
Abstract
Multiple sclerosis (MS) is a chronic autoimmune disease of the central nervous system (CNS) characterized by extensive inflammation, demyelination, axonal loss and gliosis. Evidence indicates that mast cells contribute to immunopathogenesis of both MS and experimental autoimmune encephalomyelitis (EAE), which is the most employed animal model to study this disease. Considering the inflammatory potential of mast cells, their presence at the CNS and their stabilization by certain drugs, we investigated the effect of ketotifen fumarate (Ket) on EAE development. EAE was induced in C57BL/6 mice by immunization with MOG35-55 and the animals were injected daily with Ket from the seventh to the 17th day after disease induction. This early intervention with Ket significantly reduced disease prevalence and severity. The protective effect was concomitant with less NLRP3 inflammasome activation, rebalanced oxidative stress and also reduced T cell infiltration at the CNS. Even though Ket administration did not alter mast cell percentage at the CNS, it decreased the local CPA3 and CMA1 mRNA expression that are enzymes typically produced by these cells. Evaluation of the CNS-barrier permeability indicated that Ket clearly restored the permeability levels of this barrier. Ket also triggered an evident lymphadenomegaly due to accumulation of T cells that produced higher levels of encephalitogenic cytokines in response to in vitro stimulation with MOG. Altogether these findings reinforce the concept that mast cells are particularly relevant in MS immunopathogenesis and that Ket, a known stabilizer of their activity, has the potential to be used in MS control.
Similar content being viewed by others

Introduction
Multiple sclerosis (MS) is one of the most prevalent progressive pathologies of the central nervous system (CNS) and the leading cause of non-traumatic neurological disabilities in young adults [1,2,3]. MS clinical presentation often varies among patients; some present more cognitive dysfunctions while others develop physical disabilities such as ataxia, hemiparesis, paraparesis and visual impairment. Despite this clinical variability, it is basically considered an autoimmune inflammatory disorder of the CNS characterized by inflammation, demyelination and variable extension of axonal loss [4,5,6]. Despite some controversy, it is believed that myelin specific T cells play a major role in MS immunopathogenesis [5,6,7,8].
Much of what is known from MS pathogenesis and therapy comes from data of its corresponding animal model known as experimental autoimmune encephalomyelitis (EAE), that is induced in rodents, especially in mice strains, by immunization with myelin antigens associated with Complete Freund’s Adjuvant (CFA) in the presence of pertussis toxin [9,10,11]. Data derived from both human and experimental disease indicate that their pathological features are orchestrated, in general lines, by the following steps: peripheral myelin-reactive T cell activation [12]; massive infiltration of these cells into the CNS through the blood-CNS barrier [13]; T cell local reactivation by recognition of self-antigens presented by major histocompatibility complex (MHC) molecules at the surface of antigen presenting cells (APCs) [14]; persistent inflammation associated with inflammasome activation and oxidative stress [15]; demyelination and its axonal deleterious consequences [16, 17].
Great efforts to fully understand the cellular and molecular mechanisms involved in initiation and progression of MS/EAE had, initially, revealed the pivotal role of innate immune cells, such as neutrophils and macrophages, in the processes that culminate with neurological dysfunctions [18,19,20]. More recently, an emergent role in neuroinflammatory and neurodegenerative diseases has been attributed to mast cells [21,22,23]. Besides being associated with parasitic and allergic pathologies, these cells have also been linked to a plethora of other conditions, such as metabolic disorders [24], pathological fibrosis [25], infectious [26] and autoimmune [27, 28] pathologies. Strong evidence supports the contribution of mast cells to MS immunopathogenesis, as for example, their accumulation in patient’s demyelinating plaques [29], increased mast cell transcripts within and outside patient’s lesions [30] and the presence of tryptase, which is a mast cell typical enzyme, in the patient’s cerebrospinal fluid [31]. Evidence also indicates their relevant contribution to EAE development by taking part in several pathological steps, such as peripheral immune activation, access to the CNS and neurodegenerative process [32,33,34,35,36].
Mast cells are found in practically all vascularized tissues, especially at sites that have contact with the external environment [37]. In the brain, up to 97% of the mast cells reside on the abluminal side of the blood vessels [38]. Their most well recognized characteristic is the presence of cytoplasmic granules filled with a variety of biologically active molecules such as enzymes, bioactive amines, certain cytokines, heparin and proteoglycan [39]. The secretion of this wide range of mediators seems to be the main mechanism used by mast cells to orchestrate many pathological processes, including MS [40, 41]. Considering the pathological potential of these mediators [42,43,44], we presumed that mast cell stabilizers could be useful for therapeutic interventions. Ketotifen fumarate (Ket) is a strong mast cell stabilizer [45] and a first-generation antihistamine drug widely employed to treat some allergic diseases [46, 47] and other inflammatory pathologies such as gout, diabetes, thyroiditis and uveitis [48,49,50,51,52,53]. Given this background, we assumed that this drug could be potentially therapeutic in MS. In this context and considering that this issue was not experimentally addressed yet, we investigated the effect of Ket on EAE development.
Methods
Experimental Design
To investigate the effect of Ket administration on EAE development, mice (eight to nine weeks-old) were allocated into three groups: mice submitted to EAE induction (EAE group); EAE-induced mice injected with Ket from the seventh to the 17th day after EAE induction (EAE/Ket group); and normal mice (control group - Ctrl). Body weight was evaluated daily while clinical scores were checked from day 10 on. The CNS-blood barrier permeability test was done at EAE clinical onset (11th to 12th day). Other parameters were all evaluated at the 18th day when it was expected that most of the EAE mice were already clinically sick. The following tests were performed with CNS samples: histopathology, oxidative stress status, inflammasome activation and real time RT-PCR for mast cell enzymes, T cell transcriptional factors mRNA expression and flow cytometry. At the periphery, lymphocyte subsets, percentage of activated dendritic cells (DCs) and cytokine production by cell cultures were determined. The whole experimental procedure is illustrated in Fig. 1 and CNS segments used in each assay and genes and phenotypic markers adopted for cell identification are displayed in Table 1.

Schematic timeline of the general experimental procedure
Animals
Female C57BL/6 mice (5-6 weeks old) were purchased from Ribeirão Preto Medical School (University of São Paulo, Ribeirão Preto, SP, Brazil) and housed under specific pathogen-free conditions until the age of 8-9 weeks. Sterilized food and water were available ad libitum. All experimental procedures were performed in accordance with the ethical guidelines adopted by the National Council for the Control of Animal Experimentation (CONCEA, Brazil) and approved by the Ethics Committee on use of animals (CEUA - process number 638), Institute of Biosciences, São Paulo State University (Botucatu, SP, Brazil).
EAE induction and Ket Administration
Mice from EAE and EAE/Ket groups were subcutaneously immunized with 150 μg of MOG35–55 peptide (Genemed Synthesis Inc., San Antonio, TX, USA) emulsified in Complete Freund’s Adjuvant (CFA) (Sigma-Aldrich, St. Louis, MO, USA) containing 4 mg/mL of Mycobacterium tuberculosis (Difco, Detroit, MI, USA) according to Zorzella-Pezavento et al. [54]. These animals also received two intraperitoneal doses, 0 and 48 hours after immunization, of 200 ng of Bordetella pertussis toxin (Sigma-Aldrich). Onset and severity of the disease were daily evaluated through analysis of body weight and clinical score according to the following criteria: 0 – no symptoms, 1 – limp tail, 2 – hind legs weakness, 3 – partially paralyzed hind legs, 4 – complete hind leg paralysis, and 5 – complete paralysis/death. The animals of the EAE/Ket group were injected daily, from day 7 to day 17 after EAE induction, with intraperitoneal doses of 0.4 mg/kg of ketotifen fumarate (Sigma-Aldrich) diluted in saline solution [55].
