
Abstract
For feeding the world in 2100, the global agriculture, the entire food chain, as well as the behavior of all consumers must be change fundamentally. Essential resources needed to intensify agriculture and use barren land, such as phosphorus, water, and fossil fuels, are becoming increasingly scarce and expensive. An ecological form of agriculture that uses these resources more responsibly requires more land for the same yields. Therefore, new concepts for food and feed production have to be developed, in which nutrients are recycled beyond these areas. A possible starting point could be bioreactors, since these are enormously efficient and enable resource-efficient land use. Wastewater treatment as a means of nutrient recycling will be one of the most important tasks in the future. Hereby, not only the heterotrophic bioreactors currently used for this purpose but also autotrophic photobioreactors show great potential, especially if these two reactor types would be combined. Because of the ability to use inorganic nitrogen and phosphorus for their growth as well as the ability to produce a wide range of metabolites, microalgae offer an integrated approach. This review provides an overview of the potential of microalgae as components of a sustainable, circular agricultural system for feed and food production.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
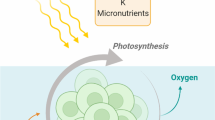
Today agriculture is based on the primary production of terrestrial plants. With the notable exception of most fish (etc.), almost every calorie a human consumes was produced in the leaf of a relatively large, soil-based vascular plant. Eukaryotic in water living organisms which operate photosynthesis are called algae. The number of species can only be estimated, but they are the major contributors of biodiversity and present in salt-, brackish- or freshwater (Graham et al. 2009; Metting 1996). On the basis of their size, a distinction is made between microalgae and macroalgae. Microalgae are single-celled organism, and they can be found individually, in chains, or groups with a size ranging from a few micrometers to a few hundred micrometers. Responsible for roughly 70% of the oxygen in the atmosphere, these single-celled algae and cyanobacteria (together referred to as microalgae) dominate in aquatic ecosystems. These organisms have long been viewed by scientists as having great potential for the agricultural system, especially with regard to biofuel but also for food, feed, and fertilizer production. The main reasons for this optimism are very high photosynthetic efficiency of microalgae and the possibility of using waste water for their cultivation, as well as flue gases, which can be fed into photobioreactors to increase productivity and mitigate CO2. However, the “hype” (Posten 2009; Walker et al. 2005) around the potential of microalgae cultivation has not yet been accompanied by large-scale success of photobioreactors.
The individual algae classes (divisions), Fig. 1, are distinguished primarily by the composition of their photosynthetic pigments and products. The microalgae diatoms (Bacillariophyceae) can be found in the oceans, freshwater, brackish water, and soils of the world. The green algae (Chlorophyceae) are abundant, especially in freshwater and have the same photosynthetic pigments (chlorophyll a and b), the same set of carotenoids (alpha, beta, and gamma carotene; lutein; zeaxanthin; violaxanthin; etc.), the same reserve substance (starch), and the same framework substance of the cell wall (cellulose) as green plants. Euglenophyta (Euglenophyceae) are predominating in freshwater, especially in eutrophic waters. Their frequent occurrence can cause a water bloom. The golden algae (Chrysophyceae) are a large group of algae, found mostly in freshwater. The dinoflagellates (Pyrrophyta) belong to the class of Dinophyceae with more than 1000 species, many of which live parasitic. Dinoflagellates occur in salt- and freshwater. In the sea, they are the second most important group of phytoplankton after diatoms. In warm waters, biodiversity is high with low numbers of individuals. In cold climates, few species with high numbers of individuals predominate. At regular intervals, mass developments of certain species occur in which the water turns red or orange (red tide) because of the large amount of carotenoids formed (Algen 1996–2004). The red algae (Rhodophyta, Rhodophyceae) are a division of algae, which are colored red by the phycobilin that is involved in photosynthesis. Beside Glaucophyta and Chloroplastida, the red algae form one of the three groups of Archaeplastida. Red algae occur in the majority in the littoral zone of the sea and some species also in freshwater and in moist soil.

The brown algae (Phaeophyceae) are a large group of multicellular algae mostly living in marine environments, and they are important for food and as habitat. The blue green algae (Cyanophyceae; Cyanobacteria, prokaryotes) are an excellent source of biologically active natural products including vitamin, protein, fine chemicals, and renewable fuel. They are largely unexplored and offer a great opportunity to discover new compounds, among others, biologically active compounds like antibacterial, antiviral, antifungal, algaecide, therapeutic agents, and cytotoxic activities (El Abed et al. 2008; van den Hoek and Jahns 1995).
Microalgae are a large, unexplored group of organisms, and only a few of the 30,000 known species are currently of commercial significance (Table 1). Even though they can be used to produce a wide range of metabolites like proteins, lipids, carbohydrates, carotenoids, and vitamins for health, food and feed additives, cosmetics, and for energy production. Microalgae can enhance the nutritional value of food and feed and play a crucial role in aquaculture (polyunsaturated fatty acids, PUFA). Three key points of microalgae can be converted into technical and commercial advantages:
-
Genetically very diverse with a wide range of physiological and biochemical characteristics
-
Cost-effectively incorporate the stable isotopes 13C, 15N, and 2H into their biomass
-
A large, unexplored group of organisms which offer an untapped source for products (Chew et al. 2017; Priyadarshani and Rath 2012; Vanthoor-Koopmans et al. 2013)
Nevertheless, the positive aspects of using microalgae also have downsides or rather challenges. High yields can only be achieved by high energy expenditure. Light energy is an important factor, therefore, either artificial lighting costs have to be borne or the strong dependence of sunlight has to be accepted. Currently, the use of algae as an alternative to fuels is not competitive (Hannon et al. 2010; Lehr and Posten 2009; Razzak et al. 2013; Richardson et al. 2012).
