
Abstract
The Indo-Pacific is recognized as a hotspot for marine diversity. The taxonomy of certain taxa, such as Discomedusae jellyfish, has been neglected, despite its importance in the fishery industry. This study documents the first records of Discomedusae for the Java Sea using an integrative approach and provide notes about its distribution in the area. We used up to 53 morphological and meristic characters and amplified one mitochondrial marker (COI). The comparison and assessment of these data resulted in the recognition of seven species of Discomedusae, from which five has been recorded for the Indo-Pacific area. Two other species require a taxonomic revision to confirm the species assignation. The distribution of jellyfish in the coast of Java Sea might be correlated with the jellyfish life history and species-specific ranges of tolerance, and not solely determined by the environmental parameters. These findings provide the foundations for extending the taxonomic research in the area; the description of the biodiversity will increase the understanding of the population dynamics and its implications in the fisheries.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Scyphozoan jellyfish are one of the most common pelagic invertebrates which inhabit all the oceanic basins (Kramp 1961; Arai 1997). The class Scyphozoa includes the subclass Discomedusae (Order Rhizostomeae, shallow water, and Semaeostomeae, shallow and deep-water medusae) and Order Coronatae (shallow and deep-water medusae, Kramp 1961; Marques and Collins 2004; Bayha et al. 2010). Today, Discomedusae are increasing in frequency and abundance (Attrill et al. 2007; Condon et al. 2012; Brotz et al. 2012), rising the public and scientific interest due its detrimental ecological and economic consequences (Purcell et al. 2007; Richardson et al. 2009; Dong et al. 2010; Roux et al. 2013) and its growing demand and profits for the jelly-fishery industry (Kitamura and Omori 2010; Brotz 2016; Brotz et al. 2017). However, most of the studies do not address the genetic and taxonomic diversity (e.g., Condon et al. 2012), even for hotspot areas, which have been proven to account for a high biodiversity (e.g., Tropical Eastern Pacific, Gómez Daglio and Dawson (2017), in part Caribbean and Indo-Pacific, Abboud et al. 2018).
The Indo-Pacific Ocean has a well-documented marine diversity for invertebrates such as corals, mollusks, and crustaceans and fishes (Moretzsohn and McShane 2004; Barber and Bellwood 2005; Barber and Boyce 2006; Allen 2007; Hubert et al. 2012; Mihaljević et al. 2017). However, documentation of scyphomedusae diversity is limited to a few expeditions and taxonomic revisions made during nineteenth and early twentieth century (Haeckel 1880; Vanhöffen 1888; Maas 1907; Mayer 1910; Rao 1931; Stiasny 1940). The Indo-Pacific is a heavily fishing area, and the jelly-fishery is a common activity in the area (Brotz 2016). Currently, only two studies describe some fished Discomedusae using an integrative approach (morphology and molecular tools, Strait of Malacca (Rizman-Idid et al. 2016), and Indian Ocean, Central Java (Nishikawa et al. 2014)). Hence, the jelly-fishery is managed without a reliable taxonomic identification of the targeted species, and estimates about its landing, abundance, and distribution are questioned (Kitamura and Omori 2010; Brotz 2016; Rizman-Idid et al. 2016).
Indonesia shows in the top three countries with the highest landing (~ 29,469 tons) and exports of jellyfish (Omori and Nakano 2001; Brotz 2016). Yet, there are 35 species of Discomedusae reported for Indonesia (Table S1; Kramp 1961; De Souza and Dawson 2018; Jarms and Morandini, 2019). However, the species identification and distribution range are uncertain for 13 of the 35 recorded species (Table S1). Only five confirmed records exist for the area: Crambione mastigophora (Asrial et al. 2015), Crambionella helmbiru (Nishikawa et al. 2014), Lobonemoides robustus (Kitamura and Omori 2010), Rhopilema esculentum (Kitamura and Omori 2010), and Versuriga anadyomene (Ohtsuka et al. 2010). Except for C. helmbiru, the identification and records rely on morphological characterization of the species, that can be problematic, due to the presence of cryptic species complexes within Discomedusae (Dawson and Jacobs 2001; Holland et al. 2004; Holst and Laakmann 2014; Gómez Daglio and Dawson 2017). It is necessary to describe the Discomedusae diversity in such important hotspot and fishery area. Herein, we aim to describe the Discomedusae diversity on an unexplored area of Indonesia (Java Sea: Strait of Surabaya), employing an integrative approach (genetic and morphologic data). In addition, we include some notes about its records along the coastline.
Materials and methods
Sample collection
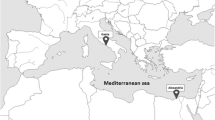
We carried out two major collection projects. The first one consisted of a survey the east coast of Surabaya at 22 stations located in two transects parallel to the shore (0.5 km and 1.5 km from the coastline, Fig. 1) between September and December 2010 every 2 weeks. At each station, we measured five physico-chemical parameters (temperature, salinity, pH, transparency, and flow velocity). Surface temperature was measured using a mercury thermometer with a precision of ± 1 °C. Salinity was measured using a hand refractometer (ATAGO ATC FG-217 with a precision of ± 1%). pH was measured using testing pH paper (Hydrion, range 0.0 to 13.0 pH). Transparency levels were measured using a Secchi disk following the protocol of English et al (1994). Flow velocity was estimated using buoys, which were held together with 2 m of rope and floated at each sampling point then timed with a chronometer.

Map of the sampling station in the Eastern Surabaya (Java Sea). We sampled two transects along the coast. Each transect consisted in 11 stations; the distance of the first transect is 0.5 km and the second one 1.5 km. Country codes as follows: AU, Australia; IN, Indonesia; MY, Malaysia; PH, Philippines; TH, Thailand. Map source: The Generic Mapping Tools (GMT)
Discomedusae were collected during mornings with Gill-nets (Ø = 1.5 cm) and Push-nets (Ø = 3 cm) at 2 m deep, with immersion time ranging from 10 to 15 min over the 5-h period. In addition, NORPAC nets (Ø = 150 μm) were laid horizontally on the surface at 2-min intervals. A piece of oral arm, tentacle, or umbrella margin were clipped and preserved in ethanol 90% for subsequent DNA analyses; the whole medusa was preserved in 10% formalin. To complete the taxonomic sampling, we carried out a different sampling; here we collected jellies from shrimp nets set by local fishermen in Kenjeran in the strait of Surabaya during the months of September through November 2010. All the samples were labeled and separated by morphotypes groups at the taxonomic family level (Pelagiidae, Catostylidae, Mastigiidae, Cyaneidae, Lychnorhizidae, and Rhizostomatidae sensu (Kramp 1961). In addition, on each site, we recorded the number of Discomedusae captured per station.
