
Abstract
Mesenchymal stem cells (MSCs) are well known for their beneficial effects, differentiation capacity and regenerative potential. Dental-derived MSCs (DSCs) are more easily accessible and have a non-invasive isolation method rather than MSCs isolated from other sources (umbilical cord, bone marrow, and adipose tissue). In addition, DSCs appear to have a relevant neuro-regenerative potential due to their neural crest origin. However, it is now known that the beneficial effects of MSCs depend, at least in part, on their secretome, referring to all the bioactive molecules (neurotrophic factors) released in the conditioned medium (CM) or in the extracellular vesicles (EVs) in particular exosomes (Exos). In this review, we described the similarities and differences between various DSCs. Our focus was on the secretome of DSCs and their applications in cell therapy for neurological disorders. For neuro-regenerative purposes, the secretome of different DSCs has been tested. Among these, the secretome of dental pulp stem cells and stem cells from human exfoliated deciduous teeth have been the most widely studied. Both CM and Exos obtained from DSCs have been shown to promote neurite outgrowth and neuroprotective effects as well as their combination with scaffold materials (to improve their functional integration in the tissue). For these reasons, the secretome obtained from DSCs in combination with scaffold materials may represent a promising tissue engineering approach for neuroprotective and neuro-regenerative treatments.
Graphical Abstract

Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Stem cells (SCs) are undifferentiated cell populations with high self-renewal capabilities and unlimited differentiation potential [1]. They are found during embryonic development (Embryonic stem cells, ESCs) and in many adult tissues and organs, where they replace dying cells and regenerate damaged tissue. ESCs are pluripotent cells. This means that they can differentiate into cell types belonging to each germ line (ectoderm, mesoderm, and endoderm). Adult stem cells (ASCs), are classified as mesenchymal stem cells (MSCs), Neural stem cells (NSCs), Epithelial stem cells (EpSCs), and Skin stem cells (SSCs) depending on the tissue which they are isolated and have limited differentiation potential. The most studied line of ASCs is the bone marrow-derived line (BM-MSCs), as they were discovered to belong to the hematopoietic niche in the 1970s [2]. However, also BM-MSCs have limited differentiation potential due to their mesodermal origin. This limitation is particularly relevant for the eventual therapeutic application in organs mainly composed of post-mitotic cells, such as the brain. Indeed, although BM-MSCs may be induced to differentiate vs a neurogenic phenotype, in this case they do not appear to be functionally active [3]. Since their initial discovery, MSCs have been isolated from a variety of tissues including dental tissues and named human dental-derived mesenchymal stem cells (DSCs). Gronthos et al., first isolated a population of MSCs from dental pulp, with similar properties to BM-MSCs [4]. DSCs in teeth have been described and classified according to their origin tissue, in dental pulp stem cells (DPSCs), stem cells from human exfoliated deciduous teeth (SHEDs), periodontal ligament stem cells (PDLSCs), dental follicle stem cells (DFSCs), stem cells from apical papilla (SCAPs), and gingival MSCs (GMSCs) [4,5,6,7,8,9]. Notably, DSCs are derived from the neural crest, a transient population of cells derived from the ectoderm germ layer, the same one that gives rise to mature neurons, and for this reason they show more potent neurogenic capabilities compared to other MSCs [10]. Thus, DSCs, may be a good source of MSCs for the treatment of neurodegenerative disorders and neural regeneration thanks to their differentiation potential and paracrine effects, [11, 12]. Indeed, most pharmacological approaches to nervous system disorders are directed against symptoms of the pathology (e.g., neurodegenerative disorders, autoimmune diseases) [13]. Thus, their beneficial effects on patients are limited. For this reason, cell therapy (CT) is receiving increased attention as a possible alternative approach to such pathologies (Fig. 1). CT involves the application of autologous or allogeneic cells into a patient [14] in order to alleviate symptoms, or to regenerate a tissue damage. Strikingly, even the administration of material derived from SCs (i.e., secretome) has been proved beneficial in preclinical models of nervous system diseases.

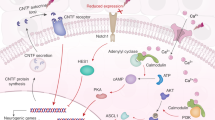
Schematic representation of DSCs in tissue engineering application for CT. DSCs can release growth factors and extracellular vesicles or be used directly with scaffolds for tissue engineering cell therapy in neurological disorders
In this review, we summarize the most recent research in the field of CT using human DSCs and their secretome. Highlighting analogies and differences between different types of DSCs, we recapitulate the main characteristics of these cells and the content of their secretome, as well as advances in materials to support and enable DSC growth. In the future, the increased use of neural crest SCs will provide invaluable therapeutic application and, ideally, the establishment of a biobank of these cells to perform autologous CT for the eradication of transplant rejection.
Dental Stem Cells
DSCs have the advantages of being easily accessible by minimally invasive procedures [15], being expandable and maintaining relative genomic stability over a long period of time and exhibiting immunomodulatory properties [16]. Moreover, they are also able to differentiate toward the mesodermal lineage, but also show the ability to transdifferentiate into ectodermal and endodermal lineages [17]. For these reasons, DSCs have been considered a promising tool for therapeutic applications [18] due in part to their ability to secrete multiple factors essential for tissue regeneration and because they are obtained with fewer ethical or legal problems compared to other procedures [11, 19, 20]. All types of DSCs show different origins and characteristics, but at the same time, share similar expression profiles of surface markers as shown in Table 1. They express not only mesenchymal and embryonic SCs markers, but also neuronal markers, as they are derived from migrating neural crest cells, which originated from the germ layer of the embryonic ectoderm [10].
Maintenance and Survival of DSCs
Differentiation Capabilities and Maintenance of DPSCs’stemness
DPSCs have a high differentiating capacity. In fact, it has been demonstrated that they can differentiate into different cell lineages including endodermal, mesodermal, and ectodermal lineages, respectively [19, 23, 48,49,50,51,52,53,54,55,56,57,58,59]. Furthermore, with the identification of functional markers and appropriate culture conditions for SC selection, it will be possible to target DPSCs for clinical applications [60]. In this regard, several authors have demonstrated that there are biological factors (such as tooth type, age, genetic background and lifestyle) that should be considered before tooth selection, and have compared traditional isolation, culture and storage techniques with improved methods or protocols such as the use of a serum/xenon-free culture medium to prolong the stemness of DPSCs [61, 62]. Diomede et al., studied the expression of proteins involved in cell proliferation/senescence and embryonic stem cell markers during early and late passages in MSCs obtained from dental pulp tissues, suggesting that the presence of embryonic and proliferation markers in late passages could potentially support the application of DSCs in clinical trials based on stem cell therapy [63].
Differentiation Capabilities and Maintenance of SHEDs’stemness
SHEDs also have a high differentiation capacity [5, 64,65,66,67,68]. SHEDs have a greater self-renewal potential than DPSCs even when subjected to adverse culture conditions [69, 70]. SHEDs, in addition to expressing the neural crest marker just like DPSCs [65], also express a wide range of lineage-specific markers and genes as shown in Table 1 [71, 72]. During long-term culture, SHEDs do not undergo spontaneous degeneration or differentiation [73]. Furthermore, SHEDs have been shown to possess higher proliferation rates and differentiation potential after cryopreservation and long-term storage (for two years) than DPSCs [29, 74] and retain similar properties to those obtained from fresh tissue [70, 75] that can be used for cell-based therapy [76]. It has been shown that the cell culture microenvironment can also influence the secretion and differentiation potential of pulp cells [77]. SHEDs and DPSCs, despite being SCs originating from the same tissue source, but from two different time points, show differences in their differentiation capacity towards major lineages [5, 65, 78, 79]. Naz et al., demonstrated that SHEDs have a better proliferation and self-renewal and better osteogenic differentiation capacity compared to DPSCs [80, 81]. Kanafi et al., further demonstrated that SHEDs have greater differentiation potential towards insulin-producing cells than DPSCs in the presence of appropriate inductive signals [70]. DPSCs and SHEDs are capable of regenerating pulp and dentin and therefore have the potential to be used as a source of pluripotent stem cells for future cell therapies in medicine and dentistry [78, 82, 83].
Differentiation Capabilities of Other DSC Types
PDLSCs and SCAPs possess the same in vitro multilineage differentiation potential like that of DPSCs and SHEDs [8, 34, 35, 40, 43, 84], making them good candidates in regenerative medicine to promote both dental and non-dental tissue regeneration [84]. DFSCs can differentiate into cementoblasts in vivo [85] and they can also be directed to differentiate into conventional multidirectional lineages [8, 86]. Instead, GMSCs can be targeted for osteogenic, adipogenic, chondrogenic, neurogenic, endothelial-like, odontogenic, and myogenic differentiation [46, 47].
Development of 3D Models to Study Neurological Disorders
Conventional in vitro models for studying SCs differentiation are usually cultured in two dimensions (2D). The in vitro three-dimensional (3D) model, which should ideally mimic the SCs microenvironment in vivo, is potentially useful for inducing stem cell-derived tissue formation. Biodegradable scaffolds play an important role in creating 3D structures to guide tissue formation. Furthermore, the diffusion of nutrients, oxygen, and bioactive factors through the 3D constructs shows increased efficiency for cell survival over long periods of time. Ideally, the scaffold should have mechanical properties that are consistent with the anatomical site where it is to be implanted. Recently, progress has been made in the development of natural and synthetic biomaterials for peripheral nerve (PN) repair to address the challenging clinical problem of damaged PN regeneration. These materials can be used as membranes useful in the repair of nerves and nerve ducts. For example, Wang et al., in 2020 demonstrated that collagen scaffold can promote the recovery of the functional facial nerve [87]. Moreover, the use of Silymarin nanoparticles loaded into the chitosan conduit was shown to improve functional recovery of transected sciatic nerve in rats [88]. On the same topic, Zorba Yildiz et al., investigated the preparation of a biohybrid hydrogel bioink containing graphene for use in peripheral tissue engineering and demonstrated that using of this bioink induced the neural differentiation of SCs [89]. Moreover, Su and Pan demonstrated the possibility to induce SHEDs’ neural differentiation by using 3D polydimethylsiloxane (PDMS) scaffolds in the perfusion system operating in the high-concentration of a rat Schwann cell (RSC)-seeded culture medium [90]. Other authors demonstrated that engineered Elastin-like proteins can support regeneration of PN [91]. The main scaffolds utilized to support cells involved in the repair process such as neurons, Schwann cells, macrophages, and blood vessels can be divided into biomaterials scaffold and biological scaffold. Both synthetic (biomaterials scaffold) and natural (biological scaffold) material show several tangible advantages and disadvantages attributed to both options [92, 93]. Due to their architectural structure, both types of scaffolds mimic the native extracellular matrix (ECM). The main structural difference is that biomaterial scaffolds are made of synthetic polymers or purified natural polymers, whereas biological scaffolds are composed of decellularized mammalian tissue [94].