Technological improvements and economic changes, as well as a gradual cost reduction when photobioreactors become more established, could lead to more large-scale operations in the coming years. However, it is not clear yet how microalgae cultivation can best be integrated into the agricultural system as a whole. Here we analyze the potential of microalgae cultivation with regard to photosynthetic and areal efficiency, as well as resource use and nutrient circulation.
Photosynthetic efficiency
Photosynthetic efficiency (PE), sometimes also referred to as photon conversion efficiency, is the conversion rate from solar energy (photons) to chemical energy (biomass), usually given as a percentage in relation to the light that hits a certain surface area. The basis of photosynthesis is the quantum-mechanical process of light absorption, which takes place at the thylakoid membranes in chloroplasts. In this process, energy is transferred from photons to pigments, which are part of large protein-pigment complexes. These either form antenna, meant exclusively for light harvesting, or reaction centers, which also absorb light but are most important for passing on the energy, to power the chemical reactions which ultimately cumulate in the production of glucose from CO2 and H2O. These pigments can only absorb light in a certain bandwidth. Most higher plants use light in the range from 400 to 700 nm, which means that 51.3% of solar energy is unavailable to them (Zhu et al. 2008). Chlorophylls are the most important pigments for photosynthesis. Different types of chlorophylls have different absorption characteristics, but in general, they absorb light in the red and blue range, leaving a “green gap” between roughly 500 and 600 nm. Microalgae and cyanobacteria have different types of chlorophyll, which enable them to absorb light above 700 nm and below 400 nm (Zhu et al. 2010). To fill the green gap, different pigments, such as the bacterial phycobilins, are used (Zhu et al. 2010). The use of a wider range of pigments is one of the reasons why microalgae usually have higher PE than vascular plants. However, too much light can be harmful and lead to photosynthetic stress and the creation of toxic reactive oxygen species (ROS). To react to this, plants and microalgae can change the interactions between pigments through conformational changes of the proteins, or they can use pigments such as carotenoids, xanthophyll, and zeaxanthin as protection. These pigments emit the energy from light absorption as heat rather than passing it on to the reaction centers (Zhu et al. 2010).
Photosynthesis also depends on a light-independent chain of reactions: the Calvin cycle. Here, one of the main factors affecting PE is the functioning of the enzyme RuBisCO. This enzyme is responsible for fixating CO2 but also has an affinity to O2. When O2 concentrations are high, usually due to oxygen production in the light-dependent reaction of photosynthesis, electrons will be transferred to O2 more frequently, which leads to energy loss. Many higher plants solve this problem by separating the light-dependent and independent reactions in space, by letting them take place in different compartments (C4-plants), or by separating them in time, letting the light-independent reaction take place at night (CAM-plants). Microalgae cannot use these strategies and are thus dependent on protection from light stress through the use of protective pigments and antioxidants. However, while higher plants will regularly experience high oxygen concentrations in their leaves when closing the stomata, the level of photorespiration in microalgae is mostly dependent on the oxygen concentration in the surrounding substrate. While this might mean that microalgae are experiencing less photorespiration than higher plants in most natural environments, in photobioreactors it represents one of the central problems and challenges, since it means that the cultivation process is only effective when oxygen concentrations are sufficiently low and CO2 concentrations are sufficiently high. In most reactor types, this requires constant gas exchange. An advantage of this circumstance is, however, that the productivity (and thus PE) can be increased through feeding excess CO2 from industrial or agricultural activities (e.g., biogas plants) into the photobioreactors. This has the added benefit of CO2 mitigation.
PE is not only affected by molecular differences, such as described above, but also by morphological traits. Microalgae, for example, have a lower surface to volume ratio, which is beneficial. Also, microalgae do not have to grow and support non-photosynthetic tissues. While a tree has to form roots, stems, fruits, etc. and keep up nutrient and water flow between these different body parts to complete its life cycle, microalgae will simply duplicate once their single cell has reached sufficient size and accumulated enough energy. It is important to note for microalgae cultivation that electric energy is needed to fulfill many of the functions that a higher plant takes care of itself: gas exchange, nutrient and water flow, as well as temperature control (Sayre 2010). To determine the PE of a photobioreactor, it is necessary to take these energy inputs into account. Sometimes it is not clear whether this was done when researchers give PE values for photobioreactors.
Estimations of the PE of different organisms vary widely, but the general consensus is that microalgae have much higher PE than higher plants. It is however difficult to pinpoint the magnitude of this difference from literature. Sometimes PE is given as a theoretical maximum, sometimes as a “realistic estimate” and very occasionally as a value that has actually been measured. Another difficulty is that some estimates of PE refer to the whole spectrum of sunlight, while others refer only to the spectrum of light available to plants.
