
Abstract
Root-knot nematodes (RKNs) are a vital pest that causes significant yield losses and economic damage to potato plants. The use of chemical pesticides to control these nematodes has led to environmental concerns and the development of resistance in the nematode populations. Endophytic fungi offer an eco-friendly alternative to control these pests and produce secondary metabolites that have nematicidal activity against RKNs. The objective of this study is to assess the efficacy of Aspergillus flavus (ON146363), an entophyte fungus isolated from Trigonella foenum-graecum seeds, against Meloidogyne incognita in filtered culture broth using GC–MS analysis. Among them, various nematicidal secondary metabolites were produced: Gadoleic acid, Oleic acid di-ethanolamide, Oleic acid, and Palmitic acid. In addition, biochemical compounds such as Gallic acid, Catechin, Protocatechuic acid, Esculatin, Vanillic acid, Pyrocatechol, Coumarine, Cinnamic acid, 4, 3-indol butyl acetic acid and Naphthyl acetic acid by HPLC. The fungus was identified through morphological and molecular analysis, including ITS 1–4 regions of ribosomal DNA. In vitro experiments showed that culture filtrate of A. flavus had a variable effect on reducing the number of egg hatchings and larval mortality, with higher concentrations showing greater efficacy than Abamectin. The fungus inhibited the development and multiplication of M. incognita in potato plants, reducing the number of galls and eggs by 90% and 89%, respectively. A. flavus increased the activity of defense-related enzymes Chitinas, Catalyse, and Peroxidase after 15, 45, and 60 days. Leaching of the concentrated culture significantly reduced the second juveniles’ stage to 97% /250 g soil and decreased the penetration of nematodes into the roots. A. flavus cultural filtrates via soil spraying improved seedling growth and reduced nematode propagation, resulting in systemic resistance to nematode infection. Therefore, A. flavus can be an effective biological control agent for root-knot nematodes in potato plants. This approach provides a sustainable solution for farmers and minimizes the environmental impact.
Similar content being viewed by others

Introduction
Potato (Solanum tuberosum L.) is considered a vital root crop, the fourth staple food, and an economically critical food crop after maize, rice, and wheat1,2. About 80% of the world's potato production is cultivated in several regions, including Europe, America, and Asia3. Potato is grown in an area of 20 million hectares and produces nearly 400 million tons annually4,5. In Egypt, 6.2 million tons of potatoes are from the cultivated area of 211 thousand hectares, with an average yield of 24.100 t ha-1 and a self-sufficiency rate of 111.4%6. Currently, there is an increasing need for food supply and food security. Unsustainable crop production approaches with monocultures, intensive planting, and expansion of crops into newly opened areas have enhanced problems related to new pests and diseases7,8; one of these essential crops is potato which infected by different pests and pathogens causing high yield losses such as Globodera spp., Meloidogyne spp., Ditylenchus spp. and Pratylenchus spp which are recognized as the main parasites infected potato9,10. Meloidogyne spp or nematodes are the most critical factors in reducing quantitative and qualitative yields in potatoes11,12. Nematodes are a restricting factor for potato production, reducing yields and chemical changes in potato tubers5,10. Nematodes caused an average yield loss of up to 12% in potatoes13,14,15. Moreover, many other species of infected potatoes, such as Belonolaimus, Longidorus, Xiphinema, Rotylenchulus spp, and Radopholus similis, are considered of minor importance in potato production16.
Recently, biological control of plant pests has increased17,18; most biological control endophyte (BCE) agents are commonly found among crops and can suppress critical pathogens19,20. Fungal pathogens have a harmful effect on plant physiology, which increases host defense responses to pathogens21. Fungi produce bioactive molecules, such as small peptide effectors, enzymes, and secondary metabolites that assist colonization and contribute to both symbiotic and defense against pathogenic relationships22. Trichoderma species, arbuscular mycorrhizas, ectomycorrhizas, endophytes, and yeasts of specific pathogens are among the significant beneficial fungi with biocontrol capacity23,24. Microbial pesticide active ingredients are microorganisms such as fungi, bacteria, viruses, protozoans, or algae with a broad spectrum of activity. However, every organism is relatively specific to its target pest25,26.
Microorganisms and nematodes interact with plant host roots to obtain their nutrients27,28. This interactive relationship is not unidirectional but multi-directional, with all these organisms interacting with each other either singly or in combination29,30. The fungus culture activity includes nematicidal effects on adult nematodes, inhibition of egg hatching and juvenile development31. Culture filtrates of numerous soil-borne fungi exhibited nematode-toxic characteristics32, though most of the initial studies were pointed to in vitro tests33,34. There is comparatively little information on the toxic effects of fungal metabolites on the development and reproduction of root-knot nematodes on plant growth35. Therefore, the objectives of the current study are to evaluate the efficacy of Aspergillus flavus, an entophyte fungus isolated from Trigonella foenum-graecum seeds, against Meloidogyne incognita.
Materials and methods
Source of endophytic fungi (Phytopathogens used for antagonistic assays)
The seeds of Trigonella foenum-graecum were sterilized using a sequential immersion method, which involved 1 min in 75% ethanol, followed by 10 min in 65% commercial Chlorox (final concentration of 3.25% aqueous sodium hypochlorite), and finally, a 30-s immersion in 75% ethanol36,37,38. The sterilized seeds were planted on potato dextrose agar (PDA) medium supplemented with cidocetine (500 mg/L). As well, the endophytic fungus was sub-cultured in demand to gain pure cultures. Identification of the fungal strain was detected using morphology of cultures or hyphae, the characteristics of the spores and reproductive structures if the characteristic, were visible. The isolated pure fungus was cultured on a potato dextrose agar plate and inoculated at 26 ± 2 °C in complete darkness After 5 to 7 day, about 5 mm diameter mycelia disc of fungus was inoculated into 250 mL of PDB in a 500 mL Erlenmeyer flask and incubated at 26 ± 2 °C for one week. The culture filtrate of A. flavus was prepared by centrifuging at 5000 rpm/15 min39. 10 ml of the supernatant was filtered by sterile 0.22 μm filter (Whatman, Clifon, NJ, USA) for the In vitro and In vivo experiment.
Molecular characterization
Preparation of DNA and ITS-PCR conditions
A. flavus mycelium was harvested from a potato dextrose liquid medium and was ground in liquid nitrogen using a pestle mortar to extract DNA40. Amplifications were carried out in PCR reactions in a total volume of 20 μl, containing 2 µl genomic DNA; 12.5 master mix PCR; and 8 pmol of specific primer ITS1 (forward): 5′-TCCGTAGGTGAACCTGCGG-3′ and ITS4 primer (Revers): 5′- TCCTCCGCTTATTGATATGC-3′ was done41.
