
Abstract
Australian tidal wetlands differ in important respects to better studied northern hemisphere systems, an artefact stable to falling sea levels over millennia. A network of Surface Elevation Table-Marker Horizon (SET-MH) monitoring stations has been established across the continent to assess accretionary and elevation responses to sea-level rise. This network currently consists of 289 SET-MH installations across all mainland Australian coastal states and territories. SET-MH installations are mostly in mangrove forests but also cover a range of tidal marsh and supratidal forest ecosystems. Mangroves were found to have higher rates of accretion and elevation gain than all the other categories of tidal wetland, a result attributable to their lower position within the tidal frame (promoting higher rates of accretion) higher biomass (with potentially higher rates of root growth), and lower rates of organic decomposition. While Australian tidal marshes in general show an increase in elevation over time, in 80% of locations, this was lower than the rate of sea-level rise. High rates of accretion did not translate into high rates of elevation gain, because the rate of subsidence in the shallow substrate increased with higher accretion rates (r2 = 0.87). The Australian SET-MH network, already in many locations spanning two decades of measurement, provides an important benchmark against which to assess wetland responses to accelerating sea-level rise in the decades ahead.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tidal wetlands (mangroves, tidal marshes and supratidal forests) are important coastal zone habitats. These three communities differ in structural characteristics and zonation in relation to the tidal frame. In Australia, mangroves (typically Rhizophoraceae and Avicenniaceae) generally grow between mean sea level and mean high water spring tides (Saintilan et al. 2019). Tidal marshes (or saltmarshes) consist of low-growing herbs and grasses (common species include Sporobolus virginicus, Sarcocornia quinqueflora, Samolus repens, Triglochin striata, Suaeda australis), salt bushes (of the genera Tecticornia, Atriplex) and in more brackish to freshwater environments rushes (e.g. Juncus, Phragmites). Australian tidal marshes are more frequently found in upper intertidal environments, inundated by spring tides (Saintilan et al. 2019), and extending beyond highest astronomical tide where substrate conditions are favourable (i.e. freshwater supply and saline substrate). Tidal forests (predominantly of the genera Casuarina and/or Melaleuca) are tolerant of infrequent tidal inundation. They occur between mean high water spring tidal levels through to elevations at or above the highest astronomical tides (Kelleway et al. 2021).
Australian mangroves, tidal marshes and supratidal forests make important contributions to a range of ecosystem services (Kelleway et al. 2017; Hagger et al. 2022). The disproportionate contribution of these habitats to natural carbon sequestration has been demonstrated for all three habitats, and they are the subject of emerging opportunities for blue carbon emission reductions (Kelleway et al. 2020; Lovelock et al. 2022). The contribution of mangroves and tidal marshes to estuarine fisheries has also been demonstrated (Mazumder et al. 2006, 2011; Sheaves et al. 2015; Prahalad et al. 2019). They are important habitat for a number of endangered and vulnerable species of birds and mammals (Gonsalves et al. 2013; Kelleway et al. 2017; Sievers et al. 2019).
The continued provision of the ecosystem services from coastal wetland environments is threatened by climate change. Several climate drivers influence survival and competitive interactions in coastal wetlands, including temperature, elevated carbon dioxide concentrations, precipitation and sea-level rise (McKee et al. 2012). The proliferation of mangroves in higher latitudes, where they compete with tidal marshes, has been demonstrated on five continents and linked to all of these drivers (Saintilan et al. 2014). In southeast Australia, tidal marsh land cover has been converting to mangrove land cover in most estuaries where long-term habitat dynamics have been studied, and the proportion of decline is consistent with sea-level trends over the period of observation (Saintilan et al. 2014). Tidal forest transitions to tidal marsh have also been observed, and the contribution of tidal forest habitats to carbon sequestration in Australia has only recently been considered (Kelleway et al. 2021).
While vulnerable to sea-level rise by virtue of their position in relation to tidal inundation, tidal wetlands can build elevation relative in response to sea-level rise, ameliorating the impacts of increased water level (Schuerch et al. 2018). Indeed, the effect of sea-level rise and increased hydroperiod is to increase the rate of sedimentation, which is proportional to the depth and duration of inundation. Also, increased frequency of inundation can promote plant growth, which may be limited higher in the tidal frame by higher porewater salinities and/or and lower nutrient concentrations (Feller et al. 2003). Increased biomass production contributes to the building of root volume, an important component of wetland elevation gain (Morris et al. 2002, 2016; Rogers et al. 2005a, b; Ola et al. 2018). Also, the more anaerobic conditions created by increased inundation can lead to greater carbon preservation, enhancing the blue carbon efficacy of tidal wetlands (Rogers et al. 2019).
