
Abstract
Background
The genomic information available for Pediococcus pentosaceus is primarily derived from fermented fruits and vegetables, with less information available from fermented meat. P. pentosaceus LL-07, a strain isolated from fermented meat, has the capability of producing exopolysaccharides (EPS). To assess the probiotic attributes of P. pentosaceus LL-07, we conducted whole-genome sequencing (WGS) using the PacBio SequelIIe and Illumina MiSeq platforms, followed by in vitro experiments to explore its probiotic potential.
Results
The genome size of P. pentosaceus LL-07 is 1,782,685 bp, comprising a circular chromosome and a circular plasmid. Our investigation revealed the absence of a CRISPR/Cas system. Sugar fermentation experiments demonstrated the characteristics of carbohydrate metabolism. P. pentosaceus LL-07 contains an EPS synthesis gene cluster consisting of 13 genes, which is different from the currently known gene cluster structure. NO genes associated with hemolysis or toxin synthesis were detected. Additionally, eighty-six genes related to antibiotic resistance were identified but not present in the prophage, transposon or plasmid. In vitro experiments demonstrated that P. pentosaceus LL-07 was comparable to the reference strain P. pentosaceus ATCC25745 in terms of tolerance to artificial digestive juice and bile, autoaggregation and antioxidation, and provided corresponding genomic evidence.
Conclusion
This study confirmed the safety and probiotic properties of P. pentosaceus LL-07 via complete genome and phenotype analysis, supporting its characterization as a potential probiotic candidate.
Similar content being viewed by others


Background
Probiotics have received considerable attention due to their positive impact on human health. They can improve the intestinal environment, enhance immunity, nutritional status, and prevent chronic diseases [1]. Probiotics have a wide range of applications in food, agriculture, and pharmaceutical industries. In order to play a beneficial role, probiotics need to colonize the intestinal tract in a live form. Therefore, candidate probiotic strains must possess specific characteristics, including digestive fluid and bile salt tolerance, adhesion, and safety [2].
Pediococcus pentosaceus, a type of lactic acid bacteria (LAB), is a generally recognized as safe (GRAS) strain. In addition to the acid-producing function of LAB, P. pentosaceus is known to significantly enhance the flavor, quality, and safety of fermented products [3,4,5,6]. Research indicates that P. pentosaceus and its bacteriocins are closely linked to human gastrointestinal tract health (GIT) [7, 8]. Experimental evidence suggests that P. pentosaceus has potential as a probiotic, in addition to its use as a food starter and biological preservative [9,10,11].
Exopolysaccharides (EPS) are secondary metabolites produced by microorganisms during their growth and metabolism. EPS has a variety of physiological functions [12] and plays a specific role in improving food texture [13]. Some strains of P. pentosaceus produce EPS with active functions during the growth and metabolism, in addition to generating active substances such as bacteriocins [14, 15]. The function of EPS derived from P. pentosaceus has been scientifically studied. Researchers are further exploring and confirming these findings, which suggest a potential connection between the probiotic function of P. pentosaceus and EPS. [16].
Advances in genome sequencing technology and the increasing availability of sequence analysis tools have made genome characterization of microorganisms possible and essential for defining accurate classification of new strains, useful information, and their safety. [17, 18]. The study of genome sequences enables a rigorous safety assessment, including the determination of antimicrobial resistance and the presence of virulence factors. These aspects are crucial when introducing new strains into food applications [19, 20]. Since the first P. pentosaceus strain was sequenced, as of November 2023, 150 data points related to its genome have been published in the NCBI database, with only 30 complete genome sequences. Of these 30 strains, 21 P. pentosaceus strains were isolated from fruits and vegetables, while one was each derived from soil, raw milk, shrimp, or Homo sapiens. The sources of the remaining five strains have not been determined. In comparison to LAB such as Lactiplantibacillus plantarum and Streptococcus thermophilus, P. pentosaceus has substantial potential for expanding source diversity and genome database coverage. Distinct from the known 30 strains of P. pentosaceus, P. pentosaceus LL-07 was isolated from fermented meat (Libo, Guizhou, China), and has demonstrated the ability to produce exopolysaccharides (EPS), underscoring its potential as a probiotic contender. A comprehensive analysis of its traits and possible underlying issues is needed for future development and application. This study aimed to perform whole-genome sequencing of P. pentosaceus LL-07 to establish its taxonomy through genetic exploration and comparative genomic analysis and to evaluate its beneficial properties, EPS synthesis pathways, and safety factors. In addition, we experimentally evaluated several probiotic properties of P. pentosaceus LL-07, such as tolerance to artificial digestive juice and bile, autoaggregation, and antioxidation.
Materials and methods
Bacterial strain and culture conditions
P. pentosaceus LL-07 was isolated from fermented meat (Libo, Guizhou, China) and characterized as a bacterium capable of producing EPS. It was initially identified as P. pentosaceus through the 16S rRNA gene sequencing. P. pentosaceus ATCC25745 was used as a reference.
P. pentosaceus LL-07 was inoculated in Man Rogosa Sharpe broth (MRS, Shanghai Bio-way Technology Co., Ltd., China) liquid medium with a 2% (v/v) inoculum and incubated at 37 ℃ for 24 h under static conditions. After two to three activation cultures were performed, the culture was set aside.
Staphylococcus aureus ATCC6538 was used as a positive control strain for the hemolysis test and inoculated in Luria–Bertani (LB, Shanghai Sangwei Science and Technology Co., Ltd., Shanghai, China) medium at a concentration of 2% (v/v) inoculum and incubated at 37 °C for 12 h.
Additionally, another thirty complete genomes of P. pentosaceus, publicly available on the National Center for Biotechnology Information (NCBI) website (https://www.ncbi.nlm.nih.gov/) (released from 1980 to 2023), were used for comparison (Table 1).
Genome sequencing, assembly and annotation
Typical individual P. pentosaceus LL-07 colonies were selected, inoculated in 200 mL of MRS liquid medium, and cultured at 37 ℃ until reaching logarithmic growth. Subsequently, centrifugation was carried out at 16.000 × g for 10 min at 4 ℃, the supernatant was discarded, the bacterial cells were collected, and the medium components were rinsed away with sterile water. Genomic DNA extraction followed the protocols of the bacterial DNA extraction kit (magnetic beads) from Majorbio (Shanghai, China).
Whole-genome sequencing of P. pentosaceus LL-07 was conducted by Majorbio BioTech Co. genomic services (Shanghai, China) using PacBio Sequel IIe (Pacbio, America) and NovaSeq6000 (Illumina, America).
Coding sequences (CDS) in the genome were predicted using Glimmer [21] (http://ccb.jhu.edu/software/glimmer/index.shtml). The P. pentosaceus LL-07 genome map was predicted using CGView (http://stothard.afns.ualberta.ca/cgview_server/) [22]. tRNAs in the genome were predicted using tRNAscan-SE v2.0 [23] software (http://trna.ucsc.edu/software/). rRNAs contained in the genome were predicted using Barrnap software (https://github.com/tseemann/barrnap). Predictive annotation of possible sRNAs in the bacterial genome was performed using Infernal [24] software (http://eddylab.org/infernal/) and the Rfam database (https://rfam.xfam.org/).
Plasmid identification was performed using PlasmidFinder [25] (https://cge.cbs.dtu.dk/services/PlasmidFinder/) on the results of bacterial genome assembly by the plasmid sequences obtained, and then BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) software and PLSDB database (https://ccb-microbe.cs.uni-saarland.de/plsdb/) for plasmid annotation.
The Kyoto Encyclopedia of Genes and Genomes (KEGG, http://www.genome.jp/kegg) database was used for the annotation and classification of metabolic pathways [26], via the KEEG Automatic Annotation Server (KAAS, https://www.genome.jp/kegg/kaas/) [27].
The average nucleotide identity (ANI) values and phylogenetic analyses
The average nucleotide identity (ANI) is based on the BLASTN algorithm. The phylogenetic tree was constructed using the TYGS server (https://tygs.dsmz.de) for taxonomic analyses based on complete genomes [28]. Specifically, the P. pentosaceus LL-07 genome was compared with conserved genomes of the same pedigree type, and the phylogenetic tree was visualized using iTol v.6.8.1.
