
Abstract
Background
The exposure of crops to a variety of fungal and bacterial pathogens leads to huge economic losses. Different strategies are being adapted to control these diseases among which the application of chemicals fungicide is common. However, these chemicals are posing a serious threat to the environment. For biological management of root rot disease of tomato and better fruit quality, studies were conducted on the possible use of endophytic yeast as a biocontrol agent.
Results
Endophytic yeasts were isolated from healthy plants and identified. Identification of selected isolates was confirmed on the basis of 18S rDNA gene sequencing. They were evaluated for suppressive effect on root rotting fungi in vitro and also in vivo on tomato plants, used alone or under neem cake soil amendment. Seventy-six isolates of yeasts were evaluated against root rotting fungi Fusarium oxysporum, F. solani, Rhizoctonia solani and Macrophomina phaseolina using dual culture plate assay. Seventy-five isolates were found to suppress radial growth of F. oxysporum, F. solani and M. phaseolina by producing zones of inhibition or lysing the fungal hyphae. However, none of the isolates was found to inhibit R. solani in vitro. Most of the isolates also caused nematicidal activity at varying degree against Meloidogyne javanica. All test isolates produced indole acetic acid in vitro and solubilized phosphorus. In pots and field plot experiments, test isolates of yeasts were able to suppress root rotting fungi on tomato in natural soil and soil amended with neem cake with enhancement of growth of tomato plants. Yeasts were also found to ameliorate the plant resistance through enhancing polyphenolic contents, salicylic acid and antioxidant activity.
Conclusions
Endophytic yeasts were found effective against root rot disease of tomato and could be used as a potential biocontrol agent for the management of soil-borne diseases of tomatoes.
Similar content being viewed by others

Background
Antagonistic microbes have emerged as a practical substitute to the chemical fungicides. They are used to suppress the survival or activity of a pathogen resulting in reduction in the incidence of diseases. Yeasts have been used in industries for thousands of years. Its several strains have now been recognized as potential biocontrol agents against plant diseases (Fareed et al. 2019). Pichia membranifaciens is reported to inhibit Rhizopus stolonifer on nectarine (Fan and Tian 2000), while Cryptococcus albidus suppressed Penicillium expansum in apples and pears (Tian et al. 2002). Another research indicated that plant growth enhancing and biocontrol ability of S. cerevisiae make it a strong competitor of Fusarium oxysporum which causes damping-off symptoms in sugar beet seedlings (Shalaby and El-Nady 2008). Species of Fusarium are necrotrophic pathogens of winter wheat that contaminate crops through their mycotoxins (Baliukoniene et al. 2011). Yeasts of genera Cryptococcus, Rhodotorula and Saccharomyces are pronounced to have deleterious effects on Fusarium sporotrichioides to some satisfactory degrees (Wachowska et al. 2013). Similarly, epiphytic yeasts have been reported to reduce post-harvest Penicillium rot of citrus (Habiba et al. 2019).
Tomato (Lycopersicon esculentum Mill.), widely consumed vegetable crop, is known for their nutritional values. The production of good quality tomatoes is highly favored by the diversified climatic conditions in Pakistan throughout the year, but natural obstacles are always there. Tomato plants are always subjected to many bacterial, fungal, viral and nematicidal attacks (Agrios 2005) but major yield reduction has resulted from fungal pathogens (Stone et al. 2000). This crop has a short life and is highly prone to mycotic diseases (Chohan et al. 2017). Greater losses were observed, when tomato crops were affected by both fungi and root knot nematodes (Parveen et al. 2020). The use of synthetic fungicide seems to be the best-known technique for the management of soil-borne diseases. Application of pesticides, no doubt, caused effective suppression of diseases resulting in the improvement of crop yield and quality (Sultana et al. 2011). However, the regular use of these chemicals may produce collateral problems. One of the major threats is the contamination of the environment with toxic elements which directly or indirectly affects human and animal health. Continuous application of fungicides to the crops also increases the pathogen resistance to such chemicals in particular soil environments (Tupe et al. 2014). These treatments are mostly non-specific, i.e., not only affect target pathogens but also other beneficial microorganisms (Ranganathswamy et al. 2013).
Aim of the application of any biocontrol agent is to not only suppress the disease but also to get high crop yield with good quality. A good and effective biocontrol agent has both the inhibitory effect on the pathogen as well as plant growth promoting ability. Many of the bioagents including yeast have been reported earlier as effective antagonists with positive impacts on plant growth (Moin et al. 2020). In a previous report, the biocontrol potential of endophytic yeasts against root rotting fungi of sunflower was reported (Fareed et al. 2019). The present research describes the efforts for further isolating the indigenous endophytic yeasts from healthy plants and determining their biocontrol potential against root rotting fungal pathogens of tomato in soil under neem cake amendment and their plant growth promoting ability. The report also describes the role of endophytic yeasts in enhancing the nutrient uptake and increasing antioxidant activity; salicylic acid and phenolic content status in plants.
