
Abstract
This paper examines the impact of animal manure on δ15N and δ13C values in a legume, Celtic Black broad bean (Vicia faba). In a field experiment, V. faba was cultivated in plots treated with farmyard manure and pure sheep manure. The results indicate that highly intensive manuring can increase δ15N values in beans, stems, leaves and pods. In comparison, manuring had a relatively small impact on δ13C values. In terms of palaeodietary reconstructions, the high δ15N values in very intensively manured beans (+3 ‰) are equivalent to the trophic-level effect. Based on the experimental results, it is suggested that high δ15N values in archaeobotanical remains of V. faba may be attributable to small-scale cultivation with intensive manuring.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Stable isotope analysis is becoming increasingly widespread in archaeobotanical research as a powerful method for investigating agricultural practices and land use patterns (Aguilera et al. 2008; Bogaard et al. 2013; Fraser et al. 2013; Heaton et al. 2009; Kanstrup et al. 2014; Masi et al. 2014; Vaiglova et al. 2014). Plant stable carbon (δ13C) and nitrogen (δ15N) isotope ratios vary in relation to a range of environmental factors (Dawson et al. 2002) including agricultural practices, and these variations can be analysed in archaeobotanical remains (Fiorentino et al. 2015; Szpak 2014). In particular, recent research has been directed towards the identification of manuring in cereals and pulses (Bogaard et al. 2007, 2013; Fraser et al. 2011; Kanstrup et al. 2011, 2014). Since manuring is closely linked to intensive land use patterns, it is an important area of research into past crop husbandry regimes (Bogaard 2012).
Field experiments and farm studies indicate that animal manuring increases cereal δ15N values (≥6 ‰) (Bogaard et al. 2007; Bol et al. 2005; Fraser et al. 2011; Kanstrup et al. 2011). In comparison, manuring has minimal effect on δ15N values in legumes, except where very intensively applied over a long duration (Fraser et al. 2011; Styring et al. 2014). There is a requirement to further examine the relationship between manuring intensity and legume δ15N values (Styring et al. 2014). In terms of palaeodietary reconstructions, it is important to assess the impact of manuring on legume δ15N values as a source of 15N enrichment in animal and human tissues (Bogaard et al. 2007; Fraser et al. 2011; Hedges and Reynard 2007).
Plant δ13C values have been primarily applied to investigate crop water management practices (Fiorentino et al. 2015). However, other environmental factors that alter δ13C need to be considered (Stokes et al. 2011). The relationship between plant δ13C values and manuring is not clearly understood with both positive and negative shifts observed in δ13C (Kanstrup et al. 2011; Senbayram et al. 2008; Szpak et al. 2012b; Wallace et al. 2013).
The aim of this study is to analyse the influence of manuring on δ13C and δ15N values in the legume, Celtic Black broad bean (Vicia faba L.). V. faba is frequently recovered in archaeobotanical assemblages across the Near East and Europe from the Neolithic and Bronze Age onwards (see Colledge and Conolly 2007; Stika and Heiss 2013). A recent study identified extensive evidence for V. faba in Britain in the Neolithic-Iron Age, totalling over 70 archaeobotanical assemblages containing Celtic Bean, and suggested that, in some areas, it may have been cultivated on a small ‘garden’ scale in intensively managed plots treated with animal manure (Treasure 2014). In order to accurately assess the relationship between δ13C and δ15N values in V. faba and manuring, it is necessary to undertake field experiments where the rates of manure application can be quantified.
Methodological background
Nitrogen isotopes (δ15N)
Plant δ15N values reflect the net effect of a range of factors, including the form of nitrogen acquired (NH4 +, NO3 −, N2) and the method of nitrogen assimilation (uptake of soil nitrogen, fixation of atmospheric N2) (Evans 2001; Högberg 1997).