Morphological identification
Morphological characterization and identification were performed in the Ecology Department Laboratory of Biological Science ITS Surabaya (Indonesia). Each medusa was photographed and examined to record morphological and meristic characters that the literature suggests are helpful for Discomedusae identification (Mayer 1910; Light 1914; Stiasny 1920, 1921, 1922, 1932; Marques and Collins 2004; Morandini and Marques 2010; Gómez Daglio and Dawson 2017). For example, macro-morphological characters, coloration patterns; bell shape, distribution, position, number, and shape of oral arms; shape and number of terminal clubs; position, insertion, and number of tentacles; detailed characters such as filaments, scapula, and exumbrella papillae; and rhopalia insertion, shape and position, manubrium, position, number and shape of the oral pillars, shape and size of the oral disk, and shape and number of velar lappets, were recorded and photographed using a Stereo Microscope Olympus SZ51 (list of measured characters is available in Gómez Daglio and Dawson (2017)). Specimens revised were deposited in the Museum of Comparative Zoology, Harvard University (MCZ Cat. No.153057–153060; 153062–153066; 153068–153072).
DNA extraction, amplification, and sequencing
Total genomic DNA was extracted from a subset from the morphotypes’ tissue samples (12 individuals) using a modified CTAB phenol–chloroform protocol (Dawson and Jacobs 2001). We amplified the mitochondrial locus cytochrome c oxidase subunit I (COI). COI is widely use within the Discomedusae to identify species (Ortman et al. 2010; Gómez Daglio and Dawson 2017). We used the primer pairs LCOjf (GGTCAACAAATCATAAAGATATTGGAAC; Dawson 2005a) and HCO2198 (TAAACTTCAGGGTGACCAAAAAATCA; Folmer et al. 1994). We set up 25µL PCR reactions containing 0.5 µL DNA template, 0.1 mM each dNTP (Gene Amp mix with dTTP, Applied Biosystems Inc., USA), 2.5 µL of 10 × PCR buffer and 2.5 µL MgCl2, 0.63 µL each primer, and 0.0.5 units of Amplitaq (Applied Biosystems). Occasionally, some samples included 0.2 mM Bovine Serum Albumin to stabilize the enzymatic reaction during the PCR reactions. All PCR conditions followed the modification proposed by Gómez Daglio and Dawson (2017), which consist in three initial holds: 95 °C for 8 min, 51–57 °C for 2 min, 72 °C for 2 min, and then 33–35 cycles that included denaturation 94 °C for 45 s, annealing 47–52 °C for 50–70 s, elongation 72 °C for 1 min, and then a final elongation 72 °C for 10 min. Amplicons were sequenced by DNA Sequencing Facility of the University of California, Berkeley (CA, USA) or Laragen, Inc. (CA, USA).
Molecular identification
All the sequences were assembled into contigs, primers trimmed, and inspected manually to check base calls and open reading frames using Sequencher v. 5.2.4 (Gene Codes Corp. Ann Arbor). Sequences were deposited in GenBank (Accession number MN395673-MN395694). We aligned congeneric sequences using ClustalX v. 2.0 (Larkin et al. 2007). Each alignment was visualized using Jalview (Waterhouse et al. 2009); we estimated the sequences similarity and export the consensus sequence. Each consensus sequence was compared by BLASTn searching GenBank (Benson et al. 2012), accessed February (2018), to affirm the correct morphological identification. In addition, a subset of the samples was barcoded under a global context by Abboud et al. (2018) and in extent under a phylogenetic context by Gómez Daglio and Dawson (2017) to corroborate the species identification.
Environmental parameter analyses
We analyzed the environmental parameters (Supplementary Material Table S2) recorded at each station. Each parameter data was tested for normality and homoscedasticity in R v. 3.2.4. (R Development Core Team 2020). All the data sets met the assumption of normality and homogeneity of variance. We performed a two-tailed t-test implemented in R v. 3.2.4. (R Development Core Team 2020) to evaluate the differences between the stations sampled close to the coastline (transect 1, 0.5 km) and the stations sampled at 1.5 km (transect 2) from the coast line.
Results
Using an integrative taxonomic approach, we record seven species of Discomedusae jellyfish in the eastern waters of Surabaya (Table 1). Four belong to the Order Rhizostomeae, represented by four families, Mastigiidae, Rhizostomatidae, Lychnorhizidae, and Catostylidae; two species are classified within the order Semaeostomeae, represented by two families, Pelagiidae and Cyaneidae.
Morphological identification and characterization
Order Semaeostomeae
Family Pelagiidae: Chrysaora chinensis Vanhöffen, 1888. Exumbrella surface fine granulated; brown-yellowish stripes radiating from the apical central up to the umbrella margin. Small (< 3 mm in diameter) flecks that covers the exumbrella (Fig. 2a). Four simple curtain-like oral arms attach to a manubrium. Bell margin with four semi-oval velar lappets per octant, three primary tentacles are inserted at the bell margin alternating between each velar lappet. Two rhopalia per quadrat, each rhopalium with two semi-oval lappets (Fig. 2b). Central stomach circular limited by the origin of the radial septae. Radial septae (four per quadrant) are bent proximally to the bell margin (Fig. 2b). Gonads are paired on the interadial axis. Subgenital ostia rounded (Fig. 2c).