Increases in water level may be balanced by increases in the elevation of the marsh, a negative feedback which may lead to marsh equilibrium with sea-level rise. Under this model, marshes rise or fall in the tidal frame to an optimal position (Pethick 1981; Cahoon et al. 2019), Fig. 1). Modelling based on accretion responses in tidal marshes has suggested a robust response even to comparably high rates of sea-level rise given sufficient sediment supply and deposition, prompting the suggestion that the vulnerability of tidal marshes has been overestimated in previous studies (Kirwan et al. 2016; Moritsch et al. 2022). However, the extent and efficacy of negative feedbacks between sea-level rise and vertical accretion are still poorly understood. Recent syntheses from palaeo-stratigraphic observations of marsh responses to sea-level rise during the early Holocene, when rates of RSLR were higher than today, suggest that while coastal wetlands can track low rates of sea-level rise, this capacity is lost under rates exceeding 5–7 mm per year (Horton et al. 2018; Saintilan et al. 2020).

Marsh equilibrium model for coastal wetlands experiencing sea-level rise. A deficit between marsh elevation gain and relative sea-level rise (RSLR) decreases the position of a marsh in the tidal frame (trajectory “a”), which enhances accretion, lowering the rate of relative sea-level rise and restoring the marsh to an optimal position in the tidal frame (trajectory “b”). However, if the position of the marsh becomes too low, or this feedback is too weak, anoxic conditions lead to marsh drowning (trajectory “c”), rapid elevation loss and conversion to open water
Contemporary observations of marsh responses to sea-level rise derived from accretion records (derived from radiometric and artificial markers) provide information on rates of sediment accumulation, but the extent to which this translates into elevation gain is critically important in determining marsh survival. Recently, deposited sediment is subject to autocompaction, and ongoing accretion contributes to the autocompaction of sediment below the surface (termed upper-level subsidence). Upper-level subsidence of accreting soil compromises the contribution of accretion to surface elevation gain and may be a key determinant of the resilience of wetlands under sea-level rise (Rogers and Saintilan 2021; Saintilan et al. 2022).
The Surface Elevation Table-Marker Horizon (SET-MH) technique measures the accretion and elevation of wetlands (Cahoon et al. 2000). A survey benchmark rod serves as a fixed benchmark against which relative changes in the elevation of the wetland surface are measured; these changes can be converted to absolute measurements when calibrated against a fixed datum, such as the Australian height datum. The rod is driven deep into the wetland substrate (up to 30 m), and measurements are periodically made using a detachable arm, from which pins are lowered to the wetland surface. As the wetland accretes and the elevation of the substrate surface increases, the pins appear higher against the level arm.
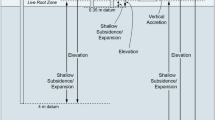
Two SET designs have been commonly deployed globally. The original SET consisted of an aluminium tube, manually driven into the wetland substrate to a maximum depth of 8 m; the depth was largely delimited by the length of aluminium tube of the depth to basement. An insert tube was concreted into the top of the SET pole, upon which the SET arm was attached (Fig. 2). A subsequent innovation of the SET utilised a solid stainless steel rod as the SET benchmark, allowing far greater depth of installation (Cahoon et al. 2002). This type of SET is termed the rod-SET, or rSET, and a lighter linear arm is used (Fig. 2). At the time of installation, feldspar marker horizons are often laid on the wetland surface, and sediment accretion can be determined by measuring sediment accumulation above the horizon over time. The accretion of sediment above the feldspar layer is gauged by sampling shallow cores in the feldspar plots. These two measurements combined are known as the Surface Elevation Table-Marker Horizon technique or SET-MH. Upper-level subsidence is inferred as the difference between vertical accretion measured against the feldspar marker horizon and elevation gain as measured from the SET benchmark rod. This defines subsidence occurring between the surface and the base of the benchmark rod. At Comerong Island, New South Wales, shallow rSETS (Cahoon et al. 2002) were also installed to measure the contribution of root zone processes to surface elevation trends.

Measuring the rod Surface Elevation Table (left) and the original Surface Elevation Table (right). (Credit: Catherine Lovelock; Neil Saintilan)
SET-MH measurements are often compared to water level changes at nearby tide gauges (Fig. 3). If the rate of water level increase exceeds the rate of elevation gain over the same period (termed the period of observation or contemporaneous sea-level rise), then the location at which SET measurements are taken may be subject to an elevation deficit; other locations, particularly those lower in the tidal frame, may not exhibit an elevation deficit (Morris 2006). Locations within a wetland that exhibit an elevation deficit are transitioning to lower positions within the tidal frame, where they may equilibrate with sea-level rise or may continue their trajectory towards submergence (Fagherazzi et al. 2020). Over time, the fate of such wetlands will depend on the timing and strength of the negative feedbacks between water level rise and vertical accretion described previously. For this reason, long-term SET-MH measures are essential in order to capture feedback responses that may occur over decadal time periods. Syntheses of SET-MH observations usually reject observation periods of less than 3 years.

