
Abstract
The Pastena cave is located in central Italy, and its best-preserved sector is Grotticella W2, which is dated radiometrically to the Early-Middle Bronze Age. The aim of this paper is to explore human diet, animal husbandry, and plant management, analysing the findings there discovered. Carbon and nitrogen stable isotope analysis was carried out on 40 charred seeds, six faunal remains, and four human individuals, investigating the whole bio-archaeological material available. To the best of our knowledge, this is one of the first papers presenting stable isotope analysis on carpological remains dated to the Italian Early-Middle Bronze Age. The obtained results are consistent with a diet based on terrestrial protein, mainly on plants, and secondly on animal products. The data suggest that plants, especially broad beans, were partially subjected to human management, while livestock was managed through different husbandry strategies. The cooperation between archaeological studies and molecular analysis allows us to contribute to clarifying the economic strategies for a Central Italian community in a scenario that is still poor in published data.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Bronze Age (BA) is one of the crucial periods of late Italian Prehistory (Bietti Sestieri 2015). Even though this phase presents different characteristics and chronologies in Europe (Fokkens and Harding 2013), it is assumed that its chronological boundaries dated around 2200–950 BCE in the Italian peninsula (Peroni 1996; Bietti Sestieri 2015; for an updated discussion about the Early (EBA) and Middle Bronze Age (MBA) chronology, see Alessandri 2019). Despite the plethora of archaeological surveys (e.g., Cocchi Genick 1995; Barfield 2007; Guidi and Rosini 2019; Skeates et al. 2021), the bioarchaeological ones are still sparse and refer to sites scattered throughout the Peninsula, with a lack of data relating to Central-South Italy. The main object of our study is the Pastena cave, which returned traces of human frequentation starting from the Late Neolithic to the Middle Bronze Age (MBA) (Guareschi and Morandini 1943; Biddittu 1987; Biddittu et al. 2007; Angle et al. 2010; Rolfo et al. 2021). Specifically, we focus on the BA bioarchaeological findings, which could act as significant pieces of evidence for supporting hypotheses related to complex dynamics in the Central Italian BA. Thus, we would contribute to the claimed need for denser and timely resolved evidence to assess the spread of BA cultures in Italy (Saupe et al. 2021; Romboni et al. 2022).
Indeed, the BA is a period of massive technological, cultural, social, and economic changes for human communities both in Europe and the Italian peninsula (Fokkens and Harding 2013; Alessandri 2013). The spread of bronze metallurgy is the most important technological innovation, and its processing required specialized roles leading to the rise of social complexity. Indeed, as the bronze items were often considered prestige goods and personal ornaments, their availability reflected the social stratification of the communities, which was supposed to be recalled, in turn, by the discovery of these items as goods in the burial grounds (Bettelli 2006; Alessandri 2013; Fokkens and Harding 2013).
A complex territorial organization also characterizes BA through the development of hierarchical residential systems, which again suggest a complex social organization: nucleated settlements, fortified villages, pile-dwellings, and Terramare hosted a growing number of inhabitants (Bettelli 2006; Fokkens and Harding 2013). Despite the increasing average lifespan of the settlements, the discovery of exotic items and recent stable isotope analysis on human bones suggest a rise in both individual and artifact mobility in the BA (e.g., Knipper et al. 2017; Cavazzuti et al. 2019; Cavazzuti et al. 2021). Goods and people were involved in complex exchange networks (Blake 2014; Cavazzuti et al. 2019), whose effects could be identified in heterogeneous lifestyles and bio-cultural characteristics (Fokkens and Harding 2013; Bietti Sestieri 2015).
A multifaceted funerary scenario also reflected the heterogenous dwelling strategy. Multiple regional specificities could be outlined across the Italian peninsula, mainly concerning pit graves, mounds, or artificial and natural cave burials, where the last were often used for ritual purposes (Bettelli 2006; Whitehouse 2007; Minniti 2012; Bietti Sestieri 2015).
Indeed, most of the knowledge for BA in Central Italy came from karst environments, as numerous caves hosting BA evidence characterize this area (Sestini 1934; Rittatore 1951a, b; Cocchi Genick 1995; Cavanna 2007; Alessandri et al. 2021; Rolfo et al. 2021).
So far, the analysis of biological remains from multiple sites complemented the archaeological findings and supported a comprehensive understanding of the complex dynamics occurring in the Central Italian BA.
The carbon and nitrogen stable isotope analysis on bio-archaeological remains is considered able to identify the dietary habits and, in turn, the economy and social behavior of past communities (e.g., Shoeninger and DeNiro 1984; Katzenberg 2008; Fontanals-coll et al. 2015; Goude et al. 2016; De Angelis et al. 2019, 2020; Varalli et al. 2021, 2022; Romboni et al. 2022). Briefly, the collagen from bones is extracted, and its carbon and nitrogen isotopic signature would relate to the dietary habits. Specifically, the estimation of the ratios of 13C/12C and 15N/14N, detectable from the collagen, reflects the diet of the last decades of life (Ambrose 1990; Hedges and Reynard 2007). The stable carbon isotopes ratio is suitable for determining the consumption of C3 (typical of temperate environments) and C4 plants (mainly from arid ecosystems), as well as marine and freshwater intake (Ambrose and Norr 1993; Krigbaum 2003; Marshall et al. 2007; Katzenberg 2008). Conversely, the analyses of stable nitrogen isotopes (14N and 15N) provide information related to the trophic position of an organism (Shoeninger and DeNiro 1984; Keegan 1989; Unkovich et al. 2001).
Human dietary habits are one of the most retained markers of the people and societies’ cultural identity (De Angelis et al. 2019) and have been extensively dissected through stable isotope analysis across the Italian BA communities (e.g., Varalli et al. 2015, 2022; Tafuri et al. 2018; Arena et al. 2020; Romboni et al. 2022). Indeed, these findings are consistent with the archaeological and archeozoological data, suggesting that the BA human groups had a mixed economy, mainly based on agriculture and breeding practices tightly related to the local ecosystem. Conversely, wild gathering through hunting and fishing was less practiced (De Grossi 1995; Cazzella 2009; Cattani and Marchesini 2010; Maini 2010; Rolfo et al. 2013; Salari and Tagliacozzo 2019).
However, new crops spread throughout the European continental areas during the BA (Miller et al. 2016). C4 plants (e.g., the common millet, Panicum miliaceum, or broomcorn millet) appeared consistently in Europe in the late 3rd millennium BC, to become established during the second half of the 2nd millennium BC (Stevens et al. 2016; Valamoti 2016). These plants are well suited to an arid environment. As fast-growing, warm-season crops, their cultivation probably reduced agricultural risk as a low-investment rain-fed crop (Stevens et al. 2016). The impact of these plants on the Italian BA diet has been previously outlined also by applying carbon and nitrogen stable isotope analysis (Tafuri et al. 2009, 2018; Varalli et al. 2015; Romboni et al. 2022), supporting the role of the human population dispersal across the peninsula (Saupe et al. 2021; Romboni et al. 2022; Varalli et al. 2022) for the dissemination of these resources. Evidence of C4 plant consumption was also reported for previous periods in Italy (Mariotti Lippi et al. 2017; Nava et al. 2021). However, whether these C4 plants were gathered as wild plants or served as animal fodder rather than extensively cultivated and consumed by people is not fully understood. Despite the local or occasional consumption of these crops (Varalli et al. 2015; Tafuri et al. 2018), the diet of people living in the Italian peninsula during the BA seems to have primarily consisted of C3 plants to be complemented by a moderate amount of animal protein (Arena et al. 2020; Varalli et al. 2022). Indeed, the prevailing horticultural regime for Peninsular Italian communities had long depended on wheat (Triticum sp.) and barley (Hordeum vulgare), whose charred remains were often recovered in archaeological surveys (e.g., Tongiorgi 1947; Guidi 1991-92; Pacciarelli 1997; Guidi and Rosini 2019). These grains were widespread in the BA contexts testifying that these plant species were broadly available. Indeed, they could be cultivated, even accounting for farming strategies implemented in productive economies to sustain their harvesting in geographical areas characterized by moist winters and meager summer precipitation.