The theoretical maximum of PE is generally higher for C4 plants than for C3 plants, with 6% compared with 4.6% (Zhu et al. 20,010). In the “real world,” however, PE of higher plants is typically below 1% in temperate and tropical climates (Blankenship et al. 2011). Microalgae are said to have theoretical maximum PE of more than 20% (Janssen et al. 2003), based on photosynthetically available light, which would amount to about 10% PE and thus roughly twice as much a higher plants. In reality, the PE of photobioreactors is also way below the theoretical maximum. A PE of above 5% can however be seen as realistic according to most sources (Schenk et al. 2008; Janssen et al. 2003; Wijffels and Barbosa 2010). All in all, the energy yield of photobioreactors is lower than that of photovoltaic solar panels (Blankenship et al. 2011) but very high compared with crops. Solar panels take advantage of a larger spectrum of light, by stacking different light-absorbing layers above each other (Blankenship et al. 2011). It would be possible to do the same with photobioreactors, for example, by positioning two flat-panel bioreactors with two different species that make use of different wavelengths of light, in front of each other. Researchers are also looking at ways of improving energy yield of photosynthesis by genetically altering organisms and for example making RuBisCO less prone to photorespiration or modifying the pigment-protein complexes (Zhu et al. 2010).
Algae for nutrient circulation
Microalgae mainly consist of proteins, carbohydrates, and lipids with varying composition depending on the microalgae species. The carbohydrate fraction (starch, sugars, glucose, and other polysaccharides) is found to be up to 64% in microalgae biomass, the protein fraction up to 71%, and the lipid fraction up to 22% of cell dry weight (Becker 2007; Razzak et al. 2013). The content is highly specific to species and depending on growth conditions (Weyer et al. 2009). Under stress conditions like low nitrogen content or in the presence of supplemental reductants like sugar or glycerol, some species, e.g., Nannochloropsis sp. F&M-M24, accumulate energy-dense storage compounds such as lipids, with a content up to 60% per gram of dry weight (Rodolfi et al. 2009; Sayre 2010). Of course, to build up biomass, microalgae need a carbon source, water, temperature control, and light but also minerals like nitrogen, phosphorus, and other essential nutrients like sulfur, iron, and magnesium (Graham et al. 2009). Therefore algae are well suited to grow on wastewater that naturally includes high amounts of nutrients (Arashiro et al. 2019; Razzak et al. 2013).
Carbon source
Microalgae are capable of using carbon in the form of carbon dioxide from the atmosphere as well as emission from industrial power plants; inorganic carbon like NaHCO3 and Na2CO3; and organic carbon in form of sodium acetate, glucose, and glycerol. Since the beginning of the industrialization, the well-balanced ecosystem, including carbon capture from photosynthesis, carbon deposition in soil and oceans, and carbon release from biological and geological sources, is out of balance. The atmospheric CO2 concentration increased from 295 to 380 ppm over the last century, which is one of the main driving factors of global warming and climate change (Sayre 2010). According to Razzak et al. (2013), producing 100 t of algal biomass fixes about 183 t of CO2. The nongaseous form of CO2, which occurs in water at a ratio over 50% at pH values between 6.4 and 10.3, is bicarbonate which can be transported and concentrated in algae. Inside the cell, the bicarbonate is reconverted to CO2 and can be fixed to RuBisCO. After several reactions these molecules are substrates for starch and oil production (Huertas et al. 2001; Sayre 2010). According to literature, flue gas can be used as a carbon source for the production of microalgae (Demirbas 2011; Doucha et al. 2005; Holdmann and Schmid-Staiger 2016; Kadam 2002). Doucha et al. provided a combined biotechnological process scheme where agricultural wastes are anaerobically digested, the produced biogas is combusted in a boiler, and, following this, the flue gases are decarbonized by microalgae (Doucha et al. 2005). In addition to autotrophic microalgae, which mainly use CO2 as a carbon source, there are several mixotrophic and heterotrophic species which can access carbon from a range of different substrates. In the cultivation of these species, light energy is not an absolutely limiting factor (Kong et al. 2013). According to Cheng et al., the algae Chlorella protothecoides achieved an oil content of 53% by cell dry weight during a heterotrophic cultivation in a media containing sugar cane juice as an alternative carbon source (Cheng et al. 2009). This strain is also able to use glycerol, crude glycerol, and a glucose/glycerol mixture as carbon sources (Kamjunke et al. 2008; O'Grady and Morgan 2011).
Nitrogen and phosphorus
Nitrogen and phosphorus are essential to all organisms. Therefore, not only the environmental burdens resulting from excessive use should be considered but also the overuse of a finite resource. Nitrogen is the essential element of proteins and DNA and therefore also the building block of all enzymes that control plant, animal, and human metabolism. Nitrogen and hydrogen can be taken from the ambient air to synthesize ammonia (Haber-Bosch process). Among other things, ammonia is used to produce urea, ammonium nitrate, ammonium sulfate, and ammonium phosphates, which are used as fertilizers and contribute to the nutrition of a large part of the world’s population. Also compounds of phosphorus are essential for all living organisms and involved in the structure and function of organisms in key areas, such as DNA and the cellular energy supply (ADP/ATP). Phosphorus is naturally present in minerals, most commonly apatite. These minerals are mined in places with high phosphate content and, after appropriate treatment, used as mineral fertilizer, thus entering the phosphate cycle. Phosphorus, with increasing annual degradation, is about to reach the peak, where production reaches its maximum. It is estimated that the world’s reserves will cover the need for about 100 years (BMEL 2011), and other sources calculate shorter periods of time out (White and Cordell 2008).