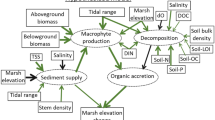
Operation of the SET-MH benchmark station, illustrating factors contributing to soil volume change
To date, approximately 1000 SET-MH stations have been installed in over 40 countries worldwide (https://www.usgs.gov/centers/eesc/science/surface-elevation-table). Extensive regional SET networks include Europe (UK, the European North Sea and Mediterranean coastlines), US Gulf of Mexico and East Coast regions, South Africa, Central America and the Caribbean, Asia (Singapore, Thailand, China, Vietnam, Indonesia), Oceania (New Zealand, Micronesia) and Australia. The technique has been described as the “global standard” for wetland monitoring against sea-level rise (Webb et al. 2013) and subject to important regional and global-scale synthesis reviews (Lovelock et al. 2015; Jankowski et al. 2017; Saintilan et al. 2022). While significant gaps in global coverage remain (Equatorial Africa, South America, Arctic coastline, Middle East), the network encompasses a range of bioclimatic zones, tidal ranges and rates of relative sea-level rise.
The Australian SET-MH network has grown over two decades to include at least 289 installations across 30 geographic locations. Habitats represented in the network include mangrove (temperate and subtropical forests of Avicennia and tropical mangroves dominated by Rhizophora and Bruguiera), saltmarshes (incorporating the saltbushes, brackish rushes and herbaceous saltmarsh) and more recently supratidal forests and some tidal restoration sites. The network covers five Australian states and territories, spanning geomorphic settings including macrotidal estuaries, drowned river valleys, microtidal barrier estuaries and coral islands, as well as restoration sites. Data from this network has hitherto never been collated or analysed.
The SET-MH technique has been extensively applied to inform estuary-scale models of sea-level rise in coastal lowlands in New South Wales (Oliver et al. 2012; Traill et al. 2011; Rogers et al. 2012), Victoria (Rogers and Saintilan 2021; Rogers et al. 2022) and Queensland (Traill et al. 2011; Runting et al. 2017; Mazor et al. 2021). However, greater insight can be gained by the collation and synthesis of information from SETs covering a range of hydrological and geomorphic settings (Saintilan et al. 2022), as a means of identifying important biological and environmental controls. We purposed therefore to (i) compile the existing SET-MH dataset for Australia, including the location of SET-MH stations, data custodian, the length of record and existing data, while identifying gaps; (ii) compile ancillary environmental data relevant to the interpretation of SET-MH trends, including climate, geomorphic setting, tide range, dominant species and the rate of local sea-level rise for the period of SET-MH measurement; and (iii) conduct analyses of SET-derived tidal wetland elevation trends in relation to key climatic and environmental drivers, including the efficacy of restoration (Table 1).
Methods
Surface Elevation Tables were first installed in Australia in 2000–2001. With assistance from the US Geological Survey, SET-MH stations were installed in 2000–2001 in New South Wales (Tweed River, Hunter River, Hawkesbury River, Parramatta River, Minnamurra River, Jervis Bay) and in Victoria (Westernport Bay). SET-MH installation in Queensland commenced in 2007, initially in Moreton Bay, expanding to the Daintree River in 2014 and most recently at the most poleward extent of mangroves at Corner Inlet, Victoria (2019) and Woody Island, of the Low Isles within the Great Barrier Reef lagoon (2022). Installation of SET-MH stations in Darwin Harbour commenced in 2016 funded under the Darwin Harbour Integrated Marine Monitoring and Research Program (IMMRP). In Western Australia, SET-MH stations were installed in the Exmouth Gulf in 2011 and in the saltmarshes of the south of the state in 2021. In South Australia, SETs were installed in 2017 within a tidal restoration site in a decommissioned salt production pond (Dittmann et al. 2019), as well as two mangrove sites close to Adelaide, with further SET-MHs installed in the west of the state since 2020.
Data were compiled with the assistance of SET-MH data custodians as set out in Table 2. Variables provided by data custodians included sampling location, the date of the initial reading and the most recent observations and the dominant species found at the site. Data custodians also provided the rate of elevation gain (in millimetres per year) as the linear trend of the SET pin measurements for the duration of the (r)SET-MH record and sediment accretion (in millimetres per year) as the linear trend of the MH observations for the duration of the accretion record (in some but not all cases). For each (r)SET, relative pin height was calculated by subtracting baseline pin height from all subsequent readings. Relative pin heights were averaged for each measurement date hierarchically within each SET arm position and then across positions to integrate small-scale spatial variation in surface elevation across the measurement area. At the time of publication, the short length of record for tidal forest SETs precluded their incorporation into the analysis.
Rainfall and temperature variables were sourced from the Bureau of Meteorology (http://www.bom.gov.au/climate/data/), and sea-level trends were calculated from the closest tide gauge in the Australian Baseline Sea Level Monitoring Network (http://www.bom.gov.au/oceanography/projects/abslmp/abslmp.shtml). A full list of variable names and their explanations is provided in Table 5 in the Appendix. Linear regression models were used to test relationships between accretion, elevation gain, shallow subsidence and water level. Paired t-tests were used to compare elevation trends between mangrove and tidal marsh SETs.
