
Abstract
Background
Information on pollen dispersal is essential for the risk assessment and management of genetically modified organisms (GMOs) such as Bt maize. We analyzed data on maize pollen deposition at 216 sites in Germany, Switzerland, and Belgium from 2001 to 2010. All data were collected using the same standardized sampling method. The distances between sampling site and the nearest maize field ranged from within the field to 4.45 km.
Results
Maize pollen deposition was negatively correlated with distance from the nearest pollen source. The highest pollen deposition was within the field, but depositions of several thousand pollen grains per square meter were recorded over the kilometer range. A power function model most accurately described the relationship between deposition and distance from the nearest pollen source, rather than the exponential model currently used in EU risk assessment and management, which underestimates exposure for distances greater than 10 m. Regression analysis confirmed the high significance of the power relationship. The large variation in pollen deposition at a given distance reflected the influences of wind direction and other meteorological and site conditions. Plausible variations of single values and the predicted mean pollen count at a given distance were expressed by confidence intervals.
Conclusions
The model described here allows estimations of pollen deposition in relation to distance from the nearest field; therefore, it will be valuable for the risk assessment and management of GMOs. Our results indicate that buffer zones in the kilometer range are required to prevent harmful exposure of non-target organisms to GMOs.
Similar content being viewed by others
Background
Wind pollination is a successful strategy for many plants, and pollen can be transported over long distances. Many agricultural or biological questions relate to gene flow mediated by pollen, and information on gene flow is crucial to understand introgression and seed purity. However, it is important to distinguish between work carried out on gene flow (including cross pollination) and that carried out on pollen deposition itself. This is also true for assessing genetically modified (GM) crops. With GM crops, gene flow may lead either to introgression into wild relatives [1]-[3] or to the presence of GM material in seeds, harvested materials, feed, or foodstuffs. Because there is a legal obligation for GM and GM-free agriculture to co-exist in Europe [4],[5], much research has been carried out to understand and manage gene flow from GM crops to other crops of the same or closely related species [6]-[13].
Pollen dispersal, independent of its ability to facilitate vertical gene flow, has been addressed in many studies related to human health, especially pollinosis. Consequently, many aerobiological studies have focused on airborne pollen. Most countries have networks to systematically monitor for the presence and prevalence of allergenic airborne pollen. However, the samplers used in this type of monitoring are commonly located on the tops of buildings in cities or villages, and the analysis focuses on those types of pollen that trigger allergies. For this reason, such data are of limited value to assess pollen deposition from agronomic crops like maize.
Information on pollen deposition is indispensable for the risk assessment of GM crops containing novel and/or toxic proteins. Because pollen grains are carried by wind into the surrounding landscape, quantitative data on pollen deposition are needed to estimate the exposure of non-target organisms that may consume the pollen deposited on their host plants. To date, almost all insect-resistant GM crops contain insecticidal proteins derived from the soil bacterium Bacillus thuringiensis (Bt). Different toxins may be produced against pest species in the Lepidoptera or Coleoptera.
Despite the assumption that Bt toxins are highly specific, many of these toxins actually show cross-order activity [14],[15]. Several Bt toxins have been reported to affect non-target taxa [16]-[21], especially non-target Lepidoptera [22]-[28].
There are increasing numbers of Bt crops that express more than one Bt toxin. Stacking of Bt genes results in increased environmental exposure to Bt toxins and, thus, in increased risks of effects on non-target organisms. Second- and third-generation GM crops, which may produce pharmaceutical or industrial compounds, are expected to add to this problem. Because of the increasing abundance and range of GM crops, it is even more important to collect background data on pollen deposition and exposure for appropriate risk assessments of genetically modified organism (GMOs).
Maize belongs to the anemophilous (wind-pollinated) grass family and produces vast amounts of pollen. Depending on the variety and growing conditions, each plant releases from approximately 5 million to 50 million pollen grains per season. Assuming an average density of 7 to 12 plants/m2, a 1-ha maize field sheds approximately 1011 to 1013 pollen grains over the flowering season [29]-[32].
Maize pollen is relatively large and heavy with a diameter ranging between 80 and 125 μm. Its settling velocity in calm air ranges from 0.15 to 0.4 m/s, with an average of approximately 0.2 m/s [33],[34]. Assuming a mean height of 2.5 to 3.5 m for a maize plant with its tassel at the top, maize pollen would settle on the ground after approximately 10 to 20 s in calm air. Considering this time, and an average wind speed of 2 m/s (e.g., that in Germany in summer), maize pollen should be deposited in the range of 20 to 40 m on average. This may be one reason why some researchers focused on short distances of a few meters from the field edge when measuring maize pollen dispersal [28],[35]-[38].
However, data from field measurements have led us to question these assumptions. Several studies have shown that maize pollen grains are dispersed over larger distances and in larger amounts following a power function of distance with a long tail, indicating long-distance pollen transport [29],[32],[39]-[43]. Also, certain factors that had been overlooked in earlier studies were reconsidered. For example, turbulence, heat-induced up-drifts, and the stronger winds that typically occur during the maize flowering period in summer can enhance the release and dispersal of maize pollen. These factors correlate with the warm, windy, dry weather conditions during the daytime that trigger the maize pollen release [29],[31],[32],[39],[40],[44],[45].
A major difficulty when assessing pollen dispersal and deposition from data in the literature is that different and/or non-standardized methods have been used in various studies. This makes it difficult, if not impossible, to compare the results.
In this study, we analyzed data on maize pollen deposition obtained using the pollen mass filter (PMF) passive aerosol particle sampler over 10 years of field measurements at 216 sites in Germany, Switzerland, and Belgium. The methodology used has been standardized according to the VDI standard 4330-3 (which gives the general norm for all pollen types) [46] with the specifications for maize pollen described in [47]. This guarantees comparable data. The aim of the study was to analyze the data on maize pollen deposition in relation to distance from the nearest maize field. This reflects the basic rule applied in regulations based on minimum distance requirements (i.e., isolation buffer zones). The variability of data represents maize pollen deposition under common cultivation conditions in Central Europe. The information presented here substantially extends the database and conclusions of an earlier report [47].
Methods
Standardized pollen sampling
There are many methods for pollen sampling, ranging from simple sticky traps such as slides or Petri dishes covered with an adhesive like Vaseline to more sophisticated devices such as high volume samplers with automatic pollen detectors [48]. The methods are usually tailored to meet the specific requirements of the research area. Simple methods like sticky microscope slides or Petri dishes are cheap, simple, and widely used [36],[49], but they are limited to semi-quantitative measurements of pollen deposition [50],[51]. For quantitative assessments, more sophisticated methods are required. Aerobiological sampling methods, their applications, and their advantages and disadvantages have been systematically reviewed by Gregory [50] and Edmonds [51], and sampling methods for pollen monitoring have been reviewed by Giesecke et al. [52]. For general aspects of aerosol measurement, we refer to Baron and Willeke [53], while the relevant methods have been reviewed by McMurry [54].
In this study, maize pollen sampling was performed using the PMF passive aerosol sampler. This method was developed in 2000 to trap airborne pollen for GMO monitoring [55]. The complete method from sampling to analysis has been validated and standardized for the measurement of pollen deposition by the Association of German Engineers [46]. Being a passive sampler, the PMF needs no power supply and can be exposed continuously for more than 4 weeks. The method generates a single number that quantifies total pollen influx over the whole exposure time (flowering period). Because this method is standardized, pollen depositions can be compared directly among different locations or years. The method is suitable for many agricultural crops.