Through traditional technologies for wastewater treatment, nitrogen, phosphorus, carbon, and other nutrients are not completely utilized and recycled (Abdel-Raouf et al. 2012; Han et al. 2019). In order to guarantee an environmentally friendly supply of these essential nutrients, continued work on economic recovery and recycling of these resources is necessary.
Microalgae offer the opportunity for the treatment of wastewater due to their ability to use inorganic nitrogen and phosphorus to build up biomass (Abdel-Raouf et al. 2012; Zhao et al. 2019). Sources of nitrogen are inorganic nitrogen sources like nitrate, nitrite, ammonia, and urea (Li et al. 2008; Xiong et al. 2008) and for some algae species also organic nitrogen sources like glycine and yeast extract (Xiong et al. 2008). Algae take up nitrate and ammonium ions directly from the surrounding water, with ammonium being preferred to build up cellular nitrogen compounds. Using the enzyme nitrate reductase, algae are able to convert nitrate to ammonium (Graham et al. 2009).
Phosphorus in the form of orthophosphate (PO43−) is the preferred uptake form of algae.
After aerobic or anaerobic biological degradation of wastewater, the content of inorganic components like nitrate, ammonium, and phosphate ions is sufficient for eutrophication of water environment. Microalgae show high capacity to take up these components and use inorganic nitrogen and phosphorus for their growth (Abdel-Raouf et al. 2012; Zhao et al. 2019).
Other nutrients
Elements like sulfur, iron, magnesium, and others are required as trace elements. Indeed, iron acts as a cofactor for several enzymes like ferredoxin, catalase, cytrocomes, glutamate synthetase, nitrogenase, nitrate, and nitrite reductase (Graham et al. 2009). Sulfur is usually taken up and assimilated as sulfate and is essential for the incorporation into a variety of sulfur-containing compounds critical for protein, lipid, and polysaccharide synthesis, as well as signaling molecules (Giordano et al. 2008; Shibagaki and Grossman 2008). Magnesium also serves as a cofactor of several enzymes and is required for the biosynthesis of chlorophyll (Brzezowski et al. 2015; Graham et al. 2009). Beside the requirement of trace elements, algae are also known for their ability to absorb and accumulate heavy metals and compounds like organochlorides intracellularly. Additionally the secretion of extracellular esterase, which degrades Deltamethrin (insecticide), and the ability to degrade a range of hydrocarbons (found in oily wastes) are known for some species of microalgae (Abdel-Raouf et al. 2012; Arunakumara and Xuecheng 2008; Worms et al. 2010).
Potential of microalgae
Due to the biodiversity, the composition of the biomass depending on the nutrient availability, use of different nutrients, and the production of different substances, microalgae show great potential for a wide variety of applications (Metting 1996).
Microalgae-assisted aquaculture is known for live feed for larvae, fish species, and zooplankton (Brown et al. 1997), as food additive to supply basic nutrients, enhance the color of salmonids or for other biological activities (Muller-Feuga 2000), stabilization and improvement of quality of culture medium (Chuntapa et al. 2003), stimulation of immune systems (Spolaore et al. 2006) as well as probiotic effects (Irianto and Austin 2002; Han et al. 2019; Roy and Pal 2014). The most used species are Spirulina, Chlorella, Scenedesmus, Dunaliella, Tetraselmis, Isochrysis, Pavlova, Skeletonema, Chaetoceros, Phaeodactylum, Nitzschia, and Thalassiosira (Beal et al. 2018; Brown et al. 1997; Han et al. 2019).
Next to the potential of microalgae in aquaculture, the potential is also given in the area of implementing an eco-friendly system using microalgae in form of, e.g., wastewater treatment and nitrogen removal (Abdel-Raouf et al. 2012; Arashiro et al. 2019; Di Termini et al. 2011; Ledda et al. 2015b; Razzak et al. 2013; Wang et al. 2019; Yu et al. 2019), treatment of heavily polluted meat processing wastewater as primary or secondary treatment option (Hu et al. 2019), recycling of animal wastewater and manure (Kim et al. 2007; Ledda et al. 2015a; Pizarro et al. 2002), and flue gas (Doucha et al. 2005; Nagase et al. 1997; Yoo et al. 2010).
