
Abstract
We present direct evidence of early grape domestication in southern Italy via a multidisciplinary study of pip assemblage from one site, shedding new light on the spread of viticulture in the western Mediterranean during the Bronze Age. This consist of 55 waterlogged pips from Grotta di Pertosa, a Middle Bronze Age settlement in the south of the Italian peninsula. Direct radiocarbon dating of pips was carried out, confirming the chronological consistency of the samples with their archaeological contexts (ca. 1450–1200 BCE). The extraordinary state of conservation of the sample allowed to perform geometric morphometric (GMM) and paleogenetic analyses (aDNA) at the same time. The combination of the two methods has irrefutably shown the presence of domestic grapevines, together with wild ones, in Southern Italy during the Middle/Late Bronze Age. The results converge towards an oriental origin of the domestic grapes, most likely arriving from the Aegean area through the Mycenaeans. A parent/offspring kinship was also recognised between a domestic/wild hybrid individual and a domestic clonal group. This data point out a little known aspect of the diffusion of the first viticulture in Italy, and therefore in the western Mediterranean, which involved the hybridization between imported domestic varieties with, likely local, wild vines.
Similar content being viewed by others


Introduction
In order to reconstruct the history of grape vines (Vitis vinifera L.) domestication, several studies have consistently shown that this process originated in Western Asia and Caucasus1,2,3,4,5,6,7,8,9,10,11, and it resulted into the differentiation of phenotypically distinct domestic (Vitis vinifera subsp. vinifera) and wild (Vitis vinifera subsp. sylvestris) subspecies. A recent study clarified that two separate domestication events took place concurrently around the advent of agriculture (~ 10.500 to 12.500 BP) in the two areas12. There is uncertainty however regarding what happened outside the primary area of domestication, especially concerning the role played by local wild populations present in those areas where viticulture developed over time. Numerous genetic studies have highlighted a complex relationship between the two subspecies, variously interpreted as the result of introgressive hybridisation following the introduction of domestic varieties from the eastern Mediterranean or secondary domestication of local wild vines4,6,10,11,13,14. In addition, the timing of grapevine domestication and of its early spread in the Mediterranean is poorly known. Observing the current geographical distribution of wild vines15,16,17, it is worth exploring the relationship between the two subspecies in the areas where viticulture spread.
Archaeobotany can potentially play a fundamental role in understanding the phenomenon as it allows us to have a direct look at the plant remains within past anthropic contexts that testify the process. Nevertheless, the spread and domestication of grapevine are not easy to read, mainly because recognition of the characteristics that enable a Vitis vinifera seed to be identified as belonging to the domestic or wild subspecies is not immediate. Over the years, various methods for achieving this aim have been developed, initially based on morphological observations and on the measurement and calculation of biometric indices on grape seeds18,19,20. These methods have however been found to suffer from various limitations. Their reliability depends on the method used, on the size and diversity of the reference collections and on the state of conservation of the archaeological remains21 and they can give contrasted results22. More elaborate methods were subsequently developed, based on the analysis of 2D images, on the statistical analysis of various morphometric features and comparison with modern reference collections23,24,25. A promising method for future research has been recently proposed for the identification of grape pips in terms of their variety, which envisages a complete 3D scanning of the geometry of the seeds and then analysis based on machine learning methods26. Geometric Morphometric (GMM) analysis of ventral and lateral outlines of the pips using the Elliptic Fourier Transform method allow the identification of modern cultivars and a powerful discrimination of wild and domesticated grape pips24,27,28,29. Paleogenetic analysis has also proved to be a powerful tool, even if strictly dependent on the taphonomic processes and the state of conservation. Up to now the best results, for both paleogenetics and morphometrics, come from the extraction and analysis from waterlogged material. This type of remains is not very common but some studies have been done, mainly regarding advanced steps in the history of viticulture in Mediterranean France30,31 but also other areas29,32, including Italy33.
For its geographic position, the Italian peninsula—especially its southern part—presumably played a major role in the spread of viticulture in the Mediterranean, and from here to western and central Europe.
Previous studies of the assemblages of grape pips discovered in Italy by means of various biometric and morphometric methods do not always have the same degree of reliability and can yield results that are not always comparable with each other. However the general opinion is that during the Middle/Late Bronze Age the first evidence of seeds with domestic or hybrid characteristics began to become evident, mainly in northern Italy and Sardinia34. The use of non-comparable methods is not the only problem with the research on the diffusion of viticulture in this important geographical region. The lack of large grape pip assemblages from Southern Italy constitutes the main limit on our ability to read the phenomenon in detail and assess its implications for the morphology of grape pips (SI. Table S1). This paper is based on the study of one of the largest pips assemblages from protohistoric southern Italy, which chronology was ascertained by direct radiocarbon dating, and the only one made up of waterlogged specimens whose preservation state is suitable to simultaneously perform geometric morphometric and paleogenetic analysis: Grotte di Pertosa e Auletta hereinafter, for the sake of brevity, Grotta di Pertosa (Fig. 1).

Bronze age sites with Vitis vinifera macro remains in southern Italy.
The site of Grotta di Pertosa and the context of discovery
The original layout of the settlement of Grotta di Pertosa seems to be dated to the early Middle Bronze Age. Its moment of greatest extension, which has been better investigated, was however in the immediately subsequent phase, which coincides with the Apennine facies and with the beginning of the Sub-Apennine facies in southern Italy, corresponding to the transition from the Middle to the Late Bronze Age35,36 (SI. Fig. S1). The site of Grotta di Pertosa lies in the hinterland of the Campania region in a strategic position dominating the northern entrance to the Vallo di Diano, a broad valley containing an ancient lake, now dried up, which forms a natural junction of several routes crossing the southern Apennines running both N-S and E-W. The decision to settle in the initial stretch of a large karst cave crossed by an underground river37 determined the type of construction. This consisted of pile-dwellings built mainly with the wood of deciduous oaks, partly constructed on the river’s muddy shore and partly directly in the water35,38 and it is the only known case of a pile-dwelling built inside a cave in the Mediterranean area. At least two overlapping levels of pile-dwellings have been recognized, but there were certainly a number of building phases39,40,41, considering that the settlement lasted until the early Iron Age42,43.
The silty-clay sediments in the cavity, saturated with water, have conserved the organic materials, and thus the plant remains, by waterlogging. However, for logistical reasons the site has not undergone systematic archaeological excavation in recent times. Despite this, during a survey in 2013, a well-conserved layer rich in plant remains was identified and interpreted as the floor of a collapsed pile-dwelling, from which a small sample (2 L of sediment) was taken41. The archaeobotanical analysis35,38 highlighted unequivocal traces of the processing of cereals and a clear preponderance of perennial plant fruits, which included the grape pips analysed in this study. In the sampled sediment 72 grape pips were found, of which 55 intact and 17 fragmentary, which is a quite remarkable density of remains. This stratigraphic unit underwent two radiocarbon datings, one of which was performed directly on the grape pips, yielding a chronological range of 1445 to 1192 (cal 2σ) BCE (SI. Table S2). All the whole pips were subjected to GMM analysis and 10 of them were selected for palaeogenetics.
