
Abstract
Ecological resource availability is crucial for the persistence and survival of local desert animal communities. Dryland resources such as shrubs and burrows positively benefit animal species by mitigating harsh abiotic factors and providing habitat. Understanding the role of native shrubs, many of which serve as foundation species within desert regions, as well as the function of underground burrows as resources, provides insights into habitat utilization. In this study, we seek to better understand the co-occurrence of these two resources as a first step in quantifying key patterns locally and regionally in drylands. We tested whether the presence of burrows increased with the density of foundational shrubs near the burrows at two scales—within a 5 m radius of every burrow recorded and at the site level—defined as discrete ecological areas. We performed fieldwork across 31 sites within the arid and semiarid regions of Central California. We used a combination of burrow field surveys and satellite imagery to document both vertebrate animal burrow frequencies and shrub densities. Additionally, the accuracy of the shrub data was verified through ground truthing. Both fine-scale and site-level shrub densities positively predicted the relative likelihood of burrows and the frequency of burrows, respectively. The existence of two highly utilized dryland resources and the relationship between them signal that areas abundant in both resources will likely better support resident animal species. This finding underscores the significance of incorporating both shrub density and burrow frequency in studies of habitat interconnectivity and quality. The co-occurrence patterns of these resources will support novel habitat management and conservation strategies designed around both conservation and restoration efforts.
Similar content being viewed by others

Introduction
Global dryland ecosystems are fundamentally shaped by semifossorial herbivorous mammals, many of which are considered ecosystem engineers and have a variety of positive impacts on the surrounding arid region [1, 2]. These impacts that ecosystem engineers exert on their environment often have a disproportionate influence on the patterns of species diversity in their communities [3, 4]. Utilizing burrows is a common behavioral adaptation for animal species that occur in dryland regions across the globe, as it can provide animals with several benefits, including a sheltered microclimate, protection from predators, and access to food [5,6,7,8]. In dryland ecosystems, these burrows provide many local species with a more favorable microclimate by buffering temperature extremes and increasing relative humidity, thus creating a more suitable microclimate [9,10,11]. Often found in large communal colonies, burrowing mammals can transform dryland regions through the creation of complex underground burrows and feeding activity patterns [12,13,14,15]. These burrows range from microsite-level to extensive regional-level systems, resulting in major impacts on soil dynamics, vegetation patterns and animal community diversity, ultimately increasing heterogeneity across a region [1, 9, 10, 16, 17]. Within southern California, the giant kangaroo rat (Dipodomys ingens) is a state and federally endangered species of burrowing rodent [18]. The extensive network of burrows that giant kangaroo rats construct provides habitat for the federally endangered blunt-nosed leopard lizard (Gambelia sila) and the state-threatened San Joaquin antelope squirrel (Ammospermophilus nelsoni) [19,20,21]. Giant kangaroo rats are also the primary prey source for the endangered San Joaquin kit fox (Vulpes macroitis mutica) and thus play an integral role in the structure and function of the ecosystem [18].
Rodent burrows serve as pivotal hubs for their activities within the region. Essential for the survival of small vertebrate species that occur in harsh arid environments, these burrows serve as a crucial resource, providing refuge from extreme temperatures and predators. As focal points for rodent activity, burrows influence the spatial distribution of vegetation, particularly in their immediate vicinity [3]. The relative frequency of rodent activity decreases with increasing distance from these burrows, leading to compositional heterogeneity in vegetation, which tends to increase closer to burrows [17]. This heightened diversity and variability in plant species abundance and distribution are attributed to localized effects such as seed dispersal, soil disturbance, and nutrient cycling facilitated by rodent activity [15, 18]. Understanding these spatial patterns of burrowing rodent activity and their interplay with other local landscape resources is crucial for elucidating the intricate dynamics within dryland regions.
Vegetation is an important part of any region because it can not only influence local vertebrate and invertebrate communities but also shape the structure of an ecosystem [22, 23]. Positive interactions associated with local vegetation are incredibly important in resource-limited environments, such as those found in dryland regions [24]. Dominant shrub species in arid environments are often beneficial to a wide range of animal and plant species, resulting in frequent positive interactions [24, 25]. In dryland regions, facilitation often occurs when certain shrubs create microclimates that provide shade, trap moisture, and offer protection, thus promoting the survival and growth of neighboring plant and animal species [22, 26]. This mutualistic interaction is beneficial for the environment because it enhances biodiversity, stabilizes ecosystems, and contributes to the overall resilience of desert regions by fostering conditions conducive to plant and animal life despite the challenging arid environment [16, 22, 27, 28]. In dryland regions, it has been shown that heterogeneous shrub density can benefit local vertebrate and invertebrate species, thus increasing habitat heterogeneity across arid regions [20, 24, 29]. The increase in shrub availability has the potential to provide more possible interactions for local species, thus increasing the frequency of facilitative interactions [8, 30]. A better understanding of how both burrows created by local vertebrates and naturally occurring foundational shrub species are spatially related will provide meaningful insight into how local animal communities utilize regional resources to persist in harsh arid environments.
