
Abstract
The socio-cultural and economic developments that took place from the Neolithic to the Bronze Age are poorly understood, despite the fact that they were essential for the establishment of fully agro-pastoral economies in Europe. In this study, we aim to assess dietary changes in communities living in southern Italy during this period by examining stable carbon and nitrogen isotope ratios on human bone collagen. In particular, we investigated skeletal remains from seven sites in the southern Italian regions of Calabria (Grotta della Monaca, Grotta di Donna Marsilia and Grotta dell’Antenato), Basilicata (Murgia Timone, Grotta Funeraria and Toppo d’Aguzzo) and Apulia (Ipogeo dei Bronzi) to explore possible variations in diet between different geographic areas and periods. The results of the analysis on bone collagen extracts from 33 human and 12 faunal (sheep, dog, cattle and pigs) specimens attest that the diets of prehistoric southern Italians were mixed and based on the consumption of terrestrial resources, including generally moderate proportions of animal protein (e.g. meat and dairy products) and of C3 plants (e.g. cereals and legumes). Minor differences in the proportion of consumed meat are mostly dependent on the nature of regional environments, with individuals from Basilicata relying more on animal protein than those from Calabria and Apulia. Our study provides insights into the dietary habits of southern Italian populations during the prehistoric period that witnessed an increase both in agriculture and in pastoralism.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In the Adriatic regions, farming spread from the Dalmatian coasts across to Italy mainly during the 6th millennium BCE, as demonstrated by the occurrence of domesticated wheat and barley, with the simultaneous appearance of pottery and the use of obsidian and ground stone (Skeates 2000; Zilhão 2001; Starnini 2002; Guilaine 2003). According to a recent meta-analysis of data from another part of the Mediterranean (Cubas et al. 2019), the definitive break with previous foraging subsistence strategies occurred during the late Neolithic period. More generally, there was an intensification of agricultural practices in the Mediterranean region during this period with a full integration of production cycles in an increasingly anthropized landscape. The shift from foraging to farming, from gathering to food production and from wild to domestic resource consumption clearly occurred at this time in southern Italy, as is demonstrated in the farming settlements of the Murge and Tavoliere (Pessina and Tiné 2008; Lelli et al. 2012). As reported by Lelli et al. (2012) for early Neolithic individuals from coastal and inland sites of Marche, Apulia and Basilicata, dietary differences were present regionally in relation to the consumption of marine and terrestrial animal and plant resources. The estimated fraction of marine fish in the total diet by dry weight was 6 to 30% for coastal sites of Marche and Apulia and 0 to 15% for inland sites of the Tavoliere in Apulia and Murge in Basilicata; both at coastal and at inland sites, however, most of the diet was centred on terrestrial resources (Lelli et al. 2012), although the first farmers of Italy adapted their diet to exploit different environments and landscapes (coastal plains, valley, high plateaus and mountains). The fact that the Neolithic (6th–4th millennium BC) in south-eastern Italy was not characterized by a marked shift to a homogeneous terrestrial diet, as suggested for northern and western Europe (Lubell et al. 1994; Richards et al. 2003), may be a difference that is worth exploring further. It should be noted, however, that another difference with the Atlantic is that the Mesolithic-Neolithic transition in the Mediterranean was not characterized by a sharp shift from intensive marine resource exploitation to fully terrestrial adaptations (Lightfoot et al. 2011; Mannino et al. 2015). Compared with other periods of European prehistory, the Bronze Age is regarded as a much more dynamic period at least in northern Italian regions (Cavazzuti et al. 2019). The first stable isotopes analyses on Bronze Age Italy were carried out by Tafuri et al. (2009) on materials from the north and south of the peninsula and by Lai et al. (2013) on specimens from Sardinia. Further isotopic analyses have been undertaken more recently on Early-Middle Bronze Age communities from sites across Italy (Varalli et al. 2016a, 2016b; Masotti et al. 2017; De Angelis et al. 2019; Rumolo et al. 2020). These studies have demonstrated that the transition from the Early Bronze Age (2300–1700 BC) to the Middle Bronze Age (1700–1350 BC) represented a moment of change, involving isotopically traceable changes in diet. In particular, the Middle Bronze Age settlements of central-northern Italy seem to have witnessed the introduction of new crops (such as millet). In contrast, the southern Italian communities were characterized by a diet inherited from the Neolithic based on C3 plant consumption (i.e. wheat and barley) (Tafuri et al. 2009). However, research examining diet in southern Italy during the Bronze Age is scarce since only a handful of sites have been investigated by means of isotope analysis (Tafuri et al. 2009; Rumolo 2020).
This study aims to provide a more comprehensive understanding of the diet for the communities living in prehistoric southern Italy, to clarify findings that emerged in previous research and to add new isotopic data to the literature. In particular, we aim to extend the area of investigation, involving seven sites belonging to different spatio-temporal contexts of southern Italy. Furthermore, we explain possible dietary changes in relation to subsistence strategies increasingly specialized on agriculture and pastoralism, which became the main source of livelihood during the Mediterranean Bronze Age (Salvadei and Santandrea 2003; Peroni 2004; Bietti Sestieri 2011).
Palaeodietary indicators
The analysis of stable isotope ratios from tissues such as bone or teeth is a direct and widely used technique for palaeodietary reconstruction because it reflects a mixture of isotopic signatures from the foods consumed during the last years of an individual’s lifetime (Lee-Thorp 2008; Schoeninger 2011; Reitsema 2013). In particular, stable isotope analysis on bone collagen reflects mainly the protein fraction of the diet. Carbon stable isotopes (δ13C) provide information on the photosynthetic pathway (i.e. C3, C4 or CAM plants) and ecosystem of provenance of the proteins consumed (i.e. terrestrial, freshwater or marine). In environmental settings with limited presence of C4 plants, such as those of prehistoric Europe, enriched δ13C values in human bone collagen have been deemed to provide ‘a direct indication of the amount of marine foods consumed’ (Lelli et al. 2012 citing Richards and Hedges 1999). Carbon and, especially, nitrogen isotope values (δ15N) reflect the trophic level of the dietary protein consumed, increasing by 0–1‰ for δ13C and 3–5‰ for δ15N at each step up the food chain (Schoeninger et al. 1983; Bocherens and Drucker 2003; Herrscher 2003; Mannino 2009; Mannino et al. 2015). In order to interpret the isotopic compositions of humans and to differentiate between terrestrial and freshwater food sources, carbon and nitrogen isotope ratios of faunal samples are necessary to understand whether the proteins consumed by humans were derived from the consumption of carnivores or herbivores (meat, milk and dairy products) (Hedges and Reynard 2007; Lee-Thorp 2008). In practice, stable isotope ratios are informative on the type of diet, allowing us to distinguish broadly the different food items that were regularly consumed by an individual (e.g. meat versus plants, terrestrial versus aquatic foods, C4 plants versus C3 plants) (DeNiro and Epstein 1978, 1981; DeNiro 1987; Lee-Thorp 2008).
The dietary patterns in European prehistoric human populations have been studied for several geographical areas (e.g.Triantaphyllou et al. 2008; Lopez-Costas et al. 2015; Goude et al. 2011; McClure et al. 2011; Fontanals-Coll et al. 2015; Alt et al. 2016; Waterman et al. 2016; Goude and Fontugne 2016; Salazar-García et al. 2013; Herrscher et al. 2013; Fernandez-Crespo and Schulting 2017; Dunne et al. 2019), but, as far as Italy is concerned and by means of isotopic analyses, only a few studies have addressed this topic on a limited number of prehistoric sites and periods. Some research has, for instance, been carried out on hunter-gatherers from the Palaeolithic and Mesolithic in western Sicily demonstrating that forager diets in the central Mediterranean were characterized by substantial consumption of animal protein, of which little generally came from marine resources (e.g. Mannino et al. 2012, 2015). The transition to farming at the beginning of the Neolithic was marked by a decline in wild resource exploitation in the Mediterranean, as elsewhere in Europe, with little or no reliance on seafood (e.g. Tauber 1981; Schulting 1998; Richards et al. 2003; Lelli et al. 2012). At the same time, the homogeneous ratios of carbon and nitrogen after the introduction of domesticated animals and plants are indicative of more terrestrial diets (Richards et al. 2003; Hedges and Reynard 2007), which in some cases became more variable at the transition from the Early to the Middle Bronze Age, when different dietary patterns were highlighted for instance in central Italy (Varalli et al. 2016 a).
Archaeological background
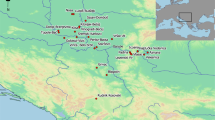
In this section, we briefly describe the archaeological sites from which the skeletal materials investigated in this study were recovered. The geographical locations of the sites are shown in Fig. 1.