Comparative genomic analysis
All P. pentosaceus genomes were annotated with Prokaryotic Genome Annotation (Prokka) v.1.13 [29]. Subsequently, OrthoFinder v.2.5.4 [30] with default settings was used to identify orthogroups of orthologous gene pairs. Flip charts were generated using R v.4.3.2 and the UpsetR package [31]. The core orthogroups shared among P. pentosaceus strains were additionally annotated to Clusters Orthologous Groups (COG) categories [32] through the online server Batch CD-Search (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) [33].
Multiple genome alignments
The genomes of strains closely related to P. pentosaceus LL-07 were selected to enhance the analysis of the evolutionary relationships between species. The MCScanX [34] tool was used for overall protein-level alignment to examine the synteny among the genome sequences of each strain. Additionally, genome-wide visualization of coding sequence identity among 31 strains was performed using BLAST Ring Image Generator (BRIG) v.0.95 [35], with the P. pentosaceus ATCC25745 strain as the reference genome.
Analysis of carbohydrates utilization and related genes
To examine the carbohydrate metabolism of P. pentosaceus LL-07, an experiment was conducted following the instructions provided by API50 CH (BioMerieux Diagnostics (Shanghai) Co., Ltd., China).
The genome of P. pentosaceus LL-07 was annotated using HMMER v.3.3.2, and the carbohydrate-active enzymes were analyzed by the Carbohydrate Active Enzymes Database (CAZy) (http://www.cazy.org/) [36, 37]. A cut-off limit of an E-value of 1e − 15 and a coverage of 0.35 were used to identify the CAZyme class.
Prediction of the EPS Gene operon
The sequence of P. pentosaceus LL-07 was aligned with known EPS coding manipulators through the Basic Local Alignment Search Tool (BLAST) program [38,39,40] to identify the presence of the genes and map the EPS synthesis gene cluster.
Mobile genetic elements
The PHASTER (PHAge Search Tool Enhanced Release) was used to identify and annotate prophages within all the strains [41]. The presence of clustered palindromic interspaced palindromic repeats (CRISPR) regions and CRISPR-associated (Cas) proteins were searched with CRISPRCasFinder (https://crisprcas.i2bc.paris-saclay.fr/) [42]. The TransposonPSI software was used to predicte the transposon [43].
Safety assessment and identification of antibiotic resistance genes
Antimicrobial resistance genes (AMR) were studied using the Comprehensive Antibiotic Resistance Database (CARD) [44] and NCBI AMRFinder Plus [45] databases. Virulence factor genes were detected using the VFDB [46] database.
Artificial saliva, gastric juice and intestinal juice tolerance
Overnight cultures of P. pentosaceus LL-07 were centrifuged and washed twice with physiological saline. The cells were resuspended in artificial saliva (pH 6.8) (Shanghai Yuanye Bio-Technology Co., Ltd, China) to tolerate 0.5 h (for a final cell concentration of approximately 108 CFU/mL). The cells were collected by centrifugation and resuspended in artificial gastric juice (pH 3.0) (Shanghai Yuanye Bio-Technology Co., Ltd, China) for 2 h. Then, the cells were collected and resuscitated in artificial intestinal juice (pH 6.8) (Shanghai Yuanye Bio-Technology Co., Ltd, China) for 3 h. The culture conditions of each step were 37 ℃ with shaking (180 rpm) to simulate chewing and gastrointestinal conditions. The number of living cells was counted by the plate count method, and the survival rate of P. pentosaceus LL-07 was calculated.
Bile salt tolerance
The collected cells of P. pentosaceus LL-07 were resuspended to a final cell concentration of approximately 108 CFU/mL in physiological saline containing 0.2% bile salt (Sigma, America). After incubating for 3 h at 37 ℃, the bile tolerance was evaluated via the plate counting method.
Autoaggregation assay [47]
The cells of P. pentosaceus LL-07 were centrifuged and washed twice with PBS, and resuspended to an optical density of 0.5 at 600 nm (A0). The suspensions were incubated at 37 ℃ for 4 h or 24 h without shaking. The aqueous phase was gently taken out to measure its absorbance at 600 nm (A1). Autoaggregation percentage was calculated as (1 − A1/A0) × 100.
Hemolytic activity
The hemolytic activity of P. pentosaceus LL-07 was assessed with 5% (v/v) sheep red blood cells on trypsin agar (TSA). The strain was incubated at 37 ℃ for 24 h, followed by the measurement of α, β, or γ hemolysis [48]. Blank experimental and S. aureus ATCC6538 control groups were set up simultaneously.
Antibiotic resistance
An antibiotic resistance test was performed on P. pentosaceus LL-07 following the instructions of the antibiotic sensitivity test paper method. The antibiotic sensitivity was assessed based on the size of the inhibition zone. The antibiotic sensitivity test paper and corresponding judgment criteria are detailed in Table 2.
Antioxidant activity
The antioxidant activity was analyzed by measuring the DPPH free radical scavenging activity according to the methods of [49], with some modifications. The strains were cultured in MRS broth at 37 ℃ for 24 h and then centrifuged. The supernatant was mixed 1:1 (v/v) with ethanol DPPH solution (0.2 mmol/L) and incubated in the dark at 37 ℃ for 30 min. The absorbance (A) at 517 nm was measured (MultiskanGO, Thermo Fisher Scientific Inc., USA). The scavenging activity of DPPH was defined as scavenging activity (%) = [1 − (As-Ab)/Ac] × 100%, where As is the A at 517 nm of the sample, Ab is the A of anhydrous ethanol instead of DPPH, and Ac is the A of water instead of sample.
Statistical analysis
All the experiments were performed in triplicate, and the data are expressed as the means ± standard deviations that were analyzed using the Statistical Package for the Social Sciences (SPSS 19.0, IBM, Inc., Armonk, NY, USA).
The P. pentosaceus LL-07 genome sequence was submitted to the GenBank database, and the accession number PRJNA904022 was obtained.
Results
General characteristics of the P. pentosaceus LL-07 genome
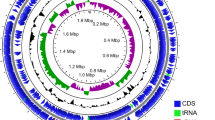
The genome size of P. pentosaceus LL-07 is 1,782,685 bp, consisting of a 1,764,987 bp circular chromosome and a 17,698 bp circular plasmid. The GC content of the chromosome and plasmid are 37.18% and 33.08%, respectively (Table 3). The basic information and genomic features are shown in Fig. 1. There are 1,739 predicted chromosomal genes in P. pentosaceus LL-07, along with three kinds of non-coding RNAs: tRNA, rRNA, and sRNA (Supplementary Tables 1, 2 and 3).

Chromosome (A) and plasmid (B) maps of P. pentosaceus LL-07. The circular illustration was visualized using the CGView server. In the cycle diagram, the first and fourth circles from the outside in represent CDS on the positive and negative strands, respectively, with different colors denoting different COG function classifications. The second and third circles represent CDS, tRNA, and rRNA on the positive and negative strands, respectively. The fifth circle indicates the GC content, and the sixth circle represents the GC-Skew value
Phylogenetic analysis
ANI analysis as a standard criterion for species classification and clustering. Previous research has shown that clear ANI divisions exist within and between species in most lineages. ANI values surpass 95% for the same species and fall below 95% for different species [50]. The results demonstrate that all 31 P. pentosaceus strains have ANI values exceeding 98%, with P. pentosaceus MGB0941 exhibiting the highest similarity to P. pentosaceus LL-07, with 99.02% ANI similarity. The phylogenetic tree (Fig. 2) also shows high similarity values among P. pentosaceus genomes. Additionally, ten strains were classified on the same branch as P. pentosaceus LL-07.