The potential of microalgae in generating biofuels has been of major interest over the last decades (Chew et al. 2017; Hussian 2018; Vanthoor-Koopmans et al. 2013). The usage of algae components like carbohydrates, mainly consisting of glucose, starch, cellulose as well as polysaccharides, show potential too (Chew et al. 2017). Microalgae are able to generate a wide variety of photosynthetic storage products including α-(1-4)-linked glucans (starches), β-(1-3)-glucans, fructans, low molecular weight carbohydrates, and fats and oils. Different types of starches are produced by different divisions of algae, e.g., red algae are known to synthesize floridean starch (amylopectin subunits), whereas blue-green algae synthesize myxophycean starch (amylopectin or glycogen-like subunits). Some species of green algae synthesize a cross-linked amylose-amylopectin starch and fructosans (inulin-like fructose oligosaccharides), which are comparable with starch in land plants. Cryptophytes and dinoflagellates generate α-(1-4)-linked glucans. Chrysophytes store oils or chrysophycean starch, a water-soluble β-(1-3) glucopyranoside (Metting 1996). According to literature, starch contents up to 60% of dry weight are achieved with Chlorella vulgaris (Branyikova et al. 2011; Dragone et al. 2011; Pruvost et al. 2011) and Tetraselmis subcordiformis (Yao et al. 2012) and up to 50% of dry weight with Phaeodactylum tricornutum (Jakob et al. 2007).
Hence, investigations (beside the usage for biofuel or as a source of bioactive compounds and pharmaceuticals, health foods, or cosmetic additives) could be of high interest maybe to yield starch (produced without the need of arable land) or fermentable monosaccharides to receive an ecological and sustainable bioresource (Chew et al. 2017; Reisky et al. 2019).
Conclusion
The versatility of algae and therefore their potential for nutrient circulation, biomitigation of carbons must be of increasing interest in the future due to existing problems including global warming, discharge of wastes, supply of nutrients (nitrogen, phosphorus), or toxic chemicals.
The possibility given through these highly diverse microalgae, capable to grow photoautotroph, heterotroph, and mixotroph on barren land, and the ability of CO2 fixation must be investigated more intensively. An ideal algae strain for tropical climates should be able to produce biomass at high solar radiance and high oxygen levels.
The challenge will be to have this ideal strain in the right place and in a perfect, yet simple combination of wastewater treatment, CO2 biomitigation and nutrient recycling to create new concepts for food and feed production in bioreactors. The implementation must be technically simple, long-term stable, and easy to handle and to care for. In order to have enough purified water, food, and feed to supply the world in 2100, the integrated usage of microalgae could be an opportunity for the global agriculture.
References
Abdel-Raouf N, Al-Homaidan AA, Ibraheem IB (2012) Microalgae and wastewater treatment. Saudi J Biol Sci 19(3):257–275. https://doi.org/10.1016/j.sjbs.2012.04.005
Algen (1996–2004) Algen. http://www1.biologie.uni-hamburg.de/b-online/e44/44.htm. Accessed 26 September 2019
Arashiro LT, Ferrer I, Rousseau DPL, Van Hulle SWH, Garfi M (2019) The effect of primary treatment of wastewater in high rate algal pond systems: biomass and bioenergy recovery. Bioresour Technol 280:27–36
Arunakumara KKIU, Xuecheng Z (2008) Heavy metal bioaccumulation and toxicity with special reference to microalgae. J Ocean U China 7(1):60–64. https://doi.org/10.1007/s11802-008-0060-y
Beal CM, Gerber LN, Thongrod S, Phromkunthong W, Kiron V, Granados J, Archibald I, Greene CH, Huntley ME (2018) Marine microalgae commercial production improves sustainability of global fisheries and aquaculture. Sci Rep-Uk 8(1):15064. https://doi.org/10.1038/s41598-018-33504-w
Becker EW (2007) Micro-algae as a source of protein. Biotechnol Adv 25(2):207–210
Blankenship RE, Tiede DM, Barber J, Brudvig GW, Fleming G, Ghirardi M et al. (2011) Comparing photosynthetic and photovoltaic efficiencies and recognizing the potential for improvement. In science (New York, N.Y.) 332 (6031), pp 805–809. doi: https://doi.org/10.1126/science.1200165
BMEL-Wissenschaftlicher Beirat für Düngungsfragen des BMEL (2011) Nachhaltiger Umgang mit der begrenzten Ressource Phosphor durch Recycling und Erhöhung der Phosphoreffizienz der Düngung. . https://www.bmel.de/SharedDocs/Downloads/Ministerium/Beiraete/Duengungsfragen/Phosphor.pdf?__blob=publicationFile. Accessed 20 Sepember 2019
Branyikova I, Marsalkova B, Doucha J, Branyik T, Bisova K, Zachleder V, Vitova M (2011) Microalgae--novel highly efficient starch producers. Biotechnol Bioeng 108(4):766–776. https://doi.org/10.1002/bit.23016
