
Abstract
Animals require sufficient intake of a variety of nutrients to support their development, somatic maintenance and reproduction. An adequate diet provides cell building blocks, chemical energy to drive cellular processes and essential nutrients that cannot be synthesised by the animal, or at least not in the required amounts. Dietary requirements of nematodes, including Caenorhabditis elegans have been extensively studied with the major aim to develop a chemically defined axenic medium that would support their growth and reproduction. At the same time, these studies helped elucidating important aspects of nutrition-related biochemistry and metabolism as well as the establishment of C. elegans as a powerful model in studying evolutionarily conserved pathways, and the influence of the diet on health.
Similar content being viewed by others

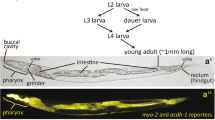

Caenorhabditis elegans ecology and diet in nature
Habitat
Caenorhabditis elegans is a free-living nematode with cosmopolitan distribution [1]. From its first isolation in 1900 by Emile Maupas, this 1-mm-long roundworm was described as a soil nematode. However, the worm is rarely found in pure soil but prefers humid patches that are rich in decaying plant material. It is often found in human-associated habitats, such as botanical gardens, orchards and compost heaps, where it prefers rotting stems, but it can occasionally be found in rotting fruits and flowers [2]. More recently, C. elegans has been isolated from forests and scrubland [3,4,5,6,7]. The main characteristic of these semi-natural and natural habitats is that they are rich in microbes and rotting vegetation. The individuals sampled occurred as a specialised larval diapause stage named dauer, which is formed due to absence of food, overcrowding or high environmental temperature, and is a main dispersal stage for colonisation of new food patches, by invertebrate carriers [1, 2]. Interestingly, in certain samples from France [5] and Northern Germany [8], C. elegans were found inhabiting the digestive tract of slugs as both dauer and feeding stages, suggesting that these worms can utilise slug intestinal microbes as a food source in the absence of decaying plants.
Food
In nature, C. elegans mainly feeds on different species of bacteria. These include soil bacteria such as Comomonas sp., Pseudomonas medocina and Bacillus megaterium [5, 9, 10]. The most commonly found bacteria in rotting fruits are Acetobacteriaceae (Acetobacter and Gluconobacter) and Enterobacteriaceae (Enterobacter) and therefore may represent a food source [11]. Moreover, intestinal extracts of freshly isolated C. elegans individuals contain also some digested eukaryotes, mostly yeast cells [5]. There is also a possibility that C. elegans takes up partially processed plant or animal material, found in decaying vegetation. This may ensure the intake of nutrients that the bacterial food sources cannot provide [1].
Feeding and food-related behaviour
Food ingestion in C. elegans is mediated by the pharynx, a neuromuscular tube-like organ that filters particulate food from a liquid suspension [12], concentrates and grinds it, and transports it further to the intestinal lumen [13], where the nutrients are taken up by intestinal cells. Similar to higher organisms, C. elegans is capable of making complex decisions based on the presence and quality of food in their environment [14]; they can learn to seek food which best supports their growth and avoid low-quality food and pathogenic bacteria [14, 15]. Additionally, worms on high-quality food display a behaviour named ‘satiety quiescence’, where they stop feeding and moving and become quiescent [16]. However, males are willing to leave the food in search for a mate [17, 18].
Dietary choices in the lab
Commonly used bacterial strains
While information about the C. elegans dietary choices in nature is scarce, its diet in the laboratory setting is quite well standardised. In the lab, C. elegans is cultured on agar petri plates seeded with bacteria. The most commonly used bacterial food source is the Escherichia coli strain OP50, a uracil auxotroph. Its use was advocated by Sydney Brenner as it grows in thin lawns which allow easier visualisation and mating of the worms [19]. Some researchers also use the E. coli wild-type K12 strain [20, 21], which forms a thick lawn and can support large worm populations on plate. The K12-derived strain HT115 (D3), which has a disrupted RNAse III gene, is widely used for RNA interference (RNAi) by feeding [22, 23]. HB101, a hybrid between E. coli strains K12 and B [24], forms a low-viscosity lawn as the cells do not adhere to each other, facilitating uptake by the worms [9, 25].
Biomass composition of E. coli, C. elegans and mammalian cells
Bacterial food provides worms with both macronutrients, which are used as sources of energy and as building blocks, and micronutrients, such as co-factors and vitamins. The average E. coli cell is very rich in nitrogen, and its dry weight comprises of approximately 55% protein, 23% nucleic acids (20% RNA and 3% DNA), 7–9% lipids and 6% carbohydrates, while vitamins, co-factors and ions comprise approximately 4% of the dry weight [26, 27]. However, E. coli strains used as food source in lab differ in their macronutrient content, especially carbohydrates [28, 29]. For instance, E. coli OP50 contains three to five times less carbohydrates compared to HT115 and HB101 [28]. Also, Comomonas aquatica DA1877 supports faster growth of C. elegans than E. coli OP50, due to its higher provision of vitamin B12 [29, 30]. Bacterial strains also differ in the amount of dietary folate or tryptophan they provide to the worms [31,32,33].
Biomass composition of C. elegans has not yet been characterised in detail. Worm dry biomass consists of roughly 60% protein, 20% lipids, 6.5% nucleic acids and 6% carbohydrates [34] (based on data from [35,36,37,38]). In mammalian cells, proteins also comprise most of the cell biomass, i.e. approximately 60%. Lipids amount to 13%, nucleic acids to 5% and sugars to roughly 6% of the dry mass [26, 39]. Overall protein and carbohydrate content seem to be similar in C. elegans and its E. coli food, as well as in mammals. However, relative lipid and nucleic acid content differ considerably between the prokaryotic E. coli and the eukaryotic C. elegans and mammalian species.
Nutritional requirements of C. elegans
Research into the dietary requirements of free-living nematodes has a long history. In the early twentieth century, Zimmerman reported successful cultivation of Turbatrix aceti in an axenic (i.e. bacteria-free) medium for numerous generations. This medium contained glucose, sodium chloride, peptone, lecithin and yeast extract [40].
Pioneering attempts to axenically culture the soil nematode Caenorhabditis briggsae highlighted a requirement for a ‘complete’ medium, containing high levels of crude organic components of undefined composition, e.g. chick embryo extract [41], liver [42,43,44] or plasma [45]. The added organic components contained unidentified, heat-labile protein-like substance(s), commonly named ‘Factor Rb’ [41, 46]. It was initially suggested that this factor was probably a vitamin source [47], but its chemical composition was only determined in subsequent fractionation and supplementation studies [48,49,50,51,52]. ‘Factor Rb’ was also shown to be a required addition to the basic medium [40] for successful growth and reproduction of T. aceti [53]. The first completely defined synthetic media for axenic culturing of C. briggsae were reported in 1956. The axenic growth medium GM-8 contained D-glucose as an energy source, amino acids, nucleotides, vitamins, salts and growth factors (such as N-acetylglucosamine, biotin and ascorbic acid). GM-9 and GM-11 were similar to GM-8 but with omission of certain amino acids and salts [54]. Unsupplemented, these media supported limited growth and reproduction of freshly-hatched larvae, while supplementation with liver medium in traces (0.032% of standard level) resulted in successful growth to adulthood and the production of a limited number of F1 offspring [54]. Subsequent media formulations were based on the composition of basal GM-8 medium (reviewed in detail in [55]).
In 1963, Sayre et al. [56] formulated a defined medium (EM1) based on the amino acid ratio found in E. coli instead of casein, as was the case in former media [54]. This medium was further modified by Buecher et al. [57] to a basal medium widely known as the Caenorhabditis briggsae Maintenance Medium (CbMM). Discoveries of nematode requirements for sterol source [48, 50], heme [49] and potassium acetate as an energy source [51] revealed the essential components of ‘Factor Rb’. By supplementation of CbMM with these substances, sustained growth of C. briggsae in a completely defined medium without added crude extracts was finally accomplished. Slightly modified, CbMM could support continuous growth of other free-living nematode species in axenic culture such as the bacteria-feeding rhabditids T. aceti [58,59,60,61], Panagrellus redivivus [58, 62], C. elegans [59, 63] and the fungi-feeding tylenchid Aphelenchus avenae [64]. C. elegans Maintenance Medium (CeMM) is a modification of the CbMM, which consists of 54 components, with either potassium acetate or glucose (albeit in higher concentration than in CbMM) as energy sources [52], [65,66,67]. Given that the chemically defined medium preparation is costly and time-consuming, there are additional variants of axenic medium available for cultivating C. elegans, one of which contains 3% soy peptone and 3% yeast extract, as sources of amino acids and vitamins, respectively, and 0.5 mg/ml of haemoglobin [68, 69]. Alternatively, C. elegans Habituation and Reproduction (CeHR) medium requires addition of milk [70], thus being referred to as semi-defined medium.
Caloric compounds and building blocks
Proteins and peptides
Early studies of nutritional requirements of free-living nematodes underlined the absolute requirement for supplementation of axenic media with a heat-labile proteinaceous factor to achieve continuous growth and reproduction [41, 44, 54]. Given that bacteria are food for C. briggsae in nature, it was tested whether they could potentially be a source of the proteinaceous factor in a liquid culture medium [47]. For this purpose, Klebsiella aerogenes was used: autoclaved and in form of a filter-sterilised supernatant of a cell homogenate. However, both treatments resulted in loss of K. aerogenes growth-promoting activity in media consisting of either salts and glucose or ‘Factor Rb’-free autoclaved liver extract [47]. After the discovery that nematodes require an exogenous sterol source, it was reported that growth could be rescued in a medium with autoclaved bacteria and sterol, by adding a liver extract. Fractionation analysis indicated that this heat-inactivated component provided by bacteria and liver is heme [48, 49], which will be discussed in more detail later. With these studies, it became clear that the proteinaceous factor was a source of sterol and heme, which were not provided in any of the chemically defined media [56, 57]. Furthermore, yeast extract also had growth-promoting activity in the form of a pellet of partially denatured ribosomes [71]. This highlighted the importance of particulate matter in a medium and that the growth-promoting activity depends on the uptake, rather than the protein nature of the extract. This was later supported by two studies in which pure precipitated proteins were used [68, 72, 73]. Likewise, C. elegans requires particulate matter to successfully take up the nutrients present in axenic medium [74]. By subjecting the medium to 0.22-μm filtration, it retains a very low concentration of small-sized particles that worms cannot ingest but expel with liquid, resulting in impediment of worm growth [74].