Phylogenetic analysis and ANI heatmap of P. pentosaceus. The proposed species cut-off boundary is above 98%, demonstrating identity within these strains
Comparative genomic analysis
Based on the phylogenetic tree results, ten bacterial strains were clustered with P. pentosaceus LL-07. A direct homologous gene analysis of these 11 strains included 19,324 genes, with 18,914 (97.88%) distributed into 2,053 orthogroups by OrthoFinder. Of these, 1,365 were identified as core orthogroups, and 688 were identified as accessory orthogroups (Fig. 3A). The orthogroups contained 1,675 (81.59%) genes of P. pentosaceus LL-07. The core orthogroups' additional details were classified into Clusters Orthologous Groups (COG) using the Batch CD-Search online server. Of the 1,365 core line types, 1,256 (92.01%) genes were categorized into the COG family (Fig. 3B), covering 21 functional classes.

Upset plot (A) and COG classification (B) of orthogroups among the eleven P. pentosaceus species. The box plot illustrates the distributions of the mean orthogroup protein lengths. The bars represent the number of orthogroups exhibiting specific conservation patterns, with the numbers provided above each bar. The differently colored chart displays the functional categories of the core orthogroups based on COG classification using the Batch CD-Search server
Multiple genome alignment
For a comprehensive visualization of the coding sequences between P. pentosaceus LL-07 and the other 30 strains, the genomes of the 30 strains were compared to the reference genome P. pentosaceus ATCC25745.. The solid section of the circle represents similarities between the genomes, while the blank areas indicate variability (Fig. 4A). The comparison result shows the presence of the open reading frame in the reference genome, which was absent in the remaining genomes, and could be attributed to differences in assembly quality [51].

Multiple genome alignment of P. pentosaceus strains. A Nucleotide alignments of eleven P. pentosaceus genomes, generated with the BRIG. Different colors represent different strains. B Multiple genome alignment of P. pentosaceus LL-07, P. pentosaceus SRCM100892, P. pentosaceus MGB0620 and P. pentosaceus MGB0941. Visualization of alignment is organized into one horizontal panel per genome sequence, with the label of the genome sequence name at the center of each panel. Homologous blocks within the genome are connected by lines
P. pentosaceus SRCM100892, P. pentosaceus MGB0620, and P. pentosaceus MGB0941 were selected for multi-genome alignment (Fig. 4B) due to their close relationships with P. pentosaceus LL-07 in the phylogenetic tree (Fig. 2). There was a relatively high sequence similarity between P. pentosaceus LL-07 and P. pentosaceus MGB0620, as well as P. pentosaceus MGB0941. The positions of similarity between P. pentosaceus LL-07 and P. pentosaceus SRCM100892 mainly involve inversions within the chromosome. Moreover, there are regions in P. pentosaceus LL-07 that do not align with P. pentosaceus SRCM100892, P. pentosaceus MGB0620, and P. pentosaceus MGB0941. This illustrates the divergence of one strain of the same species from others and provides substantial evidence for the reconstruction of P. pentosaceus LL-07.
Genotype analysis for carbohydrate utilization in P. pentosaceus LL-07
In the whole genome sequence of P. pentosaceus LL-07, 956 genes were functionally annotated by the KEGG database (54.97%), participating in six major classes of metabolic pathways (Fig. 5A). Among the metabolism-related genes, the most abundant were those associated with Global and Overview Maps (305 genes), followed by Carbohydrate Metabolism (118 genes), Nucleotide Metabolism (66 genes) and Amino Acid Metabolism (55 genes).

Genotype analysis for carbohydrate utilization in P. pentosaceus LL-07. A Kyoto Encyclopedia of Genes and Genomes (KEGG) functional annotation. The functional categories are represented as follows: Dark Blue-Cellular Processes; Purple-Environmental Information Processing; Green-Genetic Information Processing; Red-Human Diseases; Orange-Metabolism;Blue-Organismal Systems. B Distribution of CAZymes. Different colors represent different classes of CAZymes identified in the genome. The representation from the inner to outer rings are, CAZyme classes, CAZyme families, and the number of genes identified in each family, respectively. C Carbohydrate metabolism
Carbohydrate active enzymes (CAZymes) are classified based on the sequences of enzymes involved in complex carbohydrate metabolism. These enzymes were studied by comparing the predicted genes in the P. pentosaceus LL-07 genome with the CAZY database. There were forty-five genes were identified across four classes of CAZymes (Fig. 5B)..
P. pentosaceus LL-07 can utilize 25 out of 49 carbohydrates more effectively (Table 4). In addition to monosaccharides and disaccharides, which are typically degraded by Pediococcus, P. pentosaceus LL-07 has the ability to utilize lactose and galactose but not polysaccharides such as starch or glycogen. Genes related to carbohydrate metabolism were identified in the P. pentosaceus LL-07 metabolic pathway, as shown in Fig. 5C and Supplementary Table 4. The greatest number of genes (30) were involved in Amino Sugar and Nucleotide Sugar Metabolism, followed by genes in Glycolysis/Gluconeogenesis(26 genes), Pyruvate Metabolism (25 genes), Fructose and Mannose Metabolism (22 genes), Pentose Phosphate Pathway (22 genes), and Pentose-phosphate Metabolism (25 genes), Phosphate Pathway (19), Starch and Sucrose Metabolism (16), Galactose Metabolism (16), and Propanoate Metabolism (12).
Prediction of the EPS gene operon in P. pentosaceus LL-07
Amino and nucleotide sugars serve as precursors for EPS synthesis. In P. pentosaceus LL-07, aminosugars and nucleotide sugars are predominantly represented by uridine diphosphate sugars (UDP sugar). The synthesis pathways and related genes are depicted in Fig. 6A.

Prediction of the EPS Gene Operon in P. pentosaceus LL-07. A Amino sugar and nucleotide sugar metabolism in P. pentosaceus LL-07. The red box indicates potential exopolysaccharide synthesis monomers. B EPS synthesis pathway. Different colors represent different functions of genes
P. pentosaceus LL-07 contains a gene cluster consisting of 13 genes (12.77 kb) responsible for EPS synthesis (Fig. 6B). These genes included chain-length determination and export genes (gene0578, gene0579 and gene0580), genes involved in the biosynthesis of the repeating unit (gene0581, gene0584, gene0586 and gene0590), modification genes (gene0583 and gene0588), polymerization genes (gene0585 and gene0587), and export genes (gene0589).
Mobile genetic elements in P. pentosaceus LL-07
Prephages, commonly found in bacterial genomes, represent mild phage genomes integrated into the bacterial genome [52]. They often contain functional genes, such as antibiotic resistance and virulence genes, significantly to enhance bacterial competitiveness and the ability to withstand adverse environments [53]. Two potential prophages were identified in the P. pentosaceus LL-07 genome (Supplementary Table 5), with phage 2 being the largest at 29.62 Kb, encoding 37 genes. Phage2 includes genes related to phage formation and those associated with lysozyme M1, acetyl coenzyme A carboxylase, and amino acid permease. Notably, no pathogenesis-related genes were detected in either prophage 1 or prophage 2.
The CRISPR/Cas system functions as a defense mechanism in bacteria against foreign genetic material, such as phage viruses and foreign plasmids. [54, 55]. It recognizes exogenous DNA and silences the expression of exogenous genes. CRISPR usually consists of numerous short and conserved repeating sequence regions (repeat) and spacers. Cas, a double-stranded DNA nuclease, resides near CRISPR sites and cleaves the target site under the guidance of guide RNA. The CRISPR-Cas system is formed by clusters of CRISPRs along with Cas proteins. In the P. pentosaceus LL-07 genome, two dispersed CRISPRs and three dispersed Cas were identified, as detailed in Supplementary Table 6.
Transposons are DNA sequences in the genome that can move between chromosomes, plasmids, or phages in bacteria. The genome of P. pentosaceus LL-07 was analyzed, revealing the presence of three transposons (Supplementary Table 7). These transposons belong to the LINE family (Tn01, Tn03) and the helitronORF family (Tn02). Tn01, Tn02, and Tn03 encode a ribonuclease H family protein, a primosomal protein N', and an NAD-dependent DNA ligase LigA, respectively. These genes are involved in DNA replication and transcription.