The sites investigated according to the archaeological period of the sampled specimens. Two different specimens were sampled from Grotta dell’Antenato: GdA01 dated to the Late Neolithic (blue) and GdA02 dated to the Middle Bronze Age (orange)
Calabria
Grotta della Monaca (GdM)
Grotta della Monaca is a karstic cave located near Sant’Agata di Esaro (Cosenza) in north-western Calabria. During the Bronze Age, the deepest recesses of this cave site were used as a burial ground, which included five burials within niches and fractures in the rock walls. The skeletal remains were commingled and were not found in anatomical connection. The grave goods consisted of few elements in poor state of preservation and not clearly associated with the inhumated individuals (Arena et al. 2014; Arena and Gualdi-Russo 2014). Radiocarbon dating previously performed on the human skeletal remains has dated the burials to the Middle Bronze Age and specifically to the second half of the 2nd millennium BCE (GRN28721: 1770–1520 cal. BCE (95.4%); LTL5087A: 1560–1389 cal. BCE (95.4%)) (Larocca 2005; Arena et al. 2014).
Grotta dell’Antenato (GdA)
Grotta dell’Antenato is a cave located in north-eastern Calabria, near Cassano allo Ionio (Cosenza), and it was used as a burial ground for a long time during prehistory, spanning from the Neolithic to the Copper and Bronze Ages (as shown by our radiocarbon dating and by relative dating based on ceramic fragments studied by Ippolito (2015–16)). Ritual votive offerings were dedicated to the inhumated individuals, as suggested by the animal remains and vessels found at Grotta dell’Antenato (Ippolito 2015–16).
Grotta di Donna Marsilia (GdMR)
This cave is located near Morano Calabro (Cosenza), in a hilly area called ‘contrada Sassone’. The human skeletal remains recovered at the site belong to a single individual. Unpublished archaeological data indicate that they were associated with pottery of Eneolithic age (according to reports by archaeologists at the Museo Archeologico Nazionale della Sibaritide, Cosenza).
Basilicata
Murgia Timone (MT) and Grotta Funeraria (GF)
These two sites are located in the hilly area of Murgia (Matera), along the edges of the ravine of the Bradano river, where several burials were discovered. Information on the archaeological data and the results of our radiocarbon dating indicate a long frequentation of this area for funerary purposes. The published AMS radiocarbon date on a human skeleton from Grotta Funeraria indicates that the earliest burials probably date to the late Neolithic (GF01a: 5064–4894 cal. BCE (90.1%)). The little published information for these two sites mainly relates to Murgia Timone, where archaeological excavations carried out between 1800 and 1900 discovered three graves (Quagliati 1896; Patroni 1897; Ridola 1901, 1912; Grifoni Cremonesi 1976; Lo Porto 1988; Tunzi Sisto 1999; Matarese 2014). For the purpose of our study, we have only sampled faunal remains from the Middle Bronze Age contexts, as they were the only finds with a reliable chronology.
Toppo Daguzzo (TDT)
The ancient settlement of Toppo Daguzzo (Melfi) is located about 50 km from Potenza, in the fertile farmlands of the Ofanto Valley. Systematic excavations conducted at the site since 1980 have discovered three burials along the slopes of the hill. The first grave, consisting of an underground structure with two compartments and a long access corridor, dates back to the sixteenth to fifteenth centuries BCE. The second burial was a pit grave located close to the former and dating to the same chronological period. The third inhumation was discovered in a large underground chamber, dug into the tufa rock and with a long corridor (dromos), dating back to the Middle Bronze Age between the fifteenth and fourteenth centuries BCE. Inside the chamber of the third burial, the skeletal remains of about ten individuals in poor state of preservation and without grave goods were found.
A second layer in a better state of preservation was intercepted about 20 cm in depth, at which two chambers were unearthed. In the first, there was clear evidence of funerary rituals (with vessels and burned faunal remains), while in the second, there were six males with weapons (interpreted as warriors), four women (most of which with ornaments) and a child. Different vessels were placed close to the infant inhumation. This second depositional layer has been interpreted as the burial place of warrior aristocracy (Cipolloni Sampò 1986a, 1986b; Bietti Sestieri 2011).
Apulia
Ipogeo dei Bronzi (IpB)
Ipogeo dei Bronzi is located in northern Apulia near Trinitapoli (Foggia), in a flat area of the Gargano promontory around 90 m above sea level. It was a hypogeal burial ground and dates back to the Middle Bronze Age. Archaeological excavations unearthed around 200 individuals, both males and females from all age groups. Currently, part of the skeletal remains has been lost, and the remaining remains are in poor state of preservation (Bietti Sestieri 2011). The archaeological record for this hypogeum includes the remains of distinct groups of individuals, presumably belonging to different family units. The male grave goods include weapons (e.g. swords and daggers), some knives and two razors, while the grave goods associated with female individuals are bronze ornaments, bone objects and polished stones (such as amber, faïence and glass paste from the Aegean and the Eastern Mediterranean). It has been hypothesized that the presence of several collective burials mirrored a family-based social organization. However, the fact that grave goods are richer in some burials than in others seems to suggest that differences in social status were present between the inhumated individuals (Tunzi Sisto 1997, 1999; Vanzetti 1999; Mazzei and Tunzi Sisto 2005; Bietti Sestieri 2011).
Materials and methods
Samples
The study material derives from archaeological collections kept at the local storage facilities of regional archaeological agencies and museums (as listed in the acknowledgements). Anthropological analyses of the human specimens sampled for this study were carried out either directly at these institutions or at the Laboratory of Archaeo-Anthropology and Forensic Anthropology of the University of Ferrara (Italy). To date, only some preliminary anthropological studies for the site of Grotta della Monaca have been published (Scattarella et al. 2005; Arena et al. 2014).
A total of 39 specimens (29 human and 10 animal bone specimens) were analysed for stable isotope analyses (Table 1). Out of these, three human bones (2 from GdA and 1 from GF) from the sites described above were radiocarbon dated (Table 2). The first two individuals (GdA) were at first ascribed to the Bronze Age, but the necropolis also showed evidence of activities earlier than this period. AMS dating confirmed the Middle Bronze Age only for one specimen (GdA02: AAR25752), while the other one has been dated to the late Neolithic (GdA01: AAR25751). A similar late Neolithic date has been obtained on the specimen analysed from GF (GF01a: AAR25755).
We sampled specimens for isotopic analyses after establishing the biological profile of each individual and only took adults. Moreover, when skeletons were commingled, only bones used for MNI (Minimum Number of Individuals) calculation were sampled for this study to avoid multiple sampling of the same individual. When possible, age assessment was done on the basis of morphology of the auricular surface of the os coxae (Lovejoy et al. 1985; Meindl and Lovejoy 1985), pubic symphysis surface changes (Todd 1921), synchondrosis of ectocranial suture (Meindl and Lovejoy 1985; Buikstra and Ubelaker 1994) and tooth wear (Brothwell 1981; Lovejoy et al. 1985). Following the criteria by Buikstra and Ubelaker (1994), we divided the specimens into three age categories: young adults (20–35 years), middle adults (35–50 years), old adults (50+). Sex diagnosis was based on skull and pelvis morphology (Acsádi and Nemeskéri 1970) and osteometric characters of the skull (Giles and Elliot 1963; Demoulin 1972), mandible (Piquet 1956), scapula (Olivier and Pineau 1958), humerus (Ferembach et al. 1977–1979, 1980; Dittrick and Suchey 1986; France and Horn 1988), ulna (France 1998), femur (Iordanidis 1961; Dittrick and Suchey 1986), tibia (Ferembach et al. 1977–1979, 1980) and talus (Gualdi-Russo 2007).
Given the funerary context, quality of some of the excavations and state of preservation of the archaeological deposits, only a few faunal remains were available from a handful of the sites examined. For this reason, we sampled fauna from nearby coeval sites (MT, GdMR, GdA).
Analytical methods
The AMS radiocarbon dating and stable isotopes analyses were conducted at the Aarhus AMS Centre, Department of Physics and Astronomy of Aarhus University (Denmark). Collagen was extracted from 0.5 g of cleaned bone fragments (cortical bones), according to the protocol by Longin (1971) with modifications by Brown et al. (1988) and Jørkov et al. (2007). The ultrafiltration protocol, introduced by Brown et al. (1988), separates high molecular weight (MW) components of the gelatinized ‘collagen’ (˃ 30 kD) from low MW fractions (< 30 kD). The use of ultrafilters improves the quality of extracted collagen (evaluated by C/N ratios within biogenic ranges; DeNiro 1985; Higham et al. 2006). Each collagen extract was then analysed by an elemental analyser (EA) coupled to a continuous flow isotope ratio mass spectrometer (EA-IRMS). In total, 62 ‘samples’ were analysed: 39 human and faunal collagen extracts, 9 duplicates, 4 background bone extracts (a 40.000 BP old whale bone) and 10 samples of an in-house gelatine standard (‘GEL-A’). Each standard weighed 0.200–0.300 mg. The background bone was used as a secondary control sample. The carbon (δ13C) and nitrogen (δ15N) isotopes were expressed using the delta (δ) notation, as part per thousand (‰). The δ13C ratio is reported relative to the V-PDB marine fossil limestone standard (Vienna Pee Dee Belemnitella fossil) and the δ15N ratio relative to the atmospheric N2 (AIR) standard. The extracted collagen material was quality-checked in terms of collagen percentage and C/N ratios. For the purpose of this study, extracts were considered well-preserved when they had yields of ˃ 1%, C% ≥ 30% and N% ≥ 11% (wt%), as proposed by van Klinken (1999), and C/N molar ratios between 2.9 and 3.6, as originally proposed by DeNiro (1985).
