
Abstract
Excess nitrogen (N) after animal slurry application is a persistent problem of intensive agriculture, with consequences such as environmental pollution by ammonia (NH3) and nitrous oxide (N2O) emissions and nitrate (NO3−) leaching. High-carbon organic soil amendments (HCAs) with a large C:N ratio have shown the potential of mitigating unintended N losses from soil. To reduce gaseous and leaching N losses after the application of slurry, a laboratory incubation study was conducted with silt loam soil. We tested the potential of three different types of HCA—wheat straw, sawdust, and leonardite (application rate 50 g C L−1 slurry for each of the three HCAs)—to mitigate N loss after amendment of soil with pig and cattle slurry using two common application modes (slurry and HCA mixed overnight with subsequent addition to soil vs. sequential addition) at an application rate equivalent to 80 kg N ha−1. Compared to the control with only soil and slurry, the addition of leonardite reduced the NH3 emissions of both slurries by 32–64%. Leonardite also reduced the total N2O emissions by 33–58%. Wheat straw reduced N2O emissions by 40–46%, but had no effect on NH3 emission. 15 N labeling showed that the application of leonardite was associated with the highest N retention in soil (24% average slurry N recovery), followed by wheat straw (20% average slurry N recovery). The mitigation of N loss was also observed for sawdust, although the effect was less consistent compared with leonardite and wheat straw. Mixing the slurry and HCA overnight tended to reduce N losses, although the effect was not consistent across all treatments. In conclusion, leonardite improved soil N retention more effectively than wheat straw and sawdust.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Animal slurry, also known as liquid manure, contains large amounts of reactive nitrogen (N), an essential nutrient for plants and microorganisms that enhances metabolic rates and soil respiration when applied to soil. However, excessive N introduced to the environment, for example, in the form of ammonia (NH3), can be a harmful pollutant of surface water and groundwater, cause biodiversity loss, and endanger human health (Aber et al. 1995). In the form of nitrous oxide (N2O), it can exacerbate global warming (Rabalais 2002) and deplete the stratospheric ozone layer (Cowling et al. 1998).
Excessive N in the environment is a serious problem, particularly in areas with intensive livestock production such as northwest Germany, one of the areas with the most intensive livestock production in the world (Zerbe 2020). The predominant N form in animal slurry is ammonium (NH4+), which accounts for over 50% of total N (TN) in pig and cattle slurry (Jensen 2013). The calculated global median NH3 loss from the application of animal slurry amounts to 23%, which is higher than the loss resulting from the application of synthetic N fertilizer (Bouwman et al. 2002). In order to reduce NH3 emissions, the 2017 German Fertilizer Ordinance aims to gradually ban the application of liquid manure using large spreaders to crop land by 2020 and to grassland by 2025. Once in the soil, NH3 can rapidly be protonated to NH4+ and oxidized to nitrate (NO3−) by ammonia-oxidizing bacteria (AOB) and ammonia-oxidizing archaea (AOA). Any NO3− that is not taken up by plants is eventually leached into surface water and groundwater. N2O is formed during both nitrification and denitrification, and is a significant contributor to global warming as a greenhouse gas (GHG) that is 265 times more potent than carbon dioxide (CO2) (IPCC 2013). Therefore, it is important to improve soil N retention and to reduce N pollution from animal slurry.
High-carbon organic soil amendments (HCAs) can immobilize N and reduce GHG emissions and odor due to their composition and structure. Wheat straw, for example, contains considerable amounts of labile organic carbon (C), which can stimulate microbial activity and growth, and thus N immobilization in the soil (Zavalloni et al. 2011). Sawdust can eliminate the bad odor of landfills and animal or human excreta due to its high sorption capacity (Hui et al. 2003). Leonardite is a highly oxidized lignite of acidic nature that increases the degree of protonation of NH3 when applied to slurry or soil, has a high cation exchange capacity (CEC) that promotes the adsorption of NH4+, and contains up to 20% labile C that can stimulate microbial N immobilization (Manzoni and Porporato 2009). Furthermore, the addition of HCA, (e.g., straw) during slurry storage increases the amount of volatile fatty acids and may increase net N immobilization after the application of slurry to the soil (Kirchmann et al. 1993; Sørensen 1998).
HCAs have great potential to reduce N losses from soils, but it is still unclear which HCA (wheat straw, sawdust, leonardite) are best suited to effectively bind N in different types of slurry and whether recovery of N from slurry can be enhanced by the combined application of HCA and slurry to the soil.
We hypothesized that leonardite would be more effective than wheat straw and spruce sawdust in reducing N loss after slurry application due to its combination of properties. We also hypothesized that mixing HCA with slurry prior to application would increase N retention. To put these hypotheses to the test, a laboratory incubation experiment was conducted with leonardite to compare it with sawdust and wheat straw. Pig or cattle slurry was applied to a silt loam soil, either after pre-incubation of the slurry with HCA or directly to the soil amended with the respective HCA. GHG (CO2, CH4, and N2O) and NH3 emissions were monitored during the 60-day incubation period to quantify the N retention capacity of the HCA in combination with liquid manure.
2 Materials and Methods
2.1 Soil
The soil was obtained from an agricultural field close to Selhausen (Germany) and stored in a dried and 2-mm sieved state until use (Quade et al. 2018). It was classified as an orthic Luvisol with a silt loam soil texture and a pHH2O of 7.1, containing 10.0 ± 0.2 g kg−1 total organic carbon (TOC) and 1.07 ± 0.05 g kg−1 TN.
2.2 Slurries
Pig and cattle slurries were obtained from typical animal farms from the Rhineland region (Rheinisches Revier) close to Grevenbroich (North Rhine-Westphalia, Germany). Pig and cattle slurry had a density of 1.01 g cm−3 and 0.92 g cm−3, a water content of 88% and 91%, a pH of 8.4 and 8.5, and a C/N ratio of 5.9 and 6.3, respectively. Based on dry weight, the pig slurry contained 64 mg g−1 TN, of which 45 mg g−1 was NH4+, 376 mg g−1 was TOC, and 193 mg g−1 was dissolved organic carbon (DOC); the cattle slurry contained 60 mg g−1 TN, of which 32 mg g−1 was NH4+, 378 mg g−1 was TOC, and 279 mg g−1 was DOC.
