
Abstract
Endometriosis is a common condition in women of reproductive age, but its current interventions are unsatisfactory. Recent research discovered a dysregulation of the sphingosine 1-phosphate (S1P) signaling pathway in endometriosis and showed a positive outcome by targeting it. The S1P axis participates in a series of fundamental pathophysiological processes. This narrative review is trying to expound the reported and putative (due to limited reports in this area for now) interactions between the S1P axis and endometriosis in those pathophysiological processes, to provide some perspectives for future research. In short, S1P signaling pathway is highly activated in the endometriotic lesion. The S1P concentration has a surge in the endometriotic cyst fluid and the peritoneal fluid, with the downstream dysregulation of its receptors. The S1P axis plays an essential role in the migration and activation of the immune cells, fibrosis, angiogenesis, pain-related hyperalgesia, and innervation. S1P receptor (S1PR) modulators showed an impressive therapeutic effect by targeting the different S1P receptors in the endometriosis model, and many other conditions resemble endometriosis. And several of them already got approval for clinical application in many diseases, which means a drug repurposing direction and a rapid clinical translation for endometriosis treatments.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Current Treatments for Endometriosis
Endometriosis, defined as the presence of ectopic endometrium outside of the uterus, is a chronic, debilitating disease associated with pelvic pain and infertility [1]. The most common locations of this kind of dislocation are the surface of the peritoneum and ovary. Endometriosis is estimated to affect 6–10% of reproductive-age women [2]. In 2012, the estimated economic burden was over US$22 billion in the USA and 8.5 billion in the UK [3, 4]. The prevalence of endometriosis was estimated to exceed 50% of women with pelvic pain and 50% of women with infertility, which amounts to about 176 million women worldwide [5, 6]. The asymptomatic women (confirmed by microscopic histological diagnosis) are another large group of endometriosis patients (45–50%) [7].
Symptoms of endometriosis considerably harm patients’ psychological wholesome, social well-being, and quality of life [8]. Compared to the general women population, endometriosis patients have significantly high morbidity of sub-/infertility, and the mechanism of it is still poorly understood. Even if the patients finally got pregnant, it was reported that a series of poor pregnancy outcomes, such as preterm labor, pre-eclampsia, ectopic pregnancy, miscarriage, and intrauterine growth restriction, are associated with the disease [9, 10]. Pain symptoms like persistent or cyclical pelvic pain, dysmenorrhea, and dyspareunia are the other problematic annoyances for the patients. And, the same as other chronic pain conditions, endometriosis patients were reported to suffer from fatigue and depression.
Current treatments for endometriosis, including surgical and medical management, aim to improve the pain symptom and treat the infertility condition [2, 11]. Ovarian suppression is the primary strategy of medical management that can limit lesions’ activity and growth, alleviating pain symptoms. The usual way of ovarian suppression consists of oral contraceptives and gonadotrophin-releasing hormone (GnRH) agonists with add-back hormone-replacement therapy [12], while GnRH agonists have side effects such as memory loss, insomnia, and hot flushes, thus limiting long-term use. The utilization of progesterone to counteract estrogen can reduce pain symptoms, but the contraceptive effect of the treatment is not good news for women longing for conception [13]. Although non-steroidal anti-inflammatory drugs (NSAIDs) are another option for pain control, they cannot suppress the growth of endometriotic lesions, and long-time use is prone to inducing peptic ulcers. And for surgical management, laparoscopic surgery to remove lesions works for most patients in symptom relief. However, after the surgery, about 50% of patients will suffer symptom relapse within 2 years [14]. Thus, it is imperative to develop new treatments for endometriosis with fewer side effects, the ability to suppress lesion growth or even eliminate it, and, at the same time, allow patients to conceive.
There have been many attempts to develop new drugs for clinical use for endometriosis patients by repurposing drugs that have already proved effective in treating other diseases resembling endometriosis with painful inflammatory conditions [11]. Several pieces of research have already shown that this is a feasible way. For instance, it is demonstrated that immunotherapy targeting T cells or cytokines are a viable approach to handling the pain symptom of a similar disease like inflammatory bowel disease and rheumatoid arthritis [15]. And we noticed that as a biologically active molecule, sphingosine-1-phosphate (S1P) was demonstrated to play a pivotal role in endometriosis too and can be another promising direction for drug repurposing.
S1P Metabolism and Gradients
Sphingolipids, a specific class of lipids, are important components of cell membranes, which have structural and functional activities [16]. Sphingosine, 18-carbon amino alcohol with an unsaturated hydrocarbon chain, is the general characterization of sphingolipids, which means it is a constituent of all the lipids named sphingolipids, sphingomyelin, and glycosphingolipids. S1P is the most biologically active one of the sphingolipids, not only taking part in cellular homeostasis and viability [17] but also regulating immune cell migration and inflammatory responses [18]. S1P can act as the second messenger for the intracellular pathway and as a signaling molecule for the extracellular compartment [17]. As an extracellular signaling molecule, S1P mainly relies on 5 G protein-coupled receptors, S1P1–S1P5, to wield its regulatory power [19] (Fig. 1).

The metabolism of sphingosine 1-phosphate (S1P). Ceramide is the core of sphingomyelin metabolism, which can be de novo synthesized in the endoplasmic reticulum or degraded from sphingomyelin in the lysosome. And the sphingosine comes from the deacylation of ceramide which is catalyzed by ceramidase (CDase) in the lysosome or on the cell membrane. Then the sphingosine can be phosphorylated by sphingosine kinase 1 (SphK1) or SphK2 to form S1P. S1P can be exported by SPNS2, MFSD2B for red blood cells and platelets, to activate different S1PRs called “inside out signaling” to achieve a series of functions. The red blood cell, albumin, and ApoM on HDL are the main transporters of S1P in plasma. SPT, serine palmitoyltransferase; KDSR, 3-ketodihydrosphingosine reductase; CERS, ceramide synthase; DES, dihydroceramide desaturase; SMS, sphingomyelin synthase; SPP, sphingosine phosphate phosphatase; SPL, S1P lyase
The gradients are vital for most cytokines, chemokines, and developmental morphogens to implement their functions. Unlike other inflammation-related cytokines and chemokines, apart from secretion under tissue injury and pathogen invasion stimuli, S1P also acts as a “route sign” for immune cell trafficking and organ development [20]. The abundance of S1P lyase in tissues keeps S1P in a very low intercellular concentration. On the contrary, blood has a considerable concentration of S1P, about 1 μM, under the supply of red blood cells (RBCs), as might be the “route sign” of blood for immune cells. The synthesis of S1P is especially active in RBCs. However, the degradation of S1P is limited due to the lack of S1P lyase and phosphatase activity in RBCs, which makes RBCs to be the “S1P pool” [21,22,23]. And it was discovered recently that the major facilitator superfamily member MSFD2B was the primary transporter of RBCs to pump out S1P [24, 25]. Without the MSFD2B, S1P was found to accumulate in the inner layer of the RBCs’ plasma membrane of Mfsd2b−/− mice, which endows a significant proportion of RBCs a stomatocytes look [25]. As for platelets, MSFD2B is also critical for the release of S1P. The deletion of MSFD2B impairs platelet morphology and functions, reducing thrombus formation [26]. The secondary source of plasma S1P is endothelial cells, and, unlike RBCs and platelets, the major facilitator superfamily member SPNS2 is the transporter they rely on [27,28,29,30,31]. As a lipid mediator with limited aqueous solubility, S1P chaperone proteins solved this problem by stably binding and transporting S1P in plasma and interstitial fluids, helping activate S1PRs on target cells. The apolipoprotein M (ApoM) is the prototypical chaperone of S1P, which is a component of high-density lipoprotein (HDL), and HDL is used to be believed that is beneficial to vascular health [32]. Albeit at lower affinity, serum albumin is also a chaperone that binds to S1P [33].
Besides the blood plasma, lymph S1P plays an integral part in guiding immune cells out of lymph nodes and Peyer patches. Even though the circulatory loop is from plasma to interstitial fluid to lymph and then back to plasma, the concentration of S1P in tissues is extremely low due to the high activity of S1P lyase. However, the concentration of lymph S1P (∼0.1 μM) is relatively high [34], and it was demonstrated that lymphatic endothelial cells are the primary source of lymph S1P [21]. Loss of lymph S1P was observed in lymphatic endothelial cells lack of SphKs or SPNS2, which caused the block of lymphocytes to exit the lymph nodes [31, 35, 36]. And it was demonstrated recently that the inflammatory monocyte-derived S1P in the lymph node also works in regulating immune responses [37].
In conclusion, the S1P spatial gradients are created and maintained by a sophisticated system including cell secretion, tissue degradation, chaperone binding, and S1PRs, specifically activation, which guarantees that the S1P axis wields its biological function [38].
S1P in Modulating the Immune Cells
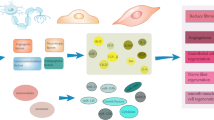
Even though the exact pathogenesis of endometriosis is still controversial, multiple lines of evidence repeatedly illustrated an altered peritoneal environment of patients with endometriosis. Not only a lot of immune cells were attracted to the lesions, but also plenty of pro-inflammatory proteins and cytokines such as IL-1, IL-6, IL-8, and IL-33, insulin-like growth factor 1 (IGF-1), and tumor necrosis factor (TNF) were aberrantly produced [11, 39,40,41]. A multiparameter single-cell technique was utilized to detect a large number of different immune cell types in the peritoneal fluid (PF) of the patients. Among these immune cells, the mononuclear phagocyte was the predominating one from the innate immune system, and it was suggested that CD69+ T cells might be associated with endometriosis [42]. Macrophages are originally the predominant immune cell type in the PF of the healthy population, and the number is further increased in endometriosis patients. Another immune cell worth noting are T cells. The activation and effector activity was increased compared to healthy control [42]. Xiao et al. recently proved the potential importance of T cell phenotype in the etiology of endometriosis, which assured the T cell dysregulation in the endometriotic lesions [43]. The S1P axis was highly associated with the migration, activation, and residence of macrophages and T cells. And perturbed S1P signaling was demonstrated both in the PF and the endometriotic lesions [44, 45]. Thus, we mainly reviewed these two groups of immune cells below (Fig. 2).

The infiltration of macrophages and T cells from blood and peritoneal fluid into the endometriotic lesions. The S1P receptor 1 (S1P1) is the most concerned one among all S1PRs in regulating T cells and macrophage migration. And the S1P2 and S1P3 are reported to take part in modulating macrophage polarization into M1 and M2, respectively. The erythrocyte lysis and platelet activation in endometriotic lesion release amounts of S1P in the lesion. And multiple cytokine and chemokine release is also regulated by the S1P axis
Macrophage
Although the ontogeny of macrophages gathering in endometriosis is still an enigma, studies show an increase in the total number and proportion of macrophages in the endometriotic lesions and the association between lesion growth and macrophage activation. It was detected that the lesion-resident macrophages contain both those from eutopic endometrium and those from the peritoneal environment [46]. An increased number of macrophages imply prolonged longevity of resident one or a rise of infiltrating one or both.
For prolonged longevity, researchers have found that S1P plays a pivotal role in the immune system homeostasis by regulating immune cell migration and their longevity, including the differentiation, migration, and survival of macrophages [47]. S1P wields an anti-apoptotic function on macrophages. It was shown that S1P prevented caspase-induced apoptosis by inducing the expression of anti-apoptotic proteins through extracellular-signal-regulated kinase (ERK1/2), phosphoinositide-3-kinase (PI3K), and Ca2+ signaling pathways or altering the metabolic balance from ceramide to sphingosine or S1P [48,49,50]. It was also shown that the anti-apoptotic function of HDL depends on S1P to induce inhibitor of apoptosis (IAP) family member survivin via STAT3 signaling in THP-1 and RAW264.7 macrophages. Still, this effect can be inhibited by antagonists of S1P2/3, indicating that this effect is mediated by S1P2/3 [51]. S1P2 was shown to suppress macrophage pyroptosis by decreasing the expression of caspase11 protein in peritoneal macrophages [52].
And for the increased infiltration of the environment macrophages, S1P can act as a chemokine to recruit macrophages into the inflammatory site [53]. Previous research showed that the S1PRs expression profile of macrophages would determine the S1P-dependent macrophage migration [54]. Among all the S1PRs, S1P1 acts pro-migratory in peritoneal and bone marrow-derived macrophages (BMDM) through Rho kinase and PI3K-Akt1 signaling to the inflammatory site [53, 55]. S1P1 is also vital for post-inflammatory macrophage emigration because macrophage-specific deletion of S1P1 resulted in tissue retention of macrophages in a chemical-induced mouse model of resolving peritoneal inflammation [56]. Studies have shown that S1P4 signaling may act as an anti-migratory factor, and consequently, the S1P1/S1P4 ratio is critical for M1 migratory behavior. The analysis of M1 and M2 macrophages showed that the S1P1/S1P4 ratio of M1 macrophages was higher than that of M2 macrophages [57]. Compared to the pro-migratory effect of S1P1, using S1pr2-knockout mice in an acute inflammatory peritonitis model demonstrated that S1P2 signaling can prevent macrophage migration by stimulating cAMP production and therefore abating the Akt phosphorylation [58]. These works illustrated that S1P1 and S1P2 have opposite properties in immune cell migration. Chemotaxis of bone macrophages, osteoclast precursors, is simultaneously regulated by S1P1/2, which is used to adjust their localization in bone, which supports the previous hypothesis to some extent [59].
Not only the number of macrophages but also the phenotypes and activation status exhibit a difference between women with and without endometriosis. Macrophages show different or diametrical roles in different stages of inflammatory response. According to their functional characteristics and differences in cytokine expression, they are generally divided into two categories with different functions: classically activated macrophage (M1) produces a large number of inflammatory cytokines, specifically removes microorganisms and defective cells, and exhibits an overall pro-inflammatory effect; and alternatively activated macrophage (M2) regulates adaptive immune responses, remove cell debris, promote angiogenesis and tissue repair, and generally exhibit anti-inflammatory effects.