Morphological characters for families Pelagiidae and Cyaneidae. Chrysaora chinensis a–c and Cyanea d–f. a Lateral view of an individual collected September 2010; b Subumbrellar octant showing the position of the tentacular insertion, septae shape, velar, and rhopaliar lappets; c gonads and gastrovascular cavity; d apical view of Cyanea sp.; e tentacle horse-shoe shape clusters; f close-up of the rhopalium embedded at ¼ distance from the bell margin. Scale bar = 10 mm. Abbreviation: cm, coronal muscles; g, gonads; m, septae; l, lobes; r, rhopalium; rl, rhopaliar lappets; rm, radial muscles; t, tentacles; vl, velar lappets
Family Cyaneidae: Cyanea sp. Medusae reddish to brownish color (Fig. 2d). Bell with 16 velar lappets (eight primary and eight secondary clefts). Four perradial curtain-like oral arms. Tentacle clusters are located in the adradial axis; the eight clusters have a horseshoe shaped (Fig. 2e). Eight rhopalia are embedded and located ¼ proximally from the lobes (Fig. 2f). Gonads are in out-folded pockets. The specimens were damaged, and features such as the number of coronal and radial muscles and pits were not accounted.
Order Rhizostomeae
Family Catostylidae: Acromitus flagellatus (Maas, 1903) and Catostylus townsendi Mayer, 1915. A. flagellatus is identified by the umbrella dome shape with fine granular protuberances that cover the entire exumbrella. Eight three-winged oral arms covered with short filaments (Fig. 3a). Each quadrant has two rophalia, each rhopalium with two pointed rhopaliar lappets (Fig. 3b) and eight rounded velar lappets per quadrant. Four subgenital ostia, each one with a heart-shaped prominent papillae arranged on the perradial axis (Fig. 3c). The canal system presents a ring canal which connects with one perradial, two interradial, and 12 adradial canal per octant. The perradial and interradial canals extend up to the stomach; adradial canals covers ½ of the surface but do not connect with the stomach. Anastomoses extend up to the bell margin and rhopalia (Fig. 3c). C. townsendi presents a fine granular exumbrella with yellowish-brownish pigmented flecks (Fig. 3d). Oral arms are three-winged with no filaments or terminal clubs. Rhopalium (two per quadrant) is embed on a furrow, with two rounded rhopaliar lappets (Fig. 3e) and 8–12 rounded and elongated velar lappets, some bifurcated. Subgenital ostia has a bone-shaped papillae. The canal system has one perradial, three interradial, and 8–10 adradial canals per octant. Adradial-adradial anastomoses do not reach the central stomach and present sinuses. The canal system extends up to the bell margin and rhopalia (Fig. 3f).

Morphological characters for families Catostylidae and Lychnorhizidae. Acromitus flagellatus a–c. a Close-up of one oral arm covered with filaments; b close-up of the rhopalium with two pointed rhopaliar lappets; c canal system (one octant): one perradial, two interradial, and 12 adradial canals that connects to the ring canal. Heart shape papilla covers the subgenital ostia. Catostylus townsendi d–f. d Apical view of the exumbrella, showing the brown flecks; e close-up of the rhopalium, showing two rounded rhopaliar lappets that are covered with brownish flecks; f canal system (one octant), showing one perradial and two interradial, and 8–10 adradial canals with sinuses. Lyhcnorhiza malayensis g–i. g Bell margin, showing velar lappets, rhopalium, and rhopaliar lappets; h cruciform gonad cavity; i canal system (one quadrat), showing one perradial and interradial and two adradial canals. Blind adradial canals arising from the ring canal; scale bar = 10 mm. Abbreviation: g, gonads; r, rhopalium; rl, rhopaliar lappets; sbp, subgenital ostia papillae; vl, velar lappets
Family Lychniorhizidae: Lychnorhiza malayensis Stiasny, 1920. Medusae with eight three-winged oral arms, without terminal clubs or filaments. Bell margin with eight bifurcated velar lappets and one rhopalium per octant. Rhopalium with two tapering lappets (Fig. 3g). Cruciform gonadal cavity (Fig. 3h). Subgenital ostia with leaf-shape papillae. Canal system with one perradial, one interradial, and two adradial canals that connects to the gastrovascular cavity per quadrant; eight adradial canals that originates in the ring canal but do not reach the gastrovascular cavity (Fig. 3i).
Family Mastigiidae: Phyllorhiza cf. pacifica (Light, 1921). Bell half-dome shaped, yellowish-brownish coloration with small crenulations and white spots that cover the exumbrella (Fig. 4a). Eight three-winged oral arms with fenestrations and intermediate filaments in the winged portion of the oral arms (Fig. 4b). Each oral arm has a single conical shaped capitate terminal club (Fig. 4a). Bell margin with two rhopalia per quadrat, with two oval rhopaliar lappets (Fig. 4d). Velar lappets are rounded variable in number (12–16 per quadrat). Canal system with one perradial, one interradial, and 8–12 adradial canals. Anastomoses reach the stomach and the ring canal (Fig. 4c).

Morphological characters for families Mastigiidae and Rhizostomidae. Phyllorhiza cf. pacifica a–d. a Individuals collected September 2010; b close-up of an oral arm, showing the intermediate filaments with white terminal bulbs; c canal system (one quadrat), showing the complex anastomoses network; d bell margin, showing the semi-oval velar lappets and tapering rhopaliar lappets. Rhopilema hispidum e–h. e Medusae showing the scapula and oral arms; f bell margin, showing the rhopalium with two rhopaliar tapering lappets and oval velar lappets; g subgenital ostia papillae; h canal system, showing the complex anastomosing network that reaches the bell margin. Scale bar = 10 mm. Abbreviation: r, rhopalium; rl, rhopaliar lappets; sbp, subgenital ostia papillae; vl, velar lappets
Family Rhizostomatidae: Rhopilema hispidum (Vanhöffen, 1888). Exumbrella with dark red spots radiating from the center up to the bell margin; pointy conical protuberances cover the entire exumbrella (Fig. 4e). Bell margin with eight semi-oval velar lappets and one rhopalia per octant, each rhopalia with two tapering lappets (Fig. 4f). Eight oral arms with multiple short spatulate shape terminal clubs, two times larger attached at the distal portion of the oral arm. Each oral arm has two scimitar-shaped attached to the smoot portion (Fig. 4e). Scapulae with filaments. Subgenital ostia with a flattened oval papillae (Fig. 4g). Canal system with one perradial, three interradial, and two adradial canals that join the gastrovascular cavity per quadrat. Adradial anastomoses make a complex mesh that includes multiple (> 179) sinuses per quadrant (Fig. 4h).