2.3 HCA
The HCA types used in the experiment were wheat straw, sawdust, and leonardite. The total C contents of the HCAs were 43.6%, 44.0%, and 46.3%; the TN contents amounted to 0.97%, 0.16%, and 0.92%; and the C:N ratios were 45, 275, and 50, respectively. The average pH in water of the leonardite was 4.2. Before application, wheat straw was cut into pieces that were 2–3 cm in length, and small particles of spruce sawdust without bark were added that were just a few millimeters in size. Leonardite was applied in powder form.
2.4 Incubations
Incubations were performed in PVC tubes (150-mm height, 50-mm diameter) containing 390 g of dried soil at a soil bulk density of 1.3 g cm−3. The dry soil was reactivated by pre-incubating at 60% water-holding capacity (WHC, 36 g water per 100 g dry soil) for 10 days. The experiment consisted of 17 treatments with three replicates each—according to the combination of two types of slurry (pig and cattle slurry)—and applied at a rate of 80 kg TN ha−1 (corresponding to 4.75 ml pig slurry or 6.22 ml cattle slurry per incubation tube). Three types of HCA (wheat straw, sawdust, leonardite) were applied at a rate of 50 g C L−1 slurry (corresponding to 62 kg C ha−1 for pig slurry and 73 kg C ha−1 for cattle slurry); the application rate of HCA was previously found to be suitable to treat animal slurry with lignite or leonardite for the purpose of maximal nutrient immobilization (German patent DE102016223352A1 by RWE Power AG). Two common agricultural application methods were simulated: either mixing slurry and the respective HCA overnight before application and adding the mixture to the soil on the following day or adding HCA to the soil first and then adding slurry directly to the HCA-amended soil. A factorial design of 2 slurries × 3 HCA × 2 application methods was used, amounting to 12 treatments in total. Five different control groups were established: (1) Soil (S); (2) Soil and Pig slurry (SP); (3) Soil and Cattle slurry (SC); (4) Pig slurry (P); and (5) Cattle slurry (C). An overview of all treatments and abbreviations is presented in Table 1. Slurry and HCA were incorporated into the first 2 cm of the soil immediately after application. Throughout the 60 days of incubation, the experiment was conducted at a room temperature of 20 °C, the tubes were weighed daily, and water loss was compensated by the addition of the corresponding amount of deionized water. Due to the large number of treatments, the experiment was divided into two batches with the pig slurry batch starting 12 days earlier than the cattle slurry batch. To follow the development of the slurry-derived N, we added and mixed 40 mg (15NH4)2SO4 (at 1 atom% 15 N) each to the 200 ml pig and cattle slurries, respectively, before application.
2.5 Gas Analysis
During the experiment, GHG and NH3 emissions of the different treatments were measured on days 1, 2, 3, 5, 7, and 10 of the experiment, and weekly thereafter, until the end of the experiment. An infrared laser gas analyzer (G2308, Picarro, Inc., Santa Clara, CA, USA) with a dynamic chamber (50-mm height, 50-mm diameter) tightly covering each tube was used in closed-loop mode to quantify the emissions of CO2, N2O, and NH3 within 10 min. Gas fluxes were calculated according to Brummer et al. (2008). On the first day of the experiment, the CO2 emissions data in the P treatment and N2O and NH3 emissions data in the P, SPW, and SPD treatments exceeded the calibrated range of the analyzer. An interpretation of this data was ruled out.
2.6 Nutrient Analysis
After the experiment, soil samples were collected and separated into a top layer (0–5 cm, including slurries and HCA) and a sublayer (5–15 cm). All samples were stored in a freezer at − 22 °C before extraction. According to Houba et al. (2000), mineral N fractions (NH4+ and NO3−) representing the available N were extracted with a 0.01 M CaCl2 solution (soil-to-solution ratio of 1:4 w:v) that had an ionic strength similar to that of the soil solution by 2-h horizontal shaking at 200 rpm and 15-min centrifugation at 3500 rpm. The supernatant was filtrated with 0.45-µm PP-membrane syringe filters (disc size 25 mm; VWR International, Darmstadt, Germany) and then subjected to continuous flow analysis and ion chromatography (Dionex DX-500, ThermoScientific, Massachusetts, USA) for NH4+ and NO3−, respectively.
2.7 Microbial Biomass
Soil microbial biomass was extracted using the chloroform-fumigation extraction method (CFE) as described in Reichel et al. (2017). A TOC analyzer (TOC-VcPH + TNM-1 + ASI-V, Shimadzu, Japan) was used to determine the DOC in CaCl2 extracts both without and after chloroform fumigation in order to quantify the microbial biomass C (Cmic). Cmic and Nmic were calculated according to Joergensen (1996) using the correction factors kEC 0.45 and kEN 0.40.
2.8 15 N Analysis
For both the top layer and the sublayer of soil samples, 2-g air dry soil were extracted with 50 ml 1 M potassium chloride (KCl) by 1-h horizontal shaking at 200 rpm and 20 min centrifugation at 3500 rpm. Here, the extraction with 1 M KCl solution was chosen to recover the maximum amount of 15NH4+ from the soil (also including the fraction adsorbed by the soil matrix) in order to analyze of the development of slurry N in soil. The N leaching potential was estimated from the NO3− content in the soil solution, which was sampled at a soil depth of 10 cm using MicroRhizon samplers (type 19.21.81, Rhizosphere Research Products, Wageningen, Netherlands) on days 0 (before application), 4, and 8, and weekly thereafter, until the end of the experiment. All solution samples were stored in a freezer at − 22 °C before analysis. For the analysis, each sample (1–2 ml) was diluted to 20 ml with deionized water. 15 N isotope signatures of NH4+ and NO3− in soil extracts and soil solutions were analyzed by applying sequential micro-diffusion and liquid–liquid extraction techniques as described in Reichel et al. (2018).