Pulses are harvested from leguminous plants which can assimilate nitrogen through fixation of atmospheric N2 via symbiotic bacteria (rhizobia) in the roots (Franche et al. 2009; Howard and Rees 1996). As N2 fixation involves minimal fractionation, legumes dependent on fixation as a source of nitrogen have δ15N values typically around 0 ‰, reflecting atmospheric N2 (i.e., δ15Nair = 0 ‰) (Kohl and Shearer 1980; Shearer and Kohl 1986; Virginia and Delwiche 1982). A range of factors, however, can influence N2 fixation, in particular, soil nitrogen availability (Liu et al. 2011). In soil N-rich environments, N2 fixation is inhibited and legumes preferentially take up nitrogen from the soil (Ledgard et al. 1996; Peoples et al. 2009; Vinther 1998). The nitrogen isotopic composition of legumes taking up nitrogen from soil (rather than N2 fixation) will reflect the δ15N value of the soil nitrogen (Andrews et al. 2011). As manuring increases soil mineral nitrogen and δ15N values (Choi et al. 2003; Simpson et al. 1999; Watzka et al. 2006), it has the potential to increase δ15N values in legumes above 0 ‰.
In cereals (non-N2-fixing plants), animal manure significantly increases δ15N values (up to +10 ‰) due to the uptake of 15N-enriched soil (Bogaard et al. 2007; Bol et al. 2005; Fraser et al. 2011; Kanstrup et al. 2011, 2012). In comparison, a recent study suggests that only very intensive animal manure application, in excess of >20–35 t/ha, alters δ15N in V. faba due to the preferential fixation of atmospheric N2 (Fraser et al. 2011; Styring et al. 2014). The largest increase in V. faba δ15N values (+2.2 ± 1.4 ‰) was observed in a farm study in Evvia, Greece, where very intensive manuring creates artificial ‘dung-soil’ (Fraser et al. 2011). However, the rate of manure application (t/ha) was not measured in the Evvia farm study.
Carbon isotopes (δ13C)
During photosynthesis, C3 plants (cereals, legumes) discriminate against 13C due to the preferential use of 12C by the enzyme RuBisCO during carbon fixation (Lloyd and Farquhar 1994; O’Leary 1981). The amount of 13C discrimination is closely linked with stomatal conductance (intrinsic water use efficiency) (Farquhar and Sharkey 1982; Farquhar et al. 1989; O’Leary 1988). Restricted water availability increases stomatal closure and reduces 13C discrimination (Chaves 1991; Farquhar et al. 1989). In comparison, during high water availability, stomata are open, increasing 13C discrimination (Chaves 1991; Farquhar et al. 1989). Studies have identified a causal link between δ13C values in plants and water availability in greenhouse and farm studies (Araus et al. 1997; Farquhar and Richards 1984; Flohr et al. 2011; Wallace et al. 2013).
However, a wide range of other environmental factors can alter δ13C values in plants including salinity, light intensity, temperature and nitrogen availability (Condon et al. 1992; Gröcke 1998, 2002; Heaton 1999; O’Leary 1981; Tieszen 1991). The relationship between plant δ13C values and manuring is complex with both positive and negative relationships observed (Kanstrup et al. 2011; Senbayram et al. 2008; Szpak et al. 2012b; Wallace et al. 2013). At present, there is no published data which specifically focuses on the impact of manuring on δ13C values in V. faba.
Materials and methods
Experimental design
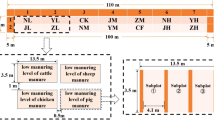
Celtic Black broad beans (V. faba L.) were cultivated in three 1-m2 outdoor plots at Durham University Botanic Gardens between May and September 2013. The Celtic Black broad beans used are a heritage variety which produces small-rounded seeds which are morphologically similar to prehistoric finds of V. faba. One plot acted as a control, and two plots were treated with manure and decomposed leaf litter (Table 1). All the available plants were harvested and the plant height, number of ripe/un-ripe pods, number of beans and dimensions of each bean were recorded.
One plant from each plot was randomly selected for isotopic analysis and air-dried. Samples of bean cotyledons, bean testae, pods, leaves and stem were analysed. All of the pods and beans available for each selected plant were analysed. The cotyledon was sampled separately, as the testa is rarely preserved in archaeobotanical remains of V. faba. The position of each pod on the plant and each bean within individual pods was recorded. This detailed sampling methodology enables analysis of within-plant δ13C and δ15N variation. Dried soil and manure samples were sieved at <1 mm. All samples were homogenised in an agate pestle and mortar.