Molecular characterization
We amplified 23 COI sequences. The resultant alignments per genus did not show sequence difference larger than 0.5%. The BlastN search result for the consensus sequences per genus confirms the morphological identification at the genus level (Table 2).
Discomedusae records
A total of 1,095 Discomedusae were recorded in the Eastern coast of Surabaya (Table 3). The largest amount of Discomedusae was recorded along the transect 1 (72.3%), with records of A. flagellatus (55.7%), L. malayensis (42.7%), and Rhopilema (1.6%). There are no records of Cyanea sp. and C. chinensis in any of the stations close to the shore (transect 1). A. flagellatus (38.6%) and L. malayensis (28.4%) represent the most recorded species along the transect 2, followed by Cyanea (14.2%), C. chinensis (11.6), and R. hispidum (7.3%). Overall, A. flagellatus cf. is the species with highest number of records (50.9%), followed by L. malayensis (38.7%). R. hispidum, Cyanea sp., and C. chinensis represent the remaining 9%.
Environmental parameters
The analyses of the environmental parameters between the two transects (T-1, 0.5 km and T-2, 1.5 km) show that pH, salinity, flow velocity, and lucidity are statistically different between both transects (Table 4). There is no statistical difference between the temperatures between both transects (p = 0.19).
Discussion
Discomedusae taxonomic diversity
There are 35 species recorded for Indonesia (Table S1; Kramp 1961; Swift et al. 2016; De Souza and Dawson 2018; Jarms and Morandini 2019). Ten of these recorded species’ distribution are not confirmed for Indonesia (Table S1). For example, Anomalorhiza shawi Light, 1921 is reported for the Philippine’s islands (Light 1921; Stiasny 1924); recent literature extends its distribution to the Western Pacific (Jarms and Morandini 2019) without any reports or records that demonstrate its presence in Indonesia or other adjacent areas. Indonesia sea is a complex oceanographic area (Pariwono et al. 2005), where the tidal patterns, surface temperature profiles, thermocline, upwellings, chlorophyll, currents, and water influx vary seasonally and yearly at a small scale due the higher complexity in the island configuration, internal waves, and topography. Also, at large scale, oceanographic conditions such as East Asian Monsoons and El Niño (Gordon 2005; Qu et al. 2005; Susanto and Marra 2005; Susanto et al. 2005) have an effect in the oceanographic conditions of the area. These exceptional oceanographic conditions increase the complexity of marine ecosystems in Indonesia sea. Hence, the distribution of Discomedusae jellyfish should not be extrapolated to entire regions without valid reports.
This study provides the first systematic records of Discomedusae in the Strait of Surabaya (Indonesia) under an integrative approach. Previously only two species (Chrysaora chinensis and (Cyanea nozakii Kishinouye, 1891) were recorded for the Straight of Surabaya (Table S1). The Strait of Surabaya as part of the Java Sea is being subject of a heavy jelly-fishery (Brotz 2016). However, the identification of the targeted species is still questioned due the lack of more thorough studies that describe the diversity of jellyfish in the area that can provide tools for a reliable identification in situ.
Discomedusae identification can be problematic due the high incidence of cryptic species (Dawson 2003, 2005b; Gómez Daglio and Dawson 2017). For example, within the order Semaeostomeae, the genera Chrysaora (Morandini and Marques 2010; Bayha et al. 2017) and Cyanea are cryptic species (Dawson 2005b; Holst and Laakmann 2014). We identify Chrysaora chinensis. C. chinensis is the only pelagiid reported for Indonesia waters (Stiasny 1921: originally named C. quinquecirrha Desor, 1848 (Morandini and Marques 2010)). Other records are from China, Vietnam, and the Philippines (Mayer 1915, 1917; Stiasny 1940). The individuals collected in this study present some morphological variations with respect to the redescription of C. chinensis published by Morandini and Marques (2010), such as gonads’ shape (rounded pockets, Fig. 2c), gastric filaments (absent), and coloration of exumbrella (yellow and brownish, Fig. 2a). The variation in these characters might be the results of the high intraspecific morphological variation of Chrysaora species, as it is demonstrated by other studies in other regions of the world and species (e.g., C. lactea (Morandini et al. 2006) and C. quinquecirrha (Bayha et al. 2017)). The identification of individuals for the family Cyaneidae is done to the genus level. The individuals collected were broken or in poor conditions, which make difficult to account for a reliable morphological characterization. Cyanea buitendijki Stiasny, 1919 is the only species described for the area (Malayan Archipelago; Stiasny and Van der Maaden 1943). The poor conditions of the samples and the lack of molecular data (our study or published) did not give enough characters to provide an accurate identification for this species.
We identify five rhizostome medusae. The specimens of C. townsendi show the diagnostic characters (Fig. 3d–c) described by Mayer (1915) and Stiasny (1921). This species has been reported for Malayan Archipelago, South East Asian ocean, and Indonesia (Stiasny 1921; Stiasny 1922; Ranson 1949; Rizman-Idid et al. 2016). The alignment from BlastN (Table 2) and the phylogenetic analyses made by Gómez Daglio and Dawson (Fig. 2 pag. 644; 2017) corroborates the identification as a Catostylus. A. flagellatus have been reported previously for the Malayan Archipelago and Java Sea (Stiasny 1921, 1922; Rizman-Idid et al. 2016). The individuals show the diagnostic characteristic of the species: intermediate filaments in the oral arms with a pointy terminal club (Fig. 3a) and canal system with a heart shape papillae on each perradial canal (Fig. 3c). The alignment comparison (Table 2) shows a ~ 90% identity from other individuals collected from China and Malayan Archipelago. That might suggest that the individuals collected in Surabaya might represent a different species from the individuals collected by Rizman-Idid et al. (2016) in the Malayan Archipelago. A description of specimens from the Malayan Archipelago and the recorded distribution of the species (China, Taiwan, Japan, Indian-Ocean, and Borneo (Kramp 1961)) are needed to clarify the identification.
The identification of Lychnorhiza malayensis relies on the comparison of the morphological data with the original description (Stiasny 1920, 1932). However, some morphological variations were noticed with respect reports from different geographic areas (India and Pakistan (Menon 1930; Gul and Osmany 2017). According to Abboud et al. (2018), there is a high likelihood that different species of Lychnorhiza might occur on small geographic ranges. To define the intraspecific variation and corroborate the identification with molecular data, we cannot discard the possibility of the presence of multiple species in the coast of Surabaya.