2.9 Calculations and Statistical Analysis
The results are expressed as a mean value of three replicates ± standard deviation on a dry soil basis. After considering the statistical requirements (normal distribution and homogeneity of variance), the differences between treatments were analyzed for significance by three-way analysis of variance (ANOVA) with Tukey’s HSD used as the post-hoc test at a significance level of p ≤ 0.05 (IBM SPSS Statistics for Windows, Version 25.0, IBM Corp., Armonk, NY, USA).
3 Results
The three-way ANOVA (Supplementary data, Table S1) showed that cumulative GHG and NH3 emissions and 15 N recovery responded significantly to the type of slurry (larger in pig than in cattle slurry) and the HCA type. In addition, application methods also affected N2O, NH3 emissions, and 15 N recovery (p < 0.05). In the top layer of the soil, the type of HCA significantly affected the DOC, Cmic, NO3−, and pH (p < 0.05). In the sublayer, NO3− responded to all three factors, while pH only responded to the application method.
3.1 pH
The pH values of the topsoil layer of all treatments were lower than in the S control at the end of the incubation period (Table 2). Leonardite significantly decreased the pH values of the soil to below 7 for both slurries. There was no significant difference between treatments in the sublayer; the pH of all samples was around 7.6.
3.2 CO2
The CO2 emission rates were extremely high on the first day (Supplementary data, Figure S1). The highest emission rates occurred in SPD (33.7 g C m−2 day−1) and SCW (25.7 g C m−2 day−1), while SPL (12.8 g C m−2 day−1) and SCL (9.8 g C m−2 day−1) had the lowest CO2 emission rates compared with the other HCAs. Comparing the two application methods, the first application method (mixing slurry and HCA overnight followed by application of the mixture on the following day) led to significantly higher CO2 emission rates for cattle slurry treatments on the first day; this difference then diminished in the following days. The CO2 emission rates rapidly declined in all treatments after the first day, but remained at an intermediate level for 10 days before levelling off at 1 g C m−2 day−1 towards the end of the experiment. In this period, the slurry-only treatments P and C always had the lowest CO2 emission rates.
The cumulative CO2 emissions of the two application methods were not significantly different from each other, while the wheat straw treatments emitted considerably more CO2 compared to the other HCA treatments. Adding pig slurry to the soil increased cumulative CO2 emissions by 39% compared to the S treatment (Fig. 1A). The addition of wheat straw increased CO2 emissions by 15% in the SP + W treatment compared to the SP treatment. Leonardite significantly decreased CO2 emissions by 16% and 15% in the SP + L and SPL treatments, respectively, compared to the corresponding control (SP). The CO2 emissions of the SP + D treatment were also reduced by 17% compared to the corresponding control (SP).

A Cumulative CO2 emissions of pig slurry treatments. B Cumulative CO2 emissions of cattle slurry treatments. C Cumulative N2O emissions of pig slurry treatments. D Cumulative N2O emissions of cattle slurry treatments. E Cumulative NH3 emissions of pig slurry treatments. F Cumulative NH3 emissions of cattle slurry treatments. The abbreviations of the treatments are as follows: S, soil; P, pig slurry; C, cattle slurry; W, wheat straw; D, sawdust; L, leonardite; “ + ”, direct application of slurry and HCA to the soil; no “ + ”, slurry and HCA mixed overnight before application. The error bars show the standard error of the mean of each treatment (n = 3). The different lowercase letters indicate significant differences at the p < 0.05 level between treatment. The CO2 emissions of the P treatment are not compared due to the absence of data for the first day. The N2O emissions of the P, SPW, and SPD treatments are not compared due to the absence of data for the first day. The NH3 emissions of the P, SPW, and SPD treatments are not compared due to the absence of data for the first day
In contrast to pig slurry, the addition of cattle slurry to soil only increased CO2 emissions by 1% (Fig. 1B). In the cattle slurry treatments, wheat straw increased CO2 emissions by 51% and 29% in SC + W and SCW treatments, respectively, compared to the SC treatment. However, the other HCA types did not significantly reduce the CO2 emissions in the cattle slurry treatments.
3.3 N2O
On the day of slurry application, the emission rates of N2O (Supplementary data, Figure S1) peaked in the C treatment with a maximum emission rate of 813 mg N m−2 day−1. Among the HCA substrates tested, only leonardite significantly reduced N2O emission rates for both slurry treatments. On day 2 after application, the rates decreased rapidly below 5 mg N m−2 day−1. A second peak was reached in the pig and cattle slurry treatments 18 days and 7 days after application, respectively, while it was reached on day 10 in the C (cattle slurry only) treatment. The application of pig slurry together with wheat straw was associated with the lowest N2O emission rates during the second peak period, in contrast to cattle slurry where this difference was not significant. Thereafter, N2O emission rates gradually declined to the level of untreated soil.
The application of pig slurry increased cumulative N2O emissions 11-fold compared to the soil-only (S) treatment. N2O emissions were reduced by 46% and 58% in SP + W and SP + L, respectively, compared to the SP treatment (Fig. 1C). The N2O emissions of SC were increased by twofold compared with the S treatment. The SCW, SC + L, and SCL treatments tended to reduce the N2O emissions by 40%, 34%, and 33%, respectively, in relation to the SC treatment (Fig. 1D).
3.4 NH3
In general, pig slurry treatments had higher NH3 emissions than cattle slurry treatments (Fig. 1E, F). The application of pig slurry to soil was associated with a large NH3 loss. The total NH3 emissions in the SP treatment were 3.9 times higher than in the SC treatment. On the first day of slurry application, the NH3 loss was extremely high, with maximum emission rates of 4000 mg N m−2 day−1 and 5000 mg N m−2 day−1 in the pig and cattle slurry treatments (Supplementary data, Figure S1), respectively. One day after slurry application, NH3 emissions declined considerably to rates below 150 mg N m−2 day−1, while in the P treatment, emissions remained high at a rate of 565 mg N m−2 day−1. NH3 strongly decreased to below 1 mg N m−2 day−1 after day 10, with the exception of P where NH3 emissions could no longer be detected after day 25.
The two application methods had different effects on cumulative NH3 emissions for the two slurries. In the pig slurry treatments, mixing slurry and HCA reduced NH3 emissions compared to adding slurry and HCA directly to the soil, but the opposite was the case in the cattle slurry.