Macrophages have been found to accumulate in the foci [42]. An abnormally elevated proportion of M2-type macrophages has been found in rhesus monkey models, and patients with endometriosis [60, 61], and M2 has a significant impact on the lesions. The promotion of growth and fibrosis has been demonstrated in mouse models [62], and the anti-inflammatory effects of M2 play a role in ectopic endometriotic lesion survival, foci formation, angiogenesis, and regulation of T-cell immunity. In addition to locally aggregated macrophages, circulating monocytes in patients have also been shown to promote the proliferation of endometrial cells, while controls exhibit an inhibitory effect [63]. Peritoneal macrophages may also play a pivotal role in infertility symptoms and pelvic pain. Macrophages release pro-fibrotic mediators such as transforming growth factor (TGF), interleukin 13 (IL-13), or platelet-derived growth factor (PDGF) [64], which is the key to fibroblast activation and proliferation. Although the effects of macrophages on lesions are determined, we still do not know the molecular mechanisms underlying macrophage aggregation at endometriosis foci and why massive M2 polarization occurs. Studies have shown that a large number of macrophages are often gathered in the foci of endometriosis, and M2 has an abnormally high proportion in it [65, 66]. By comparing patients with endometriosis and healthy people, it was found that M2 macrophages were distributed in large numbers in the lesions and increased in the peritoneal epithelium and peritoneal fluid [60, 67]. The total number and proportion of activated macrophages are both increased in endometriosis patients, and consequentially, they produce more cytokines. Inflammatory conditions, especially those with erythrocyte lysis, will release a lot of S1P, which will be sensed by macrophages. Not only macrophages but a lot of circulating progenitor cells and various immune cells are also proven to be able to sense the increased concentration of S1P in the inflammatory site [47]. It was demonstrated in the bone healing model that activation of S1P/S1PRs could activate M2 polarization of macrophages, which promoted vascularization and attracted more BMDM to the site of inflammation [68]. And recently, an endometriosis study demonstrated that the elevated S1P concentration in PF skewed M2 activation and causatively induced IL-6 and cyclooxygenase-2 (COX2) mRNA overexpression [69]. However, the exact mechanism of how S1P regulates M1/M2 differentiation is still not fully understood. When researchers use FTY720, a non-selective S1PRs agonist except for S1P2, to treat bone fractures, no difference in the total number of osteoclast (a kind of macrophage) was observed at the wound site, and no sign of improved healing [70]. In research on myocardial infarction, the S1P/S1P1 signaling pathway was activated by exosomes from adipose-derived mesenchymal stem cells to induce macrophage M2 polarization [71]. And Yang et al. showed that S1P could promote BMDMs into M1 differentiation, and it was demonstrated that the suppression of S1P2/3 expression or any part of the G(α)i/o/PI3K/JNK signaling pathway could prevent this effect, which suggests that the S1P2/3/G(α)i/o/PI3K/JNK signaling pathway is the mechanism S1P relies on to induce M1 differentiation [72]. Consistently, a single-cell analysis observed a decrease in the M1/M2 ratio and an increase of muscle stem cells that are beneficial to injury repair in an S1pr3-knockout muscle injury model, and this result can be reconstructed with VPC01091, an S1P3 antagonist, which improved tissue regeneration [73]. Due to the different main roles of macrophages in different stages of the inflammatory response, the types of macrophages in various tissues are also quite different. Therefore, different tissue sources and different in vitro macrophage differentiation protocols affect macrophage migration and activation. The regulation of S1P on macrophages is flexibly regulated according to the local environment, the difference in intracellular and extracellular concentrations of S1P, and the specific S1PR activated on macrophages [47]. That is why S1P can not only promote the polarization of macrophages to M2 type but also promote the production of M1-related macrophage markers under a different context. SphK1 is mainly distributed in the cytoplasm, and when activated, it will be transferred to the plasma membrane to catalyze sphingosine to form S1P, a cofactor involved in activating M1. M1 is primarily triggered by various microbial components, such as lipopolysaccharide (LPS), with or without IFN-γ. After activation, M1 rapidly produces large amounts of TNF-α through enhanced mRNA stability [74]. Then, TNF-α binds to its cognate receptor tumor necrosis factor receptor-associated factor 2 (TRAF-2), promoting the activation of NF-κB [75]. SphK1 is activated downstream of TNF receptor activation, which is associated with TRAF2. S1P catalyzed by SphK1 then acts as a cofactor for TRAF2 and is involved in mediating the polyubiquitination of RIP1 [76, 77].
In endometriosis, there is a decrease in the phagocytic activity of macrophages [78], resulting in the inability of macrophages to clear the ectopic endometrial cells that reach the lumen through retrograde menstruation. Beyond regulating macrophage activation and facilitating recruitment, S1P was recently proven to be involved in priming macrophages for uptaking cell debris [79]. During the studying of another chronic inflammatory disease, chronic obstructive pulmonary disease (COPD), researchers found that the activation of erythropoietin (EPO) signaling in murine macrophages by the S1P released from apoptotic cells induced upregulation of phagocytic receptors, CD36 and Mer tyrosine kinase (MerTK), and simultaneously, mice lacking the EPO receptor in myeloid cells showed delayed clearance of apoptotic cells and lupus-like autoimmune symptoms [79]. And for macrophages themselves, inhibition of SphKs by pharmacological inhibitors or cigarette smoke can both reduce the efferocytosis in human THP-1 macrophages, which can be reversed by exogenous S1P or FTY720 [80]. This means a promotive effect for S1P in the phagocytic activity of macrophages. On the contrary, stimuli like smoke exposure increased the SPNS2 expression of alveolar macrophages, and an impaired efferocytosis of macrophages seems to be associated with the upregulation of the S1P5 [81,82,83,84]. So, the in vivo models showed a dramatic opposite effect of S1P and S1PRs on the phagocytic ability of macrophages, which indicated a significant context-associated impact of S1P and S1PRs profile in inflammation.
In conclusion, the S1P axis participates in macrophages’ longevity, migration, phagocytic activity, etc., suggesting a pharmacologic potential in targeting macrophage-associated abnormalities of endometriosis.
T cell
Studies showed that T cells also participate in the formation and progression of endometriosis [85]. T lymphocytes are derived from the bone marrow and developed in the thymus. They are classified into several subtypes. The glycoproteins CD4 and CD8 expressed on the cell surface, which function as co-receptors for MHC class II and class I molecules, respectively, are the primary markers for the two groups. The CD4+ T cells can be further classified into Th1, Th2, Th9, Th17, and Treg [86].
It was reported that there is a higher CD4/CD8 ratio in patients with endometriosis, and the T lymphocytes showed an increased concentration of all subsets in the PF of the patients [87]. Compared to the eutopic endometrium, the endometriotic lesions have a higher concentration of T lymphocytes. However, there is no significant difference in the peripheral blood [85]. And Th17 is one of them, which derives from naïve CD4+ T cells under the activation of transforming growth factor alpha (TGFα), IL-6, and IL-23 [88]. Intriguingly, the abundance of Th17 cells is higher in the PF of women with severe endometriosis than those with mild ones [89]. The regulatory T cells (Treg), another important subset of the T lymphocytes, are potent suppressors of inflammatory immune responses and are responsible for maintaining antigen-specific T-cell tolerance and immune homeostasis. The expression of the forkhead box P3 transcription factor (Foxp3+) is the marker of Tregs. The production of IL-10, TGF-β, and other anti-inflammatory cytokines by Tregs are weapons to suppress T helper cells. It was shown that endometriotic lesions have the accumulation of CD4+/FoxP3+ Tregs [90]. And for patients with endometriosis, the Treg percentage is decreased in the peripheral blood but increased in the PF [91]. Additionally, it was demonstrated that the proportion of both subgroups of Tregs, CD45RA+Foxplow resting Tregs and CD45RA−Foxphigh effector Tregs, have an increased concentration in the peritoneal fluid, while no difference in the peripheral blood, which suggests a local accumulation within endometriotic lesions [92].
The S1P signaling mediates matured T cells exit from the thymus and activated T cells exit from the lymph nodes [93, 94]. The first US Food and Drug Administration (FDA)-approved drug FTY720 for the treatment of multiple sclerosis is based on the immune-regulating function, and the next-generation S1PRs-mediators have been promising in a series of diseases, including inflammatory bowel disease (IBD), systemic lupus erythematosus (SLE), and pulmonary conditions like asthma, fibrosis, and even COVID-19. [95]. The main mechanism of these drugs is believed to prevent activated T cells from exiting the lymphoid organs and infiltrating into inflammatory sites.
T cells develop in the thymus, and once they mature, they would exit the thymus by expressing S1P1 to sense the gradient of S1P between tissue and lymphatic fluid and blood [96]. Different S1PRs have been illustrated to affect different lymphocyte biology, including migration and determination of T cell phenotypes. And the expression of S1P1 on the lymphocyte surface is downregulated in the higher concentration of S1P (blood), upregulated in the lower concentration of S1P (lymphoid organs), and downregulated again in the relatively high concentration (lymphatic fluid). This concentration-associated regulation of S1P1 in circulating lymphocytes is important to set up their lymphoid organ transit time [97]. For T cells, exiting S1P1 signaling competes with CCR7 and CXCR4, which are retention signals [98, 99]. Treatment of pertussis toxin to block the Gαi signaling of T cells relieved the S1P1-deficient lymphocytes exiting from lymph nodes, indicating that the T cells have an inherent tendency to egress [97]. It was reported that under the stimuli of IFN-α/β, T cells would induce CD69 to form a complex with S1P1 to downregulate S1P1 and cause lymphocyte retention in lymphoid organs [100]. In contrast, the T cell recruitment was inhibited through S1P/S1P2 axis in mast cell-dependent acute allergic responses in the early airway of mice [101].
The increased S1P present in the inflammatory site induces T cell retention. The release of S1P by endothelial cells under the activation of a B cell peptide PEPITEM inhibited T cell recruitment, implying that an S1P concentration gradient between inflamed tissue and blood is important for T cell migration [102]. The T cell retention is mainly induced through agonism of S1P1, which inhibits the egress of inflammatory site T cells into the afferent lymph in inflammatory conditions. It was mediated partially by interactions of the integrin VLA-4 with its ligand VCAM-1 and of the integrin LFA-1 with its ligand ICAM-1 of polarized T cells at the basal surface of lymph but not blood vessel endothelium, ensuring the increased S1P present in inflammatory site to unidirectionally recruit T cells and suppress T cells egress [103]. It was reported that heterotrimeric guanine nucleotide-binding protein-coupled receptor kinase-2 (GRK2) functions in blunting S1P1 on blood-exposed lymphocytes, resulting in resisting the attraction by blood S1P [104]. It was demonstrated that without the expression of the transcription factor KLF2 and its target gene S1pr1 in CD8+-resident memory T cells allows them to stay in the peripheral tissue [105]. And CD69 also takes part in the regulation of T cell retention and local memory formation through S1P-S1P1 signaling [106]. It is conceivable that the activity of S1P lyase can regulate S1P gradient-mediated lymphocyte trafficking, which is a possible target [107].
Thus, the modulation of T cell recruitment in endometriotic lesions through the S1P axis might be a brand new direction of the T cell manipulation treatment for endometriosis.
S1P in the Endometriotic Fibrosis
As mentioned above, classical endometriotic lesions are defined as endometrial-like tissue containing glands and stroma. Still, a recent re-evaluation of the disease definition suggests that fibrosis and smooth muscle cells are more consistent features of lesions [108]. Fibrosis is a significant part of all kinds of endometriosis development. Classically, endometriotic lesions would go through a process from a soft, active, and vascularized red lesion to a hard black one because of hemoglobinolysis and fibrotic component accumulation. And then, as the fibrosis process progresses, the harder areas of white opacification and yellow–brown lesions show up, and they are considered latent stages of endometriosis [109] (Fig. 3).

The S1P axis participates in the fibrosis of endometriotic lesions. A–D The process of endometriotic lesion fibrosis from implantation, initiation, contraction, and consolidation. E The S1P axis in the process of fibroblast-to-myofibroblast transdifferentiation (FMT). FTY720-P, FTY720-phosphate; S1P2/3, S1P receptor 2/3; PI3K, phosphoinositide-3 kinase; ERK1/2, extracellular regulated protein kinase; SphK1, sphingosine kinase 1; ECM, extracellular matrix; α-SMA, α-smooth muscle actin
The pathophysiology of fibrosis follows a routine as below. Priming is the infiltration of various immune cells, such as macrophages, dendritic cells, lymphocytes, monocytes, and mast cells, into the endometriotic lesion [85]. These immune cells are attracted by chemokines and then activated by various factors like cytokines and reactive oxygen species (ROS), which induce continuing, non-resolving inflammation [110]. The next stage is the conversion and transdifferentiation into a myofibroblast [111] phenotype, which is characterized by the acquisition of expression of α-smooth muscle actin (α-SMA), a smooth muscle protein. Myofibroblasts produce and secrete an amount of extracellular matrix (ECM) and play a vital role in wound healing and tissue remodeling and the progression of fibrotic diseases [111]. Then comes a large number of ECM proteins that are synthesized, deposited, and modified to resist proteolytic degradation. That is an early stage of fibrosis, which theoretically can be treated to reverse the process of fibrosis. There are at least five sources of myofibroblasts in endometriotic lesion fibrosis, such as differentiation from pericytes, activation from interstitial fibroblasts, recruitment of fibrocytes, and from other cells by mechanisms collectively known as the epithelial-to-mesenchymal transition (EMT) and fibroblast-to-myofibroblast transdifferentiation (FMT) [112]. Solid evidence indicates that TGF-β1/Smad signaling is one of the key pathways in endometriotic lesion fibrosis and participates in the whole process of fibrogenesis because it not only mediates EMT but also modulates the production and degradation of ECM [111].