Structural Properties (Characteristics) of Scaffolds
In general, biomaterials scaffolds are considered better than biological scaffolds for several characteristics such as precise geometrical pattern, biocompatibility, porosity, and stiffness to obtain a more easily and precise architecture and they can be more easily adapted, to improve cell adhesion and tune mechanical properties [95,96,97]. However, natural materials possess innate cell binding motifs, produce harmless degradation products, and effectively mimic physiological-like tissue remodeling and activate repair pathways [98, 99].
Geometrical Pattern
Geometrical pattern is one of the features able to modulate SCs differentiation. As reported by Das and Bellare also DPSCs in Customized 3D Nanofibrous Scaffolds for regeneration of Peripheral Nervous System (PNS) can support DPSCs proliferation and their subsequent neural differentiation [100]. Together with other studies, all authors demonstrated that transplanted scaffold containing DPSCs may provide a promising strategy for neuron repair, functional recovery, and neural tissue regeneration [101, 102].
Stiffness
Stiffness is one of the characteristics that can affect in a cell-specific way cells behavior during regeneration. For example, Sridharan et al., demonstrated that upon implantation of a biomaterial, MSCs and macrophages both contribute to the regeneration cascade of events [103]. Besides, Srinivanan et al., demonstrated that stiffer substrates can modulate human neural crest derived stem cell differentiation via CD44 mediated signaling [104]. Other studies reported that Schwann cells can develop normally on both stiff and soft hydrogels but activate different intracellular pathways in response to different substrate stiffness [105]. On this topic, Liu et al., demonstrated that exosome-loaded hydrogel stiffness plays an important role in tissue regeneration by regulating exosome release behavior [106]. Also, Guo et al., fabricated highly vascularized scaffolds embedded with human DPSCs, that thanks to their paracrine-mediated angiogenic and neuro-regenerative potentials were capable of augmenting and modulating Spinal Cord Injurie (SCI) repair [107].
Porosity
Also, porosity is an important parameter affecting neural cell regeneration. For example, nanostructures surface topographies have been developed to mimic microstructures for neuronal axons and Schwann cells also incorporating matrix proteins or peptides [108]. Injectable hydrogel systems have been quite extensively studied to deliver therapeutic molecules in various neurological disorders. This system has been mainly developed for the treatment of tumors to maintain a therapeutic concentration of the drug for a prolonged time in the local area and so to avoid toxicity [109]. Also, hydrogels composite comprising of dextran dialdehyde and gelatin, loaded with dopamine have been used in animal models of Parkinson’s disease (PD) to release neurotransmitters [94]. Hydrogel matrices have also been used for embedding different neurotrophic factors by an affinity-based system. The negatively charged sulfate groups present in heparin were capable of immobilizing neurotrophic growth factors such as nerve growth factor (NGF), glial cell derived neurotrophic factor (GDNF) and Neurotrophin 3 (NT-3) that could be released in the local area of treatment [110,111,112]. Ansari et al., demonstrated that PDLSCs and GMSCs promote nerve regeneration when encapsulated in a 3D scaffold based on alginate and hyaluronic acid hydrogels that, due to its elasticity, was capable to sustain the release of NGF [113]. Ueda et al., demonstrated that chitosan scaffolds combined with Basic fibroblast growth factor (bFGF) facilitated the neural differentiation of DPSCs [114].
Viscosity
Viscous characteristics together with elastic properties are also important in brain tissues that in a process of “stress-relaxation” responds to mechanical perturbations [115]. For example, hydrogel matrix allows to reproduce several factors that are required for the axonal growth such as mechanical stress experienced by the neural membrane, neuronal cytoskeletal dynamics, and mechano-sensing ability of the neurons. Luo et al., developed a 10% GelMA hydrogel mix with recombinant human bFGF and DPSCs to fill a cellulose/soy protein isolate composite membrane (CSM) tube. They constructed a nerve-like conduit, demonstrating that CSM-GFD could be a promising tissue engineering approach to replace the conventional nerve autograft to treat the large gap defect in peripheral nerve injuries [116]. Similarly, Li et al., demonstrated that dental follicle cells (DFCs) seeded on Aligned electrospun PCL/PLGA material (AEM) could stretch along the oriented fibers and proliferate efficiently and transplanted in rats contributed to restore the defect in rat spinal cord [117]. One of the goals of tissue engineering is therefore to try to exploit the innate regenerative capacity of SCs. Designing the scaffold in such a way that the scaffold itself delivers regenerative signals to cells could be a good model. In fact, in addition to the biomechanical contribution, cellular behavior is strongly influenced by biological and biochemical signals coming from the ECM.
Conductivity
Another property able to influence the cellular behaviour in response to a regenerative stimulus could be electrical conductivity of scaffolds. Liu et al., demonstrated that a 3D electroconductive gelatin methacryloyl-multiwalled carbon nanotube/cobalt3D conductive hydrogel scaffolds accelerated the neural differentiation of SCAPs. Considering these aspects, the novel electroconductive GelMA-MWCNTs/Co hydrogel synergized with ES greatly promotes SCAPs neuronal differentiation [118]. Therefore, the use of scaffolds as delivery systems for growth factors, adhesion molecules and cytokines has recently received attention [119]. This condition would be particularly interesting for stem cell models such as DSCs. As reported so far, the combined application of technologies such as scaffolding materials and SCs is thought to have enormous potential for tissue regeneration [18].
DSCs Secretome
Currently, research is focusing on the therapeutic applications of DSCs, particularly in preventing and treating neurological diseases and nerve injuries [37]. The role played by these cells in the reparative process is mainly attributed to paracrine mechanisms mediated by their secreted factors [53]. Gnecchi et al., demonstrated that MSCs mediate their therapeutic effects through the secretion and release of trophic molecules known as “secretome”. This evidence allowed the development of the theory that the therapeutic applicability of MSCs in regenerative medicine is based on their ability to homing to the site of tissue injury and differentiate into different functional cell types, leading to tissue repair [120]. Interestingly, some studies also suggested the superiority of the secretome obtained from DSCs compared to other MSCs sources, such as BM-MSCs and adipose-derived mesenchymal stem cells (AD-MSCs), for neuroprotection [121]. The secreted factors can be found in the cell culture medium called Conditioned Medium (CM) and in the Exosomes (Exos) they secrete [122], representing an attractive, noninvasive, and acellular tool for therapeutic approaches against various disorders [123]. A summary of the main factors found in the DSCs’ secretome is given in Table 2.
Conditioned Media and Exosomes in Neurological Disorders
DPSCs
It is important to note that donor age and in vitro microenvironmental conditions may also influence secretome composition. Indeed, DPSC-CM obtained under normoxic conditions was reported to be enriched in molecules with anti-inflammatory, tissue repair, and regenerative properties compared to CM obtained under hypoxic conditions [135]. Confirming that hypoxia appears to promote neuronal differentiation of DPSCs, Delle Monache et al. demonstrated that administration of DPSC-CM under hypoxic conditions resulted in neuronal differentiation of both SH-SY5Y neuroblastoma cells and undifferentiated DPSCs [23]. DPSC-CM promoted neurite outgrowth in dorsal root ganglion (DRG) neurons. In particular, the total length and joint number of neurites increased after CM treatment. Furthermore, DPSC-CM promotes Schwann cell viability and myelin formation [136]. DPSCs’ secretome also showed beneficial effects in models of neurodegenerative diseases. Ahmed et al., described that treatment with DPSCs’secretome reduced amyloid β (Aβ) cytotoxicity in an in vitro model of Alzheimer’s disease (AD), increasing cell viability and reducing apoptosis [137]. Wang et al., demonstrated for the first time the therapeutic benefits of DPSC-CM in amyotrophic lateral sclerosis (ALS) with beneficial effects on direct motor neuron protection and lifespan prolongation [138]. The protective effect of DPSC-CM has also been demonstrated in the treatment of experimental autoimmune encephalomyelitis (EAE), the main murine model of multiple sclerosis (MS). Shimojima et al., showed that DPSC-CM administration reduced the expression of inflammatory cytokines in the spinal cord, inhibited demyelination, and improved clinical scores of EAE [139]. DPSC-Exos may be more suitable for the treatment of neurodegenerative diseases than MSCs derived from mesodermal tissues [138]. They may be an effective drug carrier for the treatment of various diseases, especially neurological disorders such as PD [140, 141].
SHEDs
The SHED-CM strongly promoted the expression of anti-inflammatory cytokines, angiogenic and anti-apoptotic factors [142], making this a potential therapeutic tool in neurological disorders. Different studies reported beneficial effects of SHED-CM both in vitro and in vivo PD models [143]. Mita et al., demonstrated that SHED-CM attenuated pro-inflammatory responses induced by β-amyloid plaques and generated an anti-inflammatory/tissue regeneration environment, which was accompanied by the induction of M2-like anti-inflammatory microglia, suggesting that SHED-CM can provide significant therapeutic benefits for AD [144]. Matsubara et al., demonstrated that SHED-CM, administered intrathecally into injured spinal cord in the rat during the acute postinjury period, caused a remarkable functional recovery related to the induction of anti-inflammatory M2 macrophage [145]. Miura-Yura et al., described that SHED-CM significantly promoted neurite outgrowth of dorsal root ganglion neurons compared to basal DMEM indicating that SHED-CM might have a therapeutic effect on diabetic polyneuropathy through the promotion of neurite outgrowth, and the increase in capillaries may contribute to the improvement of neural function [146]. In an animal model of superior laryngeal nerve injury, the systemic administration of SHED-CM induced functional recovery, increasing the degree of myelination, and promoted axonal regeneration shifting macrophages toward the M2 phenotype [147]. Li et al., injected SHED-Exos into a traumatic brain injury (TBI) rat model and observed that SHED-Exos contributed to rat motor functional restoration and cortical lesion reduction by shifting microglia polarization [148]. Narbute et al., showed that SHED-Exos significantly improved the gait impairments and contralateral rotations in the unilateral 6-hydroxydopamine (6-OHDA) rat model of PD [149]. In terms of anti-inflammatory effect, SHED-Exos significantly suppressed the carrageenan-induced acute inflammation in vivo [150]. Similarly, Luo et al., showed that SHED-Exos markedly reduced the inflammation in chondrocytes derived from the temporomandibular joint through delivering miR-100-5p [151].
