
Abstract
Fungi are responsible for many plant infections. Indeed, fungi are responsible for most plant spoilage that occurs in the field or in storage, causing symptoms of dieback, leaf spot, rot, rust and wilt. A more serious issue is that fungi produce mycotoxins, which can contaminate crops. Samples of leaf, fruit and root tissues were collected from sick crops, pre- and post-harvest within Al-Baha region. Using molecular markers, twenty fungal isolates were identified that represent eight genera and thirteen species. The most common species were Penicillium spp. (45%), followed by Aspergillus spp. (25%). The remaining 30% were comprised of multiple species (Alternaria alternata, Fusarium oxysporum, Geotrichum candidum, Lasiodiplodia theobromae, Megasporoporia minor and Rhizopus arrhizu). The majority of these isolates are recognised as spoilers of crops and producers of mycotoxins. The study evaluated which temperatures optimised fungal growth. Half of the isolates thrived best at 25 °C; six isolates favoured 30 °C, whilst four isolates grew best at 20 °C. The study also evaluated the optimal water activity (aw) for fungal growth, at four points (0.995, 0.99, 0.90 and 0.85 aw). The isolates were categorised as being either xerophilic, mesophilic or hydrophilic. The majority of isolates (70%) were xerophilic (Aspergillus and Penicillium spp.), 10% were mesophilic (Alternaria alternata and Rhizopus oryzae spp.) and 20% were hydrophilic (Lasiodiplodia theobromae, Geotrichum candidum, Megasporoporia minor and Fusarium oxysporum spp). The information derived from this study will assist in building effective strategies to control these pathogens, minimising crops losses and mycotoxin contamination.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
Much of the Kingdom of Saudi Arabia (KSA) is desert; however, there are areas of abundant and diverse vegetation [42]. According to Collenette [22] and Alzandi et al. [15], 142 plant families, representing approximately 2300 species, are found in KSA. The climate and topography of Al-Baha province make the province an important agricultural region. Located in the southwestern part of the country, Al-Baha has six municipalities: Al-Aqiq, Al-Baha (the capital), Al-Mandaq et al. [14]. The province’s varied landscape, which includes mountains, valleys and several types of forest, supports diverse habitats for flora and fauna. Around 230 species of grain, fruit, vegetables and plants with medicinal uses, from 75 families and 228 genera are found in the province [13]. The province has a long history of farming and several native techniques have been devised to support both cultivated and wild plants [22]. One of KSA’s key crops is dates. The nation’s 28 million date palms trees annually provide ~ 14% of the world’s supply of dates. However, studies conducted by El-Habbab et al. [25] and Saleh et al. [54] indicate that fungal infections ruin many of the dates grown in KSA,fungi contaminate or spoil 2.7–33% of the dates grown in KSA. Fusarium spp. have been identified as the cause of mycotoxin contamination in some Saudi date crops [7, 11]. Pencillium, Aspergillus niger, A. flavus and Colletotrichum musae, which are also mycotoxin-synthesising species, have also been identified in crops imported into KSA. [1, 2].
According to Bebber et al. [20], up to 40% of failed crops are caused by phytopathogenic microbes, with fungal infections ruining around half of all maize yields [26]. Concerns have been expressed by the agriculture sector that the virulence of pathogenic microbes could have deleterious effects upon food security. Even though fungi are considered to be the second most common plant pathogen, the level of threat they present to flora is significant. As well as having damaging impacts upon the heath of the host plants, fungi can produce mycotoxins that are harmful to the animals and humans that eat the infected plants. Mycotoxins have long been recognised as being potent carcinogens [39]. The Food and Agriculture Organization estimates that each year, mycotoxins contaminate about half of the world’s crops [26]. Researchers have identified > 400 different mycotoxins, including aflatoxins, ochratoxins, trichothecenes and fumonisins [50]. The amount of harm that mycotoxins cause varies according to the fungi producing them. Aspergillus, Fusarium and Penicillium spp. are three of the five types of fungi regarded to be most harmful in terms of their mycotoxin production, as well adversely affecting crop quality and yields. Whilst fungicides and pesticides offer crops protection against pathogens, they are not benign products. They too are harmful to animals, humans and the insects that are essential to crops and the environment. In light of this, a number of countries have banned the use of fungicides and pesticides. This action replaces one risk with another, and alternative safe and effective methods of controlling plant pathogens, is urgently needed [4, 20, 24]. Canada, Indonesia and Sweden are leading the way, by using plant extracts and biological or environmental means to mitigate phytopathogenic infections in an environmentally friendly manner [45, 49].
The invasiveness of fungi is heavily influenced by temperature and access to water. Consequently, efforts to characterise the settings in which phytopathogens to both thrive and die are warranted. Such information can be applied to modulate conditions in fields and food storage locations, thereby limiting fungal infection-induced crop failures and mycotoxins [8]. Temperature and water activity not only influence fungal growth, sporulation and the synthesis of mycotoxins, they also affect plant immune systems and a plant’s ability to fight fungal infections. There is considerable variation between and within fungal species in the optimal temperature and water activity needed for infecting and colonising hosts, mycelia growth and sporulation [28, 35]. For example, Hope et al. [29] found the optimal growth temperature of two separate strains of Fusarium colmorum sp. isolated from maize grown in different locations diverged by 10 °C. Armed with the knowledge of what temperatures optimise fungal bioactivity, agriculturists may be able to take steps to sustain conditions that do not favour fungi survival [28, 38].