Stable isotope analysis
Stable isotope measurements were performed at Durham University using a Costech Elemental Analyser (ECS 4010) coupled to a Thermo Finnigan Delta V Advantage isotope mass spectrometer. Carbon isotope ratios are Craig-corrected for 17O contribution and reported in standard delta (δ) notation in per mil (‰) relative to VPDB. Nitrogen isotope ratios are reported relative to AIR. Data accuracy is monitored through routine analyses of in-house standards, which are stringently calibrated against international standards (e.g., USGS 40, USGS 24, IAEA 600, IAEA N1, IAEA N2). Analytical uncertainty for δ13C and δ15N measurements is typically ±0.1 ‰ for replicate analyses of the international standards and typically <0.2 ‰ on replicate sample analysis. Total organic carbon and nitrogen data was obtained as part of the isotopic analysis using an internal standard (i.e., glutamic acid, 40.82 % C and 9.52 % N).
Results
Biomass analysis
The results of the biomass analysis are presented in Table 2. Six plants from the midden plot suffered insect damage and did not produce any pods. Plants in the amended plot were taller than in the control plot. In comparison, the heights of plants in the control and midden plots are identical (though this may be due to insect damage in the midden plants). Pod and bean yield was highest in plants in the amended plot. The comparative results for the midden plot are significantly lower, though, as noted above, this may be due to insect damage. Bean dimensions increased (particularly length) in the amended and midden plots.
Stable isotope analyses
Mean δ13C and δ15N values for the samples analysed are presented in Table 3. Figure 1 presents the δ13C and δ15N values for all the plant and soil samples analysed. The supplementary data includes the results for each sample in addition to C/N atomic ratio, %C and %N results.

δ15N and δ13C values for each sample (excluding the manure samples). Note the different scale on the soil graph
Soil and manure analyses
Mean soil δ15N values in the amended (5.5 ± 0.4 ‰) and midden (8.1 ± 1.7 ‰) plots are significantly higher than in the control plot (4.6 ± 0.2 ‰). Mean δ15N values for the farmyard manure were 7.7 ± 0.3 ‰ and, for the sheep manure, 7.5 ± 0.2 ‰. In the amended and midden plots, mean δ13C values are lower than in the control plot.
Plant analyses
Mean δ15N values in the control samples varied between −1 and 0.7 ‰. Mean δ15N values in cotyledons were 1.5 ± 0.2 ‰ in the amended samples and 2.6 ± 0.3 ‰ midden samples. Pods, stems and leaves were 15N enriched in the amended and midden samples. There is small variation in δ15N values between cotyledons sampled from the same pod and plant. δ15N variation between leaves, stems and pods was typically ≤1 ‰, with the largest offset (1.7 ‰) between the midden pod and midden stem samples (see Electronic supplementary material). Testa δ15N values are significantly lower than cotyledon δ15N values. Manuring intensity and δ15N values are positively correlated.
Mean δ13C values in the amended and midden samples are similar to the control samples, with small increases observed in some samples of up to 1.7 ‰. Within-plant δ13C variation between leaves, stems and pods was minimal. There is only a small variation in δ13C values between cotyledons sampled from the same pod and plant.
Discussion
Soil and manure δ13C and δ15N variability
In agreement with previous studies, manuring increased soil δ15N values (Bol et al. 2005; Kanstrup et al. 2011; Senbayram et al. 2008; Watzka et al. 2006). The mean δ15N values for the farmyard manure (7.7 ± 0.3 ‰) and sheep manure (7.5 ± 0.2 ‰) are high for animal manure (cf. Szpak 2014). Mean δ13C values were lower in amended and midden plots as animal manure is 13C-depleted (Bol et al. 2005). Previous studies have observed decreases in δ13C values in manured soils (Gerzabek et al. 1997; Senbayram et al. 2008).
Plant δ13C variability
Plant δ13C values were minimally affected by manuring, displaying only small increases, typically around +1 ‰. These results are consistent with previous studies which indicate that manuring may be a source of small variation in 13C values (Senbayram et al. 2008; Szpak et al. 2012b; Wallace et al. 2013). Small increases in δ13C values in response to nitrogen fertilisation observed in previous studies may be related to increased plant biomass which can limit stomatal conductance causing less 13C discrimination (Jenkinson et al. 1995; Serret et al. 2008).