Methods
Isolation and identification of endophytic yeast
Healthy plants (5 from each location) were collected from Karachi University and Malir, Karachi, Pakistan, and isolation of endophytic yeasts was made within 24 h in the laboratory. Stem, roots and leaves were separately cut into 2–3 cm long sections. The adhered debris and epiphytic microorganisms were removed by washing the cuttings with sterile water. They were then subjected to consecutive 1 min washes with 1% sodium hypochlorite, 70% ethanol and sterile distilled water. Surface-disinfected tissue was aseptically deliquesced with homogenizers. Serial dilution was made up to 10–6 and 0.5 mL from the last two dilutions was spread on YM medium plates, and then incubated for 5–7 days at 25 ± 1 °C. The yeast isolates grown were initially examined for their morphological characteristics based on colony color, texture and some physiological and biochemical assays (Kurtzman et al. 2011).
Molecular identification of potential yeasts
The DNA of freshly cultured yeast was extracted by using Biobasic Kit, Canada, as described by the manufacturer. ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (3'-TCC TCC GCT TAT TGA TAT GC-5′) primers were used for the amplification of rDNA on a thermal cycler (BioRad, USA). The amplified products were submitted to BGI Genomic Services (Shenzhen, Guangdong, China) for sequencing. For edition and alignment of sequences BioEdit software (Version 7.2.6) was used. Yeast isolates were identified by using BLAST and finally sequences were submitted to NCBI Genbank.
In vitro antifungal activity of endophytic yeasts
Newly isolated yeasts along with previously tested isolates (Fareed et al. 2019) were tested for antifungal activity in vitro against four root rotting fungi of tomato using dual culture plate assay. The Petri plates were poured with sterilized potato dextrose agar (PDA) and after solidification, the medium was streaked with yeast isolates on one side. A disk (5 mm) of test fungi viz., Fusarium oxysporum, F. solani, M. phaseolina and R. solani was positioned on the side of plates opposite to each yeast streak. The plates were kept at 28 °C for 3–7 days. The inhibitory effect of yeasts was assessed by measuring zones of inhibition produced by yeasts against test fungi. The experiment was conducted twice with three replicates.
Cell free culture filtrates of yeast and their nematicidal activity in vitro
The test yeast isolates were grown in YM broth in 250-ml flasks and incubated for 5 days. The broth was filtered over Whatman filter paper. The culture filtrate was then exposed to chloroform vapors to kill yeast cells, if any. Aqueous suspension (1 ml) of freshly hatched second juvenile (20 juveniles) was transferred in glass cavity blocks with 1 ml of a yeast culture filtrate. Each test was kept with 3 replicates at 26 ± 5 °C for 48 h and the nematode's mortality was recorded (Noreen et al. 2015). The experiment was repeated once.
Indole acetic acid (IAA) production by potential isolates of endophytic yeasts
The yeast isolates were grown in YM broth containing 0.1% (w/v) L-tryptophan and then incubated in the dark for 5 days. Broth was centrifuged at 4000 rpm for 15 min and one ml supernatant was mixed with 2 ml of Salkowski’s Reagent. O.D. was recorded at 530 nm on a spectrophotometer (Shimadzu, Japan, UV 1800) after 30 min, when color was developed (Gordon and Weber 1951). IAA production was calculated against the calibration curve using IAA. The experiment was conducted twice with 3 replicates.
Phosphate solubilization activity of potential yeast isolates
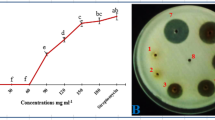
The ability of yeast to solubilize inorganic phosphorus was determined in vitro by using Pikovskaya’s agar medium (Pikovskaya 1948) in triplicate and repeated once. The fresh cultures of yeast isolates were inoculated on plates containing solid medium and left to incubate for 5 days at 25 °C. The solubilization of phosphorus was specified by the production of distinct halos around yeast colonies.
Clay pots experiment
Sandy loam soil was amended with neem cake [Sigma Energy (pvt) Ltd, Karachi] at 1% w/w and transferred to 12 cm clay pots at 1 kg per pot. A natural infestation of Fusarium spp. (3000 cfu g−1 of soil), 3–13% colonization of R. solani, and 3–9 sclerotia g−1 soil of M. phaseolina were found by using techniques described by Nash and Snyder (1962), Wilhelm (1955) and Sheikh and Ghaffar (1975). Pots were watered daily for 2 weeks. After decomposition of organic matter, 25 ml aqueous suspension (108cells/ml) of endophytic yeasts viz., Debaryomyces hansenii (Y-17), D. hansenii (Y-34), Rhodotorula mucilaginosa (Y-54) and Meyerozyma guilliermondii (Y-62), grown in YM broth for 1 week, was drenched into each pot. Four tomato (Lycopersicon esculentum Mill) seedlings were transplanted into each pot and each treatment was replicated 4 times. In another set, yeast suspensions were inoculated in un-amended soil. Carbendazim (25 mL of 200 ppm) served as positive control, while untreated plants served as control. The pots were kept with daily watering for 45 days. Observation on the effect of yeast isolates on the incidence of fungal infection and plant growth was examined after 45 days. Plant growth parameters (shoot length and shoot weight; root length and root weight) were recorded. To determine the infestation of root rot fungi, tap roots after washing were cut into pieces (1 cm), sterilize with 1% bleach and transferred onto PDA plates (amended with streptomycin (0.2 g ml−1) and Penicillin (100,000 units ml−1). Root rot fungi emerged from each piece after 5 days (at 25 °C) were identified and infection (%) for each fungus was calculated as described by Noreen et al. (2015). The experiment was repeated once, next year in similar conditions.