Sites were classified according to the geomorphic units using a typology that defines estuarine settings on the basis of dominance of river, wave and tide energy (Dalrymple et al. 1992) and using the Roy et al. (2001) model of estuary classification for southeast Australian estuaries: barrier estuarine (estuaries sheltered behind sand barriers along wave-dominated coastlines), river delta (sites associated with river systems where fluvial sedimentation is building active deltas), drowned river valley (sites of meso-macrotidal range in which tidal deposition and erosion are dominant processes), coral island (sites associated with coral reef barriers) and marine embayment (sites protected from oceanic waves by shoreline configuration but for which fluvial influence is minor). In three cases, SET-MH installations occurred in sites historically or actively subject to tidal marsh restoration. In two cases, these consisted of former commercial salt evaporation ponds subject to tidal restoration: French Island, Westernport, abandoned early in the twentieth century, and at Dry Creek, South Australia, where tidal restoration commenced in 2017. At the Tomago Wetland, New South Wales, tidal reinstatement occurred in 2015, with the intention of restoring coastal wetlands to an agricultural floodplain protected by flood control works. Many of the sites have also been modified by humans, with drainage, ditching and infill common at the margins of coastal wetlands, or wetlands were formally used for grazing purposes.
Results
Location and Site Characteristics
The Australian SET network currently consists of 289 documented benchmark installations distributed across 31 sites in all the mainland coastal states and territories (Table 1, Fig. 1). While SET installations are numerically clustered around the major SE Australian population centres of Adelaide, Melbourne, Sydney, Newcastle and Brisbane, the network has expanded in recent years to several locations in Western Australia and the Tropical north (Darwin Harbour, Daintree, and Woody Island in the Great Barrier Reef). The Australian SET network encompasses a range of geomorphic settings; it is dominated by sites in river deltas, marine embayments and drowned river valleys (Table 3, Fig. 4).

Distribution of the Australian SET-MH network and rate of elevation gain. Locations are approximate (coordinates and site numbers shown in Table 2). Yellow circles show recently installed SET-MH stations (no data). Paired sites show tidal marsh on the left and mangrove on the right
Australian SET-MH stations are equally divided between mangrove (53%) and tidal marshes (43%), with supratidal forests (4%) being under-represented in the network (Fig. 4). Being the most recent habitat sampled, supratidal forest elevation trend and accretion data are not yet available.
Rates of Elevation Gain in Relation to Sea Level
The average rate of elevation gain for mangrove and tidal marsh sites is shown in Table 4 and Figs. 4 and 5. Table 4 does not include sites recently established for which return readings have not been undertaken or for which the duration of record is too brief to derive a reliable trend (all of the Western Australian Sites, Corner Inlet in Victoria, Towra Point in New South Wales and Woody Island on the Great Barrier Reef). Of the mangrove sites, 83% showed an increased in elevation averaged across the SETs, and 63% of tidal marsh sites showed an increasing elevation trend over the period of measurement (Table 4). Mangroves have a higher rate of accretion (3.77 ± 3.04 mm yr−1) than tidal marshes (1.80 ± 1.04 mm yr−1; p < 0.001) and higher elevation gain (2.34 ± 3.97 mm yr−1) than tidal marshes (0.61 ± 1.43 mm yr−1; p < 0.001), consistent with their lower position in the tidal frame (Fig. 5).

Comparison of the rate of accretion (from the MH horizon) and elevation gain (against the SET benchmark) in mangrove compared to the three structural categories of tidal marsh
Of 44 locations (sites with specific vegetation habitat: Table 4), only three sites showed a significant loss of elevation over the period of measure: the two Tomago sites in New South Wales subject to tidal restoration (− 5.64 ± 4.60 mm yr−1 in the zone subject to greatest tidal exposure and 2.91 ± 2.86 mm yr−1 in a zone subject to lesser tidal exposure) and the Dry Creek Salt Field site in South Australia (− 3.72 ± 2.53 mm yr−1), also subject to tidal restoration. All other sites showed a stable (> 1 mm yr−1 change: 45% of sites) or increasing elevation trend (48% of sites).
Three of the New South Wales saltmarsh sites were subject to mangrove encroachment during the 20-year period of observation, and all had very low rates of elevation gain. These were Currambene Creek (0.32 ± 0.47 mm yr−1), the Minnamurra River (0.49 ± 0.57 mm yr−1) and Ukerebagh Island on the Tweed River estuary (0.24 ± 0.61 mm yr−1).
Nearly all sites in the Australian SET-MH network show a deficit in elevation gain compared to water level rises over the period of SET-MH measurement (Table 4). Only three showed a surplus of elevation gain over sea-level rise averaged across SETs: the two Giralia sites in Western Australia, an artefact of a declining relative sea-level trend over the period of measurement, and the Marramarra site in New South Wales, an artefact of a stable sea-level trend over the period of measurement. Of the 50 locations for which elevation trend data were available (Table 1), 80% show a deficit between elevation gain and water level increase. Of the mangrove locations, 72% showed an elevation deficit, a lower proportion than for tidal marshes (88%).