Leonardite had a similar effect on reducing NH3 emissions in both slurries compared to the soil + slurry control. In treatments in which leonardite was added directly to the slurry, cumulative NH3 emissions were reduced by 34% in pig slurry and by 32% in cattle slurry. Leonardite mixed with slurry overnight before application reduced the cumulative NH3 emissions from pig slurry (SPL) and cattle slurry (SCL) by 63% and 64%, respectively.
In contrast, wheat straw and sawdust increased the NH3 emissions when mixed with cattle slurry. Compared with the SC treatment, wheat straw significantly increased the NH3 emissions by 72% (SC + W) and 170% (SCW). Sawdust increased the cumulative NH3 emissions by 46% (SC + D) and by 154% (SCD) in relation to the SC treatment.
3.5 DOC and Microbial Biomass
In relation to the control S, increased DOC concentrations were found in the top soil layer of all treatments at the end of the incubation period (Fig. 2A, B). The addition of pig slurry to the soil increased the DOC concentration by 2.9 times, which was larger than after the addition of the cattle slurry (2.2 times). All the tested HCA types increased the DOC content in relation to the SC treatment: wheat straw and leonardite increased the DOC concentration by 61–68%, while for sawdust, the increase was in the range of 23–26%. In the pig slurry treatments, wheat straw increased the DOC by 42% (SP + W) and 18% (SPW). Leonardite increased the DOC by 16% (SP + L) and 30% (SPL). In contrast, the DOC decreased in sawdust treatments with pig slurry (10–12%). In the sublayer of the soil, the DOC concentrations were lower than 80 µg C g−1 without any significant difference between the treatments (Supplementary data, Figure S2).

A DOC of pig slurry treatments. B DOC of cattle slurry treatments. C Cmic of pig slurry treatments. D Cmic of cattle slurry treatments. S, soil; P, pig slurry; C, cattle slurry; W, wheat straw; D, sawdust; L, leonardite; “ + ”, direct application of slurry and HCA to the soil; no “ + ”, slurry and HCA mixed overnight before application. The error bars show the standard error of the mean of each treatment (n = 3). The different lowercase letters indicate significant differences at the p < 0.05 level between treatments
Both slurries and the tested HCA types increased the Cmic in the top layer (Fig. 2C, D). Most HCA treatments with pig slurry tended to have a larger microbial biomass compared to the SP treatment. The Cmic of the SP + W treatment was 4.4 times higher than in the SP treatment. The wheat straw treatments with cattle slurry (SC + W and SCW) tended to have a larger Cmic compared to the SC treatment. In the sublayer, none of the HCA types appeared to have a significant effect on the Cmic of the pig slurry treatments. All the measured Cmic concentrations were lower than 65 µg g−1. Most of the Cmic of the cattle slurry treatments ranged were below 70 µg g−1, with the exception of the SC + L (114 µg g−1 dry soil) (Supplementary data, Figure S2).
3.6 Mineral N in Soil
At the end of the experiment, NH4+ concentrations reached their minimum level in all treatments. Pig slurry treatments with wheat straw still had the highest NH4+ content. Compared to the SP, the NH4+ content was 9.4 and 5.9 times higher in the SP + W and SPW treatments. Soil column sublayers contained less than 2 µg NH4-N g−1 except for the SPW treatment, which contained 3.6 µg NH4-N g−1. In contrast, the HCA had no significant effect in the cattle slurry treatments (Table 2).
At the end of the experiment, the NO3− concentrations of all treatments were larger than in the S treatment (control). Most of the NO3− was retained in the top layer of the soil columns (Table 2). The addition of cattle slurry increased the NO3− concentration 1.9-fold compared to the treatments with pig slurry. Wheat straw and sawdust decreased NO3− in the top layer. However, the decrease of NO3− compared to the SC treatment was only significant for the cattle slurry treatments with wheat straw (65% for SC + W and 61% for the SCW treatment) and sawdust (53% for SC + D and 45% for the SCD treatment). In the sublayers of all treatments, NO3− was below 105 µg N g−1, but tended to be higher in the sublayer of the cattle slurry treatments with wheat straw and sawdust compared to leonardite treatments.
3.7 15 N Recovery in Soil
Wheat straw and leonardite increased the 15 N recovery in the total N fraction (15Nt) of both slurry treatments (Fig. 3). In the pig slurry treatments, wheat straw increased the 15Nt recovery by 58% (SPW) and 111% (SP + W) and leonardite by 106% (SPL) and 91% (SP + L) compared to the SP treatment. In the cattle slurry treatments, wheat straw increased the 15Nt recovery by 66% (SCW) and 28% (SC + W) and leonardite by 178% (SCL) and 37% (SC + L), respectively. However, only the increase in the SCL treatment was statistically significant. Sawdust had no apparent effect on the 15Nt recovery. Mixing cattle slurry and HCA overnight significantly increased 15Nt recovery compared to adding cattle slurry and HCA directly to soil, although not in the pig slurry treatments.

A 15 N recovery in the pig slurry treatments. B 15 N recovery in the cattle slurry treatments. The abbreviations of the treatments are as follows: S, soil; P, pig slurry; C, cattle slurry; W, wheat straw; D, sawdust; L, leonardite; “ + ”, direct application of slurry and HCA to the soil; no “ + ”, slurry and HCA mixed overnight before application. The error bars show the standard error of the mean of each treatment (n = 3). The different lowercase letters indicate significant differences at the p < 0.05 level between treatment
3.8 Mineral N and δ15N in Soil Solution
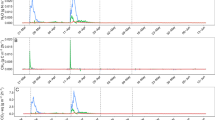
The soil solution volume of all samples was 1–2 ml. Before slurry application, NH4+ concentrations in all treatments were < 10 µg N ml−1 (Fig. 4A, B). In pig slurry treatments, the NH4+ peaked on day 8 after slurry application (> 10 µg N ml−1) and decreased to values < 5 µg N ml−1 by day 29. Most NH4+ concentrations of the cattle slurry treatments were below those of the pig slurry treatments. The NH4+ concentration of the cattle slurry treatments reached an initial peak on day 8 after slurry application (~ 8 µg N ml−1) and declined to around 2 µg N ml−1 by day 22. Interestingly, the highest NH4+ concentration in the soil solution was found in the SC + L treatment on day 22 (11 µg N ml−1), remaining at this level for longer than in all other treatments.