Safety assessment and identification of antibiotic resistance genes
There were 163 virulence factor-related genes in P. pentosaceus LL-07 (Supplementary Table 8). In comparison, the predicted virulence factors are primarily associated with antiphagocytosis (28), the iron uptake system (24), adherence (17), regulation (13), and toxins (13). Notably, the 13 toxin-related virulence genes predominantly function in self-protection and resistance, with no identified gene capable of toxin production. Prediction of resistance genes identified eighty-six genes associated with resistance to twenty-five antibiotics (Supplementary Table 9).
Assessment of probiotic properties of P. pentosaceus LL-07
The probiotics of P. pentosaceus LL-07 were evaluated, including tolerance of the simulated digestion process, autoaggregation assay, hemolytic activity, antibiotic resistance, and antioxidant activity. It was observed that P. pentosaceus LL-07 maintained better viability than P. pentosaceus ATCC25745 in artificial saliva (p < 0.05). Both P. pentosaceus LL-07 and P. pentosaceus ATCC25745 showed tolerance to artificial gastric juice, intestinal juice, and bile salt (Table 5). At 4 h, P. pentosaceus LL-07 exhibited higher autoaggregation than P. pentosaceus ATCC25745 (p < 0.05) (Table 5). There was no significant difference in autoaggregation between the two strains after 24 h (p > 0.05). The results indicate that both strains possess DPPH radical scavenging ability, with P. pentosaceus LL-07 exhibiting a higher ability than P. pentosaceus ATCC25745 (p > 0.05). This difference in scavenging ability may be attributed to the differences in the structure and composition of metabolites secreted by the two strains.
The hemolysis experiment demonstrates that P. pentosaceus LL-07 forms milky-white colonies without a hemolytic circle after 48 h of incubation on blood plates, indicating γ-hemolysis without virulence (Fig. 7A). Antibiotic resistance experiments revealed differences in P. pentosaceus LL-07 resistance to different antibiotics. Notably, within the same antibiotic class, diverse resistance patterns were observed (Fig. 7B).

Safety assessment of P. pentosaceus LL-07. A Hemolysis. B Antibiotic resistance.The above picture shows the results of antibiotic resistance experiment in each group. The bottom square column shows the diameter of antibacterial zone for each group. The letters above the bar indicate resistance to antibiotics. Where R stands for resistant, I for moderately susceptible, and S for susceptible
Discussion
This study involved subjecting P. pentosaceus LL-07 to whole genome sequencing, genome characterization, and comparative analysis. The mean G + C content of P. pentosaceus LL-07 is 37.18%, which is a standard feature of bacterial taxonomy and reflects evolutionary relatedness. This value aligns with Bergey’s Manual of Systematics of Archaea and Bacteria [56]. The bacterial genome size was 1.78 Mb with 1,739 CDS. Additionally, the bacterium contained a circular plasmid of 17 kb..
ANI has been served as the gold standard for genetic restriction in prokaryotic species [57]. In the ANI analysis, P. pentosaceus LL-07 was compared to 30 strains of P. pentosaceus, and classified to the same branch with ten strains. Notably, the ANI values of P. pentosaceus MGB0620, P. pentosaceus MGB0941, and P. pentosaceus SRCM100892 were relatively narrow with P. pentosaceus LL-07, indicating limited genetic diversity. The results suggest that P. pentosaceus LL-07 may not require an extensive set of genes to adapt to diverse ecological niches. Phylogenetic similarities between the P. pentosaceus LL-07 genome and 30 P. pentosaceus strains from NCBI were revealed through ANI and genome-wide analyses. The genome of P. pentosaceus contains 1,365 directly homologous genes, which are primarily involved in maintenance functions crucial for the growth and survival of the species. Multiple genome analyses revealed that strains resembling those in the phylogenetic tree exhibited similar levels of gene deletion..
The carbohydrate utilization capacity of bacteria is a crucial indicator of strain functionality, forming the basis for strain culture and selection. P. pentosaceus LL-07 exhibits superior carbohydrate utilization, possibly due to the presence of numerous genes encoding potential sugar transporters. Typically, the genus P. pentosaceus cannot ferment lactose [58, 59]. Sugar fermentation experiments and KEGG pathway analysis revealed a complete lactose metabolic pathway in P. pentosaceus LL-07, which can better utilize lactose and galactose. Upon entering the bacterium, lactose can be broken down into galactose (D-Gal) and glucose by the β-galactosidase LacZ (gene0193, gene0194). Under the catalysis of galactose changing enzymes GalM (gene0191, gene0210, gene1010), galactose kinases GalK (gene0116, gene0195), and glucose hexose-1-phosphate uridine transferase GalT (gene0197), galactose forms uridine diphosphate galactose (UDP Gal). UDP-Gal can be converted into two different compounds: uridine diphosphate-furanogalactose (UDP-Galf) and uridine glucose diphosphate (UDP-Glc). UDP-Galf is produced by the uridine diphosphate-furanogalactose metathesis enzyme Glf (gene0588) and is used for the biosynthesis of N-glycans. UDP-Glc, on the other hand, is formed by the catalytic action of uridine diphosphate-glucose 4-differential isomerase GalE (gene0196, gene1424) and enters a subsequent metabolic pathway. P. pentosaceus LL-07 is a natural strain capable of fermenting lactose, which contradicts the previous belief that lactose-positive strains do not exist in naturally occurring Pediococcus. This conclusion is supported by the study of Jiang [51] and overturns the notion that lactose-positive strains are absent in natural Pediococcus [59].
CAZyme analysis revealed that P. pentosaceus LL-07 predominantly contains GTs, GHs, CEs, and AAs, which are crucial for glucose metabolism. GTs are enzymes that have substrate specificity and catalyze the transfer of sugar groups from activated nucleotide sugar donors to lipids, polysaccharides, or protein acceptors. In P. pentosaceus LL-07, GTs are classified into seven different families. Enzymes from GT2 and GT4 families make up 72.22% of the total GTs. These enzymes convert glucose, mannose, xylose, galactose, trehalose, arabinose, etc., into metabolites for metabolic utilization. GHs represent key enzymes for carbohydrate metabolism and play a pivotal role in the hydrolysis of carbohydrate glycosidic bonds. P. pentosaceus LL-07 contains a total of 12 different GH families that act on glycosidic bonds formed by glucose, mannose, xylose, galactose, trehalose, arabinose, etc. This diversity proves advantageous for the strain's growth in various environments, making it versatile and critical for development [60, 61]. GTs and GHs are closely related to EPS precursor synthesis. The analysis showed that P. pentosaceus LL-07 has numerous GHs and GTs genes associated with mannose metabolism, indicating that mannose may be a significant component of the synthesized EPS. Additionally, the GHs include two lysozymes (GH25 or GH73), which enhance the bacteria's ability to adapt and resist external challenges [62, 63]. These findings align with the study conducted by Oliveira et al. [64].
In addition to bacteriocins and bacteriocin-like inhibitory substances (BLIS), EPS is an important metabolite secreted by P. pentosaceus with specific functions. P. pentosaceus DPS has been reported to synthesize heat-resistant EPS in the human gastrointestinal tract. This EPS destructs biofilms originating from harmful pathogens by inhibiting adhesion, thereby combating Listeria monocytogenes and Escherichia coli [65]. P. pentosaceus M41, sourced from marine environments, secretes EPS-M41 exhibiting antitumor activity, demonstrating high efficacy against Caco-2 and MCF-7 cells [66]. Moreover, certain strains of P. pentosaceus, such as P. pentosaceus Be1 (isolated from fermented foods) [67] and P. pentosaceus S-SU6 (isolated from the intestines of blue mackerel) [68], specialize in scavenging hydrogen peroxide. Due to the secretion of acidic EPS, P. pentosaceus MYU 759 demonstrates hydroxyl radical antioxidant capacity (HORAC), proving beneficial for its isolation from soy milk yogurt as an antioxidant product [69].