Drivers of Vertical Accretion and Elevation Gain
Elevation gain was weakly correlated with the rate of sediment accretion (p = 0.028, r2 = 0.04). The relationship between the rate of accretion and sea-level rise was weak (r2 = 0.07) but significant (p = 0.005; n = 108). Similarly, the relationship between elevation gain and sea-level rise was weak (r2 = 0.08) but significant (p = 0.0006). The rate of subsidence was directly proportional to the rate of accretion (r2 = 0.76), in a near 1:1 relationship (Fig. 6a). Because subsidence is reducing the contribution of accretion to net elevation gain in direct proportion to the rate of accretion, a deficit between elevation gain and relative sea-level increases under high rates of relative sea-level rise (Fig. 6b).

Relationship between the rate of sediment accretion above the feldspar marker horizon and the rate of upper-level subsidence (A) in Australian tidal wetlands (mangrove and tidal marsh combined: n = 108). Relationship between relative sea-level rise (RSLR) and the deficit between RSLR and elevation gain (B) in Australian tidal wetlands (n = 183)
Discussion
The Australian SET-MH network has been established to explore the vertical adjustment of tidal wetlands to sea-level rise and the processes influencing this adjustment. The collation and synthesis of the Australian SET-MH data have provided the opportunity to understand accretionary and elevation responses across a range of biogeomorphic settings. Results from the Australian SET-MH network are largely consistent with other regional and global-scale syntheses. At over 80% of sites, the rate of surface elevation gain lagged behind local sea-level rise, a proportion similar to SET-MH observations from 51 sites in Florida, USA (Feher et al. 2022). The relationship between accretion and upper-level subsidence, previously noted in Westernport (Rogers and Saintilan 2021) and in global tidal marsh analyses (Saintilan et al. 2022), extended across the Australian network and was particularly strong (r2 = 0.87). The result helps explain why Australian tidal marshes are showing a near ubiquitous deficit in relation to RSLR, and why the size of this deficit increases consistently with RSLR (r2 = 0.66). Surface accretion is correlated with subsidence of upper layers of sediment, which, in addition to the increased water burden, introduces negative feedback into the RSLR accretion response. The result underscores the importance of considering elevation gain rather than surficial sediment accretion in assessing the vulnerability of tidal wetlands to sea-level rise, in both monitoring and modelling exercises.
The lower rate of elevation gain in tidal marshes is partly partly an artefactt of their higher position in the tidal frame. Detailed consideration of the response of the Westernport SETs has shown elevation gain to be closely related to sediment supply and increased accommodation afforded by sea-level rise (Rogers et al. 2022). The deficit that emerges between elevation gain and sea-level rise may be temporary, with higher rates of elevation gain restored lower in the tidal frame. Mangroves exhibit higher rates of accretion and elevation gain, and further work is required to determine the relative contributions of autochthonous organic contributions, allochthonous sediment inputs and decomposition rates in driving these higher rates of elevation gain. Mangrove encroachment into saltmarsh may facilitate this adjustment if their more efficient elevation gain is partly intrinsic. However, in some of the instances where mangrove encroachment into tidal marsh SET plots has been observed, the initial response has been a decline in the rate of elevation gain (e.g. Rogers et al 2013). This may be a short-term response associated with saltmarsh dieback.
Two sites in the network (Dry Creek, South Australia, and Tomago Wetland in New South Wales) were measured immediately following tidal restoration. Tidal reinstatement in wetlands previously isolated from tidal exchange is a restoration mechanism successfully applied to degraded floodplains in Australia (Streever 1998; Glamore et al. 2021; Bell-James and Lovelock 2019) and has emerged as a promising opportunity for “blue carbon” emissions abatement (Hagger et al. 2022; Kelleway et al. 2020). Early results from both Tomago and Dry Creek suggest an initial response of the surface may be elevation loss, and this was quite rapid compared to elevation trajectories in natural wetlands and, in the case of Tomago, despite a low position within the tidal frame. Possible mechanisms for elevation loss in these settings may include subsidence following dieback of prior vegetation (Cahoon 2006), desiccation and erosion of the exposed surface and loading of the upper sediments from reintroduction of tidal inundation. The French Island saltmarsh site was a salt work abandoned in the early nineteenth century, and strong elevation gain at this site suggests that long-term elevation gains may be possible, and the Mutton Cove site in South Australia is a tidal reinstatement site showing elevation gains. Differences in elevation responses between these settings may relate to the time since tidal restoration but may also be influenced by available sediment, being relatively high at French Island and limited at Dry Creek.
Further work is required to better understand the response of Australian tidal wetlands to sea-level rise. The Australian SET-MH network is heavily clustered to sites easily accessible to major population centres. This has helped maintain an appropriate frequency of measures but has restricted sampling of some important wetland types. Until recently, arid zone wetlands were poorly represented, but new SET-MH installations in Western Australia and South Australia (Eyre Peninsula) will help to redress this issue. The macrotidal wetlands of northern Australia, the region of the greatest extent of tidal wetland in Australia, are represented by Darwin Harbour alone. Installations elsewhere in northern Australia would help interpret tidal wetland responses to climate change in this highly dynamic environment. Within the existing network, accurate survey of the elevation of SET-MH installations in relation to fixed tidal datum would help interpret the implications of elevation deficits in relation to “elevation capital”, the elevation of the wetland above lower survival limits.