Results
Geometric morphometrics (GMM)
The linear discriminant analysis (LDA) performed on a balanced collection of modern wild and domestic grape pips allowed to classify a majority of the archaeological pips in the wild morphotype. The domestic morphotype is nevertheless clearly identified. With a probability threshold of p ≥ 0.75, 38 grape pips were assigned to the wild group (69.1%), 9 (16.4%) to the domestic morphotype and 8 (14.5%) remained unclassified (Fig. 2). It must be stressed that, although it is in the minority, the proportion of pips assigned to the domestic type is always significantly greater than the error of the model, regardless of the probability threshold chosen (SI. Table S3). GMM thus provide a robust identification of the domestic grapevine in Middle-Late Bronze Age Pertosa.

GMM—distribution of archaeological pips according to the allocation to the wild or domesticated compartment.
Despite their small number, the 9 pips identified as domesticated by this first LDA with p-values ≥ 0.75 were submitted to a second LDA performed at cultivar level (SI. Table S4). They are allocated to 8 different modern cultivars but the p-value is over 0.75 in only 4 cases. The highlighted cultivars are mostly of eastern origins (Balkans and Caucasus/Near east areas). Only «Bourboulenc» is a traditional French white wine variety. It should be noted that a minor proportion of archaeological pips has previously been assigned to this cultivar morphotype in studies performed in Georgia29 and in the Aegean area44. Although these results are poorly representative due to the limited number of pips of the domesticated morphotype, we decided to include them in our study anyway as we have the possibility to compare them to aDNA results.
Ancient DNA
Shotgun sequencing demonstrated that nine of the ten tested seeds had at least 0.2% endogenous DNA (range = 0.00–4.48%; mean = 2.36%) and the expected patterns in cytosine deamination and DNA fragmentation (SI. Fig. S2). Damage-repaired libraries were prepared for the promising samples and enriched for SNPs present in the GrapeReSeq panel, reaching a mean depth of coverage on the SNP loci of 1.1–22.0. An analysis of the relative frequency of alleles at the SNP loci demonstrated these specimens contained < 20% paternal DNA (SI. Fig. S3), consistent with DNA preservation observed in other archaeological grape seeds30. Five samples yielded sufficient on-target data to call genotypes at > 1600 SNP loci, following a conservative approach that ignores sites with insufficient data to differentiate between true heterozygous sites and homozygous sites with limited contributions from paternal DNA31 (see SI).
A principal component analysis (PCA) of modern grapevine diversity45 projects two archaeological samples among wild V. vinifera sylvestris (pips P51 and P55), one near the boundary of wild and domesticated accessions (P52), and six within the PCA-space of domesticated grapevines (Fig. 3A). PCA was repeated using the cultivated panel and the pips affiliated with domesticated grapevines (Fig. 3B), revealing two clusters of the Grotta di Pertosa samples with none directly associated with the four groups defined by Laucou et al.46, but all near the cluster of modern grapes from the Balkans used for winemaking.

Genetic affinities between pips excavated at Grotta di Pertosa and modern V. vinifera accessions. (A) PCA with archaeological samples projected on the diversity of wild and cultivated grapevines, with domesticated grapes colored according to the four main ancestry clusters identified by Laucou et al.6. (B) PCA of putatively domesticated archaeological samples projected on the diversity of domesticated grapevines in the GrapeReSeq panel.
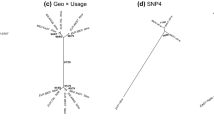
Using the called genotypes, KING found no kinship relationships between the archaeological samples and modern accessions, however it detected kinship within Grotta di Pertosa specimens: one parent–offspring relationship between P50 and P52 and a clonal relationship between P54 and P57. Kinship among all archaeological samples was further explored in a genotype likelihood framework, with ngsRelate identifying the three clusters observed in PCA to be genetic clones: P50 identical to P53, P56 and P59, P51 identical to P55, and P54 identical to P57 (Fig. 4). The software also identified all four members of the first clonal group to be in a parent–offspring relationship with P52.

Relatedness among the Grotta di Pertosa samples. Relatedness metrics were calculated using ngsRelate in a genotype likelihood framework based on allele frequencies of the GrapeReSeq diversity panel. For legibility, pairwise comparisons are shown in separate panels for each sample. The software calculates two metrics that distinguish close relationships in grapevine: the proportion of sites where samples share zero alleles identical by state (IBS0) and the KING kinship coefficient. Clonal (monozygotic twins) relationships fall within the red square(K ≥ 0.49 and IBS0 ≤ 0.001), parent–offspring relationships fall within the blue area (0.177 < K < 0.354 and IBS0 ≤ 0.001), siblings/highly related samples fall in the yellow area (0.177 < K < 0.354 and IBS0 ≤ 0.25).
Genetic distances between modern samples and the four Grotta di Pertosa genotypes were consistent with the PCA plot. The first clonal group (P50, P53, P56 and P59) has the closest genetic affinity with domesticated vines, with the closest being ‘Landroter’, a variety identified as admixed by Laucou et al.46. The most closely related non-admixed variety is ‘Gouais blanc’, which is a member of the Balkan wine group. The second clonal group (P51 and P55) has close relationships with many sylvestris accessions, with the highest in vines from Slovakia and Germany. The third clonal group is mostly closely related to admixed grapes, with the closest being Greek variety ‘Muscat à petits grains blancs’. P52, the only specimen not a part of a clonal group, has genetic affinity to many wild and domesticated grapes, with the closest being ‘Riesling bleu’, but with nearly the same genetic distance to a wild vine from Algeria. While these genetic links show affinity toward the local region, the specimens have even closer genetic links to other archaeological samples. In particular, the first clonal group and P52 show close relationships to Roman seeds from Terrasses de Montfau from southern France and the third clonal group is related to early Roman seeds from Mas de Vignoles XIV near Nîmes31 (SI Table S5).
Following the approach by Myles et al.47, the deeper history of the cultivated vines was explored in PCA by projecting cultivars and archaeological samples onto sylvestris diversity (Fig. 5). Consistent with previous observations, most modern cultivars fall in a space separating wild grapevines from Turkey and Afghanistan. Although the reference database has few sylvestris accessions from the eastern part of grapevine’s native range—notably lacking samples from Armenia, Azerbaijan, and Pakistan—the analysis recapitulates Myles et al.’s47 findings, wherein it was argued that the cultivated genepool largely descends from wild grapevines in West Asia. Grotta di Pertosa Clonal groups 1 and 3 are projected into the same space as modern cultivars, indicating they have little or no gene flow from sylvestris populations in Greece, Italy, or other parts of Western Europe. In contrast, Clonal Group 2 falls among sylvestris accessions from Slovakia, Germany, and France, potentially consistent with a local Italian origin. The admixed P52 specimen falls midway between its presumptive domesticated parent and the wild grapevines of Western Europe. P52’s position is shared by some cultivated vines used for winemaking in Western and Central Europe, and these varieties have been shown to be the product of crossing with local sylvestris, however none appear to result from an independent domestication.

PCA of domesticated and archaeological samples projected onto the diversity of sylvestris accessions.