For radiocarbon analysis, the collagen was converted to CO2 by combustion in sealed evacuated quartz tubes with 200 mg CuO. The CO2 was reduced to graphite by the H2 reduction method using an iron catalyst and MgClO4 to remove the water (Vogel et al. 1984; Santos et al. 2007). The samples were 14C dated using the HVE 1MV Tandetron accelerator AMS system at the Aarhus AMS Centre (Olsen et al. 2016). 14C dates are reported as uncalibrated 14C ages BP normalized to − 25‰ according to international convention using online 13C/12C ratios (Stuiver and Polach 1977). Dates in the article have been calibrated with OxCal v.4.3 (Bronk Ramsey 2009) using the calibration curve IntCal13 (Reimer et al. 2013) and are reported as calibrated ages BC.
All statistical analyses were performed using STATISTICA for Windows—version 11 (StatSoft Inc., Tulsa, OK, USA). Means and standard deviations (SD) were computed to describe the sample. Due to the low sample size, non-parametric tests were used to compare means by Mann-Whitney U test (2 groups) and Kruskal-Wallis test (more than 2 groups).
Multiple regression analysis was used to analyse the association between carbon and nitrogen isotope values, each of them separately tested as dependent variables, and the biological (i.e. the other isotope, sex, age category) and chronological-environmental characters (period, region), tested as independent variables. Multicollinearity among independent variables was examined by the variance inflation factor (VIF) (VIF values < 10 were considered acceptable). We performed separate models for each stable isotope.
For all statistical analyses, a significance level of p < 0.05 was used.
Results
Table 1 summarizes the anthropological evaluation carried out on the human remains and shows the results of carbon (δ13C) and nitrogen (δ15N) isotope analyses performed on faunal and human samples with the quality control indicators.
The three human bone specimens from Grotta dell’Antenato (GdA01, GdA02) and Grotta Funeraria (GF01a) produced uncalibrated dates of 6072 ± 36 BP, 3202 ± 45 BP and 6072 ± 34 BP, respectively. Calibrated dates are reported in Table 2. All the 39 specimens yielded well-preserved collagen (collagen yields ˃ 1%) according to the criteria proposed by van Klinken (1999), except specimen IpB06 with a low yield of 0.5%, and therefore this sample is to be considered with caution. Considering all 29 human specimens examined, the δ13C values are between − 20.3 and − 18.9‰, and δ15N values are between 7.0 and 10.8‰. The 10 faunal specimens show δ13C values between − 21.5 and − 19.1‰, and δ15N values are between 3.0‰ and 8.5‰. The distribution of faunal and human stable isotope ratios by site is shown in Fig. 2.

Scatterplot of δ13C and δ15N values of human and faunal samples from the southern Italian sites investigated. Humans have been sampled from southern Italian sites dated to Neolithic (GdA, GF), Copper (GdMR) and Bronze (IdB, TDT, MT, GF, GdA, GdM) Ages. Animals are from Neolithic (GdA), Copper (GdMR) and Bronze (MT) Ages
Isotope analyses on the faunal samples
Only a few faunal samples found in three of the seven archaeological sites considered and belonging to different periods were available for this study. The Neolithic faunal samples (GdA) resulted in bone collagen δ13C values from − 21.5 to − 20.5‰ (− 21.0 ± 0.7‰) and δ15N values from 5.3 to 6.9 ‰ (6.1 ± 1.1‰). The Copper Age faunal samples (GdMR) resulted in bone collagen δ13C values from − 21.4 to − 19.7‰ (− 20.6 ± 0.9‰) and δ15N values from 3.0 to 8.5‰ (5.2 ± 2.9‰). However, it should be noted that the sample GdMR04 (Sus scrofa) differs from the others in isotopic values (δ13C = − 20.7‰ and δ15N = 3.0‰), suggesting a diet characterized by a prevalent intake of vegetal protein (C3 plant–derived). The carbon and nitrogen analyses performed on the Bronze Age faunal remains from Murgia Timone (Basilicata) resulted in bone collagen δ13C values from − 20.7 to − 19.1‰ (− 19.7 ± 0.9‰) and δ15N values from 5.6 to 7.4‰ (6.8 ± 1.0‰) for the herbivores. The omnivores from this site had a bone collagen δ13C mean value of − 19.8 ± 0.9‰ and δ15N mean value of 7.9 ± 0.2‰. The faunal sample MT01 (Ovis aries) was characterized by higher δ13C values (− 20.7‰) and a slightly lower δ15N value (5.6‰) than other herbivores (cattle).
Isotope analyses on the human samples
Mean human isotopic values for different archaeological periods were estimated (Table 3), and the data from these were compared to evaluate possible dietary differences. Statistical testing indicated a significant difference for the δ15N values (Kruskal-Wallis test: H = 10.218, p = 0.0060), but not for the δ13C (Kruskal-Wallis test: H = 0.873, p = 0.65). For the Neolithic faunal specimens, the human δ13C mean value was 1.5‰ higher than the omnivore mean value (− 21.0 ± 0.7‰) and the human δ15N value 3.8‰ higher than that of the omnivores (6.1 ± 0.5‰). The Copper Age human specimen had a δ13C value 0.9‰ higher than the omnivore mean value (− 20.6 ± 0.9‰) and δ15N value 4.9‰ higher than the δ15N mean value for the omnivores (5.2 ± 2.9‰). The Bronze Age human specimens had a δ13C mean value 0.1‰ higher than the herbivore mean value (− 19.7 ± 0.9‰), and their δ15N mean value was 2.0‰ higher than that of the herbivores (6.8 ± 1.0‰). In addition, the human δ13C mean value was by 0.2‰ higher than the omnivore mean value (− 19.8 ± 0.9‰) and the human δ15N value 0.9‰ higher than the mean value for the omnivores (7.9 ± 0.2‰). Applying the trophic level isotope enrichments of δ15N = 3–5‰, δ13C = 0–1‰ (Bocherens and Drucker 2003), the comparison between animal and human mean values suggests that Neolithic and Copper Age humans were one trophic level higher than the omnivores, which is compatible with a regular consumption of animal meat and/or of their secondary products. However, the Bronze Age stable isotope values seem to suggest lower animal protein intake. Two reference animal bone samples of deer (Cervus elaphus) and sheep/goat (Ovis vel Capra), recovered from the Middle Bronze Age site of Madonna di Loreto (Foggia, Apulia) (Tafuri et al. 2009), have a δ13C value of − 20.4‰ and a δ15N value of 7.2‰ for C. elaphus and a δ13C value of − 20.4‰ and a δ15N value of 7.1‰ for Ovis/Capra. The Bronze Age human values obtained in this study are higher than these faunal values by only 0.4‰ in δ13C, while the δ15N mean value is higher than Ovis/Capra only by 1.7‰ and than Cervus elaphus only by 1.6‰, which is considerably less than a trophic level increase. Table 4 shows these and other isotopic values of faunal samples from the Neolithic to the Bronze Age for the southern continental Italy according to literature.
Stable isotopic patterns in prehistoric southern Italian communities
We checked whether there was variability in food consumption based on the sex and age of all analysed individuals. The δ13C and δ15N means are similar between sexes with slightly less δ15N variation among males (δ13C: − 19.7 ± 0.3‰ in females, − 19.5 ± 0.4‰ in males; δ15N: 9.0 ± 1.0‰ in females, 9.5 ± 0.6‰ in males), suggesting a similar diet for both sexes.
No statistical difference was found between the group of young adults (YA) and the other adults (middle adults, old adults, generic adults) (δ13C: − 19.8 ± 0.4‰ in YA, − 19.5 ± 0.4‰ in others; δ15N: 8.9 ± 0.8‰ in YA, 9.2 ± 1.0‰ in others), suggesting the consumption of similar foods between different age classes.
Statistical testing for sex (δ13C: − 19.8 ± 0.3‰ in females, − 19.4 ± 0.4‰ in males; δ15N: 8.7 ± 0.7‰ in females, 9.2 ± 0.7‰ in males) and age (δ13C: − 19.9 ± 0.3‰ in YA, − 19.5 ± 0.4‰ in others; δ15N: 8.5 ± 0.7‰ in YA, 8.9 ± 0.9‰ in others) in the Bronze Age group has also failed to detect any statistically significant differences.