PCR amplification was carried out according to the following temperature: 55 s at 94 °C as an initial step, 40 cycles of 60 s at 94 °C, 90 s at 52 °C, and 5 min at 72 °C, and a final step of 5 min at 72 °C. Electrophoresis of PCR-amplified products was performed in 1% agarose for 50 min at 80–100 Volt. PCR products were stained with 2 µl of Ethidium Bromide and visualized with 250 nm ultraviolet light. A Gel purification kit was used to amplify the purified DNA, and the purified DNA product was collected in a 2 ml Eppendorf plastic tube and stored at – 20 °C for sequencing. The PCR products were forwarded to Macrogen sequencing service (Macrogen Inc. Seoul, South Korea) to perform direct sequencing of both DNA strands.
Identification of fungal isolation
The BLAST program available from NCBI was used to identify the isolate by aligning the forward and reverse DNA strand sequences42. The amplified ITS rDNA fragment sequence was compared with NCBI non-redundant nucleotide sequence database using BLAST. Isolate identification was based on his analysis of megaBLAST output, which identified highly similar sequences. The hits of ITS rDNA sequences were then used in a phylogenetic analysis to determine the evolutionary relationship among the isolates and hits based on their ITS rDNA sequence43.
Phylogenetic analysis
The BLAST program was used to find similar sequences in GenBank using the sequence as a query. The most similar reference sequences were found and prepared for further phylogenetic analyses. These sequences were associated with the program CLUSTALX44, and the MEGA software was used to construct the relevant phylogenetic tree45. Ambiguous positions were removed, and gap positions were deleted manually. The neighbor-joining method was used to analyze the alignment data, with 1,000 iterations of random stepwise sequence addition performed for each query. Bootstrap analysis with 1,000 replications was used to estimate statistical support for the internal branches.
Phytopathogenic nematodes cultures and antagonistic assay In vitro
M. incognita was maintained on the roots of susceptible tomato plants (cultivar Alisa) in sterilized soil at 25 ± 2◦C in a greenhouse. The nematode eggs were extracted from the tomato roots. Each day, second-stage juveniles (J2s) of M. incognita were taken from the eggs and kept at a temperature of 15˚C. This process provided a consistent and abundant source of J2s and egg masses for use in the laboratory and greenhouse conducted by46,47. Freshly hatched juveniles were utilized in experiments to detect juvenile mortality activity. The J2s of M. incognita were utilized to detect the nematicidal activities of fungal filtrate. To prepare the enrichment media broth of a fungal strain, potato dextrose broth (PDB) medium was used and shaken at 180 rpm for five days at a temperature of 28 ± 2 °C. After centrifugation at 5000 × g for 10 min and filtering with blotting paper. The supernatant (spore suspensions) was prepared, and their concentrations (100, 75, 50, 25, and 10%) were modified to 108 spores ml−1 and then examined against J2s and eggs hatchability (EH) of RKN. A nematode suspension containing 50 J2s was added to each vial with 9 mL of the prepared fungal cell-free supernatant. In contrast, another vial contained only the J2s suspension mixed with PDB medium as a control. Each treatment had three vials and was incubated at 25 ± 2 °C for 72 h. The number of dead and alive J2s was observed using a stereoscopic microscope at different time intervals from exposure time. Dead nematodes were identified if they did not move when probed with a fine needle, while active ones moved or appeared as winding shapes. The nematodes in each treatment were then transferred to distilled water for another 48 h to determine whether the dead ones regained mobility. The mortality (%) of J2s activity was calculated using the formula48:
The concentrations (100, 75, 50, 25, and 10%) that showed the strongest nematicidal activity against J2s of M. incognita were selected for the greenhouse experiment, which was conducted three times by examining its effects on J2s mortality as described above.
Identification of bioactive compounds
High-performance liquid chromatography (HPLC)
To prepare the HPLC analysis standard, rutin powder was dissolved in methanol at a concentration of 1 mg/ml in a volumetric flask. The obtaining stock solution was then filtered using a 0.45 μm filter and sonicated for 2 min at 150 °C in ice before being injected into the HPLC system. Chromatography was conducted using an LC 20AD liquid chromatography system with a Shimadzu SPD-M20A PDA detector and C18G 120 (250 × 4.60) mm 5 μm column. A sample volume of twenty μl was injected, and the mobile phase consisted of a mixture of acetonitrile and 0.3% formic acid (15:85 v/v). The flow rate was set at 0.4 ml/min, and detection was performed at a wavelength of 350 nm49.
GC–MS analysis of endophytic fungal culture filtrate
The chemical constituents of the culture filtrate of A. flavus were identified through GC–MS analysis using an Agilent 7693 series GC system (United States) equipped with an OV-5 capillary column and an Agilent 5975C network selective mass detector50. The endophytic fungal culture filtrate was prepared by soaking the dry fungal culture filtrate in the eluent three consecutive times. It was subjected to GC–MS analysis using a Perkin Elmer instrument. The mass spectrometer was operated in the electron impact (El) mode at 70 eV in the scan range of 60–600 m/z.
Samples extraction for HPLC
To extract samples for HPLC analysis, 1 gm of fungal pellet was mixed with 10 mL of 80% methanol containing 1% acetic acid. The mixture was homogenized and placed in a tightly capped test tube, which was then shaken for 48 h at 200 rpm. After centrifugation and filtration through a 0.45 μm filter, the final filtrate was analyzed for phenolic compounds. This extraction process was performed in triplicate independent samples. Standard compounds such as Gallic acid, Protocatechuic acid, Catechin, Esculatin, Vanillic acid, Pyrocatechol, Coumarin, Cinnamic acid, 4,3-indol butyl acetic acid and Naphthyl acetic acid were used to detect the presence of phenolic and flavonoid compounds in the methanol extract of the A. flavus cultural filtrate pellet. The HPLC instrument and its conditions were previously described in studies51,52.
Efficacy of fungal culture filtrate against M. incognita on potato plants (greenhouse experiment)
A greenhouse experiment was conducted to determine the effect of A. flavus culture filtrate on controlling M. incognita infecting potato cv. Nicola. The potato seeds were prepared for planting by undergoing surface sterilization with 1.25% sodium hypochlorite for five minutes, followed by rinsing in sterile distilled water. The soil was autoclaved at 121 °C for two hours and aerated for a week before used to fill pots. The potato seeds sown in pots filled with a soil and sand mixture (1:1 v/v) and irrigated as needed. After one week, the top 3 cm of soil near the potato roots removed before inoculating the plants with two thousand freshly hatched juveniles of M. incognita in a juvenile suspension. Following transplantation and inoculation, the potato plants received treatments, including abamectin application and four concentrations of A. flavus culture filtrates. Growth parameters, such as root and shoot length, weight, and nematode parameters, including number of galls, egg masses per plant, and J2s per 250 cm3 of soil, assessed after 60 days of growth. Egg masses counted by examining three root samples under a stereoscope after shaking cut roots in NaClO solution. The experiment conducted with five replicates for each treatment and repeated twice at the City of Scientific Research and Technological Applications, Egypt (SRTA-City) greenhouse53,54.