The benefits associated with foundational shrub species within dryland ecosystems can promote the association of burrowing vertebrate species [24, 31]. These association can drive an increase in the abundance of burrows around shrub individuals [32]. In addition, these burrowing vertebrate species can disperse seeds throughout the landscape, promoting shrub growth at burrow areas [33]. This suggests that there should be an association between shrub densities and burrow abundance within dryland ecosystems. As these dryland ecosystems experience drier and higher temperatures associated with increasing aridity, the dependency on these foundational shrub species can potentially increase [30]. The benefits through shading and the microclimate produced from these shrubs can ameliorate these increasingly arid conditions [30, 31].
While previous studies have examined the independent utilization of burrows and foundation shrubs by local desert animal communities, there remains a gap in understanding the spatial association between these resources at both fine and site-level scales [24, 34, 35]. Here, we tested the hypothesis that there is a relationship between the fine-scale and site-level density estimates of the foundational shrub species E. californica and L. tridentata and the presence and relative frequencies of burrows formed by vertebrate animal species.
We tested the following predictions:
-
The relative frequency of burrows will increase total shrub density at each site.
-
Increasing relative aridity will increase the effects of shrub density on the total burrows per site.
-
The likelihood of burrow presence increases with fine-scale shrub density (i.e., within a 5 m radius, as tested here).
Methods
Species and Study sites
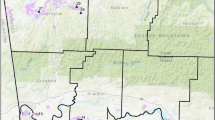
A total of 30 sites were established across southern California on an east-to-west gradient (Fig. 1; Supplementary Table S1) [36]. Key variables describing sites included foundational shrub species, geographic location, mean annual temperature (MAT), mean annual precipitation (MAP), and estimated shrub density (Supplementary Table S1). These variables were used to select and differentiate sites used within this study. A total of 30 sites were used to encompass a gradient of climate and shrub densities ranging from 0 to 166 shrubs per site (Supplementary Table S1). All sites were dominated by either the woody shrub species Ephedra californica or Larrea tridentata (Supplementary Figure S1 & S2) [8, 37, 38]. Both shrub species function as ecological foundational species for both plant [26, 38, 39] and animal communities by increasing species abundance and richness of many taxa [20, 24, 40]. These species include small to medium-sized vertebrates, including antelope squirrels (Ammospermophilus sp.) kangaroo rats (Dipodomys sp.), blunt-nosed leopard lizards (Gambelia sila), and black-tailed jack rabbits (Lepus californicus) [24, 41,42,43,44,45,46,47,48,49,50,51]. Gradients of these foundational shrubs across California influence the associations of local animal community distributions and composition [24, 30, 46].

Geographical representation of all study sites within southern California, U.S.A. A total of 30 unique sites were utilized for burrow and shrub data collection through the 2023 field season. All sampled sites are indicated with black circles. Random samples of burrows were taken at all 30 sites where the presence and density of shrubs were recorded via satellite imagery. The map was created through R version 4.3.1
Site-level shrub density effects on total burrows
The annual aridity of each site was determined using the De Martonne aridity index equation AI = P/(MAT + 10) where P represents the annual precipitation in mm within 20 years and MAT represents the mean annual temperature (°C) within 20 years [52]. These climate data were retrieved from the WorldClim version 2.1 database at a 1 km spatial resolution [52, 53]. Shrub density estimates for each of the 30 sites were derived from remotely sensed imagery. Specifically, Ephedra californica and Larrea tridentata density were estimated using Google Earth composite satellite images [8]. The base layer of these maps retrieved by google were sourced by the Airbus Earth Observation Satellite Imagery Services with a spatial resolution of 30 cm [54]. All individual shrubs at each site were geolocated and given a unique identifier. Once all E. californica and L. tridentata individuals were identified, a series of keyhole markup language files (KMLs) were extracted. The latitude and longitude coordinates for each shrub were reported [55, 56]. A total of 200 random points were ground-truthed at one of the thirty sites to confirm the accuracy of the satellite-based shrub density measures within a 5-meter plot in field [57]. Total burrows per site were then calculated for site level density effects using the detailed field survey protocol described below.