Although there was no archaeological information on specific grave goods related to the sampled skeletons, there was evidence of their presence in the examined sites (Larocca 2005; Cipolloni Sampò 1986a, 1986b; Bietti Sestieri 2011). In particular, grave goods at Toppo Daguzzo and Ipogeo dei Bronzi testified the presence of weapons in male burials (Cipolloni Sampò 1986a; Borgognini Tarli 1992; Tunzi Sisto 1999, 2005). In addition, there was a preferential differentiation among burials of males, females and sub-adults (which could correspond to roles or ranks of individuals) at Ipogeo dei Bronzi (Vanzetti 1999). A reasonable hypothesis may be that dietary practices followed the social complexity, with certain foods (i.e. meat) reserved for particular social groups (Schoeninger 1979). However, our stable isotope comparisons did not show any evidence of ‘status’ differences neither for sex nor for age. Although our results were obtained on a small group of samples, we can assume that cultural differences did not extend to the diet or at least, if any dietary differences existed, these were not isotopically measurable. Occasional consumption of ‘prestige’ foods would not be reflected in the long-term dietary stable isotope record.
We estimated the mean values of δ13C and δ15N according to the three geographical areas (regions) to which the sites belong. Differences between regions (Calabria—δ13C: − 19.8 ± 0.3‰, δ15N: 8.7 ± 1.3‰; Basilicata—δ13C: − 19.6 ± 0.4‰, δ15N: 9.6 ± 0.5‰; Apulia—δ13C: − 19.4 ± 0.3‰, δ15N: 8.8 ± 0.6‰) resulted fairly close to significance for δ13C (H = 5.264 p = 0.071) and for δ15N (H = 5.419 p = 0.067).
As part of our data analysis, we also checked whether the stable isotopes ratio could be explained on the basis of the biological characteristics of the examined individuals and of their living environment by applying multiple regression analyses (Tables 5 and 6). In these analyses, a model was tested for each of the two stable isotopes considered, thus assessing the influence of the other isotope, sex, age-at-death, archaeological period and living environment of the individual. The final model developed for δ15N explained 50% of the variance in the prehistoric individuals examined (Table 5). The final model for δ13C explained approximately one third of the variance in the same individuals (Table 6). Multicollinearity did not occur between independent variables (as indicated by the VIF values). In both models, considering individual variables, the other isotope and the living environment (region) were found to be significant determinants. In particular, δ13C values and living in Basilicata proved to be positive and significant explanatory variables of nitrogen isotopic values (Table 5); δ15N (positive association) and living in Basilicata (negative association) proved to be significant explanatory variables of carbon isotopes (Table 6).
Discussion
The analysis of chemical markers of food habits, carbon (δ13C) and nitrogen (δ15N) isotopes, was performed on faunal and human collagen extracts from seven southern Italian prehistoric sites. The general pattern observed in the examined territory indicates a mixed diet based on the consumption of terrestrial resources and mostly characterized by a vegetal protein intake (C3 plant–related) with a moderate intake of animal protein for all the periods examined.
Following the univariate analysis, we applied multiple regression analyses to identify biological and environmental explanatory variables for the stable isotope values and obtained statistically significant models for both δ15N and δ13C. Two main independent variables account for the increase of a stable isotope in the model—a dietary biomarker (the other stable isotope) and the environmental context (represented by the Italian regions). The territory of Basilicata, located between Campania and Apulia, and where three of the seven investigated sites are located (two of them having provided human specimens), significantly influenced both the increase of δ15N and the decrease of δ13C. It is likely that the physical characteristics of this region between Campania and Apulia influenced the diet of its inhabitants, particularly with the consumption of wild terrestrial fauna. The territory of Basilicata is intersected by numerous watercourses and includes large fluvio-lacustrine basins, but is overall more mountainous (92% of the territory is mountainous or hilly) than other southern regions and likely a territory where hunting could have been conducted more easily given its more isolated and less anthropized character.
Environmental comparisons
Coastal vs non-coastal sites
Both Neolithic sites examined in this study (GdA and GF) are located in inland hilly environments and have yielded similar isotopic values, indicating a homogeneous diet. Similarly, there were no significant differences in δ13C and δ15N values between these individuals of the late Neolithic and those of the early Neolithic from inland sites of southern Italy (Ripatetta in the Apulian Tavoliere; Balsignano, Masseria Maselli and Palata in Apulian Murge) (Lelli et al. 2012). Instead, the comparison with early Neolithic individuals from coastal sites of Apulia (Torre Castelluccia, Samari and Grotta delle Mura) (Lelli et al. 2012) shows a significant difference only for carbon (p = 0.0180 for δ13C; p = 0.4062 for δ15N), which was higher at coastal sites (mean values: − 18.87 ± 0.59‰; n = 7). In the absence of C4 plants (e.g. at southern Italian sites during the Neolithic), higher δ13C values on human collagen provides us with an indication of a utilization of marine food resources at the coastal sites (Richards and Hedges 1999). In this case, higher marine consumption by coastal farmers may be the most parsimonious interpretation, although the carbon and nitrogen isotope values only indicate a moderate consumption of fish (Lelli et al. 2012). Nevertheless, a certain degree of dietary variability during the Neolithic can be traced in relation to the living environment: early southern coastal farmers from Torre Castelluccia, Samari and Grotta delle Mura in Apulia consumed higher amounts of fish than the coeval individuals from Fosso Fontanaccia in Marche (p = 0.0082 for δ13C; p = 0.0080 for δ15N) (Lelli et al. 2012). As several studies of Mediterranean Neolithic sites have demonstrated, the exploitation of wild foods (e.g. fish) was not a homogeneous phenomenon (Papathanasiou 2003; Le Bras-Goude et al. 2006; Lelli et al. 2012).
The comparison of our Bronze Age human samples from the southern Italian sites located closer to the coast (distance < 20 km from the coastline: GdM, IpB) with those from inland sites (distance > 20 km from the coastline: GdA and TDT) attests no significant difference between groups (δ13C: − 19.6 ± 0.4‰ in coastal group, − 19.6 ± 0.4‰ in the inland group; δ15N: 8.7 ± 0.9‰ in the coastal group, 9.2 ± 0.8‰ in inland group) (p > 0.05), indicating a homogeneous terrestrial diet for southern Italian Bronze Age communities and essentially a lack of marine/freshwater food exploitation. A lack of consumption of aquatic resources in Mediterranean coastal sites was also found in several previous studies, indicating that aquatic foods were only a minor component within a predominantly terrestrial diet during the Bronze Age (e.g. Craig et al. 2006; Lai et al. 2013). Exceptions to this trend may be represented by the site of Grotta dello Scoglietto in central Italy (Varalli et al. 2016a) and Punta di Zambrone in Calabria (Rumolo et al. 2020). The latter study has, in our opinion, not fully justified a good collagen quality of the included samples and has not demonstrated that its results may not indicate marine resource consumption by the analysed humans (favouring the interpretation that C4 plants were consumed at the site). Comparing faunal and human isotope values suggests a relatively limited consumption of animal protein, which is in line with interpretations from previous studies (e.g. Tafuri et al. (2009) estimated that C3 plants provided around 65–70% of total dietary protein for the Toppo Daguzzo humans). However, this must be considered with caution as we were not able to study faunal remains of all species (herbivores and carnivores) or from each considered site. Nevertheless, the nitrogen isotope values from human collagen extracts (of around 9‰) seem to indicate a moderate animal protein intake, also if we cannot exclude the effects of highly intensive animal manuring (Treasure et al. 2016). The zooarchaeological record shows that animal-based Bronze Age subsistence strategies were mainly centred on livestock raising rather than hunting (De Grossi et al. 2004). In Italy, as in Mediterranean Europe, the Bronze Age animal economy was mainly based on the consumption of sheep and goats for meat and dairy products (Arrighi et al. 2007; De Grossi 2013; Maini and Curci 2013), although evidence for animal exploitation varies greatly depending on geography, environment and culture (Riedel 1996; Battisti and Marconi 2004; Harding and Fokkens 2013). As at northern Italian sites (i.e. Ledro and Fiavè) (Perini 1987; Bietti Sestieri 2011; De Grossi 2013; Maini and Curci 2013), there was a prevalence of sheep and goats among the domestic species also in southern Italy, with an increase in ovicaprid exploitation during the Late Bronze Age. This has been demonstrated, for instance, by the archaeological record at Broglio di Trebisacce in Calabria and Coppa Nevigata in Apulia, where wild species were completely absent (Bökönyi and Siracusano 1987; Tagliacozzo 1994; Bietti Sestieri 2011). It is, hence, assumed that hunting was generally infrequent during the Early and Middle Bronze Age (Tagliacozzo 1994; De Grossi 2013; De Grossi et al. 2004).