PDLSCs
PDLSCs secretome can reduce oxidative stress and inflammation in injured neurons and can increase the functionality of the PI3K/Akt/mTOR axis which results in restoring BDNF production. Moreover, the CM has a neuroprotective effect due to containing NT-3, and IL‐10, and the presence of growth factors and immunomodulatory cytokines [152]. PDLSC-CM is useful in enhancing long-term neuronal regeneration in spinal cord injury [153]. Rajan et al., showed that PDLSCs-CM obtained from patients with relapsing‐remitting MS (RR-MS) showed anti-inflammatory and antiapoptotic effects when injected in a mouse model of MS [129]. Interestingly, CM obtained from PDLSCs cultured under hypoxic conditions was efficacious in ameliorating clinical and histological disease scores in EAE mice. This treatment reduced inflammatory cell infiltration and increased remyelination in the spinal cord.
SCAPs
Yu et al., profiled the secretome of human SCAPs by comparing it to that of BM-MSCs. A total of 2,046 proteins were detected in the SCAP-CM. Chemokines were included as well as angiogenic, immunomodulatory, anti-apoptotic and neuroprotective factors and ECM proteins [133]. In another study, Yu et al., compared the osteo/odontogenic, angiogenic, and neurogenic effects of soluble factors released from SCAPs and BM-MSCs in vitro on the proliferation and differentiation of dental pulp cells (DPCs). They demonstrated that CM released from SCAPs had a greater osteo/odontogenic and neurogenic inductive effect on DPCs than BM-MSCs-CM. This indicates that SCAPs-CM may serve as an additive to enhance pulp tissue repair and regeneration [154].
GMSCs
Rajan et al., demonstrated the presence of NGF, NT-3, IL-10 and TGF-β in GMSC-CM, which provide neuroprotection in scratch-damaged motor-neuron-like NSC-34 cells suggesting a potential therapeutic application of GMSC-CM in motor neuron degenerative diseases [134]. Although few studies are currently available on the potential therapeutic applications of GMSC-Exos, they hold a great promise for tissue regeneration. Jiang and Xu demonstrated that GMSC-Exos facilitated the osteogenic differentiation of MC3T3-E1 cells [155]. In a high-lipid microenvironment, GMSC-Exos suppressed lipid accumulation, transformed pro-inflammatory macrophages to an anti-inflammatory phenotype, and decrease the secretion and expression of inflammatory factors including IL-6, IL-1β, TNF-α, and cluster of differentiation [156]. Rao et al., showed that GMSC-Exos enhanced the proliferation of Schwann cells and the growth of the dorsal root ganglion neuron axon as well as promoting the formation of nerve fibers and myelin, which subsequently contributed to the recovery of motor skills, nerve conduction function, and muscle movement [157].
Conclusion
DSCs, have been shown to possess a remarkable neuroregenerative potential due to their neural crest origin. Not only the cells but also their secretome exhibit the same enhanced neuroprotective and neuroregenerative properties. The studies evaluated in this review have shown that both CM and Exos contain neurotrophins and molecules with neuroprotective action, even at higher levels than other MSCs. DSC-CM and DSC-Exos stimulated neurite outgrowth and exhibited neuroprotective effects in preclinical models of neurological disease and neuronal injury. DSC-CM represent an attractive, non-invasive, and acellular tool for therapeutic approaches against various disorders. DSC-Exos possesses unique advantages such as high drug loading capacity, high specificity, low immunogenicity, excellent biocompatibility, ease of obtaining, low side effects and nanometer size. DSC-Exos is emerging as a promising and practical therapeutic approach for tissues repair and regeneration. Currently, DPSC and SHED secretomes are the most studied. However, several studies have also highlighted the neuroprotective effects of PDLSC and GMSC secretomes. Interestingly, some studies have also suggested the superiority of DSCs derived secretome over other MSCs sources, such as BM-MSCs and AD-MSCs, for neuroprotection. Furthermore, the mechanical properties of the substrate to which the cells are attached are fundamental fin regulating cellular mechano-transduction and the subsequent cellular behavior, especially when further technologies allow the substrate itself to become biologically active. This has important implications for development, differentiation, disease, and regeneration. The proliferation, paracrine effect, and multidirectional differentiation potential of DSCs support the application of DSCs in regenerative medicine [18]. In conclusion, DSC secretomes in combination or not with biomaterials and biological scaffold are emerging as a promising and practical therapeutic approach for the repair and regeneration of different tissues, especially in the neuroregenerative field. It is thought to be useful for the development of new neuroprotective therapies.
Data Availability
Not applicable.
Code Availability
Not applicable.
Abbreviations
- SCs:
-
Stem cells
- MSCs:
-
Mesenchymal stem cells
- DSCs:
-
Dental-derived MSCs
- DPSCs:
-
Dental pulp stem cells
- SHEDs:
-
Stem cells from human exfoliated deciduous teeth
- PDLSCs:
-
Periodontal ligament stem cells
- DFSCs:
-
Dental follicle stem cells
- SCAPs:
-
Stem cells from apical papilla
- GMSCs:
-
Gingival MSCs
- CM:
-
Conditioned medium
- Exos:
-
Exosomes
References
Zakrzewski, W., Dobrzyński, M., Szymonowicz, M., & Rybak, Z. (2019). Stem cells: Past, present, and future. Stem Cell Research & Therapy, 10(1), 68. https://doi.org/10.1186/s13287-019-1165-5
Friedenstein, A. J., Petrakova, K. V., Kurolesova, A. I., & Frolova, G. P. (1968). Heterotopic of bone marrow. Analysis of precursor cells for osteogenic and hematopoietic tissues. Transplantation, 6(2), 230–247.
Badyra, B., Sułkowski, M., Milczarek, O., & Majka, M. (2020). Mesenchymal stem cells as a multimodal treatment for nervous system diseases. Stem Cells Translational Medicine, 9(10), 1174–1189. https://doi.org/10.1002/sctm.19-0430
Gronthos, S., Mankani, M., Brahim, J., Robey, P. G., & Shi, S. (2000). Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proceedings of the National Academy of Sciences of the United States of America, 97(25), 13625–13630. https://doi.org/10.1073/pnas.240309797
Miura, M., Gronthos, S., Zhao, M., Lu, B., Fisher, L. W., Robey, P. G., & Shi, S. (2003). SHED: Stem cells from human exfoliated deciduous teeth. Proceedings of the National Academy of Sciences of the United States of America, 100(10), 5807–5812. https://doi.org/10.1073/pnas.0937635100
Seo, B. M., Miura, M., Gronthos, S., Bartold, P. M., Batouli, S., Brahim, J., Young, M., Robey, P. G., Wang, C. Y., & Shi, S. (2004). Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet (London, England), 364(9429), 149–155. https://doi.org/10.1016/S0140-6736(04)16627-0
Zhang, Q. Z., Nguyen, A. L., Yu, W. H., & Le, A. D. (2012). Human oral mucosa and gingiva: A unique reservoir for mesenchymal stem cells. Journal of Dental Research, 91(11), 1011–1018. https://doi.org/10.1177/0022034512461016
Huang, G. T., Sonoyama, W., Liu, Y., Liu, H., Wang, S., & Shi, S. (2008). The hidden treasure in apical papilla: The potential role in pulp/dentin regeneration and bioroot engineering. Journal of Endodontics, 34(6), 645–651. https://doi.org/10.1016/j.joen.2008.03.001
Morsczeck, C., Götz, W., Schierholz, J., Zeilhofer, F., Kühn, U., Möhl, C., Sippel, C., & Hoffmann, K. H. (2005). Isolation of precursor cells (PCs) from human dental follicle of wisdom teeth. Matrix Biology: Journal of the International Society for Matrix Biology, 24(2), 155–165. https://doi.org/10.1016/j.matbio.2004.12.004
Pisciotta, A., Bertoni, L., Vallarola, A., Bertani, G., Mecugni, D., & Carnevale, G. (2020). Neural crest derived stem cells from dental pulp and tooth-associated stem cells for peripheral nerve regeneration. Neural Regeneration Research, 15(3), 373–381. https://doi.org/10.4103/1673-5374.266043
Anoop, M., & Datta, I. (2021). Stem cells derived from human exfoliated deciduous teeth (SHED) in neuronal disorders: A review. Current Stem Cell Research & Therapy, 16(5), 535–550. https://doi.org/10.2174/1574888X16666201221151512
Xiao, Z., Lei, T., Liu, Y., Yang, Y., Bi, W., & Du, H. (2021). The potential therapy with dental tissue-derived mesenchymal stem cells in Parkinson’s disease. Stem Cell Research & Therapy, 12(1), 5. https://doi.org/10.1186/s13287-020-01957-4
de la Fuente, A. G., Pelucchi, S., Mertens, J., Di Luca, M., Mauceri, D., & Marcello, E. (2023). Novel therapeutic approaches to target neurodegeneration. British Journal of Pharmacology, 180(13), 1651–1673. https://doi.org/10.1111/bph.16078
El-Kadiry, A. E., Rafei, M., & Shammaa, R. (2021). Cell therapy: Types, regulation, and clinical benefits. Frontiers in Medicine, 8, 756029. https://doi.org/10.3389/fmed.2021.756029
Egusa, H., Sonoyama, W., Nishimura, M., Atsuta, I., & Akiyama, K. (2012). Stem cells in dentistry-part I: Stem cell sources. Journal of Prosthodontic Research, 56(3), 151–165. https://doi.org/10.1016/j.jpor.2012.06.001
Gan, L., Liu, Y., Cui, D., Pan, Y., Zheng, L., & Wan, M. (2020). Dental tissue-derived human mesenchymal stem cells and their potential in therapeutic application. Stem Cells International, 2020, 8864572. https://doi.org/10.1155/2020/8864572