Remarkably, other than outlining the direct trophic relationship, the stable isotope analysis could also be meaningful for disentangling these horticulture practices (Ferrio et al. 2005; Bogaard et al. 2007, 2013; Flohr et al. 2011; Fraser et al. 2011; Kanstrup et al. 2014; Szpak et al. 2014; Treasure et al. 2016; Gröcke et al. 2021). The analysis of the plant remains has often supported the archaeological identification of horticultural practices, which were critical for developing farming strategies. Specifically, fertilization and irrigation practices, recognizable with changes in carbon and nitrogen stable isotopes ratios, are two of the most significant technological improvements for boosting soil exploitation, supporting both sedentary lifestyles and the market economy (Wallace et al. 2015; Styring et al. 2016; Gron et al. 2017, 2020; Knipper et al. 2020; Mnich et al. 2020; Varalli et al. 2021).
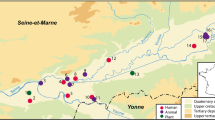
In that perspective, the stable isotope analysis of the faunal remains, which is generally carried out to establish the local isotopic variability, can also reveal, in turn, breeding strategies and the use of specific agricultural practices (Goude et al. 2016). Accordingly, this work aims to contribute to reconstructing of the economic identity of people buried in the Pastena cave (Frosinone, Latium) during the first half of the BA. We put the results in the context of recent genomic and isotopic studies, which provided suggestions for the population dynamics (Saupe et al. 2021; Romboni et al. 2022) and technological improvements (Varalli et al. 2015) running in Central Italian Early and Middle BA (Fig. 1A).

A Pastena Cave in the Italian peninsula. B East view of Grotticella W2 with its upper terrace.
Archaeological context
The Pastena cave (41°29′48.77″N, 13°29′21.91″E) is situated approximately 196 meters a.s.l. in a karst valley. The entrance presents a majestic chamber, which features a seasonal stream of the Rio Mastro Creek. The first excavations were performed in the first half of the twentieth century (Guareschi and Morandini 1943), and subsequently, surveys and excavations were carried out in several areas of the cave (Biddittu 1987; Biddittu et al. 2007; Angle et al. 2010), discovering archaeological findings ranging between the Late Neolithic and the Middle Bronze Age.
The investigation focusing on Grotticella W2 (GW2, Fig. 1B) started in 2006 and was carried out between 2012 and 2018 (Angle et al. 2014; Silvestri et al. 2019; Rolfo et al. 2021). This area is a small room opening in the west sector of the cave’s entrance hall, and it is the only area not affected by post-depositional events. The archaeological deposit has been thoroughly investigated. The radiocarbon dating on seeds and human remains (see Supplemental Material) suggested that the deposit was related to the Middle Bronze Age (MBA 1B). However, the potsherds suggest a chronology between the end of the Early Bronze Age (EBA) and the Middle Bronze Age (MBA) (Rolfo et al. 2021).
Six main layers, including multiple stratigraphic units and archaeological structures, such as hearths, reddened areas, stone paving, and pits, as well as several archaeological finds, including pottery and personal ornaments, have been identified in GW2, suggesting its intensive use related to ritual and funerary practices (Silvestri et al. 2019; Rolfo et al. 2021).
The artifacts were found intermingled along with seeds and bones. Thousands of burnt carpological remains were recovered: botanical visual analyses revealed that they mostly pertained to Vicia faba, while cereals (Triticum monococcum/dicoccum, Hordeum vulgare, Triticum aestivum) account for the residual fraction.
A few faunal remains were found in GW2 (about 100 finds): they are primarily domesticated mammals, especially sheep/goats and pigs, while wild species were rare (Silvestri et al. 2019).
Humans are represented by a minimum number of individuals (MNI) of 4, belonging to different age classes, including osteologically immature ones. The human bones primarily consist of distal limb segments, except for a mandible, some teeth, and a few long bones, which helped estimate age at death and MNI (Rolfo et al. 2021).
Materials and methods
We performed stable isotope analyses on four human bones, eight faunal remains belonging to domesticated animals (Ovis aries vel Capra hircus, Sus domesticus, and Canis familiaris), and forty burnt seeds (Vicia faba, Triticum monococcum/dicoccum, Triticum aestivum, and Hordeum vulgare) (Silvestri et al. 2019; Rolfo et al. 2021).
Carbon and nitrogen stable isotope analysis is commonly used to reconstruct past dietary habits (DeNiro 1985; Ambrose 1990; Unkovich et al. 2001; Marshall et al. 2007).
The relative abundance of 13C and 15N is expressed per mill (‰) through δ notation (respectively, δ13C and δ15N) with respect to international standards—Vienna Pee Dee Belemnite (VDB) for δ13C and atmospheric nitrogen (AIR) for δ15N—according to the relationship: δiE = (iRSA - iRREF) / iRREF, where i is the mass number of the heavier isotope of the E element, RSA is the isotope ratio of the sample, and RREF is the relevant internationally recognized reference material (Mariotti 1983).
The set of carbon and nitrogen δ values reflects the trophic position of an individual between the available resources as they reflect the values of ingested dietary protein (DeNiro 1985; Schoeninger and DeNiro 1984), with somewhat contribution from other macronutrients (Jim et al. 2007; Froehle et al. 2010; Warinner and Tuross 2009). Briefly, the trophic relations are based on a direct correlation between resources and consumers: the latter have higher values than their sources (DeNiro 1985; Schwarcz and Schoeninger 2011). Several diet-to-collagen enrichment factors for both the markers were prompted to interpret the isotopic human diet reconstructions properly via modeling (Ambrose and Norr 1993; Hedges and Van Klinken 2002; Jim et al. 2007; O'Connell et al. 2012; Fernandes et al. 2015; Piliciauskas et al. 2017; O’Connell 2017; Halffman et al. 2020; Schulting et al. 2022), even being aware that δ13C and δ15N represent complex proxy influenced by trophic position, metabolic and physiological offsets, and environmental factors. To date, the widely accepted diet-collagen offset for humans is 4.8 ± 0.5‰ for δ13C and +5.5 ± 0.5‰ for δ15N (Schulting et al. 2022).
Collagen was extracted from cortical bones for humans and faunal remains. Samples were prepared at the Centre of Molecular Anthropology for Ancient DNA Studies of the University of Rome “Tor Vergata,” following the Longin’s protocol (Longin 1971), modified by Brown et al. (1988). The cortical bone was collected and cleaned by surface abrasion. The extraction protocol was performed on about 800 mg of bone, and a modern bovine sample was used as a reference. Samples were demineralized in HCl 0.6M, rinsed with bi-distilled water, and finally gelatinized in HCl 0.001 M. An ultrafiltration step through 30 kDa Amicon Ultra-4 Centrifugal Filter Unites with Ultracel membranes (Millipore) was executed to maximize the collagen concentration, and the obtained collagen was lyophilized. Carbon and nitrogen stable isotope ratios from collagen were measured in a single run on a Delta V Advantage isotope ratio mass spectrometer coupled to a Flash 1112 Elemental Analyser via a Conflow III interface (Thermo Scientific, Milan, Italy) at Dipartimento di Scienze e Tecnologie Ambientali, Biologiche e Farmaceutiche – Università degli Studi della Campania Luigi Vanvitelli and on a Delta Plus XP isotope ratio mass spectrometer coupled with a Flash 1112 Elemental Analyser via a Conflow IV interface (Thermo Scientific, Milan, Italy) at Food Quality and Nutrition Department, Traceability Unit – Fondazione Edmund Mach. Based on replicate analyses of PDB standard for δ13C and AIR for δ15N, the measurement errors were inferred to be less than ±0.2‰ for δ13C and ± 0.3‰ for δ15N in both laboratories. Accordingly, we compared the batches, and no deviances were detected rounding the raw data to the first decimal digit. The samples were also compared against established criteria to ascertain the percentages of carbon and nitrogen, atomic C/N ratios, and collagen yields (DeNiro 1985; Ambrose 1990; van Klinken 1999). To assess the preservation state of the extracted collagen, we considered carbon and nitrogen contents between 15–51% and 5–18%, respectively, and C/N ratios within the range of 2.9 to 3.6 (DeNiro 1985; Ambrose 1990).