Southern Italian Bronze Age sites vs central-northern sites
For the Bronze Age, we expected substantial differences of isotope values between northern and southern sites because, as is known from the literature, millet was introduced in northern Italy during this period. Contrary to what has been recently proposed by Rumolo et al. (2020), it was not regularly consumed in southern Italian communities of the same period (Tafuri et al. 2009; Varalli et al. 2016a). In northern Italy, Panicum miliaceum cultivation started during the Early Bronze Age, as shown by an increase in the cultivation of this cereal in the Middle/Late Bronze Age at Monte Covolo (Pals and Vorrips 1979) and at Canár (Castiglioni et al. 1998; Castelletti et al. 2001) and as attested at Terramara di Montale by the archaeological record (Mercuri et al. 2006). During this period, there was strong economic development that resulted in a significant increase in settlements in central and northern Italy. The resulting environmental changes, such as deforestation, led to the aridification of the soils, which facilitated the introduction of millet (Valsecchi et al. 2006; Bietti Sestieri 2011; Cremaschi et al. 2016). Moreover, commercial trade with transalpine Europe may have favoured the diffusion of this new crop (Varalli et al. 2016b). The palaeobotanical record indicates that millet was imported from Western Europe with an increase in cultivation around 1700 BCE (Lake de Bourget, South-East France) (Rösch 1998; Jacob et al. 2008). According to this view, our analysis of published data (Varalli et al. 2016 b) indicates that the carbon mean value of the sites located in the Po valley in north Italy (− 18.91 ± 2.29‰) was significantly higher (U = 979.0; p = 0.0007) than the one for the southern sites presented here (− 19.44 ± 0.62‰). This difference can be explained by the consumption of different vegetal proteins: C4 plant–based for central-northern Italian sites and C3 plant–based for southern sites. There was no statistical difference between the nitrogen values obtained from our southern Italian samples (8.52 ± 0.73‰) and from their northern counterparts (8.41 ± 0.87‰) (U = 1269.5; p = 0.13).
Palaeobotanical data from coeval southern Italian sites (i.e. Broglio di Trebisacce in Calabria, Lavello in Basilicata, Coppa Nevigata, Piazza Palmieri, Scoglio di Apani, Roca and Scalo di Furno in Apulia) confirm that cultivated crops were limited to C3-based plants. Among these, archaeological finds attest the presence of einkorn wheat (Triticum monococcum) and emmer wheat (Triticum dicoccum), common wheats (Triticum aestivum, T. compactum, T. durum) and barley (Hordeum vulgare, H. distichum, H. exasticum), as well as the scarcity of spelt, rye and oats and a small variety of broad beans (Vicia faba var. minor), lentil (Lens culinaris) and chickling pea (Lathyrus cicera, L. sativus) (Caldara et al. 1999; Bietti Sestieri 2011; Primavera and Fiorentino 2014).
Comparisons between Italian prehistoric sites by period
Since our results suggested a change in the nitrogen isotopic values between Neolithic and Bronze Age samples, statistical analyses (combining current and previously published studies, but excluding infants and children) (Lelli et al. 2012 for Neolithic sites; De Angelis et al. 2019 for Copper Age; Tafuri et al. 2009, Varalli et al. 2016b and Masotti et al. 2017 for Early-Middle and Late Bronze Age) were performed to better highlight the changes in human diet that occurred during Italian prehistory (Fig. 3a, b). Significant differences were found among samples of different periods for both carbon (H = 76.17, p < 0.001) and nitrogen (H = 39.21, p < 0.001). The human mean δ15N values range from a minimum of 8.21 ± 0.96‰ in the Middle Bronze Age to a maximum of 10.14 ± 1.93‰ in the early Neolithic Age. According to paired comparisons between periods, the Middle Bronze Age sample has lower values of δ15N than the samples of all other periods. The differences were statistically significant for all groups with the exception of the Early-Middle Bronze Age sample. As shown in Fig. 3a, human nitrogen values suggested a higher animal protein intake during the Neolithic, with a gradual decrease in subsequent periods up to the Middle Bronze Age. This result may support the hypothesis that widespread intensive cereal cultivation and consumption did not occur before the Middle Bronze Age and that cereal cultivation was more important than pastoralism in Mediterranean Bronze Age economies (Salvadei and Santandrea 2003; Tafuri et al. 2009). However, an increase in nitrogen values was observed for the Middle-Late Bronze Age sites. The human mean δ13C values vary from a minimum of − 20.09 ± 0.70‰ in the Early-Middle Bronze Age to a maximum of − 15.23 ± 0.84‰ in the Middle-Late Bronze Age. Paired difference tests comparing samples of different periods indicated that the sample from the Middle-Late Bronze Age was significantly different from all the other samples. Moreover, the Middle Bronze Age sample differed from the Early-Middle Bronze Age one and the latter from the Copper Age sample (Fig. 3b). The samples from the early Neolithic to the Copper Age had similar mean values of δ13C, consistent with a diet based on terrestrial C3 resources. During the Bronze Age, there was an increase in δ13C. The trend described for Italy is confirmed by other studies carried out in Europe, in particular in Hungary, where the human carbon and nitrogen isotopic values show similar variation from the Neolithic to the Bronze Age (Gamarra et al. 2018).

a, b Box Plot of human mean values of nitrogen (a) and carbon (b) from early Neolithic to Middle-Late Bronze Age in Italy. Human isotopic values from sample of this research were combined with those from previous studies (Tafuri et al. 2009; Varalli et al. 2016a, 2016b; Lelli et al. 2012; Masotti et al. 2017; De Angelis et al. 2019) considering the following sites from different Italian regions: Early Neolithic sites (n = 19): Fosso Fontanaccia (Marche), Torre Castelluccia (Apulia), Samari (Apulia), 4 = Grotta delle Mura (Apulia), Ripatetta (n = Apulia), Balsignano (Apulia), Masseria Maselli (Apulia), Palata (Molise); Late Neolithic sites (n = 7): Grotta dell’Antenato (Calabria), Grotta Funeraria (Basilicata); Eneolithic sites (n = 47): Grotta di Donna Marsilia (Calabria), Podere Cucule (Tuscany), Le Lellere (Tuscany), Buca di Spaccasasso (Tuscany), Grotta del Fontino (Tuscany), Fontenoce di Recanati (Marche); Early-middle Bronze Age site (n = 67): Arano di Cellore (Veneto), Grotta dello Scoglietto (Tuscany), Ballabio (Lombardy), Sedegliano (Friuli); Middle Bronze Age sites (n = 55): Grotta della Monaca (Calabria); Ipogeo dei Bronzi (Apulia),Grotta dell’Antenato (Calabria), Toppo d’Aguzzo (Basilicata), Lavello (Basilicata), Grotta Misa (Latium), Felcetone (Latium); Middle-Late Bronze Age site (n = 19): Olmo di Nogara (Veneto). Horizontal lines within the box represent median values; vertical lines indicate the data range
Conclusions
In conclusion, our isotope record suggests a similar general nutritional behaviour for all analysed environments from the Neolithic to the Bronze Age in southern Italy: a terrestrial diet reliant on the consumption of C3 crops (cereals and legumes) with a moderate animal protein intake. Nevertheless, some regional and temporal variability emerged with reference to the literature. There was higher variability in the consumption of animal protein during the Neolithic, probably due to the different adoption of new subsistence strategies (i.e. agriculture) depending on the environments. A marked shift to a more homogeneous diet in Italian prehistoric communities did not occur before the Middle Bronze Age, when specialized agriculture became the main subsistence strategy in Mediterranean communities, intensifying the consumption of cereals. A terrestrial diet based on C3 plants typical of the ‘Neolithic package’ (wheat and barley) was demonstrated for southern communities, while in central-northern Italy, new crops were introduced (i.e. millet) during the Middle Bronze Age. Our data do not show dietary differentiations linked to social status or gender among the prehistoric farmers of southern Italy. Even though our results have highlighted some interesting dietary trends, new analyses including larger number of samples could provide a better characterization of the diets of prehistoric Italian communities and their variability in time and space.