Similarly, water in the environment significantly influences fungal activity and the physical and chemical properties of fungal products. Water activity represents the amount of bound or free water available in the product or in the air, in the field and in storage conditions [19, 52]. Fungi can be categorised according to the aw conditions that promotes maximum activity. As indicated by the name, hydrophilic fungi are moisture lovers, growing maximally in conditions of 0.95 aw or above. Mesophilic fungi thrive in moderate levels of moisture, with an aw of ± 0.90. Fungi that grow best in drier conditions (aw 0.65−0.85) are described as xerophilic [40]. When the water activity is at the level that is optimal for a particular type of fungus, the fungus can germinate and its pathogenic activity increase. This can result in lost or mycotoxin-tainted crops [33, 52].
In the past, taxonomists used to rely on visual evaluations of a specimen’s morphological and biological features to identify and characterise it. However, following technological advances, fungi are now usually identified by analysing its molecular mechanisms; this approach is considered to be more reliable than visual inspection. The genes known to be responsible for specific molecular mechanisms may be determined from the variable bands produced by pulse-field gel electrophoresis. Alternatively, species may be identified using basic local alignment search tool (BLAST) analysis, which compares the gene sequence of the fungal sample against sequences stored in gene repositories, such as National Centre for Biotechnology Information (NCBI) and UNITE [51]. The internal transcribed spacer (ITS) is a section of RNA that is commonly used as a marker to identify species of fungi [61]. By 2012, GenBank held over 175,000 ITS sequences obtained from > 15,000 fungal species [57]. Nonetheless, by 2017, the data in GenBank still represented < 1% of the 5.1 million species believed to exist [51]. The ITS, which is 400–800 bp long, has a highly conserved region that appears to be universal among fungi. Identification of fungi typically involves polymerase chain reaction (PCR) amplification of the ITS1 and ITS4 primers [16, 23]. Mbareche et al. [37] state that there is broad consensus among researchers that to identify a new fungal species from an ITS marker the sequence homology must exceed 96%. Recently, researchers have concentrated their efforts on using ITS sequences to differentiate regional fungal isolates. To establish the phylogenetic origin of the fungal isolate, its ITS sequences are compared against reference sequences.
2 Materials and methods
2.1 Sampling procedure
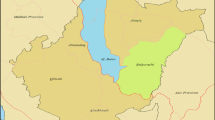
Infected root and leaf tissues were collected from twenty different pre- and post-harvested crop sites in Al-Baha province (Fig. 1). Before gathering multiple samples of the plants from each site, permission was sought from the owner. By collecting multiple samples, the prevalence and distribution of the primary fungal pathogen could be established. It also limited the potential of collecting extraneous specimens present on the soil or plant surface and wind-borne isolates.

Map showing the six principal municipalities of Al Baha province. The 20 fungal specimens collected for this study were obtained from sites et al. Baha province
Al-Baha city is sited 2400 m above sea at latitude 20°0′46″ N and longitude 41°28′4″ E [14]. Baljurashi city (19°51′34″ N, 41°33′26″ E,) is of similar altitude (2000 m), whilst Al-Mikhwah (19°46′46″ N, 41°26′8″ E) is lower to sea level (elevation 447 m) [6]. Al Aqiq is 600 m above sea level located at 20°17′41″ N, 41°38′35″ E, while Qilwah (elevation 260 m) is at located at 19°56′42.39″ N, 41°14′44.56″ E. Finally, Al Mandaq is located 1993 m above sea level at 20°13′12.3″ N, 41°15′2.64″ E.
2.2 Isolation of fungi and pure culture preparation
Fungi were removed from the surface of the plants using sterile methods. Potato dextrose agar (PDA) medium was used to culture and subculture the specimens. All of the samples obtained from a single site were reviewed then a single representative sample was selected. The representative sample was deemed to be the primary phytopathogen responsible for plant disease at that particular sampling location [1]. Consequently, twenty isolates were used in this study. Stock was cultured then stored in the stock culture bank et al.-Baha University (Table 1).
2.3 DNA extraction
Mycelia were sub-cultured in potato dextrose broth medium to create sufficient fungal material from which DNA could be harvested. Mycelial tissue was frozen with liquid nitrogen then powdered using a mortar and pestle. DNA was extracted from 150 to 200 mg of powder that had been transferred into a 2-ml Eppendorf tube. DNA was extracted using a Thermo Scientific GeneJET Plant Genomic DNA Purification kit in accordance with the manufacturer’s instructions. The extracted DNA was stored at − 20 °C until required [10].