Finally, application of the Australian SET-MH data will help the research and management community develop better models of coastal lowland responses to sea-level rise. The US National Oceanographic and Atmospheric Administration (NOAA) has recently released a sea-level rise visualisation tool that includes a marsh accretion model based on SET-MH data (https://coast.noaa.gov/slr/#). The marsh migration module showcases the potential application of dynamic elevation models in coastal planning. Differences between rates of elevation gain between coastal wetland community types are implicated in vegetation transitions between, for example, mangrove and saltmarsh in SE Australia (Rogers et al. 2005a, b, 2006). These data provide the potential to model both vegetation transition and components of soil carbon stock change under a range of climate change and planning scenarios.
References
Bell-James, J., and C.E. Lovelock. 2019. Legal barriers and enablers for reintroducing tides: An Australian case study in reconverting ponded pasture for climate change mitigation. Land Use Policy 88: 104192.
Cahoon, D.R., P.E. Marin, B.K. Black, and J.C. Lynch. 2000. A method for measuring vertical accretion, elevation, and compaction of soft, shallow-water sediments. Journal of Sedimentary Research 70 (5): 1250–1253.
Cahoon, D.R., J.C. Lynch, B.C. Perez, B. Segura, R.D. Holland, C. Stelly, and G. Stephenson. 2002. Philippe Hensel; High-precision measurements of wetland sediment elevation: II. The rod surface elevation table. Journal of Sedimentary Research 72(5): 734–739. https://doi.org/10.1306/020702720734
Cahoon, D.R. 2006. A review of major storm impacts on coastal wetland elevations. Estuaries and Coasts 29: 889–898.
Cahoon, D.R., J.C. Lynch, C.T. Roman, J.P. Schmit, and D.E. Skidds. 2019. Evaluating the relationship among wetland vertical development, elevation capital, sea-level rise, and tidal marsh sustainability. Estuaries and Coasts 42: 1–15.
Dalrymple, R.W., B.A. Zaitlin, and R. Boyd. 1992. Estuarine facies models; conceptual basis and stratigraphic implications. Journal of Sedimentary Research 62: 1130–1146.
Dittmann, S., L. Mosley, K. Beaumont, B. Clarke, E. Bestland, H. Guan, H. Sandhu, M. Clanahan, R. Baring, J. Quinn, and R. Seaman. 2019. From salt to C; carbon sequestration through ecological restoration at the dry creek salt field. Goyder Institute for Water Research Technical Report Series 19(28): 1–102.
Fagherazzi, S., Mariotti, G., Leonardi, N., Canestrelli, A., Nardin, W., & Kearney, W. S. 2020. Salt marsh dynamics in a period of accelerated sea level rise. Journal of Geophysical Research: Earth Surface, 125(8), e2019JF005200.
Feher, L.C., M.J Osland, K.L. McKee, K.R. Whelan, C. Coronado-Molina, F.H. Sklar, and L. Allain. 2022. Soil elevation change in mangrove forests and marshes of the Greater Everglades: A regional synthesis of surface elevation table-marker horizon (SET-MH) data. Estuaries and Coasts 1–30.
Feller, I.C., K.L. Mckee, D.F. Whigham, and O’neill, J. P. 2003. Nitrogen vs. phosphorus limitation across an ecotonal gradient in a mangrove forest. Biogeochemistry 62: 145–175.
Glamore, W., D. Rayner, J. Ruprecht, M. Sadat-Noori, and D. Khojasteh. 2021. Eco-hydrology as a driver for tidal restoration: Observations from a Ramsar wetland in eastern Australia. PLoS ONE 16 (8): e0254701.
Gonsalves, L., B. Law, C. Webb, and V. Monamy. 2013. Foraging ranges of insectivorous bats shift relative to changes in mosquito abundance. PLoS ONE 8: e64081.
Hagger, V., N.J. Waltham, and C. E. Lovelock. 2022. Opportunities for coastal wetland restoration for blue carbon with co-benefits for biodiversity, coastal fisheries, and water quality. Ecosystem Services 55: 101423. https://doi.org/10.1016/j.ecoser.2022.101423
Horton, B.P., I. Shennan, S.L. Bradley, N. Cahill, M. Kirwan, R.E. Kopp, and T.A. Shaw. 2018. Predicting marsh vulnerability to sea-level rise using Holocene relative sea-level data. Nature Communications 9: 1–7.
Jankowski, K.L., T.E. Törnqvist, and A.M. Fernandes. 2017. Vulnerability of Louisiana’s coastal wetlands to present-day rates of relative sea-level rise. Nature Communications 8: 1–7.
Kelleway, J.J., M.F. Adame, C. Gorham, J. Bratchell, O. Serrano, P.S. Lavery, C.J. Owers, K. Rogers, Z. Nagel‐Tynan, and N. Saintilan. 2021. Carbon storage in the coastal swamp oak forest wetlands of Australia. Wetland Carbon and Environmental Management 339–353.