Blood-CNS Barrier Permeability Assay
Assessment of blood-CNS barriers were performed based on NaFLu protocol described by Christy et al. [34]. On days 11 or 12 after EAE induction, mice were injected with 10% sodium fluorescein (NaFlu) solution (Sigma-Aldrich) by i.p route. Twenty minutes later, the animals were anesthetized with ketamine/xylazine, blood was collected by cardiac puncture with heparinized syringe and mice were then perfused with 10 mL of a cold saline solution. Brain and whole spinal cord, which were processed separately, were dissected, weighed and then macerated with 400 μL of saline solution. After centrifugation at 9000 rpm for 10 min at 22°C, supernatants and plasma were collected and placed in 96-well black plates to read in a Synergy 4 fluorimeter (BioTek Instruments, Winooski, VT, USA). Results were expressed in relative units of fluorescence (RFU). NaFlu uptake by the CNS was calculated using the following equation: (tissue sample RFU/tissue sample weight)/ (plasma RFU/blood volume).
Central Nervous System Processing
The CNS was processed in different ways according to the scheduled analyses. Seventeen days after EAE induction, animals were anesthetized with ketamine/xylazine and transcardially perfused with 10 mL of a cold saline solution. Lumbar spinal cord samples were fixed in 10% neutral buffered formalin for histopathological analysis or immediately stored at -80°C for further mRNA expression analysis. Brain and spinal cord (cervical and thoracic segments) were collected and then homogenized to determine the level of oxidative stress. Brain and spinal cord (cervical and thoracic segments) were used to isolate mononuclear cells for further cytometric analysis according to Mimura et al. [56]. This material was digested with 2.5 mg/mL of collagenase D (Roche Applied Science, Indianapolis, IN, USA) at 37°C for 45 min and macerated using cell strainers (70 μm). After washing with RPMI medium (Sigma-Aldrich), the cells were resuspended in 37% Percoll solution (GE Healthcare, Uppsala, SWE) and gently placed over 70% Percoll solution for further centrifugation at 950g for 20 min. The mononuclear cell ring was collected from Percoll interface, washed and resuspended in supplemented RPMI medium (1% gentamicin, 2% glutamine, 1% sodium pyruvate, 1% nonessential amino acids, and 10% fetal calf serum). The portions of the CNS used for each analysis are displayed in Table 1.
Histopathological Analysis
After anesthesia and perfusion, the lumbar spinal cord was carefully removed and fixed with 10% buffered formalin for 24 hours. Samples were stored in 70% alcohol. Samples were then dehydrated in progressive increasing concentrations of ethanol, cleared with two xylene solution changes and embedded with Paraplast (Sigma-Aldrich). Sections of 5 μm thickness were obtained in a semi-automatic Microtome (Leica Biosystems, Wetzlar, Germany), which were then stained with hematoxylin/eosin (H&E) or Luxol Fast Blue (LFB) protocols. To perform the H&E staining, sections were deparaffinized with xylene, hydrated with ethanol and rinsed in distilled water. Samples were then rinsed in hematoxylin solution for 90 seconds and washed with running tap water, rinsed in 100% ethanol and counterstained with eosin for 30 seconds. Slides were then dehydrated through alcohol changes and cleared in xylene. To perform LFB staining, samples were deparaffinized with three xylene changes (20 min each) and hydrated with ethanol. Sections were incubated overnight in LFB solution (Sigma-Aldrich) at 56°C. The excess stain was washed with 95% alcohol and rinsed in distilled water. Slides were differentiated in 0.05% lithium carbonate solution. The counterstaining was done using a periodic acid solution (PAS) staining protocol. Samples were oxidized in 5% PAS for 15 min, washed with tap water for 10 min and then rinsed in distilled water. Slides stained with H&E and LFB were analyzed using an optical microscope (Axiolab, Carl Zeiss, Oberkochen, GE).
Biochemical Analysis of Oxidative Stress
The level of oxidative stress in the CNS was assessed by biochemical quantification of lipid hydroxide and by determination of the enzymatic activity of catalase, superoxide dismutase and glutathione peroxidase, according to Romualdo et al. [57]. Briefly, 500 mg of CNS tissue (brain and cervical/thoracic segments of spinal cord) were mechanically homogenized in 0.1 M sodium phosphate buffer (pH 7.0) and centrifuged (12,000g, -4°C, 15 min). Supernatants were then used for biochemical analyzes. Lipid hydroxide levels were determined by Fe2+ oxidation reaction in the presence of 250 mM FeSO4, 25 mM H2SO4, 100 mM xylenol orange and 4 mM butylated hydroxytoluene in 90% methanol. Catalase activity was evaluated by sample incubation (0.5 mL) with hydrogen peroxide (30%) in phosphate buffer (pH 7.0). Superoxide dismutase activity was assessed in alkaline medium (pH 10) by measuring the inhibitory action of the enzyme over nitroblue tetrazolium reduction induced by hydroxylamine-derived free radicals. The activity of glutathione peroxidase was assessed by NADPH2 oxidation reaction in the presence of glutathione reductase and hydrogen peroxide. All measurements were performed by using a microplate reader (mQuant-Gen5 2.0 software, BioTek Instruments, Winooski, VT, USA).
Spleen and Lymph Node Assays
Spleen and lymph nodes (axillary, brachial and inguinal) were dissected 17 days after EAE induction and macerated by using cell strainers (70 μm) to obtain cell suspensions. Red blood cells from splenic samples were lysed with a solution of NH4Cl. Spleen and lymph node cells from EAE and EAE/Ket groups were plated and restimulated in vitro with MOG for further cytokine quantification. Lymph node cells were also evaluated concerning the percentage of lymphocyte populations and of activated DCs by flow cytometry and also the presence of T cell subsets by real time RT-PCR.
Cytokine Quantification
Levels of TNF-α, IL-6, IFN-γ, IL-17, IL-5 and IL-10 were measured in culture supernatants from spleen (5 x 106 cells/mL) or lymph nodes (2.5 x 106 cells/mL). Cell cultures were restimulated in vitro with MOG35–55 (20 μg/mL) for 48 hours and cytokines were quantified by enzyme-linked immunosorbent assay (BD Biosciences, San Diego, CA, USA and R&D Systems, Minneapolis, MN, USA), according to the manufacturer’s instructions. To test a possible direct Ket effect on effector anti-MOG T lymphocytes, spleen cells (5 x 106 cells/mL) from EAE mice were in vitro incubated with Ket (10 μM) for 1 hour and then stimulated with 20 μg/mL of MOG35–55. Cytokines were quantified as described above.
Analysis of mRNA Expression by Real Time PCR
Gene expression analysis using real time RT-PCR was performed for quantification of Th subset transcription factors (T-bet, GATA-3 and Foxp3), nuclear receptor RORγt, mast cell specific enzymes (CMA-1 and CPA3), as specified in Table 1, and inflammasome components (NLRP3, caspase-1 and IL-1β). Briefly, RNA extraction from lymph node cells and lumbar spinal cord was carried out using Trizol reagent (Life Technologies, Austin, TX, USA) and single strand cDNA synthesis was performed from 1μg of RNA samples by using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster city, CA, USA). Five μl of cDNA (20-fold diluted) were submitted to real time PCR reaction using TaqMantm Gene Expression Assay (Life Technologies) according to the manufacturer’s instructions. The following inventoried primers were used: GAPDH (NM_008084.2), Tbx21/ T-bet (Mm00450960_m1), GATA-3 (Mm00484683_m1), RORc/ROR-γt (Mm01261022_m1), Foxp3 (Mm00475162_m1), CMA1 (Mm00487638_m1), CPA3 (Mm00483940_m1), Nlrp3 (Mm00840904_m1), Casp1 (Mm00438023_m1) and Il1b (Mm00434228_m1). Results were expressed based on fold difference by using the delta threshold (2-∆∆Ct) method that was originally described by Livak [58]. Genes used to identify each cell type are found in Table 1.