2.4 Molecular identification and phylogenetic tree
The forward primer ITS1 (TCCGTAGGTGAACCTGCGG) and the reverse primer ITS4 (TCCTCCGCTTATTGATATGC) were used in 20-µl and 50-µl PCR amplification of the region between 18 and 28S of the small and large subunits of ribosomal ribonucleic acid (rRNA) [61]. The following PCR protocol was followed: 35 × cycles of initial denaturation for 3 min at 95 °C, followed by further denaturation for 1 min at 94 °C,annealing for 1 min at 60 °C; extension for 1 min at 72 °C and final extension for 5 min at 72 °C. The amplified DNA was stored at 16 °C. The ingredients for the 50-µl PCR reaction were: 2.5-µl DNA, 2.5 µl of 20-µM forward primer, 2.5 µl of 20-µM reverse primer, 5-µl 10 × reaction buffer, 1 µl of dNTPs, 3 µl of MgCl2 and 0.25-µl Taq DNA polymerase. To reach the 50-µl volume, the reaction ingredients were topped up with DNAase/RNAase-free water [6]. For the 20-µl reaction, lesser quantities of the same ingredients were used. The negative control samples were made up using the same quantities of PCR constituents, but the DNA was replaced with DNAase/RNAase-free water [8]. Loading dye was prepared and mixed into the PCR products. To create five aliquots of loading dye, 1 mg of agarose power was added to 100 ml of 1X tris–acetate-EDTA (TAE) buffer; the solution was cooked in a microwave oven for 2 min. To this mixture, 5 µl of 10-mg/ml ethidium bromide solution was added. The solution was poured onto a gel tray and left to set. The tray was inserted into a gel electrophoresis tank containing 1 × TAE buffer. The reaction was left to run for 1 h at 90 V [54, 63]. To purify the amplicon products, a Qiagen QIAquick PCR Purification kit was used in the manner instructed by the manufacturer. The purified products were sent to Macrogen Inc. to be sequenced. The sequencing output was provided as trace data. Sequence dimensions and standards were verified, and then ambiguous bases were eliminated. The trace data were converted into the FASTA format using BioEdit software. To facilitate identification of the fungal species, data were uploaded to NCBI or UNITE for BLAST analysis [9, 51]. A sequence homology threshold of ≥ 96% was required to perform a phylogenetic search in the gene bank. A phylogenetic tree was constructed using multiple aligned sequences obtained from the initial and benchmark sequences,creation of the tree was achieved using Geneious Prime software, version 2022.2 (Biomatters) (Fig. 3). The program’s tree builder algorithm used the distance and neighbour-joining methodology to build the tree, information and the software’s multiple sequence comparison by log- expectation (MUSCLE) alignment tool aligned multiple sequences. To calculate the genetic distances of the branches, the Tamura-Nei model was used. A bootstrap supporting value (BSV) between 70 and 100% was created to get a consensus phylogenetic tree. A general time reversible evolutionary model with 500 bootstrap replications was applied to find the best fit for the consensus phylogenetic tree [10, 12, 31]. Each of the isolates’ sequences was uploaded to the NCBI gene bank. Table 1 shows the unique accession identifier allocated to each isolate. After determining the quality and quantity of each sequence, individual sequences were filed with NCBI. Specimens that met that sequence threshold were downloaded and used as reference sequences (Table 2) [6].
2.5 Growth assessment at three different temperatures
To determine the temperature that optimised fungal growth, the isolates were exposed to three different temperatures (20 °C, 25 °C and 30 °C). Isolates were cultured and sub-cultured on 9 cm PDA Petri dishes. Sub-culturing was achieved by taking 5-mm discs of inoculum from the edge of the primary culture and transplanting them to the middle of clean Petri dishes. Five replicates were made for each isolate for each temperature; these were incubated for 6 days. At the end of the sixth day, radial growth measurements were taken for each sample and growth rates were calculated (mm/per day) (Fig. 4). The error bars indicate the variation in growth for the different isolates [28, 38].
2.6 Evaluating growth at four points of water activity
To evaluate water activity, four mixtures of PDA medium and glycerol (Sigma Aldrich G9012), were prepared. To achieve four different aw ratios (0.995, 0.99, 0.90 and 0.85) (Fig. 5), a 1-L quantity of PDA and a variable quantity of glycerol was used as follows: no glycerol for 0.99 aw (because the PDA is adjusted by the manufacturer to 0.99 aw); 22.08 ml for 0.995 aw; 404.8 ml for 0.90; 625.6 ml for 0.85 aw. An AQUALAB water activity metre was used to confirm the aw of the adjusted media. To assess the effect of temperature, five replicates of isolates were prepared for each water activity point, which were then incubated at the previously established optimal temperature for 6 days. Radial growth dimensions were assessed as described above [9, 36].
3 Results
3.1 Molecular and phylogenetic characterisation
The ITS gene marker was used to verify that samples were indeed fungal isolates. This was achieved by evaluating the electrophoresis gel bands of the PCR products (Fig. 2). To identify the species of each isolate, each of their amplicon sequences were uploaded to GenBank NCBI for BLAST analysis (Table 1).

Gel electrophoresis showing PCR products that confirm isolates’ fungal identity. The amplicons are approximately 600–750 bp based DNA ladder
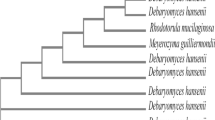
The ITS genetic markers were used to establish the genotypes of seventeen reference strains and the twenty isolates (n = 37). A phylogenetic tree was created using a neighbour-joining tree builder algorithm. The tree (Fig. 3) shows the hierarchical distribution of the 37 isolates, which includes two nodes, three clades, eight major groups, four minor groups and five sub-groups.