Discussion
Early viticulture in Bronze Age Italy
The harvesting of what were in all probability wild grapes is sporadically attested in Southern Italy since the Upper Palaeolithic48 and the Mesolithic49 within two sites. However, it is during the Neolithic that seeds of Vitis vinifera, plausibly still wild, become more common in the archaeobotanical assemblages of Italian sites34,50,51,52,53,54. This is more clear in the northern regions, but is also recorded in the south, where 5 sites are known. Domesticated grapevine was identified by morphometric and paleogenetic analyses of desiccated, uncharred pips from a Late Neolithic layer in Grotta della Serratura55, in the Campania region. However, no direct radiometric dating was performed and no explanation is provided to explain the preservation of uncharred pips in a dry site. This result should therefore be considered with caution until it is supported by direct dating and/or by matching results from other sites.
During the Early Bronze Age several pile-dwelling settlements and other wetland sites in northern Italy have yielded rich assemblages of grape pips, all regarded as wild56,57,58,59. It is in the Middle Bronze Age that a clear increase in attestations is seen throughout the peninsular territory60,61,62, and considered as possibly linked to wine production60,63,64. Basing mainly on biometric indices18,19,20, but also on simple morphological observations, some authors suggest the appearance at this time of the first domesticated types60,65 which would be more frequently attested during the Late and Final Bronze Age60,66,67,68,69,70,71,72,73,74.
This alleged general trend is primarily based on the evidence from the northern and central Tyrrhenian regions of the peninsula, while in the southern and Adriatic regions the evidence is much less abundant34 and less extensively studied. Nevertheless, in this phase, grape pips have been discovered in 21 southern sites (Fig. 1, SI Table S1). Unfortunately, the high number of sites does not correspond to a large quantity of pips discovered, and those that have been recovered are often fragmentary.
The discrepancy between the quantity of the evidence of vines between the northern/central Tyrrhenian area and the southern/central Adriatic area of the Italian peninsula is partly due to the fact that the former hosts numerous sites located in wetland environments (i.e. pile-dwelling settlements near the Alps and on the volcanic lakes of central Italy, as well as the Terramare culture in the Po plain), which, given the characteristics of their sediments, conserve organic remains even in the absence of combustion. In contrast, in the south and the islands, mainly charred grape pips are discovered, their preservation probably being the result of fortuitous and unusual conditions. However, an increase in grapevine interest has also been recorded by pollen analyses conducted on cores taken from sites in both Apulia, in the Alimini Piccolo lake75 and the Salpi lagoon near Coppa Nevigata site76, and in Campania, in the Gulf of Gaeta77. In addition, three discoveries dated to the Middle Bronze Age could constitute indirect evidence of viticulture. These are the 21 vine branches recovered from a well in the Strepparo e Cento Moggie district in Capua78, together with various anthracological remains of Vitis vinifera discovered in workshops in Coppa Nevigata79 and Tufariello80. The cited cases have been interpreted as evidence of pruning, indicating the existence of practices associated with the care and management of grape vines from as early as the Middle Bronze Age.
Diffusion of grape varieties and allochthonous input
Considering the historical period we can observe a chronological coincidence between what we believe to be the first evidences of viticulture in Southern Italy with the arrival of Mycenaean people on its coasts. Given that a gene flow between the vines of Greece and southern Italy took place at some point in history81,82, it may be wondered whether viticulture in Italy was in some way triggered or accelerated by allochthonous inputs from the eastern Mediterranean, at its very dawn. The answer is not obvious. In order to pursue this aim it is worthwhile to establish certain key points regarding Mycenaean visits to the Italian coasts during the Bronze Age (SI Fig. S1), which is a fairly complex theme83. The relationships between the Mycenaeans and the local populations differed from region to region and changed over time, ranging from occasional and sporadic visits to consolidated trade relations and more complex forms of cultural integration, with the imitation of products and models84,85,86,87. The traffic between Southern Italy and the Aegean area may have included food products that leave highly labile traces in the archaeological record which is significant and opens up new spaces for reflection. This may have included both livestock88 and agricultural products. Remains of Triticum timopheevii found in Grotta di Pertosa could constitute a clue in this sense89. Indeed this is the only case for this species to be identified in southern Italy, whereas, as far as we know, its distribution range goes from Anatolia to the Balkan peninsula up to the Alpine area and Central and Eastern Europe90,91,92,93,94,95.
Chronologically, the Mycenaean contacts start to become evident in about 1700 BCE. Significant evidence of strict contacts has been found during the subsequent phase, i.e. LH III A, corresponding to the Apennine facies in southern Italy (about 1450–1300 BCE). After this period the Mycenaean palatial system collapsed, although relations between the eastern Mediterranean and Southern Italy continued, as attested by the abundant discoveries of LH III B-C materials in Sub-Apennine contexts (Late Bronze Age). The period saw the rise of new cultural manifestations such as the imitative ceramics known as Italo-Mycenaean products83,84,85,96,97.
Grotta di Pertosa is located in an area not directly exposed to Mycenaean traffic, but several archaeological indicators (i.e. faience beads, bronze artifacts and weapons, imported lithic raw materials, timopheevii wheat) show a certain degree of integration in a network of contacts on the regional and transregional scale41,89. Furthermore sporadic Aegean ceramic fragments have been discovered in underground funerary contexts very close to Grotta di Pertosa98,99.
Thanks to new data on the remains of grape pips from Greek contexts described in a recent publication44, it is possible to compare our results with those more or less coeval sites, analysed with the same geometric morphometric method (Fig. 6). Of the four Bronze Age sites considered in that study, the one with a wild/domestic morphotypes ratio comparable to Pertosa is Agia Paraskevi, which is few centuries older. The other three sites, contemporary to Pertosa (i.e. Dikili Tash, Mitrou and Kastanas) show an already well established viticulture with domestic morphotypes representing the 55 to 95% of the total grape pips analyzed. Unfortunately the pips from Greek sites are charred so it is not possible to verify the existence of any genetic correlation with our sample. A GMM comparison of the domestic grape pips of Kastanas and Mitrou with modern varieties shows that some of them are very similar to modern cultivars grown in Turkey, the Levant and the Caucasus. The authors did not exclude the introduction of domestic varieties from the East and their potential hybridisation with local varieties44.

Distribution of archaeological pips according to the allocation to the wild or domesticated compartment (p 0.75) and to their geographic location (Greek sites data after Pagnoux et al. 202144).
Similar considerations can be made on the basis of our results from GMM and aDNA on the Pertosa samples, in which a relationship seems to emerge between the introduction of already domesticated varieties (likely from Balkans and Greece) and their hybridisation with local wild grapes. This previously unknown data for the western Mediterranean, supported both by GMM and ancient DNA results, seems to be correlated to the first contacts with Mycenaean people, who have probably already experimented with this model and spread it where there is suitable local wild material.
