lung cancer and promotes invasion and metastasis. Tumour Biol. 2016;37:729–37.
Zhou Y, Liao Q, Li X, Wang H, Wei F, Chen J, Yang J, Zeng Z, Guo X, Chen P, et al. HYOU1, Regulated by LPLUNC1, Is Up-Regulated in Nasopharyngeal Carcinoma and Associated with Poor Prognosis. J Cancer. 2016;7:367–76.
Gong Z, Zhang S, Zhang W, Huang H, Li Q, Deng H, Ma J, Zhou M, Xiang J, Wu M, et al. Long non-coding RNAs in cancer. Sci China Life Sci. 2012;55:1120–4.
Huang M, Zhong Z, Lv M, Shu J, Tian Q, Chen J. Comprehensive analysis of differentially expressed profiles of lncRNAs and circRNAs with associated co-expression and ceRNA networks in bladder carcinoma. Oncotarget. 2016;7:47186–200.
Zhang W, Huang C, Gong Z, Zhao Y, Tang K, Li X, Fan S, Shi L, Li X, Zhang P, et al. Expression of LINC00312, a long intergenic non-coding RNA, is negatively correlated with tumor size but positively correlated with lymph node metastasis in nasopharyngeal carcinoma. J Mol Histol. 2013;44:545–54.
Acknowledgements
We thank Dr. Ruoyu Zhou for the helpful discussion.
Funding
This work was supported in part by grants from the National Natural Science Foundation of China (81372907, 81301757, 81472531, 81402009, 81572787, and 81672683) and the Natural Science Foundation of Hunan Province (14JJ1010 and 2015JJ1022).
Availability of data and material
Not applicable.
Authors’ contributions
Y.W., Y.M., Z.G., S.Z., F.X., B.X., M.Z., Q.L., W.Z., X.L. and X.L. collected the related paper and drafted the manuscript. Y.L., G.L., W.X. and Z.Z. participated in the design of the review and draft the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wang, Y., Mo, Y., Gong, Z. et al. Circular RNAs in human cancer. Mol Cancer 16, 25 (2017). https://doi.org/10.1186/s12943-017-0598-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-017-0598-7