Phyllorhiza individuals were identified as P. cf. pacifica. The alignment of COI (Table 2) shows a high similarity with P. punctata von Lendenfeld, 1884 reported for the Strait of Malacca (Rizman-Idid et al. 2016). However, the morphological characters (terminal clubs and lappets number and shape, Fig. 4a–c) do not resemble the original description of P. punctata. In addition, these samples were analyzed under a phylogenetic context; the individuals from Surabaya belong to a different clade with respect to P. punctata (see Gómez Daglio and Dawson 2017: Fig. 2: pp 644).
Rhopilema hispidum is morphologically similar to those recorded in Pakistan (Gul and Morandini 2015) and the strait of Malacca (Rizman-Idid et al. 2016). The BlastN alignment shows a ~ 94% of similarity with the documented sequences from Malayan Archipelago. R. hispidum presents a broad distribution in the western Pacific (Japan, China, Philippines, Indonesia, Indian Ocean, and the Red Sea (Kitamura and Omori 2010; Gul and Morandini 2015; Rizman-Idid et al. 2016)). There are no molecular data available to compare our samples with individuals from other geographic locations.
Distribution and seasonality
The distribution and seasonality of jellyfish have been attributed to changes in environmental conditions, anthropogenic activities, and intrinsic factors (e.g., life cycle, reproduction; Suchman et al. 2012; Pikesley et al. 2014; Ceh et al. 2015; Quiñones et al. 2015). Our surveys from November to December (2010) show a higher incidence of jellyfishes close to the shoreline (Transect 1, Table 3). Transect 1 stations (0.5 km from the coast) are exposed to tidal and current conditions, whereas Transect 2 stations (1.5 km) are not highly influenced by those parameters. The environmental parameters in transect 1 are statistically different to transect 2 (offshore), except for the temperature. That suggest that the environmental conditions limit the distribution of jellyfishes close to the shore, and rhizostomes (A. flagellatus, L. malayensis, and R. hispidum) are present in transect 1 and 2 (Table 2); meanwhile the semaeostomes (C. chinensis and Cyanea sp.) only in transect 2. The correlation between the presence of jellyfish and environmental parameters shows a high correlation (Purcell et al. 2000; Lee et al. 2013; Quiñones et al. 2015; Kienberger and Prieto 2017), where the salinity, temperature, and currents are the main drivers of its presence.
The environmental factors solely cannot be the only explanation for the high incidence of rhizostomes close to the shore, and the biology of the species (feeding mechanisms and behavior, diet, life cycle, and reproduction) plays an important role in the distribution and presence of jellyfishes. Rhizostomes are shallow-water, coastal, and tropical-temperate species; their differences in lifestyle with respect to semaeostomes, such as the mutualistic association with symbiotic zooxanthella (e.g., Phyllorhiza spp. and Catostylus spp. (Dawson and Hamner 2009), their feeding behaviors (filter feeders) (Arai 1997)), could explain their presence and high occurrence close to the shore. On the other hand, semaeostome jellyfish are voracious predators on zooplankton (Purcell 2003), and their life cycle does not always follow the metagenesis—alternation between sexual and asexual generations (Ceh et al. 2015, but see Morandini et al. 2016). For the species recorded here, there are no information about the biology of the species and their seasonality. The information available about the Discomedusae distribution and abundance is scarce, and the information comes from the fisheries (landing values). However, these data are unreliable, due to the high incidence of misidentification of the species (Gul and Morandini 2015; Rizman-Idid et al. 2016).
Conclusion
Herein we identify seven species of Discomedusae and provide the first records for R. hispidum, A. flagellatus, C. townsendi, L. malayensis, and P. cf. pacifica along the coast of Surabaya. The taxonomic knowledge for this area is still patchy, and misidentifications occur oftentimes. We highlight the importance to document the taxonomic diversity in the area, using an integrative approach (e.g., morphological and molecular data), that proves to solve part of the taxonomic uncertainty. Indonesia, as a country with a high fishery activity, requires a good documentation of its unique biodiversity, tools that help to identify the species, and a continuous monitoring of the targeted species. In addition, to understand the seasonality and distribution of the jellyfish, it is necessary to grow the body knowledge of the biology of the species that should include the reproduction, feeding behavior, diet, and life cycle. This knowledge will help to document and understand the ecological patterns of these economic important species.