In the standard version, the passive sampling system consists of two passive sampling units: the PMF and the Sigma-2 (Figure 1). The Sigma-2 consists of a sheltered vessel that contains an adhesive slide or foil, which is protected from rain. This slide/foil is used for direct quantitative microscopic analysis of the deposited pollen grains. The PMF has a higher sampling efficiency than the Sigma-2 and collects larger quantities of pollen that can be used for additional analyses; for example, molecular analyses such as polymerase chain reaction (PCR) of pollen DNA or enzyme-linked immunosorbent assay (ELISA) for proteins and toxins [41],[55],[56]. The PMF sampler contains a filter unit consisting of eight stratified polypropylene filter ronds, which have low aerodynamic resistance to the ambient air passing through them. Aerosol particles larger than 10 μm, such as pollen, are retained in the filter. Pollen grains washed off by rain are collected in a 1.5-L PET flask connected to the body of the PMF sampling unit. While the Sigma-2 is restricted to particle sizes with an aerodynamic equivalent diameter of smaller than 60 μm [57], the PMF unit is well suited to capturing larger pollen grains (e.g., maize pollen, 80 to 125 μm). Therefore, it has been used for the quantitative microscopic pollen analyses and assessment of maize pollen deposition in the studies that produced the data reported here [41],[47]. The standard exposure height of the PMF sampling unit is 1.8 m above the ground.

Sampling sites and exposure time
We analyzed data on maize pollen deposition collected from 2001 to 2010, at 216 sampling sites in Germany, Switzerland, and Belgium (Table 1, Figure 2). The sampling sites used in this study relate to different research and monitoring projects and reflect the heterogeneity of common maize cultivation (different field sizes, field arrangements, varieties). They also reflect variations in environmental conditions (regional, topographical, and meteorological conditions), as well as different positions relative to the nearest maize field and different wind directions. For all sampling sites, the distance to the nearest maize field was recorded.

Location of 216 sample sites in Germany, Switzerland, and Belgium. Dot size indicates sample size (see Table 1).
Daily pollen shedding values and, consequently, the pollen deposition values vary greatly depending on meteorological and growing conditions. Therefore, to characterize the intensity of pollen deposition for any season and site in a comparable way in a single figure, the total accumulated deposition over the maize flowering period must be taken as an appropriate parameter [52],[60]-[62]. Because pollen production and maize pollen release vary considerably over time, the accumulated pollen deposition over the whole flowering period is the best measure of the overall intensity of pollen deposition at each site. This parameter is more reliable when based on the whole flowering period, rather than on shorter time intervals. For all data presented here, the exposure time of the PMF samplers covered the main maize pollen flowering period in each particular area (3 to 5 weeks and sometimes longer). Whereas any calculated mean deposition rate based on a fixed time span (daily, weekly, or other) would depend on the number of days of exposure, the total accumulated deposition covering the whole flowering period is a more stable and comparable quantity.
Sample preparation
All deposition data analyzed in this study were obtained in the same way according to the VDI standard 4330-3 [46] specific for maize [47] as follows: the pollen samples were removed from the filter pads of the PMF and from the liquid in the 1.5-L attached flask by using an ultrasonic bath (Sonorex Bandelin Super RK 102, BANDELIN electronic GmbH & Co. KG, Berlin, Germany) and vacuum filtration (Sartorius, Göttingen, Germany). These procedures sedimented the aerosol particle sample as a filter cake on an acetyl-ethylene membrane filter (12 μm pore size, 50 mm diameter, Whatman, Maidstone, UK), after pre-filtering the liquid through a 125-μm metal mesh test sieve (100 mm diameter, Fritsch, Idar-Oberstein, Germany). Sample quality was checked by examining the filter cakes under a binocular microscope. A sample from each filter cake was transferred into a 15-ml tube. After centrifugation (500 × g for 5 min), the volume was reduced to 2 ml. Then, 2 ml glycerol was added to give 4 ml 50% glycerol pollen suspension (density 1.1 g/cm3). The pollen suspension was mixed with a shaker and stored at −20°C until subsequent analysis.
Quantitative microscopic pollen analysis
The pollen suspensions were carefully mixed, and subsamples were taken to determine the number of maize pollen grains per PMF sample microscopically. Pollen was quantitatively analyzed using the dynamic counting method [41],[46],[47] under × 400 magnification using a Nikon Eclipse E200 microscope (Nikon, Tokyo, Japan).
The absolute detection limit for maize pollen counting is determined by the discrete number of one detected pollen grain in the sub-sample for counting. The relative detection limit of maize pollen depends on the total number of pollen grains counted. The dynamic counting method ensures comparable results of maize pollen counting; the detection limit remains in the range of <0.1% of the total pollen count, relatively and absolutely, until the maize pollen count is greater than 3 at least. This is achieved by adjusting the volume of the sub-sample to be analyzed until approx. 3,000 total pollen grains were counted and maize pollen grains were detected. Using a micropipette, sub-samples (volume, 10 to 50 μl) were taken from the glycerol pollen suspension and the maize pollen grains and total pollen grains were counted. Several subsamples with corresponding volumes were subsequently taken and quantified. The cumulated pollen counts of the sub-samples were then extrapolated to the total sample, giving the maize pollen count per PMF sample (Ni).
Determination of maize pollen deposition
From the maize pollen counts in the sample (Ni), the pollen flow (Fi) and deposition (Di) of maize pollen were determined as described by Hofmann [47] based on VDI 4330-3 [46]. The PMF measurement represents the horizontal pollen flow [62]a. Horizontal pollen flow is defined as the number of pollen grains transported by wind through a given cross-sectional area of 1 m2 during the entire exposure time (in this case, the maize flowering period). Pollen deposition is defined as the number of pollen grains deposited on an acceptor surface of 1 m2 during the exposure period, here obtained using a standardized deposition measurement method. Because deposition depends on the acceptor's surface conditions, standardized measurements must be used for data to be comparable. The relevant cross-sectional area of the filter is 8 × 10 cm (80 cm2). The horizontal maize pollen flow in the air was calculated using Equation 1 below, taking the PMF sampling efficiency into account:
where:
Fi, is the horizontal flow for pollen of species i in air (n/m2), N i is the number of pollen grains in the sample, i is the pollen from species i; here, maize pollen, Ei, PMF is the sampling efficiency of the PMF (0.35) for maize pollen compared with the volumetric standard pollen trap (Hirst type/Burkard).
The sampling efficiency depends on both the collector type and the characteristics and behavior of the aerosol particle, in this case, the pollen type. The average sampling efficiency of the PMF for maize pollen was estimated to be 0.35 by calibration to a Hirst-type volumetric pollen trap [63], the standard trap used by the European and German Aerobiology Networks.
The deposition of aerosol particles depends on the physical behavior of the particles in the atmosphere and the interaction with the surface of the acceptor [50],[51],[64]. For data to be comparable, the acceptor must be standardized [50],[51],[65]. For large aerosol particles such as maize pollen, gravitational sedimentation is the main factor in deposition when the receptor is an inert horizontal surface [50],[51]. The standardized deposition of maize pollen on a planar inert surface of 1 m2 is calculated from horizontal pollen flow, taking into account the mean wind speed and the deposition velocity of maize pollen [50] as expressed below:
where:
Di is the accumulated total deposition of pollen species i over the flowering period, u is the mean wind speed, and vd,i is the mean deposition velocity for maize pollen (m/s).
The values for deposition velocity of maize pollen reported in the literature range between 0.15 and 0.4 m/s, with mean values around 0.2 m/s [33],[34]. In this study, we assumed an average deposition velocity for maize pollen of 0.2 m/s and an average wind speed of 2 m/s, which is typical for July/August in Germany.