Brown MR, Jeffrey SW, Volkman JK, Dunstan GA (1997) Nutritional properties of microalgae for mariculture Aquac 151:315-331. doi.org/https://doi.org/10.1016/S0044-8486(96)01501-3
Brzezowski P, Richter AS, Grimm B (2015) Regulation and function of tetrapyrrole biosynthesis in plants and algae. Biochim Biophys Acta 1847(9):968–985. https://doi.org/10.1016/j.bbabio.2015.05.007
Cheng Y, Lu Y, Gao C, Wu Q (2009) Alga-based biodiesel production and optimization using sugarcane as the feedstock. Energy Fuel 23:4166–4173. https://doi.org/10.1021/ef9003818
Chew KW, Yap JY, Show PL, Suan NH, Juan JC, Ling TC, Lee DJ, Chang JS (2017) Microalgae biorefinery: high value products perspectives. Bioresour Technol 229:53–62. https://doi.org/10.1016/j.biortech.2017.01.006
Chuntapa B, Powtongsook S, Menasveta P (2003) Water quality control using Spirulina platensis in shrimp culture tanks Aquac 220(1-4):355-366. doi.org/https://doi.org/10.1016/S0044-8486(02)00428-3
Demirbas A (2011) Biodiesel from oilgae, biofixation of carbon dioxide by microalgae: a solution to pollution problems. Appl Energy 88(10):3541-3547. doi.org/https://doi.org/10.1016/j.apenergy.2010.12.050
Di Termini I, Prassone A, Cattaneo C, Rovatti M (2011) On the nitrogen and phosphorus removal in algal photobioreactors. Ecol Eng 37(6):976–980. https://doi.org/10.1016/j.ecoleng.2011.01.006
Doucha J, Straka F, Lívanský K (2005) Utilization of flue gas for cultivation of microalgae (Chlorella sp.) in an outdoor open thin-layer photobioreactor. J Appl Phycol 17(5):403–412. https://doi.org/10.1007/s10811-005-8701-7
Dragone G, Fernandes BD, Abreu AP, Vicente AA, Teixeira JA (2011) Nutrient limitation as a strategy for increasing starch accumulation in microalgae. Appl Energy 88(10):3331-3335. doi.org/https://doi.org/10.1016/j.apenergy.2011.03.012
El Abed MM, Marzouk B, Medhioub MN, Helal AN, Medhioub A (2008) Microalgae: a potential source of polyunsaturated fatty acids. Nutr Health 19(3):221–226. https://doi.org/10.1177/026010600801900309
Enamala MK, Enamala S, Chavali M, Donepudi J, Yadavalli R, Kolapalli B, Aradhyula TV, Velpuri J, Kuppam C (2018) Production of biofuels from microalgae - a review on cultivation, harvesting, lipid extraction, and numerous applications of microalgae. Renew Sust Energ Rev 94:49–68. https://doi.org/10.1016/j.rser.2018.05.012
Fondriest Environmental I (2014) Algae, phytoplankton and chlorophyll. https://wwwfondriestcom/environmental-measurements/parameters/water-quality/algae-phytoplankton-chlorophyll/ Accessed 16 September 2019
Giordano M., Norici A., Ratti S., J.A. R (2008) Role of sulfur for algae: acquisition, metabolism, ecology and evolution. In: Hell R., Dahl C., Knaff D., Leustek T. (eds) sulfur metabolism in phototrophic organisms. Advances in photosynthesis and respiration. Vol 27, chapter 20: 397–415. Springer, Dordrecht. doi https://doi.org/10.1007/978-1-4020-6863-8
Graham JE, Wilcox LW, Graham LE (2009) Algae, 2 edn. Pearson Education, University of Wisconsin, Madison
Han P, Lu Q, Fan L, Zhou W (2019) A review on the use of microalgae for sustainable aquaculture. Appl Sci 9(11):2377. https://doi.org/10.3390/app9112377
Hannon M, Gimpel J, Tran M, Rasala B, Mayfield S (2010) Biofuels from algae: challenges and potential. Biofuels 1(5):763–784
Holdmann C, Schmid-Staiger U (2016) Bioraffinerie auf Basis kohlenhydratreicher Algenbiomasse, Nutzung von Stärke und Protein: Teilvorhaben 1: Koordination sowie Auswahl und Prozessentwicklung für stärkereiche Mikroalgen: Schlussbericht zum Vorhaben: Teilbericht A und Teilbericht B gemeinsamer Sachbericht: Laufzeit: 12.2012. bis 11.2015. doi https://doi.org/10.2314/GBV:870178172
Hu X, Meneses YE, Stratton J, Wang B (2019) Acclimation of consortium of micro-algae help removal of organic pollutants from meat processing wastewater. J Clean Prod 214:95–102. https://doi.org/10.1016/j.jclepro.2018.12.255
Huertas IE, Colman B, Espie GS, Lubian LM (2001) Active transport of CO2 by three species of marine microalgae. J Phycol 36:314–320. https://doi.org/10.1046/j.1529-8817.2000.99142.x
Hussian AEM (2018) The role of microalgae in renewable energy production: challenges and opportunities. In: Türkoğlu M, Önal U, Ismen A (eds) Marine ecology - biotic and abiotic interactions. IntechOpen. https://doi.org/10.5772/intechopen.73573
Irianto A, Austin B (2002) Probiotics in aquaculture J Fish Dis 25(11):633-642. doi.org/https://doi.org/10.1046/j.1365-2761.2002.00422.x
Jakob T, Wagner H, Stehfest K, Wilhelm C (2007) A complete energy balance from photons to new biomass reveals a light- and nutrient-dependent variability in the metabolic costs of carbon assimilation. J Exp Bot 58(8):2101–2112. https://doi.org/10.1093/jxb/erm084