Use of local wild grapes
The results of the aDNA analyses showed no evidence of independent domestication of local wild grapes. Nonetheless, a parent–offspring relationship emerged between a domestic/wild hybrid individual (P52) and a domesticated clonal group (P50, P53, P56 and P59). Besides GMM showed that vines producing morphologically wild pips were surely exploited. This highlight the contribution of wild grapes in the early viticulture of this area. We cannot state whether the crossing was voluntary or accidental and we do not know the extent of the phenomenon. However, the presence of clonal groups and hybrids within the sample, as well as the coexistence of wild and domesticated grapes, leads us to suppose that this first stage of viticulture in Middle to the Late Bronze Age Southern Italy included both asexual and sexual propagation of the vines, as well as the introduction of non-native varieties and the use of local wild vines. Selective pressure had to be rather mild and this could explain the proportions between domesticated and wild morphotypes in the Grotta di Pertosa assemblage. Indeed, GMM analyses shows that the percentage of domestic morphotypes reaches 16.5%, while the wild ones are the 69.1%.
Such a consideration is in line with what has emerged from recent studies27,44,100, according to which, viticulture in its initial stage, requiring maintenance of the plants involving low selective pressure and occasional recourse to sexual reproduction, does not necessarily result in drastic changes in the shape of the seeds. This is seen only gradually, in a non-linear manner, as the selective pressure exerted on the plants by human beings increases. It is manifested on the archaeobotanical level by mixed assemblages composed of variable percentages of fully domestic types, wild types and weakly selected types, as in the case presented here, according to a trend that persists even in subsequent historical periods in other areas21. Our study adds a piece to this reconstruction and suggests that, in some cases, the less clear morphotypes could represent wild-domestic hybrids, rather than weakly selected types.
Conclusion
This work has allowed us to establish some fixed points on the diffusion of viticulture in the western Mediterranean. The presence of domestic vines in the south of the Italian peninsula between the Middle and Late Bronze Age, already presumed from other archaeobotanical indicators, has been confirmed beyond any doubt. In this historical phase the indigenous Italic communities experience a growing economic and social complexity and contacts with Mycenaean peoples constitute an important factor of cultural contamination. Within this framework, the introduction of domestic grape varieties from the Aegean area, as well as the growing interest in the cultivation of vines, could represent a reflection of the adhesion by local élites to a system of uses and values spread by Mycenaean sailors throughout the Mediterranean. Local societies, however, do not passively absorb the new models, but make them their own and reinterpret them. It is suggestive to note how this mechanism, observed for some aspects of material culture, is in a certain way also valid for viticulture practices: the domestic grapes introduced from the East, in fact, hybridize with the local wild ones, harvested and exploited in this region for thousands of years, originating something new.
Materials and methods
A total of 55 grape pips from Grotta di Pertosa, dated to the latter period of the Middle Bronze Age to the beginning of the Late Bronze Age (from 1440 to 1200 BCE), were analysed in this study by geometric morphometrics. Although the total number of specimens discovered in the site was greater, only the pips that were perfectly preserved could be taken into account. All the grape pips analysed were waterlogged and were from an inhabited context, precisely from a single stratigraphic unit, formed by the collapsed floor of a pile-dwelling. From these, a group of ten seeds, showing a certain degree of shape diversity, were selected for paleogenetic analysis. All plant experiments were carried out with relevant institutional, national, and international guidelines and legislation.
Geometric morphometric analyses
Geometric morphometrics were based on seed outline analysis using the Elliptic Fourier Transform (EFT) method, which is by now widely known in the literature and has been consolidated over the last decade23,28,44,95. The 55 selected grape pips were photographed in dorsal and lateral views using a stereomicroscope (Olympus SZ-ET) connected to a digital camera (Olympus DP12). Subsequently, these images were semi-automatically converted into black shapes on white backgrounds by means of photo editing software. All the morphometric analyses were then performed using the Momocs package101 in R environment (R Development Core Team 2021). The x and y coordinates of 360 equidistant points were sampled along the outline of each profile. The outlines were normalized by centring and scaling according to their centroid size and the first point was positioned above the centroid. The computation of EFT allows to decompose the seed outlines into a harmonic serie of trigonometric functions described by numerical coefficients that can be used as shape descriptors in statistical analyses. Only the 6 first harmonics were considered, as a good compromise between the need to precisely describe the shape and to minimise the errors24. Each harmonic is associated with 4 coefficients, and thus each outline is described by 24 coefficients, bringing the total number of coefficients used to describe each individual seed to 48.
GMM statistical analyses
All statistical analyses were performed with the MASS package102. Based on the 48 shape descriptors, the morphology of the 55 archaeological seeds was characterized by comparison to modern populations of grape pips using Linear Discriminant Analyses (LDA). A two-step approach was applied using two nested LDAs and the modern reference collection of wild grapevines and cultivated varieties of ISEM29,44. These wild grapes and modern cultivars are originating from various countries from Western Asia to Central and Western Europe and around the Mediterranean. A first LDA was performed to identify the wild or domesticated status of the archaeological pips. The archaeological pips were compared to a balanced modern collection of randomly selected pips of wild (N = 2430) and domesticated (N = 2430) grapes. The power of this method in discriminating between domestic and wild morphotypes has been demonstrated in previous studies24,29. Using this balanced reference collection and leave-one-out cross-validation 95.7% of the pips are correctly classified according to their wild or domesticated status29,44.
The archaeological pips identified as domesticated (p value ≥ 0.75) by this first LDA were then submitted to a second LDA designed to compare their morphology to modern varieties. This second LDA was based on a balanced collection composed of 280 traditional cultivars whose main agronomic traits and areas of origins are known, with ca 30 pips per cultivar. In this case, leave-one-out cross-validation allows the correct cultivar identification of 77.18% of the pips.
aDNA methods
Ancient DNA was recovered and analyzed following methods developed by Ramos-Madrigal, et al.31, with details described in supplementary information and data. Ultrashort DNA was extracted from ten individual seeds103, converted to Illumina libraries using a single-stranded DNA preparation104, treated with uracil–DNA–glycosylase to repair DNA damage105, and enriched using an in-solution kit which targets 10,000 single nucleotide polymorphisms (SNPs)31. Sequencing data were processed in the PALEOMIX pipeline following recommendations for degraded DNA106. Genetic structure and relatedness were investigated in the context of a grapevine reference panel of 783 cultivars46, 112 wild accessions107, and 31 archaeological samples29,31, using smartpca45, KING108, and ngsRelate109.
Data availability
The datasets generated and/or analysed during the current study are available in the NIH repository, https://dataview.ncbi.nlm.nih.gov/object/PRJNA999173?reviewer=j47gnuhdvgai0tj72ljisb78vq.
References
Zohary, D. & Spiegel-Roy, P. Beginnings of fruit growing in the old world: Olive, grape, date, and fig emerge as important Bronze Age additions to grain agriculture in the Near East. Science 187(4174), 319–327. https://doi.org/10.1126/science.187.4174.319 (1975).
McGovern, P. E. Ancient Wine (Princeton University Press, 2013).
McGovern, P. E. et al. Early Neolithic wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. 114, E10309. https://doi.org/10.1073/pnas.1714728114 (2017).
Arroyo-García, R. et al. Multiple origins of cultivated grapevine (Vitis vinifera L. ssp. sativa) based on chloroplast DNA polymorphisms. Mol. Ecol. 15(12), 3707–3714. https://doi.org/10.1111/j.1365-294X.2006.03049.x (2006).