Statistical analysis
For the statistical analyses of the data, we used Microsoft Excel® [66], the statistic tool XLSTAT-Pro 2011 [67], SegReg [68], and R software [69], specifically the mcgv package [70].
A necessary property of an adequate statistical model for the relationship between distance from the pollen source (X) and pollen deposition (Y) is that the model produces predictions that are non-negative and that tend towards zero for large distances. Simple models with these properties include the exponential model Y = eb+c·X, c <0, used for risk analysis in the EU [71]-[75], and the power model Y = a · Xm with m <0, used by Aylor et al. [29], Arrit et al. [39], and Boehm et al. [44]. Both models can be linearized to facilitate calculations. If the data follows a power model, then log10Y and log10X are linearly related (log10Y = log10a + m · log10X), with the corresponding plot showing points scattered around a straight line. If the data follows an exponential model, then log10Y and X are linearly related (log10Y = b · log10 e + c · log10 e · X), and a plot of log10Y versus X shows a linear relationship. Standard linear regression techniques were applied to (log10Y, log10X) and (log10Y, X) to obtain optimal (least squares) estimates for the regression parameters (a, m) and (b, c), respectively, together with confidence intervals for the parameters and predicted values. Residual plots were used to check the homoscedasticity of the error variance. Logarithms with base 10 were used in all cases. The goodness of fit of the models was expressed by the coefficient of determination, R2, which was also used as the criterion to select the optimal model. The consistency of the optimal model, with regard to a potential change in the model characteristics over distance, was checked by fitting a segmented linear regression (the ‘hockey stick approach’), using the open source statistical program SegReg [68]. To detect a nonlinear component in the deposition-distance relationship beyond one that might be identified by a segmented linear regression, a smooth curve based on a regression spline approach was fitted to the residuals of the optimal model using the ‘gam’ model of the mgcv package in R [69],[70]. This type of curve would uncover any reasonably smooth nonlinear component in the relationship. Because the complete dataset was obtained over a long period (2001 to 2010), a test for a change in the deposition-distance relationship over time was performed by comparing the regression line from data collected in 2001 to 2006 (122 samples) [47] with that for deposition data collected in 2007 to 2010 (94 samples). In Figure 3, the data values from these two periods are shown in different colors. The hypotheses tested were H0: ‘common regression line for both sampling periods’ against the alternative HA: ‘specific regression line per sampling period’. An F test was used for the comparison. For illustrative purposes, a linear and a cubic polynomial were fitted to the untransformed data, although these approaches do not meet the above requirements of generating non-negative predictions that tend towards zero for large distance.

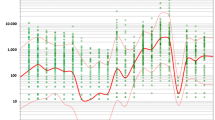
Maize pollen deposition versus distance to the next maize field. Data points from field measurements 2001 to 2010 (gray/dark, data from 2001 to 2006; light blue/light, data from 2007 to 2010; N =216) and results of regression analysis (linearized power regression equation log10Y = −0.585 · log10X + 6.104, R2 = 0.709, p <0.001; power regression equation, Y = 1.271 · 106 · X−0.585; Y, deposition in n/m2; X, distance to nearest maize field in m). Central bold blue line, expected deposition; dashed lines, confidence interval for expected deposition; red solid lines (outer lines), 95% confidence intervals for single predictions. Deposition values in n/cm2 were obtained by multiplying displayed deposition values by 0.0001.
Results and discussion
Pollen deposition and distance relationship
Pollen deposition was measured at 216 sites with distances to the nearest pollen source ranging from 0.2 m (within the field) to 4.45 km away from the nearest edge of the maize field. The values of maize pollen deposition varied from 23.3 million pollen grains/m2 (2,330 pollen/cm2) within the field to 2,857 pollen grains/m2 at further distances from the field.
The amount of maize pollen deposition over the main flowering period decreased markedly with increasing distance. Nevertheless, the spatial distribution exhibited a long-tailed shape, with pollen detected as far as 4.45 km away from the nearest field. Comparing the fits of the power model and the exponential model, the relationship between maize pollen deposition and distance to the next maize field was fitted best by the following power function:
where:
Y is the deposition of maize pollen (n/m2), X is the distance to the nearest maize pollen source/field edge [m], and n is the number of pollen grains.
The above power function was highly significant (p <0.001) with distance accounting for 70.9% of the total variation in deposition (R2 = 0.709).
Figure 3 shows the data and the fitted curve with confidence intervals on log-log transformed axes. This presentation turns the power relationship between the X and Y variable into a linear relationship. The scale on the left vertical axis denotes pollen deposition in the standard unit (n/m2). The horizontal axis shows the distance to the next maize field, ranging from within the same field (0.2 m) to 4,450 m. Table 2 shows the estimated regression parameters.
Figure 3 shows the good fit over the data range. It also shows that data points are scattered around the regression line with nearly constant variation over the distance range, as required by the assumptions for standard regression analysis. This finding is confirmed by the standardized residuals plot (Figure 4).

Standardized residuals for regression line shown in Figure3. X, distance from nearest maize field in m.
Fitting an exponential model to the data resulted in a much weaker determination coefficient of R2 = 0.222 (p <0.001) compared with the R2 = 0.709 for the power model. The corresponding equation is as follows:
A fitted purely linear regression line has the following equation:
with a coefficient of determination R2 = 0.021 (p =0.035). The least-squares fitted cubic polynomial has the following equation:
with a coefficient of determination R2 = 0.078 (p <0.001). The cubic polynomial shows a slightly better, but nevertheless poor, quality of fit with 7.8% of the variation in Y explained by X. For the linear function, all predictions for distances greater than approximately 2,400 m are negative. For the cubic polynomial, distances between 380 and 1,900 m and greater than 4,400 m generate negative predictions for pollen deposition, interrupted by positive predictions for distances between 1,900 and 4,400 m, with a local maximum at about 3,200 m. The polynomial model for the relation between deposition and distance previously published by Lang et al. [36] (equation: Y = 74.75 − 9.71 · X + 1.32 · X2 − 0.08 · X3) was similarly inacceptable, predicting negative deposition for all distances greater than 13 m. These model fits are numerical examples of the fact that (non-constant) polynomials tend to plus or minus infinity for large distances instead of converging to zero, as is required for a reasonable model. Even worse, as demonstrated by the third-order polynomial, high-order polynomials can exhibit erratic behavior outside the X data range and even in the observed X range.
Consistency of the power regression over the distance range and sampling periods
A segmented linear regression showed no improvement of fit compared with that of a single linear regression line on a log-log scale over the whole distance range, from within the same field to 4 km (p >0.050). The improvement in the goodness of fit after adding a nonlinear smooth component to the power model was practically zero (p =1), and the R2 value was increased by only 0.00014. For this reason, and because neither the graphical check of the regression line (Figure 3) nor the residual plot (Figure 4) suggested deviations from linearity, the power model was accepted and no further nonlinear models for the distance effect were investigated.
The regression equations were not significantly different between the two sampling periods of 2001 to 2007 and 2008 to 2010 (F test; p =0.107).
Summary of model selection
The results from the goodness of fit calculations and the consistency checks confirmed the superiority of the power model (Equation 3) over the exponential model (Equation 4) and confirmed the validity of the power model for pollen deposition over the whole distance range examined.
Several previous studies [29],[39],[42],[44] have arrived at the same conclusion, namely, that the decrease in pollen deposition with increasing distance follows a power function. The exponential model used in some studies [37],[76],[77] and the polynomial model used in others [36] are not appropriate in the light of the present dataset because of their poor fit and their implausible predictions of pollen behavior over large distances.