Flow Cytometry Analysis
Flow cytometry analyses were performed with lymph node cells and CNS-derived mononuclear cells collected as previously described. Red blood cells were lysed with NH4Cl solution and suspensions containing 1 x 106 cells were incubated with the following fluorochrome-labeled antibodies (eBioscience, San Diego, CA, USA): anti-CD45-PeCy7 (30-F11), anti-CD45-APC (30-F11), anti-CD3-PercPCy5.5 (145-2C11), anti-CD4-FITC (GK1.5), anti-CD8-APC (53-6.7), anti-CD8-PE (53-6.7), anti-CD19-PE (MB19-1), anti-CD11c-FITC (N418), anti-CD11b- APC efluor780 (m1/70), anti-F4/80-PerCPCy5.5 (BM8), anti-CD117-FITC (2B8), and anti-FcεRI-PECy7 (MAR-1). After being washed with the staining buffer, the cells were fixed in paraformaldehyde 1% and acquired using FACS Canto II flow cytometer (BD Biosciences) and FACS Diva software (BD Biosciences). Acquired data were analyzed with FlowJo software (Tree Star, Ashland, OR, USA). Phenotypic markers used to identify each cell type are displayed in Table 1.
Statistical Analysis
The results were initially analyzed by Kolmogorov-Smirnov normality test followed by Levene's test, except for EAE prevalence that was compared by Fischer's test (days 11 and 12) or by chi-squared test (days 13-18), and for clinical score data which was only analyzed by Mann-Whitney test. Other statistical comparisons were performed following the criteria: Student’s T-test or one-way ANOVA, respectively, for comparison of two or three groups presenting parametric data; Mann-Whitney or Kruskal-Wallis test, respectively, for comparison of two or three groups presenting non-parametric data. Significance level was p<0.05 for all statistical analyses. The following softwares were used for statistical analysis and graph composition: GraphPad Prism 5.0 (GraphPad, San Diego, CA, USA), SigmaPlot 12.0 (Systat Software, San Jose, CA, USA) and Statistica 10 (StatSoft, Tulsa, OK, USA). Data were presented as median and interquartile range. Each dot represents an animal.
Results
EAE Prevalence and Severity are Controlled by Early Intervention with Ketotifen
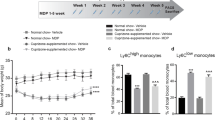
Female C57BL/6 mice were submitted to EAE induction and then injected daily with Ket from day 7 to 17 post-induction. This early intervention with Ket clearly affected prevalence and clinical signs. Disease prevalence was significantly lower in EAE/Ket than in EAE (disease control group), as depicted in Fig. 1A and B. Disease prevalence in the EAE group increased steadily until 87.5% at day 18, whereas the percentage of sick animals in the EAE/Ket group, at the same period, was only 35.7%.
This protective effect was also very clear when typical clinical signs, such as clinical score and body weight loss, were evaluated. The lower disease severity in the EAE/Ket group was ascertained by the most commonly used analyses as area determination (Fig. 2C), cumulative clinical score (Fig. 2D) and maximum clinical score (Fig. 2E). Information concerning prevalence and maximum clinical scores reached by EAE and EAE/Ket animals used in Figs. 2, 3, 4, 5, 6, 7, and 8 are indicated in the supplementary material. Body weight loss, clearly accentuated in the EAE group, was much more discrete in the EAE/Ket animals (Fig. 2F), with its variation in this group similar to the variation observed in normal mice (Fig. 2G).

Early intervention with Ket reduces EAE prevalence and severity. EAE was induced in C57BL/6 mice by subcutaneous immunization with MOG35–55 peptide plus CFA containing 4 mg/mL of Mycobacterium tuberculosis, and two intraperitoneal (ip.) doses of pertussis toxin. Ket adminstration (0.4 mg/kg) was performed by ip. route from the 7th to the 17th day after EAE induction. Body weight was measured every day, while clinical score was checked daily from day 10 until day 18 post-induction. (A) EAE prevalence data, (B) daily values of disease prevalence, (C) daily values of EAE clinical score, (D) cumulative EAE score, (E) maximum clinical score reached during the experiment, (F) daily values of body weight and (G) body weight variation during the experimental period. Results were obtained in five independent experiments (3-6 animals per group per experiment, EAE n=24 and EAE/Ket n=28). Data from 18 normal mice (Ctrl group) were also included in body weight illustrations. Data were expressed as median and interquartile range. Each dot represents an animal. *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001, vs EAE

Histopathological illustration of inflammatory and demyelinating lesions in EAE and EAE/Ket groups. EAE induction, Ket administration and perfusion were performed as previously described. Lumbar spinal cord samples were stained with H&E or LFB to analyze the inflammatory infiltration and demyelination, respectively, by means of optical microscopy. Representative photomicrographs (40x, 100x and 400x magnification) of samples from (A) EAE and (B) EAE/Ket (sick and non-sick animals)

Early intervention with Ket reduces oxidative stress status and inflammasome activation at the CNS. EAE induction, Ket administration and perfusion were performed as previously described. CNS was collected and the following tests were done: (A) mRNA expression for NLRP3, (B) caspase-1 (CASP-1) and (C) IL-1β were evaluated in lumbar spinal cord samples by real time RT-PCR. Results from two independent experiments (3-4 animals per group per experiment) are shown. (D) Levels of lipid hydroxide, (E) catalase, (F) superoxide dismutase and (G) glutathione peroxidase were assessed by biochemical analysis. Data are expressed as median and interquartile range. Each dot represents an animal. **p<0.01, ***p<0.001 and ****p<0.0001, vs EAE. a*** indicates statistical significance between EAE/Ket and Ctrl group (p<0.001)

Early intervention with Ket reduces the amount of T cells, especially Th1, in the CNS. EAE induction, Ket administration and perfusion were performed as previously described. Mononuclear cells were eluted from the brain and cervical and thoracic segments of spinal cord. (A) Number of cells with lymphocytic morphology eluted from CNS was determined by using Türk’s solution and optical microscopy. (B) Percentages of infiltrating leukocytes (CD45high), (C) total (CD45highCD3+), (D) CD4+ (CD45highCD4+CD8-) and (E) cytotoxic T lymphocytes (CD45highCD4-CD8+) were evaluated by flow cytometry (100,000 events). (F) mRNA expression for T-bet, (G) RORc, (H) GATA-3 and (I) Foxp3 was evaluated in the lumbar segment of the spinal cord by real time RT-PCR. Results were obtained from two independent experiments (3-6 animals per group per experiment). Data are expressed as median and interquartile range. Each dot represents an animal. *p<0.05 and **p<0.01, vs EAE

Early intervention with Ket downregulates local mRNA expression for mast cell enzymes and preserves blood-CNS barrier permeability. EAE induction, Ket administration and perfusion were performed as previously described. (A) Percentage of mast cells (CD45highCD117+FcεRI+) by flow cytometry (100,000 events) and mRNA expression for (B) CPA3 and (C) CMA1 in lumbar segment of spinal cord by real time RT-PCR. (D) For blood-CNS barrier permeability determination, Ket was administered from day 7 until days 11 or 12 (disease onset), when animals were intraperitoneally injected with 10% sodium fluorescein (NaFlu) solution. Cardiac blood and CNS samples were collected. NaFlu uptake was calculated by the relative fluorescence units (RFU) emitted by tissue samples normalized by their weight, and RFU emitted by corresponding blood sample normalized by the amount of cardiac blood. Readings were performed in a Synergy 4 fluorimeter (BioTek Instruments) with excitation/emission wavelengths of 485/520 nm. Results obtained from two independent experiments (3-5 animals per group per experiment) are shown. Data are expressed as median and interquartile range. Each dot represents an animal. **p<0.01, vs EAE