It remains unclear from previous studies why tissue extracts still proved to be more efficient in promoting growth in the presence of sterol and heme than any pure protein [72, 73]. A partial answer to this question was provided by a study in which supplementation of medium with glycogen (in the presence of a heme source, but devoid of any protein) had a stimulatory effect on the reproduction of C. briggsae [75]. Based on all these findings, Vanfleteren suggested that the growth factor had two roles: to provide the two essential nutrients, sterol and heme and to supply the particulate matter in a medium that would facilitate the uptake of heme [68, 76]. Finally, two studies by Lu et al. revealed that the yet unknown function of the growth factor was to provide an adequate energy source and that it could be fulfilled by adding different lipid components, potassium acetate or glucose [51, 52]. Hence, other than for the heme, C. elegans does not require any essential protein or peptide in its diet, unless as a general source of essential amino acids, and these requirements will be discussed in more detail in subsequent sections.
Amino acids
In 1912, Abderhalden classified amino acids as nutritionally essential and nutritionally non-essential [77]. The former are the amino acids that cannot be synthesised by an animal in normal physiological conditions or not at sufficient rate to support normal growth and thus must be acquired through the diet. On the other hand, non-essential amino acids can be synthesised from precursors in amounts that are sufficient to support the animal’s growth and do not require dietary supplementation. During the evolutionary adaptation to foraging on other organisms, many eukaryotes, including C. elegans, lost the ability to synthesise approximately half of the amino acids, in contrast to plants and fungi that are able to synthesise them all [78,79,80,81].
Dougherty and Hansen [54] were the first to demonstrate that C. briggsae could be successfully cultured in a semi-defined medium that contained arginine, histidine, leucine, isoleucine, lysine, phenylalanine, methionine, tryptophan, threonine and valine as only amino acid supplements. However, added liver extract [54] could have been a source of additional amino acids and/or changed their concentration in the mixture leading to non-reliable conclusions about their essentiality. Later studies on amino acid metabolism based on the use of radioactively (14C) labelled precursors showed that C. briggsae is capable of synthesising 16 amino acids [82,83,84]. However, this study could not reveal whether the rate at which these amino acids are synthesised is adequate to sustain growth and reproduction. Finally, Vanfleteren [68, 85] unambiguously showed that the dietary essential amino acids for both C. briggsae and C. elegans are arginine, histidine, lysine, tryptophan, phenylalanine, methionine, threonine, leucine, isoleucine and valine. The same set of amino acids, except for arginine, represents the dietary essential amino acids for rat and human [78, 86]. Whole-genome sequence analysis by Payne and Loomis [81] indicated that biosynthetic pathways for 10 dietary non-essential amino acids are evolutionarily conserved in rat, human and C. elegans. However, the C. elegans genome does not encode for orthologues of the enzymes involved in arginine biosynthesis in the human urea cycle (i.e. carbamoylphosphate synthetase 1 (CPS1), ornithine transcarbamoylase (OTC), argininosuccinate synthetase 1 (ASS1) and argininosuccinate lyase (ASL) [81, 87]. In humans, the main source of endogenous arginine is the kidneys. There, citrulline derived from glutamine metabolism in the small intestine is converted into arginine by ASS and ASL and released to blood [87, 88].
Carbohydrates
An early study of carbohydrate requirements of free-living nematodes reported that fecundity of C. elegans cultured in CbMM supplemented with liver growth factor devoid of glucose, inositol and choline was reduced compared to the same medium with added carbohydrates. Addition of 6.5 mg/ml of glucose or trehalose resulted in increased reproduction while supplementation with ribose and sucrose had no such effect [63]. Also, glycogen supplementation to CbMM containing hemin had a positive effect on growth and reproduction of both C. briggsae and C. elegans [75]. In contrast with the previous report [63], addition of trehalose to the hemin-supplemented medium did not support growth and reproduction of nematodes, which was also shown for other carbohydrate supplements such as rice starch, rice flour and corn starch [75]. Overall, these studies did not clearly elucidate the role of carbohydrate supplementation in a defined axenic medium. Lu et al. [51] showed that the addition of potassium acetate to CbMM results in higher population growth of C. elegans compared to CbMM supplemented with casamino acids (a casein hydrolysate that contains amino acids and peptides). This was a first study in which a chemically defined compound was characterised as an energy source able to support optimal growth and reproduction in CbMM. Later, it was shown that carbohydrates, in particular glucose and glycogen, and to a lesser extent trehalose (in contrast with [63]), fructose and sucrose, can support C. elegans maintenance as a major energy source in axenic medium [52]. Based on this study, it was recommended to increase the glucose concentration of CbMM to 32.5 mg/ml, a level that gives the highest population growth. In a medium without carbohydrate supplementation, worm growth is heavily impaired, which provides definite evidence for the worm’s nutritional requirement for sugar supplementation [52].
In addition to being an important dietary constituent, glucose also has profound effects on C. elegans physiology [89,90,91]. For instance, worms fed a high-glucose diet live shorter and this effect depends on downregulation of the transcription factor DAF-16/FOXO, heat shock factor HSF-1 and the glycerol channel aqp-1 [91]. This resembles the finding that glycerol channel Aqp7-knockout (KO) mice show increased triglyceride levels and develop obesity and insulin resistance when fed a diet rich in fat or sugar [92]. In C. elegans, high glucose exerts its detrimental effects through increased formation of reactive oxygen species (ROS), triglyceride accumulation and formation of advanced glycation end products (AGEs) [90, 93, 94]. However, the observed lifespan-shortening effect of a high-glucose diet in C. elegans can be alleviated by upregulation of the sterol regulatory element-binding protein (SREBP) homologue sbp-1 and mediator-15 (mdt-15), which induce expression of fatty acid (FA) desaturases, thus decreasing levels of saturated FAs by converting them to unsaturated FAs [95]. Similarly, toxic effects of glucose in mice are reduced by SREBP-induced lipogenesis in hepatocytes, which lowers blood glucose levels [96, 97].
Both in humans and C. elegans, excess glucose is stored as glycogen. Primary storage sites of glycogen in humans are the liver and skeletal muscle [98], while in worms, these are the intestine, and to a certain extent the hypodermis and muscle [99,100,101]. Unlike humans, who possess different isoforms of glycogen synthase in each of the storage tissues [98], C. elegans has only one, encoded by gsy-1 [99, 100]. In non-feeding C. elegans dauer larvae increased glycogen levels are important sources of energy for locomotion and nictation behaviour (i.e. dispersal behaviour characterised by upright waving motions) [102]. Moreover, in C. elegans, glucose can be stored as the non-reducing disaccharide trehalose via activity of trehalose-6-phosphate synthase, encoded by genes tps-1 and tps-2 [103, 104]. Humans, however, do not have this capacity but can break down dietary trehalose [105]. Interestingly, C. elegans with reduced gsy-1 activity live longer and healthier due to elevated storage of glucose in the form of trehalose, in a DAF-16/FOXO-dependent manner [101]. A similar positive effect on lifespan and healthspan is achieved by dietary supplementation of trehalose and inhibition of trehalose breakdown [101].
Lipids
An isotope labelling study by Perez and Van Gilst [106] revealed that E. coli OP50 is the major source of palmitate (C16:0) in lab-cultured C. elegans. Additionally, palmitoleate (C16:1ω7), vaccenate (C18:1ω7) and cyclopropane fatty acids (C17 and C19) are also predominantly absorbed from the bacterial food source, although worms can synthesise these fatty acids in very small amounts [106, 107]. The bacterial diet cannot provide stearic (C18:0), oleic (C18:1ω9), linoleic (C18:2ω6) and C20 polyunsaturated fatty acids (PUFAs) [106, 107], but C. elegans is enzymatically equipped to synthesise these fatty acids de novo [108, 109]. This ability was first demonstrated for nematodes in the free-living nematode T. aceti [110] and later in C. briggsae and P. redivivus [111] by incorporation of 14C-labelled acetate into complete chains of linoleic, linolenic and arachidonic acids in worms grown in axenic culture. Moreover, Lu et al. [51] evaluated the effect of sodium oleate, linoleate and stearate on population growth of C. briggsae in CbMM and showed that oleate supported the best growth, potentially due to more efficient utilisation of saturated and monounsaturated FAs than PUFAs by nematodes.
PUFAs are essential constituents of cell membranes conferring their fluidity and semi-permeability. They also have a range of important roles in cell signalling, endocytosis and exocytosis, immune response and pathogen defence [112,113,114]. The critical step in PUFA synthesis is the production of linoleic acid by desaturases, through introduction of a second double bond into the chain of oleic acid (a monounsaturated FA). This feature has previously been attributed only to plants, fungi and protozoans [115]. However, numerous studies have shown that this step occurs in several insect species [115], certain arachnids [116], pulmonate molluscs [117] and nematodes [108, 110] but not in vertebrates [118]. Mammals lack Δ12 and ω3 desaturases found in plants, hence, plant-derived linoleic and linolenic (C18:3ω3) acids are essential dietary FAs and required precursors for C20 PUFA synthesis [109]. C20 PUFA synthesis in mammals occurs through series of desaturation and elongation steps of essential FAs in the endoplasmic reticulum. These processes are catalysed by Δ6 and Δ5 desaturases and elongases, respectively, to produce arachidonic (20:4ω6) and eicosapentaenoic (20:5ω3) acids [119]. In contrast with plants and humans, C. elegans possesses all the enzymes necessary to produce arachidonic and eicosapentaenoic acids. The Δ12 and ω3 desaturases found in plants are encoded by fat-2 and fat-1, respectively, with fat-1 having both desaturase functions [120,121,122]. The C. elegans homologues of human Δ6 and Δ5 desaturases and ω3 and ω6 elongases are fat-3, fat-4, elo-1 and elo-2, respectively [108, 109, 123].