Enzyme activity assays
Enzymatic activities of Chitinase, Peroxidase, and Catalase were measured 15, 30, and 60 days after nematode inoculation according to the method outlined by55. Initially, twenty-five grams of chitin milled and then suspended in 250 ml of 85% phosphoric acid (H3PO4). This mixture stored at 4 °C for 24 h before the blended in 2 L of distilled water using a warning blender. The resulting suspension was centrifuged, and this washing process repeated twice. The final wash resulted in a colloidal chitin suspension that adjusted to pH 7.0 with (1 N) NaOH. This suspension separated by centrifugation, and the pelleted colloidal chitin stored at 4 °C. The determination of Chitinase activity was conducted using the56 method. Initially, 1 ml of 1% colloidal chitin in 0.05 M citrate–phosphate buffer (pH 6.6) was placed in test tubes, followed by 1 ml of enzyme extract, mixed by shaking. The exact process repeated using crude extract instead. The tubes incubated in a water bath at 37 °C for 60 min, cooled, and centrifuged before assaying. The amount of reducing sugar in 1 ml of the supernatant was determined using dinitrosalicylic acid57, with optical density measured at 540 nm. Chitinase activity was expressed as mM N-acetylglucosamine equivalent released per gram fresh weight tissue per hour. Peroxidase activity was determined spectrophotometrically by the methods of58, while, Catalase was according to59.
Statistical analysis
The statistical analysis of the data involved the use of analysis of variance (ANOVA) and a revised LSD test to determine significant differences among the means at a p-value of ≤ 0.05. The SAS system60 was utilized for this purpose.
Ethics approval and consent to participate
This article does not contain any studies with human or animal subjects. The current experimental research and field study, including collecting plant material and compliance with relevant institutional, national, and international guidelines and legislation, is used for research and development.
Results
Identification of the isolated endophytic fungus
The isolation process led to the identification of endophytic fungal isolate from the genus Aspergillus, as confirmed by initial microscopic analysis (Fig. A). The isolated Aspergillus specimen was determined to be Aspergillus sp. The colonies of A. flavus exhibited rounded shapes, a velvety texture, a wet appearance, and a yellowish-green color with a light white border on PDA media (Fig. 1). In addition, the confirmation of the fungus isolate as Aspergillus flavus was determined through analysis of ITS sequence and culture morphology. Identification of the endophytic isolate fungal was identified by DNA sequences of the purified PCR product to confirm the genus of the microscopically identified A. flavus isolate, isolated from T. foenum-graecum seeds, the Aspergillus consensus ITS1-ITS4 PCR fragment was amplified (Aspergillus flavus ON146363). ITS sequencing region and identification of endophytic fungs were aligned, and the alignment sequence was subject to homology search by NCBI BLAST. Homology of the ITS region was used to identify the isolated endophyte as a species of Aspergillus. The current sequence showed 100% homology with sequences of the ITS regions of Aspergillus available in the Gene Bank database using the BLAST search program (https://blast.ncbi.nlm.nih.gov/Blast.cgi). To check the suitability of the isolated genomic DNA in downstream molecular biology applications, fungal strains were subjected to amplification using universal primers for the ITS region of 18 S rDNA61.

(A, B) Polymerase chain reaction (PCR) of Aspergillus flavus with ITS1-ITS4 (598 bp) and (C) Neighbor-joining tree of fungal endophyte A. flavus ON146363 based on ITS1-5.8S-ITS2 rDNA sequences. Confidence values above 50% obtained from a 500-replicate bootstrap analysis shown at the branch nodes. Bootstrap values from the neighbor-joining method were detected. A. flavus (LN482443) was used as the outgroup.
Figure 1A and B presented the gel electrophoresis and phylogenetic tree of the isolated endophytic fungi. The GenBank search for similar ITS region sequences revealed that strain ON146363 had higher sequence similarities with the species A. flavus than any other reference taxa. All ten sequences of reference taxa were acquired for constructing phylogeny. In the neighbor-joining (NJ) tree (Fig. 1C), strain ON146363 and the other reference taxa, A. flavus (MW288377), formed a clade with 99% bootstrap reinforcement. In this clade, the results of similarity comparative with the ITS1- 5.8S-ITS2 part sequence show that strain ON146363 had the maximum nucleotide similarities with Aspergillus flavus. This indicates that the morphological and molecular identification results could be assured that ON146363 is A. flavus.
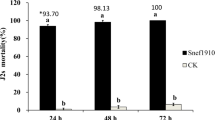
In vitro antagonistic activity against J2s mortality of M. incognita
The mortality rates of second-stage juvenile nematode odes were directly proportional to the concentration of culture filtrate, as shown in Fig. 2A. The appearance of J2s was not uniform when exposed to fungal culture filtrates, and all treatments showed a significant (P ≤ 0.05) increase in J2s mortality percentage after 12, 24, and 48 h compared to the control (DH2O). However, the response varied at different concentrations of the fungus culture filtrate treatments. The highest mortality rate for J2s was observed at the maximum concentration (100%), while the lowest concentration resulted in lower mortality rates. In most concentrations, the hatching of nematode eggs was significantly reduced compared to the control with DH2O. Culture filtrates from A. flavus caused 100% juvenile mortality after 48 h of exposure at a concentration of 100%. The differences in values between each concentration level were significant, with a stepwise increase in J2s mortality percentages with increasing concentrations and exposure time. The most aggressive concentration appeared to be A. flavus culture filtrate with a concentration of 100%, which caused 75%, 94%, and 97.7% juvenile mortality of J2s larvae in vitro after exposure times of 12, 24, and 48 h, respectively.

Effect of fungal culture filtrate on (A) larvae mortality and reduction percentages (R%) of the root-knot nematode and on (B) hatchability of M. incognita eggs (R%), under laboratory conditions after 12, 24, 48, 72 h and one week of exposure time. Bar charts are generated using data from five biological replications with two technical replicates per replication. Each bar represents the mean, and the error bar indicates the standard deviation (± SE). This figure was drawn with GraphPad Prism 8 (9.4.1, (458) Serial number: GPS-2567891- 8A130A8A228). The significant of the changes were assessed using SAS one-way ANOVA test. Means with different letters are significantly different (P ≦ 0.05).
In vitro effect on egg hatching of M. incognita
The results of the egg hatchability bioassay (Fig. 2B) demonstrate that the culture filtrates of A. flavus had a strong inhibitory effect on the hatching of M. incognita egg masses. The percentage decrease in hatchability ranged up to 95.1% and 97% after 72 h and one week of exposure, respectively. The various concentrations of the culture filtrate showed significant differences compared to the water control, with only the 10% dilution showing no significant difference. The data revealed that A. flavus culture filtrate was detrimental to egg hatching, with the lowest rate of hatchability observed at all concentrations tested. Moreover, the efficacy of the fungus culture filtrate in causing juvenile mortality was highest at 100%, followed by 75%, 50%, and 25% at the same time.