Early intervention with Ket triggers lymphocyte accumulation in draining lymph nodes. EAE induction, Ket administration and perfusion were performed as previously described. Draining lymph nodes (axillary, brachial and inguinal) were collected. (A) Photograph representing lymph node size collected from EAE and EAE/Ket mice and (B) comparison of cell number with lymphocytic morphology. Percentage of total (C) (CD3+), (D) CD4 (CD3+CD4+CD8-) and (E) cytotoxic (CD3+CD4-CD8+) T lymphocytes, (F) B (CD19+) cells and (O) DCs (F4/80-CD11c+MHCIIhigh) expressing high levels of MHCII molecules was evaluated by flow cytometry (at least 100,000 events). Estimated amount of (G) T cells, (H) Th cells, (I) Tc cells and (J) B cells. mRNA expression for (K) T-bet, (L) GATA-3, (M) RORc and (N) Foxp3 was evaluated by real time RT-PCR. Results obtained from two or three independent experiments (3-6 animals per group per experiment) are shown. Data are expressed as median and interquartile range. Each dot represents an animal. **p<0.01 and ***p<0.001, vs EAE

Cytokine production is affected by in vivo but not by in vitro Ket addition. EAE induction, Ket administration and perfusion were performed as previously described. Draining lymph nodes and spleen cells were collected. (A-F) Lymph node cells from EAE and EAE/Ket animals were restimulated in vitro with MOG35–55 (20 μg/mL) over 48 hours. (G-L) Splenocytes from EAE-induced mice were also restimulated with MOG35–55 (20 μg/mL), (M-R) in the presence or absence (C-) of Ket (10 μM) over 48 hours. Levels of (A,G,M) IL-17, (B,H,N) IFN-γ, (C,I,O) IL-6, (D,J,P) TNF-α, (E,K,Q) IL-5, and (F,L,R) IL-10 in culture supernatants were assessed by ELISA. (A-F) One representative experiment of three is shown (4-6 animals per group). (G-R) Results were obtained from two independent experiments (3-4 animals per group). Data are expressed as median and interquartile range. Each dot represents an animal. *p<0.05 and *p<0.01, vs EAE
To check if full protection was due to absence of cell infiltration at the CNS, H&E stained sections from the lumbar spinal cord were analyzed for the presence of inflammation. As expected, mononuclear cell infiltrates were observed in all animals of the EAE group that developed the disease (87.5%), predominantly on the periphery of the tissue. On the other hand, the animals of the EAE/Ket group that did not develop the disease (64.3%) presented no local inflammation. As expected, EAE/Ket mice that were not fully protected developed signs of paralysis and also presented inflammation and demyelination. Existence of inflammation in the lumbar spinal cord of EAE and EAE/Ket sick mice and its absence in EAE/Ket healthy animals is illustrated by representative photomicrographs (Fig. 3). As anticipated, the inflamed regions were co-located with demyelinated areas in the EAE animals as shown in Fig. 3A. No demyelination was observed in EAE/Ket healthy animals (Fig. 3B).
Impact of Early Intervention with Ketotifen on Inflammasome and Oxidative Stress at the CNS
Evidence from MS patients and EAE highly suggest that inflammasome activation occurs during disease and that the NLRP3 inflammasome is essential for disease development [59]. Ket administration triggered a significant reduction of mRNA expression for NLRP3, caspase-1 and IL-1β as illustrated in Fig. 4A, B and C, respectively. Since oxidative stress has been related to inflammasome activation [60], and to both disruption of the blood-CNS barrier and killing of oligodendrocytes [61], its status at the CNS was also checked. Lipid hydroxide was lower whereas antioxidative enzymes such as catalase, superoxide dismutase and glutathione peroxidase were all significantly elevated in Ket-administered mice as shown in Fig. 4D, E, F and G, respectively.
A Smaller Amount of T Cells, Especially Th1, are Found in the CNS of EAE/Ket Mice
It is well established that lymphocytes play a major role in the autoimmune process that characterizes MS [62]. In this context, we investigated if Ket administration affected the amount and the differentiation of these cells in the CNS. Initially, by using light microscopy to count the total number of cells eluted from the CNS, we found a significantly lower number in the EAE/Ket group as indicated in Fig. 5A. Flow cytometry analysis confirmed that Ket indeed decreased the percentage of leukocytes that reached the CNS as indicated by Fig. 5B and also revealed that Th, but not Tc cells, were significantly down-regulated, as shown in Fig. 5C, D and E. By RT-PCR we verified that Ket drastically reduced the amount of Th1 and Treg cells but not of Th17 and Th2 cells as indicated in Fig. 5F, I, G and H, respectively.
Ketotifen Downregulates Local Expression of Mast Cell-specific Enzymes and Preserves Blood-CNS Barrier Permeability
Mast cells are widely distributed and also found at the CNS where they are able to interfere in many local functions by signaling to blood vessels, neurons and microglia [63]. In this context, we investigated if Ket was modulating the local amount or activity of these cells. Similar percentages of mast cells (CD45highCD117+FcεRI+ cells) were found in the CNS of EAE and EAE/Ket experimental groups (Fig. 6A). However, mRNA expression encoding enzymes stored in mast cell granules as CPA3 and CMA1, was largely downregulated by Ket as illustrated at Fig. 6B and C, respectively.
Taking into consideration that mast cell enzymes have been implicated in blood-brain barrier and blood-spinal cord barrier disruption [64], the possible effect of Ket on blood-CNS barriers was tested. As already described by other authors, the NaFlu test indicated that the permeability of this barrier was highly increased in EAE mice. On the other hand, Ket administration was able to keep blood-CNS barrier function at the level found in healthy mice (Fig. 6E).
Ketotifen Triggers Lymphocyte Accumulation in Draining Lymph Nodes
To evaluate the possibility of lymphocyte sequestration at peripheral lymphoid organs as is triggered by fingolimod [65], the amount of cells present in the draining lymph nodes and the size of these organs were assessed at the 17th day after disease induction. A visible lymph node enlargement was observed at this active disease phase and the number of cells with lymphocytic morphology in these organs was significantly higher in the EAE/Ket group as depicted in Fig. 7A and B, respectively.
Flow cytometry analysis indicated similar percentages of CD3+ (Fig. 7C), CD3+CD4+CD8- (Fig. 7D), CD3+CD4-CD8+ T lymphocytes (Fig. 7E), and also of CD19+ B cells (Fig. 7F). Despite the similar proportion in these lymphocyte subsets, lymph nodes from Ket-injected mice contained a much higher estimated number (calculated by using the mathematical rule of three) of CD3+ (Fig. 7G) and CD3+CD4+CD8- T cells (Fig. 7H). CD3+CD4-CD8+ T lymphocytes (Fig. 7I), and CD19+ B cells (Fig. 7J) were slightly increased. Flow cytometry was also used to determine the percentage of DCs expressing high levels of MHC II molecules and indicated that the total percentage of these cells was, likewise, much higher in EAE/Ket mice (Fig. 7O).