Fine-scale analysis of shrub density effects on burrows
Burrow surveys were done at the same 30 sites from May to June of 2023. A single 25 m x 25 m quadrat was randomly established at each site. The quadrat was used to visually count burrows created by vertebrates. Coordinates (decimal degrees) were collected at each independent burrow opening using a handheld GPS (Garmin-GPSMAP65). Shrub density within a 5 m radius was estimated for each burrow within the 25 m x 25 m quadrate by combining in field burrow observations with the satellite shrub data. Although the resolution of burrow points and shrubs can vary between 3 m and 5 m, potentially leading to false positive associations [8], our analysis explored associations within various radii including 5 m, 10 m, and 20 m. We tested theses associations at 5 m, 10 m, and 20 m radii for all 30 sites. The observed patterns remain robust across these different spatial scales. Pseudoabsence points were spatially generated using a random sampling approach using the R package sf [58]. Field observations were then used to ensure burrow openings were correctly attributed to vertebrates (Supplementary Figure S4 & S5). Vertebrate burrows typically display visible tracks or nearby feces, that the current pattern use from invertebrate burrows (Supplementary Figures S3, S4, & S5). To further eliminate the possible misclassification during burrow surveys, instances where burrows exhibited clear indications of frequent or recent rodent activity were recorded along with detailed characteristics [59]. These data are published alongside all other attributes recorded [59]. Burrow diameters were measured and ranged from 4.4 cm to 11.2 cm. Vertebrate animal responsible for burrow formations were not identified.
Statistical analysis
Site-level shrub density effects on total burrows
A second-order polynomial regression model was used to test the effect of site-level shrub density on the total number of burrows recorded per site [60]. Total number of burrows per site was the response variable, and total shrub density per site was the main factor. Aridity was also examined as a factor. All assumptions of regression were tested for the site-level analysis including multicollinearity using the R package ‘performance’ [61].
Fine-scale analysis of shrub density effects on burrow
The relative importance of shrub density on predicted presence of burrows at fine-scales, i.e. within a 5 m radius of each burrow, was tested with a general linear mixed model with binary data - otherwise known as a logistic regression [62]. Since the response variable within the models was represented by binary data, the presence or absence of a shrub, a logistic regression best estimates the probability of the observations belonging to one of the two categories [62]. The presence-absence of burrows was the response variable while shrub density within 5 m of each burrow was the factor, nested within sites. This presence-absence data were modelled as binomial [63]. The package ‘aod’ was used [64] to calculate the predicted probabilities of burrows at the range of shrub densities calculated. The null likelihood and full model with shrub density effects at probability of burrows were contrasted using the ‘performance’ R package function test_performance [65, 66]. The probability of burrow presences and absence range based on certainty and do not necessarily assume up to 1 or 100% [67]. All statistical analyses and models were done in R version 4.3.1 [68].
Results
Site-level shrub density effects on total burrows
There were no statistically significant differences between the geolocated shrub density estimates and the ground-truthed, field shrub density counts (Supplement Table S2; paired t-test, t = -0.05, df = 389.41, p = 0.96). Total shrub density per site significantly predicted the total number of burrows (Fig. 2; regression, R2 = 0.28, df = 26, p-value = 0.03). Increasing aridity decreased the total number of burrows per site (estimate 21.1, df = 27, p-value = 0.001), but aridity across sites did not significantly influence the positive effects of shrub density on burrows (estimate = 8.09, df = 26, p-value = 0.15). Regressions to explore shrub densities within 10 m and 20 m of burrows per site expressed similar trends (Supplement Table S3).

The effect of shrub density on the total number of burrows across Central California desert sites. The total count of burrows observed in the field surveys was regressed against the total shrub density per site from satellite imagery data (r2 = 0.28, p value = 0.03)
Fine-scale analysis of shrub density effects on burrows
The probability of burrow presence significantly increased with increasing shrub density within a 5 m radius of each observed instance (Fig. 3; GLMM, estimate = 161.46, df = 26, p value = 0.001). The predictive model nested by site performed significantly better than the non-nested model (GLM; ω2 = 0.04, LR = 161.46, p-value < 0.001).

The predicted probabilities of burrows with increasing shrub density within a 5 m radius. The presence of a burrow is shown in dark blue (1), and the absence of a burrow is shown in light blue (0). Shaded areas show the 95% confidence intervals for each line of best fit
Discussion
Exploring the co-occurrence patterns between burrow abundance and shrub density can reveal some of the key forms of heterogeneity in resources within dryland regions. Here, we tested whether there is a relationship between the density of foundational shrub species and the presence of burrows across various regions within Central California. We found evidence supporting the hypothesis that sites with foundational shrubs exhibit a significantly high number of burrows. Shrub density was greater around burrows than in areas where burrows were absent, and increasing shrub density significantly predicted a greater probability of burrow presence.
Physical structure can influence ecological processes in a variety of complex ways, influencing how local species communities form, function, and persist [69, 70]. Burrow systems are more than just ‘holes in the ground’; they are a critical resource for many species [18, 21, 71]. Burrowing mammals generally have a positive impact on species richness at the regional level by increasing habitat diversity [10, 71]. The potential consequences of losing burrowing mammals in desert regions extend beyond a mere disruption of biodiversity; they may have a disproportional impact on the distribution of other organisms and the overall functioning of desert ecosystems. In particular, the absence of burrow systems could exacerbate temperature extremes and sun exposure in arid environments, creating unfavorable conditions for various species [1, 19, 30]. Moreover, the combined threats of wildfires, desertification, a decline in shrub health, and the negative stigma of shrub encroachment pose additional risks to the vertebrate populations inhabiting these regions [72,73,74]. These challenges will lead to disruptions in behavioral patterns, migration routes, and reproductive success, ultimately posing a threat to the ecological balance of these ecosystems [2, 10]. Understanding the multifaceted role of burrow systems is therefore essential for comprehending their broader implications for regional dynamics and biodiversity conservation.