Plant δ15N variability
The results presented in this study indicate that very intensive animal manuring (>70 t/ha) can increase δ15N values (+3 ‰) in V. faba. There is little δ15N variation between plant tissues, with the exception of testa samples, comparable with results from previous studies (López-Bellido et al. 2010; Nebiyu et al. 2014). Control sample δ15N values varied between 0 ± 1 ‰ and are consistent with fixation of atmospheric N2 in V. faba (Fan et al. 2006; López-Bellido et al. 2011; Nebiyu et al. 2014; Tryderman et al. 2004; Unkovich 2013). In contrast, the elevated δ15N values in the amended and midden samples indicate preferential uptake of soil mineral nitrogen in comparison to atmospheric N2 fixation. The δ15N values in the midden plot are higher than in the amended plot due to the application of pure manure compared to farmyard manure (i.e., a mixture of straw and manure).
The results presented here agree with a previous study which indicated that only very intensive manuring can significantly alter δ15N values in V. faba due to the preferential fixation of atmospheric N2 (Fraser et al. 2011; Styring et al. 2014). Fraser et al. (2011) only observed a large increase in δ15N values for V. faba in a farm study at Evvia, Greece, where very intensive manuring (sheep/goat dung) creates an artificial dung-soil. The quantity of manure applied (t/ha) in the Evvia farm study could not be measured, though the soil conditions appear to be similar to the midden plot in this study.
The relationship between N2 fixation and animal manuring in legumes is an area which requires further research. It is not clear why δ15N values in legumes are only altered by very intensive manuring as it requires less energy to take up soil mineral nitrogen compared to fixing atmospheric N2 (Andrews et al. 2009). A possible explanation may be due to ability of certain varieties of V. faba to continue atmospheric N2 fixation in the presence of high soil mineral nitrogen (Köpke and Nemecek 2010). Recently, Szpak et al. (2014) observed large 15N enrichment (+16 ‰) in a legume (common garden bean, Phaseolus vulgaris L.) amended with seabird guano which has a very high δ15N value (> +20 ‰) compared with animal manure (Szpak et al. 2012a; Szpak 2014). This indicates that manures which are high in plant available nitrogen and have high δ15N values can significantly enrich 15N in legumes and suppress N2 fixation compared to animal manure (Szpak et al. 2014). In comparison, a recent study demonstrated that animal manure increased N2 fixation in peas (Pisum sativum L.; Jannoura et al. 2014).
Archaeological implications
The results of this study suggest that intensive manuring of V. faba may be identifiable in archaeobotanical remains using nitrogen isotope analysis (cf. Fraser et al. 2011). The high intensity of manuring required to effect the nitrogen isotopic composition of pulses indicates that pulse δ15N values can reflect the scale of cultivation. In recent farming contexts, intensity of manuring is closely correlated with the scale of cultivation, with smaller plots receiving intensive manure application (Bogaard et al. 2000; Jones 2005). In Evvia, Greece, V. faba is cultivated in small infield (garden) areas, some of which are very intensively manured, creating an artificial dung-soil (Jones et al. 1999). Similarly, in Asturias, Spain, V. faba is cultivated in small intensively manured plots that are rotated with cereals (Charles et al. 2002). It is suggested that high δ15N values in archaeobotanical remains of V. faba may indicate small-scale cultivation with very intensive manuring. The results of this study should be viewed as preliminary in character, and further research is currently ongoing to explore δ13C and δ15N variability between plants cultivated in the same plot. For example, Wallace et al. (2013) have demonstrated large variation in δ13C values in V. faba cultivated under similar conditions. Variation in plant isotope values is expected in traditional farming regimes where growing conditions can be variable (Wallace et al. 2013)
In terms of palaeodietary reconstructions, measurement of plant δ15N values is necessary in order to accurately reconstruct baseline data and interpret δ15N values in animal and human tissues (Casey and Post 2011; Hedges and Reynard 2007). In particular, Fraser et al. (2013) have demonstrated that the plant component of diets can be assessed with greater accuracy through direct measurement of archaeological plant δ15N values. The impact of manuring on cereal δ15N values (up to 10 ‰) is significantly higher than the Celtic Black broad beans analysed in this study. Despite this, the enrichment in δ15N between the control and the midden samples (~3 ‰) is equivalent to the trophic-level effect (3–5 ‰, Bocherens and Drucker 2003) and, hence, could subsequently cause 15N enrichment in animal and human tissues. This is particularly significant as palaeodietary studies typically consider legumes to have δ15N values around 0 ‰ (DeNiro and Epstein 1981; van Klinken et al. 2002).