Ketotifen Effect on Cytokine Production: In vivo and In vitro Results
Cells from draining lymph nodes and spleen, obtained 18 days after EAE induction, were initially restimulated with MOG in vitro to evaluate the intensity of the cellular immune response to the specific antigen. Lymph nodes and spleen cell cultures presented a very similar pattern characterized by higher cytokine production by EAE/Ket mice as illustrated at Fig. 8A-L. Unlike these results, in vitro treatment of spleen cells from EAE mice with Ket, before stimulation with MOG, did not alter the level of cytokine production by these cells (Fig. 8M, N, O, P, Q and R).
Discussion
Mast cells stabilizers are widely employed to treat allergic diseases because they avoid the release of a plethora of proinflammatory mediators [42]. More recently, the discovery that mast cells are possibly also involved in the genesis and perpetuation of autoimmunity [28, 66], opened a new perspective in the understanding of these inflammatory pathologies. In this context, we asked if mast cells could be a relevant target for MS therapy. To test this possibility, we evaluated the effect of Ket, a largely employed mast cell stabilizer, in the evolution of experimental autoimmune encephalomyelitis (EAE). Ket administration was initiated 6 days after EAE induction when a very strong anti-MOG immune response is already established in this mice model [10]. In our opinion, this experimental protocol represents an early intervention in the disease, therefore distinct from both the prophylactic and therapeutic classical procedures.
This early intervention clearly triggered a protective effect characterized by lower disease prevalence and severity. Ket traditional protective effects have been described in various non-autoimmune conditions including both experimental and human diseases such as allergic rhinitis and conjunctivitis [67, 68], gout [69], type II diabetes [51] and, more recently, even in a dengue disease model [70]. Only a few reports suggest that it could be also effective in some autoimmune pathologies as human thyroiditis [52] and experimental uveoretinitis [53]. Our investigation is, therefore, the first report showing that Ket significantly controlled EAE development. When disease control was absolute, that is, when EAE/Ket animals did not show any paralysis sign, no inflammation or demyelination was found at the lumbar spinal cord level, as is in accordance with the pathognomonic role of these lesions in both EAE and MS [71,72,73]. Otherwise, when EAE/Ket animals were not fully protected, they presented inflammatory infiltration at the lumbar spinal cord.
Once it was established that Ket was able to reduce disease prevalence and severity, various assays were performed to better characterize this protection. These ensuing tests were always done with all the animals of the group, independently of their degree of protection; meaning therefore that the findings described in this paper represent a median of all animals tested in each experimental group.
It is well established that local oxidative stress and inflammasome activation are implicated in a self-perpetuating cycle that triggers and maintains neurodegeneration in many CNS pathologies [74, 75]. In fact, evaluation of oxidative stress at the CNS indicated that Ket was somehow able to hamper this process. EAE mice presented an expected unbalanced status characterized by high levels of lipid hydroxide, an oxidative stress marker, together with low levels of glutathione peroxidase, catalase and superoxide dismutase. Contrastingly, the EAE/Ket group showed a reversal scenario marked by an elevated production of antioxidant enzymes. Even though the start of this neurodegenerative cycle involving oxidative stress and inflammasome is not completely understood, it is accepted that reactive oxygen species (ROS) are some of the host-derived molecules that can elicit inflammasome activation [60, 76]. In this regard, it is conceivable that oxidative molecules released by mast cells are implicated in the beginning of this neurodegenerative process. Some support for this hypothesis is provided in the literature. It is known, for example, that ROS participate in mast cell intracellular signaling [77] and that mast cell degranulators such as acrolein, for example, are able to promote ROS generation [78]. In addition, it was recently demonstrated that Ket was able to control the oxidative damage determined by the compound 48/80, which is a well-known mast cell activator, in rat livers [79]. We also found out that the inflammasome platform was significantly downregulated by Ket therapy, as indicated by reduced levels of NLRP3, caspase-1 and IL-1β mRNA. This reduction in oxidative stress and in the inflammasome platform was clearly coincident with a diminished cell infiltration at the CNS, especially concerning T cells polarized towards Th1 profile that have been described as pivotal players in both MS and EAE [4, 18]. A likely connection between our findings and mast cell stabilization by Ket finds support in the enlightening model outlined by Russi et al. [80] and Russi et al. [81] to illustrate the role of mast cells in EAE. According to them, mast cell-derived IL-1β is fundamental to license T cells to become encephalitogenic. Briefly, they propose that peripheral autoreactive primed T cells transiting through the meninges interact or activate mast cells eliciting IL-1β release that, in turn, licenses T cells to produce GM-CSF that will, ultimately, allow them to traffic to the CNS and coordinate the local inflammatory process. Interestingly, we found no local increase of Th2 and Tregs in the EAE/Ket mice, suggesting therefore that this drug was not acting through induction of these potentially regulatory T cells.
It is well established that leukocyte infiltration at the CNS mainly occurs throughout two barriers: the blood-brain barrier and the blood-spinal cord barrier. Much scientific effort has been driven to clarify their dysfunction in the course of EAE and MS. The contribution of the endothelial NMDA receptors [82], micro RNAs [83] and extracellular vesicles [84] are some of the more recent investigations in this regard. Restoration of these barriers have been a very promising strategy to control MS and other neurological pathologies [85, 86]. As mast cells have been considered crucial regulators of this barrier permeability [87], the amount and activity of these cells was checked at the CNS. Ket administration did not affect their local number but clearly downmodulated their biological activity. This was ascertained by the demonstration that this drug inhibited the expression of mRNA for CPA-3 and CMA1 whose products are present in high levels in mast cell granules [88]. There are no straightforward demonstrations in the literature concerning the interaction between Ket and these CNS barriers. However, there is an expressive amount of data revealing that mast cells can release vasoactive mediators able to promote endothelial activation leading to CNS barrier dysfunction [89,90,91,92]. A plethora of mast cell-derived molecules are presumably involved in its disruption as histamine, heparine, serotonin, nitric oxide, vasoactive intestinal peptide and vascular endothelial growth factor. Mast cells-specific proteolytic enzymes such as MMP-2 and MMP-9 are also able to damage microvascular basal lamina components leading to the disruption of tight junction proteins [93]. Interestingly, MMPs are also released by polymorphonuclear neutrophils (PMN) that play a substantial role in EAE and MS immunopathogenesis [94]. In this context, Ket could control EAE via mast cell stabilization and subsequent inhibition of release of chemotactic mediators for PMN as already demonstrated in a model of inflammatory skin rejection [95]. A direct inhibitory effect of Ket in the release of MMPs by PMN cannot be excluded.
Having observed that the lymph nodes from EAE/Ket animals presented a visible hypertrophy, the possibility that autoreactive cells were being trapped and continually activated at the peripheral lymphoid organs was checked. Even though the proportion of T, Th, Tc and B cells was similar in the lymph nodes of both experimental groups, the estimated number of T and Th populations was significantly elevated in the EAE/Ket group. The percentage of DCs expressing high levels of MHC II levels, which indicates a higher capacity for antigen presentation [96], was also more elevated in the EAE/Ket group, suggesting the presence of a more active peripheral immune response in these mice. This lymphadenomegaly was, at least partially, due to local accumulation of MOG-specific cells able to produce significantly higher levels of IL-17 and IFN-γ that are considered pivotal encephalitogenic cytokines [97, 98]. Lymphocyte sequestration in peripheral lymphoid organs has already been characterized as an important mechanism of action of fingolimod that is frequently adopted in MS therapy mainly for patients with the relapsing-remitting course [99,100,101].