Emerging evidence indicates that the SphK1/S1P/S1PRs axis participates in fibrogenesis, such as liver fibrosis, cardiac fibrosis, pulmonary fibrosis, and renal fibrosis. It was reported that the S1P concentration, SK1, SPNS2, and S1P2 expression level have a positive correlation with the fibrosis extent [113, 114]. Genetical deletion of the S1PR2 mouse model showed an ameliorated liver fibrosis of the CCl4-induced model [115]. And the BMDM, which S1P could attract to the injured liver, performed as an aggravating factor in the fibrotic response [116, 117]. S1P1 expressed by hepatic sinusoidal endothelial cells regulates liver regeneration after partial hepatectomy or bile duct ligation through activation of ApoM+HDL-bound S1P. Apom-deficient mice have impaired hepatic sinusoidal vascular remodeling and recovery of hepatic tissue function, characterized by perivascular fibrosis and thrombosis. In contrast, Apom-transgenic mice exhibited minimal fibrosis through endothelial cell S1P1 function in the hepatic sinusoids, which means S1P has a positive role in alleviating liver fibrosis through S1P1 expressed on the endothelial cells of the special vascular niche of the liver [118]. On the other hand, to myocardial fibrosis, the SphK1 overexpressed mouse model showed a nearly 20-fold increase in enzymatic activity, developed detectable cardiac pathological fibrosis in the heart through Smad signaling, and it could be inhibited by Rho family G protein inhibitor or S1pr3 deletion [119]. And in normal lung fibroblast cell lines, FTY720-P induced the synthesis of ECM through the activation of S1P2 and S1P3 in vitro [120]. Conversely, FTY720-P suppressed the migration of bone marrow-derived mesenchymal stem cells both to the circulation and to the liver, and this effect had an ameliorating effect on liver fibrosis [121]. In diabetic nephropathy, S1P induced the EMT from tubular epithelial cells to myofibroblast via activating S1P2 and the downstream Rho kinases, which is associated with the development of renal fibrosis [122]. Furthermore, the expression of fibrotic markers was induced in a normal rat kidney fibroblasts cell line by S1P stimulation, including α-SMA, and its effect could be attenuated by SphK1 inhibitor, N, N-dimethylsphingosine [123].
Notably, SphK1 has already been found to be overexpressed in the endometriotic lesion (endometrial cyst of the ovary and peritoneal endometriosis), and the S1P concentration was hundreds of times higher than in control [45]. The TGF-β1-Smad signaling pathway is one of the most important mediators of endometriotic lesion fibrosis [111, 124, 125]. And the SphK1-S1P-S1PRs axis has been found to be closely associated with the TGF-β1 signaling pathway in many different diseases and fibrogenesis-associated states. Studies showed that S1P could activate the Smad signaling, independent of the TGF-β1, to contribute to myofibroblast differentiation of keratinocytes [126]. Accordingly, FTY720-P induced transdifferentiation of fibroblasts to myofibroblasts via S1P3 activation, which required Smad signaling but without TGF-β1 activation [127]. On the other hand, in the study mentioned above that used a normal human lung fibroblasts cell line, FTY720-P was reported to be capable of inducing ECM synthesis via S1P2/3, PI3K/Akt, and ERK1/2 signaling activation but could not induce the expression of α-SMA, which means FTY720-P cannot directly promote EMT like TGF-β1/Smad signaling in vitro [120]. Proto-myofibroblasts are defined as an intermediate stage between fibroblasts and myofibroblasts, and ECM is synthesized in proto-myofibroblasts without expressing α-SMA in the cells [128]. Thus, maybe the SphK1-S1P-S1PRs axis not only takes part in the transdifferentiation of fibroblasts to myofibroblasts but also stimulates ECM synthesis in both stages of proto-myofibroblasts and myofibroblasts, or the stimulation of transdifferentiation is microenvironment-dependent.
Recently, a study suggested that the SphK1-S1P-S1PRs axis mediates endometriotic lesion fibrosis induced by TGF-β1 [129]. It was demonstrated that SphK1 and calcium- and integrin-binding protein 1 (CIB1), an activating protein of SphK1 which helps SphK1 to translocate to the plasma membrane to wield its function [130], were both significantly upregulated in the endometriosis with the overexpression of EMT and fibrotic markers including α-SMA, and S1P3 was the most dominantly expressed one of all S1PRs. In vitro experiments suggested that suppressing SphK1, SphK2, and S1P2, S1P3 by siRNA could suppress the EMT and fibrotic markers expression induced by TGF-β1 [129]. As mentioned above, the SphK1-S1P-S1PRs axis contributes to the downstream TGF-β1 signaling. In two different fibrosis models, TGF-β1 upregulated the expression of SphK1 and activated S1P3 to induce the transdifferentiation of precursor cells into myofibroblasts [131, 132]. SphK1 transcription may be regulated by TGF-β1 via Smad signaling or connective tissue growth factor signaling. It is worth noting that the Rho-Rho kinase signaling pathway participates in myofibroblast differentiation, and activation of S1P2 and S1P3 can stimulate the Rho-Rho kinase signaling pathway [133, 134]. These reports support a complex interaction between the SphK1-S1P-S1PRs axis and TGF-β1/Smad signaling in endometriotic lesion fibrosis. Of course, the activation of fibrogenic signaling pathways and the immune responses are indispensable for the fibrosis process. As mentioned above in the inflammation section, for example, the S1P axis promotes the infiltration and activation of macrophages, which contributes to the transdifferentiation of myofibroblasts by secreting TGF-β1 [135].
In conclusion, fibrosis is a necessary, even defining process of endometriosis, which leads to tissue adhesions, scarring, and anatomical alterations associated with a series of symptoms. To date, effective treatment for fibrosis is still lacking, especially when fibrous tissue is formed. Thus, to prevent or slow down the process, modulating the S1P axis is a feasible way to achieve this end in endometriosis.
S1P in the Endometriotic Angiogenesis
Endometriosis is a benign disease that holds a series of malignant characteristics, including apoptosis resistance, metastasis, invasion, and angiogenesis [136]. Angiogenesis is defined as the growth of new blood vessels. The newly developed endometriotic lesion has a highly activated angiogenesis process that endows it with a red appearance. It is primarily believed that numerous vasculatures could bring ectopic lesions enough nutrients and oxygen supplies. Physiologically, angiogenesis is highly regulated in endometrial growth and remodeling in the female reproductive tract [137]. Under estrogen and progesterone control, endometrial blood vessels routinely grow and regress through the menstrual cycle. Modulation of endometrial angiogenesis is complicated and involves a variety of biologically active factors. Reports demonstrated that the role of estrogen in endometrial angiogenesis was context-dependent [138,139,140,141,142], and amounts of angiogenic factors and inhibitors are concerned in the endometrium. Vascular endothelial growth factor-A (VEGF-A) is believed to be one of the most pivotal angiogenic growth factors in the endometrium, and VEGFR-1, VEGFR-2, and neuropilin-1 (NRP-1) are its receptors. Several cytokines, including platelet-derived endothelial cell growth factor (PDGF), fibroblast growth factor (FGF), tumor necrosis factor-α (TNF-α), hepatocyte growth factor (HGF), and prokineticins (PK) are also reported to be facilitators of the angiogenic process [143, 144]. Women with endometriosis show an increased expression of VEGF-A, angiopoietin-1 (Ang-1), and Ang-2 and their receptors VEGFR-2 and Tie2 in the eutopic endometrium [145,146,147,148,149,150,151,152]. Thus, these studies suggested that patients with endometriosis have a more significant angiogenic potential in the eutopic endometrium compared to women without it, which allows the deciduous endometrial fragments to get enough nutrients from more vessels when they arrive at the peritoneum.
And for endometriotic lesions, VEGF-A is strongly expressed within it [147, 148, 150, 153,154,155]. Compared to older black or white scarred lesions, VEGF-A and other angiogenic cytokines were expressed higher in early-stage, red, and vascular, peritoneal endometriotic lesions [147, 153]. Furthermore, the primary receptor of VEGF-A, VEGFR-2, had a higher expression with consequential higher vascularization and proliferative activity in the red lesions [150, 155].
A large number of studies showed that the S1P signaling pathway plays a vital role in vascular development, and the description of a hypoxia-induced proangiogenic factor to S1P would be oversimplifying its complicated role in vascular growth and homeostasis [156]. The specific receptor subtype primarily involved, and the carrier protein that takes the lipid to the target receptor would determine the quantity and integrity of the vessels formed under the stimulation of S1P [157]. Additionally, S1P1 is able to sense fluid shear stress and be activated by it even without S1P [158]. It is worth noting that among five S1P receptors, only S1P1-3 expression was detected on endothelial cells [158, 159].
It was first found by Liu et al. that S1pr1−/− and Sphk1−/−Sphk2−/− mice were embryonic lethal due to vascular defect and severe hemorrhage [160]. While single S1pr2−/− or S1pr3−/− did not show any vascular developmental defects, S1pr2/3 double knockout mice had a reduced litter size and could not survive infancy for most of them, and S1pr1/2/3 triple-knockout mice were found to have an earlier and more severe hemorrhage than S1pr1−/− mice, which means S1P2 and S1P3 are also functioning in vascular development [161, 162]. Observation of endothelial-specific S1pr1−/− mice showed that the lack of S1P1 induced excessive sprouting of neovessels, and the activation of endothelial S1P1 suppressed the excessive sprouting and maintained vascular barrier functions to consolidate the neovessels by the blood full of S1P [163,164,165]. The cross-species evolutionary conservation is suggested with the same hypersprouting phenotype observed in S1pr1−/− zebrafish in several studies [163, 164, 166, 167].
Angiogenesis is primarily induced by hypoxia which is predominantly mediated by the oxygen-dependent transcriptional activators, and hypoxia-inducible factors (HIFs), and their expression increases in hypoxic cells in response to the hypoxic environment. S1P was released to the medium under the stimulation of hypoxia in an in vitro model of glioma cells through the upregulation of SphK1, which can be abolished by the knockdown of HIF-2α, and HIF-2α could bind to the SphK1 promoter [168]. And in adenocarcinoma cells, hypoxia-induced SphK2 expression and S1P release [169]. Intriguingly, SphK1, SPNS2, or S1P1 siRNA was found to abolish the HIF-2α upregulation in two renal cancer cell lines, suggesting that SphK1/SPNS2/S1P1 (S1P “inside-out” signaling) signaling axis can be the stimulator to HIF-2α expression too [170]. However, in another renal cancer cell line, just as in the glioma cells, HIF-2α siRNA suppressed the upregulated SphK1 expression, indicating that HIF-2α is an upstream regulator of SphK1 as also shown in glioma cells [171], which means there is a positive feedback loop between S1P and HIF-2α. And for HIF-1α, S1P2 signaling was shown to positively regulate its expression by stabilizing the protein [172].
The most prominent angiogenic promoter, VEGF, is primarily stimulated by hypoxia and mediated by HIF [173]. Unlike the S1pr1 deletion resulted in the hypersprouting phenotype, significantly reduced endothelial sprouting and vessel density were observed in the Vegfr2 deletion model [174]. VE-cadherin is very important to endothelial cell adhesive. It was reported that the VEGF signal induced VE-cadherin internalization from the plasma membrane [174], while the S1P1 activation promoted the stabilization of VE-cadherin localized at endothelial junctions [159]. All these suggest a functional antagonism between S1P1 and VEGFR2 signaling in the barrier function of vasculature through VE-cadherin localization and endothelial junctional stability. In conclusion, VEGFR2 signaling takes control of the early phase of the angiogenic process to sprout premature leaky vasculature, and S1P1 signaling is responsible for stabilizing the neovessels to mature in the later phase. And this explained the suppression of S1P1 by siRNA reduced tumor angiogenesis and vascular maturation to cause less primary tumor growth in the implantation of tumor cells [175]. Additionally, neutralization of S1P by monoclonal antibodies inhibited the proangiogenic effect of VEGF and reduced tumor progression in multiple cancer cell lines upon multiple murine models [176]. A proangiogenic role of S1P signaling is strongly suggested by all experiments in multiple systems, but S1PRs are widely expressed in most cell types, and which S1PR is expressed by what cells or what combination of them were determinants to angiogenesis remains to be elucidated. In contrast to S1P1, S1P2 receptors play an angiogenetic role that resembles that of VEGF, causing a hypersprouting phenotype in response to hypoxia [177]. In the retinal endothelium, hypoxia upregulated S1P2 receptor expression and led to the formation of blood vessel sprouts with leaky basement membranes and limited perfusion [178]. The blood vessel sprouts were competent enough to have a comparable blood flow to the mature vasculature in S1pr2-knockout mice [178]. But, the S1P2 inhibitor, JTE013 administration potentiated S1P-mediated angiogenesis in vivo in the Matrigel implant assay of mice, so the effect of S1P2 on angiogenesis may vary in a different context [179]. The balance between S1P1 and S1P2 activation determines the terminal effect of S1P on vascular endothelium (Fig. 4a).

The mechanism of the S1P axis regulates angiogenesis, innervation, and hyperalgesia. A The hypoxia within the endometriotic lesion can induce the expression of hypoxia-induced factor (HIF), then the downstream upregulation of vascular endothelial growth factor (VEGF) and sphingosine 1-phosphate (S1P) to form a VEGF and S1P gradient. The VEGF alone can induce multiple immature sprouting of neovessels, but only with S1P can consolidate the neovessels to become mature functional vessels. B The role of the S1P axis in the innervation and mechanism of hyperalgesia. S1P1, S1P receptor 1; VE-cadherin, vascular endothelial-cadherin; TNF-α, tumor necrosis factor-α; TNFR, tumor necrosis factor receptor; IL-1β, interleukin-1β; IL-1R, interleukin-1 receptor; NGF, nerve growth factor; SMase, sphingomyelinase; p-MAPK, phosphate- mitogen-activated protein kinase; p-p38, phosphate-protein 38; NF-κB, nuclear factor kappa B; NADPH, nicotinamide adenine dinucleotide phosphate; NOS, nitric oxide synthase; COX2, cyclo-oxygenase 2; NO, nitric oxide; PGE2, prostaglandin E2
As for the endometriosis research, Rudzitis-Auth et al. demonstrated recently that SKI-5C, a SphK1 inhibitor, could notably inhibit the development and vascularization of peritoneal lesions. A significantly smaller lesion size, lower functional microvessel density, smaller microvessel diameters, and reduced blood perfusion of the newly developing microvascular networks were observed in SKI-5C-treated mice [180].
In conclusion, angiogenesis is a vital part of endometriotic lesion development which has been repeatedly proven as a targetable process for endometriosis. The SphK1 inhibitor, SKI-5C-treated mouse model, demonstrated an obvious suppression of vascularization of endometriotic lesions. And it is noteworthy that the treatment did not affect the microvessel density of the ovaries and uterine horns [180]. This means the S1P axis is a promising target for endometriosis angiogenesis.