The difference between the power function and the exponential function fitted to our data lies markedly in the curve tail, where the exponential curve underestimates pollen deposition, especially at long distances. This raises questions about the accuracy of some risk assessments of genetically modified plants in the EU based on an exponential relationship [71]-[75].
The reason why different authors proposed different curve shapes for the relationship between deposition and distance may be explained by the different distance ranges of the data used in the analyses. Looking at the data from a narrow distance range, a power function, an exponential function, a polynomial, or even a straight line may fit the data quite well. However, a model derived from short distance range data of a few meters cannot reasonably be extrapolated and expected to fit long-distance data. The exponential curve for pollen deposition used in the EU risk assessment is given in Perry et al. [71], page 4, equation 2.3, 2.4, and related text], based on the data of Wraight et al. [76]. Amending the stated factors, the equation for the exponential curve of maize pollen deposition on slides (n/cm2) versus distance (m) is log10Y = 2.368 − 0.145 · X. The dataset used was limited to a distance range of 7 m from the field edge. In this range, both the exponential model of Perry et al. and the power function derived here produce acceptable predictions. However, as illustrated in Figure 5, extrapolating the exponential curve to distances greater than 10 m leads to a rapidly descending curve with increasing discrepancies from field measurement data and from the respective power regression line.

Exponential model of EU risk assessment compared with power model described in this study. For maize pollen deposition versus distance to nearest maize field. Circles, data points from field measurements 2001 to 2010 (N =216). Blue solid line, expected mean deposition calculated using power regression equation Y =1.271 · 106 · X−0.585; Y, deposition in n/m2; X, distance from nearest maize field in m; dashed lines, confidence interval for expected deposition; dotted lines, 95% confidence intervals for single predictions; red line, exponential model used in EU risk assessment (Perry et al. [71], deposition values on slides used in equation 2.3 corrected for stated factor 3); solid, predictions within database up to 7 m from field edge; dashed line, extrapolation of exponential model over distance >7 m.
Mean regression and confidence intervals
The power regression line (Equation 3) provides estimates for the mean expected pollen deposition at given distances from the pollen source. The mean values predicted from the regression equation range from 3.26 million pollen grains/m2 (326 pollen/cm2) close to the pollen source to 9,340 pollen grains/m2 (0.934 pollen/cm2) at 4,450 m away from the next field margin (see also Table 3). The uncertainty of this prediction is expressed by the 95% confidence interval (CI) of the mean regression line (Figure 3, fine dotted lines).
Also shown in Figure 3 is the 95% CI for single observations (solid red lines). This interval describes the range in which a single observation is expected to lie with a probability of 95%. The upper boundary of a single value CI at a given distance indicates the deposition, which is exceeded by a single observation with a probability of no more than 2.5%. Vice versa, to find the distance at which the exposure can be expected to lie below a defined level, while accounting for variations in the data, the intersection between the upper confidence limit and a horizontal line at the defined level must be determined. Table 3 shows predicted values for maize pollen deposition and their confidence intervals for selected distances on the usual linear scale. The standard deviations of the confidence intervals were originally calculated on the logarithmic scale (sd_log), but are shown in Table 3 as 10sd_log so that they are on the same scale as the pollen counts. It must be emphasized, however, that 10sd_log is not the standard deviation of a Y prediction, neither for mean nor for single values. In both cases, for a given X, the relationship between 10sd_log and the confidence interval (CIlow, CIhigh) on the linear scale is expressed as follows:
where the constant c depends on the number of observations and the coverage probability of the confidence interval. In contrast to the usual form of a confidence interval (‘mean value ± constant c’), here, the multiplicative forms ‘mean value/factor c’ and ‘mean value · factor c’ apply. For the present data, with n = 216 and a coverage probability of 95%, c = 1.97 holds for mean and single value predictions. The way to calculate sd_log differs for mean and single value predictions. Details are given in standard texts on linear regression, e.g. Neter et al. [78] chapter 2].
As an example, Table 3 shows that at the closest distance (within the maize field), the mean value for maize pollen deposition is expected to be 3,258,000 pollen grains/m2 with a confidence interval of (2,462,000; 4,311,000) pollen grains/m2. This confidence interval can be described as (mean prediction/1.32; mean prediction × 1.32). While this consideration is for mean values, the numbers for single observations are quite different. The factor for the confidence interval of a predicted single observation is 7.33, indicating large variations among single values. This means that overall, 95% of maize pollen deposition values from single measurement sites within the field vary between the lower boundary (444,000 n/m2) and the upper boundary (23,876,000 n/m2). Half of the remaining 5% (2.5%) will be above the upper boundary, so the probability of values not exceeding the upper boundary will be 97.5%. At 1,000-m distance, single observations of maize pollen deposition are expected to be between 3,100 and 164,000 n/m2 with a mean of 22,500 n/m2 (2.25 n/cm2). Consequently, a distance of 1,000 m would be necessary to exclude a maize pollen deposition higher than 164,000 n/m2 (16 n/cm2) with a probability of 97.5% for distinct sites.
Dispersal range
In Germany, maize flowers in summer (July to August) and pollen release is favored by warm and windy (i.e., drying and turbulent) weather conditions during the daytime [29],[45],[59],[79],[80]. In turbulent wind conditions, pollen grains are transported higher above the ground and are dispersed over further distances than they are under non-turbulent conditions [29],[32],[39],[40],[42],[44],[59],[80]. Pollen dispersal itself varies according to wind speed and direction, other climatic conditions, topography, and factors that affect the settling velocity of maize pollen, for example, dehydration of the pollen grain [33].
A considerable portion of our dataset represents the longer distance range (>100 m), which has been underrepresented in the literature so far. Compared with that stated in our earlier report [47], the maximum distance for which pollen data is reported is extended from 3.3 to 4.45 km with a total of 81 data points representing distances greater than 100 m.
The results shown in Figure 3 illustrate that maize pollen dispersal is not restricted to close distances (<100 m), but extends well beyond to longer distances up to the kilometer range. This is consistent with the findings of other studies [29],[39]-[42],[44].
Maize is a wind-pollinated plant and produces enormous amounts of pollen (1011 to 1013 pollen grains/ha). Maize pollen is relatively large (80 to 125 μm diameter; 1.25 g/cm3; approx. 500 μg) and its settling velocity in air is approximately 0.2 m/s. Assuming that the average height of the maize tassel is 2 to 3.5 m above the ground, we can estimate that maize pollen settles in the range of 20 to 40 m from the field margin on average. However, this assumption would be only true under still air conditions. In the field, still air conditions are practically non-existent. As our data from real environmental conditions (Figure 3) show, maize pollen was deposited even at the farthest distance measured (4.45 km). In fact, considerable amounts of maize pollen drift on the wind over longer distances (>100 m) and even further than 1,000 m, with deposition values of 3,000 to 164,000 maize pollen grains per square meter (Figure 3). This is consistent with observations of long-distance maize pollen dispersal in other studies [29],[39],[42]. Brunet et al. [40] observed that maize pollen was dispersed at even higher altitudes of several km and was transported over distances as far as 70 km.
The database includes distances up to 4.45 km, which covers a distance range relevant to questions related to Bt maize dispersal. The respective power function for the distance relationship expressed in Equation 3 is highly significant within the distance range of the database. We recommend not to extrapolate to distances far beyond without specific validation. One possible estimation method would be to combine the use of the deposition database with an appropriate dispersal model. The standardized PMF deposition data serve to calibrate the model within the data range, while model predictions may be used to extrapolate over larger distances [32],[80]. These predictions are open to validation based on further standardized measurements.