Mineral N concentrations in the soil solution. A NH4+ content of pig slurry treatments. B NH4+ content of cattle slurry treatments. C NO3− content of pig slurry treatments. D NO3− of cattle slurry treatments. E 15NH4+ of pig slurry treatments. F 15NH4+ of cattle slurry treatments. G 15NO3− of pig slurry treatments. H 15NO3− of cattle slurry treatments. The abbreviations of the treatments are as follows: S, soil; P, pig slurry; C, cattle slurry; W, wheat straw; D, sawdust; L, leonardite; “ + ”, direct application of slurry and HCA to the soil; no “ + ”, slurry and HCA mixed overnight before application. The error bars show the standard error of the mean of each treatment (n = 3)
Fig. 4E, F, G, and H show the development of the δ15N trends over the total incubation time of 60 days. In the pig slurry treatments, the 15NH4+ reached its peak value on day 22, slightly later than the NH4+ concentrations. After 36 days of incubation, 15NH4+ content decreased to its initial level before slurry application. In cattle slurry treatments, 15NH4+ of all HCA treatments reached peak values before day 15, except for the leonardite treatments which peaked on day 22. 15NH4+ also declined to its initial level on day 36
The NO3− concentrations of all pig slurry treatments were < 50 µg ml−1 and remained at this level until incubation on day 8 before increasing to a peak value on day 29 (~ 255 µg N ml−1). There were no significant differences between the NO3–-N concentrations among treatments. In the cattle slurry treatments, after slurry and/or HCA application, NO3− concentrations increased to a maximum concentration between 250 and 300 µg ml−1 on day 22; only the SCL treatment peaked on day 29 (322 µg ml−1).
In contrast to the trend in NO3− concentrations, 15NO3− trends were concurrent with 15NH4+ at the beginning. The 15NO3− of the leonardite treatments reached its highest value on day 29, while the other treatments reached their peak on day 22, remaining at that high level until the end of the experiment.
In all treatments, the NH4+ and NO3− developed in opposite directions, with the maximum NH4+ concentration and minimum NO3− concentration in the early incubation period and the lowest and largest concentrations at the end of the incubation, respectively.
4 Discussion
In this experiment, the GHG (CO2 and N2O) and NH3 emissions in pig slurry treatments were higher than in cattle slurry treatments. According to Kirchmann et al. (1993), the lower CO2 emissions in the cattle slurry treatments are related to a lower C mineralization potential in the soil. In contrast to pigs, the high digestive activity in cattle rumen might have removed labile C sources from the slurry, thus lowering its CO2 emissions potential in soil.
The pig slurry contained more NH4+ (70% of TN) than the cattle slurry (53% of TN), and the pig slurry infiltrated into the soil much more slowly than the cattle slurry. A restricted infiltration of the slurry into the soil was found to increase NH3 emissions (Chen 1986), as this increases the slurry’s time of exposure to the atmosphere and hence the time for NH3 outgassing. Our experiment also showed that less N2O was emitted from cattle slurry treatments. The rumen activity of cattle leads to more DOC in the cattle slurry (74% DOC in cattle slurry compared to 51% DOC in pig slurry), which could have resulted in a stronger immobilization of available N in the soil, thus reducing the N2O emissions potential (Burger and Jackson 2003). Overall, we can state that the difference in the DOC and the mineral N content in pig and cattle slurries led to the observed differences in gas emission between the treatments with pig and cattle slurries.
Wheat straw treatments emitted more CO2 than the other HCA treatments with both slurries. The larger fraction of easily available C of wheat straw most likely stimulated the growth of microorganisms after incorporation into the soil. Slurry-NH4+ further stimulated the mineralization of wheat straw, which additionally increased CO2 emissions (Chantigny et al. 2001; Firestone et al. 1980; Sørensen 1998). Leonardite also contains sources of labile C, which also significantly increased DOC and Cmic concentrations in pig slurry treatments, but without a significant increase in CO2 emissions. This is in agreement with previous studies, and is also related to the release of proton from acidic leonardite in neutral soils (Schefe et al. 2008; Tran et al. 2015). In general, the high amount of labile C released from wheat straw increased CO2 emissions, while the acidity of leonardite could have counteracted this effect.
N2O emission rates showed two peaks. Similar dynamics have previously been observed in the lab (Velthof et al. 2003) and in the field (Thomsen et al. 2010). The peak on the day of slurry application could have originated from the denitrification of soil NO3− stimulated by the input of labile organic C due to slurry application, as assumed by Velthof et al. (2003). This is supported by the findings of Nguyen et al. (2017), who provided a much more detailed insight into the NH4+, NO3−, and O2 dynamics after slurry application to a grassland soil, and who found N2O dynamics very similar to those of our findings. They used a planar optode setup to monitor the O2 concentration in the soil before, during, and after slurry application. In addition, they also implemented a treatment, in which the nitrification inhibitor 3, 4-dimethyl pyrazole phosphate was added to the slurry. Finally, they determined the 15 N site preference of the emitted N2O as an indicator of the N2O origin (nitrification/fungal denitrification vs. bacterial denitrification). For the first peak, they found no evidence of a significant contribution of nitrification to N2O emissions despite clearly measurable nitrification activity, which was indicated by the significant difference in oxygen consumption between the slurry treatments with and without the nitrification inhibitor and the much higher NH4+ and much lower NO3− concentrations in the topsoil at the end of the experiment. Nevertheless, the site preference of N2O during the first peak was much higher than is typical for bacterial denitrification, which Nguyen et al. (2017) attributed to the fungal denitrification of NO3− present in the soil before slurry application in addition to the fact that the nitrification inhibitor did not have an effect on the magnitude of the first N2O peak. In contrast, during the second N2O peak, Nguyen et al. (2017) found higher O2 concentrations, significantly higher N2O emissions in the treatments without the nitrification inhibitor, and intermediate N2O site preference, with significantly lower values for the treatment without the nitrification inhibitor, all of which point towards a close connection between nitrification and bacterial denitrification as N2O source processes. This is also backed up by the N2O, soil NH4+, and NO3− dynamics reported by Thomsen et al. (2010), who found increasing concentrations of NO3−in the soil coinciding with the second N2O peak. The oxygen concentration in the soil gradually increases again with the depletion of labile C due to the initially strong respiration activity following slurry application (Nguyen et al. 2017). This enables an increase in nitrification activity and hence an increase in NO3− concentration, which in turn serves as a substrate for denitrification. With the build-up of the soil NO3− pool, N2O emissions thus increase again to a second peak due to simultaneous nitrification–denitrification (Rochette et al. 2008; Saguer and Gispert 1996). In short, the two N2O emission peaks had different causes: while the first peak likely derived from denitrification alone, stimulated by a high soil NO3− content, the second N2O peak was likely due to simultaneous nitrification–bacteria denitrification.