Efficacies of fungus culture filtrate against M. incognita on potato plants (greenhouse experiment)
The effectiveness of fungus culture filtrate in controlling M. incognita on potato plants was evaluated through a greenhouse experiment based on the Invitro nematicidal efficiency, and the disease control activity of fungal culture filtrate was assessed with potato root-knot nematode diseases in pot experiments. Also, the tested fungal culture filtrate 100% (s) concentration was potent and influential in suppressing the formation of the galls, the number of egg masses/ plant, and the number of J2s/250 cm3 of soil with potato roots by 90, 89.5 and 95.3%, respectively, compared with the Abamectin, (Fig. 3). The influence of fungal filtering and the commercial nematicide (Abamectin) on growth parameters of nematode-infected potatoes under greenhouse conditions were illustrated in (Table 1).

The impact of A. flavus culture filtrate on Meloidogyne incognita galls, egg masses, and J2s in 250 g soil of potato plants was examined under greenhouse conditions. Bar charts are generated using data from three biological replications with two technical replicates per replication. Each bar represents the mean, and the error bar indicates the standard deviation (± SE). This figure was drawn with GraphPad Prism 8. The significant of the changes were assessed using SAS one-way ANOVA test. Means with different letters are significantly different (P ≦ 0.05).
The findings in Fig. 3 indicated that the application of filtered S, S/2 (100, 75%), and abamectin concentrations increased the length and fresh weight of the plant, as well as the shoots and roots, compared to the infected control. However, there was no significant difference in shoot and root fresh weights and lengths between those treated with nematicide abamectin. The results demonstrated a noteworthy enhancement in potato growth parameters compared to the control treatment. Furthermore, potato plants treated with fungal culture filtrate at a concentration of 100% exhibited the highest percentage increase in shoot length at 41.7% Fig. 4. In comparison, those inoculated with M. incognita and treated with fungal filtering showed an increase of 31%.

Potato plants (A) control healthy without any treatment, (B) control M. incognita, (C) treated with fungal culture filtrate at a concentration of 100% and (D): A. flavus isolated potato.
Data in Table 1 illustrated a significant improvement (P ≤ 0.05) in potato plant growth parameters compared to the control treatment. After being inoculated with M. incognita, potato plants treated with A. flavus culture filtrate at concentrations of 50% and 100% displayed the highest percentage increase in shoot length at 20.62% and 37.85%, respectively. In comparison, Abamectin recorded an increase of only 30.9%. Additionally, potato plants treated with A. flavus culture filtrate at 100% and 50% concentrations showed the highest increase in root length percentages of 48.2% and 43.1%, respectively, followed by potato plants inoculated with M. incognita and treated with Abamectin at an increase of 43.6%. Furthermore, the highest increase in root fresh and dry weights was observed with treatments of fungal culture filtrate at a percentage increase of 15.81% and 31.41%, respectively, followed by potato plants inoculated with M. incognita alone and treated with Abamectin at an increasing value of 16.22%. In addition, the highest shoot dry weight increase percentages of 23.5 and 87.5% were recorded with potato plants inoculated with M. incognita and treated with Aspergillus culture filtrated at 50 and 100% concentration compare with Abamectin at 48.5% increase, respectively. Otherwise, the most significant increase percentages of 26.0% were recorded with treatments of potato plants inoculated with100% concentration from fungal filtration.
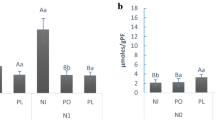
The activity of defence-related enzymes
This study aimed to determine the relative expression levels of three defence enzymes, Chitinase, Peroxidase, and Catalase, in potato plants that were inoculated with M. incognita and treated with varying concentrations of A. flavus culture filtrate. The findings presented in Fig. 5a–c indicated that the inoculation with M. incognita significantly improved the performance of Chitinase, Peroxidase, and Catalase enzymes compared to control plants. However, the two concentrations of A. flavus culture filtrate showed a more significant boost in defence enzyme activity than plants inoculated with M. incognita alone. The best treatment was observed when plants were treated with 50 mL of A. flavus one week before M. incognita inoculation as it significantly enhanced the activity of Chitinase, Peroxidase, and Catalase after 15, 45, and 60 days (9.89, 11, and 11.121), (3.89, 4.66 and 4.65), and (2.16, 2.1 and 2.81) respectively over plants inoculated with Meloidogyne only. Similarly, when A. flavus was mixed at a concentration of 50% with Chitinase, Peroxidase, and Catalase enzymes after meloidogyne inoculation; it resulted in increased enzyme activity after 15, 45, and 60 days (5.2, 8 0.23, and 7 0.23); (2 0.76, 2 0.79, and 1 0.05), and (1 0.08, 1 0.04, and 1 0.07) respectively over plants inoculated with Meloidogyne only.

Efficacy of A. flavus culture filtrate with concentrations (50 and 100%) on (A) Chitinas extracted, (B) Peroxidase enzyme and (C) Catalyse enzyme, from the fresh potato plant inoculated with M. incognita under green-house conditions. Bar charts are generated using data from five biological replications with two technical replicates per replication. Each bar represents mean and error bar indicates standard deviation (± SE). This figure was drawn with GraphPad Prism 8. The significant of the changes were assessed using SAS one-way ANOVA test. Means with different letters are significantly different (P ≦ 0.05).
GC–MS analysis
Capillary columns are utilized in GC–MS to separate complex mixtures containing low polarity or volatile substances. The increased fragmentation provided by GC/MS enables the identification of unknowns by searching a mass spectral database. GC–MS analysis was employed to uncover its compounds because the culture filters of the A. flavus fraction had low polarity. Using the NIST database with R match values exceeding 90% (Fig. 6), fifteen components were identified from the culture filter fraction of A. flavus. The area normalization method revealed that Gadoleic acid, Oleic acid di-ethanolamide, Oleic acid, and Palmitic acid with 310.5, 301, 282.5, and 256.4 g/mol, respectively.,accounted for 88% of the total content of the culture filter fraction from A. flavus.

GC–Ms analysis of A. flavus culture filtrated using an Agilent 7693 series GC system equipped with an OV-5 capillary column and an Agilent 5975C network selective mass detector.
HPLC analysis
The HPLC method aimed to initially verify the presence of common phenolic antioxidants. The chromatograms indicated that the fungal extract generated several peaks that corresponded to the retention time (RT) of genuine polyphenol antioxidants. Specifically, the chromatogram for the cultural filtrate fraction from A. flavus strain ON146363 exhibited ten peaks that matched the RTs of authentic gallic acid (2.999 min) with a quantity of 42.000 Ug/ml, catechin (3.590 min) with 126 Ug/ml, Protocatechuic acid (3.411) with 82 Ug/ml, Esculatin (4.652 min) with 93 Ug/ml, Vanillic acid (4.979 min) with 81Ug/ml, Pyrocatechol (5.336 min) with 52 Ug/ml, Coumarine (11.613 min) with 48 Ug/ml, Cinnamic acid (13.417 min) with 86 Ug/ml, 4,3-indol butyl acetic acid (14.750 min) with 84 Ug/ml and Naphthyl acetic acid (15.697 min)with 84 Ug/ml as illustrated in Fig. 7a,b.