An established protocol for stable isotope analysis is not available for seeds. Different methods have been applied using different acid-base-acid pre-treatment protocols (Bogaard et al. 2013; Fraser et al. 2013b), but even crushing and powdering raw seeds (Heaton et al. 2009; Gron et al. 2017, 2020), which indeed were the most applied approach in recent attempts. Considering the results obtained by Lightfoot and Stevens (2012), in which the statistical comparisons showed a negligible difference between pre-treated and untreated samples in either δ13C or δ15N, we did not apply chemical treatment and crashed the seeds singularly in a mortar. Due to the differing total amounts of carbon and nitrogen in the seeds, each grain was weighed and analysed twice (Lightfoot and Stevens 2012; Mueller-Bieniek et al. 2019), and we retained all the samples returning positive yields (Vaiglova et al. 2014) as no consensual criteria exist to assess the preservation of charred seeds and chaff.
Faunal and human data have been evaluated through the linear mixing model proposed by Fraser et al. (2013a). As described by the authors, this model uses the midpoint and the offsets between consecutive trophic levels—an average of 4‰ for δ15N—to identify the effect of predators on their prey. Accordingly, we implemented different models based on plants and faunal values determined in the present paper to calculate the theoretical endpoints for terrestrial resource consumption to evaluate the intake of domesticated crops in faunal remains and the animal protein consumption in the human diet.
Furthermore, we used a Bayesian mixed model through FRUITS v3.1 (Fernandes et al. 2014) to dissect the human diet. The software provides probabilistic quantification of dietary inputs, incorporating food macronutrients, and elemental and isotopic composition in its calculation. The average isotopic values of humans were used as consumer data, and all the estimated offset values were input into FRUITS with uncertainties of 0.5‰ for carbon and 1‰ for nitrogen (Fernandes et al. 2015; Fernandes 2016). Four resources (cereals, broad beans, faunal products, and freshwater fish) have been identified as putative food groups in the model. We used the data from local resources for the first three categories, while freshwater fish values are based on published values (Varalli et al. 2021). The carbon and nitrogen offset/weight ratios are based on Hedges and Reynard (2007), Fernandes et al. (2015), and Styring et al. (2017). We calculated the estimation of source values and the amount of energy and proteins in each food group following Fernandes et al. (2015), Knipper et al. (2020), and Varalli et al. (2021) (see Supplementary materials for details). Furthermore, we subtracted 0.31‰ from the determined seed values to account for the charring effect (Nitsch et al. 2015).
Seeds watering status has been established through carbon values. In order to consider CO2 fluctuations in Holocene, Δ13C values have been used to calculate water availability, comparing archaeological plant data over time (http://web.udl.es/usuaris/x3845331/AIRCO2_LOESS.xls, Ferrio et al. 2005). Δ13C values reflect plants’ water status during their life cycle, which was influenced both by natural precipitation and human management (Wallace et al. 2015). Wallace et al. (2015) provide the largest collections of Δ13C values for the archaeological crop to be used as patterns for the watering status of ancient plants. Following Ferrio et al. (2005), we calculated the plant Δ13C from δ13Cair and plant carbon isotope composition as described by Farquhar et al. (1982) (See Suppl. Mat.).
The estimation of manuring rates was addressed through the comparison with methodological approaches (e.g., Bogaard et al. 2007; Styring et al. 2016; Treasure et al. 2016) and values obtained from archaeological contexts (e.g., Kanstrup et al. 2014; Bogaard et al. 2013), showing that cereals not subjected to fertilization have a baseline δ15N around 2.5‰. Conversely, manured grains (about 35 t/ha compost applied annually) present nitrogen values equal to or greater than 6‰. Intermediate δ15N values between 2.5 and 6‰ can be related to different scenarios (Treasure et al. 2016). Differently, legumes naturally present δ15N values of 0‰, and only an intensive fertilization with animal cages (>70 t/ha) can increase nitrogen values to reach +3‰ (Treasure et al. 2016).
Results
Carbon and nitrogen isotope values of seeds, animals, and humans are listed in Table 1 and shown in Fig. 2. Isotope data were successfully obtained for all the materials. However, two samples of Sus domesticus were excluded due to their C/N ratios outside the range 2.9–3.6 (DeNiro 1985).

Isotope data dispersion for seeds, faunal remains, and humans from the Pastena cave.
A sample of 40 seeds, consisting of broad beans, emmer, barley, and wheat, was also characterized. The isotopic ratios of the C3 plants range from 0.1 to 5.2‰ for δ15N (median= 1.7‰; IQR= 1.60), and −22.1‰ to −27.4‰ for δ13C (median= −24.5‰; IQR = 1.80).
For Δ13C, calculated according to Ferrio et al. (2005), values varied between 17.5 and 21.6‰ for legumes and between 16.1 and 19.3‰ for cereals, indicating that these crops are grown under good hydric conditions. The considerable variation in legumes and grains suggests that they were cultivated on soils receiving variable water inputs and/or varying water retention capacity. Seeds values have been tested through the Kolmogorov-Smirnov test: cereals do not present significant differences (p>0.05), possibly due to the small sample size. Legumes and cereals appear significantly different in both carbon (p=5.65E-07) and nitrogen values (p=2.80E-04). Considering the lack of outlier in these two groups and consistent with archaeological information, we consider the values representative of local produce. The isotopic markers make it reliable that agricultural practices, such as manuring or watering, were present, even though they do not appear to have been as extensive as we expected, considering the vast quantities of seeds in the deposit.
The domesticated animals show δ13C values ranging between −20.8‰ and −18.2‰ (IQR = 1.60), and 3.9‰ and 5.6‰ (IQR = 0.65) for δ15N. Considering the small offset between herbivores and plant remains, especially cereals, the obtained values seem to suggest that herbivores were left to graze.
We tried to confirm the abovementioned hypothesis by evaluating the fraction of plants from the total intake and realizing two models based on the δ15N. The first one (Fig. 3A) is based on the mean value of Vicia faba, which is the starting point to estimate the intake of these crops in herbivores’ diet, as they should rely upon a +4‰ step. The returned consumption of legumes proposed by that model is very high (about 80%). The second model (Fig. 3B) considers the mean value of cereals: the result suggests a low intake of these plants (about 40%).

Fraser’s model (Fraser et al. 2013a). Modeled scenario estimating the plants protein fraction (percentage) from the total intake of faunal dietary choices. B Cereals δ15N values determine the starting point for the esteem of the % of plants protein in faunal remains. A Broad beans δ15N values determine the starting point for the estimation of the % of plants protein in the faunal remains.
We recognize that the reconstruction obtained with Fraser’s model should be interpreted cautiously, considering that it reflects only the protein fraction and is based on an offset value rather than a range. Thus, we can reasonably assume that the plant remains found in the cave might not be the appropriate baseline to assess animal diet, especially considering the cereals, even though legumes could also be used to complement the free animal feeding. Four human individuals—an adult, one skeletally immature, and two children, as reported in Rolfo et al. (2021)—have been analysed for diet reconstruction. Their values ranged between −19.9 and −20.7‰ (IQR = 0.39) for δ13C and between 7.5 and 10.7‰ (IQR=1.65) for δ15N. Adult and young individuals have similar values, suggesting no differences in the diet despite the different ages, except for the infant of about 6 months, whose values reflect the well-known breastfeeding effects (Fuller et al. 2006; Tsutaya and Yoneda 2015) and were removed from the quantitative diet reconstruction accordingly.
The δ13C and δ15N values from bone collagen of humans suggest that their dietary protein was primarily terrestrial in origin, as the δ13C values are all around −20‰ (DeNiro 1985; Schoeninger and Moore 1992). Furthermore, the diet-collagen offset for the nitrogen isotope values reflects less reliance on the fauna protein than on cereals and legumes. The consumption of freshwater fish is not excluded due to the distribution of the δ13C and δ15N data, even though the δ15N values seem to be extremely low to account for the exploitation of the freshwater ecosystem, which is roughly characterized by longer food chains and subsequent higher δ15N values (Schoeninger et al. 1983).
Fraser’s model was also applied to dissect the above observations further and quantify the fraction of animal protein exploitation (Fig. 4). Plants and herbivores values from Pastena determine the starting point for the estimation of the percentage of animal protein in the human diet, which seems moderately high (about 70%). Still, it is essential to consider that the model is an approximation and could boost the estimation as it considers a diet exclusively (100%) based on animal protein consumption, disregarding the possible plant exploitation, even though the model prompts some initial considerations on food practices.

Fraser’s model (Fraser et al. 2013a). Modeled scenario estimating the protein fraction (percentage) from the total intake in the human diet. Herbivores δ15N determine the starting point for the estimation of the percentage of animal protein.
