
Abstract
Majority of the pollination related studies are based on the diurnal pollinators, and the nocturnal pollinators received less scientific attention. We reveal the significance of settling moths in pollination of angiosperm families in Himalayan ecosystem of North-East India. The refined and novel method of pollen extraction from the proboscides provides a more robust assessment of the pollen carrying capacity. The study is based on one of the largest data sets (140 pollen transporter moth species (PTMS)), with interpretation based on seasonal as well as altitudinal data. In the present study about 65% moths (91 species) carried sufficient quantities of pollen grains to be considered as potential pollinators (PPMS). Teliphasa sp. (Crambidae) and Cuculia sp. (Noctuidae) are found to carry the highest quantity of pollen. We found pollen grains of 21 plant families and the abundant pollen are from Betulaceae, Fabaceae, Rosaceae and Ericaceae. Species composition of PTMS and PPMS in pre-monsoon, monsoon, and post-monsoon revealed the dominance of Geometridae. Maximum diversity of PTMS and PPMS is found from 2000 to 2500 m altitude. The nocturnal pollen transfer network matrices exhibited high degree of selectivity (H2ʹ = 0.86).
Similar content being viewed by others

Introduction
Pollination is an indispensable ecological process for the continuity of germplasm1 and is mainly driven by wind, water and animals as vectors1,2. About 87.5% of all angiosperms are mainly pollinated by animals3,4,5,6,7. Among these, pollinating insects have important mutualistic relationships with angiosperms, which are essential for the conservation of wild as well as agricultural landscapes2,8.
So far, most research on pollinating insects has focused on diurnal pollinators, and the nocturnal pollinators have traditionally received less scientific attention9. Despite recent research in the last many years on their role in pollination, many questions left unanswered. An important review by Macgregor and co-workers revealed that 289 species of plants (mostly angiosperms from the orders Caryophyllales, Ericales, Gentianales, and Lamiales) under 75 families are partially or entirely pollinated by 21 moth families8. Some limited studies related to the comparable importance of the moths as pollinators to that of diurnal pollinators10,11,12, advocate the benefits of moths over the diurnal pollinators through mechanisms such as (1) longer-distance dispersal of pollen13,14, (2) good seed set despite deposition of fewer pollinia15,16,17 and (3) increased pollination efficiency due to pollen deposition in a single visit18.
Among the moths, the settling moths (mainly of families other than Sphingidae, which hover when feeding from flowers) are extremely common and diverse flower visitors19 but are less studied for their role in pollination. Many studies have focused on pollination by Sphingidae (i.e. the Sphingophily)20,21,22,23,24,25,26,27,28,29,30,31. The role of settling moths as pollinators i.e. the Phalaenophily29 has also been studied, but arguably to a lesser degree19,27. Settling moths may potentially be effective pollinators of generalist plants which are not visited by hawkmoths or where the hawkmoths are not common32.
The noteworthy studies related to pollination by settling moths are sporadic, few and limited to certain regions of the world viz. in eastern England12, East Asia33,34, North Europe and America35,36. Other studies conducted on the pollination of plant families, Asteraceae, Ericaceae, Plantaginaceae, Myrtaceae, Campanulaceae, Thymelaeaceae, revealed the participation of settling moths12,27,37,38.
The Indian moths are negligibly studied for their role in pollination and only four papers could be reviewed for this aspect. Paul39 studied the pollination efficiency of noctuid moth species in urban areas of Delhi. Sarkar and Sreedevi40 studied the nocturnal insect pollinators of bottle gourd and ridge gourd crop of Andhra Pradesh and reported three species of settling moths, Arthroschista hilaralis (Crambidae), Diaphania indica (Crambidae) and Anadevidias peponis (Noctuidae), as major pollinators. The seasonal dynamics of plant-pollinator networks in agricultural landscapes of West Bengal and the pollination of medicinal plants in Tripura revealed the participation of settling moths41,42. The aim of the present study is to investigate the role of settling moths in pollination of angiosperm families distributed in Himalayan ecosystem of North-East India by quantifying various network level indices along with the effect of various seasons and altitudinal gradient on the pollen carrying capacity. The pollen transport does measure pollination success, but is a proxy whereby we can begin to assess involvement in pollination process43. The present study is based on the dataset generated from the light trapping at 24 field sites in Himalayan ecosystem of North-East India, over the period of 13 months.
Results
Settling moths as pollen vectors
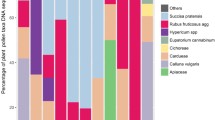
We identified 140 moth species in 18 subfamilies and 6 families of settling moths carrying pollen grains on their proboscis, and termed them as ‘pollen transporter moth species’ (PTMS). Out of the 140 PTMS, the proboscides of 91 moth species are found to carry five or more than five pollen grains of one or another plant family. Herein, we termed the 91 moth species as ‘potential pollinator moth species’ (PPMS) by following Macgregor et al.8 and Devoto et al.44 (Figs. 1, 2; see supplementary Table S1 and S2 of supplementary file 1). On the proboscides of studied moth species, we found the pollen grains of 21 plant families viz. Acanthaceae, Anacardiaceae, Apocynaceae, Asteraceae, Balsaminaceae, Betulaceae, Elaeocarpaceae, Ericaceae, Euphorbiaceae, Fabaceae, Fagaceae, Lamiaceae, Malvaceae, Myrtaceae, Oleaceae, Passifloraceae, Plantaginaceae, Poaceae, Rosaceae, Salicaceae and Verbenaceae. The pollen grains of Betulaceae are most frequently found (39.89% of total pollen grains found on proboscides of all PTMS), followed by Fabaceae (26.62%), Rosaceae (15.44%) and Ericaceae (7.66%). Further, we found the largest number of PTMS came from the Geometridae (53 species; 37.85%), followed by Erebidae (50 species; 35.71%), Crambidae and Noctuidae (14 species each; 10%), Drepanidae (7 species; 5%) and Nolidae (2 species, 1.42%). Composition of PPMS is similar, with highest number from Geometridae (36 species; 39.56%), followed by Erebidae (31 species; 34.06%), Noctuidae (11 species; 12.08%), Crambidae (8 species; 8.79%), Drepanidae (3 species; 3.29%) and Nolidae (2 species; 2.19%). Of the 91 PPMS, 60 species carried the pollen of a single plant family, and 23 species carried pollen grains of two plant families. Only six moth species i.e., Hypocala sp. (Erebidae: Arctiinae), Callindra nyctemerata (Erebidae: Arctiinae), Achaea janata (Erebidae: Erebinae), Oxyodes scrobiculata (Erebidae: Erebinae), Thysanoplusia orichalcea (Noctuidae: Plusiinae) and Cyclidia substigmaria (Drepanidae: Cyclidinae) carried pollen grains of three plant families and two moth species, Ourapteryx sp. (Geometridae: Ennominae) and Sarbanissa catacoloides (Noctuidae: Agaristinae) carried pollen of five plant families. Teliphasa sp. (Epipaschiinae: Crambidae) and Cuculia sp. (Noctuidae: Cuculliinae) are found to carry the highest quantity of pollen (more than 1000 pollen grains of Fabaceae and Betulaceae).