Leonardite’s mitigating effect on NH3 emissions was evident for both slurries, but in particular for the pig slurry. When leonardite was mixed with the slurries 1 day before application, the total NH3 emissions were reduced by more than 60% compared to the control (soil + slurry). This effect was likely due to the acidic nature and high CEC of leonardite, which led to the protonation of NH3 and the adsorption of NH4+ (Chen et al. 2015; Sommer et al. 2003). The deprotonation of the acid groups on the surface of the lignite decreased the pH of the alkaline slurry, thus enhancing the NH4+/NH3 ratio, and increased the negative charge on the surface of the leonardite, providing more adsorption sites for NH4+ (Simmler et al. 2013).
In this experiment, leonardite also reduced N2O emissions, especially in the SP + L treatment where the reduction amounted to 58%. This was in contrast to the study by Sun et al. (2016), but they applied lignite, i.e., a less oxidized form than leonardite, directly to the surface of a cattle pen, thereby probably creating a urine-soaked soil cover, in which less cation exchange sites compared to leonardite and micro-anaerobic conditions may have promoted denitrification and N2O formation.
In contrast to our results, wheat straw applied to fertilized soil without manure was previously found to increase N2O emissions (Wei et al. 2020). The application of slurry may therefore have partly suppressed potential N2O emission pathways in our study. For example, animal slurry could have stimulated the mineralization of wheat straw, which may have promoted the growth of soil microbial biomass and N immobilization, as indicated by the tendency to higher Cmic values in the wheat straw treatments (Fig. 2), which subsequently led to a reduction in the availability of N to the nitrifying microorganism (Aita et al. 2012; Wei et al. 2020). In contrast, wheat straw and sawdust reduced the infiltration velocity of slurry into the soil and lowered the water content of slurries (Vandre et al. 1997). This can lead to an increase in NH3 volatilization, as was observed in our experiment. HCAs also significantly affect N in the soil. After the 60 days of incubation, wheat straw and leonardite increased the DOC content of the topsoil layer for both slurry treatments. This is in line with the fact that wheat straw contains more labile organic compounds than sawdust; and lignite was also reported to contain up to 20% of labile C (Chen et al. 2015). The amount of labile OC determines the microbial N immobilization potential by microbial biomass growth. Leonardite therefore clearly has the potential for microbial N immobilization in addition to its physicochemical N retention properties.
Mineral N (NO3− and NH4+) was mainly retained in the top layer of the soil and was not translocated downward along the soil columns. This might have been due to the fact that the water content was kept constant throughout the experiment by adding small amounts of water every day. This prevented significant vertical water movement in the soil and was therefore different from larger natural precipitation events. Most of the NH4+ was oxidized to NO3− by the end of the incubation period, although the soil treated with cattle slurry (SC) contained less NH4+ and more NO3− than the soil treated with pig slurry (SP). The cattle slurry thus seems to be more conducive to nitrification than the pig slurry. One reason for this might be that heavy metals and antibiotic substances are more abundant in pig slurry than in cattle slurry, which affects microbial N transformations in the soil (Sommer et al. 2003). Compared to leonardite, the addition of wheat straw and sawdust led to lower NO3− concentration in the top layer of the cattle slurry treatments, which could have resulted from the higher NH3 loss than in the leonardite treatments (Fig. 1F) and the preservation of NH4+ by leonardite during the early phase of the incubation. In short, the two different slurries led to significant differences in mineral N dynamics, with a lower nitrification rate in the soil treated with pig slurry compared to the soil amended with cattle slurry. Of the three HCAs tested, leonardite resulted in the highest N retention.
Overall, the application of wheat straw and leonardite was associated with a higher 15 N recovery than in the sawdust treatments, i.e., more slurry-N was retained in the soil. The reason for this may very likely be the increase in DOC in the soil due to the application of wheat straw and leonardite, which led to an increase in N retention by stimulating microbial growth. However, compared with straw, leonardite has a more acidic nature and higher CEC, which leads to a stronger ability to adsorb slurry-NH3 in the form of NH4+. As a consequence, leonardite was characterized by lower N2O and NH3 emissions compared to the other HCA treatments, leading to a higher N retention in the topsoil layer.
The trend in δ15N showed the N pathway of the labeled slurry, i.e., NH4+ content decreased while NO3− increased due to nitrification. It is remarkable that the δ15N of the NH4+ of the leonardite treatments reached higher values, but at a later point in time compared to the other HCA treatments. The reason for this could be that leonardite adsorbed and retained more NH4+ in the soil due to its high CEC and its acidic nature and thus its potential negative effect on nitrification activity, thus preserving more NH4+ from being nitrified to NO3− in the early incubation phase.
The effects of the different slurry application methods were complex in our experiment. The overnight mixing of slurry and HCA increased the CO2 emission rate on the day of application only for the cattle slurry treatments. This may be due to the conversion of some insoluble organic matter in the cattle slurry to soluble C by fermentation during overnight storage of the slurry with the HCA (Harper and Lynch 1982). The pig slurry already contained more DOC than the cattle slurry, which is why mixing with HCA and overnight storage did not have a significant effect on the pig slurry treatments.