References
Abboud SS, Gómez Daglio L, Dawson MN (2018) A global estimate of genetic and geographic differentiation in macromedusae—implications for identifying the causes of jellyfish blooms. Mar Ecol Prog Ser 591:199–216. https://doi.org/10.3354/meps12521
Allen GR (2007) Conservation hotspots of biodiversity and endemism for Indo-Pacific coral reef fishes. Aquat Conserv: Mar Freshw Ecosyst 18:541–556. https://doi.org/10.1002/aqc.880
Arai MN (1997) A functional biology of Scyphozoa. Chapman & Hall, New York
Asrial E, Prajitmo A, Susilo E, Bintoro G (2015) Biology and blooms of the edible jellyfish (Crambione mastigophora) in the Saleh Bay, Indonesia. J Biodivers Environ Sci 6:356–369
Attrill MJ, Wright J, Edwards M (2007) Climate-related increases in jellyfish frequency suggest a more gelatinous future for the North Sea. Limnol Oceanogr 52:480–485. https://doi.org/10.4319/lo.2007.52.1.0480
Barber PH, Bellwood DR (2005) Biodiversity hotspots: evolutionary origins of biodiversity in wrasses (Halichoeres: Labridae) in the Indo-Pacific and new world tropics. Mol Phylogenet Evol 35:235–253. https://doi.org/10.1016/j.ympev.2004.10.004
Barber P, Boyce SL (2006) Estimating diversity of Indo-Pacific coral reef stomatopods through DNA barcoding of stomatopod larvae. Proc R Soc B Biol Sci 273:2053–2061. https://doi.org/10.1098/rspb.2006.3540
Bayha KM, Dawson MN, Collins AG et al (2010) Evolutionary relationships among scyphozoan jellyfish families based on complete taxon sampling and phylogenetic analyses of 18S and 28S ribosomal DNA. Integr Comp Biol 50:436–455. https://doi.org/10.1093/icb/icq074
Bayha KM, Collins AG, Gaffney PM (2017) Multigene phylogeny of the scyphozoan jellyfish family Pelagiidae reveals that the common U.S. Atlantic Sea nettle comprises two distinct species (Chrysaora quinquecirrha and C. chesapeakei). PeerJ 5:3863–3843. https://doi.org/10.7717/peerj.3863
Benson DA, Cavanaugh M, Clark K et al (2012) GenBank. Nucleic Acids Res 41:D36–D42. https://doi.org/10.1093/nar/gks1195
Brotz L (2016) Jellyfish fisheries – a global assessment. In: Pauly D, Zeller D (eds) Atlas of Marine Fisheries ecosystems impacts and analysis. Island Press, Washington DC, pp 110–124
Brotz L, Cheung WWL, Kleisner K et al (2012) Increasing jellyfish populations: trends in Large Marine Ecosystems. Hydrobiol 690:3–20. https://doi.org/10.1007/s10750-012-1039-7
Brotz L, Schiariti A, López-Martínez J et al (2017) Jellyfish fisheries in the Americas: origin, state of the art, and perspectives on new fishing grounds. Rev Fish Biol Fisheries 27:1–29. https://doi.org/10.1007/s11160-016-9445-y
Ceh J, Gonzalez J, Pacheco AS, Riascos JM (2015) The elusive life cycle of scyphozoan jellyfish – metagenesis revisited. Sci Rep 5:12037. https://doi.org/10.1038/srep12037
Condon RH, Graham WM, Duarte CM et al (2012) Questioning the rise of gelatinous zooplankton in the world’s oceans. Bioscience 62:160–169. https://doi.org/10.1525/bio.2012.62.2.9
Dawson MN (2003) Macro-morphological variation among cryptic species of the moon jellyfish, Aurelia (Cnidaria: Scyphozoa). Mar Biol 143:369–379. https://doi.org/10.1007/s00227-003-1070-3
Dawson MN (2005a) Morphologic and molecular redescription of Catostylus mosaicus conservativus (Scyphozoa: Rhizostomeae: Catostylidae) from south-east Australia. J Mar Biol Assoc U K 85:723–731. https://doi.org/10.1017/S002531540501163X
Dawson MN (2005b) Cyanea capillata is not a cosmopolitan jellyfish: morphological and molecular evidence for C. annaskala and C. rosea (Scyphozoa: Semaeostomeae: Cyaneidae) in south-eastern Australia. Invert Syst 19:361–370. https://doi.org/10.1071/IS03035
Dawson MN, Jacobs DK (2001) Molecular evidence for cryptic species of Aurelia aurita (Cnidaria, Scyphozoa). Biol Bull 200:92–96. https://doi.org/10.2307/1543089?ref=search-gateway:7315323a8a81d3fc48fe154ee3095708
Dawson MN, Hamner WM (2009) A character-based analysis of the evolution of jellyfish blooms: adaptation and exaptation. Hydrobiol 616:193–215. https://doi.org/10.1007/s10750-008-9591-x
Desor E (1848) Meeting of the November 1, 1848. Proc Boston Soc Nat Hist 3:75–76
De Souza M, Dawson MN (2018) Redescription of Mastigias papua (Scyphozoa, Rhizostomeae) with designation of a neotype and recognition of two additional species. Zootaxa 4457:520–536. https://doi.org/10.11646/zootaxa.4457.4.2
Dong Z, Liu D, Keesing JK (2010) Jellyfish blooms in China: dominant species, causes and consequences. Mar Pollut Bull 60:954–963. https://doi.org/10.1016/j.marpolbul.2010.04.022
English S, Wilkinson C, Baker V (1994) Survey manual for tropical marine research. Australian Institute of Marine Science, Townville
Folmer O, Black M, Hoeh W, et al (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3:294–299. ASEAN-Australia Matine Science Project. Australia Institute of Marine Science, Townsville
Gómez Daglio L, Dawson MN (2017) Species richness of jellyfishes (Scyphozoa: Discomedusae) in the Tropical Eastern Pacific: missed taxa, molecules, and morphology match in a biodiversity hotspot. Invertebr Syst 31:635–663. https://doi.org/10.1071/IS16055
Gordon AL (2005) Oceanography of the Indonesian seas and their throughflow. Oceanogr 4:14–21. https://doi.org/10.5670/oceanog.2005.07
Gul S, Morandini AC (2015) First record of the jellyfish Rhopilema hispidum (Cnidaria: Scyphozoa) from the coast of Pakistan. Mar Biodivers Rec 8:30–34. https://doi.org/10.1017/S175526721500007X
Gul S, Osmany HB (2017) Additions to the Scyphomedusae (Cnidaria: Scyphozoa) of Pakistan. Int J Biol Biotech 14:665–667. https://doi.org/10.1016/j.watres.2019.03.087
Haeckel E (1880) Systems der Acraspeden. Jena, Germany
Holland BS, Dawson MN, Crow GL, Hofmann DK (2004) Global phylogeography of Cassiopea (Scyphozoa: Rhizostomeae): molecular evidence for cryptic species and multiple invasions of the Hawaiian Islands. Mar Biol 145:1119–1128. https://doi.org/10.1007/s00227-004-1409-4
Holst S, Laakmann S (2014) Morphological and molecular discrimination of two closely related jellyfish species, Cyanea capillata and C. lamarckii (Cnidaria, Scyphozoa), from the northeast Atlantic. J Plankton Res 36:48–63. https://doi.org/10.1093/plankt/fbt093
Hubert N, Meyer CP, Bruggemann HJ et al (2012) Cryptic diversity in Indo-Pacific coral-reef fishes revealed by DNA-barcoding provides new support to the centre-of-overlap hypothesis. PLoS ONE 7:e28987–e28988. https://doi.org/10.1371/journal.pone.0028987
Jarms G, Morandini AC (2019) World Atlas of Jellyfish. Dölling und Galitz Verlag, Hamburg
Kienberger K, Prieto L (2017) The jellyfish Rhizostoma luteum (Quoy & Gaimard, 1827): not such a rare species after all. Mar Biodiv 219:1–8. https://doi.org/10.1007/s12526-017-0637-z
Kishinouye K (1891) Cyanea nozakii, nov. sp. Zool. Magazine (Dobutsugaku zasshi). 3:93–95.