Increasing annual temperatures during summer are a major issue for vertebrates in desert regions [75]. When faced with continuous solar radiation and high temperatures, some species of mammals visit cooler microclimates to reduce their overall exposure [74.75]. The dehydration effect of dry desert air on life forms is a second significant barrier to survival in desert and arid regions [6]. The microclimate of subterranean rodent burrows has been shown to be much more favorable than that on the surface because the relative humidity inside burrows is near saturation [6, 12, 71]. Ecosystem engineers that construct these site-level burrows can directly influence plant communities through a variety of mechanisms and thus play integral roles in shaping the surrounding vegetation and ecosystem dynamics [1, 13, 75]. Dipodomys sp. in California maintain circles of bare ground around their burrows, creating open spaces lacking annual plants [18, 71, 76]. They have also been known to inadvertently transport seeds within their fur or bury them in caches near their burrows, facilitating the dispersal and germination of plant species across the region [75, 77, 78]. The intricate design and network of these burrows exert substantial influence on the surrounding environment, shaping both direct and indirect impacts on local animal and plant communities [75, 77]. On a regional scale, the presence of burrowing animals and burrows can promote biodiversity by providing habitat and resources for a wide range of plant and animal species. Additionally, the ecosystem engineering effects of burrowing animals contribute to nutrient cycling, soil aeration, and water infiltration, thereby influencing the resilience and functioning of dryland regions [71, 79]. Foundational shrubs typically benefit animal communities in dryland regions through a variety of mechanisms, including providing a refuge from harsh temperatures, serving as a food source for some small animals, and preventing predation [8, 22, 80]. Zuliani et al. [24] reported greater numbers of burrowing mammals, such as Dipodomys sp. and Ammospermophilus sp., in areas with greater shrub densities, supporting the idea that smaller vertebrate species are reliant on the positive effects of foundational shrubs.
Although not explicitly examined in this research, one of the primary functions of burrows and shrubs is to provide shelter from intense temperatures for various animal species [20, 30]. Several studies have reported greater associations of animals near shrubs during peak times of the day, suggesting that thermal amelioration is a direct benefit to these species [22, 30, 81]. This positive association of animal species with shrubs can help us explain the greater probability of burrows being observed, as several burrowing species are more frequently observed in these areas. This further supports our findings that increasing shrub density can predict the probability of burrows being present. Having a burrow within proximity to a shrub allows for many benefits, such as acting as a refuge from predation [30, 48]. In the event of a predation attempt, the presence of both a burrow and shrub can allow for either resource to be utilized as a means of escape [21, 25, 40]. It also allows for closer proximity to a food source and the ability to spend time above or below ground while still being proximal to one’s home [71]. The intricate relationships between foundational shrubs, burrowing species, and their surrounding environment highlight the significance of structural landscape features in shaping species associations and interactions in arid regions [82]. Understanding the multifaceted benefits, ranging from thermal amelioration to refuge from predation, underscores the pivotal role that increasing shrub density plays in predicting the presence of burrows.
The presence of two key desert habitat resources, shrubs and burrows, shows that both can potentially benefit local animal species [83,84,85] provided that we deepen our understanding and empirical knowledge of co-occurrence patterns. These data will inform habitat conservation and restoration efforts. With desertification affecting at-risk and endangered species, these findings could help guide conservation efforts by emphasizing the importance of animal burrows and the facilitative interactions between foundational shrubs and target animal species [57, 86]. As these arid regions continue to deteriorate overall ecosystem health, it is critical to investigate and include other regional resources that influence the associations between plant and animal communities. These discoveries may offer valuable perspectives on safeguarding critical habitats in ecosystems that are essential for endangered species. This insight can facilitate targeted species rehabilitation with minimal human intervention, as land managers and restoration biologists can leverage these findings to enhance the habitat association of the targeted species.
Data availability
All Burrow data is available at https://doi.org/10.6084/m9.figshare.22272622.v5.
References
Mallen-Cooper M, Nakagawa S, Eldridge DJ. Global meta‐analysis of soil‐disturbing vertebrates reveals strong effects on ecosystem patterns and processes. Keith S, editor. Glob Ecol Biogeogr. 2019;28(5):661–79.
Vázquez J, Álvarez-Castañeda ST. Influence of kangaroo rat burrows in the spatial organization of the San Jose Island rodent community. J Arid Environ. 2014;111:84–90.
Louw MA, Haussmann NS, le Roux PC. Testing for consistency in the impacts of a burrowing ecosystem engineer on soil and vegetation characteristics across biomes. Sci Rep. 2019;9(1):19355.