Yin, J. Y., Luo, X. H., Feng, W. Q., Miao, S. H., Ning, T. T., Lei, Q., Jiang, T., & Ma, D. D. (2021). Multidifferentiation potential of dental-derived stem cells. World Journal of Stem Cells, 13(5), 342–365. https://doi.org/10.4252/wjsc.v13.i5.342
Song, W. P., Jin, L. Y., Zhu, M. D., Wang, H., & Xia, D. S. (2023). Clinical trials using dental stem cells: 2022 update. World Journal of Stem Cells, 15(3), 31–51. https://doi.org/10.4252/wjsc.v15.i3.31
Al Madhoun, A., Sindhu, S., Haddad, D., Atari, M., Ahmad, R., & Al-Mulla, F. (2021). Dental pulp stem cells derived from adult human third molar tooth: A brief review. Frontiers in Cell and Developmental Biology, 9, 717624. https://doi.org/10.3389/fcell.2021.717624
Vu, H. T., Han, M. R., Lee, J. H., Kim, J. S., Shin, J. S., Yoon, J. Y., Park, J. H., Dashnyam, K., Knowles, J. C., Lee, H. H., Kim, J. B., & Lee, J. H. (2022). Investigating the effects of conditioned media from stem cells of human exfoliated deciduous teeth on dental pulp stem cells. Biomedicines, 10(4), 906. https://doi.org/10.3390/biomedicines10040906
Santilli, F., Fabrizi, J., Pulcini, F., Santacroce, C., Sorice, M., Delle Monache, S., & Mattei, V. (2022). Gangliosides and their role in multilineage differentiation of mesenchymal stem cells. Biomedicines, 10(12), 3112. https://doi.org/10.3390/biomedicines10123112
Aydin, S., & Şahin, F. (2019). Stem cells derived from dental tissues. Advances in Experimental Medicine and Biology, 1144, 123–132. https://doi.org/10.1007/5584_2018_333
Delle Monache, S., Pulcini, F., Santilli, F., Martellucci, S., Santacroce, C., Fabrizi, J., Angelucci, A., Sorice, M., & Mattei, V. (2022). Hypoxia induces DPSC differentiation versus a neurogenic phenotype by the paracrine mechanism. Biomedicines, 10(5), 1056. https://doi.org/10.3390/biomedicines10051056
Shi, S., Robey, P. G., & Gronthos, S. (2001). Comparison of human dental pulp and bone marrow stromal stem cells by cDNA microarray analysis. Bone, 29(6), 532–539. https://doi.org/10.1016/s8756-3282(01)00612-3
Zhang, W., Walboomers, X. F., Shi, S., Fan, M., & Jansen, J. A. (2006). Multilineage differentiation potential of stem cells derived from human dental pulp after cryopreservation. Tissue Engineering, 12(10), 2813–2823. https://doi.org/10.1089/ten.2006.12.2813
Bar, J. K., Lis-Nawara, A., & Grelewski, P. G. (2021). Dental pulp stem cell-derived secretome and its regenerative potential. International Journal of Molecular Sciences, 22(21), 12018. https://doi.org/10.3390/ijms222112018
Hu, L., Zhao, B., Gao, Z., Xu, J., Fan, Z., Zhang, C., Wang, J., & Wang, S. (2020). Regeneration characteristics of different dental derived stem cell sheets. Journal of Oral Rehabilitation, 47(Suppl 1), 66–72. https://doi.org/10.1111/joor.12839
Gronthos, S., Brahim, J., Li, W., Fisher, L. W., Cherman, N., Boyde, A., DenBesten, P., Robey, P. G., & Shi, S. (2002). Stem cell properties of human dental pulp stem cells. Journal of Dental Research, 81(8), 531–535. https://doi.org/10.1177/154405910208100806
Martinez Saez, D., Sasaki, R. T., Neves, A. D., & da Silva, M. C. (2016). Stem cells from human exfoliated deciduous teeth: A growing literature. Cells, Tissues, Organs, 202(5–6), 269–280. https://doi.org/10.1159/000447055
Seo, B. M., Sonoyama, W., Yamaza, T., Coppe, C., Kikuiri, T., Akiyama, K., Lee, J. S., & Shi, S. (2008). SHED repair critical-size calvarial defects in mice. Oral Diseases, 14(5), 428–434. https://doi.org/10.1111/j.1601-0825.2007.01396.x
Hochuli, A. H. D., Senegaglia, A. C., Selenko, A. H., Fracaro, L., & Brofman, P. R. S. (2021). Dental Pulp from human exfoliated deciduous teeth-derived stromal cells demonstrated neuronal potential: In vivo and in vitro studies. Current Stem Cell Research & Therapy, 16(5), 495–506. https://doi.org/10.2174/1574888X16666210215160402
Trubiani, O., Di Primio, R., Traini, T., Pizzicannella, J., Scarano, A., Piattelli, A., & Caputi, S. (2005). Morphological and cytofluorimetric analysis of adult mesenchymal stem cells expanded ex vivo from periodontal ligament. International Journal of Immunopathology and Pharmacology, 18(2), 213–221. https://doi.org/10.1177/039463200501800204
Mohebichamkhorami, F., Fattahi, R., Niknam, Z., Aliashrafi, M., Khakpour Naeimi, S., Gilanchi, S., & Zali, H. (2022). Periodontal ligament stem cells as a promising therapeutic target for neural damage. Stem Cell Research & Therapy, 13(1), 273. https://doi.org/10.1186/s13287-022-02942-9
Trubiani, O., Pizzicannella, J., Caputi, S., Marchisio, M., Mazzon, E., Paganelli, R., Paganelli, A., & Diomede, F. (2019). Periodontal ligament stem cells: Current knowledge and future perspectives. Stem Cells and Development, 28(15), 995–1003. https://doi.org/10.1089/scd.2019.0025
Kawanabe, N., Murata, S., Murakami, K., Ishihara, Y., Hayano, S., Kurosaka, H., Kamioka, H., Takano-Yamamoto, T., & Yamashiro, T. (2010). Isolation of multipotent stem cells in human periodontal ligament using stage-specific embryonic antigen-4. Differentiation; Research in Biological Diversity, 79(2), 74–83. https://doi.org/10.1016/j.diff.2009.10.005
Bi, R., Lyu, P., Song, Y., Li, P., Song, D., Cui, C., & Fan, Y. (2021). Function of dental follicle progenitor/stem cells and their potential in regenerative medicine: From mechanisms to applications. Biomolecules, 11(7), 997. https://doi.org/10.3390/biom11070997
Mai, Z., Chen, H., Ye, Y., Hu, Z., Sun, W., Cui, L., & Zhao, X. (2021). Translational and clinical applications of dental stem cell-derived exosomes. Frontiers in Genetics, 12, 750990. https://doi.org/10.3389/fgene.2021.750990
Lima, R. L., Holanda-Afonso, R. C., Moura-Neto, V., Bolognese, A. M., DosSantos, M. F., & Souza, M. M. (2017). Human dental follicle cells express embryonic, mesenchymal and neural stem cells markers. Archives of Oral Biology, 73, 121–128. https://doi.org/10.1016/j.archoralbio.2016.10.003
Zhu, Y., Zhang, P., Gu, R. L., Liu, Y. S., & Zhou, Y. S. (2018). Origin and clinical applications of neural crest-derived dental stem cells. The Chinese Journal of Dental Research, 21(2), 89–100. https://doi.org/10.3290/j.cjdr.a40435
Sonoyama, W., Liu, Y., Yamaza, T., Tuan, R. S., Wang, S., Shi, S., & Huang, G. T. (2008). Characterization of the apical papilla and its residing stem cells from human immature permanent teeth: A pilot study. Journal of Endodontics, 34(2), 166–171. https://doi.org/10.1016/j.joen.2007.11.021
Han, C., Yang, Z., Zhou, W., Jin, F., Song, Y., Wang, Y., Huo, N., Chen, L., Qian, H., Hou, R., Duan, Y., & Jin, Y. (2010). Periapical follicle stem cell: A promising candidate for cementum/periodontal ligament regeneration and bio-root engineering. Stem Cells and Development, 19(9), 1405–1415. https://doi.org/10.1089/scd.2009.0277
Paganelli, A., Trubiani, O., Diomede, F., Pisciotta, A., & Paganelli, R. (2021). Immunomodulating profile of dental mesenchymal stromal cells: A Comprehensive overview. Frontiers in Oral Health, 2, 635055. https://doi.org/10.3389/froh.2021.635055
Kang, J., Fan, W., Deng, Q., He, H., & Huang, F. (2019). Stem cells from the apical papilla: A promising source for stem cell-based therapy. BioMed Research International, 2019, 6104738. https://doi.org/10.1155/2019/6104738
Zhang, Q., Shi, S., Liu, Y., Uyanne, J., Shi, Y., Shi, S., & Le, A. D. (2009). Mesenchymal stem cells derived from human gingiva are capable of immunomodulatory functions and ameliorate inflammation-related tissue destruction in experimental colitis. Journal of Immunology (Baltimore, Md.: 1950), 183(12), 7787–7798. https://doi.org/10.4049/jimmunol.0902318
Ji, X., Zhang, Z., Han, Y., Song, J., Xu, X., Jin, J., Su, S., Mu, D., Liu, X., Xu, S., Cui, H., Zhao, Z., Wang, Y., & Liu, H. (2016). Mesenchymal stem cells derived from normal gingival tissue inhibit the proliferation of oral cancer cells in vitro and in vivo. International Journal of Oncology, 49(5), 2011–2022. https://doi.org/10.3892/ijo.2016.3715
Kim, D., Lee, A. E., Xu, Q., Zhang, Q., & Le, A. D. (2021). Gingiva-derived mesenchymal stem cells: Potential application in tissue engineering and regenerative medicine - a comprehensive review. Frontiers in Immunology, 12, 667221. https://doi.org/10.3389/fimmu.2021.667221
Subba, T. A., Varma, S., Thomas, B., Rao, S., Kumar, M., Talwar, A., & Shashidhar, K. (2022). Comparison of cellular and differentiation characteristics of mesenchymal stem cells derived from human gingiva and periodontal ligament. Journal of International Society of Preventive & Community Dentistry, 12(2), 235–244. https://doi.org/10.4103/jispcd.JISPCD_259_21
Yamada, Y., Nakamura-Yamada, S., Kusano, K., & Baba, S. (2019). Clinical potential and current progress of dental pulp stem cells for various systemic diseases in regenerative medicine: A concise review. International Journal of Molecular Sciences, 20(5), 1132. https://doi.org/10.3390/ijms20051132
Martellucci, S., Santacroce, C., Manganelli, V., Santilli, F., Piccoli, L., Cassetta, M., Misasi, R., Sorice, M., & Mattei, V. (2019). Isolation, propagation, and prion protein expression during neuronal differentiation of human dental pulp stem cells. Journal of Visualized Experiments: JoVE, (145), https://doi.org/10.3791/59282. https://doi.org/10.3791/59282.