S1P in the Pain of Endometriosis
Endometriosis is a chronic disease. Chronic pain is one of the central problems of patients. Dysmenorrhea, cyclic dysuria, cyclic or acyclic lower abdominal pain, dyspareunia, dyschezia, and infertility are the typical complaints of endometriosis patients. The pain-related symptom is closely associated with inflammation, and the neuro-immune crosstalk plays a vital role in the inflammation and pain signal conduction. For example, allodynia, a phenomenon known as peripheral sensitization, makes innocuous mechanical or thermal stimuli induce pain. And hyperalgesia, which makes pain sensation associated with noxious stimulation causes drastically increased pain. They are both caused by the sensory threshold lowered by inflammation [181].
Nerve growth factor (NGF), a main neurotrophic factor released at the inflammatory site [182], is critical for the initiation and maintenance of both mechanical and thermal hypersensitivity [183]. Immunohistochemical staining of endometriotic specimens found the NGF expression in the endometriotic interstitium [184], and there is a significant increase of NGF expression in invasive lesions compared to noninvasive ones [185]. Inflammatory cytokines, including TNF-α and IL-1β, can regulate the NGF expression. Activating mast cell degranulation and cytokine production is a way to cause persistent inflammatory pain by this neurotrophin [186]. Khodorova et al. proved that NGF could activate neutral SMase to increase ceramide and S1P production via the p75 neurotrophin receptor (p75NTR), which is key signaling of mechanical allodynia [187].
The concentration of TNF-α is higher in PF and serum of endometriosis patients, especially in the early stages of the disease [188, 189]. It is worth noting that the SMase activation and resultant ceramide production can be modulated by TNF through binding to TNFR1, which causes pain and caspase-dependent apoptosis [190]. Intradermal injection of ceramide was found to induce a dose-dependent thermal and mechanical hyperalgesia in rats, mediated by the S1P/S1P1 axis [191]. S1P1 stimulation was reported to induce peroxynitrite formation through local activation of NADPH oxidase and nitric oxide synthase, which is a potent nitroxidative species implicated in many pain states [192]. Moreover, NF-κB- and MAPK-dependent COX2 pathway is also taking part in pain caused by increasing prostaglandin E2 (PGE2) production in the same model [193]. The concentration of PGE2 and PGF2α is long known to have a high level in endometriosis patients and is positively correlated with the severity of vaginal hyperalgesia and dysmenorrhea, respectively [194]. A recent study demonstrated that S1P stimulation induced a significant surge of COX2 mRNA expression in the peritoneal macrophage, which may contribute to the pain symptom [69].
The innervation within endometriotic lesions is also known as pain-related. The sympathetic, parasympathetic, and sensory nerve fiber development was repeatedly found in ectopic endometrium implants in humans and multiple rodent models [195, 196]. Many studies have demonstrated the sensory innervation and the abnormal secretion of different cytokines, which can mediate neurogenesis and subsequent peripheral neuroinflammation in endometriosis [197]. Aδ sensory, C sensory, cholinergic, and adrenergic nerves are those usually concerned in endometriosis, and the sensory nerve density within the lesion is increased [195, 198, 199]. A study showed that PF collected from endometriosis patients could induce increased sprouting of sensory neuritis from dorsal root ganglia (DRG) and decreased neurite outgrowth from sympathetic ganglia (DRG and sympathetic ganglia were dissected from the spinal cord of the Valo-specific pathogen-free egg embryos) [195]. It was proved that endometriosis patients have a high concentration of S1P both in the PF and cyst fluid [45]. And the S1P concentration and the receptor subtype that is mostly activated would alternatively affect the same population of sensory neurons. Another study showed that S1P3-mediated growth cone collapse and neurite retraction through Ras homolog gene family, member A (RhoA) activation, whereas S1P1 stimulation resulted in the elongation of neuronal processes [200]. It was also proved that the way S1P/S1P1 pathway affects pain transduction acts upon immune cells concentration, such as neutrophil infiltration, and activated CD4+ T cell persistence [201, 202]. Moreover, activation of S1P/S1P1 signaling in macrophages induced IL-6 production, which is also observed in endometriotic stromal cells (ESC), and the production of IL-6 upregulated S1P1 expression on cell membranes and vice versa to form a feed-forward cycle, contributing to the persistence of chronic inflammation and pain [45, 203].
The S1P axis plays a role in mediating peripheral sensitization too. Treating isolated sensory neurons with specific siRNAs targeted to individual S1PRs demonstrated that activation of S1P2, S1P4, or S1P5 is not sufficient to elicit significant neuronal firing [204, 205]. Thus, except for S1P1, S1P3 plays an important part in pain too [205]. S1P3 has emerged as a key contributor to S1P-induced peripheral sensitization [206]. S1pr3−/− mice showed loss of mechanical sensitivity, and intraplantar injection of TY-52156, an S1P3 selective antagonist, in normal mice caused dramatically decreased responsiveness to noxious stimulation [207], which seems at odds with the in vitro experiments (Fig. 4b).
In conclusion, pain is one of the most problematic symptoms of endometriosis patients. The innervation and peripheral nerve sensitization contribute to the exacerbation of the pain symptom, and that is the part S1P axis has a high correlation. Except for the inflammation-associated signaling pathway, studies also showed the regulation of nerve fiber infiltrating and retraction by the S1P axis. Unfortunately, up to the present, there is still no research exploring the exact impact of the S1P axis on endometriosis-associated pain symptoms, which is an imperative part of the mechanism resolving.
Conclusion and Future Outlook
Endometriosis is a problematic situation for both the patients and the physicians. For patients, a series of symptoms is annoying. Long-time heavy menstruation leads to anemia. Pelvic pain (cyclical and non-cyclical), painful periods, painful sex, and pain on defecation and urination pain brings obstacles to living a normal life and are associated with fatigue and depression. Last but not the least, sub-/infertility is significantly higher in patients with endometriosis compared to the general female population, which is also a challenge for physicians [208]. Current treatments for endometriosis are divided into medical management, surgical removement of the lesion, and a combination of them. Medical management mainly includes hormone regulation and non-steroidal anti-inflammatory drugs (NSAIDs). The most routinely prescribed treatment to endometriosis patients in clinical practice is hormonal therapies, because of their effectiveness in pain symptom control [209]. However, on the one hand, patients have to keep taking pills to maintain relief because symptoms return once the treatments stop. On the other hand, none of the hormone treatments allow patients to conceive. For NSAIDs, they can only handle the pain symptoms without suppressing the progression of lesions, and the study suggested an ovulation inhibition possibility of continuous usage [210]. The benefit of surgical removement in improving pain is still controversial, and the cumulative probability of persistent endometriosis-related pain following surgical treatment may be as high as 40–50% in 5 years [211, 212]. Therefore, current treatments for endometriosis are far from satisfying.
As a disease characterized by periodic hemorrhage and chronic inflammation, the endometriotic lesions are proven to have a significantly increased S1P concentration and dysregulated S1PRs profile expression [44, 45]. Studies of endometriosis and other abnormalities pertaining to inflammation all have fibrosis and angiogenesis processes, showing druggable active S1P signaling. SphK inhibitors, S1P monoclonal antibodies, and S1PRs modulators are the mainly pharmacologic agents targeting the S1P axis at present. Because of the vital physiological role S1P plays in maintaining multiple systems’ homeostasis, the relatively intense treatments of SphKs inhibitors are mainly under the development of malignant diseases. And the overall pro-proliferative effect of S1P on most somatic cells is another reason. As for S1PRs modulators, the ubiquitous expression of S1PRs in every organ and system brings the first approved S1P modulator for multiple sclerosis, fingolimod, which acts through multiple receptors, the cardiovascular and ocular side effects due to the activation of S1PRs in non-target systems. Thus, the next generation of S1PRs modulators is trying to fine-tune receptor selectivity to focus on specific S1PR [95]. For example, suppression of S1P2 by JTE013, an antagonist for S1P2 was demonstrated to block the pro-proliferative effect induced by S1P administration to endometriotic stromal cells [58]. On the contrary, a thioglycolate-induced peritonitis model showed the inhibition of S1P2 increased the recruitment of macrophages into the inflammatory site [58], which is considered a promotion factor for endometriotic lesion growth. The highly active interaction between immune cells, endometriotic cells, and neurocytes in different stages of inflammation makes the same S1P renders different and even completely opposite effect. Therefore, a clear elucidation of the S1P axis from the synthesis to transportation, activation, signaling transduction, and degradation is very important for precise targeting. And the crosstalk of immune cells, endometriotic cells, and neurocytes emphasizes the indispensable role of in vivo models in further research. Finally, the S1P axis closely participates in the pregnancy process and fetus development [213], which is also a vital consideration in the usage of S1P axis-associated medicine.
Data availability
Not applicable.
Code Availability
Not applicable.
References
Taylor HS, Kotlyar AM, Flores VA. Endometriosis is a chronic systemic disease: clinical challenges and novel innovations. Lancet. 2021;397(10276):839–52. https://doi.org/10.1016/s0140-6736(21)00389-5.
Zondervan KT, Becker CM, Missmer SA. Endometriosis. N Engl J Med. 2020;382(13):1244–56. https://doi.org/10.1056/NEJMra1810764.
Simoens S, Dunselman G, Dirksen C, Hummelshoj L, Bokor A, Brandes I, et al. The burden of endometriosis: costs and quality of life of women with endometriosis and treated in referral centres. Hum Reprod. 2012;27(5):1292–9. https://doi.org/10.1093/humrep/des073.
Nnoaham KE, Hummelshoj L, Webster P, d’Hooghe T, de CiccoNardone F, de CiccoNardone C, et al. Reprint of: Impact of endometriosis on quality of life and work productivity: a multicenter study across ten countries. Fertil Steril. 2019;112(4 Suppl1):e137–52. https://doi.org/10.1016/j.fertnstert.2019.08.082.
Giudice LC. Clinical practice. Endometriosis N Engl J Med. 2010;362(25):2389–98. https://doi.org/10.1056/NEJMcp1000274.
Zondervan KT, Becker CM, Koga K, Missmer SA, Taylor RN, Viganò P. Endometriosis. Nat Rev Dis Primers. 2018;4(1):9. https://doi.org/10.1038/s41572-018-0008-5.
Balasch J, Creus M, Fábregues F, Carmona F, Ordi J, Martinez-Román S, et al. Visible and non-visible endometriosis at laparoscopy in fertile and infertile women and in patients with chronic pelvic pain: a prospective study. Hum Reprod. 1996;11(2):387–91. https://doi.org/10.1093/humrep/11.2.387.
Klein S, D’Hooghe T, Meuleman C, Dirksen C, Dunselman G, Simoens S. What is the societal burden of endometriosis-associated symptoms? a prospective Belgian study. Reprod Biomed Online. 2014;28(1):116–24. https://doi.org/10.1016/j.rbmo.2013.09.020.
Kobayashi H, Kawahara N, Ogawa K, Yoshimoto C. A relationship between endometriosis and obstetric complications. Reprod Sci. 2020;27(3):771–8. https://doi.org/10.1007/s43032-019-00118-0.
Saraswat L, Ayansina DT, Cooper KG, Bhattacharya S, Miligkos D, Horne AW, et al. Pregnancy outcomes in women with endometriosis: a national record linkage study. BJOG. 2017;124(3):444–52. https://doi.org/10.1111/1471-0528.13920.
Saunders PTK, Horne AW. Endometriosis: etiology, pathobiology, and therapeutic prospects. Cell. 2021;184(11):2807–24. https://doi.org/10.1016/j.cell.2021.04.041.
Al Kadri H, Hassan S, Al-Fozan HM, Hajeer A. Hormone therapy for endometriosis and surgical menopause. Cochrane Database Syst Rev 2009;(1):CD005997. https://doi.org/10.1002/14651858.CD005997.pub2.
Gallagher JS, Missmer SA, Hornstein MD, Laufer MR, Gordon CM, DiVasta AD. Long-term effects of gonadotropin-releasing hormone agonists and add-back in adolescent endometriosis. J Pediatr Adolesc Gynecol. 2018;31(4):376–81. https://doi.org/10.1016/j.jpag.2018.03.004.
Bedaiwy MA, Alfaraj S, Yong P, Casper R. New developments in the medical treatment of endometriosis. Fertil Steril. 2017;107(3):555–65. https://doi.org/10.1016/j.fertnstert.2016.12.025.
Wei Y, Liang Y, Lin H, Dai Y, Yao S. Autonomic nervous system and inflammation interaction in endometriosis-associated pain. J Neuroinflammation. 2020;17(1):80. https://doi.org/10.1186/s12974-020-01752-1.
Lahiri S, Futerman AH. The metabolism and function of sphingolipids and glycosphingolipids. Cell Mol Life Sci. 2007;64(17):2270–84. https://doi.org/10.1007/s00018-007-7076-0.
Cartier A, Hla T. Sphingosine 1-phosphate: lipid signaling in pathology and therapy. Science 2019;366(6463). https://doi.org/10.1126/science.aar5551.
Huang WC, Nagahashi M, Terracina KP, Takabe K. Emerging role of sphingosine-1-phosphate in inflammation, cancer, and lymphangiogenesis. Biomolecules. 2013;3(3):408–34. https://doi.org/10.3390/biom3030408.
Kano K, Aoki J, Hla T. Lysophospholipid mediators in health and disease. Annu Rev Pathol. 2022;17:459–83. https://doi.org/10.1146/annurev-pathol-050420-025929.
Lander AD. Morpheus unbound: reimagining the morphogen gradient. Cell. 2007;128(2):245–56. https://doi.org/10.1016/j.cell.2007.01.004.
Pappu R, Schwab SR, Cornelissen I, Pereira JP, Regard JB, Xu Y, et al. Promotion of lymphocyte egress into blood and lymph by distinct sources of sphingosine-1-phosphate. Science. 2007;316(5822):295–8. https://doi.org/10.1126/science.1139221.
Selim S, Sunkara M, Salous AK, Leung SW, Berdyshev EV, Bailey A, et al. Plasma levels of sphingosine 1-phosphate are strongly correlated with haematocrit, but variably restored by red blood cell transfusions. Clin Sci (Lond). 2011;121(12):565–72. https://doi.org/10.1042/cs20110236.