In manured cereals, there is a large offset in δ15N values between the grain and chaff (2.4 ± 0.8 ‰), suggesting that the use of chaff for animal fodder and grain for human consumption will result in significantly different nitrogen isotopic signatures between animals and humans (Bogaard et al. 2007; Fraser et al. 2011). In comparison, the results of this study indicate a comparatively small offset in δ15N values of manured V. faba between different plant components. This is significant as ethnographic evidence indicates that V. faba was used as a source of fodder, either the seeds, chaff or whole plants (Forbes 1996; Halstead 2014).
Conclusion
The results of this study indicate that highly intensive animal manuring can increase δ15N values in legumes. Celtic Black broad beans (V. faba) displayed significantly higher δ15N values in intensively manured plots. In comparison, manure minimally affected δ13C values, indicating that manuring may only be a source of small variation in δ13C values. In terms of palaeodietary reconstructions, manuring increased δ15N values on a scale equivalent to a single step in trophic level. Based on the experimental results presented here, it is suggested that high δ15N values in archaeobotanical remains of V. faba should be attributed to small-scale cultivation with very intensive manuring.
References
Aguilera M, Araus JL, Voltas J, Rodríguez-Ariza MO, Molina F, Rovira N, Buxó R, Ferrio JP (2008) Stable carbon and nitrogen isotopes and quality traits of fossil cereal grains provide clues on sustainability at the beginnings of Mediterranean agriculture. Rapid Commun Mass Spectrom 22:1653–1663
Andrews M, Lea PJ, Raven JA, Azevedo RA (2009) Nitrogen use efficiency. 3. Nitrogen fixation: genes and costs. Ann Appl Bot 155:1–13
Andrews M, James EK, Sprent JI, Boddey RM, Gross E, Bueno dos Reis F (2011) Nitrogen fixation in legumes and actinorhizal plants in natural ecosystems: values obtained used 15N natural abundance. Plant Ecolog Divers 4:131–140
Araus JL, Febrero A, Buxó R, Rodríguez-Ariza MO, Molina F, Camalich MD, Martín D, Voltas J (1997) Identification of ancient irrigation practices based on the carbon isotope discrimination of plant seeds: a case study from the south-east Iberian Peninsula. J Archaeol Sci 24:729–740
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Bogaard A (2012) Middening and manuring in Neolithic Europe: issues of plausibility, intensity and archaeological method. In: Jones RL (ed) Manure matters: historical, archaeological and ethnographic perspectives. Ashgate, Farnham, pp 25–39
Bogaard A, Charles M, Haltead P, Jones G (2000) The scale and intensity of cultivation: evidence from weed ecology. In: Halstead P, Fredrick C (eds) Landscape and land use in postglacial Greece. Sheffield Academic Press, Sheffield, pp 129–134
Bogaard A, Heaton THE, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeobotanical implications for reconstruction of diet and crop management practices. J Archaeol Sci 34:335–343
Bogaard A, Fraser R, Heaton THE, Wallace M, Vaiglova P, Charles M, Jones G, Evershed RP, Styring AK, Andersen NH, Arbogast R-M, Bartosiewicz L, Gardeisen A, Kanstrup M, Maier U, Marinova E, Ninov L, Schäfer M, Stephan E (2013) Crop manuring and intensive land management by Europe’s first farmers. Proc Natl Acad Sci 110:12589–12594
Bol R, Eriksen J, Smith P, Garnett MH, Coleman K, Christensen BT (2005) The natural abundance of 13C, 15N, 34S, and 14C in archived (1923-2000) plant and soil samples from the Askov long-term experiments on animal manure and mineral fertilizer. Rapid Commun Mass Spectrom 19:3216–3226
Casey MM, Post DM (2011) The problem of isotopic baseline: reconstructing the diet and trophic position of fossil animals. Earth Sci Rev 106:131–148
Charles M, Bogaard A, Jones G, Hodgson J, Halstead P (2002) Towards the archaeobotanical identification of intensive cereal cultivation: present-day ecological investigation in the mountains of Asturias, northwest Spain. Veg Hist Archaeobot 11:133–142