Delle Monache, S., Pulcini, F., Frosini, R., Mattei, V., Talesa, V. N., & Antognelli, C. (2021). Methylglyoxal-dependent glycative stress is prevented by the natural antioxidant oleuropein in human dental pulp stem cells through Nrf2/Glo1 pathway. Antioxidants (Basel, Switzerland), 10(5), 716. https://doi.org/10.3390/antiox10050716
Stevens, A., Zuliani, T., Olejnik, C., LeRoy, H., Obriot, H., Kerr-Conte, J., Formstecher, P., Bailliez, Y., & Polakowska, R. R. (2008). Human dental pulp stem cells differentiate into neural crest-derived melanocytes and have label-retaining and sphere-forming abilities. Stem Cells and Development, 17(6), 1175–1184. https://doi.org/10.1089/scd.2008.0012
Ratajczak, J., Bronckaers, A., Dillen, Y., Gervois, P., Vangansewinkel, T., Driesen, R. B., Wolfs, E., Lambrichts, I., & Hilkens, P. (2016). The neurovascular properties of dental stem cells and their importance in dental tissue engineering. Stem Cells International, 2016, 9762871. https://doi.org/10.1155/2016/9762871
Mattei, V., Martellucci, S., Pulcini, F., Santilli, F., Sorice, M., & Delle Monache, S. (2021). Regenerative potential of DPSCs and revascularization: Direct, paracrine or autocrine effect? Stem Cell Reviews and Reports, 17(5), 1635–1646. https://doi.org/10.1007/s12015-021-10162-6
Nuti, N., Corallo, C., Chan, B. M., Ferrari, M., & Gerami-Naini, B. (2016). Multipotent differentiation of human dental pulp stem cells: A literature review. Stem Cell Reviews and Reports, 12(5), 511–523. https://doi.org/10.1007/s12015-016-9661-9
Ching, H. S., Luddin, N., Rahman, I. A., & Ponnuraj, K. T. (2017). Expression of odontogenic and osteogenic markers in DPSCs and SHED: A review. Current Stem Cell Research & Therapy, 12(1), 71–79. https://doi.org/10.2174/1574888x11666160815095733
Mortada, I., & Mortada, R. (2018). Dental pulp stem cells and osteogenesis: An update. Cytotechnology, 70(5), 1479–1486. https://doi.org/10.1007/s10616-018-0225-5
Hu, L., Liu, Y., & Wang, S. (2018). Stem cell-based tooth and periodontal regeneration. Oral Diseases, 24(5), 696–705. https://doi.org/10.1111/odi.12703
Martellucci, S., Manganelli, V., Santacroce, C., Santilli, F., Piccoli, L., Sorice, M., & Mattei, V. (2018). Role of Prion protein-EGFR multimolecular complex during neuronal differentiation of human dental pulp-derived stem cells. Prion, 12(2), 117–126. https://doi.org/10.1080/19336896.2018.1463797
Candelise, N., Santilli, F., Fabrizi, J., Caissutti, D., Spinello, Z., Moliterni, C., Lancia, L., Delle Monache, S., Mattei, V., & Misasi, R. (2023). The importance of stem cells isolated from human dental pulp and exfoliated deciduous teeth as therapeutic approach in nervous system pathologies. Cells, 12(13), 1686. https://doi.org/10.3390/cells12131686
Saei Arezoumand, K., Alizadeh, E., Pilehvar-Soltanahmadi, Y., Esmaeillou, M., & Zarghami, N. (2017). An overview on different strategies for the stemness maintenance of MSCs. Artificial Cells, Nanomedicine, and Biotechnology, 45(7), 1255–1271. https://doi.org/10.1080/21691401.2016.1246452
Rodas-Junco, B. A., & Villicaña, C. (2017). Dental pulp stem cells: Current advances in isolation, expansion and preservation. Tissue Engineering and Regenerative Medicine, 14(4), 333–347. https://doi.org/10.1007/s13770-017-0036-3
Abdel Moniem, E. M., El-Batran, M. M., Halawa, A. M., Gomaa, D. H., Eldeen, G. N., & Aly, R. M. (2019). Optimizing a serum-free/xeno-free culture medium for culturing and promoting the proliferation of human dental pulp stem cells. Stem Cell Investigation, 6, 15. https://doi.org/10.21037/sci.2019.06.05
Diomede, F., Rajan, T. S., Gatta, V., D’Aurora, M., Merciaro, I., Marchisio, M., Muttini, A., Caputi, S., Bramanti, P., Mazzon, E., & Trubiani, O. (2017). Stemness maintenance properties in human oral stem cells after long-term passage. Stem Cells International, 2017, 5651287. https://doi.org/10.1155/2017/5651287
Cordeiro, M. M., Dong, Z., Kaneko, T., Zhang, Z., Miyazawa, M., Shi, S., Smith, A. J., & Nör, J. E. (2008). Dental pulp tissue engineering with stem cells from exfoliated deciduous teeth. Journal of Endodontics, 34(8), 962–969. https://doi.org/10.1016/j.joen.2008.04.009
Majumdar, D., Kanafi, M., Bhonde, R., Gupta, P., & Datta, I. (2016). Differential Neuronal plasticity of dental pulp stem cells from exfoliated deciduous and permanent teeth towards dopaminergic neurons. Journal of Cellular Physiology, 231(9), 2048–2063. https://doi.org/10.1002/jcp.25314
Shi, X., Mao, J., & Liu, Y. (2020). Pulp stem cells derived from human permanent and deciduous teeth: Biological characteristics and therapeutic applications. Stem Cells Translational Medicine, 9(4), 445–464. https://doi.org/10.1002/sctm.19-0398
Lee, S. H., Looi, C. Y., Chong, P. P., Foo, J. B., Looi, Q. H., Ng, C. X., & Ibrahim, Z. (2021). Comparison of isolation, expansion and cryopreservation techniques to produce stem cells from human exfoliated deciduous teeth (SHED) with better regenerative potential. Current Stem Cell Research & Therapy, 16(5), 551–562. https://doi.org/10.2174/1574888X15666200928110923
Bhandi, S., Alkahtani, A., Mashyakhy, M., Abumelha, A. S., Albar, N. H. M., Renugalakshmi, A., Alkahtany, M. F., Robaian, A., Almeslet, A. S., Patil, V. R., Varadarajan, S., Balaji, T. M., Reda, R., Testarelli, L., & Patil, S. (2021). Effect of ascorbic acid on differentiation, secretome and stemness of stem cells from human exfoliated deciduous tooth (SHEDs). Journal of Personalized Medicine, 11(7), 589. https://doi.org/10.3390/jpm11070589
Xie, F., He, J., Chen, Y., Hu, Z., Qin, M., & Hui, T. (2020). Multi-lineage differentiation and clinical application of stem cells from exfoliated deciduous teeth. Human Cell, 33(2), 295–302. https://doi.org/10.1007/s13577-020-00323-z
Kanafi, M. M., Rajeshwari, Y. B., Gupta, S., Dadheech, N., Nair, P. D., Gupta, P. K., & Bhonde, R. R. (2013). Transplantation of islet-like cell clusters derived from human dental pulp stem cells restores normoglycemia in diabetic mice. Cytotherapy, 15(10), 1228–1236. https://doi.org/10.1016/j.jcyt.2013.05.008
Sukarawan, W., & Osathanon, T. (2017). Stem cells from human exfoliated deciduous teeth: Biology and therapeutic potential. IntechOpen. https://doi.org/10.5772/intechopen.68173
Oubenyahya, H. (2021). Stem cells from dental pulp of human exfoliated teeth: Current understanding and future challenges in dental tissue engineering. The Chinese Journal of Dental Research, 24(1), 9–20. https://doi.org/10.3290/j.cjdr.b1105867
Arora, V., Arora, P., & Munshi, A. K. (2009). Banking stem cells from human exfoliated deciduous teeth (SHED): Saving for the future. The Journal of Clinical Pediatric Dentistry, 33(4), 289–294. https://doi.org/10.17796/jcpd.33.4.y887672r0j703654
Wang, X., Sha, X. J., Li, G. H., Yang, F. S., Ji, K., Wen, L. Y., Liu, S. Y., Chen, L., Ding, Y., & Xuan, K. (2012). Comparative characterization of stem cells from human exfoliated deciduous teeth and dental pulp stem cells. Archives of Oral Biology, 57(9), 1231–1240. https://doi.org/10.1016/j.archoralbio.2012.02.014
Shi, S., Bartold, P. M., Miura, M., Seo, B. M., Robey, P. G., & Gronthos, S. (2005). The efficacy of mesenchymal stem cells to regenerate and repair dental structures. Orthodontics & Craniofacial Research, 8(3), 191–199. https://doi.org/10.1111/j.1601-6343.2005.00331.x
Ma, L., Makino, Y., Yamaza, H., Akiyama, K., Hoshino, Y., Song, G., Kukita, T., Nonaka, K., Shi, S., & Yamaza, T. (2012). Cryopreserved dental pulp tissues of exfoliated deciduous teeth is a feasible stem cell resource for regenerative medicine. PLoS ONE, 7(12), e51777. https://doi.org/10.1371/journal.pone.0051777
Zhang, X., Li, H., Sun, J., Luo, X., Yang, H., Xie, L., Yang, B., Guo, W., & Tian, W. (2017). Cell-derived micro-environment helps dental pulp stem cells promote dental pulp regeneration. Cell Proliferation, 50(5), e12361. https://doi.org/10.1111/cpr.12361
Cristaldi, M., Mauceri, R., Tomasello, L., Pizzo, G., Pizzolanti, G., Giordano, C., & Campisi, G. (2018). Dental pulp stem cells for bone tissue engineering: A review of the current literature and a look to the future. Regenerative Medicine. https://doi.org/10.2217/rme-2017-0112. Advance online publication.