Pollen deposition under common cultivation
Under common agricultural practice, maize is rarely grown in isolated fields. The pollen deposition of several fields is expected to overlap, and more than one pollen source contributes to exposure at any single site. The potential overlap with other surrounding fields, even if they are distant, should be considered as a factor affecting the distance relationship to the nearest maize field, especially for distances greater than 100 m. This assumption has been supported in various scenarios using a dispersal model [32],[59],[80]. The relative proportion of overlap will depend on several factors, such as distance, the relative position of other maize fields, wind direction, other meteorological conditions, and the flowering behavior in each field. Therefore, it is difficult or not feasible to extrapolate from single field experiments to the complex situations that exist in common cultivation, that is, those situations relevant for the risk assessment and management of GMOs.
The large number of locations and years represented in our dataset covers a broad range of environmental and agricultural conditions. In fact, the data used in our analyses reflect the variable conditions of common maize cultivation in central European countries such as Germany. The data cover small, single-field settings as well as complex field arrangements with no preferences for field size or shape, variety, topography, nor for the relative location of the pollen trap to the main wind direction nor to other maize fields. The distance relationship expressed in Equation 3, therefore, represents the expected values of maize pollen deposition under common agricultural practice in the studied countries.
The reference to common agricultural practice is important when interpreting the variability in deposition under real field conditions. As described before, the results of the regression analysis show a relatively wide confidence interval for individual values (see Figure 3, Table 3). This can be expected, as our data include variability in pollen production, release, and dispersal resulting from different relative positions of the traps to the main wind direction, and other factors such as field size, plant density, maize variety, growing conditions, agricultural management, and weather conditions [29],[45].
Consequences for risk assessment and management of Bt maize
The power curves and exponential curves fitted to our data showed different shapes over the distance range. Our results indicate that the exponential model previously used by some authors and currently used for risk assessment and management of Bt maize [37],[71]-[77] is inferior to the power function in terms of goodness of fit. It underestimates deposition, and thus underestimates the exposure of non-target organisms to GMO maize pollen for distance ranges greater than 10 m, with increasing inaccuracy over longer distances (see Figure 5).
The toxin concentration varies among different Bt maize varieties [81],[82], and the various target insect species and their different larval instars show variations in their sensitivity to the toxin [83]. Therefore, in risk assessment, event and species-specific variations should be considered when establishing dose-effect relationships between Bt maize pollen and sensitive species. Nevertheless, it is not practical to implement specific legal regulations for any single event. Therefore, more general measures based on the precautionary principle are required for risk management of Bt maize cultivation.
Bt maize pollen, even at low pollen densities, adversely affects sensitive Lepidoptera species. For example, Felke et al. reported that the LD50 value of Bt maize pollen (Bt-176) for the fourth larval instar of Plutella xystostella could be as low as nine Bt maize pollen grains/cm2 (92,000 n/m2) [24], and that a single uptake of four or more Bt pollen grains was sufficient to kill larvae, with earlier instars being even more sensitive [83],[84].
The current recommendation of the event-specific EU risk assessment for Bt maize states, for example, buffer distances of 20 to 30 m between Bt maize and protected habitats of extremely sensitive butterfly species [73]-[75]. These recommendations have been based on an exponential curve for exposure assessment (see Figure 5 and related discussion above). The results of the present study indicate that these buffer zones may be inappropriately small. Instead, conclusions on risk assessment and risk management should be updated based on the model described here for the distance relationship of maize pollen deposition under common cultivation.
With respect to a general risk management of Bt maize cultivation, and based on the precautionary principle, the upper boundary of the confidence interval of the regression for single value predictions indicates that buffer distances in the kilometer range are required to prevent exposure of protected and/or sensitive species to Bt pollen, rather than ranges of tens of meters as proposed in the actual EU risk management.
Conclusions
To our knowledge, the results presented here represent the largest dataset on maize pollen deposition under complex common cultivation conditions published to date. The data cover variations in pollen deposition related to the use of different crop varieties, field sizes, field arrangements, wind directions, regional environments, and years. A considerable proportion of the data represents a longer distance range (>100 m), which has been lacking in the literature so far. This dataset has a high informative value because all data were collected using the same standardized sampling method, the passive sampling system PMF, according to VDI standard 4330-3 [46] specific for maize pollen [47]. Therefore, the pollen deposition data are comparable among different locations and years.
The results show that under common cultivation, maize pollen is deposited in substantial quantities (thousands of grains per m2) over a range of up to several kilometers away from the next maize field. The distance relationship significantly fits to a power function, reflecting the long tail of pollen deposition over larger distances.
Our results indicate that the exponential model previously used by some authors and currently used for risk assessment in the EU [37],[71]-[77] is inferior to the power function in terms of goodness of fit. The power and the exponential curves fitted to our data have different shapes over the distance range. The exponential curve used in the EU risk assessment underestimates deposition and, thus, underestimates exposure of non-target organisms, especially over a longer distance range. Consequently, previous risk assessments and conclusions regarding distances, potential exposure, and effects on non-target organisms should be revised in the light of these findings.
The regression model presented here allows us to predict expected deposition at a given distance. Its confidence intervals for mean and single observations provide information about the variation that can be expected to result from different site/field positions and various meteorological and growing conditions. Therefore, the results are particularly useful to optimize sample size and position when designing field studies and monitoring programs [41],[59],[80]. The standardized sampling method and its results can be used to validate models of maize pollen dispersal and exposure on local and larger scales.
Finally, the results of this study are valuable for risk assessment, risk management, and environmental monitoring of GM maize. These findings will be especially useful for exposure assessments, for example, for tasks related to pollen dispersal and exposure such as co-existence and seed purity [11],[56],[85],[86], and to define buffer zones between GM maize fields and ecologically sensitive areas [41],[59],[80]. These results will be also valuable to estimate the exposure of non-target organisms to Bt maize [59]. In general, these results emphasize the need to collect exposure data using standardized methods for environmental risk assessments and monitoring of GMO [65],[87],[88].
Endnote
aThe equivalent physical term would be ‘accumulated horizontal flux density (rate)’ [62]. We used the term ‘flow’ in this study for simplicity.
Authors’ contributions
All authors made substantial contributions to the published study and drafted the manuscript. MO defined the conceptual framework and task, FH and WW designed and performed the study, FH contributed and/or collected the deposition data and compiled the dataset, and WW and FH conducted statistical analyses and produced the graphic representations. All authors read and approved the final manuscript.
Abbreviations
- GMO:
-
genetically modified organism
- GM:
-
genetically modified
- PMF:
-
pollen mass filter, a passive aerosol sampler according to VDI standard 4330-3
References
den Nijs HCM, Bartsch D, Sweet J: Introgression from genetically modified plants into wild relatives. CABI Publishing, Wallingford; 2004.
Ellstrand NC, Prentice HC, Hancock JF: Gene flow and introgression from domesticated plants into their wild relatives. Annu Rev Ecol Syst 1999, 30: 539–563. 10.1146/annurev.ecolsys.30.1.539
Stewart CN, Halfhill MD, Warwick SI: Transgene introgression from genetically modified crops to their wild relatives. Nature 2003, 4: 806–817.
Regulation (EC) No 1829/2003 of the European Parliament and of the Council of 22 September 2003 on genetically modified food and feed Off J Eur Union L 2003, 268: 1–23.
Regulation (EC) No 1830/2003 of the European Parliament and of the Council of 22 September 2003 concerning the traceability and labelling of genetically modified organisms and the traceability of food and feed products produced from genetically modified organisms and amending Directive 2001/18/EC Off J Eur Union L 2003, 268: 24–28.