Janssen M, Tramper J, Mur LR, Wijffels RH (2003) Enclosed outdoor photobioreactors: light regime, photosynthetic efficiency, scale-up, and future prospects. Biotechnol Bioeng 81(2):193–210. https://doi.org/10.1002/bit.10468
Kadam KL (2002) Environmental implications of power generation via coal-microalgae cofiring. Energ. Environ Sci 27:905–922. https://doi.org/10.1016/S0360-5442(02)00025-7
Kamjunke N, Kohler B, Wannicke N, Tittel J (2008) Algae as competitors for glucose with heterotrophic bacteria(1). J Phycol 44(3):616–623. https://doi.org/10.1111/j.1529-8817.2008.00520.x
Kim MK, Park JW, Park CS, Kim SJ, Jeune KH, Chang MU, Acreman J (2007) Enhanced production of Scenedesmus spp. (green microalgae) using a new medium containing fermented swine wastewater. Bioresour Technol 98(11):2220–2228. https://doi.org/10.1016/j.biortech.2006.08.031
Kong WB, Yang H, Cao YT, Song H, Hua SF, Xia CG (2013) Effect of glycerol and glucose on the enhancement of biomass, lipid and soluble carbohydrate production by Chlorella vulgaris in mixotrophic culture. Food Technol Biotechnol 51(1):62–69
Ledda C, Idà A, Allemand D, Mariani P, Adani F (2015a) Production of wild Chlorella sp. cultivated in digested and membrane-pretreated swine manure derived from a full-scale operation plant. Algal Res 12:68–73. https://doi.org/10.1016/j.algal.2015.08.010
Ledda C, Romero Villegas GI, Adani F, Acién Fernández FG, Molina Grima E (2015b) Utilization of centrate from wastewater treatment for the outdoor production of Nannochloropsis gaditana biomass at pilot-scale. Algal Res 12:17–25. https://doi.org/10.1016/j.algal.2015.08.002
Lehr F, Posten C (2009) Closed photo-bioreactors as tools for biofuel production. Curr Opin Biotechnol 20(3):280–285. https://doi.org/10.1016/j.copbio.2009.04.004
Li Y, Horsman M, Wang B, Wu N, Lan CQ (2008) Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Appl Microbiol Biotechnol 81(4):629–636. https://doi.org/10.1007/s00253-008-1681-1
Metting FB (1996) Biodiversity and application of microalgae. J Ind Microbiol Biotechnol 17:477–489. https://doi.org/10.1007/BF01574779
Muller-Feuga A (2000) The role of microalgae in aquaculture: situation and trends. J Appl Phycol 12(3/5):527–534. https://doi.org/10.1023/a:1008106304417
Nagase H, Yoshihara K, Eguchi K, Yokota Y, Matsui R, Hirata K, Miyamoto K (1997) Characteristics of biological NOx removal from flue gas in a Dunaliella tertiolecta culture system. J Ferment Bioeng 83(5):461–465
O'Grady J, Morgan JA (2011) Heterotrophic growth and lipid production of Chlorella protothecoides on glycerol. Bioprocess Biosyst Eng 34(1):121–125. https://doi.org/10.1007/s00449-010-0474-y
Pizarro C, Kebede-Westhead E, Mulbry W (2002) Nitrogen and phosphorus removal rates using small algal turfs grown with dairy manure. J Appl Phycol 14(6):469–473. https://doi.org/10.1023/a:1022338722952
Posten C (2009) Design principles of photo-bioreactors for cultivation of microalgae. Eng. Life Sci 9(3):165–177. https://doi.org/10.1002/elsc.200900003
Priyadarshani I, Rath B (2012) Commercial and industrial applications of micro algae – a review. J Algal Biomass Utln 3(4):89–100
Pruvost J, Van Vooren G, Le Gouic B, Couzinet-Mossion A, Legrand J (2011) Systematic investigation of biomass and lipid productivity by microalgae in photobioreactors for biodiesel application. Bioresour Technol 102(1):150–158. https://doi.org/10.1016/j.biortech.2010.06.153
Razzak SA, Hossain MM, Lucky RA, Bassi AS, de Lasa H (2013) Integrated CO2 capture, wastewater treatment and biofuel production by microalgae culturing-a review. Renew Sust Energ Rev 27:622–653. https://doi.org/10.1016/j.rser.2013.05.063
Reisky L, Préchoux A, Zühlke M-K, Bäumgen M, Robb CS, Gerlach N, Roret T, Stanetty C, Larocque R, Michel G, Song T, Markert S, Unfried F, Mihovilovic MD, Trautwein-Schult A, Becher D, Schweder T, Bornscheuer UT, Hehemann J-H (2019) A marine bacterial enzymatic cascade degrades the algal polysaccharide ulvan. Nat Chem Biol 15:803–812. https://doi.org/10.1038/s41589-019-0311-9
Richardson JW, Johnson MD, Outlaw JL (2012) Economic comparison of open pond raceways to photo bio-reactors for profitable production of algae for transportation fuels in the southwest. Algal Res 1(1):93–100. https://doi.org/10.1016/j.algal.2012.04.001
Rodolfi L, Chini Zittelli G, Bassi N, Padovani G, Biondi N, Bonini G, Tredici MR (2009) Microalgae for oil: strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol Bioeng 102(1):100–112. https://doi.org/10.1002/bit.22033