Pie-chart of ten major plant families and their pollen load percentage found on proboscides of PTMS.

Family-wise species composition of PTMS and PPMS.
Season-wise distribution of PTMS and PPMS at different altitudes in the Himalayan ecosystem
In our study, moths are found to carry pollen across the altitudinal gradient sampled up to 3000 m, however, pollen composition from different families varied at different altitude and in different seasons. The maximum number of PTMS are found in sub-tropical zone (1000–2000 m) (74 species; 52.85%), followed by temperate zone (2000–3000 m) (69 species; 49.28%), and in tropical zone (0–1000 m) (9 species; 6.42%). Drepanidae transported pollen in higher altitudes, beyond 2500 m.
Pre-monsoon season
In pre-monsoon, we found 54 and 40 PTMS and PPMS, respectively. 25 PTMS (46.29%) came from Erebidae, followed by Geometridae (13; 24.07%), Noctuidae (8; 14.81%), Crambidae (6; 11.11%) and Drepanidae (2; 3.70%) (Fig. 3; see supplementary Table S3 of supplementary file 1). No Nolidae are recorded from this season. During the pre-monsoon, moths carried the pollen of 15 plant families (Fabaceae, Betulaceae, Verbenaceae, Ericaceae, Balsaminaceae, Malvaceae, Poaceae, Rosaceae, Salicaceae, Elaeocarpaceae, Oleaceae, Asteraceae, Plantaginaceae, Fagaceae and Euphorbiaceae).

Season-wise species composition of PTMS.
Altitudinally, in tropical zone (0–1000 m), a single PTMS, Pelagodes sp. (Geometridae) is reported. In sub-tropical zone (1000–2000 m), we found 23 PTMS, of which Erebidae dominated with 11 PTMS, followed by Geometridae (6 PTMS), Noctuidae (5 PTMS) and Drepanidae (1 PTMS). No PTMS are recorded in Crambidae and Nolidae. In this zone, Cuculia sp. (Erebidae) reported to transport maximum (more than 1000) pollen grains, followed by Pericyma umbrina (223) and Abraxas sp. (103). In sub-tropical zone, PTMS transported maximum pollen grains of Rosaceae, followed by Ericaceae, Fabaceae and Betulaceae, along with pollen of other 5 plant families. In temperate zone (2000–3000 m), we report 41 PTMS, with Erebidae having the highest PTMS (18), followed by Geometridae (12 PTMS), Crambidae (6 PTMS), Noctuidae (3 PTMS) and Drepanidae (2 PTMS). No PTMS are found in Nolidae (see supplementary file 2). In this zone, Teliphasa sp. (Crambidae) transported maximum pollen (more than 1000), followed by Vamuna remelana (200) and Semiothisa sp. (175). In temperate zone, PTMS transported the pollen of 14 plant families, with Fabaceae dominated the pollen composition.
Monsoon season
In monsoon season, we found 50 and 31 PTMS and PPMS, respectively. Of which, 23 PTMS (46%) are Geometridae, followed by Erebidae (18; 36%), Noctuidae (4; 8%), Crambidae and Drepanidae (2 each; 4%) and only one PTMS from Nolidae (Fig. 3; see supplementary Table S3 of supplementary file 1). We found the pollen of 9 different plant families (Rosaceae, Fabaceae, Betulaceae, Ericaceae Oleaceae, Malvaceae, Salicaceae, Poaceae and Passifloraceae), of which first four families dominated the pollen composition. Among the three seasons, this is the lowest number of plant families represented on the proboscides of moths.
Altitudinally, in tropical zone (0–1000 m), we report 9 PTMS transporting pollen of 3 plant families, Betulaceae, Fabaceae and Rosaceae. Among pollen transporting moths, Erebidae dominated with 4 PTMS (44% ). No PTMS are found in Nolidae and Drepanidae. In sub-tropical zone (1000–2000 m), 30 PTMS are reported to transport pollen of 8 plant families. Geometridae dominated with 14 PTMS, followed by Erebidae (11 PTMS). In this zone, Sarcinodes aequilinearia (Geometridae) transported maximum pollen, followed by Percnia felinaria (Geometridae), and Siglophora ferreilutea (Nolidae). No PTMS are reported from Crambidae. In sub-tropical zone, pollen grains of Rosaceae are transported maximum in numbers followed by Ericaceae, Fabaceae and Betulaceae. In temperate zone (2000–3000 m), 15 PTMS are found to carry pollen grains of 7 plant families. In this zone, Geometridae dominated with 7 PTMS, followed by Erebidae (6 PTMS). No PTMS are recorded in Nolidae and Drepanidae (see supplementary file 2). Sarcinodes restitutaria (Geometridae) transported highest pollen, followed by Achaea janata (Erebidae). In this zone, PTMS transported highest pollen of Rosaceae, followed by Betulaceae and Ericaceae.