Prugh LR, Brashares JS. Partitioning the effects of an ecosystem engineer: kangaroo rats control community structure via multiple pathways. J Anim Ecol. 2012;81(3):667–78.
Louw MA, le Roux PC, Meyer-Milne E, Haussmann NS. Mammal burrowing in discrete landscape patches further increases soil and vegetation heterogeneity in an arid environment. J Arid Environ. 2017;141:68–75.
Baldo MB, Antenucci CD, Luna F. Effect of ambient temperature on evaporative water loss in the subterranean rodent Ctenomys talarum. J Therm Biol. 2015;53:113–8.
Whitford WG, Kay FR. Biopedturbation by mammals in deserts: a review. J Arid Environ. 1999;41(2):203–30.
Zuliani M, Ghazian N, Owen M, Westphal MF, Butterfield HS, Lortie CJ. Shrub density effects on the presence of an endangered lizard of the Carrizo Plain National Monument, California. Ecol Evol. 2023;13(5):e10128.
Davidson AD, Detling JK, Brown JH. Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world’s grasslands. Front Ecol Environ. 2012;10(9):477–86.
Davidson AD, Lightfoot DC, McIntyre JL. Engineering rodents create key habitat for lizards. J Arid Environ. 2008;72(12):2142–9.
Bongiovanni SB, Borruel Díaz NG, Borghi CE. Ecosystem engineering in a high and cold desert: Effect of a subterranean rodent on lizard abundance and behavior. Austral Ecol. 2023;48(7):1238–44.
Davidson AD, Lightfoot DC. Burrowing rodents increase landscape heterogeneity in a desert grassland. J Arid Environ. 2008;72(7):1133–45.
Miranda V, Rothen C, Yela N, Aranda-Rickert A, Barros J, Calcagno J, Fracchia S. Subterranean Desert rodents (Genus Ctenomys) create soil patches enriched in Root Endophytic Fungal Propagules. Microb Ecol. 2019;77(2):451–9.
Wada N, Narita K, Kumar S, Furukawa A. Impact of overgrazing on seed predation by rodents in the Thar desert, northwestern India. Ecol Res. 1995;10(2):217–21.
Shaw WT. The ability of the Giant Kangaroo Rat as a Harvester and Storer of seeds. J Mammal. 1934;15(4):275.
Eldridge DJ, Whitford WG, Duval BD. Animal disturbances promote shrub maintenance in a desertified grassland. J Ecol. 2009;97(6):1302–10.
Yoshihara Y, Okuro T, Buuveibaatar B, Undarmaa J, Takeuchi K. Clustered animal burrows yield higher spatial heterogeneity. Plant Ecol. 2010;206(2):211–24.
Bean WTM. Spatial Ecology of the Giant Kangaroo Rat (Dipodomys ingens): A Test of Species Distribution Models as Ecological Revealers [Internet] [Dissertation]. [California]: University of California, Berkeley; 2012. https://escholarship.org/uc/item/25390639.
Cypher BL, Kelly EC, O’Leary R, Phillips SE, Saslaw LR, Tennant EN, Westall TL. Conservation of threatened San Joaquin antelope squirrels: distribution surveys, habitat suitability, and conservation recommendations. Calif Fish Wildl J. 2021;(CESA Special Issue):345–66.
Gaudenti N, Nix E, Maier P, Westphal MF, Taylor EN. Habitat heterogeneity affects the thermal ecology of an endangered lizard. Ecol Evol. 2021;11(21):14843–56.
Germano DJ, Rathbun GB, Saslaw LR, Cypher BL. Home range and movements of San Joaquin antelope squirrels in the San Joaquin Desert of California. Calif Fish Wildl J. 2021;(CESA Special Issue):317–28.
Lortie CJ, Filazzola A, Sotomayor DA. Functional assessment of animal interactions with shrub-facilitation complexes: a formal synthesis and conceptual framework. Michalet R, editor. Funct Ecol. 2016;30(1):41–51.
Schooley RL, Sharpe PB, Horne BV. Can shrub cover increase predation risk for a desert rodent? Can J Zool. 1996;74(1):157–63.
Zuliani M, Ghazian N, Lortie CJ. Shrub density effects on the community structure and composition of a desert animal community. Wildl Biol [Internet]. 2021 Apr 1 [cited 2023 Dec 7];2021(2). https://bioone.org/journals/wildlife-biology/volume-2021/issue-2/wlb.00774/Shrub-density-effects-on-the-community-structure-and-composition-of/https://doi.org/10.2981/wlb.00774.full.
Filazzola A, Westphal M, Powers M, Liczner AR, (Smith), Woollett DA, Johnson B, Lortie CJ. Nontrophic interactions in deserts: Facilitation, interference, and an endangered lizard species. Basic Appl Ecol. 2017;20:51–61.