Miller, N. F. Sweeter than wine? The use of the grape in early western Asia. Antiquity 82(318), 937–946. https://doi.org/10.1017/S0003598X00097696 (2008).
Myles, S. et al. Genetic structure and domestication history of the grape. Proc. Natl. Acad. Sci. 108(9), 3530–3535. https://doi.org/10.1073/pnas.1009363108 (2011).
Bacilieri, R. et al. Genetic structure in cultivated grapevines is linked to geography and human selection. BMC Plant Biol. 13(1), 1–14. https://doi.org/10.1186/1471-2229-13-25 (2013).
Riaz, S. et al. Genetic diversity analysis of cultivated and wild grapevine (Vitis vinifera L.) accessions around the Mediterranean basin and Central Asia. BMC Plant Biol. 18(1), 1–14. https://doi.org/10.1186/s12870-018-1351-0 (2018).
Fuller, D. Q. & Stevens, C. J. Between domestication and civilization: The role of agriculture and arboriculture in the emergence of the first urban societies. Veg. Hist. Archaeobotany 28(3), 263–282. https://doi.org/10.1007/s00334-019-00727-4 (2019).
This, P., Lacombe, T. & Thomas, M. R. Historical origins and genetic diversity of wine grapes. Trends Genet. 22, 511–519. https://doi.org/10.1016/j.tig.2006.07.008 (2006).
Grassi, F. et al. Evidence of a secondary grapevine domestication centre detected by SSR analysis. Theor. Appl. Genet. 107(7), 1315–1320. https://doi.org/10.1007/s00122-003-1321-1 (2003).
Dong, Y. et al. Dual domestications and origin of traits in grapevine evolution. Science 379, 892–901. https://doi.org/10.1126/science.add8655 (2023).
Grassi, F. & De Lorenzis, G. Back to the origins: Background and perspectives of grapevine domestication. Int. J. Mol. Sci. 22, 4518. https://doi.org/10.3390/ijms22094518 (2021).
Imazio, S., Somaschini, A., Biagini, B., Grassi, F. & Labra, M. Progetto VINUM: metodi di analisi del genoma e primi risultati. In Archeologia della vite e del vino in Toscana e nel Lazio: dalle tecniche dell’indagine archeologica alle prospettive della biologia molecolare (eds Ciacci, A. et al.) 601–630 (All’insegna del Giglio, 2012).
Levadoux, L. Les populations sauvages et cultivées des Vitis vinifera L Vol. 1 (Institut national de la recherche agronomique, 1956).
Arnold, C., Gillet, F. & Gobat, J.-M. Situation de la vigne sauvage Vitis vinifera ssp. silvestris en Europe. Vitis 37(4), 159–170. https://doi.org/10.5073/vitis.1998.37.159-170 (1998).
Zohary, D. & Hopf, M. Domestication of Plants in the Old World: The Origin and Spread of Cultivated Plants in West Asia, Europe and the Nile Valley 3rd edn. (Oxford University Press, 2000).
Stummer, A. Zur Urgeschichte der Rebe und des Weinbaues. Mitteilungen der Anthropologischen Gesellschaft in Wien 41, 283–296 (1911).
Mangafa, M. & Kotsakis, K. A new method for the identification of wild and cultivated charred grape seeds. J. Archaeol. Sci. 23(3), 409–418. https://doi.org/10.1006/jasc.1996.0036 (1996).
Perret, M. Polymorphisme des génotypes sauvages et cultivés de Vitis vinifera L. à l’aide de marqueurs RAPD. Bulletin de la société Neuchâteloise des sciences naturelles 120, 45–54 (1997).
Bouby, L. et al. Bioarchaeological insights into the process of domestication of grapevine (Vitis vinifera L.) during Roman times in Southern France. PLoS ONE 8, 5. https://doi.org/10.1371/journal.pone.0063195 (2013).
Bonhomme, V. et al. Early viticulture in Neolithic and Bronze Age Greece: Looking for the best traditional morphometric method to distinguish wild and domestic grape pips. In Cooking with Plants in Ancient Europe and Beyond. Interdisciplinary Approaches to the Archaeology of Plant Foods (eds Valamoti, S. M. et al.) 57–70 (Sidestone Press, 2022).
Rovner, I. & Gyulai, F. Computer-assisted morphometry: A new method for assessing and distinguishing morphological variation in wild and domestic seed populations. Econ. Bot. 61(2), 154–172. https://doi.org/10.1663/0013-0001(2007)61[154:CMANMF]2.0.CO;2 (2007).
Terral, J. F. et al. Evolution and history of grapevine (Vitis vinifera) under domestication: New morphometric perspectives to understand seed domestication syndrome and reveal origins of ancient European cultivars. Ann. Bot. 105(3), 443–455. https://doi.org/10.1093/aob/mcp298 (2010).
Orrù, M. et al. Morphological characterisation of Vitis vinifera L. seeds by image analysis and comparison with archaeological remains. Veg. Hist. Archaeobotany 22(3), 231–242. https://doi.org/10.1007/s00334-012-0362-2 (2013).
Landa, V. et al. Accurate classification of fresh and charred grape seeds to the varietal level, using machine learning based classification method. Sci. Rep. 11(1), 1–15. https://doi.org/10.1038/s41598-021-92559-4 (2021).
Pagnoux, C. et al. Inferring the agrobiodiversity of Vitis vinifera L. (grapevine) in ancient Greece by comparative shape analysis of archaeological and modern seeds. Veg. Hist. Archaeobotany 24(1), 75–84. https://doi.org/10.1007/s00334-014-0482-y (2015).
Bonhomme, V. et al. Eco-evo-devo implications and archaeobiological perspectives of trait covariance in fruits of wild and domesticated grapevines. PLoS ONE 15(11), e0239863. https://doi.org/10.1371/journal.pone.0239863 (2020).
Bouby, L. et al. Tracking the history of grapevine cultivation in Georgia by combining geometric morphometrics and ancient DNA. Veg. Hist. Archaeobotany 30(1), 63–76. https://doi.org/10.1007/s00334-020-00803-0 (2021).
Bacilieri, R. et al. Potential of combining morphometry and ancient DNA information to investigate grapevine domestication. Veg. Hist. Archaeobotany 26(3), 345–356. https://doi.org/10.1007/s00334-016-0597-4 (2017).
Ramos-Madrigal, J. et al. Palaeogenomic insights into the origins of French grapevine diversity. Nat. Plants 5(6), 595–603. https://doi.org/10.1038/s41477-019-0437-5 (2019).
Guasch-Jané, M. R. Grape archaeology and ancient DNA sequencing. In The Grape Genome. Compendium of Plant Genomes (eds Cantu, D. & Walker, M.) 57–75 (Springer, 2019).
Wales, N. et al. The limits and potential of paleogenomic techniques for reconstructing grapevine domestication. J. Archaeol. Sci. 72, 57–70. https://doi.org/10.1016/j.jas.2016.05.014 (2016).