Chaves MM (1991) Effects of water deficits on carbon assimilation. J Exp Bot 42:1–16
Choi W-J, Ro H-M, Hobbie EA (2003) Patterns of natural 15N in soils and plants from chemically and organically fertilized uplands. Soil Biol Biochem 35:1493–1500
Colledge S, Conolly J (eds) (2007) The origins and spread of domestic plants in Southwest Asia. Left Coast Press, Walnut Creek
Condon AG, Richards RA, Farquhar GD (1992) The effect of variation in soil water availability, vapour pressure deficit and nitrogen nutrition on carbon isotope discrimination in wheat. Aust J Agric Res 43:935–948
Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP (2002) Stable isotopes in plant ecology. Annu Rev Ecol Syst 33:507–559
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta 45:341–351
Evans RD (2001) Physiological mechanisms influencing plant nitrogen isotope composition. Trends Plant Sci 6:121–126
Fan F, Zhang G, Song Y, Sun J, Bao X, Guo T, Li L (2006) Nitrogen fixation of faba bean (Vicia faba L.) interacting with a non-legume in two contrasting intercropping systems. Plant Soil 283:275–286
Farquhar GD, Richards RA (1984) Isotope composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust J Plant Physiol 11:539–552
Farquhar GD, Sharkey TD (1982) Stomatal conductance and photosynthesis. Annu Rev Plant Physiol 33:317–345
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annu Rev Plant Physiol Plant Mol Biol 40:503–537
Fiorentino G, Ferrio JP, Bogaard A, Araus JL, Riehl S (2015) Stable isotopes in archaeobotanical research. Veg Hist Archaeobot 24:215–227
Flohr P, Müldner G, Jenkins E (2011) Carbon stable isotope analysis of cereal remains as a way to reconstruct water availability: preliminary results. Water Hist 3:121–144
Forbes H (1996) European agriculture viewed bottom-side upwards: fodder- and forage-provision in a traditional Greek community. Environ Archaeol 1:19–34
Franche C, Lindströ K, Elmerich C (2009) Nitrogen-fixing bacteria associated with leguminous and non-leguminous plants. Plant Soil 125:1–15
Fraser RA, Bogaard A, Heaton T, Charles M, Jones G, Christensen BT, Halstead P, Merbach I, Poulton PR, Sparkes D, Styring AK (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary patterns. J Archaeol Sci 38:2790–2804
Fraser RA, Bogaard A, Schäfer M, Arbogast R, Heaton THE (2013) Integrating botanical, faunal and human stable carbon and nitrogen isotope values to reconstruct land use and palaeodiet at LBK Vaihingen van der Enz. Baden Württemb World Archaeol 45:492–517
Gerzabek MH, Pichlmayer F, Kirchmann H, Haberhauer G (1997) The response of soil organic matter to manure amendments in a long-term experiment at Ultuna, Sweden. Eur J Soil Sci 48:273–282
Gröcke DR (1998) Carbon-isotope analyses of fossil plants as a chemostratigraphic and palaeoenvironmental tool. Lethaia 31:1–13
Gröcke DR (2002) The carbon isotope composition of ancient CO2 based on higher-plant organic matter. Philos Trans R Soc Lond Ser A 360:633–658
Halstead P (2014) Two oxen ahead: pre-mechanized farming in the Mediterranean. Wiley Blackwell, Oxford
Heaton THE (1999) Spatial, species, and temporal variations in the 13C/12C ratios of C3 plants: implication for palaeodiet studies. J Archaeol Sci 26:637–649
Heaton THE, Jones HP, Tsipropoulus T (2009) Variations in the 13C/12C ratios of modern wheat grain, and implications for interpreting data from Bronze Age Assiros Toumba, Greece. J Archaeol Sci 36:2224–2233
Hedges REM, Reynard LM (2007) Nitrogen isotopes and trophic level of humans in archaeology. J Archaeol Sci 34:1240–1251
Högberg P (1997) 15N natural abundance in soil-plant systems. New Phytol 137:179–203
Howard JB, Rees DC (1996) Structural basis of biological nitrogen fixation. Chem Rev 96:2965–2982
Jannoura R, Joergensen R, Bruns C (2014) Organic fertilizer effects on growth, crop yield, and soil microbial biomass indices in sole and intercropped peas and oats under organic farming conditions. Eur J Agron 52:259–270