The prediction of the EPS synthesis gene cluster in P. pentosaceus LL-07 using antiSMASH did not yield a matching gene cluster model. However, upon relaxing the detection requirements, genes related to EPS synthesis were identified between chromosomes 596,709 and 609,483. Moreover, by utilizing Orthomcl software and BlastN to predict the gene operons of EPS, it was determined that the 13 ORFs in the EPS cluster of P. pentosaceus LL-07 are related to the EPS biosynthesis process. It can be inferred that the genes on the P. pentosaceus LL-07 EPS synthesis gene cluster exhibit specific structural differences from known gene clusters but encompass essential elements of EPS synthesis, including regulatory genes, chain length-determining genes, repeat unit biosynthesis genes, and aggregation output genes.
The predicted products of gene0578, gene0579, and gene0580 are homologous to Wzz, the CpsD/CapB family tyrosine protein kinase, and the CpsB/CapC family capsule biosynthesis tyrosine phosphatase, respectively. The literature reports indicate that Wzz is one of the factors influencing the heterogeneity of the E. coli O antigen chain length [70]. The CpsD/CapB family tyrosine protein kinase is a critical regulatory device in bacterial physiology and acts as a domain protein involved in EPS biosynthesis [71]. The CpsB/CapC family of capsule biosynthetic tyrosine phosphatases belongs to the protein tyrosine phosphatase (PTP) superfamily and can synergistically interact with protein tyrosine kinases. In E. coli K-30,the tyrosine kinase Wzc and its homologous phosphatase Wzb are critical factors in the synthesis and assembly of capsule polysaccharides. In the gram-positive bacterium Streptococcus pneumoniae, the CpsCD complex is similar to Wzc, while the phosphatase CpsB is the corresponding homologous phosphatase [72]. In S. pneumoniae, CpsB regulates the biosynthesis of capsule polysaccharides through tyrosine dephosphorylation of its homologous tyrosine kinase CpsD [73]. The predicted products of gene0581 and gene0586 are homologous to sugar transfer. These enzymes can catalyze the transfer of sugars from donors (such as UDP-glucose or UDP-galactose) to lipid carriers (such as undecyl phosphate) [74] and sugar transferases similar to the E. coli beta-1,6-galactofuranosyltransferase WbbI, which is involved in transferring galactofuranose (Galf) onto an alpha-D-gluco-configured acceptor substrate to form a beta-1,6-linkage [75]. The biosynthesis of EPS repeat units also requires multiple glycosyltransferases (GTs), whose coding genes typically reside in the middle of EPS gene clusters [76]. Gene0584 and gene0590 were identified as GT genes. Although predicted functions of the GT genes in P. pentosaceus LL-07 are the same (glycosyltransferase), their nucleotide sequences exhibit some variation. Therefore, the activity of these glycosyltransferases may vary [77], offering potential avenues for enhancing EPS production through genetic modification. Gene0585 has been identified as a Stealth CR1 domain-containing protein, and research indicates that this protein may be a D-hexose-1-physiological transfer involved in EPS biosynthesis [78]. The predicted product of the precursor gene (gene0588) is associated with UDP-galactopyranose mutase. This enzyme is involved in converting UDP-GALP into UDP-GALF through a 2-keto intermediate. UDP-galactopyranose mutase regulates the biosynthesis of precursors in lipopolysaccharide production in Klebsiella pneumoniae [79]. Gene0587 and gene 0589 are associated with polysaccharide polymerase and polysaccharide biosynthesis C-terminal domain-containing proteins associated with polysaccharide polymerization and export [80, 81].
Mobile genetic elements, such as prephages and transposons, are frequently present in microorganisms. They can serve as carriers of pathogenic genes or facilitate horizontal gene transfer (HGT) from antimicrobial-resistant (AMR) microorganisms to others [82]. Therefore, it is necessary to conduct further analysis of these regions to identify any undiscovered pathogenic or AMR genes in the genome of P. pentosaceus LL-07. The genome contains two potential prophages and three transposons. However, no pathogenicity-related genes were detected. Phage 2 includes a complete prophage region and genes related to lysozyme M1, acetyl-CoA carboxylase, and amino acid permease. The transposon genes are mainly related to DNA replication and transcription.
The CRISPR-Cas system serves as an adaptive immune role against invasive elements, such as phages in bacterial cells, and regulates gene activity, DNA repair, and genome recombination [55, 83]. Despite the presence of two isolated CRISPRs and three isolated clusters of Cas3 in its genome, the predicted results suggest that the CRISPR-Cas system may not be present in the genome of P. pentosaceus LL-07. According to Zhang and Ye [84], CRISPR regions without cas genes may not be functional or work with distant cas motifs in the same genome.
During proliferation, certain bacteria can produce hemolysin, which specifically binds to red blood cells, causing severe hemolytic reactions and resulting in conditions such as anemia. Hemolysis test results indicated that in comparison to the S. aureus ATCC6538 control group, P. pentosaceus LL-07 exhibited general growth on the blood plate, without the formation of a hemolytic circle, indicating nontoxic γ-hemolysis. Accordingly, in P. pentosaceus LL-07, potential virulence factor genes were identified using the Virulence Factor Database (VFDB), revealing the presence of 163 putative virulence factor genes. Most of these genes exhibited less than 70% similarity in the VFDB, indicating low resemblance to genes in the database. Additionally, a search and Blast comparison revealed the absence of genes encoding toxins, such as hemolysin, in P. pentosaceus LL-07, consistent with the experimental findings.
AMR analysis revealed 86 antibiotic resistance (AR) genes in the P. pentosaceus LL-07 genome, corresponding to 25 antibiotic. The results of the antibiotic resistance experiment were generally consistent with the genetic analysis. Aminoglycoside and fluoroquinolone antibiotic genes were present in the P. pentosaceus LL-07 genome, resulting in resistance to gentamicin, kanamycin, and ofloxacin. Notably, despite the presence of macrolide and tetracycline antibiotic genes in the P. pentosaceus LL-07 genome, the strain exhibited moderate sensitivity to erythromycin and doxycycline, and no sensitivity to azithromycin and tetracycline. No genes related to penicillin antibiotics were identified, indicating sensitivity and moderate sensitivity to ampicillin and penicillin, respectively. This variation could be due to multiple factors, such as gene expression levels and substrate specificity of the expression products [85].
Further analysis of the P. pentosaceus LL-07 genome revealed that theresistance mechanism of this strain predominantly involves antibiotic efflux, antibiotic target alteration, and antibiotic target protection. These systems may contribute to bacterial resistance to these antibiotics. The main concern about genes that confer AR in LAB is their potential to be transferred to other bacteria, mainly pathogens, which could lead to complications and reduce the effectiveness of antibiotic treatment. No AR genes were identified by analyzing the prophage region, transposons and plasmid genes in the P. pentosaceus LL-07 genome, indicating a low risk of transfer to other bacteria.
P. Pentosaceus LL-07 exhibits bile salt and digestive juice tolerance, adhesion, and antioxidation. These characteristics are supported by 1 Choloylglycine hydrolase family protein gene, eight F0F1 ATP synthase genes, nine adhesion-related genes, and 31 antioxidant-related genes (Supplementary Table 10). Additionally, P. pentosaceus LL-07 contains a gene that encodes bifunctional folate polyglutamate synthase/dihydrofolate synthetase, which is associated with folate synthesis. The gene can code bifunctional folate polyglutamate synthase/dihydrofolate synthetase, which can catalyze the synthesis of dihydrofolate and folylpolyglutamate. Dihydrofolate is a precursor to folate, and folylpolyglutamate is the primary form of folate in the body.
Conclusion
In this study, the probiotic potential of the meat-derived P. pentosaceus LL-07 was studied via whole genome analysis combined with experimental studies. In this study, we determined the accurate taxonomic location, carbohydrate utilization capacity, safety and probiotic characteristics of P. pentosaceus LL-07. The whole genome sequence of P. pentosaceus LL-07 provides potential for new research to determine the genetic factors and mechanisms related to its beneficial role as a probiotic. The in vitro experimental results of the probiotic characteristics revealed its high tolerance to saliva, gastric juice, intestinal juice and bile, auto-aggregation, safety, antibiotic resistance and antioxidant activity. Compared with the reference strain, P. pentosaceus LL-07 has better saliva and bile tolerance, faster autoaggregation and greater antioxidant activity. In conclusion, genomic analysis and in vitro experiments revealed that P. pentosaceus LL-07 is a strong candidate for probiotics. Future research will focus on the relationship between EPS and probiotic activity to provide a deeper understanding of its probiotic mechanism, promoting the future development of probiotic products.