Kitamura M, Omori M (2010) Synopsis of edible jellyfishes collected from Southeast Asia, with notes on jellyfish fisheries. Plankton Benthos Res 5:106–118. https://doi.org/10.3800/pbr.5.106
Kramp PL (1961) Synopsis of the medusae of the world. J Mar Biol Assoc 40:1–496
Larkin MA, Blackshields G, Brown NP et al (2007) Clustal W and Clustal X version 2.0. Bioinformatics 23:2947–2948. https://doi.org/10.1093/bioinformatics/btm404
Lee PLM, Dawson MN, Neill SP et al (2013) Identification of genetically and oceanographically distinct blooms of jellyfish. J Royal Soc Interface 10:20120920–20120920. https://doi.org/10.1016/j.tree.2009.01.010
Light SF (1914) Some Philippine Scyphomedusae, including two new genera, five new species, and one new variety. Philippine J Sci 9: 195–231
Light SF (1921) Further notes on Philippine scyphomedusan jellyfishes. Philippine J Sci 18:25–48
Maas O (1903) Die scyphomedusen der Siboga-expedition. Sigboa Expedition Monograph 11:1–93
Maas O (1907) Die Scyphomedusen. Ergebn Fortsch Zool II:13–238. https://doi.org/10.5962/bhl.title.11300
Marques A, Collins A (2004) Cladistic analysis of Medusozoa and cnidarian evolution. Invertebr Biol 123:23–42. https://doi.org/10.1111/j.1744-7410.2004.tb00139.x
Mayer AG (1910) Medusae of the world. Carnegie Institution of Washington, Washington DC
Mayer AG (1915) Medusae of the Philippines and of Torres Straits. Being a report upon the Scyphomedusae collected by the United States Fisheries Bureau Steamer “Albatross” in the Philippine islands and Malay Archipelago, 1901–1910, and upon the Medusae collected by the expedition of the Carnegie Institution of Washington to Torres Straits, Australia, in 1913. Carnegie Institution of Washington Pub 8:157–202
Mayer AG (1917) Report upon the scyphomedusae collected by the United States Bureau of fisheries steamer “Albatross” in the Philippine Islands and Malay Archipelago. Bull US Nat Mus 1:181–233
Menon MGK (1930) The scyphomedusae of Madras and the neighbouring coast. Bull Madras Gov Mus 3:1–28
Mihaljević M, Korpanty C, Renema W (2017) (2017) Identifying patterns and drivers of coral diversity in the Central Indo-Pacific marine biodiversity hotspot. Paleobiology 43:343–364. https://doi.org/10.5061/dryad.3k5v6
Morandini AC, Marques AC (2010) Revision of the genus Chrysaora Peron & Lesueur, 1810 (Cnidaria: Scyphozoa). Zootaxa 2464:1–97. https://doi.org/10.11646/zootaxa.2464.1.1
Morandini AC, De Silveira FL, Cornelius PF (2006) Redescription of Chrysaora lactea Eschscholtz, 1829 (Cnidaria, Scyphozoa) from the Brazilian coast, with designation of a neotype. Zootaxa 1135:29–48. https://doi.org/10.11646/ZOOTAXA.1135.1.2
Morandini AC, Schiariti A, Stampar SN, Maronna MM, Straehler-Pohl I, Marques AC (2016) Succession of generations is still the general paradigm for scyphozoan life cycles. Bull Mar Sci 92:342–351. https://doi.org/10.5343/bms.2016.1018
Moretzsohn F, McShane MK (2004) Mapping marine invertebrate biodiversity Hotspots in the Indo-Pacific Ocean using GIS. Bishop Museum, Honolulu
Nishikawa J, Ohtsuka S, Mulyadi et al (2014) A new species of the commercially harvested jellyfish Crambionella (Scyphozoa) from central Java, Indonesia with remarks on the fisheries. J Mar Biol Assoc UK 95:471–481. https://doi.org/10.1017/S002531541400157X
Ohtsuka S, Kondo Y, Sakai Y et al (2010) In-situ observations of symbionts on Medusae occurring in Japan, Thailand, Indonesia and Malaysia. Bull Hiroshima Univer Mus 2:9–18. https://doi.org/10.15027/32060
Omori M, Nakano E (2001) Jellyfish Fisheries in Southeast. Asia Hydrobiol 451:19–26. https://doi.org/10.1023/A:1011879821323
Ortman BD, Bucklin A, Pages F, Youngbluth M (2010) DNA barcoding the Medusozoa using mtCOI. Deep-Sea Res II 57:2148–2156. https://doi.org/10.1016/j.dsr2.2010.09.017
Pariwono JI, Ilahude AG, Hutomo M (2005) Oceanography of the Indonesian seas and their throughflow. Oceanogr 4:42–29. https://doi.org/10.5670/oceanog.2005.04
Pikesley SK, Godley BJ, Ranger S et al (2014) Cnidaria in UK coastal waters: description of spatio-temporal patterns and inter-annual variability. J Mar Biol Assoc UK 94:1401–1408. https://doi.org/10.1017/S0025315414000137
Purcell JE (2003) Predation on zooplankton by large jellyfish, Aurelia labiata, Cyanea capillata and Aequorea aequorea, in Prince William Sound, Alaska. Mar Ecol Prog Ser 246:137–152. https://doi.org/10.3354/meps246137
Purcell JE, Brown ED, Stokesbury KDE et al (2000) Aggregations of the jellyfish Aurelia labiata: abundance, distribution, association with age-0 walleye pollock, and behaviors promoting aggregation in Prince William Sound, Alaska, USA. Mar Ecol Prog Ser 195:145–158. https://doi.org/10.3354/meps195145
Purcell JE, Uye S, Lo W (2007) Anthropogenic causes of jellyfish blooms and their direct consequences for humans: a review. Mar Ecol Prog Ser 350:153–174. https://doi.org/10.3354/meps07093
Qu T, Du Y, Strachan J, Meyers G, Slingo J (2005) Sea surface temperature and its variability in the Indonesian region. Oceanogr 18:50–61. https://doi.org/10.5670/oceanog.2005.05
Quiñones J, Mianzan H, Purca S et al (2015) Climate-driven population size fluctuations of jellyfish (Chrysaora plocamia) off Peru. Mar Biol 162:1–12. https://doi.org/10.1007/s00227-015-2751-4