The Phylogenetic tree built using ITS genetic markers and Neighbour-joining consensus tree-based method. The isolates code been used for the 20 de-novo isolates, the NCBI accession number been used for the reference sequences. The species name abbreviation was in bracket
Node 1 represents three monophyletic groups (1.1, 1.2 and 1.3); each group is comprised of two taxa. The taxa of group 1.1 represent Geotrichum candidum species. The taxa of group 1.2 belong to Rhizopus oryzae, while those of group 1.3 represent Megasporoporia minor species.
Node two holds 31 of the 37 taxa. This are separated into three clades (2.1, 2.2 and 2.3), with BSV affinities of 89–100%. The largest clade is 2.1, which encompasses 25 taxa distributed between three major groups (A, B and C), with BSV of 97–100%.
Group A is composed of one isolate of Penicillium expansum species. Meanwhile, group B is representing fourteen taxa, which are divided into the minor groups of B1 and B2; these are further divided into three sub-groups (B1.1, B2.1 and B2.2). Minor group B1 is comprised of one polyphyletic group of two taxa belonging to Penicillium glabrum and one monophyletic sub-group B1.1; the latter represents three further Penicillium glabrum species. The two sub-groups of minor group B2 represent Penicillium brevicompactum species (sub-group B2.1), Penicillium polonicum and Penicillium expansum species (sub-group B2.2). The BSV affinity values range from 62 to 98%.
Major group C, consists of ten taxa representing the Aspergillus genus. This is divided into minor groups C1 and C2; the BSV affinity was 63–100%. Minor group C1, is comprised of eight taxa, of which two represent Aspergillus awamori, while the remaining six taxa belong to Aspergillus niger species. Minor group C2 has two taxa that represent Aspergillus terreus species. Clade 2.2 has two taxa that reflect Alternaria alternata species. Meanwhile, the two groups (D and E of clade 2.3) have a BSV affinity of 89–100%. The two taxa of group D belong to Fusarium oxysporum species, and the two taxa of E group represent Lasiodiplodia theobromae species.
3.2 The effect of temperature upon growth
The radial growth data of the twenty isolates, representing thirteen species and eight genera, at different temperatures are presented in Fig. 4 and Table 3.

This bar chart reflects the growth rate at 20 °C, 25 °C and 30 °C. The error bar indicates the extent of variation among the five replicates. These new 20 isolates represent different species, host and geographical locations with in Al Baha province
Isolates were categorised according to the ambient temperature at which optimal growth occurred. The first group were isolates that grew best 20 °C. There were four species in this group: Penicillium expansum (BHU125), Alternaria alternata (BHU002), Megasporoporia minor species (BHU005) and Penicillium polonicum species (BHU069). There were ten isolates in the second group, which grew optimally at 25 °C. These were Penicillium expansum (BHU123 and BHU126), Lasiodiplodia theobroma (BHU101), Rhizopus arrhizus (BHU100), Aspergillus terreus (BHU025), Aspergillus awamori (BHU031), Aspergillus niger (BHU012) and Penicillium glabrum (BHU068, BHU137 and BHU132). The final group had six isolates. These were fungi that grew best at 30 °C. They were Geotrichum candidum (BHU162), Fusarium oxysporum (BHU164), Penicillium expansum (BHU070), Penicillium brevicompactum (BHU072) and Aspergillus niger (BHU102).
3.3 The effect of water activity upon growth
The radial growth data reveal the optimal growth condition for the twenty isolates at four different points of water activity (0.995, 0.99, 0.90 and 0.85 aw) are presented in Fig. 5 and Table 3.

This bar chart reflects the growth rate at four ratios of water activity (0.995, 0.99, 0.90 and 0.85 aw). The error bar reflects the scope of variability among the five replicates. These new 20 isolates represent different species, host and geographical locations with in Al Baha province
The optimal growth condition of the four Penicillium expansum spp. isolates (BHU123, BHU125, BHU126 and BHU070) occurred at 0.85 aw. This ratio of water y activity was also preferred by three isolates of Penicillium glabrum species (BHU068, BHU137 and BHU132), one isolate of Penicillium polonicum species (BHU069), one isolate of Penicillium brevicompactum species (BHU072), three Aspergillus niger species (BHU017, BHU102 and BHU012), one isolate of Aspergillus terreus species (BHU025) and one Aspergillus awamori species (BHU031). Two isolates (Alternaria alternata sp. (BHU002) and Rhizopus arrhizus sp. (BHU100)), preferred 0.90 aw. Meanwhile, 0.995 aw was the optimal ratio for Geotrichum candidum sp. (BHU162), Fusarium oxysporum sp. (BHU164), Megasporoporia minor sp. (BHU005) and Lasiodiplodia theobromae sp. (BHU101) (Fig. 5).