Monomethyl branched-chain fatty acids (mmBCFA) C15ISO and C17ISO represent another group of FAs that are essential for C. elegans growth and development and are synthesised de novo [124, 125]. Unlike the biosynthetic pathway of straight-chain FAs, where the primer is acetyl-CoA [126], biosynthesis of mmBCFAs utilises primers derived from branched-chain amino acids (BCAA; leucine, isoleucine and valine), while the chain extender for both pathways is malonyl-CoA [127]. Primer synthesis is catalysed by the branched-chain α-ketoacid dehydrogenase (BCKDH) multi-subunit complex and elongation of the mmBCFA backbone is performed by elongases ELO-5 and ELO-6 [124]. Disruption of BCKDH in humans causes a metabolic disorder named maple syrup urine disease (MSUD), which symptoms are mainly attributed to accumulation of BCAA [128, 129]. Loss of function of dbt-1 (encoding for the C. elegans homologue of the human BCKDH E2 subunit) results in embryonic lethality and arrested larval development, predominantly due to lack of mmBCFAs rather than the accumulation of BCAAs [130]. Hence, this may give new insights into the importance of mmBCFAs for human health.
Fats are sequestered in lipid droplets, organelles that are evolutionarily conserved in C. elegans [131], D. melanogaster [132], S. cerevisiae [133] and mammals [134]. Unlike humans and other vertebrates, C. elegans does not possess specialised cells called adipocytes [135], which are the main constituents of the adipose tissue, an organ with endocrine and immune functions [136, 137]. Instead, C. elegans stores fats in the intestine, which is a major energy storage organ, similar in function to the liver and adipose tissue. To some extent, fat also accumulates in the hypodermis [131, 135] and muscles [138]. Like glycogen, triglycerides are accumulated in large amounts in C. elegans dauer larvae, where they serve as a primary energy source for starvation survival [139, 140].
Small organic compounds
The ability of free-living nematodes to utilise two-carbon compounds (e.g. acetic acid and ethanol) as an energy source in axenic medium was first demonstrated in three rhabditid species: C. briggsae, T. aceti and P. redivivus [141,142,143]. Supplementation of ethanol, n-propanol and potassium acetate has stimulatory effects on population growth of C. briggsae in CbMM [51]. These short carbon-chain compounds can be readily utilised by free-living nematodes, including C. elegans, due to the presence of a set of enzymes for conversion of alcohols into acetyl-CoA, i.e. alcohol dehydrogenase, aldehyde dehydrogenase and acetyl-CoA synthetase [144]. Starved L1 larvae of C. elegans live twice as long when treated with 1 mM ethanol. The underlying mechanism is not clear [145] but likely involves conversion of ethanol into acetyl-CoA. This could be an important strategy for starvation survival in harsh environmental conditions.
The fully functional glyoxylate shunt allows C. elegans to convert acetyl-coA into succinate, which can fuel the citric acid cycle and the resulting excess of oxaloacetate may be utilised for gluconeogenesis [146]. In addition to nematodes, an operational glyoxylate cycle is present in bacteria, fungi, plants and protozoans, but not in vertebrates. However, some studies reported the activity of one or both glyoxylate cycle enzymes, isocitrate lyase (ICL) and malate synthase (MS), in different tissues of amphibians [147], birds [148] and mammals [149]. Moreover, a comparative genomic study identified a tandem of ICL and MS genes in a cnidarian genome [150]. No study so far managed to find a fully functional glyoxylate cycle in any metazoan besides nematodes.
Vitamins
Vitamins are defined as a group of organic compounds essential in small amounts for organismal function that cannot be synthesised by the body and thus must be provided through the diet (or via the biosynthetic activity of intestinal bacteria). Unlike macromolecules that are classified based on similarities in their chemical properties, vitamins are grouped based on the function they serve. Different organisms vary in their capacity to synthesise these organic compounds. Hence, what is considered a vitamin for one organism might not serve that function for the other.
Sterols
Early studies into the dietary requirements of free-living nematodes and attempts to formulate a chemically defined medium that would support growth and reproduction gradually resulted in a defined medium (CbMM) that required supplementation with crude substances for sustainable cultures [54, 56, 57]. One of the essential growth factors that CbMM lacked and the crude extracts could potentially provide were sterols [57]. A study on Steinernema feltiae was the first one to report the requirement of sterols for growth and reproduction of nematodes in axenic medium [151]. The same requirement was later demonstrated for C. briggsae, T. aceti and P. redivivus [48, 152]. Parallel biochemical studies revealed that these species are unable to synthesise sterols de novo from 14C-labelled acetate and mevalonate [153, 154].
In order to find the missing step(s) in nematode sterol biosynthesis, Lu et al. added five sterol precursors (acetic acid, DL-mevalonic acid lactone, farnesol, squalene and lanosterol) and cholesterol to T. aceti, C. briggsae and C. elegans populations cultured in CbMM [50]. Supplementation with acetic acid, DL-mevalonic acid lactone and farnesol did not have any significant effect on population growth while squalene, lanosterol and cholesterol showed dose-dependent positive effects. Hence, it was proposed that the metabolic block in de novo sterol biosynthesis in nematodes likely occurs between farnesol and squalene and at any of the steps prior to farnesol synthesis [50]. However, these conclusions were in contrast with a previous study reporting that squalene had no effect on growth and reproduction of E. coli-fed C. elegans. Also, another study indicated no effect of lanosterol on C. elegans development in a chemically defined medium [48, 155]. On the contrary, C. elegans growth is fully supported only by exogenous provision of ergosterol, β-sitosterol, stigmasterol or cholesterol, which are the final products of sterol biosynthesis in plants and mammals [48, 66]. Indeed, the C. elegans genome encodes for homologues of mammalian enzymes involved in the initial steps of sterol synthesis (up to farnesyl pyrophosphate), but no downstream enzymes, including squalene synthase and squalene cyclase [156, 157]. Unlike nematodes, mammals possess a complete set of enzymes required for de novo synthesis of sterols from acetyl-CoA, under the tight regulation of SREBP and thus do not require exogenous sterol supplementation [158].
Under standard laboratory conditions, C. elegans sterol requirement is fulfilled by addition of cholesterol to the culture, even though it was shown not to be the essential dietary sterol for C. elegans [159, 160]. Similarly, in nature, bacterial food cannot provide dietary sterols; thus, this requirement is likely met by feeding on decaying plant or fungal material or on animal faeces [1, 156].
In mammals, cholesterol is an important component of cellular membranes required for their fluidity and semi-permeability and a precursor for the synthesis of bile acids and steroid hormones. In C. elegans, its role is not entirely clear. Given that C. elegans membranes contain almost no cholesterol, its structural role was proposed to be less likely. Instead, it was suggested that it could have roles in cellular signalling, related to moulting and dauer formation [156, 160, 161]. Indeed, C. elegans uses cholesterol as a precursor for the synthesis of dafachronic acids (DAs), bile-like steroids that bind to the nuclear receptor DAF-12, a homologue of pregnane-X and vitamin D receptors in vertebrates that controls dauer entry and modulates lifespan [162,163,164,165]. The key role in DA synthesis belongs to daf-9-encoded cytochrome P450, of which the human homologue is sterol 27-hydroxylase (CYP27A1) known to be involved in bile acid synthesis in the liver [163, 166, 167].
Heme
Supplementation of chemically defined axenic media with organic substances could possibly provide nematodes with yet another essential growth factor which these media lacked: heme [54, 56, 57]. C. briggsae can be indefinitely cultured in a salt-buffered medium containing only live E. coli supplemented with sterols [48]. When bacteria are heat-killed in such medium, C. briggsae cannot grow, suggesting that a certain essential component had been destroyed by autoclaving bacteria. However, C. briggsae can be successfully cultured in a medium with autoclaved E. coli and sterols supplemented with a heated liver extract that contains heme. The same effect was achieved by supplementation of cytochrome c, haemoglobin, myoglobin or hemin chloride [49]. Addition of hemin chloride to CbMM and EM1 allowed for repeated subculturing of four free-living nematode species—T. aceti, P. redivivus, C. briggsae and C. elegans—but the final population size was small, unless the concentration of hemin chloride was high (250 μg/ml) or it was provided in an adequate precipitated form at lower concentration (50 μg/ml) [76, 168]. Under standard axenic culture conditions, the dietary requirement for heme is met by supplementation of the medium with haemoglobin [68, 69] or cytochrome c [66]. Standard culturing on agar plates seeded with E. coli does not require addition of heme to the medium, since it is provided by the live bacteria. C. elegans utilises dietary heme to provide this prosthetic group to endogenous heme proteins and potentially as an iron source in conditions of environmental iron deficiency [169].
Genome sequence analysis indicated that C. elegans lacks orthologues of all mammalian genes involved in synthesis of heme from δ-aminolevulinic acid. These enzymes are cytosolic δ-aminolevulinic acid dehydratase, porphobilinogen deaminase, uroporphyrinogen III synthase, uroporphyrinogen decarboxylase, mitochondrial coproporphyrinogen oxidase, protoporphyrinogen oxidase and ferrochetalase [169, 170].