Eastham K, Sweet J: Genetically modified organisms (GMOs): The significance of gene flow through pollen transfer. Environ Issue Rep 2002, 28. Copenhagen: European Environment Agency (EEA).
Goggi AS, Lopez-Sanchez H, Caragea P, Westgate M, Arritt R, Clark CA: Gene flow in maize fields with different local pollen densities. Int J Biometeorol 2007, 51: 493–503. 10.1007/s00484-007-0088-5
Knispel AL, McLachlan SM, Van Acker RC, Friesen LF: Gene flow and multiple herbicide resistance in escaped Canola populations. Weed Sci 2008, 56: 72–80. 10.1614/WS-07-097.1
Messean A, Angevin F, Gómez-Barbero M, Menrad K, Rodriguez-Cerezo E: New case studies on the coexistence of GM and non-GM crops in European agriculture.Technical Report Series, EUR 22102 EN. Joint Research Centre of the European Commission, 2006. , [http://ftp.jrc.es/EURdoc/eur22102en.pdf]
Poppy GM, Wilkinson MJ: Gene Flow from GM Plants. Blackwell, Oxford; 2005.
Reuter H, Böckmann S, Breckling B: Analysing cross-pollination studies in maize. In Implications of GM-crop Cultivation at Large Spatial Scales. Edited by: Breckling B, Verhoeven R. Peter Lang, Frankfurt; 2008:59–65.
Rieben S, Kalinina O, Schmid B, Zeller SL: Gene flow in genetically modified wheat. PLoS ONE 2011, 6(12):e29730. 10.1371/journal.pone.0029730
Riesgo L, Areal FJ, Sanvido O, Rodriguez-Cerezo E: Distances needed to limit cross-fertilization between GM and conventional maize in Europe. Nature Biotechnol 2011, 28: 780–782. 10.1038/nbt0810-780
van Frankenhuyzen K: Insecticidal activity of Bacillus thuringiensis crystal proteins. J Invertebr Pathol 2009, 101: 1–16. 10.1016/j.jip.2009.02.009
van Frankenhuyzen K: Cross-order and cross-phylum activity of Bacillus thuringiensis pesticidal proteins. J Invertebr Pathol 2013, 114: 76–85. 10.1016/j.jip.2013.05.010
Bøhn T, Primicerio R, Hessen DO, Traavik T: Reduced Fitness of Daphnia magna Fed a Bt-Transgenic Maize Variety. Arch Environ Contam Toxicol 2008, 55: 584–592. 10.1007/s00244-008-9150-5
Erban T, Nesvorna M, Erbanova M, Hubert J: Bacillus thuringiensis var. tenebrionis control of synanthropic mites (Acari: Acaridida) under laboratory conditions. Exp Appl Acarol 2009, 49: 339–346. 10.1007/s10493-009-9265-z
Hilbeck A, McMillan JM, Meier M, Humbel A, Schlaepfer-Miller J, Trtikova M: A controversy re-visited: is the coccinellid Adalia bipunctata adversely affected by Bt toxins? Environ Sci Eur 2012, 24: 10. 10.1186/2190-4715-24-10
Perez-Guerrero S, Aldebis HK, Vargas-Osuna E: Toxicity of six Bacillus thuringiensis Cry proteins against the olive moth Prays oleae . Bull Insectology 2012, 65: 119–121.
Rosi-Marshall EJ, Tank LJ, Royer TV, Whiles MR, Evans-White M, Chambers C, Griffiths NA, Pokelsek J, Stephen ML: Toxins in transgenic crop byproducts may affect headwater stream ecosystems. Proc Natl Acad Sci USA 2007, 104: 16204–16208. 10.1073/pnas.0707177104
Schmidt JEU, Braun CU, Whitehouse LP, Hilbeck A: Effects of activated Bt transgene products (Cry1Ab, Cry3Bb) on immature stages of the Ladybird Adalia bipunctata in laboratory ecotoxicity testing. Arch Environ Contam Toxicol 2009, 56: 221–228. 10.1007/s00244-008-9191-9
Anderson PL, Hellmich RL, Prasifka JR, Lewis LC: Effects on fitness and behavior of monarch butterfly larvae exposed to a combination of Cry1Ab-expressing corn anthers and pollen. Environ Entomol 2005, 34: 944–952. 10.1603/0046-225X-34.4.944
Dively GP, Rose R, Sears MK, Hellmich RL, Stanley-Horn DE, Calvin DD, Russo JM, Anderson PL: Effects on monarch butterfly larvae (Lepidoptera: Danaidae) after continuous exposure to Cry1Ab-expressing corn during anthesis. Environ Entomol 2004, 33: 1116–1125. 10.1603/0046-225X-33.4.1116
Felke M, Lorenz N, Langenbruch GA: Laboratory studies on the effects of pollen from Bt-maize on larvae of some butterfly species. J Appl Entomol 2002, 126: 320–325. 10.1046/j.1439-0418.2002.00668.x
Jesse LCH, Obrycki JJ: Field deposition of Bt transgenic corn pollen: lethal effects on the monarch butterfly. Oecologia 2000, 125: 241–248. 10.1007/s004420000502
Lang A, Vojtech E: The effects of pollen consumption of transgenic Bt maize on the common swallowtail, Papilio machaon L . (Lepidoptera, Papilionidae). Basic Appl Ecol 2006, 7: 296–306. 10.1016/j.baae.2005.10.003
Losey JE, Rayor LS, Carter ME: Transgenic pollen harms monarch larvae. Nature 1999, 399: 214. 10.1038/20338
Zangerl AR, McKenna D, Wraight CL, Carroll MJ, Ficarello P, Warner P, Berenbaum MR: Effects of exposure to event 176 Bacillus thuringiensis corn pollen on monarch and black swallowtail caterpillars under field conditions. Proc Natl Acad Sci USA 2001, 98: 11908–11912. 10.1073/pnas.171315698
Aylor DE, Schultes NP, Shields EJ: An aerobiological framework for assessing cross-pollination in maize. Agric For Meteorol 2003, 119: 111–129. 10.1016/S0168-1923(03)00159-X
Fonseca AE, Westgate ME, Grass L, Dornbos DL: Tassel morphology as an indicator of potential pollen production in maize. Crop Manage 2003. doi:10.1094/CM-2003–0804–01-RS 20.4.2008.
Fonseca AE, Lizaso JI, Westgate ME, Grass L, Dornbros DL: Simulating potential kernel production in maize hybrid seed fields. Crop Sci 2004, 44: 1696–1709. 10.2135/cropsci2004.1696
Hofmann F, Janicke L, Janicke U, Wachter R, Kuhn U: Modellrechnungen zur Ausbreitung von Maispollen unter Worst-Case-Annahmen mit Vergleich von Freilandmessdaten. Federal Agency for Nature Conservation.. Bonn – Bad Godesberg: 2009. ., [http://www.bfn.de/fileadmin/MDB/documents/service/Hofmann_et_al_2009_Maispollen_WorstCase_Modell.pdf]
Aylor DE: Settling speed of corn ( Zea mays ) pollen. J Aerosol Sci 2002, 33: 1601–1607. 10.1016/S0021-8502(02)00105-2
Di Giovanni F, Kevan PG, Nasr MR: The variability in settling velocities of some pollen and spores. Grana 1995, 34: 39–44. 10.1080/00173139509429031
Gathmann A, Wirooks L, Hothorn LA, Bartsch D, Schuphan I: Impact of Bt maize pollen (MON810) on lepidopteran larvae living on accompanying weeds. J Mol Biol 2006, 15: 2677–2685.
Lang A, Ludy C, Vojtech E: Dispersion and deposition of Bt maize pollen in field margins. J Plant Dis Protect 2004, 111: 417–428.