4 Discussion
4.1 Limitations of characterisation using ITS markers
ITS genetic markers were used to identify the eight genera and thirteen species and to establish their phylogenetic relationships. Although ITS markers are reliable for differentiating between specimens at the interspecies level (e.g. 1.1, 1.2, 1.3, 2.2, B1, C2, D and E), the technique is less effective at distinguishing at the intraspecies level. For example, all the isolates in sub-group B2.2 belong to Penicillium spp., yet the BSVs indicate reduced genetic heterogeneity.
Several researchers have reported upon the intraspecies limitation of the ITS markers identification method being particularly pronounced with Fusarium spp. and Penicillium spp. [41, 43, 44, 58]. It is noteworthy that the limitation does not apply equally to species of fungi. Some species of fungi (e.g. Alternia spp. and Aspergillus spp.) can be differentiated at the inter- and intraspecies level using the ITS marker method [5, 9, 17, 27, 62].
4.2 Optimal growth temperatures
Pietikäinen et al. [48] and Ma et al. [34] state that the prevailing temperature in a geographical location greatly influences the optimal temperature at which local fungi thrive. In this study, maximum growth of ten of the isolates occurred at the median temperature (25 °C) and six isolates grew best at 30 °C. Meanwhile the remaining four isolates favoured the relative coolness of 20 °C. That the majority of the isolates grew optimally at the median and high temperatures is not surprising, as they are consistent with the temperatures prevalent in the Al-Baha region. Whilst the majority of strains prefer temperatures in the 20–30 °C range, some species thrive in temperatures as low as 15 °C and others as high as 42 °C.
This observation is also consistent with other research into optimal growth temperatures for European isolates. These studies report that cold-to-moderate temperatures produce maximum growth. For example, various Fusarium species isolates from different parts of Europe grow maximally in temperatures of 15–25 °C [29, 55]. Meanwhile, the optimal temperature for A. alternata species and P. camemberti and P. roqueforti is reported to be 25 °C [18, 30]. On the other hand, Aspergillus species, A. flavus and A. niger, thrive in temperatures of 30 °C and 35 °C [5, 27]. Inevitably, fungi in Saudi Arabia have been habituated to thriving in the Kingdom’s moderate to high temperatures (25–30 °C).
4.3 Optimal water activity
The growth rates of isolates revealed increased variation between isolates from different genera and greater homology between isolates at the species level.
The majority of fungal strains appear to prefer water activity ratios of 0.85–0.99 aw. However, growth is significantly limited in some hydrophilic species when the water activity ratio is 0.85 aw. Four hydrophilic species were found this study Lasiodiplodia theobromae, Geotrichum candidum, Megasporoporia minor and Fusarium oxysporum, which grew best at 0.995 aw. While Alternaria alternata and Rhizopus oryzae species are categorised as being mesophilic, as their maximum growth was achieved at a water activity ratio of 0.90 aw. Fourteen of the isolates, which were members of Aspergillus and Penicillium spp., grew at all of the levels of water activity evaluated, but maximum growth occurred at 0.85 aw, defining these species as xerophilic. The findings presented here are consistent with those of other published studies. Almiman [8], Akbar and Alam [3], Verheecke-Vaessen et al. [60], Koňuchová and Valík [32] and Phan et al. [47] describe species such as Trichoderma, Neofusicoccum, Mucor, Lasiodiplodia theobromae, Geotrichum candidum, Megasporoporia minor and Fusarium to be hydrophilic, because their optimal growth occurred at 0.99 aw. Meanwhile, Yoder et al. [64] and Romero Donato et al. [53] categorised Alternaria alternata and Rhizopus oryzae species, as mesophilic because their favoured water activity ratio is 0.90 aw. As stated above, the majority of the Penicillium and Aspergillus isolates grew at all levels of water activity tested. They grew at low levels of moisture ± 0.85 aw. Therefore, these genera fall between xerophilic and mesophilic. These findings are consistent with those of Sautour et al. [56] and Park et al. [46], who categorised Penicillium and Aspergillus species as xerophilic.
Growth is the bedrock that influences sporulation and the synthesis of mycotoxins, which in turn correlates with fungal pathogenic activity such as dispersal, colonisation and contamination [9]. Some of the species isolated in this study were recognised to be mycotoxigenic, such as; Alternaria, Aspergillus, Fusarium and Penicillium [3, 9, 21].
The information presented in Table 3 might assist with the development of farming practices and schedules that can mitigate fungal infections of crops, thereby minimising crop loss and contamination [8, 12].
5 Conclusion and future perspectives
Phytopathogenic fungi pose a significant threat to food security, with many crops being lost each year to fungal diseases. Tackling fungal infections is challenging, as many fungicides are hazardous to animals, humans and the wider environment. One of the issues to treating fungal infections is the considerable diversity of fungal species and the optimal conditions for their growth. This study identified the optimal ambient temperature and water activity for the growth of phytopathogenic and mycotoxigenic fungal isolates recovered from various fruit and vegetable crops grown in Al-Baha province.
Based upon an analysis of the ITS markers and GenBank data for the new twenty isolates, indicated that the major fungal candidates for vegetable and fruits infestation were Penicillium and Aspergillus spp at 45% and 25% respectively. Whereas the remaining 30% of isolates were solitary representatives of Alternaria alternata, Fusarium oxysporum, Geotrichum candidum, Lasiodiplodia theobromae, Megasporoporia minor and Rhizopus arrhizus species. Some of isolated species in this study were recognised as mycotoxigenic species such as; Alternaria, Aspergillus, Fusarium and Penicillium [3, 11, 21].