Field experiment
The antagonistic ability of the endophytic yeasts was also investigated under field conditions, where (2 × 2 m) plots of the experimental field of Crop Diseases Research Institute (CDRI) (PARC, Karachi University Campus, Karachi) was amended by neem cake. The neem cake was mixed in the soil at 70 g per 2 m row and allowed to decompose by watering on alternate days. Twelve healthy and equal sized seedlings of tomato were planted in each row. Each row was inoculated by 250 ml of yeast suspensions viz; Debaryomyces hansenii (Y-17), D. hansenii (Y-34) and Meyerozyma guilliermondii (Y-62). All the treatments were replicated 4 times. Plots without any treatment served as control, while carbendazim (100 ml of 200 ppm per meter row was drenched) served as positive control. Data were recorded after 45 and 90 days by uprooting four tomato plants from each treatment.
Estimation of biochemical parameters of leaf
Determination of phosphorus from leaf samples
Leaf phosphorus was measured by the following method used by Noreen et al. (2019). One g oven-dried leaf sample (dried at 120 °C for 24 h) was crushed and homogenized in 2N-HCl. The extract was digested for 60 min and then filtered (Whatman No.1 filter paper). Digested filtrate (10 ml) and freshly prepared Barton reagent (10 ml) were mixed in a 100-ml flask. Final volume was made 100 ml with distilled water. Absorbance of phosphorus vanado-molybdate complex was recorded against blank at 420 nm after 30 min and concentration of phosphorus was calculated in mg/g (Crous et al. 2019).
Estimation of polyphenols
The extraction of oven-dried leaves was done with ethanol (96% v/v). After centrifuging the extracts at 3000 rpm for 20 min, supernatants were used for analyzing polyphenols and antioxidants.
The Folin–Ciocalteu assay described by Rahman et al. (2017) was applied for the quantification of total phenols from plant leaves. Sodium bicarbonate (Na2CO3) 2% of 2 ml was added to 100 mL of extract (10 mg/ml) and left untouched for 2 min at room temperature. After adding 100 µL Folin–Ciocalteu reagents (50%), the mixture was incubated in a dark place for 30 min. The test samples were subjected to a spectrophotometer for determining their absorbance at 720 nm. Standard curve obtained from Gallic acid was used for the estimation of total phenol contents in the samples.
Estimation of salicylic acid (SA)
Salicylic acid in leaf samples was determined by using the method of Warrier et al. (2013), where 0.1 ml of chilled ethanolic leaf extracts was mixed with 3 mL of 0.1% ferric chloride. The absorbance was noted at 540 nm on the spectrophotometer. The quantity of SA (μg ml−1) was calculated as mg g−1 dried samples, where 100 mg of SA in 100 mL of ethanol was used for the standard curve.
Estimation of antioxidant activity
An aliquot of leaf sample (200 µl) was mixed with 800 µl of 10 Mm Tris HCl buffer (pH 7.4), then 30 µM DPPH (dissolved in DMSO) was added to the mixture. After mixing, the mixture was kept in the dark at room temperature. DPPH in ethanol served as control. The absorbance was taken at 517 nm against blank after 1 min and then at 30 min after keeping in dark. Free radical scavenging ability was calculated as described by Rahman et al. (2016).
Data analysis
Statistical software, CoStat (CoHort Software, USA) was used to determine the analysis of variance (ANOVA) and the significant level at (p < 0.05) was calculated using least significant difference (LSD).
Results
Isolation and identification of endophytic yeasts
A total of 76 isolates of endophytic yeasts were isolated and attentively identified (Additional file 1: Table S1).
Molecular identification of promising isolates
The potential isolates of endophytic yeasts were identified as Rhodotorula mucilaginosa (MT949216), Debaryomyces hansenii (MT949217), Debaryomyces hansenii (MT949218), Debaryomyces hansenii (MT949219), Debaryomyces hansenii (MT949220), Meyerozyma guilliermondii (MT949221), Debaryomyces hansenii (MT949222), Meyerozyma guilliermondii (MT949223) and Debaryomyces hansenii (MT949224) by molecular biology technique (Fig. 1).