Even though spleen cells from the EAE/Ket group also released higher cytokine amounts than in the EAE group, the in vitro addition of Ket to spleen cell cultures from EAE mice did not change the types nor the amount of cytokines. These findings suggest that this elevated cytokine production observed in the EAE/Ket mice is not the consequence of a direct interaction between Ket and lymphocytes. Otherwise, these results reinforce the accumulation of MOG-specific cells at these peripheral lymphoid organs.
A challenging feature of our findings is some variability in terms of protection, that is, some animals were fully protected while others showed some signs of paralysis. This could be interpreted as a consequence of the intrinsic variability of the EAE model. However, other factors such as Ket optimal concentration for each animal, mainly the level achieved at the CNS, and the severity of the experimental disease, could all contribute to the protection level. The complete understanding of this phenomenon certainly deserves further and thorough investigation.
The control of inflammation in MS by targeting mast cells has been proposed and scientifically supported by recognized experts in this field [21, 34, 64, 81, 89]. We believe that our results further strengthen the rationale for exploring this and other mast cell stabilizers in the therapy of MS. As suggested by our findings and highlighted by Hart [102], administration of this and other mast cell targeted drugs will probably be more beneficial at earlier disease phases. Association of this or similar pharmaceuticals and other already classical drugs used in MS are worthwhile to be tested, considering that their associations could have a much stronger therapeutic potential. Another aspect to be considered is the expressive experience with the use of this and other related drugs in human beings. Ket and other mast cell stabilizers have successfully controlled other pathologies such as asthma, allergic conjunctivitis, atopic eczema and chronic urticaria [103] without eliciting major side effects such as, for example, global immunosuppression [70, 104].
Considering the literature and these experimental findings, it seems clear that mast cells play a pivotal contribution to EAE and likely MS, and should, therefore, be considered as potential targets for therapeutic intervention in this pathology.
References
Howard J, Trevick S, Younger DS. Epidemiology of multiple sclerosis. Neurol Clin. 2016;34: 919-939.
Reich DS, Lucchinetti CF, Calabresi PA. Multiple sclerosis. N Engl J Med. 2018;378:169-180.
Browne P, Chandraratna D, Angood C, et al. Atlas of multiple sclerosis 2013: a growing global problem with widespread inequity. Neurology 2014;83:1022-1024.
Dendrou CA, Fugger L, Friese MA. Immunopathology of multiple sclerosis. Nat Rev Immunol. 2015;15:545-558.
Garg N, Smith TW. An update on immunopathogenesis, diagnosis, and treatment of multiple sclerosis. Brain Behav. 2015;5:e00362.
Filippi M, Bar-Or A, Piehl F, Preziosa P, Solari A, Vukusic S, Rocca MA. Multiple sclerosis. Nat Rev Dis Primers. 2018;4:43.
Holmoy T. Immunopathogenesis of multiple sclerosis: concepts and controversies. Acta Neurol Scand. 2007;187:39-45.
Fletcher JM, Lalor SJ, Sweeney CM, et al. T cells in multiple sclerosis and experimental autoimmune encephalomyelitis. Clin Exp Immunol. 2010;162:1-11.
Miller SD, Karpus WJ, Davidson TS. Experimental autoimmune encephalomyelitis in the mouse. Curr Protoc Immun 2010;88:15.1.1-15.1.20.
Constantinescu CS, Farooqi N, O'Brien K, et al. Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Br J Pharmacol. 2011;164:1079-1106.
Lassmann H, Bradl M. Multiple sclerosis: experimental models and reality. Acta Neuropathol. 2017;133:223-244.
Almolda B, Gonzalez B, Castellano B. Activated microglial cells acquire an immature dendritic cell phenotype and may terminate the immune response in an acute model of EAE. J Neuroimmunol. 2010;223:39-54.
Schulz M, Engelhardt B. The circumventricular organs participate in the immunopathogenesis of experimental autoimmune encephalomyelitis. Cerebrospinal Fluid Res. 2005;2:8.
Flach AC, Litke T, Strauss J, et al. Autoantibody-boosted T-cell reactivation in the target organ triggers manifestation of autoimmune CNS disease. Proc Natl Acad Sci. USA. 2016;113:3323-3328.
Freeman LC, Ting JP. The pathogenic role of the inflammasome in neurodegenerative diseases. J Neurochem. 2016;1:29-38.
Osorio-Querejeta I, Saenz-Cuesta M, Munoz-Culla M, et al. Models for studying myelination, demyelination and remyelination. Neuromolecular Med. 2017;19:181-192.
Lubetzki C, Stankoff B. Demyelination in multiple sclerosis. Handb Clin Neurol. 2014;122: 89-99.
Hemmer B, Kerschensteiner M, Korn T. Role of the innate and adaptive immune responses in the course of multiple sclerosis. Lancet Neurol. 2015;14:406-419.
Hernandez-Pedro NY, Espinosa-Ramirez G, de la Cruz VP, Pineda B, Sotelo J. Initial immunopathogenesis of multiple sclerosis: innate immune response. Clin Dev Immunol. 2013;2013:413465.
Gandhi R, Laroni A, Weiner HL. Role of the innate immune system in the pathogenesis of multiple sclerosis. J Neuroimmunol. 2010;221:7-14.
Girolamo F, Coppola C, Ribatti D. Immunoregulatory effect of mast cells influenced by microbes in neurodegenerative diseases. Brain Behav Immun. 2017;65:68-89.
Hendriksen E, van Bergeijk D, Oosting RS, et al. Mast cells in neuroinflammation and brain disorders. Neurosci Biobehav Rev. 2017;79:119-133.
Kempuraj D, Selvakumar GP, Zaheer S, et al. Cross-talk between glia, neurons and mast cells in neuroinflammation associated with Parkinson's disease. J Neuroimmune Pharmacol. 2017;13:100-112.
Bais S, Kumari R, Prashar Y, Gill NS. Review of various molecular targets on mast cells and its relation to obesity: A future perspective. Diabetes Metab Syndr. 2017;2:S1001-S1007.
Gieseck RL, Wilson MS, Wynn TA. Type 2 immunity in tissue repair and fibrosis. Nat Rev Immunol. 2018;18:62-76.
Nascimento CR, Andrade D, Carvalho-Pinto CE, Serra RR, et al. Mast cell coupling to the Kallikrein-Kinin system fuels intracardiac parasitism and worsens heart pathology in experimental Chagas disease. Front Immunol. 2017;8:840.
Yu X, Kasprick A, Petersen F. Revisiting the role of mast cells in autoimmunity. Autoimmun Rev. 2015;14:751-759.
Xu Y, Chen G. Mast cell and autoimmune diseases. Mediators Inflamm. 2015;2015:246126.
Kruger PG. Mast cells and multiple sclerosis: a quantitative analysis. Neuropathol Appl Neurobiol. 27;2001:275-280.
Couturier N, Zappulla JP, Lauwers-Cances V, et al. Mast cell transcripts are increased within and outside multiple sclerosis lesions. J Neuroimmunol. 2008;195:176-185.
Rozniecki JJ, Hauser SL, Stein M, Lincoln R, Theoharides TC. Elevated mast cell tryptase in cerebrospinal fluid of multiple sclerosis patients. Ann Neurol. 1995;37:63-66.