Discussion
The GW2 sector of the Pastena cave is characterized by several archaeological finds, including hundreds of thousands of burnt carpological remains, mainly broad beans (Vicia faba). Due to the high quantity, it is possible to assume that seeds resulted from abundant production, probably due to human care and management. The amount of remains, involving remarkable energy expenditure by the human community, seems to be beyond the sole dietary consumption and could be related reliably to specific rituals and, at least partially, to foraging purposes.
As previously reported (Rolfo et al. 2021), Pastena GW2 was a funerary area initially. Afterward, it appears to become devoted solely to non-burial ritual practices, as corroborated by archaeological features such as stone-paving, hearths, pits, and evidence of meals of most likely ritual nature. From that perspective, the relationship between broad beans and the after-life world seems worth mentioning. Indeed, multiple ancient cultures, such as Egyptians, Greeks, and even Romans, attested such a close link between broad beans and the soul of the dead, with the latter offering beans to keep the evil spirits away (De Cleene and Lejeune 2004; Beer 2010; Silvestri 2016).
However, the seeds should have also been part of the dietary habits of people buried inside the cave. The stable isotope data from the individuals suggest a diet based principally on the consumption of C3 sources and the exploitation of local resources. Accordingly, we quantitatively reconstructed the human diet through a Bayesian mixed model (Fernandes et al. 2014). We implemented two different models, first using the raw data and then implementing the following prioritizations: legumes > cereals; legumes > animal products; legumes > freshwater fish; cereals > animal products; and cereals > freshwater fish.
In the first model (Fig. 5A, Suppl. Mat.), the contribution of animal products is high (median 50.7%), followed by cereals (36.3%), legumes (10.9%), and freshwater fish (2.0%). The model is consistent with the hypothesis formulated through the qualitative results obtained from stable isotope analysis and Fraser’s model and highlights the high consumption of terrestrial resources and the rare exploitation of freshwater fish in the community. Furthermore, considering the high number of seeds in the cave, we decided to prioritize the broad beans exploitation, even though we are aware that they could be selectively put in the cave by human selection strategy.

Estimated contributions of cereals, legumes, animal products, and freshwater fish to the diets of adult individuals from the Pastena cave using the Bayesian model FRUITS: A estimations without priors, B estimations with prioritizations (legumes > cereals, cereals and legumes > fauna, freshwater fish).
The prioritization of legumes and cereals led to significant changes in the identification of plants as the primary edible source (Fig. 5B, Suppl. Mat.): the intake of legumes is 37.6%, followed by cereals (32.1%), terrestrial animal products (26.25%), and freshwater fish (4.0%).
The dietary landscape outlined for Pastena is worth being reported considering the diet reconstructions for coeval Italian sites. Only a few Central Italian sites reveal the consumption of C4 plants in the Middle Bronze Age (Varalli et al. 2015; Romboni et al. 2022). Genomics (Saupe et al. 2021) showed how changes occurred in central Italy during the MBA, probably related to the arrival of populations from northern areas throughout the Italian peninsula. The burial area we are considering was radiometrically dated between the end of the Early Bronze Age and the beginning of the Middle Bronze Age (Rolfo et al. 2021). Considering the geographical proximity between the Pastena cave, the Misa cave, and the La Sassa cave—where C4 plants consumption was identified—as well as the tight chronological frames, it is worth noting that Pastena does not seem to be involved in those complex dynamics.
Multiple factors could be responsible for that odd scenario for Pastena.
First, the slightly different chronologies of all these contexts should be considered. Indeed, the Misa cave is dated—chrono-typologically—to the MBA 1-2 (Cocchi Genick 1995), while La Sassa dates back from the Copper Age up to the MBA 3 (Alessandri et al. 2020, 2021). Remarkably, the individual from La Sassa supporting the consumption of C4 plants (Romboni et al. 2022) dates back to the MBA 2, which outdates the upper chronological boundary for Pastena. Additionally, the Misa cave is located in northern Latium; conversely, Pastena and La Sassa caves are both located in southern Latium and close to one another. It is worth considering that the putative demic flow responsible for introducing new crops arriving in northern Latium in the first phases of MBA might spread to southern Latium in MBA 2 (Varalli et al. 2015; Romboni et al. 2022), excluding people buried in the Pastena cave from these complex dynamics.
Similarly to Pastena, people buried in Sepolcreto di Felcetone (northern Latium), Collepardo Cave (southern Latium), and Scoglietto cave (southern Tuscany), dated to MBA3 (Cocchi Genick 1995; Skeates et al. 2021) and EBA (Cocchi and Ceccanti 1978), respectively, did not show C4 consumption (Varalli et al. 2015; Skeates et al. 2021), suggesting a not continuous southward cline for the spread of the newly introduced plants as edible resources.
If people buried in the Pastena cave had a diet based on C3 plants, it seems reasonable to consider that they could gather these resources by tuning of the horticultural practices. So far, the agricultural practices have been attested in Europe since the Neolithic (e.g., Buurman 1988; Fokkens 1982), and genetic studies have demonstrated how human genetic adaptation to an agricultural diet occurred during the Bronze Age (Mathieson and Mathieson 2018).
Archaeological and bioarchaeological data suggest an increase in cereal cultivation in Italy, especially from the Middle Bronze Age, when intensive agriculture spread (Salvadei and Santandrea 2003; Tafuri et al. 2009; Arena et al. 2020).
Our data suggest that artificial management was applied to the growing plants. Specifically, the cereals values indicate that they grow under similar conditions. Cereals appear to be moderate-to-poorly watered (Wallace et al. 2015; Knipper et al. 2020; Varalli et al. 2021; Fig. 6A). Conversely, legumes (Fig. 6B) were well-watered (Wallace et al. 2015). However, it should be borne in mind that water availability differences might also be related to seasonal rainfall and cultivation.

Estimations of watering status based on Wallace et al. (2015): A cereals, B broad beans
Published data (Bogaard et al. 2007, 2013; Fraser et al. 2011; Kanstrup et al. 2014) demonstrated that fertilization increases nitrogen values in cereals and legumes depending on the intensity and duration of manuring. Cereals from the Pastena cave do not appear to be extensively manured (Bogaard et al. 2013), contrary to the legumes. Fraser et al. (2011) show that legumes have a low nitrogen isotopic signature (about 0‰), and only intensive fertilization can alter these values. Vicia faba δ15N values from the Pastena cave range from 0.1 to 3.2‰; thus, it is possible to hypothesize that some broad beans have been subject to fertilization. The massive presence of legumes on the site and the fact that none of them suffered from water stress led to considering these crops’ volunteer management.
Finally, it is worth mentioning that the seeds analysed were charred, which could impact the managing identification’s reliability. However, previous studies (DeNiro and Hastorf 1985; Bogaard et al. 2007; Kanstrup et al. 2012) demonstrated that charring until 220 °C does not significantly alter the pristine isotope values, which otherwise would be disrupted. We have not yet identified the temperature to which the grains were exposed; however, the influence of charring is highly questionable as some scholars claimed that the charring is not a significant modifier for δ13C and δ15N (Kanstrup 2012; Bogaard et al. 2013; Fraser et al. 2013b; Styring et al. 2013). Specifically, the carbon values were demonstrated only minimally or not affected (Fraser et al. 2013b), while δ15N may rise by about 1‰ (Fraser et al. 2013b). Additionally, recent research showed that the offset might be tiny and accountable by subtracting 0.31‰ from the determined values (Nitsch et al. 2015), not impacting our considerations based on the model development (Suppl. Mat.).
Conclusion
The GW2 sector of Pastena cave (Latium) allowed for the overall reconstruction of the subsistence strategies for a Central Italian Early-Middle Bronze Age community. Carbon and nitrogen stable isotope data confirm that people who exploited the cave were devoted to an agricultural economy based on terrestrial resources, mainly including plants and animal proteins, consistently with what was hypothesized through the archaeological evaluation.
Despite the limited sample size for humans, this paper presents, to the best of our knowledge, the first stable isotope analysis carried out on carpological remains dated to the Italian Bronze Age, contributing to filling a gap in the biomolecular research of the Italian peninsular prehistoric communities. These data allow for hypothesizing a human role in crop management as broad beans were grown under better watering conditions than cereals. Similarly, differential fertilization can be hypothesized for these plants.
Most of the faunal remains in GW2 belong to herbivorous and omnivorous animals, whose diet suggests that animals were probably fed, at least occasionally, with agricultural output.