Endophytic yeasts identified on the basis of 18S rDNA sequencing showing their ancestral relationship. Phylogenetic tree was generated using parsimony, neighbor-joining and maximum likelihood analysis of aligned sequences with different sets of cluster
In vitro antifungal activity of endophytic yeasts against root rotting fungi
The yeast isolates tested for their antifungal activity, showed varying degrees of inhibition against test fungi. Out of 76 isolates, 75 inhibited the mycelial growth of 3 tested fungi viz; F. solani, F. oxysporum and M. phaseolina, while none of the isolates inhibited R. solani. Among these isolates, some produced significant zones of inhibition around fungal growth. Lysis of fungal hyphae was also caused by some yeast isolates like: Y-37, Y-39, Y-41, Y-42, Y-43, Y-44, Y-45, Y-46, Y-47, Y-48, Y-49, Y-51, Y-53, Y-64 and Y-69 (Additional file 1: Table S1). Maximum inhibition of F. solani growth was observed by the yeast culture Y-34, Y-62 and Y-66. Yeast isolates Y-9, Y-25, Y-34, Y-38 and Y-62 produced maximum growth inhibition of F. oxysporum, whereas proliferation of M. phaseolina was greatly affected by isolates Y-5, Y-9, Y-34, Y-38 and Y-62 (Fig. 2; Additional file 1: Table S1).

Box-and-Whisker plot for inhibition of three fungal species. It shows median (middle line in the box), upper quartile (end of black box), lower quartile (end of gray box), maximum and minimum values
In vitro nematicidal activity of cell free culture filtrates of endophytic yeasts
Culture filtrates of different yeast isolates showed a nematicidal activity against root knot nematodes, juveniles at varying degrees. Complete killing of juvenile’s (100%) was exhibited by yeast isolates viz; Y- 5, Y-9, Y-29, Y-30, Y-34 and Y-66 within 24 h, of exposure, while yeast isolates Y-10, Y-21, Y-31, Y-65 and Y-67 killed the juveniles completely (100%) after 48 h, whereas several yeast isolates were able to kill more than 50 nematodes within 48 h (Fig. 3; Additional file 1: Table S1).

Box-and-Whisker plot for nematicidal activity. It shows median (middle line in the box), upper quartile (end of black box), lower quartile (end of gray box), maximum and minimum values
In vitro indole acetic acid (IAA) production by potential isolates of endophytic yeasts
The concentration of IAA produced by the yeast isolates was within the range from1.29 µg/ml to 9.21 µg/ml. Maximum IAA (9.2 µg/ml) was found to be produced by Y-17 and Y-62 as compared to other yeasts. Y-31 produced relatively the lowest IAA concentration (Table 1).
Phosphate solubilization activity by potential yeast isolates
Out of 15 yeast isolates tested for phosphate solubilizing ability, 12 were found to be efficient in this activity by producing clear halos around yeast colonies (Table 1).
Clay pots experiment
Plant growth was significantly enhanced by the application of endophytic yeasts either alone or along with neem cake. Significant increase at p < 0.05 in shoot length, shoot weight, root length and root weight were acquired by the plants treated with M. guilliermondii along with neem as compared to the non-treated control plants and plants having individual treatments of carbendazim and neem (Table 2). Treatments of plants with yeasts and neem cake significantly reduced the incidence of fungal pathogens in the root system. Complete suppression of F. solani was observed by the treatment of M. guilliermondii alone, followed by the minimum infection in D. hansenii (Y-17) treatment alone and in D. hansenii (Y-34) combination with neem cake. M. phaseolina and F. oxysporum were completely suppressed by M. guilliermondii and combined treatment of carbendazim and neem cake, respectively. Minimum infection by R. solani was produced by the individual treatment of neem cake followed by M. guilliermondii (Table 3).
In season II (repeated experiment), M. guilliermondii combined with neem cake and alone produced significantly highest shoot length, shoot weight, root length and root weight (Table 2). Treatments of plants with yeasts in neem cake amended soil significantly reduced the incidence of fungal pathogens in the root system. Significant suppression of F. solani was observed by the treatment of M. guilliermondii with neem cake. D. hansenii (Y-17) with neem cake showed significant reduction in infection of F. oxysporum. R. mucilaginosa (Y-54) with neem cake showed significant suppression of M. phaseolina (Table 4).
Field plot experiment
Tomato plants treated with endophytic yeasts either alone or along with neem cake showed better plant growth than control plants. Combined treatment of M. guilliermondii and neem resulted in maximum positive effect on plant growth. These plants also produced the greatest number of fruits as compared to untreated plants after 90 days but it was statistically non-significant. Maximum weight was gained by the fruits of plants treated with D. hansenii (Y-17) alone (Table 5). Infection by root rotting fungi was also significantly reduced by the application of endophytic yeasts. Within 45 days, minimum incidence of F. solani was detected in the roots of plants treated with D. hansenii (Y-17) alone followed by the individual treatment of M. guilliermondii but after 90 days combined treatment of M. guilliermondii and neem produced least F. solani infection. No infection by F. oxysporum was found in the treatments of D. hansenii (Y-17 and Y-34) and M. guilliermondii alone in 45 days but after 90 days it was little increased but still the least infection as compared to control plants. D. hansenii (Y-17) and combined treatment of M. guilliermondii and neem completely suppressed M. phaseolina within 45 days but after 90 days its least incidence was observed by the treatment of M. guilliermondii and D. hansenii (Y-34) with neem. Rhizoctonia solani was greatly suppressed by carbendazim alone and with neem in 45 days but after 90 days least infection was observed in the treatment of M. guilliermondii alone followed by the combined treatment of D. hansenii (Y-17) and neem (Table 6).