Pleasants JM, Hellmich RL, Dively GP, Sears MK, Stanley-Horn DE, Mattila HR, Foster JE, Clark TL, Jones GD: Corn pollen deposition on milkweeds in and near cornfields. Proc Natl Acad Sci USA 2001, 98: 11919–11924. 10.1073/pnas.211287498
Stanley-Horn DE, Dively GP, Hellmich RL, Mattila HR, Sears MK, Rose R, Jesse LCH, Losey JE, Obrycki JJ, Lewis L: Assessing the impact of Cry1Ab-expressing corn pollen on monarch butterfly larvae in field studies. Proc Natl Acad Sci USA 2001, 98: 11931–11936. 10.1073/pnas.211277798
Arritt RW, Clark CA, Goggi AS, Lopez-Sanchez H, Westgate ME, Riese JM: Lagrangian numerical simulations of canopy air flow effects on maize pollen dispersal. Field Crops Res 2007, 102: 151–162. 10.1016/j.fcr.2007.03.008
Brunet Y, Foueillassar X, Audran A, Garrigou D, Dayau S, Tardieu L: Evidence for long-range transport of viable maize pollen. In 1stEuropean Conference on the Coexistence of Genetically Modified Crops with Conventional and Organic Crops, 13–14 November 2003, Helsingor, Denmark. Edited by Boelt B. Slagelse: Danish Institute of Agricultural Sciences; 2003:74–76.
Hofmann F, Epp R, Kalchschmid A, Kratz W, Kruse L, Kuhn U, Maisch B, Müller E, Ober S, Radtke J, Schlechtriemen U, Schmidt G, Schröder W, von der Ohe W, Vögel R, Wedl N, Wosniok W: Monitoring of Bt-Maize pollen exposure in the vicinity of the nature reserve Ruhlsdorfer Bruch in northeast Germany 2007 to 2008. Umweltwiss Schadst Forsch 2010, 22: 229–251. 10.1007/s12302-010-0133-6
Loos C, Seppelt R, Meier-Bethke S, Schiemann J, Richter O: Spatially explicit modelling of transgenic maize pollen dispersal and cross-pollination. J Theor Biol 2003, 225: 241–255. 10.1016/S0022-5193(03)00243-1
Lourbet B, Jarosz N, Durand B, Foueillassar X, Huber L: Measured airborne concentration and deposition rate of maize pollen ( Zea mays L .) downwind of an experimental field. In Proceedings of the 25th Conference on Agricultural and Forest Meteorology. Norfolk: American Meteorological Society; 2002:24–25.
Boehm MT, Aylor DE, Shields EJ: Maize pollen dispersal under convective conditions. J Appl Meteorol Climatol 2008, 47: 291–307. 10.1175/2007JAMC1554.1
Viner BJ, Westgate ME, Arrit RW: A model to predict diurnal pollen shed in maize. Crop Sci 2010, 50: 235–245. 10.2135/cropsci2008.11.0670
VDI-Standard: 4330 Blatt 3 Monitoring the effects of genetically modified organisms (GMO) - Pollen monitoring - Technical pollen sampling using pollen mass filter (PMF) and Sigma-2 sampler. Beuth, Berlin; 2007.
Hofmann F: Kurzgutachten zur Abschätzung der Maispollendeposition in Relation zur Entfernung von Maispollenquellen mittels technischem Pollensammler PMF. Federal Agency for Nature Conservation. Bonn – Bad Godesberg: 2007. , [http://www.bfn.de/fileadmin/MDB/documents/themen/agrogentechnik/07–05–31_Gutachten_Pollendeposition_end.pdf]
Kawashima S, Clot B, Fujita T, Takahashi Y, Nakamura K: An algorithm and a device for counting airborne pollen automatically using laser optics. Atmos Environ 2007, 41: 7987–7993. 10.1016/j.atmosenv.2007.09.019
Schuppener M, Mühlhause J, Müller A-K, Rauschen S: Environmental risk assessment for the small tortoiseshell Aglais urticae and a stacked Bt-maize with combined resistances against Lepidoptera and Chrysomelidae in central European agrarian landscapes. Mol Ecol 2012, 21: 4646–4662. 10.1111/j.1365-294X.2012.05716.x
Gregory PH: The Microbiology of the Atmosphere. A Plant Science Monograph. Hill Books, Aylesbury; 1973.
Edmonds RL: Aerobiology: The Ecological Systems Approach. Dowden, Hutchinson & Ross, Stroudsburg; 1979.
Giesecke T, Fontana SL, van der Knaap WO, Pardoe HS, Pidek IA: From early pollen trapping experiments to the pollen monitoring programme. Veg Hist Archaeobot 2010, 19: 247–258. 10.1007/s00334-010-0261-3
Baron AB, Willeke K: Aerosol Measurement: Principles, Techniques, and Applications. John Wiley & Sons Inc., London; 2005.
McMurry PH: A review of atmospheric aerosol measurements. Atmos Environ 2000, 34: 1959–1599. 10.1016/S1352-2310(99)00455-0
Hofmann F, Schlechtriemen U, Wosniok W, Foth M: GVO - Pollenmonitoring - Technische und biologische Pollenakkumulatoren und PCR-Screening für ein Monitoring von gentechnisch veränderten Organismen. BfN Skripten 2005, 139: 274.
Folloni S, Kagkli DM, Rajcevic B, Guimaraes NCC, Van Droogenbroeck B, Valicente FH, Van den Eede G, Van den Bulcke M: Detection of airborne genetically modified maize pollen by real-time PCR. Mol Ecol Resour 2012, 12: 810–821. 10.1111/j.1755-0998.2012.03168.x
Association of German Engineers: VDI-Standard 2119 Blatt 4 - Ambient air measurements - Sampling of atmospheric particles bigger than 2,5 μm on an acceptor surface using the Sigma-2 passive sampler - Characterisation by optical microscopy and calculation of number settling rate and mass concentration. Berlin: Beuth; 2013.
Beismann H, Kuhlmann M: Raumrepräsentativität technischer Pollensammler für ein Langzeitmonitoring von gentechnisch veränderten Pflanzen (GVP). BfN Skripten 2006, 169: 305.
Hofmann F, Ober S, Janicke L, Janicke U, Kuhn U, Schlechtriemen U, Unselt C, Wittich KP: Eintrag von Maispflanzenteilen in die Umwelt: Abschätzung der Umweltexposition für die Risikobewertung transgener Pflanzen. BfN Skripten 2013, 353: 253.
Davis MB, Deevey ES: Pollen accumulation rates: estimates from late-glacial sediment of Rogers Lake. Science 1964, 145: 1293–1295. 10.1126/science.145.3638.1293
Kawashima S, Matsuo K, Du Ming Y, Takahashi Y, Inoue S, Yonemura S: An algorithm for estimating potential deposition of corn pollen for environmental assessment. Environ Biosafety Res 2004, 3: 197–207. 10.1051/ebr:2005003
Thompson R: Use of the word "influx" in paleolimnological studies. Quat Res 1980, 14: 269–270. 10.1016/0033-5894(80)90054-X
Hirst JM: An automatic volumetric spore trap. Ann Appl Biol 1952, 39: 257–265. 10.1111/j.1744-7348.1952.tb00904.x
McCartney HA, Aylor DE: Relative contributions of sedimentation and impaction to deposition of particles in a crop canopy. Agricult Forest Meteorol 1987, 40: 343–358. 10.1016/0168-1923(87)90046-3
VDI-Standard 4330 Blatt 1 - Monitoring the Ecological Effects of Genetically Modified Organisms (GMO). Basic Principles and Strategies. Beuth, Berlin; 2006.