Thus, despite the limited bio-archaeological materials in the cave, the synergic evaluation of bio-archaeological data leads to reconstructing the trophic net—from plants to humans—aiming to obtain a reliable reconstruction of the subsistence strategy for one of the communities living in an area characterized by complex demographic dynamics in Central Italian Bronze Age.
References
Alessandri L (2013) Latium Vetus in the Bronze Age and Early Iron Age /Il Latium Vetus nell’età del Bronzo e nella prima età del Ferro. BAR International Series, Oxford, p 2565
Alessandri L (2019) The early and Middle Bronze Age (1/2) in South and central Tyrrhenian Italy and their connections with the Avellino eruption: An overview. Quat Int 499:161–185. https://doi.org/10.1016/j.quaint.2018.08.002
Alessandri L, Baiocchi V, Del Pizzo S, Di Ciaccio F, Onori M, Rolfo MF, Troisi S (2020) The fusion of external and internal 3D photogrammetric models as a tool to investigate the ancient human/cave interaction. ISPRS Arch 43:1443–1450. https://doi.org/10.5194/isprs-archives-XLIII-B2-2020-1443-2020
Alessandri L, Cardello GL, Attema PAJ, Baiocchi V, De Angelis F, Del Pizzo S, Di Ciaccio F, Fiorillo A, Gatta M, Monti F, Onori M, Rolfo MF, Romboni M, Sottili G, Troisi S (2021) Reconstructing the Late Pleistocene – Anthropocene interaction between the neotectonic and archaeological landscape evolution in the Apennines (La Sassa cave, Italy). Quat Sci Rev 265:107067. https://doi.org/10.1016/j.quascirev.2021.107067
Ambrose SH (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. J Archaeol Sci 17:431–451. https://doi.org/10.1016/0305-4403(90)90007-R
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein of bone collagen and carbonate. In: Lambert JB, Grupe G (eds) Prehistoric Human Bone. Springer, Berlin, pp 1–37
Angle M, Bruni N, Cerqua M, Riva A, Cavazzuti C, Celletti P (2010) Nuovi scavi nella grotta di Pastena (Frosinone). In: Ghini G (ed) Atti del convegno sesto incontro di studi sul Lazio e la Sabina. Quasar, pp 369–380
Angle M, Rolfo MF, Fusco I, Silvestri L (2014) New investigations at the Cave of Pastena (Frosinone): Report 2012. In: Ghini G, Mari Z (eds) Lazio e Sabina, Scoperte Scavi e Ricerche, Atti del Decimo Incontro di Studi sul Lazio e la Sabina. Quasar, pp 205–211
Arena F, Gualdi-Russo E, Olsen J, Philippsen B, Mannino MA (2020) New data on agro-pastoral diets in southern Italy from the Neolithic to the Bronze Age. Archaeol Anthropol Sci 12:245. https://doi.org/10.1007/s12520-020-01209-9
Barfield LH (2007) Excavations in the Riparo Valtenesi, Manerba 1976-1994. Istituto Italiano di Preistoria e Protostoria, Florence
Beer M (2010) Taste or Taboo: dietary choices in Antiquity. Prospect books, Totnes
Bettelli M (2006) L’età del Bronzo e la prima età del Ferro in Europa. Storia d'Europa e del Mediterraneo – Dalla Preistoria alla Storia 1:365–411
Biddittu I (1987) Insediamento dell'età del bronzo nelle Grotte di Pastena. Latium 4:237–239
Biddittu I, Bruni N, Carancini GL, Cerqua M, Riva A (2007) La frequentazione delle grotte di Pastena in età preistorica e protostorica. In: Cocchi GD (ed) Atti della XL Riunione Scientifica dell’Istituto Italiano Preistoria e Protostoria: Strategie di insediamento fra Lazio e Campania in età preistorica e protostorica, II edn. L’Erma di Bretschneider, Firenze, pp 683–693
Bietti Sestieri AM (2015) L’Italia nell’età del Bronzo e del Ferro: dalle palafitte a Romolo (2200-700 a.C.). Carocci Editore, Roma
Blake E (2014) Social networks and regional identity in Bronze Age Italy. Cambridge University Press, Cambridge
Bogaard A, Heaton THE, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J Archeol Sci 34:335–343. https://doi.org/10.1016/j.jas.2006.04.009
Bogaard A, Fraser R, Heaton THE, Wallace M, Vaiglova P, Charles M, Jones G, Evershed RP, Styring AK, Andersen NH, Arbogast RM, Bartosiewicz L, Gardeisen A, Kanstrup M, Maier U, Marinova E, Ninov L, Schäfer M, Stephan E (2013) Crop manuring and intensive land management by Europe’s first farmers. Proc Natl Acad Sci 110:12589–12594. https://doi.org/10.1073/pnas.1305918110
Brown TA, Nelson DE, Vogel JS, Southon JR (1988) Improved collagen extraction by modified Longin method. Radiocarbon 30:171–177. https://doi.org/10.1017/S0033822200044118
Buurman J (1988) Economy and environment in Bronze Age West-Friesland, Nord Holland. In: Murphy P, French C (eds) The exploitation of wetlands. British Archeological reports, Oxford, pp 268–292
Cattani M, Marchesini M (2010) Economia e gestione del territorio. In: Cattani M, Marchesini M, Marchesini S (eds) Paesaggio ed economia nell’Età del Bronzo: la pianura bolognese tra Samoggia e Panaro. Museo Archeologico Ambientale, Bologna, pp 233–243
Cavanna C (2007) La grotta dello Scoglietto. Indagini 2005-2006. In: Cavanna C (ed) La preistoria nelle grotte del parco naturale della Maremma, suppl. n. 22. Grosseto, pp. 67-98
Cavazzuti C, Skeates R, Millard AR, Nowell G, Peterkin J, Bernabò Brea M, Cardarelli A, Salzani L (2019) Flows of people in villages and large centres in Bronze Age Italy through strontium and oxygen isotopes. PLoS One 14(1):e0209693. https://doi.org/10.1371/journal.pone.0209693
Cavazzuti C, Hajdu T, Lugli F, Sperduti A, Vicze M, Horváth A, Major I, Molnár M, Palcsu L, Kiss V (2021) Human mobility in a Bronze Age Vatya “urnfield” and the life history of a high-status woman. PLoS One 16(7):e0254360. https://doi.org/10.1371/journal.pone.0254360
Cazzella A (2009) La formazione di centri specializzati dell’Italia sud-orientale durante l’età del Bronzo. Scienze dell’Antichità: Storia, Archeologia, Antropologia 15:293–310
Cocchi D, Ceccanti M (1978) La grotta dello Scoglietto (Grosseto) – Studio dei materiali conservati al Museo Fiorentino di Preistoria. Rivista di Scienze preistoriche 33:187–214
Cocchi Genick D (1995) Aspetti culturali della media età del Bronzo nell’Italia centro-meridionale. Octavo, Firenze
De Angelis F, Scorrano G, Martinez-Labarga C, Giustini F, Brilli M, Pacciani E, Silvestrini M, Calattini M, Volante N, Martini F, Sarti L, Rickards O (2019) Eneolithic subsistence economy in Central Italy: first dietary reconstructions through stable isotopes. Archaeol Anthropol Sci 11:4171–4186
De Angelis F, Varano S, Battistini A, Di Giannantonio S, Ricci P, Lubritto C, Facchin G, Brancazi L, Santangeli-Valenzani R, Catalano P, Gazzaniga V, Rickards O, Martinez-Labarga C (2020) Food at the heart of the Empire: dietary reconstruction for Imperial Rome inhabitants. Archeol Anthropol Sci 12:244. https://doi.org/10.1007/s12520-020-01194-z
De Cleene M, Lejeune MC (2004) Compendium of symbolic and ritual plants in Europe. Mens & Cultuur Uitgevers, Ghent
De Grossi MJ (1995) Economie di allevamento in Italia centrale dalla media età del Bronzo alla fine dell’età del Ferro. In: Christie N (ed) Settlement and economy in Italy 1500 BC to AD 1500: Papers of the Fifth Conference of Italian Archaeology, 41st edn. Oxbow Monograph, Oxford, pp 167–177
DeNiro MJ (1985) Post-mortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature 317:806–809. https://doi.org/10.1038/317806a0