Jenkinson DS, Coleman K, Harkness DD (1995) The influence of fertilizer nitrogen and season on the carbon-13 abundance of wheat straw. Plant Soil 171:365–367
Jones G (2005) Garden cultivation of staple crops and its implications for settlement location and continuity. World Archaeol 37:164–176
Jones G, Bogaard A, Halstead P, Charles M, Smith H (1999) Identifying the intensity of crop husbandry practices on the basis of weed floras. Annu Br Sch Athens 94:167–189
Kanstrup M, Thomsen IK, Andersen AJ, Bogaard A, Christensen BT (2011) Abundance of 13C and 15N in emmer, spelt and naked barley grown on differently manured soils: towards a method for identifying past manuring practice. Rapid Commun Mass Spectrom 25:2879–2887
Kanstrup M, Thomsen IK, Mikkelsen PH, Christensen BT (2012) Impact of charring on cereal grain characteristics: linking prehistoric manuring practice to δ15N signatures in archaeobotanical material. J Archaeol Sci 39:25–40
Kanstrup M, Holst MK, Jensen PM, Thomsen IL, Christensen BT (2014) Searching for long-term trend in prehistoric manuring practice. δ15N analyses of charred cereal grains from the 4th to the 1st millennium BC. J Archaeol Sci 51:115–125
Kohl DH, Shearer G (1980) Isotopic fractionation associated with symbiotic N2 fixation and uptake of NO-3 by plants. Plant Physiol 66:51–56
Köpke U, Nemecek T (2010) Ecological services of faba bean. Field Crop Res 115:217–238
Ledgard SF, Sprosen M, Steele K (1996) Nitrogen fixation by nine white clover cultivars in grazed pasture, as affected by nitrogen fertilization. Plant Soil 178:193–203
Liu Y, Wu L, Baddeley JA, Watson CA (2011) Models of biological nitrogen fixation of legumes. A review. Agron Sustain Dev 31:155–172
Lloyd J, Farquhar GD (1994) 13C Discrimination during CO2 assimilation by the terrestrial biosphere. Oecologia 99:201–215
López-Bellido FJ, López-Bellido RJ, Redondo R, López-Bellido L (2010) B value and isotopic fraction in N2 fixation by chickpea (Cicer arietinum L.) and faba bean (Vicia faba L.). Plant Soil 337:425–434
López-Bellido RJ, López-Bellido L, Benítez-Vega J, Muñoz-Romero V, López-Bellido FJ, Redondo R (2011) Chickpea and faba bean nitrogen fixation in a Mediterranean rainfed Vertisol: effect of the tillage system. Eur J Agron 34:222–230
Masi A, Sadori L, Restelli FB, Baneschi I, Zanchetta G (2014) Stable carbon isotope analysis as a crop management indicator at Arslantepe (Malayta, Turkey) during the Late Chalcolithic and Early Bronze Age. Veg Hist Archaeobot 23:751–760
Nebiyu A, Huygens D, Ram Upadhayay H, Diels J, Boeckx P (2014) Importance of correct B value determination to quantified biological N2 fixation and N balances of faba beans (Vicia faba L.) via 15N natural abundance. Biol Fertil Soils 50:517–525
O’Leary MH (1981) Carbon isotope fractionation in plants. Phytochemistry 20:553–567
O’Leary MH (1988) Carbon isotopes in photosynthesis. BioScience 38:328–336
Peoples MB, Brockwell J, Herridge DF, Rochester IJ, Alves BJR, Urquiaga S, Boddey RM, Dakora FD, Bhattarai S, Maskey SL, Sampet C, Rerkasem B, Khan DF, Hauggaard-Nielsen H, Jensen ES (2009) The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 48:1–17
Senbayram M, Dixon L, Goulding KWT, Bol R (2008) Long-term influence of manure and mineral nitrogen applications on plant and soil 15N and 13C values from the Broadbalk wheat experiment. Rapid Commun Mass Spectrom 22:1735–1740
Serret MD, Ortiz-Monasterio I, Pardo A, Araus JL (2008) The effects of urea fertilisation and genotype on yield, nitrogen use efficiency, δ15N and δ13C in wheat. Ann Appl Biol 153:243–257
Shearer G, Kohl DH (1986) N2-fixation in field settings: estimations based on natural 15N abundance. Aust J Plant Physiol 13:699–756
Simpson IA, Bol R, Bull ID, Evershed RP, Petzke KJ, Dockrill SJ (1999) Interpreting early land management through compound specific stable isotope analyses of archaeological soils. Rapid Commun Mass Spectrom 13:1315–1319
Stika H-P, Heiss A (2013) Plant cultivation in the bronze age. In: Fokkens H, Harding A (eds) The Oxford handbook of the European bronze age. Oxford University Press, Oxford, p 348–369