(A) HPLC chromatogram of A. flavus cultural filtrate the mobile phase consisted of a mixture of acetonitrile and 0.3% formic acid (15:85 v/v). The flow rate was set at 0.4 ml/min, and detection was performed at a wavelength of 350 nm. (B) The concentrations of the detected compounds. This figure was drawn with GraphPad Prism 8.
Discussion
In this investigation, the culture filtrate of A. flavus revealed that nematicidal activity increased the mortality of J2s and decreased the egg hatchability of M. incognita. However, the specific percentage decrease observed in this study may be influenced by factors such as the concentration of the culture filtrate used and the specific nematode species tested. The finding that only the 10% dilution showed no significant difference compared to the water control suggests that lower concentrations of A. flavus culture filtrate may not have a significant impact on nematode egg hatching. This result is in agreement with what comes by62 who reported that the lower concentrations of fungal culture filtrates may have less pronounced effects on nematodes . The observation that juvenile mortality was highest at 100% concentration and decreased with decreasing concentrations is also consistent with previous research62 . This suggests that higher concentrations of A. flavus culture filtrate may be more effective at reducing nematode populations in lab experiment as reported by63.
The results of the tested fungal culture filtrate at 100% concentration in suppressing the formation of galls, egg masses, and J2s in potato plants infected with M. incognita are consistent with many studies that have reported the effectiveness of fungal metabolites in controlling plant parasitic nematodes. For instance, a study by64,65 reported that the culture filtrate of Trichoderma harzianum significantly reduced the galls and egg masses of M. incognita on tomato plants. Similarly, a study by66 found that the culture filtrate of A. niger significantly reduced the number of J2s and galls on tomato plants infected with M. incognita.
The effectiveness of fungal metabolites in controlling plant parasitic nematodes has been attributed to their ability to produce secondary metabolites such as enzymes, antibiotics, and volatile organic compounds that can directly or indirectly inhibit nematode growth and development67,68. This study's results suggest that applying filtered S, S/2 (100, 75%), and abamectin concentrations can improve the growth of potato plants infected with M. incognita. This is consistent with previous studies that have shown the effectiveness of these treatments in controlling nematode infestations and promoting plant growth69,70. The findings regarding using A. flavus culture filtrate are also interesting, as they suggest that this treatment can significantly increase shoot length in infected potato plants. This is consistent with previous studies that have shown the potential of fungal culture filtrates as biocontrol agents against plant pathogens71,72. Data from GC–MS makes it clear that A. flavus produces a variety of secondary metabolites with nematicidal activity that may detect it. High nematicidal activity against M. incognita is found within the four fatty acids identified: gadolic acid, oleic acid diethanolamide, oleic acid, and palmitic acid. The nematicidal activity of these fatty acids has been reported in many studies. A study conducted by73 found that gadoleic acid effectively reduced the population of Meloidogyne incognita, a common root-knot nematode, in tomato plants. Additionally, oleic acid di-ethanolamide, a derivative of oleic acid, has been shown to exhibit strong nematicidal activity against various nematode species. Whereas73 demonstrated its effectiveness against the soybean cyst nematode Heterodera glycines. Palmitic acid, a saturated fatty acid, was also found to have significant nematicidal effects on the root-knot nematode Meloidogyne javanica in a study by74, in this regard, various modes of action have been discussed, including overall disruption of the cell membrane permeability leading to electrolyte leakage, cell death and inhibition of protein synthesis and fatty acids metabolism75. Together, the combination of these substances gave rise to A. flavus more aggressive behavior towards nematodes. The HPLC analysis results indicated an increase in the concentration of important compounds in the A. flavus filtrate. This finding aligns with previous studies by76, who demonstrated the strong nematicidal activity of Gallic acid extracted from Rubus niveus against M. incognita, as well as the nematicidal activity of kojic acid isolated from Aspergillus oryzae77 and oxalic acid produced by Aspergillus niger F2278 against M. incognita. The culture filtrate of A. flavus JCK-4087 also exhibited potent nematicidal activity against M. incognita, consistent with findings by63. Compared to another A. flavus strain studied by63, JCK-4087 showed significantly stronger nematicidal activity with 100% J2s mortality at a concentration of 20%, whereas the other strain achieved only about 40% mortality at a concentration of 25%. The use of chemical nematicides has been effective in controlling RKNs, but prolonged use of carbamate and organophosphorus nematicides has led to nematode resistance and reduced field efficacy, prompting the development of novel chemical nematicides with different action mechanisms79.
The study observed an increase in the expression of defense-related genes POD, CHI, and CAT in potato plants colonized by A. flavus. CHI is responsible for breaking down chitin, a key component of fungal cell walls80. The overexpression of the antioxidant stress marker gene POD helps to reduce reactive oxygen species (ROS) in plant tissues under various stresses. This defense response was supported by the higher activities of POD and PPO enzymes. Up-regulation of these genes helped to reduce disease severity and incidence in potato plants colonized by A. flavus. Additionally, the accumulation of nematotoxic phenolic compounds in potato plants may hinder the growth and spread of M. incognita between cell81.
Overall, when a plant is colonized by endophytic fungi, it triggers the production of defensive secondary metabolites that help defend against pathogens. These mechanisms include cell wall lignification, production of nematicidal substances, stimulation of phytoalexins and PR-proteins, and increased expression of defense-related genes. The fungal endophyte also produces indole acetic acid (IAA) in significant amounts. Utilizing beneficial endophytic microorganisms as bio-fertilizers can enhance plant growth and production for sustainable agriculture and food safety81. These results are consistent with previous studies that have examined the impact of plant–microbe interactions on defense enzyme activity such as82. For instance34,83, research demonstrated that introducing the beneficial fungus Trichoderma harzianum increased Peroxidase and Catalase activity in tomato plants. At the same time84 studies found that introducing the beneficial bacterium B. subtilis increased Chitinase and Peroxidase activity in some plants. The enzyme Catalase breaks down hydrogen peroxide into water and oxygen, as noted by85. It has an incredibly high turnover rate, with one molecule of Catalase able to convert 40 million molecules of hydrogen peroxide into water and oxygen every second, according to86. Peroxidases are involved in several physiological processes, including auxin catabolism, defense against pathogens, respiratory processes, and the synthesis of lignin and other terpenoids that contribute to phytoalexin production87. The activity of oxidative enzymes has been tested with nematode infection and has proven useful as a biochemical marker in various plant species, such as tomato, as demonstrated by88.
Conclusion
In summary, the findings suggest that the A. flavus (ON146363) cultural filtrate may positively boost the activity of defensive enzymes and improve a plant's ability to resist plant pathogenic nematodes. Fungal filtrate is also rich in components that prevent or effectively control nematodes. This makes it a promising solution for controlling nematodes, particularly in severe cases. Additionally, it is eco-friendly and serves as a bio-agent for the ecosystem. In conclusion, the tested fungal culture filtrate suppresses M. incognita infestation in potato plants, consistent with many studies that have reported the potential of fungal metabolites as an alternative to synthetic nematicides.