Wang, H., Zhong, Q., Yang, T., Qi, Y., Fu, M., Yang, X., Qiao, L., Ling, Q., Liu, S., & Zhao, Y. (2018). Comparative characterization of SHED and DPSCs during extended cultivation in vitro. Molecular Medicine Reports, 17(5), 6551–6559. https://doi.org/10.3892/mmr.2018.8725
Naz, S., Khan, F. R., Khan, I., Zohra, R. R., Salim, A., Mohammed, N., & Ahmad, T. (2022). Comparative analysis of dental pulp stem cells and stem cells from human exfoliated teeth in terms of growth kinetics, immunophenotype, self-renewal and multi lineage differentiation potential for future perspective of calcified tissue regeneration. Pakistan Journal of Medical Sciences, 38(5), 1228–1237. https://doi.org/10.12669/pjms.38.5.5187
Lu, B. W., Liu, N., Xu, L. L., Shi, H. G., Zhang, Y., & Zhang, W. (2016). Difference of in vitro osteogenic differentiation and osteoclast capacity between stem cells from human exfoliated deciduous teeth and dental pulp stem cells. Nan Fang Yi Ke Da Xue Xue Bao = Journal of Southern Medical University, 36(2), 180–185.
Vishwanath, V. R., Nadig, R. R., Nadig, R., Prasanna, J. S., Karthik, J., & Pai, V. S. (2013). Differentiation of isolated and characterized human dental pulp stem cells and stem cells from human exfoliated deciduous teeth: An in vitro study. Journal of Conservative Dentistry: JCD, 16(5), 423–428. https://doi.org/10.4103/0972-0707.117509
Brar, G. S., & Toor, R. S. (2012). Dental stem cells: Dentinogenic, osteogenic, and neurogenic differentiation and its clinical cell-based therapies. Indian Journal of Dental Research: Official Publication of Indian Society for Dental Research, 23(3), 393–397. https://doi.org/10.4103/0970-9290.102239
Queiroz, A., Albuquerque-Souza, E., Gasparoni, L. M., de França, B. N., Pelissari, C., Trierveiler, M., & Holzhausen, M. (2021). Therapeutic potential of periodontal ligament stem cells. World Journal of Stem Cells, 13(6), 605–618. https://doi.org/10.4252/wjsc.v13.i6.605
Mata, M., Peydró, S., de Llano, J. J. M., Sancho-Tello, M., & Carda, C. (2022). Human dental pulp stem cells differentiate into cementoid-like-secreting cells on decellularized teeth scaffolds. International Journal of Molecular Sciences, 23(24), 15588. https://doi.org/10.3390/ijms232415588
Kumar, A., Kumar, V., Rattan, V., Jha, V., Pal, A., & Bhattacharyya, S. (2017). Molecular spectrum of secretome regulates the relative hepatogenic potential of mesenchymal stem cells from bone marrow and dental tissue. Scientific Reports, 7(1), 15015. https://doi.org/10.1038/s41598-017-14358-0
Wang, P., Zhao, H., Yao, Y., Lu, C., Ma, J., Chen, R., & Pan, J. (2020). Repair of facial nerve crush injury in rabbits using collagen plus basic fibroblast growth factor. Journal of Biomedical Materials Research Part A, 108(6), 1329–1337. https://doi.org/10.1002/jbm.a.36905
Askarzadeh, N., Nazarpak, M. H., Mansoori, K., Farokhi, M., Gholami, M., Mohammadi, J., & Mottaghitalab, F. (2020). Bilayer cylindrical conduit consisting of electrospun polycaprolactone nanofibers and DSC cross-linked sodium alginate hydrogel to bridge peripheral nerve gaps. Macromolecular Bioscience, 20(9), e2000149. https://doi.org/10.1002/mabi.202000149
Zorba Yildiz, A. P., Darici, H., Yavuz, B., Abamor, E. S., Ozdemir, C., Yasin, M. E., Bagirova, M., Allahverdiyev, A., & Karaoz, E. (2022). Preparation and characterization of graphene-based 3D biohybrid hydrogel bioink for peripheral neuroengineering. Journal of Visualized Experiments: JoVE, (183). https://doi.org/10.3791/63622. https://doi.org/10.3791/63622.
Su, W. T., & Pan, Y. J. (2016). Stem cells from human exfoliated deciduous teeth differentiate toward neural cells in a medium dynamically cultured with Schwann cells in a series of polydimethylsiloxanes scaffolds. Journal of Neural Engineering, 13(4), 046005. https://doi.org/10.1088/1741-2560/13/4/046005
Suhar, R. A., Marquardt, L. M., Song, S., Buabbas, H., Doulames, V. M., Johansson, P. K., Klett, K. C., Dewi, R. E., Enejder, A. M. K., Plant, G. W., George, P. M., & Heilshorn, S. C. (2021). Elastin-like proteins to support peripheral nerve regeneration in guidance conduits. ACS Biomaterials Science & Engineering, 7(9), 4209–4220. https://doi.org/10.1021/acsbiomaterials.0c01053
Luca, A. C., Raffoul, W., Giacalone, F., Bertolini, M., & Summa, P. G. (2015). Tissue-engineered constructs for peripheral nerve repair: Current research concepts and future perspectives. Plastic and Aesthetic Research, 2, 213–219.
Gregory, H., & Phillips, J. B. (2021). Materials for peripheral nerve repair constructs: Natural proteins or synthetic polymers? Neurochemistry International, 143, 104953. https://doi.org/10.1016/j.neuint.2020.104953
Wang, Y., Tan, H., & Hui, X. (2018). Biomaterial scaffolds in regenerative therapy of the central nervous system. BioMed Research International, 2018, 7848901. https://doi.org/10.1155/2018/7848901
Li, G., Zhao, X., Zhao, W., Zhang, L., Wang, C., Jiang, M., Gu, X., & Yang, Y. (2014). Porous chitosan scaffolds with surface micropatterning and inner porosity and their effects on Schwann cells. Biomaterials, 35(30), 8503–8513. https://doi.org/10.1016/j.biomaterials.2014.05.093
Mobasseri, S. A., Terenghi, G., & Downes, S. (2014). Schwann cell interactions with polymer films are affected by groove geometry and film hydrophilicity. Biomedical Materials (Bristol, England), 9(5), 055004. https://doi.org/10.1088/1748-6041/9/5/055004
Shahriari, D., Shibayama, M., Lynam, D. A., Wolf, K. J., Kubota, G., Koffler, J. Y., Tuszynski, M. H., Campana, W. M., & Sakamoto, J. S. (2017). Peripheral nerve growth within a hydrogel microchannel scaffold supported by a kink-resistant conduit. Journal of Biomedical Materials Research Part A, 105(12), 3392–3399. https://doi.org/10.1002/jbm.a.36186
Bonnans, C., Chou, J., & Werb, Z. (2014). Remodelling the extracellular matrix in development and disease. Nature Reviews. Molecular Cell Biology, 15(12), 786–801. https://doi.org/10.1038/nrm3904
Nicolas, J., Magli, S., Rabbachin, L., Sampaolesi, S., Nicotra, F., & Russo, L. (2020). 3D extracellular matrix mimics: Fundamental concepts and role of materials chemistry to influence stem cell fate. Biomacromolecules, 21(6), 1968–1994. https://doi.org/10.1021/acs.biomac.0c00045
Das, S., & Bellare, J. R. (2020). Dental pulp stem cells in customized 3D nanofibrous scaffolds for regeneration of peripheral nervous system. Methods in Molecular Biology (Clifton, N.J.), 2125, 157–166. https://doi.org/10.1007/7651_2018_194
Mansouri, N., Al-Sarawi, S., Losic, D., Mazumdar, J., Clark, J., Gronthos, S., & O’Hare Doig, R. (2021). Biodegradable and biocompatible graphene-based scaffolds for functional neural tissue engineering: A strategy approach using dental pulp stem cells and biomaterials. Biotechnology and Bioengineering, 118(11), 4217–4230. https://doi.org/10.1002/bit.27891
Luo, L., Albashari, A. A., Wang, X., Jin, L., Zhang, Y., Zheng, L., Xia, J., Xu, H., Zhao, Y., Xiao, J., He, Y., & Ye, Q. (2018). Effects of transplanted heparin-poloxamer hydrogel combining dental pulp stem cells and bFGF on spinal cord injury repair. Stem Cells International, 2018, 2398521. https://doi.org/10.1155/2018/2398521
Sridharan, R., Kelly, D. J., & O’Brien, F. J. (2021). Substrate stiffness modulates the crosstalk between mesenchymal stem cells and macrophages. Journal of Biomechanical Engineering, 143(3), 031001. https://doi.org/10.1115/1.4048809
Srinivasan, A., Chang, S. Y., Zhang, S., Toh, W. S., & Toh, Y. C. (2018). Substrate stiffness modulates the multipotency of human neural crest derived ectomesenchymal stem cells via CD44 mediated PDGFR signaling. Biomaterials, 167, 153–167. https://doi.org/10.1016/j.biomaterials.2018.03.022
Powell, R., Eleftheriadou, D., Kellaway, S., & Phillips, J. B. (2021). Natural biomaterials as instructive engineered microenvironments that direct cellular function in peripheral nerve tissue engineering. Frontiers in Bioengineering and Biotechnology, 9, 674473. https://doi.org/10.3389/fbioe.2021.674473
Liu, Z., Tong, H., Li, J., Wang, L., Fan, X., Song, H., Yang, M., Wang, H., Jiang, X., Zhou, X., Yuan, H., & Wang, Y. (2022). Low-stiffness hydrogels promote peripheral nerve regeneration through the rapid release of exosomes. Frontiers in Bioengineering and Biotechnology, 10, 922570. https://doi.org/10.3389/fbioe.2022.922570
Guo, S., Redenski, I., Landau, S., Szklanny, A., Merdler, U., & Levenberg, S. (2020). Prevascularized Scaffolds bearing human dental pulp stem cells for treating complete spinal cord injury. Advanced Healthcare Materials, 9(20), e2000974. https://doi.org/10.1002/adhm.202000974