Post-monsoon season
In post-monsoon season, of the 56 PTMS and 29 PPMS respectively, 25 PTMS (44.64%) are Geometridae, followed by Erebidae (17; 30.35%), Crambidae (7; 12.5%), Drepanidae (4; 7.14%), Noctuidae (2; 3.57%) and Nolidae (1; 1.78%) (Fig. 3; see supplementary Table S3 of supplementary file 1). During the season, moths carried the pollen of 16 plant families, of which, pollen of Euphorbiaceae, Salicaceae, Acanthaceae, Anacardiaceae, and Lamiaceae are very few.
Altitudinally, in post-monsoon, no PTMS are reported from tropical zone (0–1000 m). In sub-tropical zone (1000–2000 m), 32 PTMS are reported to carry pollen of 14 plant families. Geometridae dominated with 23 PTMS, followed by Erebidae (7 PTMS). In this zone, Dindica olivacea (Geometridae) transported maximum pollen, followed by Ectropis sp. (Geometridae), and Cyana bellissima (Erebidae). No PTMS are recorded from Crambidae, Nolidae and Drepanidae. Pollen of Betulaceae are transported in maximum, followed by Fabaceae, Poaceae and Myrtaceae. In temperate zone (2000–3000 m), we found 15 PTMS transporting pollen of 9 plant families, with Geometridae dominating the PTMS composition (7 PTMS), followed by Erebidae (6 PTMS). In this zone, major PTMS are Tyana sp. (Nolidae), and Hesudra divisa (Erebidae). No PTMS are recorded from Drepanidae (see supplementary file 2). PTMS are found to carry maximum pollen of Betulaceae followed by Malvaceae and Fabaceae.
Pollen transfer networks
The settling moth-plant pollen transfer network comprises 140 moth species and 21 plant families (Figs. 4, 5; Based upon supplementary file 2; the script for the bipartite network construction is given as supplementary file 3). In the first network (Fig. 5) we present the consolidated data (for all seasons and all altitudes) and in the second network (Fig. 6) the data is analysed seasonally and altitudinally. The consolidated network has 1.31 links per species with a linkage density of 5.91 and 0.48 as the interaction evenness. The interaction richness is found to be 3.90. The network shows the high degree of selectivity (H2ʹ = 0.86) with nested interaction (nestedness = 5.41). All the calculated network matrices are provided in Table 1. The PDI score (supplementary file 4) indicate that 9 plant families viz. Acanthaceae, Anacardiaceae, Apocynaceae, Balsaminaceae, Fagaceae, Lamiaceae, Myrtaceae, Passifloraceae and Plantaginaceae show complete specialism in relation with moths. Similarly in case of moths we found that 98 settling moth species exhibit specialism and rest are generalists .

Circos plot to visualise the interaction of moth families with the plant families (based upon supplementary file 2).

Consolidated settling moth-plant pollen transfer network (width of higher-level boxes indicates number of pollen grain carried by moths whereas width of lower level boxes indicate total pollen grain of a plant family transported).

Seasonal and altitudinal wise settling moth-plant pollen transfer network (dominating plant families and moth families are coloured).
Season-wise and altitudinal zone-wise pollen transfer network of settling moths
The season-wise and altitudinal zone-wise settling moth-plant pollen transfer network is constructed to get the information on variation in pollen transfer networks in various seasons across different altitudinal zones in Himalayan ecosystem of North-East India (Fig. 6 based upon supplementary file 2; for network matrices see supplementary file 4). Due to lesser number of PTMS as well as plant families, the matrices for data of tropical zone are not analysed.
Pre-monsoon season
During this season the bipartite moth-plant pollen transfer network in sub-tropical zone (1000–2000 m) shows higher values of connectance (0.15), nestedness (20.28), interactive evenness (0.36) and linkage density (2.04), whereas, in temperate zone (2000–3000 m), the network shows higher values of links per species (0.94), Shannon’s diversity (2.21), and interaction strength asymmetry (0.53). The PDI score indicate that the number of specialist and generalist moth species, and the number of specialist and generalist plant families are higher in temperate zone. Interestingly the selectivity (H2’) remained same for both zones i.e., 0.98 (Table 1; Based upon supplementary file 2; see supplementary file 4).
Monsoon season
In monsoon, the bipartite network in sub-tropical zone (1000–2000 m) shows higher values of interactive evenness (0.47), linkage density (2.74), links per species (1), and Shannon’s diversity (2.55) whereas, in temperate zone (2000–3000 m), the network shows higher values of connectance (0.17), nestedness (31.12) (Table 1). The PDI score indicate that the number of specialist and generalist moth species and generalist plant families is higher in sub-tropical zone (see supplementary file 4). Selectivity (0.96) is higher in sub-tropical zone.
Post-monsoon season
In post-monsoon season, values of the network matrices viz. links per species (1), Shannon’s diversity (2.68) and linkage density (2.75) are higher in sub-tropical zone (1000–2000 m) whereas, the values of connectance (0.14), nestedness (25.64), interaction evenness (0.45) and interaction strength asymmetry (0.40) are higher in temperate zone (2000–3000 m). The number of specialist moth species, specialist plant families, generalist moth species are higher in sub-tropical zone but the number of generalist plant families remain same for both sub-tropical and temperate zone. Selectivity (H2ʹ = 0.89) is higher in temperate zone (Table 1 based upon supplementary file 2; see supplementary file 4).