The most noteworthy result was that leonardite reduced total NH3 emissions by more than 60% compared to the control when mixed with slurry the day before application. However, when the HCA and slurry were added sequentially, the reduction was only about 33%. Although significant differences were found only in the pig slurry treatments, there was a tendency for mixing the slurries and the HCA before application to reduce NH3 emissions more than the sequential method for both slurry treatments. Mixing slurries and leonardite overnight before application led to more NH4+ adsorption to the leonardite surface (Sommer et al. 2003), while also likely increasing microbial N immobilization due to labile C released from leonardite (Sørensen 1998).
However, despite the trends described above, the effects of the different application methods for the different slurries and HCAs were not consistent, which were likely due to the short mixing time. Overnight mixing enabled the leonardite to adsorb NH4+ through a physicochemical process, but the time was too short to obtain significant effects from biochemical processes. It has been reported in the literature, that this usually takes several weeks or months (Amon et al. 2006; van der Weerden et al. 2014).
5 Conclusion
In this laboratory study with silt loam soil, leonardite combined with animal slurry reduced N2O and NH3 emissions, increased the retention of slurry-derived N in the soil, and reduced the formation of leachable NO3− more effectively than the same amount of wheat straw or sawdust. Overnight mixing of the slurry and the HCA reduced N loss in some treatments, but the effect was not consistent. The type of slurry (pig or cattle slurry) and application methods had a strong influence on the effect of the tested HCA on GHG emissions and N retention. We conclude that leonardite may be a suitable additive for reducing nutrient losses after organic fertilization with animal slurries under similar soil conditions, thus improving the efficiency of agricultural N use.
Our study proves that it is possible to reduce N losses after slurry application by using leonardite. However, since this experiment was conducted under laboratory conditions without plants, further research is required to confirm this under more realistic soil conditions with crops. Future research should also investigate the potential beneficial effects of leonardite on the long-term storage of animal slurries and evaluate whether the positive effects on soil N retention and N cycling can be harnessed in practice.
References
Aber JD, Magill A, McNulty SG, Boone RD, Nadelhoffer KJ, Downs M, Hallett R (1995) Forest biogeochemistry and primary production altered by nitrogen saturation Water. Air, Soil Pollut 85:1665–1670. https://doi.org/10.1007/bf00477219
Aita C, Recous S, Cargnin RHO, da Luz LP, Giacomini SJ (2012) Impact on C and N dynamics of simultaneous application of pig slurry and wheat straw, as affected by their initial locations in soil. Biol Fertil Soils 48:633–642. https://doi.org/10.1007/s00374-011-0658-x
Amon B, Kryvoruchko V, Amon T, Zechmeister-Boltenstern S (2006) Methane, nitrous oxide and ammonia emissions during storage and after application of dairy cattle slurry and influence of slurry treatment Agric. Ecosyst Environ 112:153–162. https://doi.org/10.1016/j.agee.2005.08.030
Bouwman AF, Boumans LJM, Batjes NH (2002) Estimation of global NH3 volatilization loss from synthetic fertilizers and animal manure applied to arable lands and grasslands Global Biogeochem Cycles 16 doi:https://doi.org/10.1029/2000gb001389
Brummer C et al (2008) Soil-atmosphere exchange of N2O and NO in near-natural savanna and agricultural land in Burkina Faso (W. Africa). Ecosystems 11:582–600. https://doi.org/10.1007/s10021-008-9144-1
Burger M, Jackson LE (2003) Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems. Soil Biol Biochem 35:29–36. https://doi.org/10.1016/s0038-0717(02)00233-x
Chantigny MH, Rochette P, Angers DA (2001) Short-term C and N dynamics in a soil amended with pig slurry and barley straw: a field experiment Can J. Soil Sci 81:131–137. https://doi.org/10.4141/s00-046
Chen DL, Sun JL, Bai M, Dassanayake KB, Denmead OT, Hill J (2015) A new cost-effective method to mitigate ammonia loss from intensive cattle feedlots: application of lignite Sci Rep 5 doi:https://doi.org/10.1038/srep16689
Chen YR (1986) Rheological Properties of Sieved Beef-Cattle Manure Slurry - Rheological Model and Effects of Temperature and Solids Concentration Agric Wastes 15:17–33. https://doi.org/10.1016/0141-4607(86)90123-x
Cowling EB, Erisman JW, Smeulders SM, Holman SC, Nicholson BM (1998) Optimizing air quality management in Europe and North America: justification for integrated management of both oxidized and reduced forms of nitrogen. Nitrogen, the Confer-N-S.
Firestone MK, Firestone RB, Tiedje JM (1980) Nitrous-oxide from soil denitrification - factors controlling its biological production. Science 208:749–751. https://doi.org/10.1126/science.208.4445.749
Harper SHT, Lynch JM (1982) The role of water-soluble components in phytotoxicity from decomposing straw. Plant Soil 65:11–17. https://doi.org/10.1007/bf02376798
Houba VJG, Temminghoff EJM, Gaikhorst GA, van Vark W (2000) Soil analysis procedures using 0.01 M calcium chloride as extraction reagent. Commun Soil Sci Plant Anal 31:1299–1396. https://doi.org/10.1080/00103620009370514
Hui CH, So MK, Lee CM, Chan GYS (2003) Nitrous oxide flux from landfill leachate-sawdust nitrogenous compost. Chemosphere 52:1547–1551. https://doi.org/10.1016/s0045-6535(03)00494-6
IPCC (2013) Climate Change 2013: The physical science basis. Climate Change 2013: The Physical Science Basis. doi:https://doi.org/10.1017/cbo9781107415324
Jensen LS (2013) Animal manure fertiliser value, crop utilisation and soil quality impacts. Animal manure recycling: treatment and management.