References
Acsádi G, Nemeskéri J (1970) History of human life, span and mortality. Akadémiai Kiadó, Budapest
Alt KW, Zesch S, Garrido-Pena R, Knipper C, Szeacseanyi-Nagy A, Roth C, Tejedor-Rodríguez C, Held P, García-Martínez-de-Lagrán I, Navitainuck D, Arcusa Magallón H, Rojo-Guerra MA (2016) A community in life and death: the late Neolithic Megalithic tomb at Alto de Reinoso (Burgos, Spain). PLoS One 11(1):e0146176. https://doi.org/10.1371/journal.pone.0146176
Arena F, Gualdi-Russo E (2014) Taphonomy and post-depositional movements of a Bronze Age mass grave in the archaeological site of Grotta della Monaca (Calabria). 7th International Meeting on Taphonomy and Fossilization Ferrara, Sept.10-13th, 2014, Annali online dell’Università degli Studi di Ferrara - Sezione di Fisica e Scienze della Terra,1:10 (http://annali.unife.it/FST/article/view/919/815). Accessed 10 Jan 2020
Arena F, Larocca F, Onisto N, Gualdi Russo E (2014) Il sepolcreto protostorico di Grotta della Monaca in Calabria. Aspetti antropologici. XX Congresso AAI Ferrara Sept.11-13th, 2013. Annali online dell’Università degli Studi di Ferrara - Museologia Scientifica e Naturalistica 10(2):74–80 (http://annali.unife.it/museologia/issue/viewIssue/200/33). Accessed 10 Jan 2020
Arrighi S, Benvenuti M, Crezzini J, Gonnelli T, Mariotti-Lippi M, Milanesi C, Moroni Lanfredini A (2007) L’abitato della media età del Bronzo di Gorgo del Ciliegio (Sansepolcro-AR). Dati preliminari sul contesto paleoambientale Rivista di Scienze Preistoriche 57:263–276
Battisti M, Marconi S (2004) La fauna dell'insediamento dei Pizzini di Castellano (TN) e l'allevamento nell'Italia nord orientale nel corso dell'antica età del Bronzo. Padusa - Bollettino del Centro Polesano di Studi Storici Archeologici ed Etnografici Rovigo 39(2003):45–59
Bietti Sestieri AM (2011) L’Italia nell’età del Bronzo e del Ferro. Carocci editore, Roma
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Bökönyi S, Siracusano G (1987) Reperti faunistici dell’età del Bronzo del sito di Coppa Nevigata: un commento preliminare. In: Cassano SM, Cazzella A, Manfredinio A, Moscoloni M (eds) Coppa Nevigata e il suo Territorio. Edizioni Quasar, Roma, pp 205–210
Borgognini Tarli SM (1992) Aspetti antropologici e paleodemografici dal Paleolitico superiore alla prima età del Ferro. In: GUIDI A, PIPERNO M (eds) Italia Preistorica. Laterza, Bari, pp 239–273
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51(1):337–360
Brothwell DR (1981) Digging Up Bones. Oxford University Press, Oxford
Brown TA, Nelson DE, Vogel JS, Southon JR (1988) Improved collagen extraction by modified Longin method. Radiocarbon 30:171–177
Buikstra JE, Ubelaker DH (1994) Standards for data collection from human skeletal remains. Arkansas Archaeological Survey Research Series n. 44, Fayetteville
Caldara M, Cazzella A, Fiorentino G, Lopez R, Magri D, Simone O (1999) Primi risultati di una ricerca paleoambientale nell'area di Coppa Nevigata (Foggia). Atti del 19° Convegno Nazionale sulla Preistoria, Protostoria e Storia della Daunia, Stab. Lit. Centrografico Francescano, Foggia, pp.199-236
Castelletti I, Castiglioni F, Rottoli M (2001) L’agricoltura nell’Italia settentrionale dal Neolitico al Medioevo. In: Falla O, Forni G (Eds) Le piante coltivate e la loro storia. Dalle origini al transgenico in Lombardia nel centenario della riscoperta della genetica di Mendel, Atti del convegno organizzato dal Museo Lombardo di Storia dell’Agricoltura (1999), Milano, pp. 33-84
Castiglioni E, Motella De Carlo S, Nisbet R (1998) Indagini sui resti vegetali macroscopici a Canàr. In: Balista C, Bellintani P (Eds.) Canàr di San Pietro Polesine. Ricerche archeo-ambientali sul sito palafitticolo. Padusa Quaderni 2: 115–130
Cavazzuti C, Skeates R, Millard AR, Nowell G, Peterkin J, Bernabò Brea M, Cardarelli A, Salzani L (2019) Flows of people in villages and large centres in Bronze Age Italy through strontium and oxygen isotopes. PLoS One 14(1):e0209693. https://doi.org/10.1371/journal.pone.0209693
Cipolloni Sampò M (1986a) La Tomba 3 dell’Acropoli di Toppo Daguzzo (Potenza). Elementi per uno studio preliminare. Annali dell’Istituto Orientale, Archeologia e Storia Antica 8:1–40
Cipolloni Sampò M (1986b) La tomba di Toppo Daguzzo (Basilicata nord-orientale). Considerazioni sulle comunità della media età del Bronzo nel sud-est italiano. In: Marazzi M, Tusa S, Vagnetti L (eds) Traffici micenei nel Mediterraneo. Problemi storici e documentazione archeologica, Atti Convegno Palermo, pp 27–40
Craig OE, Biazzo M, Tafuri MA (2006) Paleodietary records of coastal Mediterranean populations. J Mediter Stud 16:63–77
Cremaschi M, Mercuri AM, Torri P, Florenzano A, Pizzi C (2016) Climate change versus land management in the Po Plain (Northern Italy) during the Bronze Age: new insights from the VP/VG sequence of the Terramara Santa Rosa di Poviglio. Quat Sci Rev 136:153–172
Cubas M, Peyroteo-Stjerna R, Fontanals-Coll M, Llorente-Rodriguez L, Lucquin A, Craig OE, Colonese AC (2019) Long-term dietary change in Atlantic and Mediterranean Iberia with the introduction of agriculture: a stable isotope perspective. Archaeol Anthropol Sci 11:3825–3836
De Angelis F, Scorrano G, Martinez-Labarga C, Giustini F, Brilli M, Pacciani E, Silvestrini M, Calattini M, Volante N, Martini F, Sarti L, Rickards O (2019) Eneolithic subsistence economy in Central Italy: first dietary reconstructions through stable isotopes. Archaeol Anthropol Sci 11:4171–4186
De Grossi MJ (2013) Considerazioni sullo sfruttamento animale in ambito terramaricolo. Economia e ambiente nell’Italia padana dell’età del Bronzo. Le indagini bioarcheologiche. In: De Grossi MJ, Curci A, Giacobini G (eds) Beni Archeologici- Conoscenza e Tecnologie, Quaderno 11. Edipuglia, Bari, pp 257–263
De Grossi MJ, Riedel A, Tagliacozzo A (2004) L’evoluzione delle popolazioni animali e dell’economia nell’età del Bronzo recente. In: Cocchi Genick D (ed) L’età del Bronzo Recente in Italia: Atti del Congresso Nazionale di Lido di Camaiore, 26-28th October, 2000. Baroni, Viareggio, pp 227–232
Demoulin F (1972) Importance des Certaines Mesures Craniennes (en Particulier de la Longueur Sagittale de la Mastoid) dans la Détermination Sexuelle des Cranes. Bull et Mém de la Soc d’Anthrop de Paris 9:259–264
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317:806–809
DeNiro MJ (1987) Stable isotope and archaeology. Am Sci 75:182–191
DeNiro MJ, Epstein S (1978) Influence of the diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta 42:495–506
DeNiro MJ, Epstein S (1981) Influence of the diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta 45:341–351
Dittrick J, Suchey JM (1986) Sex determination of prehistoric central California skeletal remains using discriminant analysis of the femur and humerus. Am J Phys Anthropol 70:3–9
Dunne J, Rebay-Salisbury K, Salisbury RB, Frisch A, Walton-Doyle C, Evershed RP (2019) Milk of ruminants in ceramic baby bottles from prehistoric child graves. Nature 574:246–248
Ferembach D, Schwidetzky I, Stloukal M (1977–1979) Raccomandazioni per la determinazione dell’età e del sesso sullo scheletro. Rivista di Antropologia 60:5–51
Ferembach D, Schwidetzky I, Stloukal M (1980) Recommendations for age and sex diagnoses of skeletons. J Hum Evol 9:517–549
Fernandez-Crespo T, Schulting RJ (2017) Living different lives: early social differentiation identified through linking mortuary and isotopic variability in Late Neolithic/ Early Chalcolithic north-central Spain. PLoS One 12(9):e0177881. https://doi.org/10.1371/journal.pone.0177881
Fontanals-Coll M, Subirà ME, Dìaz-Zorita M, Duboscq S, Gibaja JF (2015) Investigating palaeodietary and social differences between two differentiated sectors of a Neolithic community, La Bòbila Madurell-Can Gambùs (north-east Iberian Peninsula). J Archaeol Sci Rep 3:160–170
France DL (1998) Observational and metric analysis of sex in the skeleton. In: Reichs KJ (ed) Forensic osteology: advances in the identification of human remains. Charles C. Thomas Publisher, Springfield, pp 163–186
France DL, Horn AD (1988) Lab manual and workbook for physical anthropology. West Publishing Company, San Francisco