Secor VH, Secor WE, Gutekunst CA, Brown MA. Mast cells are essential for early onset and severe disease in a murine model of multiple sclerosis. J Exp Med. 2000;191:813-822.
Gregory GD, Robbie-Ryan M, Secor VH, et al. Mast cells are required for optimal autoreactive T cell responses in a murine model of multiple sclerosis. Eur J Immunol. 2005;35:3478-3486.
Christy AL, Walker ME, Hessner MJ, et al. Mast cell activation and neutrophil recruitment promotes early and robust inflammation in the meninges in EAE. J Autoimmun. 2013;42:50-61.
Hong GU, Kim NG, Jeoung D, et al. Anti-CD40 Ab- or 8-oxo-dG-enhanced Treg cells reduce development of experimental autoimmune encephalomyelitis via down-regulating migration and activation of mast cells. J Neuroimmunol. 2013;260:60-73.
Yin JJ, Hu XQ, Mao ZF, et al. Neutralization of Interleukin-9 Decreasing Mast Cells Infiltration in Experimental Autoimmune Encephalomyelitis. Chin Med J (Engl). 2017;130:964-971.
Voehringer D. Protective and pathological roles of mast cells and basophils. Nat Rev Immunol. 2013;13:362-375.
Khalil M, Ronda J, Weintraub M, Jain K, Silver R, Silverman AJ. Brain mast cell relationship to neurovasculature during development. Brain Res. 2007;1171:18-29.
Cildir G, Pant H, Lopez AF, Tergaonkar V. The transcriptional program, functional heterogeneity, and clinical targeting of mast cells. J Exp Med. 2017;214:2491-2506.
Vukman KV, Forsonits A, Oszvald A, et al. Mast cell secretome: Soluble and vesicular components. Semin Cell Dev Biol. 2017;67:65-73.
Theoharides TC, Alysandratos KD, Angelidou A, et al. Mast cells and inflammation. Biochem Biophys Acta 2012;1822:21-33.
Finn DF, Walsh JJ. Twenty-first century mast cell stabilizers. Br J Pharmacol. 2013;170:23-37.
Borriello F, Granata F, Varricchi G, et al. Immunopharmacological modulation of mast cells. Curr Opin Pharmacol. 2014;17:45-57.
Zhang T, Finn DF, Barlow JW, et al. Mast cell stabilisers. Eur J Pharmacol. 2016;778:158-168.
Baba A, Tachi M, Ejima Y, et al. Anti-allergic drugs tranilast and ketotifen dose-dependently exert mast cell-stabilizing properties. Cell Physiol Biochem. 2016;38:15-27.
Grant SM, Goa KL, Fitton A, et al. Ketotifen. A review of its pharmacodynamic and pharmacokinetic properties, and therapeutic use in asthma and allergic disorders. Drugs 1990;40:412-448.
Wade L, Bielory L, Rudner S. Ophthalmic antihistamines and H1-H4 receptors. Curr Opin Allergy Clin Immunol. 2012;12:510-516.
Hsu DZ, Chu PY, Chen SJ, et al. Mast cell stabilizer ketotifen inhibits gouty inflammation in rats. Am J Ther. 2016;23:1009-1015.
Lai DS, Lue KH, Hsieh JC, et al. The comparison of the efficacy and safety of cetirizine, oxatomide, ketotifen, and a placebo for the treatment of childhood perennial allergic rhinitis. Ann Allergy Asthma Immunol. 2002;89:589-598.
Chen Z, Sun H, Wang J, et al. Role of ketotifen on metabolic profiles, inflammation and oxidative stress in diabetic rats. Endocr J. 2017;64:411-416.
El-Haggar SM, Farrag WF, Kotkata FA. Effect of ketotifen in obese patients with type 2 diabetes mellitus. J Diabetes Complications 2015;29:427-432.
Karaayvaz M, Caliskaner Z, Turan M, et al. Levothyroxine versus ketotifen in the treatment of patients with chronic urticaria and thyroid autoimmunity. J Dermatolog Treat. 2002;13:165-172.
de Kozak Y, Sakai J, Sainte-Laudy J, et al. Pharmacological modulation of IgE-dependent mast cell degranulation in experimental autoimmune uveoretinitis. Jpn J Ophthalmol. 1983;27:598-608.
Zorzella-Pezavento SFG, Mimura LAN, et al. Experimental autoimmune encephalomyelitis is successfully controlled by epicutaneous administration of MOG plus vitamin D analog. Front Immunol. 2017;8:1198.
Kim MS, Lee DH, Lee CW, et al. Mast cell stabilizer, ketotifen, prevents UV-induced wrinkle formation. J Invest Dermatol. 2013;133:1104-1107.
Mimura LA, Chiuso-Minicucci F, Fraga-Silva TF, et al. Association of myelin peptide with vitamin D prevents autoimmune encephalomyelitis development. Neuroscience. 2016;317:130-140.
Romualdo GR, Goto RL, Fernandes AAH, et al. Vitamin D3 supplementation attenuates the early stage of mouse hepatocarcinogenesis promoted by hexachlorobenzene fungicide. Food Chem Toxicol. 2017;107:27-36.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real time quantitative PCR and the 22DDCT method. Methods. 2001;25:402-408.
Barclay W, Shinohara ML. Inflammasome activation in multiple sclerosis and experimental autoimmune encephalomyelitis (EAE). Brain Pathol. 2017;27:213-219.
Harijith A, Ebenezer DL, Natarajan V. Reactive oxygen species at the crossroads of inflammasome and inflammation. Front Physiol. 2014;5:352.
Wang HF, Liu XK, Li R, et al. Effect of glial cells on remyelination after spinal cord injury. Neural Regen Res. 2017;12:1724-1732.
Legroux L, Arbour N. Multiple sclerosis and T lymphocytes: An entangled story. J Neuroimmune Pharmacol. 2015;10:528-546.
Skaper SD, Giusti P, Facci L. Microglia and mast cells: two tracks on the road to neuroinflammation. FASEB J. 2012;26:3103-3117.
Elieh-Ali-Komi D, Cao Y. Role of mast cells in the pathogenesis of multiple sclerosis and experimental autoimmune encephalomyelitis. Clin Rev Allergy Immunol. 2017;52:436-445.
Gonzalez-Cabrera PJ, Cahalan SM, Nguyen N, et al. S1P(1) receptor modulation with cyclical recovery from lymphopenia ameliorates mouse model of multiple sclerosis. Mol Pharmacol. 2012;81:166-174.
Christy AL, Brown MA. The multitasking mast cell: positive and negative roles in the progression of autoimmunity. J Immunol. 2007;179:2673-2679.
Greiner JV, Minno G. A placebo-controlled comparison of ketotifen fumarate and nedocromil sodium ophthalmic solutions for the prevention of ocular itching with the conjunctival allergen challenge model. Clin Ther. 2003;25:1988-2005.
Akhavan A, Karimi-Sari H, Khosravi MH, Arefzadeh E, Yavarahmadi M. Comparing the effects of ketotifen fumarate eye drops and ketotifen oral pills on symptom severity and quality of life in patients with allergic rhinitis: a double-blind randomized clinical trial. Int Forum Allergy Rhinol. 2015;5:386-391.
Hsu DZ, Chu PY, Chen SJ, Liu MY. Mast cell stabilizer ketotifen inhibits gouty inflammation in rats. Am J Ther. 2016;23:e1009-1015.