Kelleway, J.J., K. Cavanaugh, K. Rogers, I.C. Feller, E. Ens, C. Doughty, and N. Saintilan. 2017. Review of the ecosystem service implications of mangrove encroachment into salt marshes. Global Change Biology 23: 3967–3983.
Kelleway, J.J., O. Serrano, J.A. Baldock, R. Burgess, T. Cannard, P.S. Lavery, C.E. Lovelock, P.I. Macreadie, P. Masqué, and M. Newnham. 2020. A national approach to greenhouse gas abatement through blue carbon management. Global Environmental Change 63: 102083.
Kirwan, M.L., S. Temmerman, E.E. Skeehan, G.R. Guntenspergen, and S. Fagherazzi. 2016. Overestimation of marsh vulnerability to sea level rise. Nature Climate Change 6: 253–260.
Lovelock, C.E., M.F. Adame, J. Bradley, S. Dittmann, V. Hagger, S.M. Hickey, L.B Hutley, A. Jones, J.J. Kelleway, and P.S. Lavery. 2022. An Australian blue carbon method to estimate climate change mitigation benefits of coastal wetland restoration. Restoration Ecology e13739.
Lovelock, C.E., D.R. Cahoon, D.A. Friess, G.R. Guntenspergen, K.W. Krauss, R. Reef, K. Rogers, M.L. Saunders, F. Sidik, and A. Swales. 2015. The vulnerability of Indo-Pacific mangrove forests to sea-level rise. Nature 526: 559–563.
Mazor, T., R.K. Runting, M.I. Saunders, D. Huang, D.A. Friess, N.T. Nguyen, R.J. Lowe, J.P. Gilmour, P.A. Todd, and C.E. Lovelock. 2021. Future-proofing conservation priorities for sea level rise in coastal urban ecosystems. Biological Conservation 260: 109190.
Mazumder, D., N. Saintilan, and R.J. Williams. 2006. Trophic relationships between itinerant fish and crab larvae in a temperate Australian saltmarsh. Marine and Freshwater Research 57: 193–199.
Mazumder, D., N. Saintilan, R.J. Williams, and R. Szymczak. 2011. Trophic importance of a temperate intertidal wetland to resident and itinerant taxa: Evidence from multiple stable isotope analyses. Marine and Freshwater Research 62: 11–19.
McKee, K., K. Rogers, and N. Saintilan. 2012. Response of salt marsh and mangrove wetlands to changes in atmospheric CO2, climate, and sea level. Global change and the function and distribution of wetlands 63–96.
Moritsch, M.M., K.B. Byrd, M. Davis, A. Good, J.Z. Drexler, J.T. Morris, and J.M. Rybczyk. (2022). Can coastal habitats rise to the challenge? Resilience of estuarine habitats, carbon accumulation, and economic value to sea-level rise in a Puget Sound estuary. Estuaries and Coasts 45(8): 2293–2309.
Morris, J.T. 2006. Competition among marsh macrophytes by means of geomorphological displacement in the intertidal zone. Estuarine, Coastal and Shelf Science 69: 395–402.
Morris, J.T., D.C. Barber, J.C. Callaway, R. Chambers, S.C. Hagen, C.S. Hopkinson, B.J. Johnson, P. Megonigal, S.C. Neubauer, and T. Troxler. 2016. Contributions of organic and inorganic matter to sediment volume and accretion in tidal wetlands at steady state. Earth’ s Future 4: 110–121.
Morris, J.T., P. Sundareshwar, C.T. Nietch, B. Kjerfve, and D.R. Cahoon. 2002. Responses of coastal wetlands to rising sea level. Ecology 83: 2869–2877.
Ola, A., S. Schmidt, and C.E. Lovelock. 2018. The effect of heterogeneous soil bulk density on root growth of field-grown mangrove species. Plant and Soil 432: 91–105.
Oliver, T.S., K. Rogers, C.J. Chafer, and C.D. Woodroffe. 2012. Measuring, mapping and modelling: An integrated approach to the management of mangrove and saltmarsh in the Minnamurra River estuary, southeast Australia. Wetlands Ecology and Management 20: 353–371.
Pethick, J.S. 1981. Long-term accretion rates on tidal salt marshes. Journal of Sedimentary Research 51 (2): 571–577.
Prahalad, V., V. Harrison-Day, P. McQuillan, and C. Creighton. 2019. Expanding fish productivity in Tasmanian saltmarsh wetlands through tidal reconnection and habitat repair. Marine and Freshwater Research 70 (1): 140–151.
Rogers, K., N. Saintilan, and D. Cahoon. 2005a. Surface elevation dynamics in a regenerating mangrove forest at Homebush Bay, Australia. Wetlands Ecology and Management 13: 587–598.
Rogers, K., N. Saintilan, and H. Heijnis. 2005b. Mangrove encroachment of salt marsh in Western Port Bay, Victoria: The role of sedimentation, subsidence, and sea level rise. Estuaries 28: 551–559.
Rogers, K., K.M. Wilton, and N. Saintilan. 2006. Vegetation change and surface elevation dynamics in estuarine wetlands of southeast Australia. Estuarine, Coastal and Shelf Science 66 (3–4): 559–569.