Filazzola A, Liczner AR, Westphal M, Lortie CJ. Shrubs indirectly increase desert seedbanks through facilitation of the plant community. Liu J, editor. PLOS ONE. 2019;14(4):e0215988.
Holzapfel C, Mahall BE. Bidirectional facilitation and interference between shrubs and annuals in the Mojave desert. Ecology. 1999;80(5):1747–61.
Rolhauser AG, Pucheta E. Annual plant functional traits explain shrub facilitation in a desert community. Kikvidze Z, editor. J Veg Sci. 2016;27(1):60–8.
Cosentino BJ, Schooley RL, Bestelmeyer BT, Coffman JM. Response of lizard community structure to desert grassland restoration mediated by a keystone rodent. Biodivers Conserv. 2013;22(4):921–35.
Ivey KN, Cornwall M, Crowell H, Ghazian N, Nix E, Owen M, Zuliani M, Lortie CJ, Westphal M, Taylor E. Thermal ecology of the federally endangered blunt-nosed leopard lizard (Gambelia sila). Cooke S, editor. Conserv Physiol. 2020;8(1):coaa014.
Zuliani M, Ghazian N, MacDonald S, Lortie CJ. The relative effects of artificial shrubs on animal community assembly. Restor Ecol. 2024;e14121. https://doi.org/10.1111/rec.14121.
Reichman OJ, Smith SC. Impact of Pocket Gopher Burrows on overlying Vegetation. J Mammal. 1985;66:720–5.
Godó L, Valkó O, Borza S, Deák B. A global review on the role of small rodents and lagomorphs (clade glires) in seed dispersal and plant establishment. Global Ecol Conserv. 2022;33:e01982.
Baumberger KL, Eitzel MV, Kirby ME, Horn MH. Movement and habitat selection of the western spadefoot (Spea hammondii) in southern California. Halliday WD, editor. PLOS ONE. 2019;14(10):e0222532.
McNew LB, Dahlgren DK, Beck JL, editors. Rangeland Wildlife Ecology and Conservation [Internet]. Cham: Springer International Publishing; 2023 [cited 2024 Mar 20]. https://link.springer.com/https://doi.org/10.1007/978-3-031-34037-6.
Lortie C, Zuliani M, Ghazian N, Braun J, Haas S, King R. A list of ecological study sites within Central California Drylands [Internet]. [object Object]; 2022 [cited 2024 Mar 20]. https://knb.ecoinformatics.org/view/doi:https://doi.org/10.5063/F1X34VXT.
Filazzola A, Brown C, Westphal M, Lortie CJ. Establishment of a desert foundation species is limited by exotic plants and light but not herbivory or water. Hölzel N, editor. Appl Veg Sci. 2020;23(4):586–97.
Lortie CJ, Gruber E, Filazzola A, Noble T, Westphal M. The Groot Effect: Plant facilitation and desert shrub regrowth following extensive damage. Ecol Evol. 2018;8(1):706–15.
Lucero JE, Seifan M, Callaway RM, Lortie CJ. Positive associations with native shrubs are intense and important for an exotic invader but not the native annual community across an aridity gradient. Lambrinos J, editor. Divers Distrib. 2020;26(9):1177–97.
Owen E, Zuliani M, Goldgisser M, Lortie C. The importance of native shrubs on the distribution and diversity of reptiles and amphibians in the central deserts of California [Internet]. In Review; 2023 Oct [cited 2024 Mar 20]. https://www.researchsquare.com/article/rs-3482635/v1.
Braun J, Westphal M, Lortie CJ. The shrub Ephedra californica facilitates arthropod communities along a regional desert climatic gradient. Ecosphere. 2021;12(9):e03760.
Ruttan A, Lortie CJ, Haas SM. Shrubs as magnets for pollination: a test of facilitation and reciprocity in a shrub-annual facilitation system. Curr Res Insect Sci. 2021;1:100008.
Hollander JL, Vander Wall SB, Baguley JG. Evolution of seed dispersal in North American Ephedra. Evol Ecol. 2010;24(2):333–45.
Loew SS, Williams DF, Ralls K, Pilgrim K, Fleischer RC. Population structure and genetic variation in the endangered giant Kangaroo Rat (Dipodomys ingens). Conserv Genet. 2006;6(4):495–510.
Bean WTM. Spatial Ecology of the Giant Kangaroo Rat (Dipodomys ingens): A Test of Species Distribution Models as Ecological Revealers.
Westphal MF, Noble T, Butterfield HS, Lortie CJ. A test of desert shrub facilitation via radiotelemetric monitoring of a diurnal lizard. Ecol Evol. 2018;8(23):12153–62.
Zuliani M, Ghazian N, MacDonald SE, Lortie CJ. The Importance of Variation Shrub Densities on Local Vertebrate Association Across an Aridity Gradient [Internet]. SSRN; 2023 [cited 2024 Mar 21]. https://www.ssrn.com/abstract=4615528.