Ito K, Anada Y, Tani M, Ikeda M, Sano T, Kihara A, et al. Lack of sphingosine 1-phosphate-degrading enzymes in erythrocytes. Biochem Biophys Res Commun. 2007;357(1):212–7. https://doi.org/10.1016/j.bbrc.2007.03.123.
Kobayashi N, Kawasaki-Nishi S, Otsuka M, Hisano Y, Yamaguchi A, Nishi T. MFSD2B is a sphingosine 1-phosphate transporter in erythroid cells. Sci Rep. 2018;8(1):4969. https://doi.org/10.1038/s41598-018-23300-x.
Vu TM, Ishizu AN, Foo JC, Toh XR, Zhang F, Whee DM, et al. Mfsd2b is essential for the sphingosine-1-phosphate export in erythrocytes and platelets. Nature. 2017;550(7677):524–8. https://doi.org/10.1038/nature24053.
Chandrakanthan M, Nguyen TQ, Hasan Z, Muralidharan S, Vu TM, Li AWL, et al. Deletion of Mfsd2b impairs thrombotic functions of platelets. Nat Commun. 2021;12(1):2286. https://doi.org/10.1038/s41467-021-22642-x.
Hisano Y, Kobayashi N, Yamaguchi A, Nishi T. Mouse SPNS2 functions as a sphingosine-1-phosphate transporter in vascular endothelial cells. PLoS One. 2012;7(6):e38941. https://doi.org/10.1371/journal.pone.0038941.
Nijnik A, Clare S, Hale C, Chen J, Raisen C, Mottram L, et al. The role of sphingosine-1-phosphate transporter Spns2 in immune system function. J Immunol. 2012;189(1):102–11. https://doi.org/10.4049/jimmunol.1200282.
Fukuhara S, Simmons S, Kawamura S, Inoue A, Orba Y, Tokudome T, et al. The sphingosine-1-phosphate transporter Spns2 expressed on endothelial cells regulates lymphocyte trafficking in mice. J Clin Invest. 2012;122(4):1416–26. https://doi.org/10.1172/jci60746.
Nagahashi M, Kim EY, Yamada A, Ramachandran S, Allegood JC, Hait NC, et al. Spns2, a transporter of phosphorylated sphingoid bases, regulates their blood and lymph levels, and the lymphatic network. Faseb j. 2013;27(3):1001–11. https://doi.org/10.1096/fj.12-219618.
Mendoza A, Bréart B, Ramos-Perez WD, Pitt LA, Gobert M, Sunkara M, et al. The transporter Spns2 is required for secretion of lymph but not plasma sphingosine-1-phosphate. Cell Rep. 2012;2(5):1104–10. https://doi.org/10.1016/j.celrep.2012.09.021.
Christoffersen C, Obinata H, Kumaraswamy SB, Galvani S, Ahnström J, Sevvana M, et al. Endothelium-protective sphingosine-1-phosphate provided by HDL-associated apolipoprotein M. Proc Natl Acad Sci U S A. 2011;108(23):9613–8. https://doi.org/10.1073/pnas.1103187108.
Fleming JK, Wojciak JM. Measuring sphingosine-1-phosphate/protein interactions with the kinetic exclusion assay. Methods Mol Biol. 2018;1697:1–8. https://doi.org/10.1007/7651_2017_5.
Yanagida K, Hla T. Vascular and immunobiology of the circulatory sphingosine 1-phosphate gradient. Annu Rev Physiol. 2017;79:67–91. https://doi.org/10.1146/annurev-physiol-021014-071635.
Pham TH, Baluk P, Xu Y, Grigorova I, Bankovich AJ, Pappu R, et al. Lymphatic endothelial cell sphingosine kinase activity is required for lymphocyte egress and lymphatic patterning. J Exp Med. 2010;207(1):17–27. https://doi.org/10.1084/jem.20091619.
Mendoza A, Fang V, Chen C, Serasinghe M, Verma A, Muller J, et al. Lymphatic endothelial S1P promotes mitochondrial function and survival in naive T cells. Nature. 2017;546(7656):158–61. https://doi.org/10.1038/nature22352.
Baeyens A, Bracero S, Chaluvadi VS, Khodadadi-Jamayran A, Cammer M, Schwab SR. Monocyte-derived S1P in the lymph node regulates immune responses. Nature. 2021;592(7853):290–5. https://doi.org/10.1038/s41586-021-03227-6.
Maceyka M, Spiegel S. Sphingolipid metabolites in inflammatory disease. Nature. 2014;510(7503):58–67. https://doi.org/10.1038/nature13475.
Cheong YC, Shelton JB, Laird SM, Richmond M, Kudesia G, Li TC, et al. IL-1, IL-6 and TNF-alpha concentrations in the peritoneal fluid of women with pelvic adhesions. Hum Reprod. 2002;17(1):69–75. https://doi.org/10.1093/humrep/17.1.69.
Forster R, Sarginson A, Velichkova A, Hogg C, Dorning A, Horne AW, et al. Macrophage-derived insulin-like growth factor-1 is a key neurotrophic and nerve-sensitizing factor in pain associated with endometriosis. Faseb j. 2019;33(10):11210–22. https://doi.org/10.1096/fj.201900797R.
Kato T, Yasuda K, Matsushita K, Ishii KJ, Hirota S, Yoshimoto T, et al. Interleukin-1/-33 signaling pathways as therapeutic targets for endometriosis. Front Immunol. 2019;10:2021. https://doi.org/10.3389/fimmu.2019.02021.
Guo M, Bafligil C, Tapmeier T, Hubbard C, Manek S, Shang C, et al. Mass cytometry analysis reveals a distinct immune environment in peritoneal fluid in endometriosis: a characterisation study. BMC Med. 2020;18(1):3. https://doi.org/10.1186/s12916-019-1470-y.
Xiao F, Liu X, Guo SW. Platelets and regulatory T cells may induce a type 2 immunity that is conducive to the progression and fibrogenesis of endometriosis. Front Immunol. 2020;11:610963. https://doi.org/10.3389/fimmu.2020.610963.
Santulli P, Marcellin L, Noël JC, Borghese B, Fayt I, Vaiman D, et al. Sphingosine pathway deregulation in endometriotic tissues. Fertil Steril. 2012;97(4):904–11. https://doi.org/10.1016/j.fertnstert.2011.12.051.
Yoshino O, Yamada-Nomoto K, Kano K, Ono Y, Kobayashi M, Ito M, et al. Sphingosine 1 phosphate (S1P) increased IL-6 expression and cell growth in endometriotic Cells. Reprod Sci. 2019;26(11):1460–7. https://doi.org/10.1177/1933719119828112.
Greaves E, Cousins FL, Murray A, Esnal-Zufiaurre A, Fassbender A, Horne AW, et al. A novel mouse model of endometriosis mimics human phenotype and reveals insights into the inflammatory contribution of shed endometrium. Am J Pathol. 2014;184(7):1930–9. https://doi.org/10.1016/j.ajpath.2014.03.011.
Weigert A, Olesch C, Brüne B. Sphingosine-1-phosphate and macrophage biology-how the sphinx tames the big eater. Front Immunol. 2019;10:1706. https://doi.org/10.3389/fimmu.2019.01706.
Weigert A, Weis N, Brüne B. Regulation of macrophage function by sphingosine-1-phosphate. Immunobiology. 2009;214(9–10):748–60. https://doi.org/10.1016/j.imbio.2009.06.003.
Weis N, Weigert A, von Knethen A, Brüne B. Heme oxygenase-1 contributes to an alternative macrophage activation profile induced by apoptotic cell supernatants. Mol Biol Cell. 2009;20(5):1280–8. https://doi.org/10.1091/mbc.e08-10-1005.
Gómez-Muñoz A, Kong J, Salh B, Steinbrecher UP. Sphingosine-1-phosphate inhibits acid sphingomyelinase and blocks apoptosis in macrophages. FEBS Lett. 2003;539(1–3):56–60. https://doi.org/10.1016/s0014-5793(03)00197-2.
Feuerborn R, Becker S, Potì F, Nagel P, Brodde M, Schmidt H, et al. High density lipoprotein (HDL)-associated sphingosine 1-phosphate (S1P) inhibits macrophage apoptosis by stimulating STAT3 activity and survivin expression. Atherosclerosis. 2017;257:29–37. https://doi.org/10.1016/j.atherosclerosis.2016.12.009.
Song F, Hou J, Chen Z, Cheng B, Lei R, Cui P, et al. Sphingosine-1-phosphate receptor 2 signaling promotes caspase-11-dependent macrophage pyroptosis and worsens Escherichia coli sepsis outcome. Anesthesiology. 2018;129(2):311–20. https://doi.org/10.1097/aln.0000000000002196.
Al-Jarallah A, Chen X, González L, Trigatti BL. High density lipoprotein stimulated migration of macrophages depends on the scavenger receptor class B, type I, PDZK1 and Akt1 and is blocked by sphingosine 1 phosphate receptor antagonists. PLoS One. 2014;9(9):e106487. https://doi.org/10.1371/journal.pone.0106487.
Gude DR, Alvarez SE, Paugh SW, Mitra P, Yu J, Griffiths R, et al. Apoptosis induces expression of sphingosine kinase 1 to release sphingosine-1-phosphate as a “come-and-get-me” signal. Faseb j. 2008;22(8):2629–38. https://doi.org/10.1096/fj.08-107169.
Liao CY, Song MJ, Gao Y, Mauer AS, Revzin A, Malhi H. Hepatocyte-derived lipotoxic extracellular vesicle sphingosine 1-phosphate induces macrophage chemotaxis. Front Immunol. 2018;9:2980. https://doi.org/10.3389/fimmu.2018.02980.
Weichand B, Weis N, Weigert A, Grossmann N, Levkau B, Brüne B. Apoptotic cells enhance sphingosine-1-phosphate receptor 1 dependent macrophage migration. Eur J Immunol. 2013;43(12):3306–13. https://doi.org/10.1002/eji.201343441.
Müller J, von Bernstorff W, Heidecke CD, Schulze T. Differential S1P receptor profiles on M1- and M2-polarized macrophages affect macrophage cytokine production and migration. Biomed Res Int. 2017;2017:7584621. https://doi.org/10.1155/2017/7584621.
Michaud J, Im DS, Hla T. Inhibitory role of sphingosine 1-phosphate receptor 2 in macrophage recruitment during inflammation. J Immunol. 2010;184(3):1475–83. https://doi.org/10.4049/jimmunol.0901586.
Ishii M, Kikuta J, Shimazu Y, Meier-Schellersheim M, Germain RN. Chemorepulsion by blood S1P regulates osteoclast precursor mobilization and bone remodeling in vivo. J Exp Med. 2010;207(13):2793–8. https://doi.org/10.1084/jem.20101474.
Bacci M, Capobianco A, Monno A, Cottone L, Di Puppo F, Camisa B, et al. Macrophages are alternatively activated in patients with endometriosis and required for growth and vascularization of lesions in a mouse model of disease. Am J Pathol. 2009;175(2):547–56. https://doi.org/10.2353/ajpath.2009.081011.
Smith KA, Pearson CB, Hachey AM, Xia DL, Wachtman LM. Alternative activation of macrophages in rhesus macaques (Macaca mulatta) with endometriosis. Comp Med. 2012;62(4):303–10.
Duan J, Liu X, Wang H, Guo SW. The M2a macrophage subset may be critically involved in the fibrogenesis of endometriosis in mice. Reprod Biomed Online. 2018;37(3):254–68. https://doi.org/10.1016/j.rbmo.2018.05.017.
Braun DP, Muriana A, Gebel H, Rotman C, Rana N, Dmowski WP. Monocyte-mediated enhancement of endometrial cell proliferation in women with endometriosis. Fertil Steril. 1994;61(1):78–84. https://doi.org/10.1016/s0015-0282(16)56456-5.
Lech M. Anders HJ (2013) Macrophages and fibrosis: how resident and infiltrating mononuclear phagocytes orchestrate all phases of tissue injury and repair. Biochim Biophys Acta. 1832;7:989–97. https://doi.org/10.1016/j.bbadis.2012.12.001.
Berbic M, Schulke L, Markham R, Tokushige N, Russell P, Fraser IS. Macrophage expression in endometrium of women with and without endometriosis. Hum Reprod. 2009;24(2):325–32. https://doi.org/10.1093/humrep/den393.
Takebayashi A, Kimura F, Kishi Y, Ishida M, Takahashi A, Yamanaka A, et al. Subpopulations of macrophages within eutopic endometrium of endometriosis patients. Am J Reprod Immunol. 2015;73(3):221–31. https://doi.org/10.1111/aji.12331.
Khan KN, Masuzaki H, Fujishita A, Kitajima M, Sekine I, Ishimaru T. Differential macrophage infiltration in early and advanced endometriosis and adjacent peritoneum. Fertil Steril. 2004;81(3):652–61. https://doi.org/10.1016/j.fertnstert.2003.07.037.
Das A, Segar CE, Hughley BB, Bowers DT, Botchwey EA. The promotion of mandibular defect healing by the targeting of S1P receptors and the recruitment of alternatively activated macrophages. Biomaterials. 2013;34(38):9853–62. https://doi.org/10.1016/j.biomaterials.2013.08.015.
Ono Y, Kawakita T, Yoshino O, Sato E, Kano K, Ohba M et al. Sphingosine 1-phosphate (S1P) in the peritoneal fluid skews M2 macrophage and contributes to the development of endometriosis. Biomedicines 2021;9(11). https://doi.org/10.3390/biomedicines9111519.
Heilmann A, Schinke T, Bindl R, Wehner T, Rapp A, Haffner-Luntzer M, et al. Systemic treatment with the sphingosine-1-phosphate analog FTY720 does not improve fracture healing in mice. J Orthop Res. 2013;31(11):1845–50. https://doi.org/10.1002/jor.22426.
Deng S, Zhou X, Ge Z, Song Y, Wang H, Liu X, et al. Exosomes from adipose-derived mesenchymal stem cells ameliorate cardiac damage after myocardial infarction by activating S1P/SK1/S1PR1 signaling and promoting macrophage M2 polarization. Int J Biochem Cell Biol. 2019;114:105564. https://doi.org/10.1016/j.biocel.2019.105564.