Data availability
All data generated or analyzed during this study are included in this article.
References
Steglińska, A. et al. Metabolite formation by fungal pathogens of potatoes (Solanum tuberosum L.) in the presence of bioprotective agents. Int. J. Environ. Res. Public Health 20, 5221 (2023).
Kang, Y. et al. Exogenous Glutathione enhances tolerance of the potato (Solanum tuberosum L.) to cadmium stress by regulating the biosynthesis of phenylpropanoid and the signal transduction of plant hormones. Chem. Biol. Technol. Agric. 10, 24. https://doi.org/10.1186/s40538-023-00400-z (2023).
Hijmans, R. J. Global distribution of the potato crop. Am. J. Potato Res. 78, 403–412. https://doi.org/10.1007/BF02896371 (2001).
Orlando, V. et al. Root-lesion nematodes of potato: Current status of diagnostics, pathogenicity and management. Plant Pathol. 69, 405–417. https://doi.org/10.1111/ppa.13144 (2020).
Medina, I. L. et al. Genetic diversity of Meloidogyne spp. parasitising potato in Brazil and aggressiveness of M. javanica populations on susceptible cultivars. Nematology 19, 69–80 (2017).
Mohy, S., El Aasar, K. & Sakr, Y. Decomposition analysis of virtual water outflows for major Egyptian exporting crops to the European union. Sustainability 15, 4943 (2023).
Lima, F. S., Correa, V. R., Nogueira, S. R., & Santos, P. R. Nematodes affecting soybean and sustainable practices for their management. Soybean–basis of yield, biomass and productivity, 95–110 (2017).
Akrofi-Atitianti, F., Ifejika Speranza, C., Bockel, L. & Asare, R. Assessing climate smart agriculture and its determinants of practice in Ghana: A case of the cocoa production system. Land 7, 30 (2018).
Jones, J. T. et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 14, 946–961. https://doi.org/10.1111/mpp.12057 (2013).
Holgado, R. & Magnusson, C. Nematodes as a limiting factor in potato production in Scandinavia. Potato Res. 55, 269–278. https://doi.org/10.1007/s11540-012-9209-6 (2012).
Abd-Elgawad, M. M. M. Biological control agents in the integrated nematode management of potato in Egypt. Egypt. J. Biol. Pest Control 30, 121. https://doi.org/10.1186/s41938-020-00325-x (2020).
Karuri, H. W., Olago, D., Neilson, R., Mararo, E. & Villinger, J. A survey of root knot nematodes and resistance to Meloidogyne incognita in sweet potato varieties from Kenyan fields. Crop Protect. 92, 114–121. https://doi.org/10.1016/j.cropro.2016.10.020 (2017).
Fouda, M. M. G. et al. Utilization of High throughput microcrystalline cellulose decorated silver nanoparticles as an eco-nematicide on root-knot nematodes. Colloids Surf. B Biointerfaces 188, 110805. https://doi.org/10.1016/j.colsurfb.2020.110805 (2020).
Elkobrosy, D. H. et al. Quantitative detection of induced systemic resistance genes of potato roots upon ethylene treatment and cyst nematode, Globodera rostochiensis, infection during plant–nematode interactions. Saudi J. Biol. Sci. 29, 3617–3625. https://doi.org/10.1016/j.sjbs.2022.02.045 (2022).
Abdel-Salam, M. S., Ameen, H. H., Soliman, G. M., Elkelany, U. S. & Asar, A. M. Improving the nematicidal potential of Bacillus amyloliquefaciens and Lysinibacillus sphaericus against the root-knot nematode Meloidogyne incognita using protoplast fusion technique. Egypt. J. Biol. Pest Control 28, 31. https://doi.org/10.1186/s41938-018-0034-3 (2018).
Ravichandra, N. G. in Horticultural Nematology (ed N. G. Ravichandra) 207–238 (Springer India, 2014).
van Lenteren, J. C., Bolckmans, K., Köhl, J., Ravensberg, W. J. & Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 63, 39–59. https://doi.org/10.1007/s10526-017-9801-4 (2018).
Jamal, Q. M. S. & Ahmad, V. Lysinibacilli: A biological factories intended for bio-insecticidal, bio-control, and bioremediation activities. J. Fungi 8, 1288 (2022).
Fontana, D. C. et al. Endophytic fungi: Biological control and induced resistance to phytopathogens and abiotic stresses. Pathogens 10, 570 (2021).
Larran, S. et al. Endophytic Trichoderma strains increase soya bean growth and promote charcoal rot control. J. Saudi Soc. Agric. Sci. https://doi.org/10.1016/j.jssas.2023.03.005 (2023).
Noman, M. et al. Bio-functionalized manganese nanoparticles suppress fusarium wilt in watermelon (Citrullus lanatus L.) by infection disruption, host defense response potentiation, and soil microbial community modulation. Small 19, 2205687. https://doi.org/10.1002/smll.202205687 (2023).
Zeilinger, S. et al. Friends or foes? Emerging insights from fungal interactions with plants. FEMS Microbiol. Rev. 40, 182–207. https://doi.org/10.1093/femsre/fuv045 (2015).
Martinez-Medina, A. et al. Recognizing plant defense priming. Trends Plant Sci. 21, 818–822. https://doi.org/10.1016/j.tplants.2016.07.009 (2016).
Giauque, H. & Hawkes, C. V. Historical and current climate drive spatial and temporal patterns in fungal endophyte diversity. Fungal Ecol. 20, 108–114. https://doi.org/10.1016/j.funeco.2015.12.005 (2016).
Lord, J. C. Low humidity, moderate temperature, and desiccant dust favor efficacy of Beauveria bassiana (Hyphomycetes: Moniliales) for the lesser grain borer, Rhyzopertha dominica (Coleoptera: Bruchidae). Biol. Control 34, 180–186. https://doi.org/10.1016/j.biocontrol.2005.05.004 (2005).
Luz, C., Rodrigues, J. & Rocha, L. F. N. Diatomaceous earth and oil enhance effectiveness of Metarhizium anisopliae against Triatoma infestans. Acta Tropica 122, 29–35. https://doi.org/10.1016/j.actatropica.2011.11.014 (2012).
Gortari, M. C. & Hours, R. A. Fungal chitinases and their biological role in the antagonism onto nematode eggs A review. Mycol. Progress 7, 221–238. https://doi.org/10.1007/s11557-008-0571-3 (2008).
Li, Y. et al. Microbiota and functional analyses of nitrogen-fixing bacteria in root-knot nematode parasitism of plants. Microbiome 11, 48. https://doi.org/10.1186/s40168-023-01484-3 (2023).
Mahmud, K. et al. Rhizosphere microbiome manipulation for sustainable crop production. Curr. Plant Biol. 27, 100210. https://doi.org/10.1016/j.cpb.2021.100210 (2021).