Being a heme auxotroph, C. elegans has been a powerful model to study mechanisms of heme uptake, transport and homeostasis that are evolutionary conserved between worms and mammals [171,172,173,174,175,176]. Uptake of dietary heme in C. elegans is mediated by the activity of the two heme transporters, HRG-1 (heme-responsive gene 1) and HRG-4, which import heme into the intestine [171, 174]. Subsequently, heme is delivered to other tissues and the embryos by the secreted transporter HRG-3 and to the hypodermis by the transmembrane transporter HRG-2 [173, 175]. Finally, the multidrug resistance protein MRP-5 is the key player in regulating heme homeostasis in C. elegans by acting as a heme exporter, which is genetically conserved between worms, yeast, zebrafish and mammals [176].
Other vitamins
First insights into the vitamin requirements of nematodes emerged in the 1950s in a study in which was shown that continuous growth of C. briggsae in axenic medium containing autoclaved liver extract could be accomplished by the addition of folic acid [177]. This finding was later confirmed in experiments with aminopterin, a folic acid antagonist, which had severe effects on C. briggsae growth and development due to thymine deficiency [178]. In addition to being important for thymine biosynthesis, folic acid is also required by C. briggsae for histidine catabolism [178, 179]. These findings were not surprising, given that a biochemical pathway for de novo folate synthesis is present only in plants and microorganisms, while animals require dietary folate to maintain physiological functions. In humans, folate deficiency leads to neural tube defects during embryogenesis and dietary supplementation of folic acid has proven successful in decreasing the occurrence of such birth defects [180, 181]. C. elegans feeding on E. coli that are mutant in folate biosynthesis show a lifespan extension of 30–50%. However, this is not a consequence of changes in folate uptake in C. elegans but probably occurs due to reduction in toxin-based virulence of E. coli, related to excess folate that these bacteria produce [31, 33]. In humans and other mammals, folate is taken up by the reduced folate carrier (RFC), which is similar to the folate uptake in C. elegans via the carrier encoded by folt-1 [182, 183].
In addition to folate, all the other vitamins of the B complex were also reported to be essential for normal growth and development of C. briggsae in axenic medium: riboflavin, thiamine, pyridoxine, niacinamide, pantothenic acid, biotin and cobalamin [55, 184,185,186]. Among these, cobalamin (vitamin B12) was most extensively studied in the C. elegans model system [30, 187,188,189]. This vitamin has the unique property of being synthesised solely by archaea and bacteria [190]. However, the common food source for C. elegans in the lab, E. coli, lacks the vitamin B12 biosynthetic pathway. Hence, worms take it up by ingesting bacteria that absorb the vitamin from the culturing medium [191]. Vitamin B12 deficiency was shown to severely impair C. elegans biology, leading to growth retardation, lifespan shortening and reduced egg-laying capacity, which is consistent with results obtained in mice and humans [187, 192, 193]. Also, memory retention in C. elegans is impaired, partially due to severe oxidative stress [188]. MRP-5, previously shown to be the heme exporter through the intestinal membrane to other tissues, was also identified as the exporter of vitamin B12 from the hermaphrodite intestine to the embryos [176, 189]. This finding can be explained by the structural similarity of vitamin B12 and heme; both contain a protoporphyrin ring with a cobalt or iron ion in its centre, respectively. Additionally, based on sequence homology, MRP-5 is likely a functional ortholog of the human MRP1 [189]. Apart from vitamins of the B complex, no information is available on C. elegans nutritional requirements for other vitamins.
Minerals
Dietary minerals represent a class of inorganic nutrients that are essential for many metabolic and physiological processes in the body, usually required in small amounts naturally found in different types of food. Minerals are required for signal transduction, maintenance of osmotic balance and acid-base equilibrium, energy metabolism, enzyme functions and, in vertebrates, for formation and maintenance of bones [194]. Importantly, certain minerals are required by all animals, but the amounts in which they must be provided can vary greatly depending on the species and the function of the mineral.
In humans, dietary minerals are classified into principal or macroelements and trace or microelements. The former are required in amounts greater than 200 mg per day and comprise of calcium, phosphorus, sulphur, potassium, chlorine, sodium and magnesium. The latter are required in minute amounts; represent only 0.02% of the total body weight; and include zinc, iron, chromium, copper, cobalt, manganese, molybdenum, selenium, iodine and fluorine [195]. Importantly, both mineral deficiency as well as excess mineral intake can have detrimental effects on health. For instance, iron deficiency can result in anaemia and problems with the immune system, while excess intake causes liver damage [195].
Mineral deficiency studies in nematodes were difficult to perform until a completely chemically defined medium was developed [51]. Deficiency of individual minerals was achieved by depleting them from the basal medium. After depletion, different concentrations of each mineral were added to the medium to determine the quantitative requirements of C. elegans [196]. Potassium and magnesium were shown to be absolutely essential minerals for C. elegans since no growth was observed when these minerals were removed from CbMM. C. elegans could survive copper, calcium and manganese depletion. However, complete deficiency of these minerals was difficult to induce. Similarly, sodium was present in different components of the medium and effects of its absolute deficiency could not be tested. Hence, the authors suggested optimal concentrations of these minerals in CbMM to achieve maximal population growth but gave no conclusion on their essentiality [196]. Moreover, this study could not provide any evidence for the requirement of zinc in C. elegans, nor could it create iron deficiency, given that heme was an essential part of the CbMM. A recent study on zinc deficiency in C. elegans revealed that lack of zinc reduces worm fertility by causing aberrations in oocyte development and meiotic division [197]. These results are consistent with the role of zinc in mammalian gametogenesis where it is involved in the production of sperm and maturation of oocytes. This establishes C. elegans as a suitable model for the study of zinc as a factor in animal fertility [198, 199]. Furthermore, reducing zinc levels in vivo has been shown to extend C. elegans lifespan and reduce age-related protein aggregation, partially by inducing DAF-16/FOXO nuclear localization [200].
Concluding remarks
For decades, C. elegans has been a preferred model organism to study fundamental metazoan biology, due it its genetic amenability, inexpensive maintenance, easy experimentation and fully annotated genome encoding for homologues of many human disease-associated genes. Therefore, it can also be considered a suitable model for the genetics of animal nutrition and metabolism. Indeed, many dietary requirements and metabolic responses are evolutionarily conserved, such as the fat accumulation as a result of decreased Ins/IGF-like signalling. Yet, one should be well aware that nematodes and vertebrates do not share all enzymatic pathways and thus show some important differences in their dietary requirements. The most profound differences are sterol and heme auxotrophy of nematodes and the types of essential amino acids and fatty acids required by worm and human (outlined in Fig. 1).

Known common and specific essential nutrients of C. elegans and humans
References
Schulenburg H, Félix M-A. The natural biotic environment of Caenorhabditis elegans. Genetics. 2017;206(1):55–86.
Félix M-A, Braendle C. The natural history of Caenorhabditis elegans. Curr Biol. 2010;20(22):R965–9.
Barrière A, Félix M-A. High local genetic diversity and low outcrossing rate in Caenorhabditis elegans natural populations. Curr Biol. 2005;15(13):1176–84.
Kiontke KC, Félix M-A, Ailion M, Rockman MV, Braendle C, Pénigault J-B, Fitch DHA. A phylogeny and molecular barcodes for Caenorhabditis, with numerous new species from rotting fruits. BMC Evol Biol. 2011;11(1):339.
Félix M-A, Duveau F. Population dynamics and habitat sharing of natural populations of Caenorhabditis elegans and C. briggsae. BMC Biol. 2012;10(1):59.
Petersen C, Dirksen P, Prahl S, Strathmann EA, Schulenburg H. The prevalence of Caenorhabditis elegans across 1.5 years in selected North German locations: the importance of substrate type, abiotic parameters, and Caenorhabditis competitors. BMC Ecol. 2014;14(1):4.
Cook DE, Zdraljevic S, Tanny RE, Seo B, Riccardi DD, Noble LM, Rockman MV, Alkema MJ, Braendle C, Kammenga JE, et al. The genetic basis of natural variation in Caenorhabditis elegans telomere length. Genetics. 2016;204(1):371.
Petersen C, Hermann RJ, Barg M-C, Schalkowski R, Dirksen P, Barbosa C, Schulenburg H. Travelling at a slug’s pace: possible invertebrate vectors of Caenorhabditis nematodes. BMC Ecol. 2015;15(1):19.
Avery L, Shtonda BB. Food transport in the C. elegans pharynx. J Exp Biol. 2003;206(14):2441.
Montalvo-Katz S, Huang H, Appel MD, Berg M, Shapira M. Association with soil bacteria enhances p38-dependent infection resistance in Caenorhabditis elegans. Infect Immun. 2013;81(2):514.
Samuel BS, Rowedder H, Braendle C, Félix M-A, Ruvkun G. Caenorhabditis elegans responses to bacteria from its natural habitats. Proc Natl Acad Sci. 2016;113(27):3941–9.
Avery L, Thomas JH. Feeding and defecation. In: Riddle DLBT, Meyer BJ, et al., editors. C elegans II. 2nd ed. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 1997. p. 679–716.
Doncaster CC. Nematode feeding mechanisms. 1. Observations On Rhabditis and Pelodera. Nematologica. 1962;8(4):313.
Shtonda BB, Avery L. Dietary choice behavior in Caenorhabditis elegans. J Exp Biol. 2006;209(1):89.
Zhang Y, Lu H, Bargmann CI. Pathogenic bacteria induce aversive olfactory learning in Caenorhabditis elegans. Nature. 2005;438:179.
You Y-j, Kim J, Raizen DM, Avery L. Insulin, cGMP, and TGF-β signals regulate food intake and quiescence in C. elegans: a model for satiety. Cell Metab. 2008;7(3):249–57.