Yang J, Yang L, Tian L, Ji X, Yang L, Li L. Sphingosine 1-phosphate (S1P)/S1P Receptor2/3 axis promotes inflammatory M1 polarization of bone marrow-derived monocyte/macrophage via G(α)i/o/PI3K/JNK pathway. Cell Physiol Biochem. 2018;49(5):1677–93. https://doi.org/10.1159/000493611.
Hymel LA, Ogle ME, Anderson SE, San Emeterio CL, Turner TC, York WY, et al. Modulating local S1P receptor signaling as a regenerative immunotherapy after volumetric muscle loss injury. J Biomed Mater Res A. 2021;109(5):695–712. https://doi.org/10.1002/jbm.a.37053.
Schott J, Reitter S, Philipp J, Haneke K, Schäfer H, Stoecklin G. Translational regulation of specific mRNAs controls feedback inhibition and survival during macrophage activation. PLoS Genet. 2014;10(6):e1004368. https://doi.org/10.1371/journal.pgen.1004368.
Hayden MS, Ghosh S. Regulation of NF-κB by TNF family cytokines. Semin Immunol. 2014;26(3):253–66. https://doi.org/10.1016/j.smim.2014.05.004.
Alvarez SE, Harikumar KB, Hait NC, Allegood J, Strub GM, Kim EY, et al. Sphingosine-1-phosphate is a missing cofactor for the E3 ubiquitin ligase TRAF2. Nature. 2010;465(7301):1084–8. https://doi.org/10.1038/nature09128.
Xia P, Wang L, Moretti PA, Albanese N, Chai F, Pitson SM, et al. Sphingosine kinase interacts with TRAF2 and dissects tumor necrosis factor-alpha signaling. J Biol Chem. 2002;277(10):7996–8003. https://doi.org/10.1074/jbc.M111423200.
Králíčková M, Vetvicka V. Immunological aspects of endometriosis: a review. Ann Transl Med. 2015;3(11):153. https://doi.org/10.3978/j.issn.2305-5839.2015.06.08.
Luo B, Gan W, Liu Z, Shen Z, Wang J, Shi R, et al. Erythropoeitin signaling in macrophages promotes dying cell clearance and immune tolerance. Immunity. 2016;44(2):287–302. https://doi.org/10.1016/j.immuni.2016.01.002.
Tran HB, Barnawi J, Ween M, Hamon R, Roscioli E, Hodge G, et al. Cigarette smoke inhibits efferocytosis via deregulation of sphingosine kinase signaling: reversal with exogenous S1P and the S1P analogue FTY720. J Leukoc Biol. 2016;100(1):195–202. https://doi.org/10.1189/jlb.3A1015-471R.
Tran HB, Jersmann H, Truong TT, Hamon R, Roscioli E, Ween M, et al. Disrupted epithelial/macrophage crosstalk via Spinster homologue 2-mediated S1P signaling may drive defective macrophage phagocytic function in COPD. PLoS One. 2017;12(11):e0179577. https://doi.org/10.1371/journal.pone.0179577.
Barnawi J, Tran H, Jersmann H, Pitson S, Roscioli E, Hodge G, et al. Potential link between the sphingosine-1-phosphate (S1P) system and defective alveolar macrophage phagocytic function in chronic obstructive pulmonary disease (COPD). PLoS One. 2015;10(10):e0122771. https://doi.org/10.1371/journal.pone.0122771.
Barnawi J, Jersmann H, Haberberger R, Hodge S, Meech R. Reduced DNA methylation of sphingosine-1 phosphate receptor 5 in alveolar macrophages in COPD: a potential link to failed efferocytosis. Respirology. 2017;22(2):315–21. https://doi.org/10.1111/resp.12949.
Barnawi J, Tran HB, Roscioli E, Hodge G, Jersmann H, Haberberger R, et al. Pro-phagocytic effects of thymoquinone on cigarette smoke-exposed macrophages occur by modulation of the sphingosine-1-phosphate signalling system. COPD. 2016;13(5):653–61. https://doi.org/10.3109/15412555.2016.1153614.
Vallvé-Juanico J, Houshdaran S, Giudice LC. The endometrial immune environment of women with endometriosis. Hum Reprod Update. 2019;25(5):564–91. https://doi.org/10.1093/humupd/dmz018.
Zhou L, Chong MM, Littman DR. Plasticity of CD4+ T cell lineage differentiation. Immunity. 2009;30(5):646–55. https://doi.org/10.1016/j.immuni.2009.05.001.
Riccio L, Santulli P, Marcellin L, Abrão MS, Batteux F, Chapron C. Immunology of endometriosis. Best Pract Res Clin Obstet Gynaecol. 2018;50:39–49. https://doi.org/10.1016/j.bpobgyn.2018.01.010.
Takamura M, Koga K, Izumi G, Hirata T, Harada M, Hirota Y, et al. Simultaneous detection and evaluation of four subsets of CD4+ T lymphocyte in lesions and peripheral blood in endometriosis. Am J Reprod Immunol. 2015;74(6):480–6. https://doi.org/10.1111/aji.12426.
Gogacz M, Winkler I, Bojarska-Junak A, Tabarkiewicz J, Semczuk A, Rechberger T, et al. Increased percentage of Th17 cells in peritoneal fluid is associated with severity of endometriosis. J Reprod Immunol. 2016;117:39–44. https://doi.org/10.1016/j.jri.2016.04.289.
Maridas DE, Hey-Cunningham AJ, Ng CHM, Markham R, Fraser IS, Berbic M. Peripheral and endometrial dendritic cell populations during the normal cycle and in the presence of endometriosis. J Endometr Pelvic Pain Disord. 2014;6(2):67–119. https://doi.org/10.5301/je.5000180.
Olkowska-Truchanowicz J, Bocian K, Maksym RB, Białoszewska A, Włodarczyk D, Baranowski W, et al. CD4+ CD25+ FOXP3+ regulatory T cells in peripheral blood and peritoneal fluid of patients with endometriosis. Hum Reprod. 2013;28(1):119–24. https://doi.org/10.1093/humrep/des346.
Hanada T, Tsuji S, Nakayama M, Wakinoue S, Kasahara K, Kimura F, et al. Suppressive regulatory T cells and latent transforming growth factor-β-expressing macrophages are altered in the peritoneal fluid of patients with endometriosis. Reprod Biol Endocrinol. 2018;16(1):9. https://doi.org/10.1186/s12958-018-0325-2.
Matloubian M, Lo CG, Cinamon G, Lesneski MJ, Xu Y, Brinkmann V, et al. Lymphocyte egress from thymus and peripheral lymphoid organs is dependent on S1P receptor 1. Nature. 2004;427(6972):355–60. https://doi.org/10.1038/nature02284.
Allende ML, Dreier JL, Mandala S, Proia RL. Expression of the sphingosine 1-phosphate receptor, S1P1, on T-cells controls thymic emigration. J Biol Chem. 2004;279(15):15396–401. https://doi.org/10.1074/jbc.M314291200.
McGinley MP, Cohen JA. Sphingosine 1-phosphate receptor modulators in multiple sclerosis and other conditions. Lancet. 2021;398(10306):1184–94. https://doi.org/10.1016/s0140-6736(21)00244-0.
Rivera J, Proia RL, Olivera A. The alliance of sphingosine-1-phosphate and its receptors in immunity. Nat Rev Immunol. 2008;8(10):753–63. https://doi.org/10.1038/nri2400.
Lo CG, Xu Y, Proia RL, Cyster JG. Cyclical modulation of sphingosine-1-phosphate receptor 1 surface expression during lymphocyte recirculation and relationship to lymphoid organ transit. J Exp Med. 2005;201(2):291–301. https://doi.org/10.1084/jem.20041509.
Nakai A, Hayano Y, Furuta F, Noda M, Suzuki K. Control of lymphocyte egress from lymph nodes through β2-adrenergic receptors. J Exp Med. 2014;211(13):2583–98. https://doi.org/10.1084/jem.20141132.
Pham TH, Okada T, Matloubian M, Lo CG, Cyster JG. S1P1 receptor signaling overrides retention mediated by G alpha i-coupled receptors to promote T cell egress. Immunity. 2008;28(1):122–33. https://doi.org/10.1016/j.immuni.2007.11.017.
Shiow LR, Rosen DB, Brdicková N, Xu Y, An J, Lanier LL, et al. CD69 acts downstream of interferon-alpha/beta to inhibit S1P1 and lymphocyte egress from lymphoid organs. Nature. 2006;440(7083):540–4. https://doi.org/10.1038/nature04606.
Oskeritzian CA, Hait NC, Wedman P, Chumanevich A, Kolawole EM, Price MM, et al. The sphingosine-1-phosphate/sphingosine-1-phosphate receptor 2 axis regulates early airway T-cell infiltration in murine mast cell-dependent acute allergic responses. J Allergy Clin Immunol. 2015;135(4):1008-1018.e1001. https://doi.org/10.1016/j.jaci.2014.10.044.
Chimen M, McGettrick HM, Apta B, Kuravi SJ, Yates CM, Kennedy A, et al. Homeostatic regulation of T cell trafficking by a B cell-derived peptide is impaired in autoimmune and chronic inflammatory disease. Nat Med. 2015;21(5):467–75. https://doi.org/10.1038/nm.3842.
Ledgerwood LG, Lal G, Zhang N, Garin A, Esses SJ, Ginhoux F, et al. The sphingosine 1-phosphate receptor 1 causes tissue retention by inhibiting the entry of peripheral tissue T lymphocytes into afferent lymphatics. Nat Immunol. 2008;9(1):42–53. https://doi.org/10.1038/ni1534.
Arnon TI, Xu Y, Lo C, Pham T, An J, Coughlin S, et al. GRK2-dependent S1PR1 desensitization is required for lymphocytes to overcome their attraction to blood. Science. 2011;333(6051):1898–903. https://doi.org/10.1126/science.1208248.
Skon CN, Lee JY, Anderson KG, Masopust D, Hogquist KA, Jameson SC. Transcriptional downregulation of S1pr1 is required for the establishment of resident memory CD8+ T cells. Nat Immunol. 2013;14(12):1285–93. https://doi.org/10.1038/ni.2745.
Mackay LK, Braun A, Macleod BL, Collins N, Tebartz C, Bedoui S, et al. Cutting edge: CD69 interference with sphingosine-1-phosphate receptor function regulates peripheral T cell retention. J Immunol. 2015;194(5):2059–63. https://doi.org/10.4049/jimmunol.1402256.
Schwab SR, Pereira JP, Matloubian M, Xu Y, Huang Y, Cyster JG. Lymphocyte sequestration through S1P lyase inhibition and disruption of S1P gradients. Science. 2005;309(5741):1735–9. https://doi.org/10.1126/science.1113640.
Vigano P, Candiani M, Monno A, Giacomini E, Vercellini P, Somigliana E. Time to redefine endometriosis including its pro-fibrotic nature. Hum Reprod. 2018;33(3):347–52. https://doi.org/10.1093/humrep/dex354.
Nisolle M, Donnez J. Peritoneal endometriosis, ovarian endometriosis, and adenomyotic nodules of the rectovaginal septum are three different entities. Fertil Steril. 1997;68(4):585–96. https://doi.org/10.1016/s0015-0282(97)00191-x.
Machairiotis N, Vasilakaki S, Thomakos N. Inflammatory mediators and pain in endometriosis: a systematic review. Biomedicines 2021;9(1). https://doi.org/10.3390/biomedicines9010054.
Zhang Q, Duan J, Olson M, Fazleabas A, Guo SW. Cellular changes consistent with epithelial-mesenchymal transition and fibroblast-to-myofibroblast transdifferentiation in the progression of experimental endometriosis in baboons. Reprod Sci. 2016;23(10):1409–21. https://doi.org/10.1177/1933719116641763.
Pakshir P, Noskovicova N, Lodyga M, Son DO, Schuster R, Goodwin A et al. The myofibroblast at a glance. J Cell Sci. 2020;133(13). https://doi.org/10.1242/jcs.227900.
Sato M, Ikeda H, Uranbileg B, Kurano M, Saigusa D, Aoki J, et al. Sphingosine kinase-1, S1P transporter spinster homolog 2 and S1P2 mRNA expressions are increased in liver with advanced fibrosis in human. Sci Rep. 2016;6:32119. https://doi.org/10.1038/srep32119.
Yang L, Yue S, Yang L, Liu X, Han Z, Zhang Y, et al. Sphingosine kinase/sphingosine 1-phosphate (S1P)/S1P receptor axis is involved in liver fibrosis-associated angiogenesis. J Hepatol. 2013;59(1):114–23. https://doi.org/10.1016/j.jhep.2013.02.021.
Ikeda H, Watanabe N, Ishii I, Shimosawa T, Kume Y, Tomiya T, et al. Sphingosine 1-phosphate regulates regeneration and fibrosis after liver injury via sphingosine 1-phosphate receptor 2. J Lipid Res. 2009;50(3):556–64. https://doi.org/10.1194/jlr.M800496-JLR200.
Stelling A, Hashwah H, Bertram K, Manz MG, Tzankov A, Müller A. The tumor suppressive TGF-β/SMAD1/S1PR2 signaling axis is recurrently inactivated in diffuse large B-cell lymphoma. Blood. 2018;131(20):2235–46. https://doi.org/10.1182/blood-2017-10-810630.
Yang L, Han Z, Tian L, Mai P, Zhang Y, Wang L, et al. Sphingosine 1-phosphate receptor 2 and 3 mediate bone marrow-derived monocyte/macrophage motility in cholestatic liver injury in mice. Sci Rep. 2015;5:13423. https://doi.org/10.1038/srep13423.
Ding BS, Liu CH, Sun Y, Chen Y, Swendeman SL, Jung B, et al. HDL activation of endothelial sphingosine-1-phosphate receptor-1 (S1P(1)) promotes regeneration and suppresses fibrosis in the liver. JCI Insight. 2016;1(21):e87058. https://doi.org/10.1172/jci.insight.87058.
Takuwa N, Ohkura S, Takashima S, Ohtani K, Okamoto Y, Tanaka T, et al. S1P3-mediated cardiac fibrosis in sphingosine kinase 1 transgenic mice involves reactive oxygen species. Cardiovasc Res. 2010;85(3):484–93. https://doi.org/10.1093/cvr/cvp312.