R Core Team (2020) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ranson G (1949) Résultats Scientifiques des Croisières du Navire-école Belge ''Mercator'' Meduses. Mém Mus Royal d'histoire naturelle de Belgique 33:123–158
Rao HS (1931) Notes on scyphomedusae in the Indian museum. Rec Indian Museum 33:25–62
Richardson AJ, Bakun A, Hays GC, Gibbons MJ (2009) The jellyfish joyride: causes, consequences and management responses to a more gelatinous future. Trends Ecol Evol (amst) 24:312–322. https://doi.org/10.1016/j.tree.2009.01.010
Rizman-Idid M, Farrah-Azwa AB, Chong VC (2016) Preliminary taxonomic survey and molecular documentation of jellyfish species (Cnidaria: Scyphozoa and Cubozoa) in Malaysia. Zool Studies 55:1–19. https://doi.org/10.6620/ZS.2016.55-35
Roux J-P, van der Lingen CD, Gibbons MJ et al (2013) Jellyfication of marine ecosystems as a likely consequence of overfishing small pelagic fishes: lessons from the Benguela. B Mar Sci 89:249–284. https://doi.org/10.5343/bms.2011.1145
Suchman CL, Brodeur RD, Daly EA, Emmett RL (2012) Large medusae in surface waters of the Northern California Current: variability in relation to environmental conditions. Hydrobiol 690:113–125. https://doi.org/10.1007/s10750-012-1055-7
Stiasny G (1919) Die Scyphomedusen-Sammlung des Naturhistorischen Reichsmuseums in Leiden: II. Stauromedusae, Coronatae. Semaeostomeae. Zool. Mededeelingen 5:66–99
Stiasny G (1920) Die scyphomedusen-sammlung des naturhistorischen reichs museums in Leiden. III, Rhizostomae. Zool Mededeelingen 5:213–230
Stiasny G (1921) Mittheilungen über Scyphomedusen I. Zool. Mededeelingen 6:109–114
Stiasny G (1922) Die scyphomedusen-sammlung von Dr. Th. Mortensen nebst anderen medusen aus dem zoologischen Museum der Universität in Kopenhagen. Vidensk Meddel Dansk Naturhist Foren Kjøbenhavn 73:513–558
Stiasny G (1924) Rhizostomeen von Manila. Zool Mededeelingen 5:39–54
Stiasny G (1932) Üeber Lychnorhiza malayensis Stiasny. Zool Mededeelingen 19:89–95
Stiasny G (1940) Die Scyphomedusen. The Carlsberg foundations oceanographical expedition round the world and previous Dana-expeditions 18:1–27
Stiasny G, Van der Maaden H (1943) Über Scyphomedusen aus dem Ochotskischen und Kamtschatka meer nebst einer kritik der genera Cyanea und Desmonema. Zool Jahrb Abt Syst Oekol Geog Tiere 76:228–266
Susanto RD, Marra J (2005) Effect of the 1997/98 El Niño on chlorophyll a variability along the southern coasts of Java and Sumatra. Oceanogr 18:124–127. https://doi.org/10.5670/oceanog.2005.13
Susanto RD, Mitnik L, Zheng Q (2005) Ocean internal waves observed in the Lombok Strait. Oceanogr 18:80–87. https://doi.org/10.5670/oceanog.2005.08
Swift HF, Gomez Daglio L, Dawson M (2016) Three routes to crypsis: stasis, convergence, and parallelism in the Mastigias species complex (Scyphozoa, Rhizostomeae). Mol Phylogenet Evol 99:103–115. https://doi.org/10.1016/j.ympev.2016.02.013
Vanhöffen E (1888) Untersuchungen über semaeostome und rhizostome Medusen. Bibliotheca Zoologica I:52
von Lendenfeld R (1884) The scyphomedusae of the southern hemisphere. Part III. Proc Linn Soc new South Wales 9:259–306
Waterhouse AM, Procter JB, Martin DMA et al (2009) Jalview Version 2–a multiple sequence alignment editor and analysis workbench. Bioinformatics 25:1189–1191. https://doi.org/10.1093/bioinformatics/btp033
Acknowledgements
This research paper was possible through the help and dedication from numerous parties. It is with great appreciation that we thank the members of the Laboratory of Ecology—Biology ITS and the numerous fishermen from Kenjeran, Surabaya, especially Pak Malik and Pak Aripin who contributed to the completion of this study. We thank Dr. Michael Dawson to facilitate his lab for molecular and morphological analyses. We thank the Ministry of Home Affairs Echelons of Indonesia government for the collecting and exporting permits (1551/FRP/SM/VI).
We are grateful for all the curatorial assistance provided by the Museum of Comparative Zoology, specially to Adam Baldinger and Penny Benson.
We thank all the comments and feedback provided by the anonymous reviewers.
Funding
This study was partially funded by the Fulbright Senior Research Scholarship, the NIH/NINDS AREA (1R15-NS060099-01 and NIH-MBRS SCORE-5S06GM048680-13) to MEdB.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Ethical approval
No animal testing was performed during this study.
Sampling and field studies
All necessary permits for sampling have been obtained by the authors from the competent authorities and are mentioned in the acknowledgements.
Data availability
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Author contribution
Dian Saptarini and MEdB conceived and designed the research. Ria Hayati and Farid Muzaki conducted experiments. Dian Saptarini, Ria Hayati, Aunurohim, and Farid Muzaki analyzed the data. Liza Gómez Daglio conducted the PCR and identification of species. Liza Gómez Daglio, Tiffany Coleman, Yoo-Min Han, and MEdB wrote the manuscript. All authors read and approved the manuscript.
Additional information
Communicated by S. Piraino
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gómez Daglio, L., Hayati, R., Coleman, T. et al. Species composition of Discomedusae jellyfish (Scyphozoa) in the coastal waters of Eastern Surabaya, East Java. Mar. Biodivers. 52, 23 (2022). https://doi.org/10.1007/s12526-021-01253-1
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12526-021-01253-1