Roy SS, Pal R (2014) Microalgae in aquaculture: a review with special references to nutritional value and fish dietetics. P Zool Soc 68(1):1–8. https://doi.org/10.1007/s12595-013-0089-9
Sayre R (2010) Microalgae: the potential for carbon capture. Bioscience 60(9):722–727. https://doi.org/10.1525/bio.2010.60.9.9
Schenk PM, Thomas-Hall SR, Stephens E, Marx UC, Mussgnug JH, Posten C, Kruse O, Hankamer B (2008) Second generation biofuels: high-efficiency microalgae for biodiesel production. Bioenerg Res 1(1):20–43. https://doi.org/10.1007/s12155-008-9008-8
Shibagaki N, Grossman A (2008) The state of sulfur metabolism in algae: from ecology to genomics. In: Hell R, Dahl C, Knaff D, Leustek T (eds) Sulfur metabolism in phototrophic organisms. Advances in photosynthesis and respiration,. vol 27. Chapter 13: 231–267. Springer, Dordrecht. https://doi.org/10.1007/978-1-4020-6863-8_13
Spolaore P, Joannis-Cassan C, Duran E, Isambert A (2006) Commercial applications of microalgae. J Biosci Bioeng 101(2):87–96. https://doi.org/10.1263/jbb.101.87
van den Hoek CMD, Jahns HM (1995) Algae: an introduction to phycology. Cambridge University Press 32:203–205. https://doi.org/10.1017/S096702629621100X
Vanthoor-Koopmans M, Wijffels RH, Barbosa MJ, Eppink MH (2013) Biorefinery of microalgae for food and fuel. Bioresour Technol 135:142–149. https://doi.org/10.1016/j.biortech.2012.10.135
Walker TL, Purton S, Becker DK, Collet C (2005) Microalgae as bioreactors. Plant Cell Rep 24(11):629–641. https://doi.org/10.1007/s00299-005-0004-6
Wang L, Addy M, Lu Q, Cobb K, Chen P, Chen X, Liu Y, Wang H, Ruan R (2019) Cultivation of Chlorella vulgaris in sludge extracts: nutrient removal and algal utilization. Bioresour Technol 280:505–510. https://doi.org/10.1016/j.biortech.2019.02.017
Weyer KM, Bush DR, Darzins A, Willson BD (2009) Theoretical maximum algal oil production. Bioenerg Res 3(2):204–213. https://doi.org/10.1007/s12155-009-9046-x
White S, Cordell D (2008) Peak Phopshorus: the sequel to peak oil. http://phosphorusfuturesnet/the-phosphorus-challenge/peak-phosphorus-the-sequel-to-peak-oil/ Accessed 20 September 2019
Wijffels RH, Barbosa MJ (2010) An outlook on microalgal biofuels. Science (New York, NY) 329(5993):793–796. https://doi.org/10.1126/science.1189139
Worms IA, Traber J, Kistler D, Sigg L, Slaveykova VI (2010) Uptake of cd(II) and Pb(II) by microalgae in presence of colloidal organic matter from wastewater treatment plant effluents. Environ Pollut 158(2):369–374. https://doi.org/10.1016/j.envpol.2009.09.007
Xiong W, Li XF, Xiang JY, Wu QY (2008) High-density fermentation of microalga Chlorella protothecoides in bioreactor for microbio-diesel production. Appl Microbiol Biotechnol 78(1):29–36. https://doi.org/10.1007/s00253-007-1285-1
Yao CH, Ai JN, Cao XP, Xue S, Zhang W (2012) Enhancing starch production of a marine green microalga Tetraselmis subcordiformis through nutrient limitation. Bioresour Technol 118:438–444. https://doi.org/10.1016/j.biortech.2012.05.030
Yoo C, Jun S-Y, Lee J-Y, Ahn C-Y, Oh H-M (2010) Selection of microalgae for lipid production under high levels carbon dioxide. Bioresour Technol 101(1,Supplement):S71–S74. https://doi.org/10.1016/j.biortech.2009.03.030
Yu H, Kim J, Lee C (2019) Potential of mixed-culture microalgae enriched from aerobic and anaerobic sludges for nutrient removal and biomass production from anaerobic effluents. Bioresour Technol 280:325–336. https://doi.org/10.1016/j.biortech.2019.02.054
Zhao Z, Liu S, Yang X, Lei Z, Shimizu K, Zhang Z, Lee D-J, Adachi Y (2019) Stability and performance of algal-bacterial granular sludge in shaking photo-sequencing batch reactors with special focus on phosphorus accumulation. Bioresour Technol 280:497–501. https://doi.org/10.1016/j.biortech.2019.02.071
Zhu XG, Long SP, Ort DR (2008) What is the maximum efficiency with which photosynthesis can convert solar energy into biomass? Curr Opin Biotechnol 19(2):153–159. https://doi.org/10.1016/j.copbio.2008.02.004
Zhu XG, Long SP, Ort DR (2010) Improving photosynthetic efficiency for greater yield. Annu Rev Plant Biol 61:235–261. https://doi.org/10.1146/annurev-arplant-042809-112206
Availability of data and material
Not applicable.
Code availability
Not applicable.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethics approval
Not applicable.
Consent to participate
I consent.
Consent for publication
I consent.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised due to a retrospective Open Access order.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kuenz, A., Grimm, D. & Rahmann, G. Versatility of algae—exploring the potential of algae for nutrient circulation. Org. Agr. 11, 251–260 (2021). https://doi.org/10.1007/s13165-020-00308-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13165-020-00308-0