Estimation of biochemical parameters of leaf
Phosphorus content
Phosphorus content was significantly increased in the plants treated with yeast alone or along with neem cake. After 45 days, maximum phosphorus absorption was detected in the plants that were treated with M. guilliermondii along with neem cake, followed by the combined treatment of D. hansenii (Y-17) and neem. After 90 days, D. hansenii (Y-17) plus neem showed the highest uptake of phosphorus in contrast to control plants and other treatments (Table 7).
Salicylic acid
After 45 days, the level of salicylic acid (SA) was much increased in the plants treated with M. guilliermondii alone, followed by the combined treatment of D. hansenii (Y-34) and neem. Highest level of SA was produced by the treatment of M. guilliermondii alone after 90 days, followed by the combined treatment of D. hansenii (Y-17) and neem cake (Table 7).
Total phenolic contents
Meyerozyma guilliermondii alone treatment produced the highest phenolic contents within the 45 and 90 days interval, followed by the treatment of neem cake alone (Table 8).
Antioxidant activity
At day 45, antioxidant activity was much enhanced in the plants treated with M. guilliermondii alone, followed by the combined treatment of M. guilliermondii and cake neem. The same treatments exhibited the highest antioxidant activity in the plants after 90 days (Table 8).
Discussion
Among the major threats to food security are soil-borne pathogens including fungal phytopathogens and parasitic nematodes (Parveen et al. 2020). Emergence of endophytes as potential biocontrol agents for the management of plant diseases has opened a new window for research. In the present study, endophytic yeasts isolated from different healthy plants showed suppressive effect against root rotting fungi including F. solani, F. oxysporum, M. phaseolina, R. solani, and M. javanica, a root knot nematode in vitro. There are reports that yeast strains produce certain glycoproteins with antagonistic and fungicidal properties (Suzzi et al. 1995). They produced toxins, inhibitory to other microorganisms also known as “Killer Phenomenon.” This killer property was previously associated with only Saccharomyces cerevisiae but now many other yeast genera have been reported to produce killer toxins, such as Cryptococcus, Candida, Hanseniaspora, Debaryomyces, Hansenula, Pichia, Kluyveromyces, Tilletiopsis, Sporidiobolus and Zygosaccharomyces (Santos et al. 2000). The ability of several killer yeasts to antagonize filamentous pathogenic fungi is also well recorded (Li et al. 2016). The well-known killer strain of Saccharomyces cerevisiae has been used as a positive biocontrol agent against F. oxysporum (Shalaby and Nady 2008), F. solani and M. phaseolina (Attyia and Youssry 2001). Liu et al. (2018) reported the successful antagonistic activity of S. cerevisiae against Colletotrichum gloeosporioides, a per-harvest anthracnose agent of grapes. The killer strains may possibly produce certain metabolites like volatile compounds and hydrolytic enzymes which contribute to their antagonistic activity (Li et al. 2016). Antagonistic metabolites produced by endophytic yeasts need further investigation. In this study, several isolates on endophytic yeasts besides producing zones of inhibition also caused lysis of fungal hyphae. Yeasts produce different levels of hydrolytic enzymes such as chitinases and glucanases (Saligkarias et al. 2002), which have antagonistic effects on phytopathogenic fungi (Langner and Göhre 2016). Similarly, Candida and Kluyveromyces species have been reported to produce alkaline proteases (Agrawal and Kotasthane 2012). Alkaline proteases inhibitor is known to possess activity against phytopathogenic fungi (Vernekar et al. 1999).
In the present study, the endophytic yeasts applied as soil-drench individually or in soil amended with neem cake suppressed root rot pathogens of tomato. Akhtyamova and Sattarova (2013) reported antagonistic activity of yeasts against different pathogens. Abd El-Kader et al. (2012) reported S. cerevisiae as an effective bio-agent against root rot pathogens under green-house conditions. Similarly, Fareed et al. (2019) reported suppression of root rot of sunflower by the endophytic yeasts. The antagonistic properties of yeasts have opened the door for their use as natural biocontrol agents both in pre- and post-harvest crops diseases (Habiba et al. 2019).
Phosphorus, a major micronutrient, is required by both plants and microorganisms for their growth and productivity. Unfortunately, this phosphorus is not found in free form in soil (Yang et al. 2010). Microorganisms play their important role in solubilizing inorganic phosphate to soluble form and making it available for plant uptake. Use of phosphate solubilizing microagents provides an eco-friendly and sustainable method for dissolving both fertilizer phosphorus and bond phosphorus (Khan et al. 2007). In the present study, all the tested endophytic yeast isolates showed their phosphate solubilizing ability. Many yeasts belonging to different genera have previously been reported as positive phosphate solubilizers like: Saccharomyces, Rhodotorula, Hansenula, Kloeckera and Debaryomyces species (Varsha et al. 2010).