Marvelli, S., De Siena, S., Rizzoli, E. & Marchesini, M. The origin of grapevine cultivation in Italy: The archaeobotanical evidence. Annali di Botanica 3, 155–163. https://doi.org/10.4462/annbotrm-10326 (2013).
Breglia, F. Il sito palafitticolo ipogeo di Grotta di Pertosa (SA) nell’età del Bronzo: ricerche archeobotaniche. Doctoral dissertation (University of Salento, 2020).
Larocca, F., Breglia, F., Calcagnile, L., D’Elia, M. & Quarta, G. A radiocarbon chronology for “Grotte di Pertosa” in Campania, Southern Italy. In 24th Radiocarbon Conference 10th 14C & Archaeology Conference (Zurich, 11–16 September 2022). https://radiocarbon24.ethz.ch/wp-content/uploads/2022/08/A05_P12.htm.
Larocca, F. & Breglia, F. Grotte di Pertosa (Salerno). Un contesto archeologico sommerso in ambiente sotterraneo. In Il patrimonio culturale sommerso Ricerche e proposte per il futuro dell’archeologia subacquea in Italia (ed. Capulli, M.) 327–335 (Forum, 2018).
Breglia, F. & Fiorentino, G. La palafitta protostorica. Analisi archeobotaniche in Tra pietra e acqua. In Archeologia delle Grotte di Pertosa-Auletta Studi e ricerche 2004–2016 (ed. Larocca, F.) 40–55 (Fondazione MidA, 2017).
Patroni, G. Caverna naturale con avanzi preistorici in provincia di Salerno, Monumenti Antichi della Reale Accademia dei Lincei IX, 545–616 (1899).
Carucci, P. La grotta preistorica di Pertosa (Salerno). Contribuzione alla Paletnologia, Speleologia ed Idrografia, (Napoli, 1907).
Larocca, F. Tra pietra e acqua. Archeologia delle grotte di Pertosa-Auletta. Studi e ricerche 2004–2016 (Fondazione MIdA, 2017).
Fuscone, A. Grotta Pertosa: prima nota sui materiali ceramici del Bronzo Tardo e Primo Ferro dagli scavi Patroni. In Archeologi in progress. Il cantiere dell’archeologia di domani (eds Brancato, R. et al.) 60–70 (BraDypUS Communicating Cultural Heritage, 2015).
De Falco, M. & Larocca, F. Il luogo sacro. Aspetti e caratteri dell’utilizzo cultuale della cavità. In Tra pietra e acqua. Archeologia delle grotte di Pertosa-Auletta Studi e ricerche 2004–2016 (ed. Larocca, F.) 120–135 (Fondazione MIdA, 2017).
Pagnoux, C. et al. Local domestication or diffusion? Insights into viticulture in Greece from Neolithic to Archaic times, using geometric morphometric analyses of archaeological grape seeds. J. Archaeol. Sci. 125, 105263. https://doi.org/10.1016/j.jas.2020.105263 (2021).
Price, A. L. et al. Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 38, 904–909. https://doi.org/10.1038/ng1847 (2006).
Laucou, V. et al. Extended diversity analysis of cultivated grapevine Vitis vinifera with 10K genome-wide SNPs. PLoS ONE 13(2), e0192540. https://doi.org/10.1371/journal.pone.0192540 (2018).
Myles, S. et al. Rapid genomic characterization of the genus Vitis. PLoS ONE 5(1), e8219. https://doi.org/10.1371/journal.pone.0008219 (2010).
Cattani, L., Colonese, A. & Ricciardi, S. Ricostruzione climatica e ambientale di Grotta del Romito: analisi palinologiche e malacofaunistiche dei livelli epigravettiani C e D. In Atti della XXXVII Riunione Scientifica Preistoria e Protostoria della Calabria 637–640 (Istituto Italiano di Preistoria e Protostoria, Firenze, 2004).
Costantini, L. Semi e carboni del Mesolitico e Neolitico della Grotta dell’Uzzo, Trapani. Quaternaria 23, 233–247 (1982).
Acconcia, V. & Piergrossi, A. L’archeologia del vino nella penisola italiana e nelle grandi isole del Tirreno tra il neolitico e la romanizzazione: tematiche, dati e approcci possibili. Bollettino di Archeologia on line, XII (2021).
Accorsi, C. A. Bandini Mazzanti, M. Fiorentino, G. Gorgoglione, M. A. & Mercuri A. M. Archaeological and archaeobotanical data on the mesolithic/ancient–medium neolithic site of Terragne (Taranto-Southern Italy, 96 m a.s.l.,40º24′N 17º38′E). In Proceedings of 1st International Congress Science and Technology for Safeguard of Cultural Heritage in the Mediterranean Basin (Catania, 1995), 1521–1527 (Tipolitogr. Luxograph, 1998).
Brun, J. P. Archéologie du vin et de l’huile: de la préhistoire à l’époque Hellénistique (Errance, 2004).
Costantini, L. & Costantini Biasini, L. La viticoltura dalla Grecia alla Magna Grecia: la documentazione archeobotanica. In Alle radici della civiltà del vino in Sicilia: atti del convegno Alle radici della civiltà del vino nel Mediterraneo: 3000 anni di storia (Menfi, 6 luglio 1996) (eds Failla, O. & Forni, G.) 169–192 (Cantine Settesoli, 1999).
Morter, J. The Chora of Croton 1: the Neolithic settlement at Capo Alfiere (ed Robb, J.) (University of Texas Press, 2010).
Gismondi, A. et al. Grapevine carpological remains revealed the existence of a Neolithic domesticated Vitis vinifera L. specimen containing ancient DNA partially preserved in modern ecotypes. J. Archaeol. Sci. 69, 75–84. https://doi.org/10.1016/j.jas.2016.04.014 (2016).
Carra, M. Ambiente ed economia di sussistenza nell’età del bronzo. Analisi paleocarpologica dei siti perilacustri di “Villaggio delle Macine” (Castel Gandolfo, Roma) e Castellaro Lagusello (Mantova): due realtà a confronto. In Atti del I Convegno Nazionale degli Studenti di Antropologia, Preistoria e Protostoria (Ferrara, 8–10 Maggio 2004) (ed. Hohenstein, U. T.) 79–82 (Annali dell’Università di Ferrara, 2007).
Castiglioni, E., Motella De Carlo, S. & Nisbet, I. Indagini sui resti vegetali macroscopici a Canàr. In Canàr di S. Pietro Polesine Ricerche archeo-ambientali sul sito palafitticolo (eds Balista, C. & Bellintani, P.) 115–130 (Rovigo, 1998).
Costantini, L., Laria, M. & Tecchiati, U. I resti carpologici dell’antica e media età del Bronzo del riparo del Santuario di Lasino (Trento)-scavi 1996. Annali del Museo Civico di Rovereto, Sezione Archeologia, Storia e Scienze Naturali 17, 3–40 (2001).
De Marinis, R. C. et al. Lavagnone (Desenzano del Garda): new excavations and palaeoecology of a Bronze Age pile dwelling in northern Italy In Wetland economies and Societies. Proceedings of the International Conference in Zürich, 10–13 March 2004. (eds. Della Casa, P. & Trachsel, M.) 221–232 (Collectio Archaeologica 3, 2005).