Discussion
Our results are based on one of the largest data sets (140 species) interpreting the role of settling moths in pollination and is the first of its kind to understand the settling moths-plant pollination interactions in the central and eastern Himalayan ecosystem of India. Our approach of using the proboscis by isolating from the moth head and relaxing it for pollen extraction, provides a more robust assessment of the pollen carrying capacity of the moths, because (1) by using the proboscis for the pollen collection, the likelihood of contamination due to pollen rain is reduced (as swabbing of other parts of body result in collection of extra pollen grains from the body, unrelated to flower visitation), and (2) de-coiling the proboscis enables the study of pollen grains stuck within the coiling of the proboscis (which may remain there during swabbing).
We found that substantial proportion (about 65%) of PTMS carried sufficient quantities of pollen grains (five or more than five) to be considered as potential pollinators (PPMS). Geometridae and Erebidae turned out to be the most important for the pollen transportation in the Himalayan region and on the other hand, pollen grains of Betulaceae, predominantly a wind-pollinated plant family, are transported in maximum. In our analysis, we include the Betulaceae related data as some wind-pollinated plant families may benefit from enhanced dispersal by insects, although the extent of this relationship is unknown45. Some recent studies reveal that some of the insect pollinators, particularly bees and syrphid fly species, collect pollen from a broad range of wind-pollinated plant species46,47,48, and also, some other studies49,50,51,52,53,54,55,56,57 identified insect pollination in plant species which were presumed to be wind-pollinated. Our study may open some new horizons in the field of plant-pollinator interactions, a field with large gaps in knowledge. The present study reveals that the season and altitude affect the role of moths as PTMS greatly. In sub-tropical zone (1000–2000 m), we found highest number of PTMS as well as the diversity of pollen grains in post-monsoon whereas in temperate zone, the maximum diversity of PTMS and pollen grain is reported in pre-monsoon. The number of PTMS, PPMS and the diversity of pollen grains are lowest in the monsoon season. This may be due to the heavy rainfall with tropical rainstorms restricting the moth activities, as the weather parametres like rainfall, daily temperature plays a significant role in moth abundance, richness in a particular habitat58. Overall, we found maximum diversity of PTMS from higher sub-tropical zone (1500–2000 m) (44 PTMS) to lower temperate zone (2000–2500 m) (65 PTMS), a Himalayan zone with high floral diversity59,60,61. The tropical zone of Himalaya is mainly dominated with plant families like Fabaceae, Betulaceae, Euphorbiaceae, Theaceae and Asteraceae61, among which most of the species are trees. Whereas, in sub-tropical and temperate zone the floral component is characterized by plant families like Elaeocarpaceae, Oleaceae, Solanaceae, Rubiaceae, Zingiberaceae, Verbenaceae, Ericaceae, Asteraceae, Rosaceae and Orchidaceae59,60,62,63 with dominance of shrubs and herbs. In our study, higher proportion of pollen from four plant families i.e., Fabaceae, Betulaceae, Ericaceae and Rosaceae is a possible indication of high preferences of settling moths towards these plant families, mainly for nectaring or may be for some non-nectar resource. Some non-lepidopteran pollinators visit wind-pollinated plants to collect or feed on pollen, and for non-floral resources i.e. insect honey dew, plant resins or resinous secretions45,64,65,66,67 but our understanding about the attraction of settling moths towards small flowers with a very low quantity of nectar is very limited and requires further exploration.
We constructed the pollen number based network following Banza et al.68 (with selectivity value of 0.79), and our network matrices also revealed high degree of selectivity (0.86) of settling moths in Himalayan ecosystem, which is highest value as compared to the other studies conducted on nocturnal moths in several ecosystems around the world12,68,69,70, thus, showing the importance of our study conducted in a larger spatial scale. The PDI score revealed that major proportion (98 species; 70%) of PTMS are specialist and only 42 species (30%) are generalists. High degree of specialism in moths is a strong indication of inter-dependency, and there are high possibilities that the dwindling moth diversity will adversely affect the plant diversity, and the other way round also. In our consolidated network, the value of interaction strength asymmetry (0.50) indicates that the specialised species are interacting with the generalised species and the vice versa.
We found significant correlation between the polyphagous nature of Achaea janata (Erebidae: Erebinae) and its ability to transfer pollen of different plant families. A. janata, commonly known as Castor oil semi-looper or Croton caterpillar, is a widely distributed species in the region and more broadly, it is a well-known pest of various economically important plants like Arachis hypogaea (Fabaceae), Rosa chinensis (Rosaceae), Tamarindus indica (Fabaceae), Glycine max (Fabaceae), Vigna mungo (Fabaceae), and Dalbergia sisso (Fabaceae)71. However, our results establish A. janata as a potential pollinator of three plant families, an addition to the knowledge provided by few such studies72 indicating that the moths can provide net benefits as pollinators even when acting as larval herbivores of the same species. The phenomenon revealed that the species-interactions are much more complicated than we think of.
We believe that our results will lay a strong foundation for the studies related to moth-plant interaction, particularly in the Indian context and will further strengthen the concepts of ecosystem conservation rather than concentrating on few taxa. The present dataset is a tip of an iceberg as there are about 12,000 moth species in India73 and about 160,000 in the world74 and their abundance is still unknown to the science. We are quite sure that data generation and accumulation of knowledge through such type of studies will definitely strengthen our understanding about the role of nocturnal insect pollinators and will help in a better way regarding the management decisions for conservation, biodiversity, and agriculture.