Madl, C. M., Katz, L. M., & Heilshorn, S. C. (2018). Tuning bulk hydrogel degradation by simultaneous control of proteolytic cleavage kinetics and hydrogel network architecture. ACS Macro Letters, 7(11), 1302–1307. https://doi.org/10.1021/acsmacrolett.8b00664
Fourniols, T., Randolph, L. D., Staub, A., Vanvarenberg, K., Leprince, J. G., Préat, V., des Rieux, A., & Danhier, F. (2015). Temozolomide-loaded photopolymerizable PEG-DMA-based hydrogel for the treatment of glioblastoma. Journal of Controlled Release: Official Journal of the Controlled Release Society, 210, 95–104. https://doi.org/10.1016/j.jconrel.2015.05.272
Johnson, P. J., Parker, S. R., & Sakiyama-Elbert, S. E. (2009). Controlled release of neurotrophin-3 from fibrin-based tissue engineering scaffolds enhances neural fiber sprouting following subacute spinal cord injury. Biotechnology and Bioengineering, 104(6), 1207–1214. https://doi.org/10.1002/bit.22476
Taylor, S. J., McDonald, J. W., 3rd., & Sakiyama-Elbert, S. E. (2004). Controlled release of neurotrophin-3 from fibrin gels for spinal cord injury. Journal of Controlled Release: Official Journal of the Controlled Release Society, 98(2), 281–294. https://doi.org/10.1016/j.jconrel.2004.05.003
Perez-Puyana, V., Wieringa, P., Yuste, Y., de la Portilla, F., Guererro, A., Romero, A., & Moroni, L. (2021). Fabrication of hybrid scaffolds obtained from combinations of PCL with gelatin or collagen via electrospinning for skeletal muscle tissue engineering. Journal of Biomedical Materials Research Part A, 109(9), 1600–1612. https://doi.org/10.1002/jbm.a.37156
Ansari, S., Chen, C., Xu, X., Annabi, N., Zadeh, H. H., Wu, B. M., Khademhosseini, A., Shi, S., & Moshaverinia, A. (2016). Muscle tissue engineering using gingival mesenchymal stem cells encapsulated in alginate hydrogels containing multiple growth factors. Annals of Biomedical Engineering, 44(6), 1908–1920. https://doi.org/10.1007/s10439-016-1594-6
Ueda, T., Inden, M., Ito, T., Kurita, H., & Hozumi, I. (2020). Characteristics and therapeutic potential of dental pulp stem cells on neurodegenerative diseases. Frontiers in Neuroscience, 14, 407. https://doi.org/10.3389/fnins.2020.00407
Procès, A., Luciano, M., Kalukula, Y., Ris, L., & Gabriele, S. (2022). Multiscale mechanobiology in brain physiology and diseases. Frontiers in Cell and Developmental Biology, 10, 823857. https://doi.org/10.3389/fcell.2022.823857
Luo, L., He, Y., Jin, L., Zhang, Y., Guastaldi, F. P., Albashari, A. A., Hu, F., Wang, X., Wang, L., Xiao, J., Li, L., Wang, J., Higuchi, A., & Ye, Q. (2020). Application of bioactive hydrogels combined with dental pulp stem cells for the repair of large gap peripheral nerve injuries. Bioactive Materials, 6(3), 638–654. https://doi.org/10.1016/j.bioactmat.2020.08.028
Li, X., Yang, C., Li, L., Xiong, J., Xie, L., Yang, B., Yu, M., Feng, L., Jiang, Z., Guo, W., & Tian, W. (2015). A therapeutic strategy for spinal cord defect: Human dental follicle cells combined with aligned PCL/PLGA electrospun material. BioMed Research International, 2015, 197183. https://doi.org/10.1155/2015/197183
Liu, J., Zou, T., Zhang, Y., Koh, J., Li, H., Wang, Y., Zhao, Y., & Zhang, C. (2022). Three-dimensional electroconductive carbon nanotube-based hydrogel scaffolds enhance neural differentiation of stem cells from apical papilla. Biomaterials Advances, 138, 212868. https://doi.org/10.1016/j.bioadv.2022.212868
Hosseinkhani, M., Mehrabani, D., Karimfar, M. H., Bakhtiyari, S., Manafi, A., & Shirazi, R. (2014). Tissue engineered scaffolds in regenerative medicine. World Journal of Plastic Surgery, 3(1), 3–7.
Gnecchi, M., He, H., Liang, O. D., Melo, L. G., Morello, F., Mu, H., Noiseux, N., Zhang, L., Pratt, R. E., Ingwall, J. S., & Dzau, V. J. (2005). Paracrine action accounts for marked protection of ischemic heart by Akt-modified mesenchymal stem cells. Nature Medicine, 11(4), 367–368. https://doi.org/10.1038/nm0405-367
Gugliandolo, A., & Mazzon, E. (2021). Dental mesenchymal stem cell secretome: An intriguing approach for neuroprotection and neuroregeneration. International Journal of Molecular Sciences, 23(1), 456. https://doi.org/10.3390/ijms23010456
Kichenbrand, C., Velot, E., Menu, P., & Moby, V. (2019). Dental pulp stem cell-derived conditioned medium: An attractive alternative for regenerative therapy. Tissue Engineering Part B: Reviews, 25(1), 78–88. https://doi.org/10.1089/ten.TEB.2018.0168
Ogata, K., Moriyama, M., Matsumura-Kawashima, M., Kawado, T., Yano, A., & Nakamura, S. (2022). The therapeutic potential of secreted factors from dental pulp stem cells for various diseases. Biomedicines, 10(5), 1049. https://doi.org/10.3390/biomedicines10051049
Caseiro, A. R., Santos Pedrosa, S., Ivanova, G., Vieira Branquinho, M., Almeida, A., Faria, F., Amorim, I., Pereira, T., & Maurício, A. C. (2019). Mesenchymal Stem/ Stromal Cells metabolomic and bioactive factors profiles: A comparative analysis on the umbilical cord and dental pulp derived Stem/ Stromal Cells secretome. PLoS ONE, 14(11), e0221378. https://doi.org/10.1371/journal.pone.0221378
Joo, K. H., Song, J. S., Kim, S., Lee, H. S., Jeon, M., Kim, S. O., & Lee, J. H. (2018). cytokine expression of stem cells originating from the apical complex and coronal pulp of immature teeth. Journal of Endodontics, 44(1), 87-92.e1. https://doi.org/10.1016/j.joen.2017.08.018
Konala, V. B. R., Bhonde, R., & Pal, R. (2020). Secretome studies of mesenchymal stromal cells (MSCs) isolated from three tissue sources reveal subtle differences in potency. In Vitro Cellular & Developmental Biology Animal, 56(9), 689–700. https://doi.org/10.1007/s11626-020-00501-1
Rajan, T. S., Giacoppo, S., Trubiani, O., Diomede, F., Piattelli, A., Bramanti, P., & Mazzon, E. (2016). Conditioned medium of periodontal ligament mesenchymal stem cells exert anti-inflammatory effects in lipopolysaccharide-activated mouse motoneurons. Experimental Cell Research, 349(1), 152–161. https://doi.org/10.1016/j.yexcr.2016.10.008
Ballerini, P., Diomede, F., Petragnani, N., Cicchitti, S., Merciaro, I., Cavalcanti, M. F. X. B., & Trubiani, O. (2017). Conditioned medium from relapsing-remitting multiple sclerosis patients reduces the expression and release of inflammatory cytokines induced by LPS-gingivalis in THP-1 and MO3.13 cell lines. Cytokine, 96, 261–272. https://doi.org/10.1016/j.cyto.2017.04.022
Rajan, T. S., Giacoppo, S., Diomede, F., Ballerini, P., Paolantonio, M., Marchisio, M., Piattelli, A., Bramanti, P., Mazzon, E., & Trubiani, O. (2016). The secretome of periodontal ligament stem cells from MS patients protects against EAE. Scientific Reports, 6, 38743. https://doi.org/10.1038/srep38743
Soundara Rajan, T., Giacoppo, S., Diomede, F., Bramanti, P., Trubiani, O., & Mazzon, E. (2017). Human periodontal ligament stem cells secretome from multiple sclerosis patients suppresses NALP3 inflammasome activation in experimental autoimmune encephalomyelitis. International Journal of Immunopathology and Pharmacology, 30(3), 238–252. https://doi.org/10.1177/0394632017722332
Giacoppo, S., Thangavelu, S. R., Diomede, F., Bramanti, P., Conti, P., Trubiani, O., & Mazzon, E. (2017). Anti-inflammatory effects of hypoxia-preconditioned human periodontal ligament cell secretome in an experimental model of multiple sclerosis: A key role of IL-37. FASEB Journal: Official Publication of the Federation of American Societies for Experimental Biology, 31(12), 5592–5608. https://doi.org/10.1096/fj.201700524R
Nagata, M., Iwasaki, K., Akazawa, K., Komaki, M., Yokoyama, N., Izumi, Y., & Morita, I. (2017). Conditioned medium from periodontal ligament stem cells enhances periodontal regeneration. Tissue Engineering Part A, 23(9–10), 367–377. https://doi.org/10.1089/ten.TEA.2016.0274
Yu, S., Zhao, Y., Ma, Y., & Ge, L. (2016). Profiling the secretome of human stem cells from dental apical papilla. Stem Cells and Development, 25(6), 499–508. https://doi.org/10.1089/scd.2015.0298
Rajan, T. S., Diomede, F., Bramanti, P., Trubiani, O., & Mazzon, E. (2017). Conditioned medium from human gingival mesenchymal stem cells protects motor-neuron-like NSC-34 cells against scratch-injury-induced cell death. International Journal of Immunopathology and Pharmacology, 30(4), 383–394. https://doi.org/10.1177/0394632017740976