Noble TJ, Lortie CJ, Westphal M, Butterfield HS. A picture is worth a thousand data points: an imagery dataset of paired shrub-open microsites within the Carrizo Plain National Monument. GigaScience. 2016;5(1):40.
Zuliani M, Ghazian N, Lortie CJ. A meta-analysis of shrub density as a predictor of animal abundance. Wildl Biol. 2023;2023(3):e01042.
Lapinski W. Tarantulas and Their Habitats. In: Pérez-Miles F, editor. New World Tarantulas [Internet]. Cham: Springer International Publishing; 2020 [cited 2024 Mar 22]. pp. 191–236. (Zoological Monographs; vol. 6). http://link.springer.com/https://doi.org/10.1007/978-3-030-48644-0_7.
Polis GA, Myers C, Quinlan M. Burrowing biology and spatial distribution of desert scorpions. J Arid Environ. 1986;10(2):137–46.
Oliver JE, editor. Encyclopedia of World Climatology [Internet]. Springer Netherlands; 2005 [cited 2024 Mar 22]. (Encyclopedia of Earth Sciences Series). http://link.springer.com/https://doi.org/10.1007/1-4020-3266-8.
Fick SE, Hijmans RJ. WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int J Climatol. 2017;37(12):4302–15.
Hestrio YF, et al. Satellite data receiving antenna system for pleiades neo observation satellite. J Phys : Conf Ser. 2021;1763:012019.
Zuliani M, Lortie C. Geolocations of all Ephedra californica shrubs at key ecological areas within the San Joaquin Desert. [Internet]. [object Object]; 2022 [cited 2024 Mar 20]. https://knb.ecoinformatics.org/view/doi:https://doi.org/10.5063/F1HT2MRF.
Zuliani M, Lortie C. Geolocations of all Ephedra californica shrubs at key ecological areas within the Mojave Desert. [Internet]. [object Object]; 2022 [cited 2024 Mar 20]. https://knb.ecoinformatics.org/view/doi:https://doi.org/10.5063/F1D21W2W.
lortie cj, Noble T, Zuliani M, Westphal F, Nix M, Ghazian E, Owen N, Scott Butterfield M, Braun H. J. Telemetry of the lizard species Gambelia sila at Carrizo Plain National Monument [Internet]. [object Object]; 2020 [cited 2024 Mar 23]. p. 196242 Bytes. https://figshare.com/articles/dataset/Telemetry_of_the_lizard_species_Gambelia_sila_at_Carrizo_Plain_National_Monument/8239667/2.
Barbet-Massin M, Jiguet F, Albert CH, Thuiller W. Selecting pseudo‐absences for species distribution models: how, where and how many? Methods Ecol Evol. 2012;3(2):327–38.
Owen E. lortie cj. A survey of burrows in the drylands of Central California dataset. [Internet]. [object Object]; 2023 [cited 2024 Mar 22]. p. 40372 Bytes. https://figshare.com/articles/dataset/February_2023_burrow_survey_data/22272622/2.
Ostertagová E. Modelling using Polynomial Regression. Procedia Eng. 2012;48:500–6.
Harrell FE. Regression modeling strategies: with applications to Linear models, logistic and ordinal regression, and Survival Analysis. Cham: Springer International Publishing; 2015. https://doi.org/10.1007/978-3-319-19425-7.
Jamil T, Ozinga WA, Kleyer M. Ter Braak, C. J. F. selecting traits that explain species–environment relationships: a generalized linear mixed model approach. J Veg Sci. 2013;24:988–1000.
Manel S, Williams HC, Ormerod SJ. Evaluating presence–absence models in ecology: the need to account for prevalence. J Appl Ecol. 2001;38:921–31.
CRAN - Package. aod [Internet]. [cited 2024 Mar 21]. https://cran.r-project.org/web/packages/aod/index.html.
Bivand R, Pebesma EJ, Gómez-Rubio V. Applied spatial data analysis with R. Second edition. New York: Springer; 2013. 405 p. (Use R!).
Lüdecke D, Ben-Shachar M, Patil I, Waggoner P, Makowski D. Performance: an R Package for Assessment, comparison and testing of statistical models. JOSS. 2021;6:3139.
Starbuck C. The fundamentals of People Analytics: with applications. Cham: Springer International Publishing; 2023. https://doi.org/10.1007/978-3-031-28674-2.
R. A Language and Environment for Statistical Computing [Internet]. The R Project for Statistical Computing. [cited 2024 Mar 20]. https://www.r-project.org/.
Nicolai N. Ecological engineers’ nests benefit plant recovery following fire in a semiarid grassland, New Mexico, USA. Overbeck G, editor. J Veg Sci. 2019;30(4):709–19.
Surkova E, Popov S, Tchabovsky A. Rodent burrow network dynamics under human-induced landscape transformation from desert to steppe in Kalmykian rangelands. Integr Zool. 2019;14(4):410–20.