Hodge, A., Berta, G., Doussan, C., Merchan, F. & Crespi, M. Plant root growth, architecture and function. Plant Soil 321, 153–187. https://doi.org/10.1007/s11104-009-9929-9 (2009).
Twomey, U., Warrior, P., Kerry, B. R. & Perry, R. N. Effects of the biological nematicide, DiTera, on hatching of Globodera rostochiensis and G. pallida. Nematology 2, 355–362. https://doi.org/10.1163/156854100509114 (2000).
Akhtar, M. & Malik, A. Roles of organic soil amendments and soil organisms in the biological control of plant-parasitic nematodes: a review. Bioresour. Technol. 74, 35–47. https://doi.org/10.1016/S0960-8524(99)00154-6 (2000).
Bhat, M. Y. & Wani, A. Bio-activity of fungal culture filtrates against root-knot nematode egg hatch and juvenile motility and their effects on growth of mung bean (Vigna radiata L. Wilczek) infected with the root-knot nematode, Meloidogyne incognita. Arch. Phytopathol. Plant Protect. 45, 1059–1069 (2012).
Akhter, G. & Khan, T. A. Evaluation of some plant extracts for nemato-toxic potential against juveniles of Meloidogyne incognita in vitro. J Phytopharmacol 7, 141–145 (2018).
Shinya, R. et al. Secretome analysis of the pine wood nematode bursaphelenchus xylophilus reveals the tangled roots of parasitism and its potential for molecular mimicry. PLOS ONE 8, e67377. https://doi.org/10.1371/journal.pone.0067377 (2013).
Guo, L. D., Hyde, K. D. & Liew, E. C. Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 147, 617–630. https://doi.org/10.1046/j.1469-8137.2000.00716.x (2000).
Patel, M. et al. Bacillus subtilis ER-08, a multifunctional plant growth-promoting rhizobacterium, promotes the growth of fenugreek (Trigonella foenum-graecum L.) plants under salt and drought stress. Front. Microbiol. 14, 1208743 (2023).
Khawar, K. et al. In vitro crown galls induced by Agrobacterium tumefaciens strain A281 (pTiBo542) in Trigonella foenum-graecum. Biol. Plant. 48, 441–444 (2004).
Gams, W., Bissett, J. (Taylor & Francis, London, 1998).
Mylroie, J. E. et al. Identification and quantification of a toxigenic and non-toxigenic Aspergillus flavus strain in contaminated maize using quantitative real-time PCR. Toxins 8, 15 (2016).
Kraus, G. F. et al. Trichoderma brevicompactum sp. nov. Mycologia 96, 1059–1073 (2004).
Yang, L. et al. Species identification through mitochondrial rRNA genetic analysis. Sci. Rep. 4, 4089 (2014).
Mira, A., Pushker, R., Legault, B. A., Moreira, D. & Rodríguez-Valera, F. Evolutionary relationships of Fusobacterium nucleatum based on phylogenetic analysis and comparative genomics. BMC Evol. Biol. 4, 1–17 (2004).
Williams, S. T. et al. Numerical classification of streptomyces and related genera. Microbiology 129, 1743–1813. https://doi.org/10.1099/00221287-129-6-1743 (1983).
Tamura, K., Dudley, J., Nei, M. & Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) Software Version 40. Mol. Biol. Evol. 24, 1596–1599. https://doi.org/10.1093/molbev/msm092 (2007).
Hussey, R., & Barker, K. Comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Disease Reporter (1973).
Ghareeb, R. Y. et al. Nematicidal activity of seaweed-synthesized silver nanoparticles and extracts against Meloidogyne incognita on tomato plants. Sci. Rep. 12, 3841. https://doi.org/10.1038/s41598-022-06600-1 (2022).
Nguyen, D.-M.-C. et al. Nematicidal activity of 3,4-dihydroxybenzoic acid purified from Terminalia nigrovenulosa bark against Meloidogyne incognita. Microb. Pathogene. 59–60, 52–59. https://doi.org/10.1016/j.micpath.2013.04.005 (2013).
Dubber, M.-J. & Kanfer, I. High-performance liquid chromatographic determination of selected flavonols in Ginkgo biloba solid oral dosage forms. J. Pharm. Pharm. Sci. 7, 303–309 (2004).
Abdullah, S. M., Kolo, K. & Sajadi, S. M. Greener pathway toward the synthesis of lichen-based ZnO@ TiO2@ SiO2 and Fe3O4@ SiO2 nanocomposites and investigation of their biological activities. Food Sci. Nutrit. 8, 4044–4054 (2020).
Sarker, U. & Oba, S. Phenolic profiles and antioxidant activities in selected drought-tolerant leafy vegetable amaranth. Sci. Rep. 10, 18287. https://doi.org/10.1038/s41598-020-71727-y (2020).
Pappritz, K. et al. Speckle-tracking echocardiography combined with imaging mass spectrometry assesses region-dependent alterations. Sci. Rep. 10, 3629. https://doi.org/10.1038/s41598-020-60594-2 (2020).
Ramadhan, M., Amin, N. & Nasruddin, A. in IOP Conference Series: Earth and Environmental Science. 022083 (IOP Publishing).
Regaieg, H., Ciancio, A., Raouani, N. H., Grasso, G. & Rosso, L. Effects of culture filtrates from the nematophagous fungus Verticillium leptobactrum on viability of the root-knot nematode Meloidogyne incognita. World J. Microbiol. Biotechnol. 26, 2285–2289. https://doi.org/10.1007/s11274-010-0397-4 (2010).
Reid, J. D. & Ogrydziak, D. M. Chitinase-overproducing mutant of Serratia marcescens. Appl. Environ. Microbiol. 41, 664–669 (1981).
Boller, T. & Mauch, F. in Methods in Enzymology Vol. 161 430–435 (Academic Press, 1988).
Monreal, J. & Reese, E. T. The chitinase of Serratia marcescens. Can. J. Microbiol. 15, 689–696. https://doi.org/10.1139/m69-122 (1969).
Rowan-Nash, A. D., Korry, B. J., Mylonakis, E. & Belenky, P. Cross-Domain and viral interactions in the microbiome. Microbiol. Mol. Biol. Rev. 83, e00044-e18. https://doi.org/10.1128/MMBR.00044-18 (2019).
Chance, B. & Maehly, A. C. in Methods in Enzymology Vol. 2 764–775 (Academic Press, 1955).
Institute, S. SAS Procedures Guide. (Sas Inst, 1988).
van Burik, J.-A.H., Schreckhise, R. W., White, T. C., Bowden, R. A. & Myerson, D. Comparison of six extraction techniques for isolation of DNA from filamentous fungi. Med. Mycol. 36, 299–303. https://doi.org/10.1080/02681219880000471 (1998).
Mukhtar, T. & Pervaz, I. In vitro evaluation of ovicidal and larvicidal effects of culture filtrate of Verticillium chlamydosporium against Meloidogyne javanica. Int. J. Agric. Biol. 5, 576–579 (2003).