DeNiro MJ, Hastorf CA (1985) Alteration of 15N/14N and 13C/12C ratios of plant matter during the initial stages of diagenesis: studies utilizing archaeological specimens from Peru. Geochim Cosmochim Acta 45:341–351. https://doi.org/10.1016/0016-7037(85)90194-2
Farquhar GD, O’Leaey MH, Berry JA (1982) On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust J Plant Physiol 9:121–137. https://doi.org/10.1071/PP9820121
Fernandes R (2016) A simple (R) model to predict the source of dietary carbon in individual consumers. Archaeometry 58:500–512. https://doi.org/10.1111/arcm.12193
Fernandes R, Millard AR, Brabec M, Nadeau MJ, Grootes P (2014) Food reconstruction using isotopic transferred signals (FRUITS): a Bayesian model for diet reconstruction. PLoS One 9(2):e87436. https://doi.org/10.1371/journal.pone.0087436
Fernandes R, Grootes P, Nadeau M-J, Nehlich O (2015) Quantitative diet reconstruction of a Neolithic population using a Bayesian mixing model (FRUITS): the case study of Ostorf (Germany). Am J Phys Anthropol 158(2):325–340. https://doi.org/10.1002/ajpa.22788
Ferrio JP, Araus JL, Buxò R, Voltas J, Bort J (2005) Water management practices and climate in ancient agriculture: inferences from the stable isotope composition of archaeobotanical remains. Veg Hist Archaeobotany 14:510–517. https://doi.org/10.1007/s00334-005-0062-2
Flohr P, Müldner G, Jenkins E (2011) Carbon stable isotope analysis of cereal remains as a way to reconstruct water availability: preliminary results. Water Hist 3:121–144. https://doi.org/10.1007/s12685-011-0036-5
Fokkens H (1982) Late Neolithic occupation near Bornwird. Palaeohistoria 24:91–113
Fokkens H, Harding A (2013) The Oxford handbook of the European Bronze Age. Oxford University press, Oxford, pp 17–369
Fontanals-Coll M, Díaz-Zorita Bonilla M, Subirà M (2015) A paleodietary study of stable isotope analysis from a high-status burial in the Copper Age: the Montelirio megalithic structure at Valencina de la Concepción-Castilleja de Guzmán, Spain. Int J Osteoarchaeol 26(3):447–459. https://doi.org/10.1002/oa.2435
Fraser RA, Bogaard A, Heaton T, Charles M, Jones G, Christensen BT, Halstead P, Merbach I, Poulton PR, Sparkes D, Styring AK (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38:2790–2804. https://doi.org/10.1016/j.jas.2011.06.024
Fraser RA, Bogaard A, Schäfer M, Arbogast R, Heaton THE (2013a) Integrating botanical faunal and human stable carbon and nitrogen isotope values to reconstruct land use and palaeodiet at LBK Vaihingen an der Enz Baden-Wurttemberg. World Archaeol 45:492–517. https://doi.org/10.1080/00438243.2013.820649
Fraser RA, Bogaard A, Charles M, Styring AK, Wallace M, Jones G, Ditchfield P, Heaton THE (2013b) Assessing natural variation and the effects of charring, burial and pre-treatment on the stable carbon and nitrogen isotope values of archaeobotanical cereals and pulses. J Archaeol Sci 40:4754–4766. https://doi.org/10.1016/j.jas.2013.01.032
Froehle AW, Kellner CM, Schoeninger MJ (2010) FOCUS: effect of diet and protein source on carbon stable isotope ratios in collagen: follow up to Warinner and Tuross. J Archaeol Sci 37:2662–2670. https://doi.org/10.1016/j.jas.2010.06.003
Fuller BT, Fuller JL, Harris DA, Hedges REM (2006) Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios. Am J Phys Anthropol 129(2):279–293. https://doi.org/10.1002/ajpa.20249
Goude G, Rey L, Toulemonde F, Cervel M, Rottier S (2016) Dietary changes and millet consumption in northern France at the end of prehistory: evidence from archaeobotanical and stable isotope data. Environ Archaeol 22(3):268–282. https://doi.org/10.1080/14614103.2016.1215799
Gröcke DR, Treasure ER, Lester JJ, Gron KJ, Church MJ (2021) Effects of marine biofertilization on Celtic bean carbon, nitrogen and sulphur isotopes: implications for reconstructing past diet and farming practices. RCM 35:e8985. https://doi.org/10.1002/rcm.8985
Gron KJ, Gröcke DR, Larsson M, Sørensen L, Larsson L, Rowley-Conwy P, Church MJ (2017) Nitrogen isotope evidence for manuring of early Neolithic Funnel Beaker Culture cereals from Stensborg, Sweden. J Archaeol Sci Rep 14:575–579. https://doi.org/10.1016/j.jasrep.2017.06.042
Gron KJ, Larsson M, Gröcke DR, Andersen NH, Andreasen MH, Bech J-H, Henriksen PS, Hilton RG, Jessen MD, Møller NA, Nielsen FO, Nielsen PO, Pihl A, Sørensen L, Westphal J, Rowly-Conwy P, Church MJ (2020) Archaeological cereals as an isotope record of long-term soil health and anthropogenic amendment in southern Scandinavia. Quat Sci Rev 253:1–8. https://doi.org/10.1016/j.quascirev.2020.106762
Guareschi C, Morandini G (1943) Ricerche nella Grotta di Pastena, Luppa e Pietrasecca, Lazio. Bollettino della Società Veneziana di Storia Naturale e Museo Civico di Storia Naturale 3:43–62
Guidi A (1991-92) Recenti ritrovamenti in grotta nel Lazio: un riesame critico del problema dell’utilizzazione delle cavità naturali. RdA 10:427–437
Guidi A, Rosini L (2019) Materiali protostorici dalla grotta Vittorio Vecchi (Sezze Romano, LT). BAR, Oxford
Halffman CM, Potter BA, McKinney HJ, Tsutaya T, Finney BP, Kemp BM, Bartelink EJ, Wooller MJ, Buckley M, Clark CT, Johnson JJ, Bingham BL, Lanöe FB, Sattler RA, Reuther JD (2020) Ancient Beringian paleodiets revealed through multiproxy stable isotope analyses. Sci Adv 6(36):eabc1968. https://doi.org/10.1126/sciadv.abc196
Heaton THE, Jones G, Halstead P, Tsipropoulos T (2009) Variation in the 13C/12C ratios of modern wheat grain, and implications for interpreting data from Bronze Age Assiros Toumba, Greece. J Archaeol Sci 36:2224–2233. https://doi.org/10.1016/j.jas.2009.06.007
Hedges R, Reynard L (2007) Nitrogen isotopes and the trophic level of humans in archaeology. J Archaeol Sci 34:1240–1251. https://doi.org/10.1016/j.jas.2006.10.01515
Hedges REM, Van Klinken GJ (2002) “Consider a spherical cow …” — on modeling and diet. In: Ambrose SH, Katzenberg MA (eds) Biogeochemical approaches to paleodietary analysis. Springer, Boston, pp 211–241. https://doi.org/10.1007/0-306-47194-9_11
Jim S, Jones V, Ambrose SH, Evershed RP (2007) Quantifying dietary macronutrient sources of carbon for bone collagen biosynthesis using natural abundance stable carbon isotope analysis. BJN 95(6):1055–1062. https://doi.org/10.1079/BJN20051685
Kanstrup M (2012) When δ15N values reveal manuring practice: empirical evidence from fieldwork, charring experiments and archaeobotanical remains. (PhD Thesis). Århus University, Århus
Kanstrup M, Thomsen IK, Mikkelsen PH, Christensen BT (2012) Impact of charring on cereal grain characteristics: linking prehistoric manuring practice to δ15N signatures in archaeobotanical material. J Archaeol Sci 39(7):2533–2540. https://doi.org/10.1016/j.jas.2012.03.007
Kanstrup M, Holst MK, Jensen PM, Thomsen IK, Christensen BT (2014) Searching for long-term trends in prehistoric manuring practice. δ15N analyses of charred cereal grains from the 4th to the 1st millennium BC. J Archaeol Sci 51:115–125. https://doi.org/10.1016/j.jas.2013.04.018
Katzenberg MA (2008) Stable isotope analysis: a tool for studying past diet, demography, and life history. In: Katzenberg MA, Saunders SR (eds) Biological Anthropology of the human skeleton. Wiley-Liss, New York, pp 413–442