It is noteworthy that sixteen of the twenty isolates grew best at moderate (25 °C) and high (30 °C) temperatures. The remaining four isolates grew optimally at 20 °C, this could regard to the nature of biological strains variabilities (van den [59]). Water activity has an important impact upon growth. The isolates were divided in three groups according to their preferential water activity ratio. The first group contained 14 belonging to Aspergillus and Penicillium spp. These species preferred low levels of moisture 0.85 aw, thus were categorised as xerophilic. However, they did grow moderately well at the other water activity ratios. Two of twenty isolates belong to Alternaria alternata and Rhizopus oryzae, were mesophilic, preferring a moderate level of moisture at 0.90 aw. While the remaining four isolates, representing Lasiodiplodia theobromae, Geotrichum candidum, Megasporoporia minor and Fusarium oxysporum species were hydrophilic, growing optimally in high moisture levels 0.995 aw.
These information’s will initiate an effective strategy to control their expansion and reduce their adverse impacts toward agriculture, economics and consumers. Future research that explores the conditions in which mycotoxigenic species optimally synthesise mycotoxins is warranted. Factors that such studies could consider include the effect that pH, temperature and water activity have upon the virulence of phytopathogenic fungi. Other potential research could focus upon the various types of mycotoxins produced and establish whether there is a level of their consumption that is deemed safe. Such studies could be conducted alongside research into the potential of using plant extracts and biological controls in place of fungicides.
Data availability statement
The molecular datasets for this work have been deposited in NCBI gene bank and their corresponding accession numbers are available in tables of this work. Also, the datasets for the optimal growth temperatures are available and attached in this work.
References
Abd-Elsalam KA, Roshdy S, Amin OE, Rabani M (2010) First morphogenetic identification of the fungal pathogen Colletotrichum musae (Phyllachoraceae) from imported bananas in Saudi Arabia. Genet Mol Res 9:2335–2342
Ahmed RA, Mohammed SA (2014) Isolation and classification of fungi associated with spoilage of post-harvest mango (Mangifera indica L.) in Saudi Arabia. Afr J Microbiol Res 8:685–688
Akbar F, Alam S (2019) Influence of water activity and time duration on fusarium mycotoxins production in maize grains during post-harvest storage. Sarhad J Agric 35:1326–1335
Albogami A, Naguib DM (2024) Agricultural wastes: a new promising source for phenylalanine ammonia-lyase as anticancer agent. 3 Biotech 14:22
Alborch L, Bragulat MR, Abarca ML, Cabañes FJ (2011) Effect of water activity, temperature and incubation time on growth and ochratoxin A production by Aspergillus niger and Aspergillus carbonarius on maize kernels. Int J Food Microbiol 147:53–57
Al-Ghamdi SA (2021) Analysis of wind power and wind power characteristics: Al-Aqiq City, Saudi Arabia. Int J Adv Res Eng Technol 12:870–887
Almiman B (2023) Glimpse into phytopathogenic fungal species in Al Baha Province, Saudi Arabia; identification from molecular and morphological characteristics. J Taibah Univ Sci 17:2164458
Almiman B (2023b) Effects of temperature and water activity on 25 de novo strains of pathogenic plant fungi in Al-Baha and Baljurashi cities in Saudi Arabia. J Umm Al-Qura Univ Appl Sci 1–12
Almiman BF (2018) Molecular genetic and genomic characterization of an emerging mycotoxigenic pathogen Fusarium proliferatum. Doctoral dissertation, University of Bedfordshire UK
Almiman BF (2023) Identifying phytopathogenic fungi in Albaha province, Saudi Arabia through their molecular and morphological features: an overview. Saudi J Biol Sci 30:103572
Almiman BF, Shittu TA, Muthumeenakshi S, Baroncelli R, Sreenivasaprasad S (2018) Genome sequence of the mycotoxigenic crop pathogen Fusarium proliferatum strain ITEM 2341 from date palm. Microbiol Resour Announc 7:964
Al-Mutarrafi M, Elsharawy NT, Al-Ayafi A, Almatrafi A, Abdelkader H (2019) Molecular identification of some fungi associated with soft dates (Phoenix dactylifera L.) in Saudi Arabia. Adv Med Plant Res 7:97–106
Al-Namazi AA, Al-Khulaidi AWA, Algarni S, Al-Sagheer NA (2021) Natural plant species inventory of hotspot areas in Arabian Peninsula: Southwest Al-Baha region, Saudi Arabia. Saudi J Biol Sci 28:3309–3324
Alshehri KM (2020) Anticancer plants naturally growing in Al-Baha Region, Saudi Arabia. Int J Pharm Res Allied Sci 9:92–101