Utilization of organic amendments along with biological agents has become a promising practical approach. These organic amendments not only enhance crop productivity but also exhibit antagonistic behavior to plant pathogens. Several crop management programs have successfully improved the crop yields by the application of mustard cake, cotton cake, neem cake, castor cake and sesame cake as organic amendments (Shafique et al. 2016). In the present study, the neem cake amendment to the soil, either alone or in combination with endophytic yeast significantly enhanced vegetative growth and reduced fungal infections. Neem cake is used for its fungicidal and nematicidal effects as it has been reported (Shafique et al. 2016).
In this research, the antioxidant activity in the crop plants after treatment with yeasts was examined. The plants grown in soil amended with neem cake and yeast strains showed high levels of phenolic compounds and antioxidant activity than in the untreated plants. According to reports, total phenolic compounds increased in tomatoes by organic fertilizers (Toor et al. 2006) and there is a direct relationship of phenolic content with the antioxidant activity of plants (Kumar et al. 2008). The improved status of these compounds by the activity of antagonistic yeasts resulted in amelioration of plant defense mechanisms against the root rotting fungi and resulted in the improved growth of tomato plants. SA is a phenolic compound which behaves like a plant hormone by stimulating many physiological, biochemical and molecular processes and protecting it under stress conditions (Khan et al. 2013). Salicylic acid is a multifaceted hormone to combat diseases (Vlot et al. 2009). Application of P. churashimaensis as foliar spray was proved to be effective in disease suppression by stimulating plant defense mechanisms in pepper (Lee et al. 2017). Most probably, high SA activity in infected plants may increase H2O2 levels which results in the activation of systemic acquired resistance (SAR) against pathogens (Tian et al. 2007).
Conclusions
Many mycelial fungi and numerous bacteria have been reported as best biological antagonists against various plant pathogens. It is concluded from the study that endophytic yeasts can be utilized as potential biological antagonists against many root rotting fungi. The yeasts not only showed significant suppressive effects against these plant pathogens but also promoted plant growth. The improved status of antioxidant activity, phosphorus uptake, phenolic compounds and salicylic acid in treated plants ensured the potentiality of yeasts to combat successfully with the pathogens by inducing systemic acquired resistance in the host plants.
Availability of data and materials
Data will be provided on request.
Abbreviations
- rDNA:
-
Ribosomal deoxyribonucleic acid
- IAA:
-
Indole acetic acid
- FAO:
-
Food and Agriculture Organization
- YM:
-
Yeast malt
- DNA:
-
Deoxyribonucleic acid
- BGI:
-
Beijing Genomics Institute
- BLAST:
-
Basic Local Alignment Search Tool
- NCBI:
-
National Center for Biotechnology Information
- PDA:
-
Potato dextrose agar
- O.D:
-
Optical density
- Cfu:
-
Colony-forming unit
- CDRI:
-
Crop Diseases Research Institute
- PARC:
-
Pakistan Agricultural Research Council
- Min:
-
Minutes
- ml :
-
Milliliter
- v/v:
-
Volume/volume
- rpm:
-
Revolutions per minute
- µl :
-
Microliter
- nm:
-
Nanometer
- SA:
-
Salicylic acid
- DPPH:
-
2,2-Diphenyl-1-picrylhydrazyl
- LSD:
-
Least significant difference
References
Abd El-Kader MM, El-Mougy NS, Aly M, Lashin DE (2012) Different approaches of bio-control agents for controlling root rot incidence of some vegetables under greenhouse conditions. IJAFP 2(1):115–127. https://doi.org/10.5923/j.ijaf.20120201.18
Agrawal T, Kotasthane AS (2012) Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in Central India. Springer plus 1(1):1–10
Agrios GN (2005) Plant pathology, 5th edn. Academic Press, New York, p 922
Akhtyamova N, Sattarova RK (2013) Endophytic yeast Rhodotorula rubra strain TG-1: antagonistic and plant protection activities. Biochem Physiol 2(1):104. https://doi.org/10.4172/2168-9652.1000104
Attyia SH, Youssry AA (2001) Application of Saccharomyces cerevisiae as a biocontrol agent against some diseases of Solanaceae caused by Macrophomina phaseolina and Fusarium solani. Egypt J Biol 3:79–87
Baliukoniene V, Bakutis B, Januseviciene G, Miseikine R (2011) Fungal contamination and Fusarium mycotoxins in cereal grown in different tillage systems. J. Anim. Feed Sci 20(4):629–637. https://doi.org/10.22358/jafs/66222/2011
Chohan S, Perveen R, Abid M, Naqvi AH, Naz S (2017) Management of seed borne fungal diseases of tomato: a review. Pak. J. Phytopathol. 29(01):193–200