Availability of data and materials
The genome sequence was submitted to the GenBank database, and the accession number PRJNA904022 was obtained. All other data generated are included in the supplemental material files.
References
Lu Y, Yuan X, Wang M, He Z, Li H, Wang J, et al. Gut microbiota influence immunotherapy responses: mechanisms and therapeutic strategies. J Hematol Oncol. 2022;15(1):47. https://doi.org/10.1186/s13045-022-01273-9.
Byakika S, Mukisa IM, Byaruhanga YB, et al. A Review of Criteria and Methods for Evaluating the Probiotic Potential of Microorganisms. Food Rev Intl. 2019;35(5):427–66. https://doi.org/10.1080/87559129.2019.1584815.
Xu X, Bi S, Lao T, Chen F, Liao X, Wu J. Comprehensive investigation on volatile and non-volatile metabolites in broccoli juices fermented by animal- and plant-derived Pediococcus pentosaceus. Food Chem. 2020;341:128118. https://doi.org/10.1016/j.foodchem.2020.128118.
Song G, He Z, Wang XY, Zhao M, Cao X, Lin X, et al. Improving the quality of Suancai by inoculating with Lactobacillus plantarum and Pediococcus pentosaceus. Food Res Int. 2021;148:110581. https://doi.org/10.1016/j.foodres.2021.110581.
Cavicchioli VQ, Camargo AC, Todorov SD, Nero LA. Potential Control of Listeria monocytogenes by Bacteriocinogenic Enterococcus hirae ST57ACC and Pediococcus pentosaceus ST65ACC Strains Isolated From Artisanal Cheese. Probiotics Antimicrob Proteins. 2019;11(2):696–704. https://doi.org/10.1007/s12602-018-9449-0.
Pato U, Yusuf Y, Fitriani S, Tartila, Yeni R, Fadillah F, et al. Optimization of the Growth of Pediococcus pentosaceus Strain 2397 in Inhibiting Pathogenic Listeria monocytogenes IOP Conference Series. Earth Environ Sci. 2021;757(1):012056. https://doi.org/10.1088/1755-1315/757/1/012056. (8pp).
Huang J, Li S, Wang Q, Guan X, Qian L, Li J, et al. Pediococcus pentosaceus B49 from human colostrum ameliorates constipation in mice. Food Funct. 2020;11:5607–20. https://doi.org/10.1039/d0fo00208a.
Todorov SD, Wachsman M, Tomé E, Vaz-Velho M, Ivanova IV. Plasmid-Associated Bacteriocin Produced by Pediococcus pentosaceus Isolated from Smoked Salmon: Partial Characterization and Some Aspects of his Mode of Action. Probiotics Antimicrob Proteins. 2023. https://doi.org/10.1007/s12602-023-10059-5.
Jiang S, Cai L, Lv L, Li L. Pediococcus pentosaceus, a future additive or probiotic candidate. Microb Cell Fact. 2021;20(1):45. https://doi.org/10.1186/s12934-021-01537-y.
Zaki RM, Ramasamy K, Alwi NAA, Yusoff RM, Lim SM. Pediococcus pentosaceus LAB6- and Lactiplantibacillus plantarum LAB12-Derived Cell Free Supernatant Inhibited RhoA Activation and Reduced Amyloid-B In Vitro. Probiotics Antimicrob Proteins. 2022. https://doi.org/10.1007/s12602-022-10009-7.
Bhukya KK, Bhukya B. Exploration of Antidiabetic, Cholesterol-Lowering, and Anticancer Upshot of Probiotic Bacterium Pediococcus pentosaceus OBK05 Strain of Buttermilk. Probiotics Antimicrob Proteins. 2023;15(6):1484–500. https://doi.org/10.1007/s12602-022-10002-0.
Avci GA, Cagatay G, Cilak GO, Avci E. Probable novel probiotics: Eps production, cholesterol removal and glycocholate deconjugation of Lactobacillus plantarum ga06 and ga11 isolated from local handmade- cheese. J Microbiol Biotech Food Sci. 2020;10(1):83–6. https://doi.org/10.15414/jmbfs.2020.10.1.83-86.
Zhao X, Liang Q. EPS-Producing Lactobacillus plantarum MC5 as a Compound Starter Improves Rheology, Texture, and Antioxidant Activity of Yogurt during Storage. Foods. 2022;11(11):1660. https://doi.org/10.3390/foods11111660.
Yin H, Ye P, Lei Q, Cheng Y, Yu H, Du J, et al. In vitro probiotic properties of Pediococcus pentosaceus L1 and its effects on enterotoxigenic Escherichia coli-induced inflammatory responses in porcine intestinal epithelial cells. Microb Pathog. 2020;144:104163. https://doi.org/10.1016/j.micpath.2020.104163.
Adnan M, Siddiqui AJ, Hamadou WS, Ashraf SA, Hassan MI, Snoussi M, et al. Functional and Structural Characterization of Pediococcus pentosaceus-Derived Biosurfactant and Its Biomedical Potential against Bacterial Adhesion, Quorum Sensing, and Biofilm Formation. Antibiotics (Basel). 2021;10(11):1371. https://doi.org/10.3390/antibiotics10111371.
Jiang G, He J, Gan L, Li X, Xu Z, Yang L, et al. Exopolysaccharide Produced by Pediococcus pentosaceus E8: Structure, Bio-Activities, and Its Potential Application. Front Microbiol. 2022;13:923522. https://doi.org/10.3389/fmicb.2022.923522.
EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP), Rychen G, Aquilna G, Azimonti G, Bampidis V, Bastos ML, et al. Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA J. 2018;16(3):e05206. https://doi.org/10.2903/j.efsa.2018.5206.
Wu C, Huang J, Zhou R. Genomics of lactic acid bacteria: current status and potential applications. Crit Rev Microbiol. 2017;43:393–404. https://doi.org/10.1080/1040841X.2016.1179623.
Coton M, Lebreton M, Salas ML, Garnier L, Navarri M, et al. Biogenic amine and antibiotic resistance profiles determined for lactic acid bacteria and a Propionibacterium prior to use as antifungal bioprotective cultures. Int Dairy J. 2018;85:21–6. https://doi.org/10.1016/j.idairyj.2018.05.001.
Cavicchioli VQ, Camargo AC, Todorov SD, Nero LA, et al. Novel bacteriocinogenic Enterococcus hirae and Pediococcus pentosaceus strains with antilisterial activity isolated from Brazilian artisanal cheese. J Dairy Sci. 2017;100:2526–35. https://doi.org/10.3168/jds.2016-12049.
Li Y, Lei L, Zheng L, Xiao X, Tang H, Luo C. Genome sequencing of gut symbiotic Bacillus velezensis LC1 for bioethanol production from bamboo shoots. Biotechnol Biofuels. 2020;13:34. https://doi.org/10.1186/s13068-020-1671-9.
Grant JR, Stothard P. The CGView Server: a comparative genomics tool for circular genomes. Nucleic Acids Res. 2008;36:W181–4. https://doi.org/10.1093/nar/gkn179.
Chan PP, Lowe TM. tRNAscan-SE: Searching for tRNA Genes in Genomic Sequences. Methods Mol Biol. 2019;1962:1–14. https://doi.org/10.1007/978-1-4939-9173-0_1.
Nawrocki EP, Eddy SR. Infernal 1.1:100-fold faster RNA homology searches. Bioinformatics. 2013;29(22):2933–5. https://doi.org/10.1093/bioinformatics/btt509.
Carattoli A, Hasman H. PlasmidFinder and In SilicopMLST: identification and typing of plasmid replicons in wholegenome sequencing (WGS). Methods Mol Biol. 2020;2075:285–94. https://doi.org/10.1007/978-1-4939-9877-7_20.