Alzandi AA, Taher EA, Al-Sagheer NA, Al-Khulaidi AW, Azizi M, Naguib DM (2021) Phytochemical components, antioxidant and anticancer activity of 18 major medicinal plants in Al Baha region, Saudi Arabia. Biocatal Agric Biotechnol 34:102020
Arif IA, Bakir MA, Khan HA, Al Farhan AH, Al Homaidan AA, Bahkali AH, Al Sadoon M, Shobrak M (2010) A brief review of molecular techniques to assess plant diversity. Int J Mol Sci 11:2079–2096
Armitage AD, Barbara DJ, Harrison RJ, Lane CR, Sreenivasaprasad S, Woodhall JW, Clarkson JP (2015) Discrete lineages within Alternaria alternata species group: identification using new highly variable loci and support from morphological characters. Fungal Biol 119:994–1006
Balai LP, Ahir RR (2013) Role of temperature and relative humidity on mycelial growth of Alternaria alternata infecting brinjal. Trends Biosci 6:307–308
Barbosa-Cánovas GV, Fontana Jr AJ, Schmidt SJ, Labuza TP (2008) Water activity in foods: fundamentals and applications John Wiley & Sons. 1, 15–28
Bebber DP, Ramotowski MAT, Gurr SJ (2013) Crop pests and pathogens move poleward in a warming world. Nat Clim Chang 3:985–988
Bennett J, Klich M (2003) chotoxins. C. lin. Microbiol Rev 16:497–516
Collenette S (1999) Wild Flowers of Saudi Arabia, National Commission for Wildlife Conservation and Development (NCWCD). Riyadh, Saudi Arabia, 703–707
Cowan RS, Fay MF (2012) Challenges in the DNA barcoding of plant material. Plant DNA Fingerpr Barcoding Methods Protoc 862:23–33
Dean R, Van Kan JA, Pretorius ZA, Hammond-Kosack KE, Di Pietro A, Spanu PD, Rudd JJ, Dickman M, Kahmann R, Ellis J (2012) The Top 10 fungal pathogens in molecular plant pathology. Mol Plant Pathol 13:414–430
El-Habbab MS, Al-Mulhim F, Al-Eid S, Abo El-Saad M, Aljassas F, Sallam A, Ghazzawy H (2017) Assessment of post-harvest loss and waste for date palms in the Kingdom of Saudi Arabia. Int J Environ Agric Res 3:1–11
Fandohan P, Hell K, Marasas W, Wingfield M (2003) Infection of maize by Fusarium species and contamination with fumonisin in Africa. Afr J Biotech 2:570–579
Gasperini AM, Medina A, Magan N (2022) Comparison of growth and aflatoxin B1 production profiles of Aspergillus flavus strains on conventional and isogenic GM-maize-based nutritional matrices. Fungal Biol 126:82–90
Gock MA, Hocking AD, Pitt JI, Poulos PG (2003) Influence of temperature, water activity and pH on growth of some xerophilic fungi. Int J Food Microbiol 81:11–19
Hope R, Aldred D, Magan N (2005) Comparison of environmental profiles for growth and deoxynivalenol production by Fusarium culmorum and F. graminearum on wheat grain. Lett Appl Microbiol 40:295–300
Kalai S, Anzala L, Bensoussan M, Dantigny P (2017) Modelling the effect of temperature, pH, water activity, and organic acids on the germination time of Penicillium camemberti and Penicillium roqueforti conidia. Int J Food Microbiol 240:124–130
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, Thierer T, Ashton B, Meintjes P, Drummond A (2012) Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28:1647–1649
Koňuchová M, Valík Ľ (2021) Modelling the radial growth of Geotrichum candidum: effects of temperature and water activity. Microorganisms 9:532
Llorens A, Mateo R, Hinojo M, Valle-Algarra F, Jiménez M (2004) Influence of environmental factors on the biosynthesis of type B trichothecenes by isolates of Fusarium spp. from Spanish crops. Int J Food Microbiol 94(1):43–54
Ma H, Snook LA, Tian C, Kaminskyj SG, Dahms TE (2006) Fungal surface remodelling visualized by atomic force microscopy. Mycol Res 110:879–886
Magan N, Lacey J (1984) Effect of water activity, temperature and substrate on interactions between field and storage fungi. Trans Br Mycol Soc 82:83–93
Marín S, Companys E, Sanchis V, Ramos A, Magan N (1998) Effect of water activity and temperature on competing abilities of common maize fungi. Mycol Res 102:959–964
Mbareche H, Veillette M, Bilodeau GJ (2021) In silico study suggesting the bias of primers choice in the molecular identification of fungal aerosols. J Fungi 7:99
Moore ML, Six DL (2015) Effects of temperature on growth, sporulation, and competition of mountain pine beetle fungal symbionts. Microb Ecol 70:336–347
Nelson PE, Desjardins AE, Plattner RD (1993) Fumonisins, mycotoxins produced by Fusarium species: biology, chemistry, and significance. Annu Rev Phytopathol 31:233–252
Nielsen KF (2003) Mycotoxin production by indoor molds. Fungal Genet Biol 39:103–117
O’Donnell K, Ward TJ, Robert VA, Crous PW, Geiser DM, Kang S (2015) DNA sequence-based identification of Fusarium: current status and future directions. Phytoparasitica 43:583–595
Obaid WA, Boutraa T, Abdel-Hameed UK (2020) Floristics and soil characteristics of Ohud mountain, Al-Madinah Al-Munawarah, Western Saudi Arabia. J Mt Sci 17:2405–2417