Joergensen RG (1996) The fumigation-extraction method to estimate soil microbial biomass: calibration of the k EC value Soil Biology. Biochemistry 28:25–31
Kirchmann H, Lundvall AJB (1993) Soils Fo. Relationship between N immobilization and volatile fatty acids in soil after application of pig and cattle slurry 15:161–164. https://doi.org/10.1007/bf00361605
Manzoni S, Porporato A (2009) Soil carbon and nitrogen mineralization: theory and models across scales. Soil Biol Biochem 41:1355–1379. https://doi.org/10.1016/j.soilbio.2009.02.031
Nguyen QV et al (2017) Effects of cattle slurry and nitrification inhibitor application on spatial soil O2 dynamics and N2O production pathways. Soil Biol Biochem 114:200–209. https://doi.org/10.1016/j.soilbio.2017.07.012
Quade M, Brüggemann N, Graf A, Vanderborght J, Vereecken H, Rothfuss Y (2018) Investigation of kinetic isotopic fractionation of water during bare soil evaporation. Water Resour Res 54:6909–6928
Rabalais NN (2002) Nitrogen in Aquatic Ecosystems Ambio 31(102–112):111
Reichel R, Hansch M, Bruggemann N (2017) Indication of rapid soil food web recovery by nematode-derived indices in restored agricultural soil after open-cast lignite mining. Soil Biol Biochem 115:261–264. https://doi.org/10.1016/j.soilbio.2017.08.020
Reichel R et al. (2018) Potential of wheat straw, spruce sawdust, and lignin as high organic carbon soil amendments to improve agricultural nitrogen retention capacity: an incubation study Frontiers in Plant Science 9 doi:https://doi.org/10.3389/fpls.2018.00900
Rochette P, Angers DA, Chantigny MH, Gagnon B, Bertrand N (2008) N2O fluxes in soils of contrasting textures fertilized with liquid and solid dairy cattle manures. Can J Soil Sci 88:175–187. https://doi.org/10.4141/cjss06016
Saguer E, Gispert MA (1996) N2O Emission in selected representative soils of the North-Eastern Iberian Peninsula. In: Van Cleemput O, Hofman G, Vermoesen A (eds) Progress in nitrogen cycling studies: proceedings of the 8th nitrogen workshop held at the University of Ghent, 5–8 September, 1994. Springer Netherlands, Dordrecht, pp 613–619. doi:https://doi.org/10.1007/978-94-011-5450-5_100
Schefe CR, Patti AF, Clune TS, Jackson R (2008) Organic amendments increase soil solution phosphate concentrations in an acid soil: a controlled environment study. Soil Sci 173:267–276. https://doi.org/10.1097/SS.0b013e31816d1e3b
Simmler M et al (2013) Lignite reduces the solubility and plant uptake of cadmium in pasturelands. Environ Sci Technol 47:4497–4504. https://doi.org/10.1021/es303118a
Sommer SG, Genermont S, Cellier P, Hutchings NJ, Olesen JE, Morvan T (2003) Processes controlling ammonia emission from livestock slurry in the field. Eur J Agron 19:465–486. https://doi.org/10.1016/s1161-0301(03)00037-6
Sørensen P (1998) Effects of Storage Time and Straw Content of Cattle Slurry on the Mineralization of Nitrogen and Carbon in Soil Biol Fertil Soils 27:85–91
Sun JL et al (2016) Effects of lignite application on ammonia and nitrous oxide emissions from cattle pens. Sci Total Environ 565:148–154. https://doi.org/10.1016/j.scitotenv.2016.04.156
Thomsen IK, Pedersen AR, Nyord T, Petersen SO (2010) Effects of slurry pre-treatment and application technique on short-term N2O emissions as determined by a new non-linear approach Agric. Ecosyst Environ 136:227–235. https://doi.org/10.1016/j.agee.2009.12.001
Tran CKT, Rose MT, Cavagnaro TR, Patti AF (2015) Lignite amendment has limited impacts on soil microbial communities and mineral nitrogen availability. Appl Soil Ecol 95:140–150. https://doi.org/10.1016/j.apsoil.2015.06.020
van der Weerden TJ, Luo JF, Dexter M (2014) Addition of straw or sawdust to mitigate greenhouse gas emissions from slurry produced by housed cattle: a field incubation study. J Environ Qual 43:1345–1355. https://doi.org/10.2134/jeq2013.11.0452
Vandre R, Clemens J, Goldbach H, Kaupenjohann M (1997) NH3 and N2O emissions after landspreading of slurry as influenced by application technique and dry matter-reduction. 1. NH3 emissions. Zeitschrift Fur Pflanzenernahrung Und Bodenkunde 160:303–307. https://doi.org/10.1002/jpln.19971600226
Velthof GL, Kuikman PJ, Oenema O (2003) Nitrous Oxide Emission from Animal Manures Applied to Soil under Controlled Conditions Biol Fertil Soils 37:221–230. https://doi.org/10.1007/s00374-003-0589-2
Wei J, Reichel R, Islam MS, Wissel H, Amelung W, Bruggemann N (2020) Chemical composition of high organic carbon soil amendments affects fertilizer-derived N2O emission and nitrogen immobilization in an oxic sandy loam Front Environ Sci 8 doi:https://doi.org/10.3389/fenvs.2020.00015
Zavalloni C, Alberti G, Biasiol S, DelleVedove G, Fornasier F, Liu J, Peressotti A (2011) Microbial mineralization of biochar and wheat straw mixture in soil: a short-term study. Appl Soil Ecol 50:45–51. https://doi.org/10.1016/j.apsoil.2011.07.012
Zerbe S (2020) Ecology of Central European forest. Vegetation ecology of Central Europe, vol 1. Plant Biosyst 154:129–129. https://doi.org/10.1080/11263504.2019.1701579
Acknowledgements
We would like to thank Franz Leistner, Martina Krause, and Andrea Ecker for their technical support.
Funding
Open Access funding enabled and organized by Projekt DEAL. This study was conducted as part of the INPLAMINT project of the BonaRes initiative, which is funded by the German Federal Ministry of Education and Research (BMBF, FKZ 031B0508A) and supported by the Chinese Scholarship Council (scholarship no. 201706340157).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cao, X., Reichel, R., Wissel, H. et al. High Carbon Amendments Increase Nitrogen Retention in Soil After Slurry Application—an Incubation Study with Silty Loam Soil. J Soil Sci Plant Nutr 22, 1277–1289 (2022). https://doi.org/10.1007/s42729-021-00730-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-021-00730-7