Kanehisa M, Sato Y, Morishima K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J Mol Biol. 2016;428(4):726–31. https://doi.org/10.1016/j.jmb.2015.11.006.
Moriya Y, Itoh M, Okuda S, Yoshizawa AC, Kanehisa M. KAAS: an automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007;35:W182–5. https://doi.org/10.1093/nar/gkm321.
Meier-Kolthoff JP, Göker M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat Commun. 2019;10(1):2182. https://doi.org/10.1038/s41467-019-10210-3.
Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30(14):2068–9. https://doi.org/10.1093/bioinformatics/btu153.
Emms DM, Kelly S. OrthoFinder: solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015;16(1):157. https://doi.org/10.1186/s13059-015-0721-2.
Conway JR, Lex A, Gehlenborg N. UpSetR: an R package for the visualization of intersecting sets and their properties. Bioinformatics. 2017;33(18):2938–40. https://doi.org/10.1093/bioinformatics/btx364.
Tatusov RL, Galperin MY, Natale DA, Koonin EV. The COG database: a tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000;28(1):33–6. https://doi.org/10.1093/nar/28.1.33.
Marchler-Bauer A, Bryant SH. CD-Search: protein domain annotations on the fy. Nucleic Acids Res. 2004;32:W327–31. https://doi.org/10.1093/nar/gkh454.
Wang Y, Tang H, Debarry JD, Tan X, Li J, Wang X, et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012;40(7):e49. https://doi.org/10.1093/nar/gkr1293.
Alikhan NF, Petty NK, Zakour NLB, Beatson S. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics. 2011;12:402. https://doi.org/10.1186/1471-2164-12-402.
Besemer J, Lomsadze A, Borodovsky M. GeneMarkS A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001;29(12):2607–18. https://doi.org/10.1093/nar/29.12.2607.
Lombard V, Ramulu HG, Drula E, Coutinho PM, Henrissat B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014;42:D490–5. https://doi.org/10.1093/nar/gkt1178.
Yasutake T, Kumagai T, Inoue A, Kobayashi K, Noda M, Orikawa A, et al. Characterization of the LP28 strain-specific exopolysaccharide biosynthetic gene cluster found in the whole circular genome of Pediococcus pentosaceus. Biochemistry and Biophysics Reports. 2016;5:266–71. https://doi.org/10.1016/j.bbrep.2016.01.004.
Petrova MI, Macklaim JM, Wuyts S, Verhoeven T, Vanderleyden J, Gloor GB, et al. Comparative genomic and phenotypic analysis of the vaginal probiotic Lactobacillus rhamnosus GR-1. Front Microbiol. 2018;9:1278. https://doi.org/10.3389/fmicb.2018.01278.
Xiao L, Yang Y, Han S, Rui X, Ma K, Zhang C, et al. Effects of genes required for exopolysaccharides biosynthesis in Lacticaseibacillus paracasei S-NB on cell surface characteristics and probiotic properties. Int J Biol Macromol. 2023;224:292–305. https://doi.org/10.1016/j.ijbiomac.2022.10.124.
Arndt D, Grant JR, Marcu A, Sajed T, Pon A, Liang Y, et al. PHASTER: a better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016;44(W1):W16–21. https://doi.org/10.1093/nar/gkw387.
Couvin D, Bernheim A, Tofano-Nioche C, Touchon M, Michalik J, Neron B, et al. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018;46(W1):W246–51. https://doi.org/10.1093/nar/gky425.
Jiang BW, Ji X, Lyu Z, Liang B, Li J, Zhu L, et al. Detection of Two Copies of a blaNDM-1-Encoding Plasmid in Escherichia coli Isolates from a Pediatric Patient with Diarrhea. Infect Drug Resist. 2022;15:223–32. https://doi.org/10.2147/IDR.S346111.
Alcock BP, Raphenya AR, Lau T, Tsang KK, Bouchard M, Edalatmand A, et al. CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020;48(D1):D517–25. https://doi.org/10.1093/nar/gkz935.
Feldgarden M, Brover V, Haft DH, Prasad AB, Slotta DJ, Tolstoy I, et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob Agents Chemother. 2019;63(11):e00483-e519. https://doi.org/10.1128/AAC.00483-19.
Liu B, Zheng D, Jin Q, Chen L, Yang J. VFDB 2019: a comparative pathogenomic platform with an interactive web interface. Nucleic Acids. 2019;47(D1):D687–92. https://doi.org/10.1093/nar/gky1080.
Kos B, Suskovi’c J, Vukovi’c S, Simpraga M, Frece J, Matosi’c S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J Appl Microbiol. 2003;94:981–7. https://doi.org/10.1046/j.1365-2672.2003.01915.x.
Borges S, Barbosa J, Silva J, Teixerira P. Evaluation of characteristics of Pediococcus spp to be used as a vaginal probiotic. J Appl Microbiol. 2013;115(2):527–38. https://doi.org/10.1111/jam.12232.
Tsai CE, Lin LH. DPPH scavenging capacity of extracts from Camellia seed dregs using polyol compounds as solvents. Heliyon. 2019;5(8):e02315. https://doi.org/10.1016/j.heliyon.2019.e02315.
Jain C, Rodriguez-R LM, Phillippy AM, Konstantinidis KT, Aluru S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat Commun. 2018;9(1):5114. https://doi.org/10.1038/s41467-018-07641-9.
Jiang J, Yang B, Ross RP, Stanton C, Zhao J, Zhang H, et al. Comparative genomics of Pediococcus pentosaceus isolated from diferent niches reveals genetic diversity in carbohydrate metabolism and immune system. Front Microbiol. 2020;11:253. https://doi.org/10.3389/fmicb.2020.00253.
Correa AMS, Howard-Varona C, Coy SR, Buchan A, Sullivan MB, Weitz JS. Revisiting the rules of life for viruses of microorganisms. Nat Rev Microbiol. 2021;19(8):501–13. https://doi.org/10.1038/s41579-021-00530-x.
Obeng N, Pratama AA, Elsas JD. The significance of mutualistic phages for bacterial ecology and evolution. Trends Microbiol. 2016;24(6):440–9. https://doi.org/10.1016/j.tim.2015.12.009.
Westra ER, Buckling A, Fineran PC. CRISPR-Cas systems: beyond adaptive immunity. Nat Rev Microbiol. 2014;12(5):317–26. https://doi.org/10.1038/nrmicro3241.
Lander ES. The Heroes of CRISPR. Cell. 2016;164(1–2):18–28. https://doi.org/10.1016/j.cell.2015.12.041.
Whitman WB, Rainey F, Kämpfer P, et al. Bergey’s Manual of Systematics of Archaea and Bacteria. NJ, USA: Wiley; 2016.
Richter M, Rosselló-Móra R. Shifting the genomic gold standard for the prokaryotic species definition. Proc Natl Acad Sci USA. 2009;106(45):19126–31. https://doi.org/10.1073/pnas.0906412106.
Porto MCW, Kuniyoshi TM, Azevedo POS, Vitolo M, Oliveira RPS. Pediococcus spp.: An important genus of lactic acid bacteria and pediocin producers. Biotechnol Adv. 2017;35(3):361–74. https://doi.org/10.1016/j.biotechadv.2017.03.004.
Semjonovs P, Zikmanis P. Evaluation of novel lactose-positive and exopolysaccharide-producing strain of Pediococcus pentosaceus for fermented foods. Eur Food Res Technol. 2008;227:851–6. https://doi.org/10.1007/s00217-007-0796-4.
Vera C, Guerrero C, Aburto C, Cordova A, Illanes A. Conventional and non-conventional applications of β-galactosidases. Biochim Biophys Acta Proteins Proteom. 2020;1868(1):140271. https://doi.org/10.1016/j.bbapap.2019.140271.
Michlmayr H, Kneifel W. β-Glucosidase activities of lactic acid bacteria: mechanisms, impact on fermented food and human health. FEMS Microbiol Lett. 2014;352(1):1–10. https://doi.org/10.1111/1574-6968.12348.