O’Donnell K, Cigelnik E (1997) Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the Fungus Fusarium Are nonorthologous. Mol Phylogenet Evol 7:103–116
O’Donnell K, Cigelnik E, Nirenberg HI (1998) Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia 90:465–493
Pal KK, Gardener BM (2006) Biological control of plant pathogens. Plant Health Instr 2:1117–1142
Park J, Cox-Ganser JM, Kreiss K, White SK, Rao CY (2008) Hydrophilic fungi and ergosterol associated with respiratory illness in a water-damaged building. Environ Health Perspect 116:45–50
Phan LTK, Nguyen HX, De Saeger S, Jacxsens L, Eeckhout M, Devlieghere F (2022) Predictive modelling of the radial growth of Aspergillus flavus and Fusarium proliferatum on paddy and white rice (Oryza sativa). Int J Food Microbiol 375:109743
Pietikäinen J, Pettersson M, Bååth E (2005) Comparison of temperature effects on soil respiration and bacterial and fungal growth rates. FEMS Microbiol Ecol 52:49–58
Pimentel D, Hepperly P, Hanson J, Douds D, Seidel R (2005) Environmental, energetic, and economic comparisons of organic and conventional farming systems. Bioscience 55:573–582
Rahman HU, Yue X, Ren X, Zhang W, Zhang Q, Li P (2020) Multiplex PCR assay to detect Aspergillus, Penicillium and Fusarium species simultaneously. Food Addit Contam Part A 37:1939–1950
Raja HA, Miller AN, Pearce CJ, Oberlies NH (2017) Fungal identification using molecular tools: a primer for the natural products research community. J Nat Prod 80:756–770
Ramirez ML, Chulze S, Magan N (2006) Temperature and water activity effects on growth and temporal deoxynivalenol production by two Argentinean strains of Fusarium graminearum on irradiated wheat grain. Int J Food Microbiol 106:291–296
Romero Donato CJ, Nichea MJ, Cendoya E, Zachetti VG, Ramirez ML (2023) Interacting Abiotic factors affect growth and mycotoxin production profiles of Alternaria section Alternaria strains on chickpea-based media. Pathogens 12:565
Saleh AA, Sharafaddin AH, El-Komy MH, Ibrahim YE, Hamad YK, Molan YY (2017) Fusarium species associated with date palm in Saudi Arabia. Eur J Plant Pathol 148:367–377
Samapundo S, Devlieghere F, De Meulenaer B, Geeraerd A, Van Impe J, Debevere J (2005) Predictive modelling of the individual and combined effect of water activity and temperature on the radial growth of Fusarium verticilliodes and F. proliferatum on corn. Int J Food Microbiol 105:35–52
Sautour M, Soares Mansur C, Divies C, Bensoussan M, Dantigny P (2002) Comparison of the effects of temperature and water activity on growth rate of food spoilage moulds. J Ind Microbiol Biotechnol 28:311–315
Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, Levesque CA, Chen W, Fungal Barcoding Consortium, Fungal Barcoding Consortium Author List (2012) Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc Natl Acad Sci USA 109:6241–6246
Skouboe P, Frisvad JC, Taylor JW, Lauritsen D, Boysen M, Rossen L (1999) Phylogenetic analysis of nucleotide sequences from the ITS region of Terverticillate Penicillium species. Mycol Res 103:873–881
van den Brule T, Punt M, Seekles SJ, Segers FJ, Houbraken J, Hazeleger WC, Ram AF, Wösten HA, Zwietering MH, Dijksterhuis J (2022) Intraspecific variability in heat resistance of fungal conidia. Food Res Int 156:111302
Verheecke-Vaessen C, Garcia-Cela E, Lopez-Prieto A, Jonsdottir IO, Medina A, Magan N (2021) Water and temperature relations of Fusarium langsethiae strains and modelling of growth and T-2 and HT-2 mycotoxin production on oat-based matrices. Int J Food Microbiol 348:109203
White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc Guide Methods Appl 18:315–322
Woudenberg J, Groenewald J, Binder M, Crous P (2013) Alternaria redefined. Stud Mycol 75:171–212
Yang R, Su J, Shang J, Wu Y, Li Y, Bao D, Yao Y (2018) Evaluation of the ribosomal DNA internal transcribed spacer (ITS), specifically ITS1 and ITS2, for the analysis of fungal diversity by deep sequencing. PLoS ONE 13:e0206428
Yoder JA, Klever LA, Dobrotka CJ (2018) Temperature-water and competitive interactions in vitro of two toxic black molds, Fusarium oxysporum and Rhizopus stolonifer isolated from a shower setting. Stud Fungi 3:248–255
Acknowledgements
I would like to thank the deanship of scientific research in Al-Baha University.
Funding
Not applicable right now.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
I have no conflicts of interest to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Almiman, B. Identifying the optimal temperature and water activity conditions of phytopathogenic fungi recovered from Al-Baha province. J.Umm Al-Qura Univ. Appll. Sci. (2024). https://doi.org/10.1007/s43994-024-00131-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s43994-024-00131-3