Lipton J, Kleemann G, Ghosh R, Lints R, Emmons SW. Mate searching in Caenorhabditis elegans: a genetic model for sex drive in a simple invertebrate. J Neurosci. 2004;24(34):7427.
Ryan Deborah A, Miller Renee M, Lee K, Neal SJ, Fagan Kelli A, Sengupta P, Portman Douglas S. Sex, age, and hunger regulate behavioral prioritization through dynamic modulation of chemoreceptor expression. Curr Biol. 2014;24(21):2509–17.
Brenner S. The genetics of Caenorhabditis elegans. Genetics. 1974;77(1):71.
Browning DF, Wells TJ, França FLS, Morris FC, Sevastsyanovich YR, Bryant JA, Johnson MD, Lund PA, Cunningham AF, Hobman JL, et al. Laboratory adapted Escherichia coli K-12 becomes a pathogen of Caenorhabditis elegans upon restoration of O antigen biosynthesis. Mol Microbiol. 2013;87(5):939–50.
Depuydt G, Xie F, Petyuk VA, Shanmugam N, Smolders A, Dhondt I, Brewer HM, Camp DG 2nd, Smith RD, Braeckman BP. Reduced insulin/insulin-like growth factor-1 signaling and dietary restriction inhibit translation but preserve muscle mass in Caenorhabditis elegans. Mol Cell Proteomics. 2013;12(12):3624–39.
Kamath RS, Fraser AG, Dong Y, Poulin G, Durbin R, Gotta M, Kanapin A, Le Bot N, Moreno S, Sohrmann M, et al. Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature. 2003;421:231.
Rual J-F, Ceron J, Koreth J, Hao T, Nicot A-S, Hirozane-Kishikawa T, Vandenhaute J, Orkin SH, Hill DE, van den Heuvel S, et al. Toward improving Caenorhabditis elegans phenome mapping with an ORFeome-based RNAi library. Genome Res. 2004;14(10B):2162–8.
Boyer HW, Roulland-dussoix D. A complementation analysis of the restriction and modification of DNA in Escherichia coli. J Mol Biol Mol. 1969;41(3):459–72.
Davis MW, Somerville D, Lee RY, Lockery S, Avery L, Fambrough DM. Mutations in the Caenorhabditis elegans Na,K-ATPase alpha-subunit gene, eat-6, disrupt excitable cell function. J Neurosci. 1995;15(12):8408–18.
Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular biology of the cell. 5th ed. New York: Garland Science; 2008.
Bremer H, Dennis P. Modulation of chemical composition and other parameters of the cell at different exponential growth rates. EcoSal Plus. 2008;3(1):1553–69
Brooks KK, Liang B, Watts JL. The influence of bacterial diet on fat storage in C. elegans. PLoS One. 2009;4(10):e7545.
MacNeil Lesley T, Watson E, Arda HE, Zhu Lihua J, Walhout Albertha JM. Diet-induced developmental acceleration independent of TOR and insulin in C. elegans. Cell. 2013;153(1):240–52.
Watson E, MacNeil Lesley T, Ritter Ashlyn D, Yilmaz LS, Rosebrock Adam P, Caudy Amy A, Walhout Albertha JM. Interspecies systems biology uncovers metabolites affecting C. elegans gene expression and life history traits. Cell. 2014;156(4):759–70.
Virk B, Correia G, Dixon DP, Feyst I, Jia J, Oberleitner N, Briggs Z, Hodge E, Edwards R, Ward J, et al. Excessive folate synthesis limits lifespan in the C. elegans: E. coli aging model. BMC Biol. 2012;10(1):67.
Gracida X, Eckmann Christian R. Fertility and germline stem cell maintenance under different diets requires nhr-114/HNF4 in C. elegans. Curr Biol. 2013;23(7):607–13.
Virk B, Jia J, Maynard CA, Raimundo A, Lefebvre J, Richards SA, Chetina N, Liang Y, Helliwell N, Cipinska M, et al. Folate acts in E. coli to accelerate C. elegans aging independently of bacterial biosynthesis. Cell Rep. 2016;14(7):1611–20.
Yilmaz LS, Walhout Albertha JM. A Caenorhabditis elegans genome-scale metabolic network model. Cell Syst. 2016;2(5):297–311.
Cooper AF, Van Gundy SD. Metabolism of glycogen and neutral lipids by Aphelenchus avenae and Caenorhabditis sp. in aerobic, microaerobic and anaerobic environments. J Nematol. 1970;2(4):305–15.
Brock TJ, Browse J, Watts JL. Fatty acid desaturation and the regulation of adiposity in Caenorhabditis elegans. Genetics. 2007;176(2):865–75.
Miersch C, Döring F. Sex differences in carbohydrate metabolism are linked to gene expression in Caenorhabditis elegans. PLoS One. 2012;7(9):e44748.
Heavner BD, Smallbone K, Barker B, Mendes P, Walker LP. Yeast 5 – an expanded reconstruction of the Saccharomyces cerevisiae metabolic network. BMC Syst Biol. 2012;6(1):55.
Delgado FF, Cermak N, Hecht VC, Son S, Li Y, Knudsen SM, Olcum S, Higgins JM, Chen J, Grover WH, et al. Intracellular water exchange for measuring the dry mass, water mass and changes in chemical composition of living cells. PLOS ONE. 2013;8(7):e67590.
Zimmermann A. Recherches experimentales sur l’dlevage aseptique de l’anguillule du vinaigre (Anguillula oxophila Schneider). Rev Suisse Zool. 1921;28:357–80.
Dougherty EC, Raphael JC, Alton CH. The axenic cultivation of Rhabditis briggsae Dougherty and Nigon, 1949 (Nematoda: Rhabditidae). I. Experiments with chick embryo juice and chemically defined media. Proc Helminthol Soc Wash. 1950;17:1–10.
Dougherty EC. The axenic cultivation of Rhabditis briggsae Dougherty and Nigon, 1949 (Nematoda: Rhabditidae). II. Some sources and characteristics of “factor Rb”. Exp Parasitol. 1951b;1(1):34–45.
Dougherty EC. The axenic cultivation of Rhabditis briggsae Dougherty and Nigon, 1949 (Nematoda, Rhabditidae). 3. Liver preparations with various supplementation. J Parasitol. 1953b;39:371–80.
Dougherty EC. Some effects of urea on the liver protein used in the nutrition of Caenorhabditis briggsae (Nematoda: Rhabditidae). Anat Rec. 1954;120(3):804–5.
Dougherty EC, Keith DF. The axenic cultivation of Rhabditis briggsae Dougherty and Nigon, 1949 (Nematoda: Rhabditidae). IV. Plasma protein fractions with various supplementation. J Parasitol. 1953;39(4):381–4.
Dougherty EC. Factor Rb activity in human plasma. Nature. 1951a;168(4281):880.
Nicholas WL, Dougherty EC, Hansen EL. Axenic cultivation of Caenorhabditis briggsae (Nematoda: Rhabditidae) with chemically undefined supplements: comparative studies with related nematodes. Ann N Y Acad Sci. 1959;77(2):218–36.
Hieb WF, Rothstein M. Sterol requirement for reproduction of a free-living nematode. Science. 1968;160(3829):778.
Hieb WF, Stokstad ELR, Rothstein M. Heme requirement for reproduction of a free-living nematode. Science. 1970;168(3927):143.
Lu NC, Newton C, Stokstad ELR. The requirement of sterol and various sterol precursors in f-living nematodes. Nematologica. 1977;23(1):57–61.
Lu NC, Hugenberg G, Briggs GM, Stokstad ELR. The growth-promoting activity of several lipid-related compounds in the free-living nematode Caenorhabditis briggsae. Proc Soc Exp Biol Med. 1978;158(2):187–91.
Lu NC, Goetsch KM. Carbohydrate requirement of Caenorhabditis elegans and the final development of a chemically defined medium. Nematologica. 1993;39(1–4):303–11.
Nicholas WL. The axenic culture of Turbatrix aceti (the vinegar eelworm) 1. Nematologica. 1956;1(4):337–40.
Dougherty EC, Hansen EL. Axenic cultivation of Caenorhabditis briggsae (Nematoda: Rhabditidae). V. Maturation on synthetic media. Proc Soc Exp Biol Med. 1956c;93(2):223–7.
Dougherty EC, Hansen EL, Nicholas WL, Mollett JA, Yarwood EA. Axenic cultivation of Caenorhabditis briggsae (Nematoda: Rhabditidae) with unsupplemented and supplemented chemically defined media. Ann N Y Acad Sci. 1959;77(2):176–217.
Sayre FW, Hansen EL, Yarwood EA. Biochemical aspects of the nutrition of Caenorhabditis briggsae. Exp Parasitol. 1963;13(2):98–107.
Buecher EJ, Hansen E, Yarwood EA. Ficoll activation of a protein essential for maturation of the free-living nematode Caenorhabditis briggsae. Proc Soc Exp Biol Med. 1966;121(2):390–3.
Rothstein M, Cook E. Nematode biochemistry—VI. Conditions for axenic culture of Turbatrix aceti, Panagrellus redivivus, Rhabditis anomala and Caenorhabditis briggsae. Comp Biochem Physiol. 1966;17(2):683–92.
Vanfleteren JR, Roets DE. The influence of some anthelmintic drugs on the population growth of the free-living nematodes Caenorhabditis briggsae and Turbatrix aceti (Nematoda : Rhabditida). Nematologica. 1972;18(3):325–38.
Rothstein M. Practical methods for the axenic culture of the free-living nematodes Turbatrix aceti and Caenorhabditis briggsae. Comp Biochem Physiol B. 1974;49(4):669–78.
Hieb WF, Rothstein M. Aging in the free-living nematode Turbatrix aceti. Techniques for synchronization and aging of large-scale axenic cultures. Exp Gerontol. 1975;10(2):145–53.