Crous KY, Wujeska-Klause A, Jiang M, Medlyn BE, Ellsworth DS (2019) Nitrogen and phosphorus retranslocation of leaves and stemwood in a mature Eucalyptus forest exposed to 5 years of elevated CO2. Front Pl Sci 10:664. https://doi.org/10.3389/fpls.2019.00664
Fan Q, Tian SP (2000) Postharvest biological control of Rhizopus rot on nectarine fruit by Pichia membranifaciens Hansen. Plant Dis 84(11):1212–1216. https://doi.org/10.1094/PDIS.2000.84.11.1212
Fareed A, Ali SA, Hasan KA, Sultana V, Ehteshamul-Haque S (2019) Evaluation of biocontrol and plant growth promoting potential of endophytic yeasts isolated from healthy plants. Pak J Bot 51(6):2283–2289
Gordon SA, Weber RP (1951) Colorimetric estimation of indoleacetic acid. Plant Physiol 26(4):192. https://doi.org/10.1104/pp.26.1.192
Habiba NR, Ali SA, Hasan KA, Sultana V, Ara J, Ehteshamul-Haque S (2019) Evaluation of biocontrol potential of epiphytic yeast against postharvest Penicillium digitatum rot of stored Kinnow fruit (Citrus reticulata) and their effect on its physiochemical properties. Postharvest Biol Technol 148:38–48. https://doi.org/10.1016/j.postharvbio.2018.10.007
Khan MS, Zaidi A, Wani PA (2007) Role of phosphate-solubilizing microorganisms in sustainable agriculture—a review. Agron Sustain Dev 27(1):29–43
Khan MI, Iqbal N, Masood A, Per TS, Khan NA (2013) Salicylic acid alleviates adverse effects of heat stress on photosynthesis through changes in proline production and ethylene formation. Plant Signal Behav 8(11):263–274. https://doi.org/10.4161/psb.26374
Kumar KS, Ganesan K, Subba Rao PV (2008) Antioxidant potential of solvent extracts of Kappaphycus alvarezii Doty-An edible seaweed. Food Chem 107(1):289–295. https://doi.org/10.1016/j.foodchem.2007.08.016
Kurtzman C, Fell JW, Boekhout T (2011) The Yeasts: A Taxonomic Study, 5th edn. Elsevier
Langner T, Göhre V (2016) Fungal chitinases: function, regulation, and potential roles in plant/ pathogen interactions. Curr Genet 62(20):243–254. https://doi.org/10.1007/s00294-015-0530-x
Lee G, Lee SH, Kim KM, Ryu CM (2017) Foliar application of the leaf-colonizing yeast Pseudozyma churashimaensis elicits systemic defense of pepper against bacterial and viral pathogens. Sci Rep 7(1):39432
Li B, Peng H, Tian S (2016) Attachment capability of antagonistic yeast Rhodotorula glutinis to Botrytis cinerea contributes to biocontrol efficacy. Front Microbiol 7:601. https://doi.org/10.3389/fmicb.2016.00601
Liu Z, Du S, Ren Y, Liu Y (2018) Biocontrol ability of killer yeasts (Saccharomyces cerevisiae) isolated from wine against Colletotrichum gloeosporioides on grape. J Basic Microbiol 58(1):60–67. https://doi.org/10.1002/jobm.201700264
Moin S, Ali SA, Hasan KA, Tariq A, Sultana V, Ara J, Ehteshamul-Haque S (2020) Managing the root rot disease of sunflower with endophytic fluorescent Pseudomonas associated with healthy plants. Crop Protect 130:105066. https://doi.org/10.1016/j.cropro.2019.105066
Nash SM, Snyder WC (1962) Quantitative estimations by plate counts of propagules of the bean root rot Fusarium in field soils. Phytopath 52(6):567–572
Noreen R, Ali SA, Hasan KA, Habiba UF, Tariq A, Ara J, Ehteshamul-Haque S (2019) Role of fluorescent Pseudomonas associated with root nodules of mungbean in the induction of nodulation by the rhizobia in mungbean. Pak J Bot 51(3):1161–1168. https://doi.org/10.30848/PJB2019-3(44)
Noreen R, Ali SS, Hasan KA, Sultana V, Ara J, Ehteshamul-Haque S (2015) Evaluation of biocontrol potential of fluorescent Pseudomonas associated with root nodules of mungbean. Crop Protect 75:18–24
Parveen G, Urooj F, Moin S, Farhat H, Fahim MF, Ehteshamul-Haque S (2020) Estimation of losses caused by root rotting fungi and root knot nematodes infecting some important crops in Lower Sindh and Hub, Balochistan of Pakistan. Pak J Bot 52(2):673–678. https://doi.org/10.30848/PJB2020-2(15)
Pikovskaya RI (1948) Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 17:362–370
Rahman A, Sultana V, Ara J, Ehteshamul-Haque S (2016) Induction of systemic resistance in cotton by the neem cake and Pseudomonas aeruginosa under salinity stress and Macrophomina phaseolina infection. Pak J Bot 48(4):1681–1689
Rahman A, Korejo F, SultanaV AJ, Ehteshamul-Haque S (2017) Induction of systemic resistance in cotton by the plant growth promoting rhizobacterium and seaweed against charcoal rot disease. Pak J Bot 49:347–353
Ranganathswamy M, Patibanda AK, Nageswara Rao G (2013) Evaluation of toxicity of agrochemicals on Trichoderma isolates in vitro. J Mycopathol Res 51(2):289–293