Kinlaw A. A review of burrowing by semi-fossorial vertebrates in arid environments. J Arid Environ. 1999;41(2):127–45.
Doblas-Miranda E, Alonso R, Arnan X, Bermejo V, Brotons L, De Las Heras J, Estiarte M, Hódar JA, Llorens P, Lloret F, López-Serrano FR, Martínez-Vilalta J, Moya D, Peñuelas J, Pino J, Rodrigo A, Roura-Pascual N, Valladares F, Vilà M, Zamora R, Retana J. A review of the combination among global change factors in forests, shrublands and pastures of the Mediterranean Region: beyond drought effects. Glob Planet Change. 2017;148:42–54.
Sivakumar MVK. Interactions between climate and desertification. Agric Meteorol. 2007;142(2–4):143–55.
Stavi I. Wildfires in grasslands and shrublands: a review of impacts on Vegetation, Soil, Hydrology, and Geomorphology. Water. 2019;11(5):1042.
Edelman AJ. Kangaroo Rats Remodel Burrows in response to Seasonal Changes in Environmental conditions: Seasonal changes in Burrow Architecture. Ethology. 2011;117(5):430–9.
Tracy RL, Walsberg GE. Kangaroo rats revisited: re-evaluating a classic case of desert survival. Oecologia. 2002;133(4):449–57.
Edelman AJ. Dispersal, facilitation, and burrow architecture in banner-tailed kangaroo rats [Internet] [Dissertation]. The University of New Mexico; 2010. https://www.proquest.com/openview/53aa1187e1c6a23085b3bf5df678d643/1?pq-origsite=gscholar&cbl=18750
Miles DC, Burrus KR, Shoemaker KT. First records of California kangaroo rats, Dipodomys californicus, in Nevada. Northwest Nat. 2020;101(1):61.
Lacher TE, Davidson AD, Fleming TH, Gómez-Ruiz EP, McCracken GF, Owen-Smith N, Peres CA, Vander Wall SB. The functional roles of mammals in ecosystems. J Mammal. 2019;100(3):942–64.
Guo YY, Wang SS, Wang X, Liu W, Xu D. Rodents inhabiting the Southeastern Mu us Desert May not have experienced prolonged heat stress in summer 2022. Animals. 2023;13(13):2114.
Ghazian N, Zuliani M, Lortie C. Micro-climatic amelioration in a California Desert: Artificial Shelter Versus Shrub Canopy. J Ecol Eng. 2020;21(8):216–28.
Grossman BF, Hayward MW, Gibb H. An experimental test of the multi-scalar impacts of digging mammal reintroductions on invertebrate burrows. Soil Biol Biochem. 2019;132:101–10.
Jiang L, Wang X, Li L, Shi Z, Yang X. Spatial association of shrubs and their interrelation to burrowing site preference of subterranean rodents on dune slope in the Otindag Sandy Land, China. Sustainability. 2017;9(10):1729.
Machicote M, Branch LC, Villarreal D. Burrowing owls and burrowing mammals: are ecosystem engineers interchangeable as facilitators? Oikos. 2004;106(3):527–35.
Parmenter RR, MacMahon JA. Factors determining the abundance and distribution of rodents in a shrub-steppe ecosystem: the role of shrubs. Oecologia. 1983;59(2–3):145–56.
Esque TC, DeFalco LA, Tyree GL, Drake KK, Nussear KE, Wilson JS. Priority Species Lists to Restore Desert Tortoise and Pollinator Habitats in Mojave Desert Shrublands. Nat Areas J [Internet]. 2021 Apr 22 [cited 2024 Mar 22];41(2). https://bioone.org/journals/natural-areas-journal/volume-41/issue-2/043.041.0209/Priority-Species-Lists-to-Restore-Desert-Tortoise-andhttps://doi.org/10.3375/043.041.0209.full.
Acknowledgements
The authors are grateful to the Bureau of Land Management for allowing us to conducted burrow surveys at several locations across California; and to York University for supporting and facilitation the conduction of this study.
Funding
The study was funded both by York University’s Academic Excellence Fund for EO and through the Natural Sciences and Engineering Research Council of Canada (NSERC) awarded to CJL. The funding bodies did not have any role in the design of the study; collection, analysis, or interpretation of the data; or writing of the manuscript.
Author information
Authors and Affiliations
Contributions
EO, MZ, and CJL conceptualized and designed the study. EO, MZ, and CJL collected and shared data. MZ and CJL cleaned and analyzed data. EO wrote the first draft of the manuscript. EO, MZ, and CJL thoroughly edited and reviewed final the version of the manuscript. All authors provide final manuscript review and revisions.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Owen, E., Lortie, C.J. & Zuliani, M. Native shrub densities predict burrow co-occurrence patterns in Central California Drylands. BMC Ecol Evo 24, 68 (2024). https://doi.org/10.1186/s12862-024-02259-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12862-024-02259-6