Naz, I. et al. Biological control of root knot nematode, Meloidogyne incognita, in vitro, greenhouse and field in cucumber. Biol. Control 152, 104429 (2021).
Sayed, M., Abdel-rahman, T., Ragab, A. & Abdellatif, A. Biocontrol of root-knot nematode Meloidogyne incognita by Chitinolytic Trichoderma spp. Egypt. J. Agronematol. 18, 30–47 (2019).
Ghareeb, R. Y. et al. Oscillatoria sp. as a potent anti-phytopathogenic agent and plant immune stimulator against root-knot nematode of soybean cv. Giza 111. Front Plant Sci 13, 870518. https://doi.org/10.3389/fpls.2022.870518 (2022).
Abd-Elgawad, M. M. M. & Askary, T. H. Fungal and bacterial nematicides in integrated nematode management strategies. Egypt. J. Biol. Pest Control 28, 74. https://doi.org/10.1186/s41938-018-0080-x (2018).
Pires, D. et al. The fight against plant-parasitic nematodes: current status of bacterial and fungal biocontrol agents. Pathogens 11, 1178 (2022).
Patil, S. & Sriram, S. Biological control of Fusarium wilt in crop plants using non-pathogenic isolates of Fusarium species. Indian Phytopathol. 73, 11–19. https://doi.org/10.1007/s42360-020-00202-5 (2020).
Kaur, T., Jasrotia, S., Ohri, P. & Manhas, R. K. Evaluation of in vitro and in vivo nematicidal potential of a multifunctional streptomycete, Streptomyces hydrogenans strain DH16 against Meloidogyne incognita. Microbiol. Res. 192, 247–252. https://doi.org/10.1016/j.micres.2016.07.009 (2016).
Yadav, A. N. et al. Plant growth promoting bacteria: biodiversity and multifunctional attributes for sustainable agriculture. Adv. Biotechnol. Microbiol. 5, 1–16 (2017).
Tonelli, M., Taurian, T., Ibáñez, F., Angelini, J. & Fabra, A. Selection and in vitro characterization of biocontrol agents with potential to protect peanut plants against fungal pathogens. J. Plant Pathol., 73–82 (2010).
Binod, P., Sukumaran, R. K., Shirke, S., Rajput, J. & Pandey, A. Evaluation of fungal culture filtrate containing chitinase as a biocontrol agent against Helicoverpa armigera. J. Appl. Microbiol. 103, 1845–1852 (2007).
Chen, J.-X. & Song, B.-A. Natural nematicidal active compounds: Recent research progress and outlook. J. Integr. Agric. 20, 2015–2031. https://doi.org/10.1016/S2095-3119(21)63617-1 (2021).
Lu, Q. et al. Nematicidal effect of methyl palmitate and methyl stearate against Meloidogyne incognita in bananas. J. Agric. Food Chem. 68, 6502–6510 (2020).
Baloni, P. et al. Metabolic network analysis reveals altered bile acid synthesis and metabolism in Alzheimer’s disease. Cell Rep. Med. 1 (2020).
Sultana, N., Akhter, M. & Khatoon, Z. Nematicidal natural products from the aerial parts of Rubus niveus. Nat. Prod. Res. 24, 407–415 (2010).
Nguyen, V. T. et al. Nematicidal activity of cyclopiazonic acid derived from Penicillium commune against root-knot nematodes and optimization of the culture fermentation process. Front. Microbiol. 12, 726504 (2021).
Kim, T. Y. et al. Nematicidal activity of grammicin produced by Xylaria grammica KCTC 13121BP against Meloidogyne incognita. Pest Manag. Sci. 74, 384–391 (2018).
Ebone, L. A., Caverzan, A. & Chavarria, G. Physiologic alterations in orthodox seeds due to deterioration processes. Plant Physiol. Biochem. 145, 34–42 (2019).
Zarei, M. et al. Characterization of a chitinase with antifungal activity from a native Serratia marcescens B4A. Brazil. J. Microbiol. 42, 1017–1029 (2011).
Zandi, P. & Schnug, E. Reactive oxygen species, antioxidant responses and implications from a microbial modulation perspective. Biology 11, 155 (2022).
Mishra, S. et al. Harnessing plant-microbe interactions for enhanced protection against phytopathogens. Plant Microbes Symbiosis: Applied Facets, 111–125 (2015).
Awasthi, A. K., Pandey, A. K. & Khan, J. Potential of fungus Trichoderma harzianum for toxicity reduction in municipal solid waste leachate. Int. J. Environ. Sci. Technol. 14, 2015–2022. https://doi.org/10.1007/s13762-017-1271-9 (2017).
Chen, F. et al. Quantitative changes of plant defense enzymes and phytohormone in biocontrol of cucumber Fusarium wilt by Bacillus subtilis B579. World J. Microbiol. Biotechnol. 26, 675–684. https://doi.org/10.1007/s11274-009-0222-0 (2010).
Chelikani, P., Fita, I. & Loewen, P. C. Diversity of structures and properties among catalases. Cell. Mol. Life Sci. CMLS 61, 192–208. https://doi.org/10.1007/s00018-003-3206-5 (2004).
Schoonen, M. A. A., Harrington, A. D., Laffers, R. & Strongin, D. R. Role of hydrogen peroxide and hydroxyl radical in pyrite oxidation by molecular oxygen. Geochimica et Cosmochimica Acta 74, 4971–4987. https://doi.org/10.1016/j.gca.2010.05.028 (2010).
Bashan, Y. & Levanony, H. Horizontal and vertical movement of Azospirillum brasilense Cd in the soil and along the rhizosphere of wheat and weeds in controlled and field environments. Microbiology 133, 3473–3480. https://doi.org/10.1099/00221287-133-12-3473 (1987).
Molinari, A. et al. Synthesis and cytotoxic evaluation of 6-(3-Pyrazolylpropyl) derivatives of 1,4-Naphthohydroquinone-1,4-diacetate. Archiv der Pharmazie 342, 591–599. https://doi.org/10.1002/ardp.200900041 (2009).
Acknowledgements
The authors would like to acknowledge their universities for supporting the research.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Contributions
R.Y.G.: Formal analysis, investigation, methodology, resources, writing – original draft, writing – review & editing. M. J.: writing – review & editing. N.A.: writing – original draft, writing – review & editing. M.A.: data curation, formal analysis, resources, writing – original draft. E.E.-A.: conceptualization, formal analysis, resources, software, writing – original draft. M.G.: formal analysis, investigation, resources, software, writing – original draft. All authors reviewed the manuscript."
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ghareeb, R.Y., Jaremko, M., Abdelsalam, N.R. et al. Biocontrol potential of endophytic fungi against phytopathogenic nematodes on potato (Solanum tuberosum L.). Sci Rep 14, 15547 (2024). https://doi.org/10.1038/s41598-024-64056-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-64056-x
- Springer Nature Limited