Saligkarias ID, Gravanis FT, Epton HAS (2002) Biological control of Botrytis cinerea on tomato plants by the use of epiphytic yeasts Candida guilliermondii strains 101 and US7 and Candida oleophila strain I-182: II. A study on mode of action. Biol Control 25(2):151–161. https://doi.org/10.1016/S1049-9644(02)00052-X
Santos A, Marquina D, Leal JA, Peinado JM (2000) (1,6)-β-D-glucan as cell wall receptor for Pichia membranifaciens killer toxin. Appl Environ Microbiol 66(5):1809–1813. https://doi.org/10.1128/AEM.66.5.1809-1813.2000
Shafique HA, Sultana V, Ehteshamul-Haque S, Athar M (2016) Management of soil-borne diseases of organic vegetables. J Pl Protect Res 56(3):221–230. https://doi.org/10.1515/jppr-2016-0043
Shalaby ME, El-Nady MF (2008) Application of Saccharomyces cerevisiae as a biocontrol agent against Fusarium infection of sugar beet plants. Acta Biologica Szegediensis 52(2):271–275
Sheikh AH, Ghaffar A (1975) Population study of sclerotia of Macrophomina phaseolina in cotton fields. Pak J Bot 7:13–17
Stone JK, Bacon CW, White J (2000) An overview of endophytic microbes: endophytism defined. Micro Endophytes 3:29–33
Sultana V, Baloch AJ, Ehteshamul-Haque S, Tariq RM, Athar M (2011) Seaweeds as an alternative to chemical pesticides for the management of root diseases of sunflower and tomato. J Appl Bot Food Qual 84(2):162–168
Suzzi G, Romano P, Ponti I, Montuschi C (1995) Natural wine yeasts as biocontrol agents. J Appl Bacteriol 78(3):304–308. https://doi.org/10.1111/j.1365-2672.1995.tb05030.x
Tian SP, Fan Q, Xu Y, Liu HB (2002) Biocontrol efficacy of antagonist yeasts to grey mold and blue mold on apples and pears in controlled atmospheres. Plant Dis 86(8):848–853. https://doi.org/10.1094/PDIS.2002.86.8.848
Tian S, Qin S, Li B, Wang Q, Meng X (2007) Effects of salicylic acid on disease resistance and postharvest decay control of fruits. Stewart Postharvest Rev 3:1–7. https://doi.org/10.2212/spr.2007.6.2
Toor RK, Savage GP, Heeb A (2006) Influence of different types of fertilizers on the major antioxidant components of tomatoes. J Food Compos Anal 19(1):20–27. https://doi.org/10.1016/j.jfca.2005.03.003
Tupe SG, Chaudhary PM, Deshpande SR, Deshpande MV (2014) Development of novel molecules for the control of plant pathogenic fungi in agriculture. In: Kharwar RN, Upadhyay RS, Dubey NK, Raghuwanshi R (eds) Microbial diversity and biotechnology in food security. Springer, New Delhi, pp 315–325
Varsha N, Ahmed Abu Samaha SM, Patel HH (2010) Rock phosphate dissolution by specific yeast. Indian J Microbiol 50(1):57–62
Vernekar JV, Ghatge MS, Deshpande VV (1999) Alkaline protease inhibitor: a novel class of antifungal proteins against phytopathogenic fungi. Biochem Biophys Res Commun 262(3):702–707. https://doi.org/10.1006/bbrc.1999.1269
Vlot AC, Dempsey DA, Klessig DF (2009) Salicylic acid, a multifaceted hormone to combat disease. Annu Rev Phytopathol 47:177–206. https://doi.org/10.1146/annurev.phyto.050908.135202
Wachowska U, Kucharska K, Jedryczka M, Lobik N (2013) Microorganisms as biological control agents against Fusarium pathogens in winter wheat. Pol J Envion Stud 22(2):591–597
Warrier R, Paul M, Vineetha MV (2013) Estimation of salicylic acid in Eucalyptus leaves using spectrophotometric methods Genet. Plant Physiol 3(1–2):90–97
Wilhelm S (1955) Longevity of the Verticillium wilt fungus in the laboratory and field. Phytopath 45(3):180–181
Yang M, Ding G, ShiL FJ, Xu F (2010) Quantitative trait loci for root morphology in response to low phosphorus stress in Brassica napus. Theor Appl Genet 121(1):181–193
Funding
Financial assistance was provided by the Dean, Faculty of Science, University of Karachi.
Author information
Authors and Affiliations
Contributions
AJ, FU and AR isolated endophytic yeasts and performed experiments on tomatoes. HF and MI confirmed the identification of yeast using molecular biology techniques. AJ and HF analyzed and interpreted the results and wrote the manuscript. SE conceived and designed experiments, supervised research work and improved the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
No conflict of interest declared.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Growth inhibition of root rotting fungi Macrophomina phaseolina, Rhizoctonia solani, Fusarium solani and F. oxysporum in dual culture plate assay and mortality of root knot nematode Meloidogyne javanica by the endophytic yeast isolates
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jamal, A., Farhat, H., Urooj, F. et al. Characterization of endophytic yeast and its suppressive effect on root rotting fungi of tomato under neem cake soil amendment. Egypt J Biol Pest Control 31, 152 (2021). https://doi.org/10.1186/s41938-021-00493-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-021-00493-4