Morrison J, Rathore APS, Mantri CK, Aman SAB, Nishida A, St John AL. Transcriptional profiling confirms the therapeutic effects of mast cell stabilization in a Dengue disease model. J Virol. 2017;91.
Gilmore CP, DeLuca GC, Bö L, et al. Spinal cord neuronal pathology in multiple sclerosis. Brain Pathol. 2009;19:642-649.
Gibson-Corley KN, Boyden AW, Leidinger MR, et al. A method for histopathological study of the multifocal nature of spinal cord lesions in murine experimental autoimmune encephalomyelitis. Peer J. 2016;26;4:e1600.
Schmierer K, Miquel ME. Magnetic resonance imaging correlates of neuro-axonal pathology in the MS spinal cord. Brain Pathol. 2018;28:765-772.
Ortiz GG, Pacheco-Moises FP, Bitzer-Quintero OK, et al. Immunology and oxidative stress in multiple sclerosis: clinical and basic approach. Clin Dev Immunol. 2013;2013:708659.
Inoue M, Shinohara ML. NLRP3 inflammasome and MS/EAE. Autoimmune Dis. 2013; 859145.
Heid ME, Keyel PA, Kamga C, et al. Mitochondrial reactive oxygen species induces NLRP3-dependent lysosomal damage and inflammasome activation. J Immunol. 2013;191:5230-5238.
Chelombitko MA, Fedorov AV, Ilyinskaya OP, Zinovkin RA, Chernyak BV. Role of reactive oxygen species in mast cell degranulation. Biochemistry (Mosc). 2016;81:1564-1577.
Hochman DJ, Collaco CR, Brooks EG. Acrolein induction of oxidative stress and degranulation in mast cells. Environ Toxicol. 2014;29:908-915.
Ohta Y, Yashiro K, Ohashi K, Horikoshi Y, Kusumoto C, Matsura T. Compound 48/80, a mast cell degranulator, causes oxidative damage by enhancing vitamin C synthesis via reduced glutathione depletion and lipid peroxidation through neutrophil infiltration in rat livers. J Clin Biochem Nutr. 2017;60:187-198.
Russi AE, Walker-Caulfield ME, Guo Y, Lucchinetti CF, Brown MA. Meningeal mast cell-T cell crosstalk regulates T cell encephalitogenicity. J Autoimmun. 2016;73:100-110.
Russi AE, Walker-Caulfield ME, Brown MA. Mast cell inflammasome activity in the meninges regulates EAE disease severity. Clin Immunol. 2018;189:14-22.
Macrez R, Vivien D, Docagne F. Role of endothelial NMDA receptors in a mouse model of multiple sclerosis. Med Sci (Paris). 2016;32:1068-1071.
Kamphuis WW, Derada Troletti C, Reijerkerk A, Romero IA, de Vries HE. The blood-brain barrier in multiple sclerosis: microRNAs as key regulators. CNS Neurol Disord Drug Targets. 2015;14:157-167.
Ramirez SH, Andrews AM, Paul D, Pachter JS. Extracellular vesicles: mediators and biomarkers of pathology along CNS barriers. Fluids Barriers CNS. 2018;15:19.
Kuwahara H, Nishina K, Yokota T. Blood–brain barrier: A novel therapeutic target in multiple sclerosis. Clin Exp Neuroim 2015;6:129-138.
Macrez R, Stys PK, Vivien D, Lipton SA, Docagne F. Mechanisms of glutamate toxicity in multiple sclerosis: biomarker and therapeutic opportunities. Lancet Neurol. 2016;15:1089-1102.
Ribatti, D. The crucial role of mast cells in blood-brain barrier alterations. Exp Cell Res. 2015;338:119-125.
Méndez-Enríquez E, Hallgren J. Mast cells and their progenitors in allergic asthma. Front Immunol. 2019;10:821.
Brown MA, Weinberg RB. Mast cells and innate lymphoid cells: underappreciated players in CNS autoimmune demyelinating disease. Front Immunol. 2018;9:514.
Presta I, Vismara M, Novellino F, et al. Innate immunity cells and the neurovascular unit. Int J Mol Sci. 2018;19.
Ocak U, Ocak PE, Wang A, et al. Targeting mast cell as a neuroprotective strategy. Brain Inj. 2019;33:723-733.
Tran H, Mittal A, Sagi V, et al. Mast cells induce blood brain barrier damage in SCD by causing endoplasmic reticulum stress in the endothelium. Front Cell Neurosci. 2019;13:56.
Mattila OS, Strbian D, Saksi J, et al. Cerebral mast cells mediate blood-brain barrier disruption in acute experimental ischemic stroke through perivascular gelatinase activation. Stroke 2011;42:3600-3605.
Woodberry T, Bouffler SE, Wilson AS, Buckland RL, Brüstle A. The emerging role of neutrophil granulocytes in multiple sclerosis. J Clin Med 2018; 7.
Nyekel N, Pacreau E, Benadda S, et al. Mast cell degranulation exacerbates skin rejection by enhancing neutrophil recruitment. Front Immunol. 2018; 20:2690.
Shin H, Kumamoto Y, Gopinath S, Iwasaki A. CD301b+ dendritic cells stimulate tissue-resident memory CD8+ T cells to protect against genital HSV-2. Nat Commun. 2016;7:13346.
Sonar SA, Lal G. Differentiation and transmigration of CD4 T cells in neuroinflammation and autoimmunity. Front Immunol. 2017;8:1695.
McGinley AM, Edwards SC, Raverdeau M, Mills KHG. Th17 cells, γδ T cells and their interplay in EAE and multiple sclerosis. J Autoimmun. 2018.
Castro-Borrero W, Graves D, Frohman TC, et al. Current and emerging therapies in multiple sclerosis: a systematic review. Ther Adv Neurol Disord. 2012;5:205-220.
Haghikia A, Hohlfeld R, Gold R, Fugger L. Therapies for multiple sclerosis: translational achievements and outstanding needs. Trends Mol Med. 2013;19:309-319.
La Mantia L, Tramacere I, Firwana B, Pacchetti I, Palumbo R, Filippini G. Fingolimod for relapsing-remitting multiple sclerosis. Cochrane Database Syst Rev. 2016;4:CD009371.
Hart DA. Curbing inflammation in multiple sclerosis and endometriosis: should mast cells be targeted? Int J Inflam. 2015;2015:452095.
Sokol KC, Amar NK, Starkey J, Grant JA. Ketotifen in the management of chronic urticaria: resurrection of an old drug. Ann Allergy Asthma Immunol. 2013;111:433-436.
Schwarzer G, Bassler D, Mitra A, Ducharme FM, Forster J. Ketotifen alone or as additional medication for long-term control of asthma and wheeze in children. Cochrane Database Syst Rev. 2004;1:CD001384.
Acknowledgements
The authors are grateful to researchers Camila Peres Buzalaf and Ana Paula Campanelli for their help concerning cytometry data analysis. The authors also thank Fatima Silveira, Luiz Carlos da Silva and Hélio Kushima for technical support. Funding was provided by São Paulo Research Foundation – FAPESP [grant number 2014/00239-6]. The authors declare no conflicts of interest.
Required Author Forms Disclosure forms provided by the authors are available with the online version of this article.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Rights and permissions
About this article

Cite this article
Pinke, K.H., Zorzella-Pezavento, S.F.G., de Campos Fraga-Silva, T.F. et al. Calming Down Mast Cells with Ketotifen: A Potential Strategy for Multiple Sclerosis Therapy?. Neurotherapeutics 17, 218–234 (2020). https://doi.org/10.1007/s13311-019-00775-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13311-019-00775-8