Methods
Moth collection and sampling design
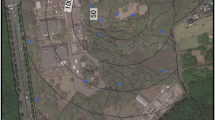
Representatives of settling moth families were collected from September, 2018 to October, 2019 in three different seasons i.e., pre-monsoon (May–June), monsoon (July–September) and post-monsoon season (October–November), using vertical sheet light traps operating with 160-W mercury vapour lamps from 24 randomly chosen sampling sites of Arunachal Pradesh, North Bengal and Sikkim (Fig. 7; see Table S4 of supplementary file 1). For collection, a total of 7 surveys were conducted. Each locality was surveyed for two days per season. Light trapping was done from 6PM to 6AM. The moths were collected individually in the jars euthanized with ethyl acetate. A maximum of 2–3 specimens of each species were collected. Moths were processed and preserved as per standard techniques in Lepidopterology75.

Localities sampled in this study (ArcGIS version 10.5, URL: https://www.arcgis.com/index.html).
The three clearly distinct seasons of Himalayan region were sampled as they greatly affect the floral and faunal components of the area under study76. The collections were made from a very large altitudinal gradient covering up to 3000 m and due to which, we interpreted our data in consolidate, as well as seasonally (pre-monsoon, monsoon and post-monsoon) and altitudinally (tropical zone (0–1000 m), sub-tropical zone (1000–2000 m) and temperate zone (2000–3000 m)).
Isolation of moth proboscis and light microscopy of Fuchsin jelly slides
We used proboscides (c.a. 1800) to investigate the pollen loads (not the other parts of the body like head or legs) and also, for the first time, we separated the proboscis of each moth and directly processed it for the study of pollen (rather than swabbing). Our method reduced the chances of contamination due to pollen rain and it also enabled the study of those particular pollen grains which are found within the coiling of the proboscis, which may remain unstudied during swabbing. We isolated the proboscis from each moth using sterilised forceps. Each proboscis was kept in a separate vial with code number. To relax, the proboscis of each moth was placed on a glass slide and treated with few drops of 1:1 ratio of phenol and glycerol solution for 1–2 min. Afterwards, one to two drops of 50% solution of basic Fuchsin dye were added and the slide was mounted with cover slips and sealed with nail varnish. A Nikon 50i microscope with DP–25 digital camera (40 X magnification) was used to photograph the slides and to count the pollen grains.
SEM analysis of isolated proboscis and pollen
The isolated proboscides of some specimens were also scanned with SEM, and photographs of pollen were taken using ZEISS Evo18 image acquiring software v5.09.
Moth and pollen grain identification
Moths were identified using various available literature, publications77,78,79,80,81,82 and websites viz. www.mothsofindia.org83, www.inaturalist.org84. The classification followed here is given by Nieukerken85. Pollen grains were identified from palynological literature, books and websites including: www.paldat.org86, www.globalpollenproject.org87,88,89,90,91.
Pollen transfer network construction and calculation of network matrices
Seven matrices (one consolidated, and two separate matrices (for sub-tropical and temperate zone) for each season) (see supplementary file 2) are prepared by keeping all the settling moth species along the rows and the plant families along the columns. The number of pollen grains from each plant family found on individual moth species is taken as the value for constructing the mutualistic network44. For quantification of all the mutualistic network parametres ‘bipartite’ package of RStudio is used70. The network-level metrics are (1) connectance (reflects the possible recorded links in the network or standardised number of species combinations)92, higher value indicates more connectance, (2) nestedness (range 0–100, higher value reflects more complexity of the network)93, (3) specialization or H2' (range 0–1, higher value indicates more specialist species are interacting against the generalist species)93,94, (4) links per species (quantify links of species present in a mutualistic network), (5) interaction evenness (evenness of interactions of animals in a network based on Shannon’s diversity, higher value reflects more evenness)95, (6) Shannon’s diversity (to calculate the interaction richness of the network, high value indicates higher diversity), (7) interaction strength asymmetry (quantify weather the specialised species are interacting with generalised species in other level or vice versa)96, (8) linkage density (measures the density of linkage in a network)97 and (9) species-level paired difference index (PDI) (measured to show the specialization of individual species for commonly visited plant family)98. The PDI score ranges from 0 to 1, 0 indicate complete generalism and 1 indicate total specialism in the plants’ relationship with moths.
References
Potts, S. G. et al. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 25, 345–353 (2010).
Kearns, C. A., Inouye, D. W. & Waser, N. M. ENDANGERED MUTUALISMS: The conservation of plant-pollinator interactions. Annu. Rev. Ecol. Syst. 29, 83–112 (1998).
Ollerton, J., Winfree, R. & Tarrant, S. How many flowering plants are pollinated by animals?. Oikos 120, 321–326 (2011).
Labandeira, C. C. A paleobiologic perspective on plant–insect interactions. Curr. Opin. Plant Biol. 16, 414–421 (2013).
Faegri, K. & Van Der Pijl, L. Principles of Pollination Ecology. (Elsevier Science, 2014).
Bhutia, J. & Sharma, B. Diversity of Pollinators/ Visitors in Namchi, South Sikkim, India. 487–498 (2020).
Torres-Vanegas, F. et al. Tropical deforestation reduces plant mating quality by shifting the functional composition of pollinator communities. J. Ecol. 109, 1730–1746 (2021).
Macgregor, C. J., Pocock, M. J. O., Fox, R. & Evans, D. M. Pollination by nocturnal Lepidoptera, and the effects of light pollution: A review. Ecol. Entomol. 40, 187–198 (2015).
Macgregor, C. J., Williams, J. H., Bell, J. R. & Thomas, C. D. Moth biomass increases and decreases over 50 years in Britain. Nat. Ecol. Evol. 3, 1645–1649 (2019).
Chamorro, S., Heleno, R., Olesen, J. M., McMullen, C. K. & Traveset, A. Pollination patterns and plant breeding systems in the Galápagos: A review. Ann. Bot. 110, 1489–1501 (2012).
Ramirez, N. Pollination specialization and time of pollination on a tropical Venezuelan plain: Variations in time and space. Bot. J. Linn. Soc. 145, 1–16 (2004).