Aranguren, B. & Perazzi, P. L. struttura interrata della media età del bronzo di San Lorenzo a Greve a Firenze e l’inizio della coltivazione della vite in Toscana. Rivista di Scienze Preistoriche LVII, 243–262 (2007).
Carra, M. Alimentazione, ambiente ed economia di sussistenza su base vegetale. Studio archeobotanico preliminare dei macroresti provenienti dal sito di Solarolo. Ipotesi di Preistoria 2, 282–291 https://doi.org/10.6092/issn.1974-7985/1613 (2009).
Castelletti, L. & Motella De Carlo, S. L’uomo e le piante nella preistoria. L’analisi dei resti macroscopici vegetali. In Archeologia in Piemonte. La preistoria Vol. 1 (eds Mercando, L. & VenturinoGambari, M.) 41–56 (Umberto Allemandi C., 1998).
Bellini, C., Mariotti Lippi, M., Mori Secci, M., Aranguren, B. & Perazzi, P. Plant gathering and cultivation in prehistoric Tuscany (Italy). Veg. Hist. Archaeobotany 17(1), 103–112. https://doi.org/10.1007/s00334-008-0189-z (2008).
Mariotti Lippi, M., Bellini, C. & Mori Secci, M. Paleovegetational reconstruction based on pollen and seeds/fruits from a Bronze Age archeological site in Tuscany (Italy). Plant Biosyst. 144(4), 902–908. https://doi.org/10.1080/11263504.2010.491978 (2010).
Fiorentino, G., Colaianni, G., Grasso, A. M. & Stellati, A. Caratteristiche del paleoambiente e modalità di sfruttamento dei vegetali a Salina nel corso dell’Età del Bronzo. In Il villaggio dell’età del Bronzo di Portella nelle Isole Eolie (scavi 2006–2008) (ed. Martinelli, M. C.) 234–241 (Rebus Edizioni, 2011).
Bakels, C. Plant remains from Sardinia, Italy, with notes on barley and grape. Veg. Hist. Archaeobotany 11, 3–8. https://doi.org/10.1007/s003340200334 (2002).
Bettini, M.C. & Zanini, A. I Forti: abitato della fine dell’età del bronzo nell’area urbana di Chiusi. In Preistoria e Protostoria in Etruria, Atti incontro di studio, Manciano, 17–19 maggio 1991. (ed. Negroni Catacchio, N.) 315–324 (1993).
Carra, M., Cattani, L. & Zanni, C. Aspetti paletnobotanici dell’area insediativa protostorica di S. Maria in Belvedere sul Monte Cetona (Siena). Rivista di Scienze Preistoriche LIII, 505–518 (2003).
Fiorentino, G. Caratteristiche della vegetazione e abitudini alimentari durante la preistoria. In Le Isole Chéradi fra natura, leggenda e storia (eds Mastronuzzi, G. & Marzo, P.) 69–78 (Stampasud, 1999).
Primavera, M. Roca e le dinamiche uomo-ambiente in Puglia durante l’età del Bronzo. Le piante e l’uomo tra ambiente paesaggio economia e culto (Edizioni Esperidi, 2018).
Primavera, M. & Fiorentino, G. Lo sfruttamento dei frutti arborei spontanei e l’arboricoltura in Puglia durante l’età del Bronzo. Studi di preistoria e protostoria 6, 527–534 (2021).
Rottoli, M. I resti vegetali. In Tarquinia. Testimonianze archeologiche e ricostruzione storica. Scavi sistematici nell’abitato (Campagne 1982–1988) (eds Bonghi Jovino, M. & Chiaramonte Trerè, C.) 92–99 (1997).
Ucchesu, M. et al. Earliest evidence of a primitive cultivar of Vitis vinifera L. during the bronze age in Sardinia (Italy). Veg. Hist. Archaeobotany 24, 587–600. https://doi.org/10.1007/s00334-014-0512-9 (2015).
Usai, A. et al. L’insediamento nuragico di Sa Osa (Cabras, OR). Il sito e i materiali archeobotanici. Rivista di Storia dell’Agricoltura LVI, 109–121 (2016).
Di Rita, F. & Magri, D. Holocene drought, deforestation, and evergreen vegetation development in the central Mediterranean: A 5500 year record from Lago Alimini Piccolo, Apulia, southeast Italy. Holocene 19, 295–306. https://doi.org/10.1177/0959683608100574 (2009).
Caldara M. et al. Primi risultati di una ricerca paleoambientale nell’area di Coppa Nevigata (Foggia). In Atti del 19° Convegno Nazionale sulla Preistoria–Protostoria–Storia della Daunia (San Severo 27–29 novembre 1998), v1 (ed. Gravina A.) 199–236 (Archeoclub San Severo, 1999).
Di Rita, F. et al. Holocene forest dynamics in central and western Mediterranean: Periodicity, spatio-temporal patterns and climate influence. Sci. Rep. 8, 8929. https://doi.org/10.1038/s41598-018-27056-2 (2018).
Castiglioni, E. & Rottoli, M. Capua (Caserta). Località Strepparo e Cento Moggie. Scavi nell’area del CIRA. Resti botanici da un pozzo dell’età del Bronzo. Bollettino di Archeologia 37–38, 62–67 (1996).
Fiorentino, G. & D’Oronzo, C. Analisi dei macroresti vegetali: strategie agronomiche, alimentazione e caratteristiche del paleoambiente a Coppa Nevigata nel corso dell’età del Bronzo. In Coppa Nevigata e l’area umida alla foce del Candelaro durante l’età del Bronzo (eds Cazzella, A. et al.) 327–337 (Foggia, 2012).
Ross Holloway, R. Buccino: The early bronze age village of Tufariello. J. Field Archaeol. 2(1/2), 11–81. https://doi.org/10.2307/529618 (1975).
De Lorenzis, G. et al. SNP genotyping elucidates the genetic diversity of Magna Graecia grapevine germplasm and its historical origin and dissemination. BMC Plant Biol. 19, 7. https://doi.org/10.1186/s12870-018-1576-y (2019).
Mercati, F. et al. Integrated bayesian approaches shed light on the dissemination routes of the Eurasian Grapevine Germplasm. Front. Plant Sci. 12, 692661. https://doi.org/10.3389/fpls.2021.692661 (2021).
Tanasi, D. Sicily before the Greeks. The interaction with Aegean and the Levant in the Pre-colonial Era. Open Archaeol. 6, 172–205. https://doi.org/10.1515/opar-2020-0107 (2020).
Buxeda, I. et al. Technology transfer at the periphery of the Mycenaean world: The cases of Mycenaean pottery found in Central Macedonia (Greece) and the plain of Sybaris (Italy). Archaeometry 45, 263–284. https://doi.org/10.1111/1475-4754.00108 (2003).
Blake, E. The Mycenaeans in Italy: A minimalist position. Pap. Br. School Rome 76, 1–34. https://doi.org/10.1017/S0068246200000398 (2008).