Keegan WF (1989) Stable isotope analysis of prehistoric diet. In: Işcan MY, Kennedy KAR (eds) Reconstruction of life from skeleton. Wiley-Liss, Florida, pp 223–236
Knipper C, Mittnik A, Massy K, Kociumaka C, Kucukkalipci I, Maus M, Wittenborn F, Metz SE, Staskiewicz A, Krause J, Stockhammer P (2017) Female exogamy and gene pool diversification at the transition from the Final Neolithic to the Early Bronze Age in central Europe. PNAS 114(38):10083–10088
Knipper C, Rihuete-Herrada C, Voltas J, Held P, Lull V, Micó R, Risch R, Alt KW (2020) Reconstructing Bronze Age diets and farming strategies at the early Bronze Age sites of La Bastida and Gatas (southeast Iberia) using stable isotope analysis. PLoS One 15(3):e0229398. https://doi.org/10.1371/journal.pone.0229398
Krigbaum J (2003) Neolithic subsistence patterns in northern Borneo reconstructed with stable carbon isotopes of enamel. J Anthropol Archaeol 22:292–304. https://doi.org/10.1016/S0278-4165(03)00041-2
Lightfoot E, Stevens RE (2012) Stable isotope investigations of charred barley (Hordeum vulgare) and wheat (Triticum spelta) grains from Danebury Hillfort: implications for palaeodietary reconstructions. J Archaeol Sci 39:656–662. https://doi.org/10.1016/j.jas.2011.10.026
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature 230:241–242. https://doi.org/10.1038/230241a0
Maini E (2010) L’allevamento e popolamento animale. In: Cattani M, Marchesini M, Marchesini S (eds) Paesaggio ed economia nell’Età del Bronzo: la pianura bolognese tra Samoggia e Panaro. Museo Archeologico Ambientale, Bologna, pp 217–229
Mariotti A (1983) Atmospheric nitrogen is a reliable standard for natural 15N abundance measurements. Nature 303:685–687. https://doi.org/10.1038/303685a0
Mariotti Lippi M, Pisaneschi L, Sarti L, Lari M, Moggi-Cecchi J (2017) Insights into the Copper-Bronze Age diet in Central Italy: plant microremains in dental calculus from Grotta dello Scoglietto (Southern Tuscany, Italy). J Archaeol Sci Rep 15:30–39. https://doi.org/10.1016/j.jasrep.2017.07.005
Marshall JD, Brooks JR, Lajtha K (2007) In stable isotopes in ecology and environmental science. Blackwell Publishers, Boston
Mathieson S, Mathieson I (2018) FADS1 and the timing of Human adaptation to agriculture. Mol Biol Evol 35:2957–2970. https://doi.org/10.1093/molbev/msy180
Miller NF, Spengler RN, Frachetti M (2016) Millet cultivation across Eurasia: origins, spread, and the influence of seasonal climate. The Holocene 26(10):1566–1575. https://doi.org/10.1177/0959683616641742
Minniti C (2012) Ambiente, sussistenza e articolazione sociale nell’Italia centrale tra Bronzo medio e Primo Ferro. BAR British Archaeological Series, Oxford
Mnich B, Mueller-Bieniek A, Nowak M, Wilczyński J, Pospuła S, Szostek K (2020) Terrestrial diet in prehistoric human groups from southern Poland based on human, faunal and botanical stable isotope evidence. JAS Rep 32:102382. https://doi.org/10.1016/j.jasrep.2020.102382
Mueller-Bieniek A, Nowak M, Styring A, Lityńska-Zając M, Moskal-del Hoyo M, Sojka A, Paszko B, Tunia K, Bogaard A (2019) Spatial and temporal patterns in Neolithic and Bronze Age agriculture in Poland based on the stable carbon and nitrogen isotopic composition of cereal grains. J Archaeol Sci Rep 27:101993. https://doi.org/10.1016/j.jasrep.2019.101993
Nava A, Fiorin E, Zupancich A, Carra M, Ottoni C, Di Carlo G, Vozza I, Brugnoletti O, Alhaique F, Grifoni Cremonesi R, Coppa A, Bondioli L, Borić S, Cristiani E (2021) Multipronged dental analyses reveal dietary differences in last foragers and first farmers at Grotta Continenza, central Italy (15,500–7000 BP). Sci Rep 11:4261. https://doi.org/10.1038/s41598-021-82401-2
Nitsch EK, Charles M, Bogaard A (2015) Calculating a statistically robust δ13C and δ15N offset for charred cereal and pulse seeds. Sci Technol Archaeol Res 1:1–8. https://doi.org/10.1179/2054892315Y.0000000001
O’Connell TC (2017) ‘Trophic’ and ‘source’ amino acids in trophic estimation: a likely metabolic explanation. Oecologia 184(2):317–326. https://doi.org/10.1007/s00442-017-3881-9
O'Connell TC, Kneale CJ, Tasevska N, Kuhnle GG (2012) The diet-body offset in human nitrogen isotopic values: a controlled dietary study. Am J Phys Anthropol 149:426–434. https://doi.org/10.1002/ajpa.22140
Pacciarelli M (1997) Acque, Grotte, Dei. 3000 anni di culti preromani in Romagna, Marche, Abruzzo. Musei Civici di Imola, Imola
Peroni R (1996) L’Italia alle soglie della storia. Laterza, Roma
Piliciauskas G, Jankauskas R, Piliciauskiene G, Dupras TL (2017) Reconstructing Subneolithic and Neolithic diets of the inhabitants of the SE Baltic coast (3100–2500 cal BC) using stable isotope analysis. Archaeol Anthropol Sci 9(7):1421–1437. https://doi.org/10.1007/s12520-017-0463-z
Rittatore F (1951a) Scoperte di età eneolitica e del bronzo nella Maremma Tosco-laziale. RSP 5(1-2):3–31
Rittatore F (1951b) Nuove scoperte dell’Età del bronzo lungo la valle del fiume Fiora. RSP 5(2):151–175
Rolfo MF, Achino KF, Fusco I, Salari L, Silvestri L (2013) La Grotta Mora Cavorso a Jenne (Roma): i livelli dell’antica-media età del Bronzo. RSP 63:95–123
Rolfo MF, Achino KF, Cortese F, Gatta M, Salari L, Silvestri L (2021) Cave rituals under the magnifying glass: methodological resources to detect unicity at the Italian Middle Bronze Age site of Grotta di Pastena. IpoTESI di Preistoria 14:1–14. https://doi.org/10.6092/ISSN.1974-7985/14323
Romboni M, Arienzo I, Di Vito MA, Lubritto C, Piochi M, Di Cicco MR, Rickards O, Rolfo MF, Sevink J, De Angelis F, Alessandri L (2022) Isotopic evidence for population dynamics in the Central Italian Copper Age and Bronze Age. BioRxiv Preprint. https://doi.org/10.1101/2021.09.30.462554
Salari L, Tagliacozzo A (2019) Le faune. Considerazioni ambientali, economiche e culturali. In: De Angelis MC (ed) La Grotta dei Cocci di Narni, Scavi 1989-2001. All’Insegna del Giglio, Firenze, pp 73–89
Salvadei L, Santandrea E (2003) Condizioni di vita e stato di salute nel campione neolitico di Masseria Candelaro (FG). Atti della XXXV Riunione Scientifica dell’Istituto Italiano di Preistoria e Protostoria, Firenze 35:829–834
Saupe T, Montinaro F, Scaggion C, Carrara N, Kivisild T, D’Atanasio E, Hui R, Solnik A, Lebrasseur O, Larson G, Alessandri L, Arienzo I, De Angelis F, Rolfo MF, Skeates R, Silvestri L, Beckett J, Talamo S, Dolfini A et al (2021) Ancient genomes reveal structural shifts after the arrival of Steppe-related ancestry in the Italian Peninsula. Curr Biol 31:1–16. https://doi.org/10.1016/j.cub.2021.04.022
Schoeninger MJ, Moore K (1992) Bone stable isotope studies in archaeology. J World Prehist 6:247–296. https://doi.org/10.1007/BF00975551
Schoeninger MJ, DeNiro MJ, Tauber H (1983) Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220:1381–1383. https://doi.org/10.1126/science.6344217
Schulting RJ, MacDonald R, Richards MP (2022) FRUITS of the sea? A cautionary tale regarding Bayesian modelling of palaeodiets using stable isotope data. Quat Int. https://doi.org/10.1016/j.quaint.2022.02.012
Schwarcz HP, Schoeninger MJ (2011) Stable isotopes of carbon and nitrogen as tracers for paleo-diet reconstruction. In: Baskaran M (ed) Handbook of environmental isotope geochemistry, advances in isotope geochemistry, vol I. Springer Berlin, pp 725–742. https://doi.org/10.1007/978-3-642-10637-8_34