Sobel K, Menyhart K, Killer N, Renault B, Bauer Y, Studer R, et al. Sphingosine 1-phosphate (S1P) receptor agonists mediate pro-fibrotic responses in normal human lung fibroblasts via S1P2 and S1P3 receptors and Smad-independent signaling. J Biol Chem. 2013;288(21):14839–51. https://doi.org/10.1074/jbc.M112.426726.
Kong Y, Wang H, Wang S, Tang N. FTY720, a sphingosine-1 phosphate receptor modulator, improves liver fibrosis in a mouse model by impairing the motility of bone marrow-derived mesenchymal stem cells. Inflammation. 2014;37(4):1326–36. https://doi.org/10.1007/s10753-014-9877-2.
Ishizawa S, Takahashi-Fujigasaki J, Kanazawa Y, Matoba K, Kawanami D, Yokota T, et al. Sphingosine-1-phosphate induces differentiation of cultured renal tubular epithelial cells under Rho kinase activation via the S1P2 receptor. Clin Exp Nephrol. 2014;18(6):844–52. https://doi.org/10.1007/s10157-014-0933-x.
Shiohira S, Yoshida T, Sugiura H, Nishida M, Nitta K, Tsuchiya K. Sphingosine-1-phosphate acts as a key molecule in the direct mediation of renal fibrosis. Physiol Rep. 2013;1(7):e00172. https://doi.org/10.1002/phy2.172.
Lin X, Dai Y, Xu W, Shi L, Jin X, Li C, et al. Hypoxia promotes ectopic adhesion ability of endometrial stromal cells via TGF-β1/Smad signaling in endometriosis. Endocrinology. 2018;159(4):1630–41. https://doi.org/10.1210/en.2017-03227.
Shi LB, Zhou F, Zhu HY, Huang D, Jin XY, Li C, et al. Transforming growth factor beta1 from endometriomas promotes fibrosis in surrounding ovarian tissues via Smad2/3 signaling. Biol Reprod. 2017;97(6):873–82. https://doi.org/10.1093/biolre/iox140.
Sauer B, Vogler R, von Wenckstern H, Fujii M, Anzano MB, Glick AB, et al. Involvement of Smad signaling in sphingosine 1-phosphate-mediated biological responses of keratinocytes. J Biol Chem. 2004;279(37):38471–9. https://doi.org/10.1074/jbc.M313557200.
Keller CD, Rivera Gil P, Tölle M, van der Giet M, Chun J, Radeke HH, et al. Immunomodulator FTY720 induces myofibroblast differentiation via the lysophospholipid receptor S1P3 and Smad3 signaling. Am J Pathol. 2007;170(1):281–92. https://doi.org/10.2353/ajpath.2007.060485.
Tomasek JJ, Gabbiani G, Hinz B, Chaponnier C, Brown RA. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat Rev Mol Cell Biol. 2002;3(5):349–63. https://doi.org/10.1038/nrm809.
Bernacchioni C, Capezzuoli T, Vannuzzi V, Malentacchi F, Castiglione F, Cencetti F, et al. Sphingosine 1-phosphate receptors are dysregulated in endometriosis: possible implication in transforming growth factor β-induced fibrosis. Fertil Steril. 2021;115(2):501–11. https://doi.org/10.1016/j.fertnstert.2020.08.012.
Jarman KE, Moretti PA, Zebol JR, Pitson SM. Translocation of sphingosine kinase 1 to the plasma membrane is mediated by calcium- and integrin-binding protein 1. J Biol Chem. 2010;285(1):483–92. https://doi.org/10.1074/jbc.M109.068395.
Yang L, Chang N, Liu X, Han Z, Zhu T, Li C, et al. Bone marrow-derived mesenchymal stem cells differentiate to hepatic myofibroblasts by transforming growth factor-β1 via sphingosine kinase/sphingosine 1-phosphate (S1P)/S1P receptor axis. Am J Pathol. 2012;181(1):85–97. https://doi.org/10.1016/j.ajpath.2012.03.014.
Cencetti F, Bernacchioni C, Nincheri P, Donati C, Bruni P. Transforming growth factor-beta1 induces transdifferentiation of myoblasts into myofibroblasts via up-regulation of sphingosine kinase-1/S1P3 axis. Mol Biol Cell. 2010;21(6):1111–24. https://doi.org/10.1091/mbc.e09-09-0812.
Bruno G, Cencetti F, Pertici I, Japtok L, Bernacchioni C, Donati C, et al. (2015) CTGF/CCN2 exerts profibrotic action in myoblasts via the up-regulation of sphingosine kinase-1/S1P3 signaling axis: implications in the action mechanism of TGFβ. Biochim Biophys Acta. 1851;2:194–202. https://doi.org/10.1016/j.bbalip.2014.11.011.
Kono Y, Nishiuma T, Nishimura Y, Kotani Y, Okada T, Nakamura S, et al. Sphingosine kinase 1 regulates differentiation of human and mouse lung fibroblasts mediated by TGF-beta1. Am J Respir Cell Mol Biol. 2007;37(4):395–404. https://doi.org/10.1165/rcmb.2007-0065OC.
An M, Li D, Yuan M, Li Q, Zhang L, Wang G. Interaction of macrophages and endometrial cells induces epithelial-mesenchymal transition-like processes in adenomyosis. Biol Reprod. 2017;96(1):46–57. https://doi.org/10.1095/biolreprod.116.144071.
Pollacco J, Sacco K, Portelli M, Schembri-Wismayer P, Calleja-Agius J. Molecular links between endometriosis and cancer. Gynecol Endocrinol. 2012;28(8):577–81. https://doi.org/10.3109/09513590.2011.650761.
Girling JE, Rogers PA. Recent advances in endometrial angiogenesis research. Angiogenesis. 2005;8(2):89–99. https://doi.org/10.1007/s10456-005-9006-9.
Iruela-Arispe ML, Rodriguez-Manzaneque JC, Abu-Jawdeh G. Endometrial endothelial cells express estrogen and progesterone receptors and exhibit a tissue specific response to angiogenic growth factors. Microcirculation. 1999;6(2):127–40.
Nayak NR, Brenner RM. Vascular proliferation and vascular endothelial growth factor expression in the rhesus macaque endometrium. J Clin Endocrinol Metab. 2002;87(4):1845–55. https://doi.org/10.1210/jcem.87.4.8413.
Kayisli UA, Luk J, Guzeloglu-Kayisli O, Seval Y, Demir R, Arici A. Regulation of angiogenic activity of human endometrial endothelial cells in culture by ovarian steroids. J Clin Endocrinol Metab. 2004;89(11):5794–802. https://doi.org/10.1210/jc.2003-030820.
Ma W, Tan J, Matsumoto H, Robert B, Abrahamson DR, Das SK, et al. Adult tissue angiogenesis: evidence for negative regulation by estrogen in the uterus. Mol Endocrinol. 2001;15(11):1983–92. https://doi.org/10.1210/mend.15.11.0734.
Heryanto B, Rogers PA. Regulation of endometrial endothelial cell proliferation by oestrogen and progesterone in the ovariectomized mouse. Reproduction. 2002;123(1):107–13. https://doi.org/10.1530/rep.0.1230107.
Surrey ES, Halme J. Effect of platelet-derived growth factor on endometrial stromal cell proliferation in vitro: a model for endometriosis? Fertil Steril. 1991;56(4):672–9. https://doi.org/10.1016/s0015-0282(16)54598-1.
Rudzitis-Auth J, Christoffel A, Menger MD, Laschke MW. Targeting sphingosine kinase 1 (SPHK1) with the small molecule inhibitor SKI-5C suppresses the development of endometriotic lesions. Br J Pharmacol. 2021. https://doi.org/10.1111/bph.15601.
Gilabert-Estellés J, Ramón LA, España F, Gilabert J, Vila V, Réganon E, et al. Expression of angiogenic factors in endometriosis: relationship to fibrinolytic and metalloproteinase systems. Hum Reprod. 2007;22(8):2120–7. https://doi.org/10.1093/humrep/dem149.
Khan KN, Masuzaki H, Fujishita A, Kitajima M, Sekine I, Ishimaru T. Immunoexpression of hepatocyte growth factor and c-Met receptor in the eutopic endometrium predicts the activity of ectopic endometrium. Fertil Steril. 2003;79(1):173–81. https://doi.org/10.1016/s0015-0282(02)04535-1.
Tan XJ, Lang JH, Liu DY, Shen K, Leng JH, Zhu L. Expression of vascular endothelial growth factor and thrombospondin-1 mRNA in patients with endometriosis. Fertil Steril. 2002;78(1):148–53. https://doi.org/10.1016/s0015-0282(02)03187-4.
Takehara M, Ueda M, Yamashita Y, Terai Y, Hung YC, Ueki M. Vascular endothelial growth factor A and C gene expression in endometriosis. Hum Pathol. 2004;35(11):1369–75. https://doi.org/10.1016/j.humpath.2004.07.020.
Burlev VA, Il’yasova NA, Dubinskaya ED. Proliferative activity of microvessels and angiogenesis in eutopic endometrium in patients with peritoneal endometriosis. Bull Exp Biol Med. 2005;139(6):727–31. https://doi.org/10.1007/s10517-005-0391-2.
Bourlev V, Volkov N, Pavlovitch S, Lets N, Larsson A, Olovsson M. The relationship between microvessel density, proliferative activity and expression of vascular endothelial growth factor-A and its receptors in eutopic endometrium and endometriotic lesions. Reproduction. 2006;132(3):501–9. https://doi.org/10.1530/rep.1.01110.
Hur SE, Lee JY, Moon HS, Chung HW. Angiopoietin-1, angiopoietin-2 and Tie-2 expression in eutopic endometrium in advanced endometriosis. Mol Hum Reprod. 2006;12(7):421–6. https://doi.org/10.1093/molehr/gal049.
Cosín R, Gilabert-Estellés J, Ramón LA, España F, Gilabert J, Romeu A, et al. Vascular endothelial growth factor polymorphisms (-460C/T, +405G/C, and 936C/T) and endometriosis: their influence on vascular endothelial growth factor expression. Fertil Steril. 2009;92(4):1214–20. https://doi.org/10.1016/j.fertnstert.2008.08.079.
Donnez J, Smoes P, Gillerot S, Casanas-Roux F, Nisolle M. Vascular endothelial growth factor (VEGF) in endometriosis. Hum Reprod. 1998;13(6):1686–90. https://doi.org/10.1093/humrep/13.6.1686.
McLaren J, Prentice A, Charnock-Jones DS, Millican SA, Müller KH, Sharkey AM, et al. Vascular endothelial growth factor is produced by peritoneal fluid macrophages in endometriosis and is regulated by ovarian steroids. J Clin Invest. 1996;98(2):482–9. https://doi.org/10.1172/jci118815.
Machado DE, Abrao MS, Berardo PT, Takiya CM, Nasciutti LE. Vascular density and distribution of vascular endothelial growth factor (VEGF) and its receptor VEGFR-2 (Flk-1) are significantly higher in patients with deeply infiltrating endometriosis affecting the rectum. Fertil Steril. 2008;90(1):148–55. https://doi.org/10.1016/j.fertnstert.2007.05.076.
Alshaikh RA, Ryan KB, Waeber C. Sphingosine 1-phosphate, a potential target in neovascular retinal disease. Br J Ophthalmol. 2021. https://doi.org/10.1136/bjophthalmol-2021-319115.
Obinata H, Kuo A, Wada Y, Swendeman S, Liu CH, Blaho VA, et al. Identification of ApoA4 as a sphingosine 1-phosphate chaperone in ApoM- and albumin-deficient mice. J Lipid Res. 2019;60(11):1912–21. https://doi.org/10.1194/jlr.RA119000277.
Jung B, Obinata H, Galvani S, Mendelson K, Ding BS, Skoura A, et al. Flow-regulated endothelial S1P receptor-1 signaling sustains vascular development. Dev Cell. 2012;23(3):600–10. https://doi.org/10.1016/j.devcel.2012.07.015.
Lee MJ, Thangada S, Claffey KP, Ancellin N, Liu CH, Kluk M, et al. Vascular endothelial cell adherens junction assembly and morphogenesis induced by sphingosine-1-phosphate. Cell. 1999;99(3):301–12. https://doi.org/10.1016/s0092-8674(00)81661-x.
Liu Y, Wada R, Yamashita T, Mi Y, Deng CX, Hobson JP, et al. Edg-1, the G protein-coupled receptor for sphingosine-1-phosphate, is essential for vascular maturation. J Clin Invest. 2000;106(8):951–61. https://doi.org/10.1172/jci10905.
Ishii I, Ye X, Friedman B, Kawamura S, Contos JJ, Kingsbury MA, et al. Marked perinatal lethality and cellular signaling deficits in mice null for the two sphingosine 1-phosphate (S1P) receptors, S1P(2)/LP(B2)/EDG-5 and S1P(3)/LP(B3)/EDG-3. J Biol Chem. 2002;277(28):25152–9. https://doi.org/10.1074/jbc.M200137200.
Kono M, Mi Y, Liu Y, Sasaki T, Allende ML, Wu YP, et al. The sphingosine-1-phosphate receptors S1P1, S1P2, and S1P3 function coordinately during embryonic angiogenesis. J Biol Chem. 2004;279(28):29367–73. https://doi.org/10.1074/jbc.M403937200.
Ben Shoham A, Malkinson G, Krief S, Shwartz Y, Ely Y, Ferrara N, et al. S1P1 inhibits sprouting angiogenesis during vascular development. Development. 2012;139(20):3859–69. https://doi.org/10.1242/dev.078550.
Gaengel K, Niaudet C, Hagikura K, Laviña B, Muhl L, Hofmann JJ, et al. The sphingosine-1-phosphate receptor S1PR1 restricts sprouting angiogenesis by regulating the interplay between VE-cadherin and VEGFR2. Dev Cell. 2012;23(3):587–99. https://doi.org/10.1016/j.devcel.2012.08.005.
Burg N, Swendeman S, Worgall S, Hla T, Salmon JE. Sphingosine 1-phosphate receptor 1 signaling maintains endothelial cell barrier function and protects against immune complex-induced vascular injury. Arthritis Rheumatol. 2018;70(11):1879–89. https://doi.org/10.1002/art.40558.