Jung, R. & Mehofer, M. Mycenaean Greece and Bronze Age Italy: Cooperation, trade or war?. Archäologisches Korrespondenzblatt 43(2), 175–193. https://doi.org/10.11588/ak.2013.2.51814 (2013).
Jones, R. E. et al. Italo-Mycenaean and other Aegean-influenced pottery in Late Bronze Age Italy: The case for regional production. J. Archaeol. Anthropol. Sci. 13, 23. https://doi.org/10.1007/s12520-020-01245-5 (2021).
Meiri, M. P., Stockhammer, W., Morgenstern, P. & Maran, J. Mobility and trade in Mediterranean antiquity: Evidence for an ‘Italian connection’ in Mycenaean Greece revealed by ancient DNA of livestock. J. Archaeol. Sci. Rep. 23, 98–103. https://doi.org/10.1016/j.jasrep.2018.10.011 (2019).
Breglia, F., Sellitto, A. & Fiorentino, G. Human-environment interaction during the protohistory in the Vallo di Diano (Southern Italy): New data from archaeobotanical analyses. In 18th Conference of the International Workgroup for Palaeoethnobotany (Lecce, 3–8 june 2019) Abstract book (eds. Fiorentino, G. et al.) 167 (University of Salento Press, 2019).
Jones, G., Valamoti, S. & Charles, M. Early crop diversity: A “new” glume wheat from northern Greece. Veg. Hist. Archaebotany 9, 133–146. https://doi.org/10.1007/BF01299798 (2000).
Kenéz, Á., Pető, Á. & Gyulai, F. Evidence of ‘new glume wheat’ from the Late Neolithic (Copper Age) of south-eastern Hungary (4th millennium cal. B.C.). Veg. Hist. Archaeobotany 23, 551–566. https://doi.org/10.1007/s00334-013-0405-3 (2014).
Toulemonde, F. et al. Records of “new” glume wheat in France: A review. Veg. Hist. Archaeobotany 24, 197–206. https://doi.org/10.1007/s00334-014-0479-6 (2015).
Perego, R. Contribution to the development of the Bronze Age plant economy in the surrounding of the Alps: an archaeobotanical case study of two Early and Middle Bronze Age sites in northern Italy (Lake Garda region). Doctoral dissertation (University of Basel, 2015).
Ulaş, B. & Fiorentino, G. Recent attestations of “new” glume wheat in Turkey: A reassessment of its role in the reconstruction of Neolithic agriculture. Veg. Hist. Archaeobotany 30(5), 685–701. https://doi.org/10.1007/s00334-020-00807-w (2021).
Roushannafas, T., Bogaard, A. & Charles, M. Geometric morphometrics sheds new light on the identification and domestication status of ‘new glume wheat’ at Neolithic Çatalhöyük. J. Archaeol. Sci. 142, 105599. https://doi.org/10.1016/j.jas.2022.105599 (2022).
Bettelli, M., Levi, S. T., Jones, R. E. & Vagnetti, L. L. ceramiche micenee in area medio tirrenica: nuove prospettive. In Studi di protostoria in onore di Renato Peroni (eds Cardarelli, A. et al.) 399–406 (All’Insegna del Giglio, 2006).
Matricardi, E., Jung, R., Mommsen, H., Pacciarelli, M. & Sterba, J. H. Aegean-type and aegeanizing pottery from Ionian Apulia. New studies and provenance analyses. Origini 44, 111–148 (2021).
Piperno, M. & Pellegrini, E. Risultati delle ricerche alla grotta del Pino (Sassano, Salerno) 1997–1998. Bullettino di Paletnologia Italiana 91–92, 121–206, (2000–2001).
D’Agostino, B. Un frammento del Vallo di Diano. Dialoghi di Archeologia VI, 5–10 (1972).
Bonhomme, V. et al. Seed morphology uncovers 1500 years of vine agrobiodiversity before the advent of the Champagne wine. Sci. Rep. 11, 2305. https://doi.org/10.1038/s41598-021-81787-3 (2021).
Bonhomme, V., Picq, S., Gaucherel, C. & Claude, J. Momocs: Outline analysis using R. J. Stat. Softw. 56, 1–24. https://doi.org/10.18637/jss.v056.i13 (2014).
Venables, W. N. & Ripley, B. D. Random and mixed effects. In Modern Applied Statistics with S (eds Venables, W. N. & Ripley, B. D.) 271–300 (Springer, 2002).
Dabney, J. et al. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proc. Natl. Acad. Sci. 110, 15758–15763. https://doi.org/10.1073/pnas.1314445110 (2013).
Gansauge, M.-T. & Meyer, M. Single-stranded DNA library preparation for the sequencing of ancient or damaged DNA. Nat. Protocols 8, 737–748. https://doi.org/10.1038/nprot.2013.038 (2013).
Rohland, N., Harney, E., Mallick, S., Nordenfelt, S. & Reich, D. Partial uracil–DNA–glycosylase treatment for screening of ancient DNA. Philos. Trans. R. Soc. B Biol. Sci. 370, 20130624. https://doi.org/10.1098/rstb.2013.0624 (2015).
Schubert, M. et al. Characterization of ancient and modern genomes by SNP detection and phylogenomic and metagenomic analysis using PALEOMIX. Nat. Protocols 9, 1056–1082. https://doi.org/10.1038/nprot.2014.063 (2014).
Le Paslier M-C. et al. A dataset of 9.896 single nuclear polymorphisms for 112 wild grapes, obtained with the GrapeReSeq 18K Vitis chip. Recherche Data Gouv. https://doi.org/10.15454/9RUCEP, Deposited 19 April 2019.
Manichaikul, A. et al. Robust relationship inference in genome-wide association studies. Bioinformatics 26, 2867–2873. https://doi.org/10.1093/bioinformatics/btq559 (2010).
Hanghøj, K., Moltke, I., Andersen, P. A., Manica, A. & Korneliussen, T. S. Fast and accurate relatedness estimation from high-throughput sequencing data in the presence of inbreeding. GigaScience 8, giz034. https://doi.org/10.1093/gigascience/giz034 (2019).
Acknowledgements
A portion of this project was undertaken on the Viking Cluster, which is a high performance compute facility provided by the University of York. We are grateful for computational support from the University of York High Performance Computing service, Viking and the Research Computing team. This project has received funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 842577, the ANR MICA project (agreement No ANR-22-CE27-0026).
Author information
Authors and Affiliations
Contributions
F.B. carried out the field research, the study of the archaeobotanical and archaeological aspects in the historic-geographic context, as well as setting up the overall research work and writing the introduction, the description of the site and the discussion paragraphs. L.B. and S.I. performed the geometric morphometric analysis and its statistical processing, as well as writing the related results and methods paragraphs. N.W. performed the paleogenetic analysis and wrote the related results and methods paragraphs. G.F. supervised the overall work and, together with the other authors, contributed to an organic reading, writing and interpretation of the results in the light of the archaeobotanical and archaeological context.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Breglia, F., Bouby, L., Wales, N. et al. Disentangling the origins of viticulture in the western Mediterranean. Sci Rep 13, 17284 (2023). https://doi.org/10.1038/s41598-023-44445-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-44445-4
- Springer Nature Limited