Gamarra B, Howcroft R, McCall A, Dani J, Hajdù Z, Nagy EG, Szabò LD, Domboròczki L, Pap I, RaczkyP MA, Zoffmann ZK, Hajdu T, Feeney RNM, Pinhasi R (2018) 5000 years of dietary variations of prehistoric farmers in the Great Hungarian Plain. PLoS One 13:e0197214. https://doi.org/10.1371/journal.pone.0197214
Giles E, Elliot O (1963) Sex determination by discriminant function analysis of the mandible. Am J Phys Anthropol 22:129–136
Goude G, Fontugne M (2016) Carbon and nitrogen isotopic variability in bone collagen during the Neolithic period: influence of environmental factors and diet. J Archaeol Sci 70:117–131
Goude G, Lisfranc R, Loison G, Forest V, Herrscher E (2011) The Bronze Age diet in Auvergne (France): a stable isotope approach. Am J Phys Anthropol 144(S52):195–196
Grifoni Cremonesi R (1976) Il neolitico e l’età dei metalli. In: Soprintendenza Archeologica della Basilicata (Ed) Il Museo Nazionale Ridola di Matera. Ed. Meta, Matera, pp 21–31
Gualdi-Russo E (2007) Sex determination from the talus and calcaneus measurements. Forensic Sci Int 171:151–156
Guilaine J (2003) Art et symboles du Néolithique à la Protohistoire. Errance, Paris
Harding AF, Fokkens H (2013) The Oxford handbook of the European Bronze Age. Oxford University Press, Oxford
Hedges R, Reynard L (2007) Nitrogen isotopes and the trophic level of humans in archaeology. J Archaeol Sci 34:1240–1251
Herrscher E (2003) Alimentation d'une population historique. Analyse des données isotopiques de la nécropole Saint-Laurent de Grenoble (XIII-XV siècles, France). Bull Mem Soc Anthropol Paris 15:149–269
Herrscher E, Goude G, Lisfranc R, Forest V, Vermeulen C (2013) Pratiques alimentaires au Bronze ancien en Auvergne: Essai de restitution par l’analyse isotopique. Préhistoires Méditerranéennes 4: http://pm.revues.org/782. Accessed 9 Jan 2020
Higham TFG, Jacobi RM, Bronk Ramsey C (2006) AMS radiocarbon datings of ancient bone using ultrafiltration. Radiocarbon 48:179–195
Iordanidis P (1961) Determination du sexe par les os du squelette (atlas, axis, clavicule, omoplate, sternum). Annales de Medecine legale 41:280–291
Ippolito F (2015–16). Late Eneolithic and Early Bronze Age funerary evidence from the Sant’Angelo IV cave (Northeastern Calabria, Italy). Palaeohistoria, University of Groningen Press, 57/58: 111-116
Jacob J, Disnar JR, Arnaud F, Chapron E, Debret M, Lallier-Vergès E, Desmet M, Revel-Rolland M (2008) Millet cultivation history in the French Alps as evidenced by a sedimentary molecule. J Archaeol Sci 35:814–820
Jørkov MLS, Heinemeier J, Lynnerup N (2007) Evaluating bone collagen extraction methods for stable isotope analysis in dietary studies. J Archaeol Sci 34:1824–1829
Lai L, Tykot R, Usai E, Beckett JF, Floris R, Fonzo O, Goddard E, Hollander D, Manunza MR, Usai A (2013) Diet in the Sardinian Bronze Age: models, collagen isotopic data, issues and perspectives. Préhistoires Méditerranéennes 4 https://journals.openedition.org/pm/795?lang=en
Larocca F (2005) La miniera pre-protostorica di Grotta della Monaca (Sant’Agata di Esaro - Cosenza). Centro Regionale di Speleologia “Enzo dei Medici”, Roseto Capo Spulico (CS)
Le Bras-Goude G, Binder D, Formicola V, Duday H, Couture-Veschambre C, Hublin J-J, Richards M (2006) Stratégies desubsistance et analyse culturelle de populations néolithiques de Ligurie: approche par l’étude isotopique (δ13C et δ15N) des restes osseux. Bull Mém Soc Anthropol Paris 18:45–55
Lee-Thorp JA (2008) On isotopes and old bones. Archaeometry 50:925–950
Lelli R, Allen R, Biondi G, Calattini M, Conati Barbaro C, Gorgoglione MA, Manfredini A, Martinez-Labarga C, Radina F, Silvestrini M, Tozzi C, Rickards O, Craig O (2012) Examining dietary variability of the earliest farmers of south-eastern Italy. Am J Phys Anthropol 149:380–390
Lightfoot E, Boneva B, Miracle PT, Slaus M, O’Connell TC (2011) Exploring the Mesolithic and Neolithic transition in Croatia through isotopic investigations. Antiquity 85:73–86
Lo Porto FG (1988) Testimonianze archeologiche di culti metapontini. Xenia 16:5–28
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature 230:241–242
Lopez-Costas O, Müldner G, Martínez Cortizas A (2015) Diet and lifestyle in Bronze Age Northwest Spain: the collective burial of Cova do Santo. J Archaeol Sci 55:209–218
Lovejoy CO, Meindl RS, Mensforth RP, Barton TJ (1985) Multifactorial determination of skeletal age at death: a method with blind tests of its accuracy. Am J Phys Anthropol 68:1–14
Lubell D, Jackes M, Schwarcz H, Knyf M, Meiklejohn C (1994) The Mesolithic-Neolithic transition in Portugal: isotopic and dental evidence of diet. J Archaeol Sci 21:201–216
Maini E, Curci A (2013) Considerazioni sull’economia di allevamento nella Romagna durante l’età del bronzo. Economia e ambiente nell’Italia padana dell’età del bronzo. Le indagini bioarcheologiche. In: De Grossi MJ, Curci A, Giacobini G (eds) Beni Archeologici - Conoscenza e Tecnologie, Quaderno 11. Edipuglia, Bari, pp 357–376
Mannino MA (2009) Analisi isotopiche sui resti umani. In: Mallegni F, Lippi B (eds) Non omnis moriar. Edizioni CISU, Rome, pp 395–424
Mannino MA, Catalano G, Talamo S, Mannino G, Di Salvo R, Schimmenti V, Lalueza-Fox C, Messina A, Petruso D, Caramelli D, Richards MP, Sineo L (2012) Origin and diet of the prehistoric hunter-gatherers on the Mediterranean Island of Favignana (Egadi Islands, Sicily). PLoS One 7(11):e49802. https://doi.org/10.1371/journal.pone.0049802
Mannino MA, Talamo S, Tagliacozzo A, Fiore I, Nehlich O, Piperno M, Tusa S, Collina C, Di Salvo R, Schimmenti V, Richards MP (2015) Climate-driven environmental changes around 8,200 years ago favoured increases in cetacean strandings and Mediterranean hunter-gatherers exploited them. Sci Rep 5:16288. https://doi.org/10.1038/srep16288
Masotti S, Varalli A, Goude G, Moggi-Cecchi J, Gualdi-Russo E (2017) A combined analysis of dietary habits in the Bronze Age site of Ballabio (northern Italy). Archaeol Anthropol Sci 11:1029–1047
Matarese I (2014) Gli ornamenti della tomba 1 di Murgia Timone (Matera) nel quadro del Bronzo medio in Italia. Atti XII Incontro di Studi “Preistoria e Protostoria in Etruria: Ornarsi per comunicare agli uomini e agli dei”, 12-14th Sept.2014, Manciano-Pitigliano-Valentano
Mazzei M, Tunzi Sisto A (2005) Gargano antico: testimonianze archeologiche dalla preistoria al tardoantico. Claudio Grenzi Ed, Foggia
McClure SB, Garcia García O, Roca de Togores C, Culleton BJ, Kennett DJ (2011) Osteological and paleodietary investigation of burials from Cova de la Pastora, Alicante, Spain. J Archaeol Sci 38:420–428
Meindl RS, Lovejoy CO (1985) Ectocranial suture closure: a revised method for the determination of skeletal age at death based on the lateral anterior sutures. Am J Phys Anthropol 68:57–66
Mercuri AM, Baroni R, Mariotti Lippi M (2006) Workshop “Archeobotanica e alimentazione”, 18 Dec, 2006, Florence. Atti della Societàdei Naturalisti e dei Matematici di Modena 137:240–456
Olivier G, Pineau H (1958) Détermination de l’age de foetus et de l’embryon. Archives d’Anatomie Pathologique 6:21–28
Olsen J, Tikhomirov D, Grosen C, Heinemeier J, Klein M (2016) Radiocarbon analysis on the new AARAMS 1MV Tandetron. Radiocarbon 59:1–9
Pals JP, Vorrips A (1979) Seeds, fruits and charcoals from two prehistoric sites in Northern Italy. Archaeo-Physica 8:217–235
Papathanasiou A (2003) Stable isotope analysis in Neolithic Greece and possible implications on human health. Int J Osteoarchaeol 13:314–324
Patroni G (1897) Matera, antichità preistoriche. Notizie degli Scavi di Antichità, pp 203–211
Perini R (1987) Scavi archeologici nella zona palafitticola di Fiavé- Carera II, Campagne 1969-1976. Resti della cultura materiale, metallo-osso-litica-legno. Patrimonio storico e artistico del Trentino 9, Servizio Beni Culturali della Provincia autonoma di Trento, Trento
Peroni R (2004) L'Italia alle soglie della Storia. Edizioni Laterza, Bari
Pessina A, Tiné V (2008) Le ceramiche impresse nel Neolitico antico. Italia e Mediterraneo. Ministero per i Beni e le Attività Culturali. Istituto Poligrafico e Zecca dello Stato, Roma
Piquet MM (1956) Etude de la Robustesse de la Mandibule. Bull Mém Soc Anthrop de Paris 7:204–224
Primavera M, Fiorentino G (2014) Acorn gatherers: fruit storage and processing in south-East Italy during the Bronze Age. Origini, Preistoria e protostoria delle civiltà antiche 35:153–168