Sayre FW, Cryan WS, Yarwood EA, Hansen E, Martin M. Axenic cultivation of the dioecious nematode Panagrellus redivivus. Nematologica. 1963;9(3):313–9.
Hansen EL, Buecher EJ. Biochemical approach to systematic studies with axenic nematodes. J Nematol. 1970;2(1):1–6.
Hansen EL, Buecher EJ, Evans AAF. Axenic culture of Aphelenchus avenae. Nematologica. 1970;16(2):328–9.
Perelman D, Lu NC. Requirements for branched chain amino acids and their interactions in the nematode Caenorhabditis elegans. Nematology. 2000;2(5):501–6.
Szewczyk NJ, Kozak E, Conley CA. Chemically defined medium and Caenorhabditis elegans. BMC Biotechnol. 2003;3(1):19.
Zhao Y, Lu CN. Requirements of non-haem iron and haem iron in the nematode Caenorhabditis elegans. Nematology. 2011;13(7):853–8.
Vanfleteren JR. Nematodes as nutritional models. In: Zuckerman BM, editor. Nematodes as biological models, vol. 2. New York: Academic Press; 1980.
Houthoofd K, Braeckman BP, Lenaerts I, Brys K, De Vreese A, Van Eygen S, Vanfleteren JR. Axenic growth up-regulates mass-specific metabolic rate, stress resistance, and extends life span in Caenorhabditis elegans. Exp Gerontol. 2002;37(12):1371–8.
Clegg E, Lapenotiere H, French D, Szilagyi M. Use of CeHR axenic medium for exposure and gene expression studies. In: East Coast Worm Meeting; 2002.
Buecher EJ, Hansen EL. Yeast extract as a supplement to chemically defined medium for axenic culture of Caenorhabditis briggsae. Experientia. 1969;25(6):656.
Buecher EJ, Hansen EL, Yarwood EA. Growth of nematodes in defined medium containing hemin and supplemented with commercially available proteins. Nematologica. 1970c;16(3):403–9.
Buecher EJ, Hansen EL, Yarwood EA. Cultivation of Caenorhabditis briggsae and Turbatrix aceti with defined proteins. J Nematol. 1971;3(1):89–90.
Flavel MR, Mechler A, Shahmiri M, Mathews ER, Franks AE, Chen W, Zanker D, Xian B, Gao S, Luo J, et al. Growth of Caenorhabditis elegans in defined media is dependent on presence of particulate matter. G3 (Bethesda, Md). 2017;8(2):567–75.
Hansen EL, Perez-Mendez G, Buecher EJ. Glycogen as a supplement in media for axenic cultivation of nematodes. Proc Soc Exp Biol Med. 1971;137(4):1352–4.
Vanfleteren JR. Nematode growth factor. Nature. 1974;248(5445):255–7.
Abderhalden E. Synthese der Zellbausteine in Pflanze und Tier. In: Synthese der Zellbausteine in Pflanze und Tier. Berlin: Springer-Verlag; 1921. p. 1–112.
Rose WC. The nutritive significance of the amino acids. Physiol Rev. 1938;18(1):109–36.
Miflin BJ, Lea PJ. Amino acid metabolism. Annu Rev Plant Physiol. 1977;28(1):299–329.
Cooper T. Nitrogen metabolism in Saccharomyces cerevisiae. In: SJ N, JE W, BJ R, editors. The molecular biology of the yeast saccharomyces: metabolism and gene expression. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 1982a. p. 39–100.
Payne SH, Loomis WF. Retention and loss of amino acid biosynthetic pathways based on analysis of whole-genome sequences. Eukaryot Cell. 2006;5(2):272–6.
Nicholas WL, Dougherty EC, Hansen EL, Holm-Hansen O, Moses V. The incorporation of 14C from sodium acetate-2-14C into the amino acids of the soil-inhabiting nematode, Caenorhabditis briggsae. J Exp Biol. 1960;37(3):435.
Rothstein M, Tomlinson GA. Biosynthesis of amino acids by the nematode Caenorhabditis briggsae. Biochim Biophys Acta. 1961;49(3):625–7.
Rothstein M, Tomlinson G. Nematode biochemistry: II. Biosynthesis of amino acids. Biochim Biophys Acta. 1962;63(3):471–80.
Vanfleteren JR. Amino acid requirements of the free-living nematode Caenorhabditis Briggsae. Nematologica. 1973;19(1):93–9.
Albanese AA. The protein and amino acid requirements of man. In: Albanese AA, editor. Protein and amino acid requirements of mammals. New York: Academic Press; 1950. p. 115–51.
Cynober L, Boucher JL, Vasson M-P. Arginine metabolism in mammals. J Nutr Biochem. 1995;6(8):402–13.
Morris SM. Regulation of enzymes of the urea cycle and arginine metabolism. Annu Rev Nutr. 2002;22(1):87–105.
Schulz TJ, Zarse K, Voigt A, Urban N, Birringer M, Ristow M. Glucose restriction extends Caenorhabditis elegans life span by inducing mitochondrial respiration and increasing oxidative stress. Cell Metab. 2007;6(4):280–93.
Schlotterer A, Kukudov G, Bozorgmehr F, Hutter H, Du X, Oikonomou D, Ibrahim Y, Pfisterer F, Rabbani N, Thornalley P, et al. C. elegans as model for the study of high glucose-mediated life span reduction. Diabetes. 2009;58(11):2450.
Lee S-J, Murphy CT, Kenyon C. Glucose shortens the life span of C. elegans by downregulating DAF-16/FOXO activity and aquaporin gene expression. Cell Metab. 2009;10(5):379–91.
Hibuse T, Maeda N, Funahashi T, Yamamoto K, Nagasawa A, Mizunoya W, Kishida K, Inoue K, Kuriyama H, Nakamura T, et al. Aquaporin 7 deficiency is associated with development of obesity through activation of adipose glycerol kinase. Proc Natl Acad Sci U S A. 2005;102(31):10993.
Mondoux MA, Love DC, Ghosh SK, Fukushige T, Bond M, Weerasinghe GR, Hanover JA, Krause MW. O-linked-N-acetylglucosamine cycling and insulin signaling are required for the glucose stress response in Caenorhabditis elegans. Genetics. 2011;188(2):369.
Alcántar-Fernández J, Navarro RE, Salazar-Martínez AM, Pérez-Andrade ME, Miranda-Ríos J. Caenorhabditis elegans respond to high-glucose diets through a network of stress-responsive transcription factors. PLOS ONE. 2018;13(7):e0199888.
Lee D, Jeong D-E, Son HG, Yamaoka Y, Kim H, Seo K, Khan AA, Roh T-Y, Moon DW, Lee Y, et al. SREBP and MDT-15 protect C. elegans from glucose-induced accelerated aging by preventing accumulation of saturated fat. Genes Dev. 2015;29(23):2490–503.
Bécard D, Hainault I, Azzout-Marniche D, Bertry-Coussot L, Ferré P, Foufelle F. Adenovirus-mediated overexpression of sterol regulatory element binding protein-1c mimics insulin effects on hepatic gene expression and glucose homeostasis in diabetic mice. Diabetes. 2001;50(11):2425.
Matsuzaka T, Shimano H, Yahagi N, Amemiya-Kudo M, Okazaki H, Tamura Y, Iizuka Y, Ohashi K, Tomita S, Sekiya M, et al. Insulin-independent induction of sterol regulatory element-binding protein-1c expression in the livers of streptozotocin-treated mice. Diabetes. 2004;53(3):560.
Bollen M, Keppens S, Stalmans W. Specific features of glycogen metabolism in the liver. Biochem J. 1998;336(Pt 1) (Pt 1):19–31.
Frazier HN III, Roth MB. Adaptive sugar provisioning controls survival of C. elegans embryos in adverse environments. Curr Biol. 2009;19(10):859–63.
Possik E, Ajisebutu A, Manteghi S, Gingras M-C, Vijayaraghavan T, Flamand M, Coull B, Schmeisser K, Duchaine T, van Steensel M, et al. FLCN and AMPK confer resistance to hyperosmotic stress via remodeling of glycogen stores. PLoS Genet. 2015;11(10):e1005520.
Seo Y, Kingsley S, Walker G, Mondoux MA, Tissenbaum HA. Metabolic shift from glycogen to trehalose promotes lifespan and healthspan in Caenorhabditis elegans. Proc Natl Acad Sci. 2018;115(12):E2791.
Burnell AM, Houthoofd K, O'Hanlon K, Vanfleteren JR. Alternate metabolism during the dauer stage of the nematode Caenorhabditis elegans. Exp Gerontol. 2005;40(11):850–6.
Pellerone FI, Archer SK, Behm CA, Grant WN, Lacey MJ, Somerville AC. Trehalose metabolism genes in Caenorhabditis elegans and filarial nematodes. Int J Parasitol. 2003;33(11):1195–206.
Hanover JA, Forsythe ME, Hennessey PT, Brodigan TM, Love DC, Ashwell G, Krause M. A Caenorhabditis elegans model of insulin resistance: altered macronutrient storage and dauer formation in an OGT-1 knockout. Proc Natl Acad Sci U S A. 2005;102(32):11266.
Richards AB, Krakowka S, Dexter LB, Schmid H, Wolterbeek APM, Waalkens-Berendsen DH, Shigoyuki A, Kurimoto M. Trehalose: a review of properties, history of use and human tolerance, and results of multiple safety studies. Food Chem Toxicol. 2002;40(7):871–98.
Perez CL, Van Gilst MR. A 13C isotope labeling strategy reveals the influence of insulin signaling on lipogenesis in C. elegans. Cell Metab. 2008;8(3):266–74.
Satouchi K, Hirano K, Sakaguchi M, Takehara H, Matsuura F. Phospholipids from the free-living nematode Caenorhabditis elegans. Lipids. 1993;28(9):837–40.