Walton, R. E., Sayer, C. D., Bennion, H. & Axmacher, J. C. Nocturnal pollinators strongly contribute to pollen transport of wild flowers in an agricultural landscape. Biol. Lett. 16, 20190877 (2020).
Young, H. J. Diurnal and nocturnal pollination of Silene alba (Caryophyllaceae). Am. J. Bot. 89, 433–440 (2002).
Maeda, M., Maguchi, S., Nakamaru, Y., Takagi, D. & Fukuda, S. Prospective study of pollen dispersal prediction and identifying the usefulness of different parameters. Nihon Jibiinkoka Gakkai Kaiho 109, 455–460 (2006).
Bertin, R. I. & Willson, M. F. Effectiveness of diurnal and nocturnal pollination of two milkweeds. Can. J. Bot. 58, 1744–1746 (1980).
Morse, D. H. & Fritz, R. S. Contributions of diurnal and nocturnal insects to the pollination of common milkweed (Asclepias syriaca L.) in a pollen-limited system. Oecologia 60, 190–197 (1983).
Jennersten, O. & Morse, D. H. The quality of pollination by diurnal and nocturnal insects visiting common milkweed Asclepias syriaca. Am. Midl. Nat. 125, 18 (1991).
Miyake, T. & Yahara, T. Why does the flower of Lonicera japonica open at dusk?. Can. J. Bot. 76, 1806–1811 (1998).
Atwater, M. M. Diversity and nectar hosts of flower-settling moths within a Florida sandhill ecosystem. J. Nat. Hist. 47, 2719–2734 (2013).
Grant, V. & Grant, K. A. Hawkmoth pollination of Mirabilis longiflora (Nyctaginaceae). Proc. Natl. Acad. Sci. 80, 1298–1299 (1983).
Willmott, A. P. & Burquez, A. The pollination of Merremia palmeri (Convolvulaceae): Can Hawk moths be trusted?. Am. J. Bot. 83, 1050 (1996).
Wasserthal, L. T. The Pollinators of the Malagasy Star Orchids Angraecum sesquipedale, A. sororium and A. compactum and the Evolution of Extremely Long Spurs by Pollinator Shift. Bot. Acta 110, 343–359 (1997).
Miyake, T., Yamaoka, R. & Yahara, T. Floral scents of hawkmoth-pollinated flowers in Japan. J. Plant Res. 111, 199–205 (1998).
Luyt, R. & Johnson, S. D. Hawkmoth pollination of the African epiphytic orchid Mystacidium venosum, with special reference to flower and pollen longevity. Plant Syst. Evol. 228, 49–62 (2001).
Rust, R. W., Vaissire, B. E. & Westrich, P. Pollinator biodiversity and floral resource use in Ecballium elaterium (Cucurbitaceae), a Mediterranean endemic. Apidologie 34, 29–42 (2003).
Jürgens, A., Witt, T. & Gottsberger, G. Flower scent composition in Dianthus and Saponaria species (Caryophyllaceae) and its relevance for pollination biology and taxonomy. Biochem. Syst. Ecol. 31, 345–357 (2003).
Oliveira, P. E., Gibbs, P. E. & Barbosa, A. A. Moth pollination of woody species in the Cerrados of Central Brazil: A case of so much owed to so few?. Plant Syst. Evol. 245, 41–54 (2004).
Morimoto, Y., Gikungu, M. & Maundu, P. Pollinators of the bottle gourd (Lagenaria siceraria) observed in Kenya. Int. J. Trop. Insect Sci. 24, (2004).
Willmer, P. Pollination and floral ecology. (Princeton University Press, 2011). https://doi.org/10.1515/9781400838943.
Mitchell, T. C., Dötterl, S. & Schaefer, H. Hawk-moth pollination and elaborate petals in Cucurbitaceae: The case of the Caribbean endemic Linnaeosicyos amara. Flora Morphol. Distrib. Funct. Ecol. Plants 216, 50–56 (2015).
Chakraborty, P., Smith, B. & Basu, P. Pollen transport in the dark: Hawkmoths prefer non crop plants to crop plants in an agricultural landscape. Proc. Zool. Soc. 71, 299–303 (2018).
Proctor, M., Yeo, P. & Lack, A. The natural history of pollination. (Timber Press, 1996).
Funamoto, D. & Sugiura, S. Settling moths as potential pollinators of Uncaria rhynchophylla (Rubiaceae). Eur. J. Entomol. 113, 497–501 (2016).
Funamoto, D. & Sugiura, S. Relative importance of diurnal and nocturnal pollinators for reproduction in the early spring flowering shrub Stachyurus praecox (Stachyuraceae). Plant Species Biol. 36, 94–101 (2021).
Buxton, M. N., Anderson, B. J. & Lord, J. M. The secret service—analysis of the available knowledge on moths as pollinators in New Zealand / Te pepe huna—he tātarihaka o te mātauraka rakahau ki kā pepe hai whakaaiai ki Aotearoa me Te Waipounamu. N. Z. J. Ecol. 42, 1–9 (2018).
Hahn, M. & Brühl, C. A. The secret pollinators: An overview of moth pollination with a focus on Europe and North America. Arthropod-Plant Interact. 10, 21–28 (2016).
Makholela, T. & Manning, J. C. First report of moth pollination in Struthiola ciliata (Thymelaeaceae) in southern Africa. South Afr. J. Bot. 72, 597–603 (2006).
Okamoto, T., Kawakita, A. & Kato, M. Floral adaptations to nocturnal moth pollination in Diplomorpha (Thymelaeaceae). Plant Species Biol. 23, 192–201 (2008).
Paul, M. Impact of urbanization on moth (Insecta: Lepidoptera: Heterocera) diversity across different urban landscapes of Delhi India. Acta Ecol. Sin. 41, 204–209 (2021).