Hisano Y, Inoue A, Taimatsu K, Ota S, Ohga R, Kotani H, et al. Comprehensive analysis of sphingosine-1-phosphate receptor mutants during zebrafish embryogenesis. Genes Cells. 2015;20(8):647–58. https://doi.org/10.1111/gtc.12259.
Mendelson K, Zygmunt T, Torres-Vázquez J, Evans T, Hla T. Sphingosine 1-phosphate receptor signaling regulates proper embryonic vascular patterning. J Biol Chem. 2013;288(4):2143–56. https://doi.org/10.1074/jbc.M112.427344.
Anelli V, Gault CR, Cheng AB, Obeid LM. Sphingosine kinase 1 is up-regulated during hypoxia in U87MG glioma cells. Role of hypoxia-inducible factors 1 and 2. J Biol Chem. 2008;283(6):3365–75. https://doi.org/10.1074/jbc.M708241200.
Schnitzer SE, Weigert A, Zhou J, Brüne B. Hypoxia enhances sphingosine kinase 2 activity and provokes sphingosine-1-phosphate-mediated chemoresistance in A549 lung cancer cells. Mol Cancer Res. 2009;7(3):393–401. https://doi.org/10.1158/1541-7786.Mcr-08-0156.
Bouquerel P, Gstalder C, Müller D, Laurent J, Brizuela L, Sabbadini RA, et al. Essential role for SphK1/S1P signaling to regulate hypoxia-inducible factor 2α expression and activity in cancer. Oncogenesis. 2016;5(3):e209. https://doi.org/10.1038/oncsis.2016.13.
Salama MF, Carroll B, Adada M, Pulkoski-Gross M, Hannun YA, Obeid LM. A novel role of sphingosine kinase-1 in the invasion and angiogenesis of VHL mutant clear cell renal cell carcinoma. Faseb j. 2015;29(7):2803–13. https://doi.org/10.1096/fj.15-270413.
Michaud MD, Robitaille GA, Gratton JP, Richard DE. Sphingosine-1-phosphate: a novel nonhypoxic activator of hypoxia-inducible factor-1 in vascular cells. Arterioscler Thromb Vasc Biol. 2009;29(6):902–8. https://doi.org/10.1161/atvbaha.109.185280.
Semenza GL. Angiogenesis in ischemic and neoplastic disorders. Annu Rev Med. 2003;54:17–28. https://doi.org/10.1146/annurev.med.54.101601.152418.
Gavard J, Gutkind JS. VEGF controls endothelial-cell permeability by promoting the beta-arrestin-dependent endocytosis of VE-cadherin. Nat Cell Biol. 2006;8(11):1223–34. https://doi.org/10.1038/ncb1486.
Chae SS, Paik JH, Furneaux H, Hla T. Requirement for sphingosine 1-phosphate receptor-1 in tumor angiogenesis demonstrated by in vivo RNA interference. J Clin Invest. 2004;114(8):1082–9. https://doi.org/10.1172/jci22716.
Visentin B, Vekich JA, Sibbald BJ, Cavalli AL, Moreno KM, Matteo RG, et al. Validation of an anti-sphingosine-1-phosphate antibody as a potential therapeutic in reducing growth, invasion, and angiogenesis in multiple tumor lineages. Cancer Cell. 2006;9(3):225–38. https://doi.org/10.1016/j.ccr.2006.02.023.
Sanchez T, Skoura A, Wu MT, Casserly B, Harrington EO, Hla T. Induction of vascular permeability by the sphingosine-1-phosphate receptor-2 (S1P2R) and its downstream effectors ROCK and PTEN. Arterioscler Thromb Vasc Biol. 2007;27(6):1312–8. https://doi.org/10.1161/atvbaha.107.143735.
Skoura A, Sanchez T, Claffey K, Mandala SM, Proia RL, Hla T. Essential role of sphingosine 1-phosphate receptor 2 in pathological angiogenesis of the mouse retina. J Clin Invest. 2007;117(9):2506–16. https://doi.org/10.1172/jci31123.
Inoki I, Takuwa N, Sugimoto N, Yoshioka K, Takata S, Kaneko S, et al. Negative regulation of endothelial morphogenesis and angiogenesis by S1P2 receptor. Biochem Biophys Res Commun. 2006;346(1):293–300. https://doi.org/10.1016/j.bbrc.2006.05.119.
Rudzitis-Auth J, Christoffel A, Menger MD, Laschke MW. Targeting sphingosine kinase-1 with the low MW inhibitor SKI-5C suppresses the development of endometriotic lesions in mice. Br J Pharmacol. 2021;178(20):4104–18. https://doi.org/10.1111/bph.15601.
Sandkühler J. Models and mechanisms of hyperalgesia and allodynia. Physiol Rev. 2009;89(2):707–58. https://doi.org/10.1152/physrev.00025.2008.
Chang HM, Wu HC, Sun ZG, Lian F, Leung PCK. Neurotrophins and glial cell line-derived neurotrophic factor in the ovary: physiological and pathophysiological implications. Hum Reprod Update. 2019;25(2):224–42. https://doi.org/10.1093/humupd/dmy047.
Denk F, Bennett DL, McMahon SB. Nerve growth factor and pain mechanisms. Annu Rev Neurosci. 2017;40:307–25. https://doi.org/10.1146/annurev-neuro-072116-031121.
Odagiri K, Konno R, Fujiwara H, Netsu S, Yang C, Suzuki M. Smooth muscle metaplasia and innervation in interstitium of endometriotic lesions related to pain. Fertil Steril. 2009;92(5):1525–31. https://doi.org/10.1016/j.fertnstert.2008.08.101.
Orellana R, García-Solares J, Donnez J, van Kerk O, Dolmans MM, Donnez O. Important role of collective cell migration and nerve fiber density in the development of deep nodular endometriosis. Fertil Steril. 2017;107(4):987-995.e985. https://doi.org/10.1016/j.fertnstert.2017.01.005.
Mita S, Shimizu Y, Sato A, Notsu T, Imada K, Kyo S. Dienogest inhibits nerve growth factor expression induced by tumor necrosis factor-α or interleukin-1β. Fertil Steril. 2014;101(2):595–601. https://doi.org/10.1016/j.fertnstert.2013.10.038.
Khodorova A, Nicol GD, Strichartz G. The p75NTR signaling cascade mediates mechanical hyperalgesia induced by nerve growth factor injected into the rat hind paw. Neuroscience. 2013;254:312–23. https://doi.org/10.1016/j.neuroscience.2013.09.046.
Vercellini P, De Benedetti F, Rossi E, Colombo A, Trespidi L, Crosignani PG. Tumor necrosis factor in plasma and peritoneal fluid of women with and without endometriosis. Gynecol Obstet Invest. 1993;36(1):39–41. https://doi.org/10.1159/000292591.
Howard FM. Endometriosis and mechanisms of pelvic pain. J Minim Invasive Gynecol. 2009;16(5):540–50. https://doi.org/10.1016/j.jmig.2009.06.017.
Joseph EK, Levine JD. Caspase signalling in neuropathic and inflammatory pain in the rat. Eur J Neurosci. 2004;20(11):2896–902. https://doi.org/10.1111/j.1460-9568.2004.03750.x.
Doyle T, Chen Z, Obeid LM, Salvemini D. Sphingosine-1-phosphate acting via the S1P1 receptor is a downstream signaling pathway in ceramide-induced hyperalgesia. Neurosci Lett. 2011;499(1):4–8. https://doi.org/10.1016/j.neulet.2011.05.018.
Doyle T, Finley A, Chen Z, Salvemini D. Role for peroxynitrite in sphingosine-1-phosphate-induced hyperalgesia in rats. Pain. 2011;152(3):643–8. https://doi.org/10.1016/j.pain.2010.12.011.
Doyle T, Chen Z, Muscoli C, Obeid LM, Salvemini D. Intraplantar-injected ceramide in rats induces hyperalgesia through an NF-κB- and p38 kinase-dependent cyclooxygenase 2/prostaglandin E2 pathway. Faseb j. 2011;25(8):2782–91. https://doi.org/10.1096/fj.10-178095.
McAllister SL, Giourgas BK, Faircloth EK, Leishman E, Bradshaw HB, Gross ER. Prostaglandin levels, vaginal innervation, and cyst innervation as peripheral contributors to endometriosis-associated vaginal hyperalgesia in rodents. Mol Cell Endocrinol. 2016;437:120–9. https://doi.org/10.1016/j.mce.2016.08.017.
Arnold J, Barcena de Arellano ML, Rüster C, Vercellino GF, Chiantera V, Schneider A, et al. Imbalance between sympathetic and sensory innervation in peritoneal endometriosis. Brain Behav Immun. 2012;26(1):132–41. https://doi.org/10.1016/j.bbi.2011.08.004.
Wang G, Tokushige N, Markham R, Fraser IS. Rich innervation of deep infiltrating endometriosis. Hum Reprod. 2009;24(4):827–34. https://doi.org/10.1093/humrep/den464.
Wu J, Xie H, Yao S, Liang Y. Macrophage and nerve interaction in endometriosis. J Neuroinflammation. 2017;14(1):53. https://doi.org/10.1186/s12974-017-0828-3.
Mechsner S, Kaiser A, Kopf A, Gericke C, Ebert A, Bartley J. A pilot study to evaluate the clinical relevance of endometriosis-associated nerve fibers in peritoneal endometriotic lesions. Fertil Steril. 2009;92(6):1856–61. https://doi.org/10.1016/j.fertnstert.2008.09.006.
Coxon L, Horne AW, Vincent K. Pathophysiology of endometriosis-associated pain: a review of pelvic and central nervous system mechanisms. Best Pract Res Clin Obstet Gynaecol. 2018;51:53–67. https://doi.org/10.1016/j.bpobgyn.2018.01.014.
Quarta S, Camprubí-Robles M, Schweigreiter R, Matusica D, Haberberger RV, Proia RL, et al. Sphingosine-1-phosphate and the S1P(3) receptor initiate neuronal retraction via RhoA/ROCK associated with CRMP2 phosphorylation. Front Mol Neurosci. 2017;10:317. https://doi.org/10.3389/fnmol.2017.00317.
Aoki M, Aoki H, Ramanathan R, Hait NC, Takabe K. Sphingosine-1-phosphate signaling in immune cells and inflammation: roles and therapeutic potential. Mediators Inflamm. 2016;2016:8606878. https://doi.org/10.1155/2016/8606878.
Jaigirdar SA, Benson RA, Elmesmari A, Kurowska-Stolarska MS, McInnes IB, Garside P, et al. Sphingosine-1-phosphate promotes the persistence of activated CD4 T cells in inflamed sites. Front Immunol. 2017;8:1627. https://doi.org/10.3389/fimmu.2017.01627.
Zhao S, Adebiyi MG, Zhang Y, Couturier JP, Fan X, Zhang H, et al. Sphingosine-1-phosphate receptor 1 mediates elevated IL-6 signaling to promote chronic inflammation and multitissue damage in sickle cell disease. Faseb j. 2018;32(5):2855–65. https://doi.org/10.1096/fj.201600788RR.
Zhang YH, Fehrenbacher JC, Vasko MR, Nicol GD. Sphingosine-1-phosphate via activation of a G-protein-coupled receptor(s) enhances the excitability of rat sensory neurons. J Neurophysiol. 2006;96(3):1042–52. https://doi.org/10.1152/jn.00120.2006.
Li C, Li JN, Kays J, Guerrero M, Nicol GD. Sphingosine 1-phosphate enhances the excitability of rat sensory neurons through activation of sphingosine 1-phosphate receptors 1 and/or 3. J Neuroinflammation. 2015;12:70. https://doi.org/10.1186/s12974-015-0286-8.
Camprubí-Robles M, Mair N, Andratsch M, Benetti C, Beroukas D, Rukwied R, et al. Sphingosine-1-phosphate-induced nociceptor excitation and ongoing pain behavior in mice and humans is largely mediated by S1P3 receptor. J Neurosci. 2013;33(6):2582–92. https://doi.org/10.1523/jneurosci.4479-12.2013.
Hill RZ, Hoffman BU, Morita T, Campos SM, Lumpkin EA, Brem RB et al. The signaling lipid sphingosine 1-phosphate regulates mechanical pain. eLife 2018;7. https://doi.org/10.7554/eLife.33285.
Vercellini P, Fedele L, Aimi G, Pietropaolo G, Consonni D, Crosignani PG. Association between endometriosis stage, lesion type, patient characteristics and severity of pelvic pain symptoms: a multivariate analysis of over 1000 patients. Hum Reprod. 2007;22(1):266–71. https://doi.org/10.1093/humrep/del339.
Becker CM, Bokor A, Heikinheimo O, Horne A, Jansen F, Kiesel L, et al. ESHRE guideline: endometriosis. Hum Reprod Open. 2022;2022(2):hoac009. https://doi.org/10.1093/hropen/hoac009.
Norman RJ, Wu R. The potential danger of COX-2 inhibitors. Fertil Steril. 2004;81(3):493–4. https://doi.org/10.1016/j.fertnstert.2003.10.016.
Duffy JM, Arambage K, Correa FJ, Olive D, Farquhar C, Garry R et al. Laparoscopic surgery for endometriosis. Cochrane Database Syst Rev 2014;(4):CD011031. https://doi.org/10.1002/14651858.CD011031.pub2.
Guo SW. Recurrence of endometriosis and its control. Hum Reprod Update. 2009;15(4):441–61. https://doi.org/10.1093/humupd/dmp007.
Fakhr Y, Brindley DN, Hemmings DG. Physiological and pathological functions of sphingolipids in pregnancy. Cell Signal. 2021;85:110041. https://doi.org/10.1016/j.cellsig.2021.110041.
Author information
Authors and Affiliations
Contributions
Y. L. and F. Z. proposed the topic of the paper together, and both participated in searching and reviewing of relevant papers for the review. F. Z. was primarily responsible for the drafting, and Y. L. revised it. Both authors approved the final version.
Corresponding author
Ethics declarations
Ethics Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Yes. All authors approved for publication.
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Zhang, F., Lu, Y. The Sphingosine 1-Phosphate Axis: an Emerging Therapeutic Opportunity for Endometriosis. Reprod. Sci. 30, 2040–2059 (2023). https://doi.org/10.1007/s43032-023-01167-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s43032-023-01167-2