Sestini A (1934) Tra Bocca d’Ombrone e i Monti dell’Uccellina. L’Universo 15(9)
Shoeninger M, DeNiro M (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639. https://doi.org/10.1016/0016-7037(84)90091-7
Silvestri L (2016) Caves and human lifeways in Middle Bronze Age Central Italy: a social bioarchaeology approach. Doctoral thesis, Durham University
Silvestri L, Rolfo MF, Achino KF, Gatta M, Angle M (2019) How to detect ritual in Middle Bronze Age Central Italy? A contextual approach at Pastena Cave. In: Buster L, Warmenhol E, Mlekuž D (eds) Between worlds: understanding ritual cave use in later prehistory. Springer, Berlin, pp 87–112. https://doi.org/10.1007/978-3-319-99022-4_6
Skeates R, Beckett J, Mancini D, Cavazzuti C, Silvestri L, Hamilton WD, Sayle KL, Crowder KD, Rolfo M, Angle M (2021) Rethinking collective burial in Mediterranean Caves: Middle Bronze Age Grotta Regina Margherita, Central Italy. J Field Archaeol:1–17. https://doi.org/10.1080/00934690.2021.1917137
Stevens CJ, Murphy C, Roberts R, Lucas L, Silva F, Fuller DQ (2016) Between China and South Asia: a Middle Asian corridor of crop dispersal and agricultural innovation in the Bronze Age. The Holocene 26(10):1541–1555. https://doi.org/10.1177/0959683616650268
Styring AK, Manning H, Fraser RA, Wallace M, Jones G, Charles M, Heaton THE, Bogaard A, Evershed RP (2013) The effect of charring and burial on the biochemical composition of cereal grains: investigating the integrity of archaeological plant material. J Archaeol Sci 40:4767–4779. https://doi.org/10.1016/j.jas.2013.03.024
Styring AK, Ater M, Hmimsa Y, Fraser R, Miller H, Neef R, Pearson JA, Bogaard A (2016) Disentangling the effect of farming practice from aridity on crop stable isotope values: a present-day model from Morocco and its application to early farming sites in the eastern Mediterranean. Anthropol Rev 3(1):2–22. https://doi.org/10.1177/2053019616630762
Styring AK, Charles M, Fantone F, Hald MM, McMahon A, Meadow RH, Nicholls GK, Patel AK, Pitre MC, Smith A, Sołtysiak SG, Weber JA, Weiss H, Bogaard A (2017) Isotope evidence for agricultural extensification reveals how the world’s first cities were fed. Nat Plants 3:17076. https://doi.org/10.1038/nplants.2017.76
Szpak P, Longstaffe FJ, Millaire J-F, White CD (2014) Large variation in nitrogen isotopic composition of a fertilized legume. J Archaeol Sci 45:72–79. https://doi.org/10.1016/j.jas.2014.02.007
Tafuri MA, Craig OE, Canci A (2009) Stable isotope evidence for the consumption of millet and other plants in Bronze Age Italy. Am J Phys Anthropol 139:146–153. https://doi.org/10.1002/ajpa.20955
Tafuri MA, Rottoli M, Cupitò M, Pulcini ML, Tasca G, Carrara N, Bonfanti F, Salzani L, Canci A (2018) Estimating C4 plant consumption in Bronze Age Northeastern Italy through stable carbon and nitrogen isotopes in bone collagen. Int J Osteoarchaeol 28:131–142. https://doi.org/10.1002/oa.2639
Tongiorgi E (1947) Grano, miglio e fave in un focolare rituale del Bronzo a Grotta Misa (bassa valle della Fiora). Nuovo Giorn Bot Ital 54:804–806
Treasure ER, Church MJ, Gröcke DR (2016) The influence of manuring on stable isotopes (δ13C and δ15N) in Celtic bean (Vicia faba L.): archaeobotanical and palaeodietary implications. Archaeol Anthropol Sci 8:555–562. https://doi.org/10.1007/s12520-015-0243-6
Tsutaya T, Yoneda M (2015) Reconstruction of breastfeeding and weaning practices using stable isotope and trace element analyses: a review. Am J Phys Anthropol 156:2–21. https://doi.org/10.1002/ajpa.22657
Unkovich M, Pate J, McNeill A, Gibbs DJ (2001) Stable isotope techniques in the study of biological processes and functioning of ecosystems. Kluwer Academic Publishers, Dordrecht
Vaiglova P, Bogaard A, Collins M, Cavanagh W, Mee C, Renard J, Lamb A, Gardeisen A, Fraser R (2014) An integrated stable isotope study of plants and animals from Kouphovouno, southern Greece: a new look at Neolithic farming. J Archaeol Sci 42:201–215. https://doi.org/10.1016/j.jas.2013.10.023
Valamoti SM (2016) Millet, the late comer: on the track of Panicum miliaceum in prehistoric Greece. Archaeol Anthropol Sci 8:51–63
van Klinken GJ (1999) Bone collagen quality indicators for Palaeodietary and radiocarbon measurements. J Archaeol Sci 26:687–695. https://doi.org/10.1006/jasc.1998.0385
Varalli A, Moggi-Cecchi J, Moroni A, Goude G (2015) Dietary variability in during Bronze Age in Central Italy: First results. Int J Osteoarchaeol 26:431–446. https://doi.org/10.1002/oa.2434
Varalli A, Desideri J, David-Elbiali M, Goude G, Honegger M, Besse M (2021) Bronze Age innovations and impact on human diet: a multi-isotopic and multi-proxy study of western Switzerland. PLoS One 16(1):e0245726. https://doi.org/10.1371/journal.pone.0245726
Varalli A, Moggi-Cecchi J, Goude G (2022) A multi-proxy bioarchaeological approach reveals new trends in Bronze Age diet in Italy. Sci Rep 12(1):12203. https://doi.org/10.1038/s41598-022-15581-0
Wallace M, Jones G, Charles M, Fraser R, Heaton THE, Bogaard A (2015) Stable carbon isotope evidence for Neolithic and Bronze Age crop water management in the Eastern Mediterranean and Southwest Asia. PLoS One 10(6):e0127085. https://doi.org/10.1371/journal.pone
Whitehouse RD (2007) Underground religion revisited. In: Malone C, Barrowclough D (eds) Cult in Context. Reconsidering Ritual in Archaeology. Oxbow, Oxford, pp 97-106.
Warinner C, Tuross N (2009) Alkaline cooking and stable isotope tissue-diet spacing in swine: archaeological implications. J Archaeol Sci 36:1690–1697. https://doi.org/10.1016/j.jas.2009.03.034
Acknowledgements
We would thank Arturo Gnesi, Mayor of Pastena, and the community for their support since 2012, and Robin Skeates for his cooperation. The leading authors are very grateful to Marco Romboni for his support in collagen extraction. The permits to execute the study were provided by Micaela Angle (Ministero della Cultura, Istituto Autonomo Villa Adriana e Villa d’Este) and Carlo Molle (Soprintendenza Archeologica Belle Arti e Paesaggio Frosinone e Latina).
Funding
Open access funding provided by Università degli Studi di Roma Tor Vergata within the CRUI-CARE Agreement. This work was supported by the Italian Ministry of Education, Universities and Research (MIUR) through PRIN 2017 action (1000 Ancient Italian Genomes: Evidence from ancient biomolecules for unravelling past human population Dynamics, GrantID: 20177PJ9XF) allotted to OR and by a 2022 Research Fund Grant from The Prehistoric Society, Institute of Archaeology, UCL, allotted to FC. This research is part of the FC’s PhD thesis in Cultural Heritage, Education and Territory.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
(XLSX 69.5 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cortese, F., De Angelis, F., Achino, K.F. et al. Isotopic reconstruction of the subsistence strategy for a Central Italian Bronze Age community (Pastena cave, 2nd millennium BCE). Archaeol Anthropol Sci 14, 201 (2022). https://doi.org/10.1007/s12520-022-01673-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-022-01673-5