Rogers, K., N. Saintilan, and C. Copeland. 2012. Modelling wetland surface elevation dynamics and its application to forecasting the effects of sea-level rise on estuarine wetlands. Ecological Modelling 244: 148–157.
Rogers, K., N. Saintilan, A.J. Howe, and J.F. Rodríguez. 2013. Sedimentation, elevation and marsh evolution in a southeastern Australian estuary during changing climatic conditions. Estuarine, Coastal and Shelf Science 133: 172–181.
Rogers, K., J.J. Kelleway, N. Saintilan, J.P. Megonigal, J.B. Adams, J.R. Holmquist, M. Lu, L. Schile-Beers, A. Zawadzki, and D. Mazumder. 2019. Wetland carbon storage controlled by millennial-scale variation in relative sea-level rise. Nature 567: 91–95.
Rogers, K., and N. Saintilan. 2021. Processes influencing autocompaction modulate coastal wetland surface elevation adjustment with sea-level rise. Frontiers in Marine Science 879.
Rogers, K., A. Zawadzki, L.A. Mogensen, and N. Saintilan. 2022. Coastal wetland surface elevation change is dynamically related to accommodation space and influenced by sedimentation and sea-level rise over decadal timescales. Frontiers in Marine Science 9: 807588.
Roy, P. S., R.J. Williams, A.R. Jones, I. Yassini, P.J. Gibbs, B. Coates, and S. Nichol. (2001). Structure and function of south-east Australian estuaries. Estuarine, coastal and shelf science 53(3): 351–384.
Runting, R.K., C.E. Lovelock, H.L. Beyer, and J.R. Rhodes. 2017. Costs and opportunities for preserving coastal wetlands under sea level rise. Conservation Letters 10 (1): 49–57.
Saintilan, N., N. Khan, E. Ashe, J. Kelleway, K. Rogers, C.D. Woodroffe, and B. Horton. 2020. Thresholds of mangrove survival under rapid sea level rise. Science 368: 1118–1121.
Saintilan, N., K.E. Kovalenko, G. Guntenspergen, K. Rogers, J.C. Lynch, D.R. Cahoon, C.E. Lovelock, et al. 2022. Constraints on the adjustment of tidal marshes to accelerating sea level rise. Science 377: 523–527.
Saintilan, N., K. Rogers, J. Kelleway, E. Ens, and D. Sloane. 2019. Climate change impacts on the coastal wetlands of Australia. Wetlands 39: 1145–1154.
Saintilan, N., N.C. Wilson, K. Rogers, A. Rajkaran, and K.W. Krauss. 2014. Mangrove expansion and salt marsh decline at mangrove poleward limits. Global Change Biology 20: 147–157.
Schuerch, M., T. Spencer, S. Temmerman, M.L. Kirwan, C. Wolff, D. Lincke, and S. Brown. 2018. Future response of global coastal wetlands to sea-level rise. Nature 561(7722): 231–234.
Sheaves, M., R. Baker, I. Nagelkerken, and R.M. Connolly. 2015. True value of estuarine and coastal nurseries for fish: Incorporating complexity and dynamics. Estuaries and Coasts 38: 401–414.
Sievers, M., C.J. Brown, V.J. Tulloch, R.M. Pearson, J.A. Haig, M.P. Turschwell, and R.M. Connolly. 2019. The role of vegetated coastal wetlands for marine megafauna conservation. Trends in Ecology & Evolution 34 (9): 807–817.
Streever, W.J. 1998. Kooragang Wetland Rehabilitation Project: Opportunities and constraints in an urban wetland rehabilitation project. Urban Ecosystems 2: 205–218.
Traill, L.W., K. Perhans, C.E. Lovelock, A. Prohaska, S. Mcfallan, J.R. Rhodes, and K.A. Wilson. 2011. Managing for change: Wetland transitions under sea-level rise and outcomes for threatened species. Diversity and Distributions 17: 1225–1233.
Webb, E.L., D.A. Friess, K.W. Krauss, D.R. Cahoon, G.R. Guntenspergen, and J. Phelps. 2013. A global standard for monitoring coastal wetland vulnerability to accelerated sea-level rise. Nature Climate Change 3: 458–465.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions This work was undertaken for the Marine and Coastal Hub, a collaborative partnership supported through funding from the Australian Government’s National Environmental Science Program (NESP). Funding and support in SA from the Eyre Peninsula Landscape Board, the SA Department for Environment and Water, the SA Coast Protection Board and the Goyder Institute for Water Research. Thanks to the Acraman Creek Conservation Park Co-management Board and the Barngarla Determination Aboriginal Corporation for permission to install and monitor SETs on country.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Meagan Eagle
Appendix
Appendix
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Saintilan, N., Sun, Y., Lovelock, C.E. et al. Vertical Accretion Trends in Australian Tidal Wetlands. Estuaries and Coasts (2023). https://doi.org/10.1007/s12237-023-01267-x
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12237-023-01267-x