Watts JL, Browse J. Genetic dissection of polyunsaturated fatty acid synthesis in Caenorhabditis elegans. Proc Natl Acad Sci U S A. 2002;99(9):5854–9.
Wallis JG, Watts JL, Browse J. Polyunsaturated fatty acid synthesis: what will they think of next? Trends Biochem Sci. 2002;27(9):467–73.
Rothstein M, Götz P. Biosynthesis of fatty acids in the free-living nematode, Turbatrix aceti. Arch Biochem Biophys. 1968;126(1):131–40.
Rothstein M. Nematode biochemistry: XI, biosynthesis of fatty acids by Caenorhabditis briggsae and Panagrellus redivivus. Int J BioChemiPhysics. 1970;1(4):422–8.
Wallis JG, Browse J. Mutants of Arabidopsis reveal many roles for membrane lipids. Prog Lipid Res. 2002;41(3):254–78.
Schmidt A, Wolde M, Thiele C, Fest W, Kratzin H, Podtelejnikov AV, Witke W, Huttner WB, Söling H-D. Endophilin I mediates synaptic vesicle formation by transfer of arachidonate to lysophosphatidic acid. Nature. 1999;401:133.
El-Yassimi A, Hichami A, Besnard P, Khan NA. Linoleic acid induces calcium signaling, Src kinase phosphorylation, and neurotransmitter release in mouse CD36-positive gustatory cells. J Biol Chem. 2008;283(19):12949–59.
De Renobales M, Ryan RO, Heisler CR, McLean DL, Blomquist GJ. Linoleic acid biosynthesis in the pea aphid, Acyrthosiphon pisum (Harris). Arch Insect Biochem Physiol. 1986;3(2):193–203.
Shimizu N, Naito M, Mori N, Kuwahara Y. De novo biosynthesis of linoleic acid and its conversion to the hydrocarbon (Z,Z)-6,9-heptadecadiene in the astigmatid mite, Carpoglyphus lactis: incorporation experiments with 13C-labeled glucose. Insect Biochem Mol Biol. 2014;45:51–7.
Weinert J, Blomquist GJ, Borgeson CE. De novo biosynthesis of linoleic acid in two non-insect invertebrates: the land slug and the garden snail. Experientia. 1993;49(10):919–21.
Malcicka M, Visser B, Ellers J. An evolutionary perspective on linoleic acid synthesis in animals. Evol Biol. 2018;45(1):15–26.
Tocher DR, Leaver MJ, Hodgson PA. Recent advances in the biochemistry and molecular biology of fatty acyl desaturases. Prog Lipid Res. 1998;37(2):73–117.
Spychalla JP, Kinney AJ, Browse J. Identification of an animal omega-3 fatty acid desaturase by heterologous expression in Arabidopsis. Proc Natl Acad Sci U S A. 1997;94(4):1142–7.
Peyou-Ndi MM, Watts JL, Browse J. Identification and characterization of an animal Δ12 fatty acid desaturase gene by heterologous expression in Saccharomyces cerevisiae. Arch Biochem Biophys. 2000;376(2):399–408.
Zhou X-R, Green AG, Singh SP. Caenorhabditis elegans Δ12-desaturase FAT-2 is a bifunctional desaturase able to desaturate a diverse range of fatty acid substrates at the Δ12 and Δ15 positions. J Biol Chem. 2011;286(51):43644–50.
Lesa GM, Palfreyman M, Hall DH, Clandinin MT, Rudolph C, Jorgensen EM, Schiavo G. Long chain polyunsaturated fatty acids are required for efficient neurotransmission in C. elegans. J Cell Sci. 2003;116(24):4965.
Kniazeva M, Crawford QT, Seiber M, Wang C-Y, Han M. Monomethyl branched-chain fatty acids play an essential role in Caenorhabditis elegans development. PLoS Biol. 2004;2(9):e257.
Entchev EV, Schwudke D, Zagoriy V, Matyash V, Bogdanova A, Habermann B, Zhu L, Shevchenko A, Kurzchalia TV. LET-767 is required for the production of branched chain and long chain fatty acids in Caenorhabditis elegans. J Biol Chem. 2008;283(25):17550–60.
Volpe JJ, Vagelos PR. Mechanisms and regulation of biosynthesis of saturated fatty acids. Physiol Rev. 1976;56(2):339–417.
Oku H, Kaneda T. Biosynthesis of branched-chain fatty acids in Bacillus subtilis. A decarboxylase is essential for branched-chain fatty acid synthetase. J Biol Chem. 1988;263(34):18386–96.
Zhang B, Zhao Y, Harris RA, Crabb DW. Molecular defects in the E1 alpha subunit of the branched-chain alpha-ketoacid dehydrogenase complex that cause maple syrup urine disease. Mol Biol Med. 1991;8(1):39–47.
Strauss KA, Morton DH. Branched-chain ketoacyl dehydrogenase deficiency: maple syrup disease. Curr Treat Options Neurol. 2003;5(4):329–41.
Jia F, Cui M, Than MT, Han M. Developmental defects of Caenorhabditis elegans lacking branched-chain α-Ketoacid dehydrogenase are mainly caused by monomethyl branched-chain fatty acid deficiency. J Biol Chem. 2016;291(6):2967–73.
Mak HY. Lipid droplets as fat storage organelles in Caenorhabditis elegans: thematic review series: lipid droplet synthesis and metabolism: from yeast to man. J Lipid Res. 2012;53(1):28–33.
Kühnlein RP. Thematic review series: lipid droplet synthesis and metabolism: from yeast to man. Lipid droplet-based storage fat metabolism in Drosophila. J Lipid Res. 2012;53(8):1430–6.
Radulovic M, Knittelfelder O, Cristobal-Sarramian A, Kolb D, Wolinski H, Kohlwein SD. The emergence of lipid droplets in yeast: current status and experimental approaches. Curr Genet. 2013;59(4):231–42.
Murphy DJ. The dynamic roles of intracellular lipid droplets: from archaea to mammals. Protoplasma. 2012;249(3):541–85.
Mullaney BC, Ashrafi K. C. elegans fat storage and metabolic regulation. Biochim Biophys Acta. 2009;1791(6):474–8.
Coelho M, Oliveira T, Fernandes R. Biochemistry of adipose tissue: an endocrine organ. Arch Med Sci. 2013;9(2):191–200.
Birsoy K, Festuccia WT, Laplante M. A comparative perspective on lipid storage in animals. J Cell Sci. 2013;126(7):1541.
Liu Z, Li X, Ge Q, Ding M, Huang X. A lipid droplet-associated GFP reporter-based screen identifies new fat storage regulators in C. elegans. J Genet Genomics. 2014;41(5):305–13.
O'Riordan VB, Burnell AM. Intermediary metabolism in the dauer larva of the nematode Caenorhabditis elegans— 1. Glycolysis, gluconeogenesis, oxidative phosphorylation and the tricarboxylic acid cycle. Comp Biochem Physiol B. 1989;92(2):233–8.
Kimura KD, Tissenbaum HA, Liu Y, Ruvkun G. daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science. 1997;277(5328):942.
Rothstein M, Mayoh H. Nematode biochemistry: IV. On isocitrate lyase in Caenorhabditis briggsae. Arch Biochem Biophys. 1964;108(1):134–42.
Rothstein M, Mayoh H. Nematode biochemistry—VII. Presence of isocitrate lyase in Panagrellus redivivus, turbatrix aceti, and Rhabditis anomala. Comp Biochem Physiol. 1965;16(4):361–5.
Rothstein M, Mayoh H. Nematode biochemistry—VIII malate synthetase. Comp Biochem Physiol. 1966;17(4):1181–6.
Alaimo JT, Davis SJ, Song SS, Burnette CR, Grotewiel M, Shelton KL, Pierce-Shimomura JT, Davies AG, Bettinger JC. Ethanol metabolism and osmolarity modify behavioral responses to ethanol in C. elegans. Alcohol Clin Exp Res. 2012;36(11):1840–50.
Castro PV, Khare S, Young BD, Clarke SG. Caenorhabditis elegans battling starvation stress: low levels of ethanol prolong lifespan in L1 larvae. PLoS One. 2012;7(1):e29984.
Braeckman BP, Houthoofd K, Vanfleteren JR. Intermediary metabolism. WormBook; 2009.
Davis WL, Jones RG, Goodman DB. Cytochemical localization of malate synthase in amphibian fat body adipocytes: possible glyoxylate cycle in a vertebrate. J Histochem Cytochem. 1986;34(5):689–92.
Davis WL, Jones RG, Farmer GR, Dickerson T, Cortinas E, Cooper OJ, Crawford L, Goodman DBP. Identification of glyoxylate cycle enzymes in chick liver—the effect of vitamin D3: Cytochemistry and biochemistry. Anat Rec. 1990;227(3):271–84.
Morgunov IG, Kondrashova MN, Kamzolova SV, Sokolov AP, Fedotcheva NI, Finogenova TV. Evidence of the glyoxylate cycle in the liver of newborn rats. Med Sci Monit. 2005;11(2):BR57–60.
Kondrashov FA, Koonin EV, Morgunov IG, Finogenova TV, Kondrashova MN. Evolution of glyoxylate cycle enzymes in metazoa: evidence of multiple horizontal transfer events and pseudogene formation. Biol Direct. 2006;1:31.
Dutky SR, Robbins WF, Thompson JV. The demonstration of sterols as requirements for the growth, development and reproduction of the DD- 136 nematode. Nematologica. 1967;13:140.
Cole RJ, Dutky SR. A sterol requirement in Turbatrix aceti and Panagrellus redivivus. J Nematol. 1969;1(1):72–5.
Cole RJ, Krusberg LR. Sterol composition of the nematodes Ditylenchus triformis and Ditylenchus dipsaci, and host tissues. Exp Parasitol. 1967;21(2):232–9.