Quagliati P (1896) Appunti delle scoperte paletnologiche di Domenico Ridola nel materano. Bullettino di Paletnologia Italiana 22:287–289
Reimer PJ, Bard E, Bayliss A, Beck JW, Blackwell PG, Bronk Ramsey C, Buck CE, Cheng H, Edwards RL, Friedrich M, Grootes PM, Guilderson TP, Haflidason H, Hajdas I, Hatté C, Heaton TJ, Hoffmann DL, Hogg AG, Hughen KA, Kaiser KF, Kromer B, Manning SW, Niu M, Reimer RW, Richards DA, Scott EM, Southon JR, Staff RA, Turney CSM, van der Plicht J (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55(4):1869–1887
Reitsema LJ (2013) Beyond diet reconstruction: stable isotope applications to human physiology, health and nutrition. Am J Hum Biol 25:445–456
Richards MP, Hedges REM (1999) Stable isotope evidence for similarities in the types of marine foods used by Late Mesolithic humans at sites along the Atlantic coast of Europe. J Archaeol Sci 26:717–722
Richards MP, Price TD, Koch E (2003) Mesolithic and Neolithic subsistence in Denmark: new stable isotope data. Curr Anthropol 44:288–294
Ridola D (1901) La Paletnologia del Materano. Bullettino di Paletnologia Italiana 27:27–41
Ridola D (1912) La Grotta dei Pipistrelli e la Grotta Funeraria. Conti Ed, Matera
Riedel A (1996) Archaeozoological investigations in North-eastern Italy: the exploitation of animals since the Neolithic. Preistoria Alpina 30:43–94
Rösch M (1998) The history of crops and crop weeds in south-western Germany from the Neolithic period to modern times, as shown by archaeobotanical evidence. Veget Hist Archaeobot 7:109–125
Rumolo A, Forstenpointner G, Rumolo P, Jung R (2020) Palaeodiet reconstruction inferred by stable isotopes analysis of faunal and human remains at Bronze Age Punta di Zambrone (Calabria, Italy). Int J Osteoarchaeol 30:90–98
Salazar-García DC, Benítez De Lugo Enrich LB, Álvarez García HJ, Benito Sánchez M (2013) Estudio diacrónico de la dieta de los pobladores antiguos de Terrinches (Ciudad Real) a partir del análisis de isótopos estables sobre restos óseos humanos. Rev Esp Antrop Fís 34:6–14
Salvadei L, Santandrea E (2003) Condizioni di vita e stato di salute nel campione neolitico di Masseria Candelaro (FG). Atti della XXXV Riunione Scientifica dell’Istituto Italiano di Preistoria e Protostoria, Firenze 35:829–834
Santos GM, Mazon M, Southon JR, Rifai S, Moore R (2007) Evaluation of iron and cobalt powders as catalysts for 14C-AMS target preparation. Nucl Instrum Methods Phys Res, Sect B 259(1):308–315
Scattarella V, Sublimi Saponetti S, Emanuel P (2005) Il Sepolcreto Ipogeo: Aspetti Antropologici. In: Larocca F (Ed) La Miniera Pre-Protostorica di Grotta della Monaca (Sant’Agata di Esaro - Cosenza), Centro Regionale di Speleologia “Enzo dei Medici”, Roseto Capo Spulico (CS), pp. 67-71
Schoeninger MJ (1979) Diet and status at Chalcatzingo: some empirical and technical aspects of strontium analysis. Am J Phys Anthropol 51:295–310
Schoeninger MJ (2011) Diet reconstruction and ecology using stable isotope ratios. In: Larsen CS (ed) A companion to biological anthropology. Wiley-Blackwell, Chichester
Schoeninger MJ, DeNiro MJ, Tauber H (1983) Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220:1381–1383
Schulting RJ (1998) Slighting the sea: stable isotope evidence for the transition to farming in northwestern Europe. Documenta Praehistorica 25:203–218
Skeates R (2000) The social dynamics of enclosure in the Neolithic of the Tavoliere, South-East Italy. J Mediterr Archaeol 13:155–188
Starnini E (2002) La ceramica Impressa dell’Adriatico orientale e dei Balcani. In: Fugazzola Delpino MA, Pessina A, Tiné V (eds) Le ceramiche impresse nel Neolitico antico. Italia e Mediterraneo. Ministero per i Beni e le Attività Culturali. Istituto Poligrafico e Zecca dello Stato, Roma, pp 29–35
Stuiver M, Polach HA (1977) Reporting of C-14 data. Radiocarbon 19(3):355–363
Tafuri MA, Craig OE, Canci A (2009) Stable isotope evidence for the consumption of millet and other plants in Bronze Age Italy. Am J Phys Anthropol 139:146–153
Tagliacozzo A (1994) I dati archeologici. In: Peroni R, Trucco F (eds) Enotri e Micenei nella Sibaritide. Ed. ISAMG, Taranto, pp 587–652
Tauber H (1981) 13C evidence for dietary habits of prehistoric man in Denmark. Nature 292:332–333
Treasure ER, Church MJ, Gröcke DR (2016) The influence of manuring on stable isotopes (δ13C and δ15N) in Celtic bean (Vicia faba L.): archaeobotanical and palaeodietary implications. Archaeol Anthropol Sci 8:555–562
Triantaphyllou S, Richards MP, Zerner C, Voutsaki S (2008) Isotopic dietary reconstruction of humans from Μiddle Bronze Age Lerna, Argolid, Greece. J Archaeol Sci 35:3028–3034
Tunzi Sisto AM (1997) Ipogei della Daunia. Culti e riti funerari della media età del Bronzo, Catalogo della mostra, Comunità Montana del Gargano, Foggia
Tunzi Sisto AM (1999) Ipogeo dei Bronzi. In: Tunzi Sisto AM (Ed) Ipogei della Daunia. Preistoria di un territorio, Banca del Monte, Foggia, pp 184-216
Tunzi Sisto AM (2005) Gli Avori di Trinitapoli. In: Vagnetti L, Bettelli M, Damiani I (eds) L’avorio in Italia nell’età del Bronzo. CNR-ISMA, Roma, pp 47–58
Valsecchi V, Tinner W, Finsinger W, Ammann B (2006) Human impact during the Bronze Age on the vegetation at Lago Lucone (Northern Italy). Veg Hist Archaeobotany 15:99–113
Van Klinken GJ (1999) Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J Archaeol Sci 26:687–695
Vanzetti A (1999) Combinazioni di corredo delle sepolture all’interno dell’Ipogeo dei Bronzi di Trinitapoli. In: Tunzi Sisto AM (Ed) Ipogei della Daunia. Preistoria di un territorio, Banca del Monte, Foggia, pp 222-226
Varalli A, Moggi-Cecchi J, Moroni A, Goude G (2016a) Dietary variability during Bronze Age in Central Italy: first results. Int J Osteoarchaeol 26:431–446
Varalli A, Moggi-Cecchi J, Dori I, Bocconi S, Bortoluzzi S, Salzani P, Tafuri MA (2016b) Dietary continuity vs. discontinuity in Bronze Age Italy. The isotopic evidence from Arano di Cellore (Illasi, Verona, Italy). J Archaeol Sci Rep 7:104–113
Vogel JS, Southon JR, Nelson DE, Brown TA (1984) Performance of catalytical condensed carbon for use in accelerator mass spectrometry. Nuclear Instruments & Methods in Physics Research Section B-Beam Interactions with Materials and Atoms 5(2):289–293
Waterman AJ, Tykot RH, Silva AM (2016) Stable isotope analysis of diet-based social differentiation at Late Prehistoric collective burials in South-Western Portugal. Archaeometry 58:131–151
Zilhão J (2001) Radiocarbon evidence for maritime pioneer colonization at the origins of farming in west Mediterranean Europe. Proc Natl Acad Sci 98:14180–14185. https://doi.org/10.1073/pnas.241522898
Acknowledgements
We are grateful to the archaeological superintendencies of Calabria and Basilicata, the Museo Archeologico Nazionale della Sibaritide (Sibari, Cosenza), the Centro Regionale di Speleologia ‘Enzo dei Medici’ (Roseto Capo Spulico, Cosenza) and the Museo Archeologico Nazionale di Potenza (Potenza) for providing the skeletal remains used in this study. Furthermore, we thank Damiano Marchi (University of Pisa, Department of Biology) for allowing us to study the remains from Ipogeo dei Bronzi. We are also indebted to the archaeologists and conservators of the institutions that hosted us, especially to F. Larocca, A. Patrone, I. Matarese and M. Savarese, for their support and cooperation. We would like to thank the staff of the Aarhus AMS Centre for teaching and helping Fabiola Arena with the sample preparation in the laboratory.
Funding
Open access funding provided by Università degli Studi di Ferrara within the CRUI-CARE Agreement. The isotope analyses and AMS radiocarbon dating were funded by the Aarhus University Research Foundation (Denmark) through the project titled ‘Danish and European Diets in Time’ awarded to Marcello A. Mannino (project number: AUFF-E-2015-FLS-8-2). This work was also supported by a grant from the University of Ferrara (Italy).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Arena, F., Gualdi-Russo, E., Olsen, J. et al. New data on agro-pastoral diets in southern Italy from the Neolithic to the Bronze Age. Archaeol Anthropol Sci 12, 245 (2020). https://doi.org/10.1007/s12520-020-01209-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-020-01209-9