Subhakar, G. & Sreedevi, K. Nocturnal insect pollinator diversity in bottle gourd and ridge gourd in southern Andhra Pradesh. Curr. Biot. 9, 137–144 (2015).
Chakraborty, P., Chatterjee, S., Smith, B. M. & Basu, P. Seasonal dynamics of plant pollinator networks in agricultural landscapes: How important is connector species identity in the network?. Oecologia 196, 825–837 (2021).
Chakraborty, P., Mukherjee, P. A., Laha, S. & Gupta, S. K. The influence of floral traits on insect foraging behaviour on medicinal plants in an urban garden of eastern India. J. Trop. Ecol. 37, 200–207 (2021).
King, C., Ballantyne, G. & Willmer, P. G. Why flower visitation is a poor proxy for pollination: Measuring single-visit pollen deposition, with implications for pollination networks and conservation. Methods Ecol. Evol. 4, 811–818 (2013).
Devoto, M., Bailey, S., Craze, P. & Memmott, J. Understanding and planning ecological restoration of plant–pollinator networks. Ecol. Lett. 15, 319–328 (2012).
Saunders, M. E. Insect pollinators collect pollen from wind-pollinated plants: Implications for pollination ecology and sustainable agriculture. Insect Conserv. Divers. 11, 13–31 (2018).
Ssymank, A., Kearns, C. A., Pape, T. & Thompson, F. C. Pollinating Flies (Diptera): A major contribution to plant diversity and agricultural production. Biodiversity 9, 86–89 (2008).
Rader, R. et al. Non-bee insects are important contributors to global crop pollination. Proc. Natl. Acad. Sci. 113, 146–151 (2016).
Garibaldi, L. A. et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 339, 1608–1611 (2013).
Gong, Y.-B. et al. Wind or insect pollination? Ambophily in a subtropical gymnosperm Gnetum parvifolium (Gnetales): Ambophily in Gnetum. Plant Species Biol. 31, 272–279 (2016).
Niklas, K. J. A Biophysical Perspective on the Pollination Biology of Ephedra nevadensis and E. trifurca. Bot. Rev. 81, 28–41 (2015).
Kato, M., Inoue, T. & Nagamitsu, T. Pollination biology of Gnetum (Gnetaceae) in a LOWLAND MIXED DIPTEROCARP forest in Sarawak. Am. J. Bot. 82, 862–868 (1995).
Celedón-Neghme, C., Santamaría, L. & González-Teuber, M. The role of pollination drops in animal pollination in the Mediterranean gymnosperm Ephedra fragilis (Gnetales). Plant Ecol. 217, 1545–1552 (2016).
Costa, A. C. G. & Machado, I. C. Flowering dynamics and pollination system of the sedge Rhynchospora ciliata (Vahl) Kükenth (Cyperaceae): does ambophily enhance its reproductive success?: Ambophily in Rhynchospora ciliata. Plant Biol. 14, 881–887 (2012).
Huang, L. et al. Beta diversity partitioning and drivers of variations in fish assemblages in a headwater stream: Lijiang River China. Water 11, 680 (2019).
Schneider, D., Wink, M., Sporer, F. & Lounibos, P. Cycads: their evolution, toxins, herbivores and insect pollinators. Naturwissenschaften 89, 281–294 (2002).
Wilson, G. W. Insect Pollination in the Cycad Genus Bowenia Hook, ex Hook. f. (Stangeriaceae)1. Biotropica 34, 438–441 (2002).
Terry, L. I. et al. Pollination of Australian Macrozamia cycads (Zamiaceae): effectiveness and behavior of specialist vectors in a dependent mutualism. Am. J. Bot. 92, 931–940 (2005).
Intachat, J., Holloway, J. D. & Staines, H. Effects of weather and phenology on the abundance and diversity of geometroid moths in a natural Malaysian tropical rain forest. J. Trop. Ecol. 17, 411–429 (2001).
Shaheen, H., Ullah, Z., Khan, S. M. & Harper, D. M. Species composition and community structure of western Himalayan moist temperate forests in Kashmir. For. Ecol. Manag. 278, 138–145 (2012).
Shaheen, H., Mallik, N. M. & Dar, M. E. U. I. Species composition and community structure of subtropical forest stands in western himalayan foothills of kashmir. Pak. J. Bot. 47, 2151–2160 (2015).
Bhutia, Y., Gudasalamani, R., Ganesan, R. & Saha, S. Assessing forest structure and composition along the altitudinal gradient in the State of Sikkim, Eastern Himalayas India. Forests 10, 633 (2019).
Dar, J. A. & Sundarapandian, S. Variation of biomass and carbon pools with forest type in temperate forests of Kashmir Himalaya India. Environ. Monit. Assess. 187, 55 (2015).
Kandel, P. et al. Plant diversity of the Kangchenjunga Landscape, Eastern Himalayas. Plant Divers. 41, 153–165 (2019).
Leonhardt, S. D. & Blüthgen, N. A sticky affair: Resin collection by bornean stingless bees: resin collection by stingless bees. Biotropica 41, 730–736 (2009).
Nyeko, P., Edwards-Jones, G. & Day, R. K. Honeybee, Apis mellifera (Hymenoptera: Apidae), leaf damage on Alnus species in Uganda: A blessing or curse in agroforestry?. Bull. Entomol. Res. 92, 405–412 (2002).
Koch, H., Corcoran, C. & Jonker, M. Honeydew collecting in malagasy stingless bees (Hymenoptera: Apidae: Meliponini) and observations on competition with invasive ants. Afr. Entomol. 19, 36–41 (2011).
Santas, L. A. Insects producing honeydew exploited by bees in Greece. Apidologie 14, 93–103 (1983).