Katharina E, Priyamedha S, Samuel K, Kemen E, Doehlemann G. A fungal member of the Arabidopsis thaliana phyllosphere antagonizes Albugo laibachii via a GH25 lysozyme. Elife. 2021;10:e65306. https://doi.org/10.7554/eLife.65306.
Hashimoto W, Ochiai A, Momma K, Itoh K, Mikami B, Maruyama Y, et al. Crystal structure of the glycosidase family 73 peptidoglycan hydrolase FlgJ. Biochem Biophys Res Commun. 2009;381(1):16–21. https://doi.org/10.1016/j.bbrc.2009.01.186.
Oliveira FS, Rodrigues RS, Carvalho AF, Nero LA. Genomic Analyses of Pediococcus pentosaceus ST65ACC, a Bacteriocinogenic Strain Isolated from Artisanal Raw-Milk Cheese. Probiotics Antimicrob Proteins. 2023;15(3):630–45. https://doi.org/10.1007/s12602-021-09894-1.
Abid Y, Casillo A, Gharsallah H, Joulak I, Lanzetta R, Corsaro MM, et al. Production and structural characterization of exopolysaccharides from newly isolated probiotic lactic acid bacteria. Int J Biol Macromol. 2018;108:719–28. https://doi.org/10.1016/j.ijbiomac.2017.10.155.
Ayyash M, Abu-Jdayil B, Olaimat A, Esposito G, Itsaranuwat P, Osaili T, et al. Physicochemical, bioactive and rheological properties of an exopolysaccharide produced by a probiotic Pediococcus pentosaceus M41. Carbohydr Polym. 2020;229:115462. https://doi.org/10.1016/j.carbpol.2019.115462.
Watanabe A, Kaneko C, Hamada Y, Takeda K, Kimata S, Matsumoto T, et al. Isolation of lactic acid bacteria exhibiting high scavenging activity for environmental hydrogen peroxide from fermented foods and its two scavenging enzymes for hydrogen peroxide. J Gen Appl Microbiol. 2016;62(2):75–82. https://doi.org/10.2323/jgam.62.75.
Kuda T, Kawahara M, Nemoto M, Takahashi H, Kimura B. In vitro antioxidant and anti-inflammation properties of lactic acid bacteria isolated from fish intestines and fermented fish from the Sanriku Satoumi region in Japan. Food Res Int. 2014;64:248–55. https://doi.org/10.1016/j.foodres.2014.06.028.
Yamamoto N, Shoji M, Hoshigami H, Watanabe K, Watanabe K, Takatsuzu T, et al. Antioxidant capacity of soymilk yogurt and exopolysaccharides produced by lactic acid bacteria. Biosci Microbiota Food Health. 2019;38(3):97–104. https://doi.org/10.12938/bmfh.18-017.
Franco AV, Liu D, Reeves PR. The Wzz (Cld) protein in Escherichia coli: amino acid sequence variation determines O-Antigen chain length specificity. J Bacteriol. 1998;180(10):2670–5. https://doi.org/10.1128/JB.180.10.2670-2675.1998.
Grangeasse C, Nessler S, Mijakovic I. Bacterial tyrosine kinases: evolution, biological function and structural insights. Phil Trans R Soc Lond B Biol Sci. 2012;367(1602):2640–55. https://doi.org/10.1098/rstb.2011.0424.
Hagelueken G, Huang H, Mainprize IL, Whitfield C, Naismith JH. Crystal Structures of Wzb of Escherichia coli and CpsB of Streptococcus pneumoniae, representatives of two families of tyrosine phosphatases that regulate capsule assembly. J Mol Biol. 2009;392(3):678–88. https://doi.org/10.1016/j.jmb.2009.07.026.
Kim HS, Lee SJ, Yoon HJ, An DR, Kim DJ, Kim SJ, et al. Crystal structures of YwqE from Bacillus subtilis and CpsB from Streptococcus pneumoniae, unique metal-dependent tyrosine phosphatases. J Struct Biol. 2011;175(3):442–50. https://doi.org/10.1016/j.jsb.2011.05.007.
Ray LC, Das D, Entova S, Lukose V, Lynch AJ, Imperiali B, et al. Membrane association of monotopic phosphoglycosyl transferase underpins function. Nat Chem Biol. 2018;14(6):538–41. https://doi.org/10.1038/s41589-018-0054-z.
Roy B, Field RA, Mukhopadhyay B. Synthesis of a tetrasaccharide related to the repeating unit of the Oantigen from Escherichia coli K12. Carbohydr Res. 2009;344(17):2311–6. https://doi.org/10.1016/j.carres.2009.09.026.
Schmid J, Sieber V, Rehm B. Bacterial exopolysaccharides:biosynthesis pathways and engineering strategies. Front Microbiol. 2015;6:496. https://doi.org/10.3389/fmicb.2015.00496.
Coutinho PM, Deleury E, Davie GJ, Henrissat B. An Evolving Hierarchical Family Classification for Glycosyltransferases. J Mol Biol. 2003;328(2):307–17. https://doi.org/10.1016/S0022-2836(03)00307-3.
Sperisen P, Schmid CD, Bucher P, Zilian O. Stealth proteins in silico identification of a novel protein family rendering bacterial pathogens invisible to host immune defense. PLoS Comput Biol. 2005;1(6):0492–9. https://doi.org/10.1371/journal.pcbi.0010063.
KOplin R, Brisson JR, Whitfield C. Udp-Galactofuranose precursor required for formation Of Thelipopolysaccharide O Antigen of Klebsiella Pneumoniae serotype 01 is synthesized by the product Ofthe Rfbd (Kp01) Gene. J Biol Chem. 1997;272(7):4121–8. https://doi.org/10.1074/jbc.272.7.4121.
Wiseman B, Widmalm G, Högbom M. Alternating L4 loop architecture of the bacterial polysaccharide co-polymerase WzzE. Commun Biol. 2023;6(1):802. https://doi.org/10.1038/s42003-023-05157-7.
Tamburrini KC, Terrapon N, Lombard V, Bissaro B, Longhi S, Berrin JG. Bioinformatic Analysis of Lytic Polysaccharide Monooxygenases Reveals the Pan-Families Occurrence of Intrinsically Disordered C-Terminal Extensions. Biomolecules. 2021;11(11):1632. https://doi.org/10.3390/biom11111632.
Ferri M, Ranucci E, Romagnoli P, Giaccone V. Antimicrobial resistance: A global emerging threat to public health systems. Crit Rev Food Sci Nutr. 2017;57(13):2857–76. https://doi.org/10.1080/10408398.2015.1077192.
Westra ER, Buckling A, Fineran PC. CRISPR-Cas systems: beyond adaptive immunity. Nat Rev Microbiol. 2014;12:317–26. https://doi.org/10.1038/nrmicro3241.
Zhang Q, Ye Y. Not all predicted CRISPR-Cas systems are equal: isolated cas genes and classes of CRISPR like elements. BMC Bioinformatics. 2017;18(1):92. https://doi.org/10.1186/s12859-017-1512-4.
Mathur S, Singh R. Antibiotic resistance in food lactic acid bacteria-a review. Int J Food Microbiol. 2005;105(3):281–95. https://doi.org/10.1016/j.ijfoodmicro.2005.03.008.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 32272359) and Guizhou Provincial Basic Research Program (Natural Science) (Grant No. ZK [2023] 091).
Author information
Authors and Affiliations
Contributions
K.L.: Methodology, Investigation, Data curation, Software, Writing original draft. X.W.: Investigation, Data curation, Software. Y.Z.: Writingreview and editing, Supervision, Funding the project. Q.Z.: Conceptualization, Writing-review and editing, Resources, Supervision, Funding the project. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Lu, K., Wang, X., Zhou, Y. et al. Genomic characterization and probiotic potential assessment of an exopolysaccharide-producing strain Pediococcus pentosaceus LL-07 isolated from fermented meat. BMC Microbiol 24, 142 (2024). https://doi.org/10.1186/s12866-024-03304-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12866-024-03304-6