Microsoft Office Excel®. 2010.
Addinsoft: , date accessed 13-12-2013., [http://www.xlstat.com/en/]
Oosterbaan RJ: ., [http://www.waterlog.info/segreg.htm]
R Core Team: R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna: 2013. ., [http://www.R-project.org/]
Wood SN: Generalized Additive Models: An Introduction with R. Chapman and Hall/CRC, London; 2006.
Perry JN, Devos Y, Arpaia S, Bartsch D, Gathmann A, Hails RS, Kiss J, Lheureux K, Manachini B, Mestdagh S, Mestdagh S, Neemann G, Ortego F, Schiemann J, Sweet J: A mathematical model of exposure of non-target Lepidoptera to Bt-maize pollen expressing Cry1Ab within Europe. Proc Biol Sci 2010, 277: 1417–1425. 10.1098/rspb.2009.2091
Perry JN, Devos Y, Arpaia S, Bartsch D, Ehlert C, Gathmann A, Hails RS, Hendriksen NB, Kiss J, Messéan A, Mestdagh S, Neemann G, Nuti M, Sweet J, Tebbe CC: Estimating the effects of Cry1F Bt-maize pollen on non-target Lepidoptera using a mathematical model of exposure. J Appl Ecol 2012, 49: 29–37. 10.1111/j.1365-2664.2011.02083.x
Scientific opinion supplementing previous conclusions and recommendations on maize 1507 for cultivation EFSA J 2012, 10(11):2934.
Scientific opinion updating the evaluation of the environmental risk assessment and risk management recommendations on insect resistant genetically modified maize 1507 for cultivation EFSA J 2011, 9(11):2429.
Scientific opinion supplementing the conclusions of the environmental risk assessment and risk management recommendations for the cultivation of the genetically modified insect resistant maize Bt11 and MON 810 EFSA J 2012, 10(12):3016.
Wraight CL, Zangerl AR, Carroll MJ, Berenbaum MR: Absence of toxicity of Bacillus thuringiensis pollen to black swallowtails under field conditions. Proc Natl Acad Sci 2000, 97: 7700–7703. 10.1073/pnas.130202097
Sears MK, Hellmich RL, Stanley-Horn DE, Oberhauser KS, Pleasants JM, Mattila HR, Siegfried BD, Dively GP: Impact of Bt corn pollen on monarch butterfly populations: a risk assessment. Proc Natl Acad Sci 2001, 98: 11937–11942. 10.1073/pnas.211329998
Neter J, Kutner MH, Nachtsheim CJ, Wasserman W: Applied linear statistical models. McGraw-Hill/Irwin, New York; 1996.
Hofmann F, Schlechtriemen U, Kuhn U, Wittich K-P, Koch W, Ober S, Vögel R, Otto M: Variation of maize pollen shedding in North Germany and its relevance for GMO-monitoring. In GM-Crop Cultivation – Ecological Effects on a Landscape Scale. Edited by: Breckling B, Verhoeven R. Peter Lang, Frankfurt; 2013:19–25.
Vögel R, Ober S, Hofmann F, Unselt C: Agrogentechnik und Naturschutz - Wie können Schutzgebiete und aquatische Systeme wirksam vor dem Eintrag von Bt-Toxin geschützt werden? Vorstellung der Ergebnisse eines dreijährigen Projektes mit konkreten Handlungsempfehlungen an das Land Brandenburg. Fachbeiträge des LUGV 133. Potsdam: LUGV; 2013. ., [http://www.mugv.brandenburg.de/sixcms/media.php/4055/fb_133_agrogen.pdf]
Nguyen HT, Jehle JA: Quantitative analysis of the seasonal and tissue-specific expression of Cry1Ab in transgenic maize Mon810. J Plant Dis Protect 2007, 114(2):82–87.
Takács E, Darvas B, Székács A: Analytical difficulties and certain biological aspects of Cry1Ab toxin determination in MON 810 genetically modified maize. Acta Phytopathol Entomol Hung 2012, 47(2):293–306. 10.1556/APhyt.47.2012.2.10
Felke M, Langebruch G-A: Auswirkungen des Pollens von transgenem Bt-Mais auf ausgewählte Schmetterlingslarven. BfN Skripten 157. Bonn: BfN; 2005.
Hellmich RL, Siegfried BD, Sears MK, Stanley-Horn DE, Daniels MJ, Mattila HR, Spencer T, Bidne KG, Lewis LC: Monarch larvae sensitivity to Bacillus turingiensis -purified proteins and pollen. Proc Natl Acad Sci 2001, 9: 11925–11930. 10.1073/pnas.211297698
Bøhn T, Aheto DW, Mwangala FS, Bones IL, Simoloka C, Mbeule I, Wikmark O-G, Schmidt G, Chapela I: Co-existence challenges in small-scale farming when farmers share and save seeds. In GM-Crop Cultivation - Ecological Effects on a Landscape Scale. Edited by: Breckling B, Verhoeven R. Peter Lang, Frankfurt; 2013:104–109.
Ernsing M, Breckling B, Reuter R, Schmidt G: Maize gene flow simulation for intensively used agrarian areas in Lower Saxony (Germany). In GM-Crop Cultivation - Ecological Effects on a Landscape Scale. Edited by: Breckling B, Verhoeven R. Peter Lang, Frankfurt; 2013:71–77.
Schröder W, Hofmann F: Wissenschaftstheoretische Grundlagen der Beobachtung von GVO-Umweltwirkungen. Umweltwiss Schadst-Forsch 2008, 20: 2–8. 10.1065/uwsf2007.06.194
Schröder W, Schmidt G: Overview of principles and implementations to deal with spatial issues in monitoring environmental effects of genetically modified organisms. Environ Sci Eur 2012, 24(6):1–10.
Acknowledgements
This study was supported by the Federal Agency for Nature Conservation (BfN) with funds provided by the Federal Ministry for Environment, Nature Conservation and Nuclear Safety. The authors thank all of the researchers, institutions, and federal states who generated, verified, and/or released the data used in this study. We thank Prof. Dr. H. Beismann, formerly TU München, Mr M. Debrus, Landwirtschaftskammer NRW; Dipl.-Met. V. Dietze, Freiburg; Dr. J. Eder, LfL Freising; Dr. B. Köllner, LANUV NRW Essen; PD Dr. Kratz, FU Berlin; Dr. M. Kruse-Plaß, Wölsauerhammer; Dr. W. Kumpfer, Breege; Dr. M. Kuhlmann, TU München; Dr. B. Oehen, FIBL Frick (Switzerland); Dr. L. Peichl and Mrs A. Wilk, LfU Augsburg; Prof. S. Rogg, TU München; Herr S. Salzmann, LGLN NDS; Dipl.-Forstwirt U. Schlechtriemen, TIEM Northeim; Mrs R. Schmidt, SfWuH, Bremen; Prof. Dr. B. Schröder, formerly University Potsdam, Dr. M. van den Bulcke and Dr. S. Folloni, formerly JRC, Ispra (Italien); Mr R. Vögel, LUGV Brandenburg; Dr. R. Wachter, Ganderkesee. We also thank Dr. J. Smith and Prof. Dr. W. Schröder for their support and the reviewers for their useful comments.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Hofmann, F., Otto, M. & Wosniok, W. Maize pollen deposition in relation to distance from the nearest pollen source under common cultivation - results of 10 years of monitoring (2001 to 2010). Environ Sci Eur 26, 24 (2014). https://doi.org/10.1186/s12302-014-0024-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12302-014-0024-3