Bousnaki, M., Bakopoulou, A., Pich, A., Papachristou, E., Kritis, A., & Koidis, P. (2022). Mapping the secretome of dental pulp stem cells under variable microenvironmental conditions. Stem Cell Reviews and Reports, 18(4), 1372–1407. https://doi.org/10.1007/s12015-021-10255-2
Omi, M., Hata, M., Nakamura, N., Miyabe, M., Ozawa, S., Nukada, H., Tsukamoto, M., Sango, K., Himeno, T., Kamiya, H., Nakamura, J., Takebe, J., Matsubara, T., & Naruse, K. (2017). Transplantation of dental pulp stem cells improves long-term diabetic polyneuropathy together with improvement of nerve morphometrical evaluation. Stem Cell Research & Therapy, 8(1), 279. https://doi.org/10.1186/s13287-017-0729-5
Ahmed, Nel-M., Murakami, M., Hirose, Y., & Nakashima, M. (2016). Therapeutic potential of dental pulp stem cell secretome for Alzheimer’s disease treatment: An in vitro study. Stem Cells International, 2016, 8102478. https://doi.org/10.1155/2016/8102478
Wang, J., Zuzzio, K., & Walker, C. L. (2019). Systemic dental pulp stem cell secretome therapy in a mouse model of amyotrophic lateral sclerosis. Brain Sciences, 9(7), 165. https://doi.org/10.3390/brainsci9070165
Shimojima, C., Takeuchi, H., Jin, S., Parajuli, B., Hattori, H., Suzumura, A., Hibi, H., Ueda, M., & Yamamoto, A. (2016). Conditioned medium from the stem cells of human exfoliated deciduous teeth ameliorates experimental autoimmune encephalomyelitis. Journal of Immunology (Baltimore, Md.: 1950), 196(10), 4164–4171. https://doi.org/10.4049/jimmunol.1501457
Haney, M. J., Klyachko, N. L., Zhao, Y., Gupta, R., Plotnikova, E. G., He, Z., Patel, T., Piroyan, A., Sokolsky, M., Kabanov, A. V., & Batrakova, E. V. (2015). Exosomes as drug delivery vehicles for Parkinson’s disease therapy. Journal of Controlled Release: Official Journal of the Controlled Release Society, 207, 18–30. https://doi.org/10.1016/j.jconrel.2015.03.033
Iezzi, I., Pagella, P., Mattioli-Belmonte, M., & Mitsiadis, T. A. (2019). The effects of ageing on dental pulp stem cells, the tooth longevity elixir. European Cells & Materials, 37, 175–185. https://doi.org/10.22203/eCM.v037a11
Matsushita, Y., Ishigami, M., Matsubara, K., Kondo, M., Wakayama, H., Goto, H., Ueda, M., & Yamamoto, A. (2017). Multifaceted therapeutic benefits of factors derived from stem cells from human exfoliated deciduous teeth for acute liver failure in rats. Journal of Tissue Engineering and Regenerative Medicine, 11(6), 1888–1896. https://doi.org/10.1002/term.2086
Fujii, H., Matsubara, K., Sakai, K., Ito, M., Ohno, K., Ueda, M., & Yamamoto, A. (2015). Dopaminergic differentiation of stem cells from human deciduous teeth and their therapeutic benefits for Parkinsonian rats. Brain Research, 1613, 59–72. https://doi.org/10.1016/j.brainres.2015.04.001
Mita, T., Furukawa-Hibi, Y., Takeuchi, H., Hattori, H., Yamada, K., Hibi, H., Ueda, M., & Yamamoto, A. (2015). Conditioned medium from the stem cells of human dental pulp improves cognitive function in a mouse model of Alzheimer’s disease. Behavioural Brain Research, 293, 189–197. https://doi.org/10.1016/j.bbr.2015.07.043
Matsubara, K., Matsushita, Y., Sakai, K., Kano, F., Kondo, M., Noda, M., Hashimoto, N., Imagama, S., Ishiguro, N., Suzumura, A., Ueda, M., Furukawa, K., & Yamamoto, A. (2015). Secreted ectodomain of sialic acid-binding Ig-like lectin-9 and monocyte chemoattractant protein-1 promote recovery after rat spinal cord injury by altering macrophage polarity. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 35(6), 2452–2464. https://doi.org/10.1523/JNEUROSCI.4088-14.2015
Miura-Yura, E., Tsunekawa, S., Naruse, K., Nakamura, N., Motegi, M., Nakai-Shimoda, H., Asano, S., Kato, M., Yamada, Y., Izumoto-Akita, T., Yamamoto, A., Himeno, T., Kondo, M., Kato, Y., Nakamura, J., & Kamiya, H. (2020). Secreted factors from cultured dental pulp stem cells promoted neurite outgrowth of dorsal root ganglion neurons and ameliorated neural functions in streptozotocin-induced diabetic mice. Journal of Diabetes Investigation, 11(1), 28–38. https://doi.org/10.1111/jdi.13085
Tsuruta, T., Sakai, K., Watanabe, J., Katagiri, W., & Hibi, H. (2018). Dental pulp-derived stem cell conditioned medium to regenerate peripheral nerves in a novel animal model of dysphagia. PLoS ONE, 13(12), e0208938. https://doi.org/10.1371/journal.pone.0208938
Li, Y., Yang, Y. Y., Ren, J. L., Xu, F., Chen, F. M., & Li, A. (2017). Exosomes secreted by stem cells from human exfoliated deciduous teeth contribute to functional recovery after traumatic brain injury by shifting microglia M1/M2 polarization in rats. Stem Cell Research & Therapy, 8(1), 198. https://doi.org/10.1186/s13287-017-0648-5
Narbute, K., Piļipenko, V., Pupure, J., Dzirkale, Z., Jonavičė, U., Tunaitis, V., Kriaučiūnaitė, K., Jarmalavičiūtė, A., Jansone, B., Kluša, V., & Pivoriūnas, A. (2019). Intranasal administration of extracellular vesicles derived from human teeth stem cells improves motor symptoms and normalizes tyrosine hydroxylase expression in the substantia Nigra and striatum of the 6-hydroxydopamine-treated rats. Stem Cells Translational Medicine, 8(5), 490–499. https://doi.org/10.1002/sctm.18-0162
Pivoraitė, U., Jarmalavičiūtė, A., Tunaitis, V., Ramanauskaitė, G., Vaitkuvienė, A., Kašėta, V., Biziulevičienė, G., Venalis, A., & Pivoriūnas, A. (2015). Exosomes from human dental pulp stem cells suppress carrageenan-induced acute inflammation in mice. Inflammation, 38(5), 1933–1941. https://doi.org/10.1007/s10753-015-0173-6
Luo, P., Jiang, C., Ji, P., Wang, M., & Xu, J. (2019). Exosomes of stem cells from human exfoliated deciduous teeth as an anti-inflammatory agent in temporomandibular joint chondrocytes via miR-100-5p/mTOR. Stem Cell Research & Therapy, 10(1), 216. https://doi.org/10.1186/s13287-019-1341-7
Pizzicannella, J., Gugliandolo, A., Orsini, T., Fontana, A., Ventrella, A., Mazzon, E., Bramanti, P., Diomede, F., & Trubiani, O. (2019). Engineered extracellular vesicles from human periodontal-ligament stem cells increase VEGF/VEGFR2 expression during bone regeneration. Frontiers in Physiology, 10, 512. https://doi.org/10.3389/fphys.2019.00512
Man, R. C., Sulaiman, N., Idrus, R. B. H., Ariffin, S. H. Z., Wahab, R. M. A., & Yazid, M. D. (2019). Insights into the effects of the dental stem cell secretome on nerve regeneration: Towards cell-free treatment. Stem Cells International, 2019, 4596150. https://doi.org/10.1155/2019/4596150
Yu, S., Zhao, Y., Fang, T. J., & Ge, L. (2020). Effect of the soluble factors released by dental apical papilla-derived stem cells on the osteo/odontogenic, angiogenic, and neurogenic differentiation of dental pulp cells. Stem Cells and Development, 29(12), 795–805. https://doi.org/10.1089/scd.2019.0262
Jiang, S., & Xu, L. (2020). Exosomes from gingival mesenchymal stem cells enhance migration and osteogenic differentiation of pre-osteoblasts. Die Pharmazie, 75(11), 576–580. https://doi.org/10.1691/ph.2020.0652
Zhang, Y., Shi, S., Xu, Q., Zhang, Q., Shanti, R. M., & Le, A. D. (2019). SIS-ECM laden with GMSC-derived exosomes promote taste bud regeneration. Journal of Dental Research, 98(2), 225–233. https://doi.org/10.1177/0022034518804531
Rao, F., Zhang, D., Fang, T., Lu, C., Wang, B., Ding, X., Wei, S., Zhang, Y., Pi, W., Xu, H., Wang, Y., Jiang, B., & Zhang, P. (2019). Exosomes from human gingiva-derived mesenchymal stem cells combined with biodegradable chitin conduits promote rat sciatic nerve regeneration. Stem Cells International, 2019, 2546367. https://doi.org/10.1155/2019/2546367
Funding
Open access funding provided by Università degli Studi dell’Aquila within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Contributions
Conceptualization, S.D.M., V.M.; writing-original draft preparation, F.S., J.F., C.S., D.C., Z.S., N.C., L.L., S.D.M., V.M.; writing-review and editing, F.S., S.D.M., V.M.; supervision, V.M, S.D.M.; All authors have read and agreed to the published version of the manuscript.” All the authors have contributed to its realization and have approved the submitted version.
Corresponding authors
Ethics declarations
Ethics Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Conflicts of Interest/Competing Interests
The authors declare that they have no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Santilli, F., Fabrizi, J., Santacroce, C. et al. Analogies and Differences Between Dental Stem Cells: Focus on Secretome in Combination with Scaffolds in Neurological Disorders. Stem Cell Rev and Rep 20, 159–174 (2024). https://doi.org/10.1007/s12015-023-10652-9
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12015-023-10652-9