Rothstein M. Nematode biochemistry—IX. Lack of sterol biosynthesis in free-living nematodes. Comp Biochem Physiol. 1968;27(1):309–17.
Chitwood DJ, Lozano R, Lusby WR. Recent developments in nematode steroid biochemistry. J Nematol. 1986;18(1):9–17.
Kurzchalia TV, Ward S. Why do worms need cholesterol? Nat Cell Biol. 2003;5:684.
Chitwood DJ. Biochemistry and function of nematode steroids. Crit Rev Biochem Mol Biol. 1999;34(4):273–84.
Espenshade PJ, Hughes AL. Regulation of sterol synthesis in eukaryotes. Annu Rev Genet. 2007;41(1):401–27.
Lozano R, Salt TA, Chitwood DJ, Lusby WR, Thompson MJ. Metabolism of sterols of varying ring unsaturation and methylation by Caenorhabditis elegans. Lipids. 1987;22(2):84–7.
Merris M, Wadsworth WG, Khamrai U, Bittman R, Chitwood DJ, Lenard J. Sterol effects and sites of sterol accumulation in Caenorhabditis elegans: developmental requirement for 4α-methyl sterols. J Lipid Res. 2003;44(1):172–81.
Matyash V, Geier C, Henske A, Mukherjee S, Hirsh D, Thiele C, Grant B, Maxfield FR, Kurzchalia TV. Distribution and transport of cholesterol in Caenorhabditis elegans. Mol Biol Cell. 2001;12(6):1725–36.
Antebi A, Yeh WH, Tait D, Hedgecock EM, Riddle DL. daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans. Genes Dev. 2000;14(12):1512–27.
Motola DL, Cummins CL, Rottiers V, Sharma KK, Li T, Li Y, Suino-Powell K, Xu HE, Auchus RJ, Antebi A, et al. Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell. 2006;124(6):1209–23.
Dumas KJ, Guo C, Shih H-J, Hu PJ. Influence of steroid hormone signaling on life span control by Caenorhabditis elegans insulin-like signaling. G3. 2013;3(5):841.
Aguilaniu H, Fabrizio P, Witting M. The role of dafachronic acid signaling in development and longevity in Caenorhabditis elegans: digging deeper using cutting-edge analytical chemistry. Front Endocrinol. 2016;7:12.
Jia K, Albert PS, Riddle DL. DAF-9, a cytochrome P450 regulating C. elegans larval development and adult longevity. Development. 2002;129(1):221.
Sharma V, Hiller M. Loss of enzymes in the bile acid synthesis pathway explains differences in bile composition among mammals. Genome Biol Evol. 2018;10(12):3211–7.
Rothstein M, Coppens M. Nutritional factors and conditions for the axenic culture of free-living nematodes. Comp Biochem Physiol B. 1978;61(1):99–104.
Rao AU, Carta LK, Lesuisse E, Hamza I. Lack of heme synthesis in a free-living eukaryote. Proc Natl Acad Sci U S A. 2005;102(12):4270–5.
Ajioka RS, Phillips JD, Kushner JP. Biosynthesis of heme in mammals. BBA - Mol Cell Res. 2006;1763(7):723–36.
Rajagopal A, Rao AU, Amigo J, Tian M, Upadhyay SK, Hall C, Uhm S, Mathew MK, Fleming MD, Paw BH, et al. Haem homeostasis is regulated by the conserved and concerted functions of HRG-1 proteins. Nature. 2008;453(7198):1127–31.
Severance S, Hamza I. Trafficking of heme and porphyrins in metazoa. Chem Rev. 2009;109(10):4596–616.
Chen C, Samuel TK, Sinclair J, Dailey HA, Hamza I. An intercellular heme-trafficking protein delivers maternal heme to the embryo during development in C. elegans. Cell. 2011;145(5):720–31.
Yuan X, Protchenko O, Philpott CC, Hamza I. Topologically conserved residues direct heme transport in HRG-1-related proteins. J Biol Chem. 2012;287(7):4914–24.
Chen C, Samuel TK, Krause M, Dailey HA, Hamza I. Heme utilization in the Caenorhabditis elegans hypodermal cells is facilitated by heme-responsive gene-2. J Biol Chem. 2012;287(12):9601–12.
Korolnek T, Zhang J, Beardsley S, Scheffer GL, Hamza I. Control of metazoan heme homeostasis by a conserved multidrug resistance protein. Cell Metab. 2014;19(6):1008–19.
Dougherty EC, Hansen E. He folic acid requirement and its antagonism by aminopterin in the nematode Caenorhabditis briggsae (Rhabditidae). Anat Rec. 1957a;128:541–2.
Vanfleteren JR, Avau H. Selective inhibition of reproduction in aminopterin-treated nematodes. Experientia. 1977;33(7):902–4.
Lu NC, Hieb WF, Stokstad ELR. Accumulation of Formimino-l-glutamic acid in the free-living nematode Caenorhabditis briggsae as related to folic acid deficiency. Proc Soc Exp Biol Med. 1974;145(1):67–9.
Ducker GS, Rabinowitz JD. One-carbon metabolism in health and disease. Cell Metab. 2017;25(1):27–42.
Bailey LB, Stover PJ, McNulty H, Fenech MF, Gregory JF 3rd, Mills JL, Pfeiffer CM, Fazili Z, Zhang M, Ueland PM, et al. Biomarkers of nutrition for development-folate review. J Nutr. 2015;145(7):1636S–80S.
Nguyen TT, Dyer DL, Dunning DD, Rubin SA, Grant KE, Said HM. Human intestinal folate transport: cloning, expression, and distribution of complementary RNA. Gastroenterology. 1997;112(3):783–91.
Balamurugan K, Ashokkumar B, Moussaif M, Sze JY, Said HM. Cloning and functional characterization of a folate transporter from the nematode Caenorhabditis elegans. Am J Phys Cell Physiol. 2007;293(2):C670–81.
Nicholas WL, Hansen E, Dougherty EC. The B-vitamins required by Caenorhabditis briggsae (Rhabditidae). Nematologica. 1962;8(2):129–35.
Nicholas WL, Jantunen R. A biotin requirement for Caenorhabditis Briggsae (Rhabditidae). Nematologica. 1963;9(3):332–6.
Lu CN, Hieb WF, Stokstad ELR. Effect of vitamin B12 and folate on biosynthesis of methionine from homocysteine in the nematode Caenorhabditis briggsae. Proc Soc Exp Biol Med. 1976;151:701–6.
Bito T, Matsunaga Y, Yabuta Y, Kawano T, Watanabe F. Vitamin B12 deficiency in Caenorhabditis elegans results in loss of fertility, extended life cycle, and reduced lifespan. FEBS Open Bio. 2013;3:112–7.
Bito T, Misaki T, Yabuta Y, Ishikawa T, Kawano T, Watanabe F. Vitamin B(12) deficiency results in severe oxidative stress, leading to memory retention impairment in Caenorhabditis elegans. Redox Biol. 2016;11:21–9.
Na H, Ponomarova O, Giese GE, Walhout AJM. C. elegans MRP-5 exports vitamin B12 from mother to offspring to support embryonic development. Cell Rep. 2018;22(12):3126–33.
Rodionov DA, Vitreschak AG, Mironov AA, Gelfand MS. Comparative genomics of the vitamin B12 metabolism and regulation in prokaryotes. J Biol Chem. 2003;278(42):41148–59.
Yilmaz LS, Walhout AJM. Worms, bacteria, and micronutrients: an elegant model of our diet. Trends Genet. 2014;30(11):496–503.
Watanabe T, Ohkawa K, Kasai S, Ebara S, Nakano Y, Watanabe Y. The effects of dietary vitamin B12 deficiency on sperm maturation in developing and growing male rats. Congenit Anom. 2003;43(1):57–64.
Pront R, Margalioth EJ, Green R, Eldar-Geva T, Maimoni Z, Zimran A, Elstein D. Prevalence of low serum cobalamin in infertile couples. Andrologia. 2009;41(1):46–50.
Fleet JC, Replogle R, Salt DE. Systems genetics of mineral metabolism. J Nutr. 2011;141(3):520–5.
Wada O. What are trace elements? Their deficiency and excess states. Japan Med Assoc J. 2004;47(8):351–8.
Lu CN, Cheng AC, Briggs GM. A study of mineral requirements in Caenorhabditis elegans. Nematologica. 1983;29(4):425–34.
Hester J, Hanna-Rose W, Diaz F. Zinc deficiency reduces fertility in C. elegans hermaphrodites and disrupts oogenesis and meiotic progression. Comp Biochem Physiol C Toxicol Pharmacol. 2017;191:203–9.
Hidiroglou M, Knipfel JE. Zinc in mammalian sperm: a review. J Dairy Sci. 1984;67(6):1147–56.
Kim AM, Vogt S, O'Halloran TV, Woodruff TK. Zinc availability regulates exit from meiosis in maturing mammalian oocytes. Nat Chem Biol. 2010;6(9):674–81.
Kumar J, Barhydt T, Awasthi A, Lithgow GJ, Killilea DW, Kapahi P. Zinc levels modulate lifespan through multiple longevity pathways in Caenorhabditis elegans. PLoS One. 2016;11(4):e0153513.
Acknowledgements
Not applicable
Funding
AZ is funded by Special Research Fund (BOF) of Ghent University, project BOF2-4J. ID is funded by H2020 project ‘Ageing with elegans’, GA633589.
Availability of data and materials
Not applicable
Author information
Authors and Affiliations
Contributions
This manuscript was drafted by AZ. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Zečić, A., Dhondt, I. & Braeckman, B.P. The nutritional requirements of Caenorhabditis elegans. Genes Nutr 14, 15 (2019). https://doi.org/10.1186/s12263-019-0637-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12263-019-0637-7