Stokes H, Müldner G, Jenkins E (2011) An investigation into the archaeological application of carbon stable isotope analysis used to establish water crop availability: solutions and ways forward. In: Mithen S, Black E (eds) Water, life and civilisation: climate, environment, and society in the Jordan valley. Cambridge University Press, Cambridge, pp 373–380
Styring AK, Fraser RA, Bogaard A, Evershed RP (2014) The effect of manuring on cereal and pulse amino acid δ15N values. Phytochemistry 102:40–45
Szpak P (2014) Complexities of nitrogen isotope biogeochemistry in plant-soil systems: implications for the study on ancient agricultural and animal management practices. Front Plant Sci 5:1–19
Szpak P, Longstaffe FJ, Millaire J-F, White CD (2012a) Stable isotope biogeochemistry of seabird Gauno fertilization: results from growth chamber studies with maize (Zea mays). PLoS ONE 7, e33741. doi:10.1371/journal.pone.0033741
Szpak P, Millaire J-F, White CD, Longstaffe J (2012b) Influence of seabird guano and camelid dung fertilization on the nitrogen isotopic composition of field-grown maize (Zea mays). J Archaeol Sci 39:3721–3740
Szpak P, Longstaffe FJ, Millaire J-F, White CD (2014) Large variation in nitrogen isotopic composition of a fertilized legume. J Archaeol Sci 45:72–79
Tieszen LL (1991) Natural variations in the carbon isotope values of plants: implications for archaeology, ecology, and palaeoecology. J Archaeol Sci 18:227–248
Treasure ER (2014) ‘Evidently carbonised by fire action’: an archaeobotanical and experimental study of Vicia faba L. (broad bean, horsebean) in prehistoric Britain. Dissertation, Durham University
Tryderman MK, Hauggaard-Nielsen H, Jørnsgård B, Steen Jensen E (2004) Comparison of interspecific competition and N use in pea-barley, faba bean-barley and lupin-barley intercrops grown at two temperate locations. J Agric Sci 142:617–627
Unkovich M (2013) Isotope discrimination provides new insight into biological nitrogen fixation. New Phytol 198:643–646
Vaiglova P, Bogaard A, Collins M, Cavanagh W, Mee C, Renard J, Lamb A, Gardeisen A, Fraser R (2014) An integrated stable isotope study of plants and animals from Kouphovouno, southern Greece: a new look at Neolithic farming. J Archaeol Sci 42:201–215
van Klinken GT, Richards MP, Hedges REM (2002) An overview of causes for stable isotopic variations in past European human populations: environmental, ecophysiological, and cultural effects. In: Ambrose SH, Katezenberg MA (eds) Biogeochemical approaches to palaeodietary analysis. Kluwer Academic, New York, pp 39–63
Vinther F (1998) Biological nitrogen fixation in grass-clove affected by animal excreta. Plant Soil 203:207–215
Virginia RA, Delwiche CC (1982) Natural 15N abundance of presumed N2-fixing and non-N2-fixing plants from selected ecosystems. Oecologia 54:317–325
Wallace M, Jones G, Charles M, Fraser R, Halstead P, Heaton THE, Bogaard A (2013) Stable carbon isotope analysis as a direct means of inferring crop water status and water management practices. World Archaeol 45:388–409
Watzka M, Buchgraber K, Wanke W (2006) Natural 15N abundance of plants and soils under different management practices in a montane grassland. Soil Biol Biochem 38:1564–1576
Acknowledgments
Edward Treasure holds a Northern Bridge AHRC studentship (Grant Number: AH/L002558/1). We thank Mike Hughes (Durham University Botanic Garden) for logistical support when growing the plants. We also thank the reviewers for their constructive comments.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 18.5 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Treasure, E.R., Church, M.J. & Gröcke, D.R. The influence of manuring on stable isotopes (δ13C and δ15N) in Celtic bean (Vicia faba L.): archaeobotanical and palaeodietary implications. Archaeol Anthropol Sci 8, 555–562 (2016). https://doi.org/10.1007/s12520-015-0243-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-015-0243-6