Banza, P., Belo, A. D. F. & Evans, D. M. The structure and robustness of nocturnal Lepidopteran pollen-transfer networks in a Biodiversity Hotspot. Insect Conserv. Divers. 8, 538–546 (2015).
Walton, R. E., Sayer, C. D., Bennion, H. & Axmacher, J. C. Improving the pollinator pantry: Restoration and management of open farmland ponds enhances the complexity of plant-pollinator networks. Agric. Ecosyst. Environ. 320, 107611 (2021).
Dormann, C. F. et al. bipartite: Visualising Bipartite Networks and Calculating Some (Ecological) Indices. (2021).
Karmawati, E. & Tobing, S. L. Laboratory biology of Achaea janata L. castor large semi-loopers. Ind. Crops Res. J. 1, 37–42 (1988).
Labouche, A. & Bernasconi, G. Cost limitation through constrained oviposition site in a plant-pollinator/seed predator mutualism. Funct. Ecol. 27, 509–521 (2013).
Ramakrishna & Alfred, J. R. B. Faunal resources of India. (Zoological Survey of India, 2007).
Lees, D. C. & Zilli, A. Moths: Their Biology, Diversity and Evolution | NHBS Field Guides & Natural History. (London Natural History Museum, 2020).
Holloway, J. D. Moths of Borneo. (Malayan Nature Journal, 2001).
Plant diversity in the Himalaya hotspot region: a volume to celebrate the completion of university service of Dr. Abhaya Prasad Das. (Bishen Singh Mahendra Pal Singh, 2018).
Hampson, G. F. The Fauna of British India, including Ceylon and Burma. vol. 1 1–560 (Taylor and Francis, 1892).
Hampson, G. F. The Fauna of British India, including Ceylon and Burma. vol. 2 1–640 (Taylor and Francis, 1894).
Hampson, G. F. The Fauna of British India, including Ceylon and Burma. vol. 3 1–582 (Taylor and Francis, 1895).
Hampson, G. F. The Fauna of British India, including Ceylon and Burma. vol. 4 1–632 (Taylor and Francis, 1896).
Kirti, J. S. & Singh, N. Arctiid moths of India. (Nature Books India, 2015).
Kirti, J. S. & Singh, N. Arctiid moths of India. vol. 2 (Nature Books India, 2016).
Moths of India. https://www.mothsofindia.org/.
iNaturalist. iNaturalist. iNaturalist https://www.inaturalist.org/users/sign_in.
Nieukerken, E. J. V. et al. Order Lepidoptera Linnaeus, 1758. In : Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa 3148, 212–221 (2011).
PalDat. https://www.paldat.org/.
Global Pollen Project. Global Pollen Project. https://globalpollenproject.org/.
Agashe, S. N. & Caulton, E. Pollen and spores: applications with special emphasis on aerobiology and allergy. (Science Publishers, 2009).
Bhattacharya, K. et al. A textbook of palynology. (2014).
Stephen, A. Pollen—A microscopic wonder of plant kingdom. Int. J. Adv. Res. Biol. Sci. 1, 45–62 (2014).
Halbritter, H. et al. Illustrated Pollen Terminology. (Springer International Publishing, 2018). doi:https://doi.org/10.1007/978-3-319-71365-6.
Dunne, J. A., Williams, R. J. & Martinez, N. D. Food-web structure and network theory: The role of connectance and size. Proc. Natl. Acad. Sci. 99, 12917–12922 (2002).
Rodriguez-Girones, M. A. & Santamaria, L. A new algorithm to calculate the nestedness temperature of presence-absence matrices. J. Biogeogr. 33, 924–935 (2006).
Blüthgen, N., Menzel, F. & Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 6, 9 (2006).
Tylianakis, J. M., Tscharntke, T. & Lewis, O. T. Habitat modification alters the structure of tropical host–parasitoid food webs. Nature 445, 202–205 (2007).
Blüthgen, N., Menzel, F., Hovestadt, T., Fiala, B. & Blüthgen, N. Specialization, constraints, and conflicting interests in mutualistic networks. Curr. Biol. 17, 341–346 (2007).
Bersier, L.-F., Banašek-Richter, C. & Cattin, M.-F. Quantitative descriptors of food-web matrices. Ecology 83, 2394–2407 (2002).
Poisot, T., Lepennetier, G., Martinez, E., Ramsayer, J. & Hochberg, M. E. Resource availability affects the structure of a natural bacteria–bacteriophage community. Biol. Lett. 7, 201–204 (2011).
Acknowledgements
We are thankful to Dr. Dhriti Banerjee, Director and Dr. Kailash Chandra, former Director, Zoological Survey of India for permission and facilities to carry out this research work. The present research work is funded by the Ministry of Environment, Forest and Climate Change, Govt. of India through the G.B. Pant Institute of Himalayan Environment and Sustainable Development, Almora under NMHS (National Mission on Himalayan Studies) programme. We thank PCCF (Principal Chief Conservator of Forests) of the states of Sikkim, Arunachal Pradesh and West Bengal for the permission to undertake field work for data collection. We thank Dr. Callum J. Macgregor (University of Hull) for his critical comments on the original manuscript.
Funding
This research work is funded by Ministry of Environment, Forest and Climate Change through the G.B. Pant Institute of Himalayan Environment and Sustainable Development under National Mission on Himalayan Studies (NMHS). (Grant ID: NMHS/2017-18/MG41/25).
Author information
Authors and Affiliations
Contributions
All authors contributed to interpretation of results and writing the manuscript. R.L., P.C. and D.M. conducted the field surveys for data collection. The research and analysis are performed by N.S. and R.L.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Singh, N., Lenka, R., Chatterjee, P. et al. Settling moths are the vital component of pollination in Himalayan ecosystem of North-East India, pollen transfer network approach revealed. Sci Rep 12, 2716 (2022). https://doi.org/10.1038/s41598-022-06635-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-06635-4
- Springer Nature Limited