
Abstract
The comparative responses of ten spring wheat cultivars to water stress were investigated. Wheat plants were cultured under hydroponics conditions (Hoagland nutrient) to the stage of three-leaf seedlings. Then, the water medium was supplemented with PEG (drought) or NaCl (salinity) to obtain a water status equal to −1.5 MPa. After a 2-day treatment, the changes in the following parameters were determined: fresh and dry weight, macro- and microelement accumulation, membrane injury (electrolyte leakage, lipid peroxidation) and fatty acid content of the phospholipid fraction of plasmalemma (in comparison to plants not stressed, taken as a control). Generally, the plants were more significantly influenced by water stress stimulated by PEG than by NaCl treatment, as compared to the plants cultivated in the control media. The results of the decrease in water content in leaves and electrolyte leakage from cells corresponded well with the intensity of lipid peroxidation (determined by malondialdehyde—MDA-content) and were chosen for the selection of investigated genotypes for tolerance to both stresses. The more tolerant genotypes exhibited the opposite changes in phospholipid fatty acid unsaturation for two applied stresses i.e. NaCl treatment caused a decrease in unsaturation whereas in PEG-treated plants an increase in unsaturation was observed. These changes were reversed for less tolerant plants, i.e. NaCl treatment influenced an increase in fatty acid unsaturation whereas in PEG-treated plants a decrease in unsaturation was measured. The ratio of U/S (unsaturated to saturated fatty acids) correlated with the total amount of accumulated macroelements. The content of Mg, Ca and S in leaves of plants undergoing both stress factors (NaCl and PEG) dropped whereas the K and P content increased in leaves of wheat seedlings cultured on media containing NaCl only. For microelements, a decrease in the accumulation of these nutrients was detected in all investigated seedlings. However, a greater reduction in the level of these elements occurred in seedlings grown on media with PEG in comparison to those grown on NaCl containing media.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Drought and high salinity have been identified as two of the major invasive environmental stresses triggering many common reactions in plants that lead to cellular dehydration with concomitant osmotic changes (Wang et al. 2009). However, under drought stress, the osmotic regulation in cells is mainly achieved by the synthesis of organic solutes, while under salt stress the accumulated solutes are predominantly inorganic sodium and chloride ions absorbed from soil (Parida and Das 2005). Osmotic stress is associated with oxidative stress and reactive oxygen species (ROS) production. At low concentrations, ROS can serve as signalling molecules in the redox signalling transduction pathway in plants, however, overproduction of ROS in stress conditions can damage cellular components including DNA, proteins and membrane lipids (Silva et al. 2010). In response, plants have evolved an efficient antioxidant system that protects them from the damaging effects caused by the oxidative stress (Asada 1999). The severity of stress depends on its magnitude and duration. Acute stress may cause a shock response while chronic stress may do more permanent damage. The first visual symptoms of osmotic stress are leaf wilting and rolling due to stomatal closure which leads to leaf death by desiccation during prolonged deficit of water (Kramer 1988). Thus, both factors stimulating osmotic stress (drought and salinity) can have similar negative effects on the yield and quality of the crops. Even though Poland and the neighbouring part of Europe are not generally characterized as drought-prone regions, periodic water deficit (and flooding) occur frequently. The frequency and scale of regional drought periods have increased, with an exacerbation of the situation projected for many parts of the world (Schar et al. 2004; Walter et al. 2011). The yield reduction due to these stress factors depends on the type of crop and its phenological phase (Martyniak et al. 2007). In Poland, the losses in agricultural production caused by environmental stresses reach 30% of grain yield. Spring varieties are more sensitive to drought occurring during spring season than winter cereals, which develop the root system before winter season and are thus able to penetrate the soil, reaching water reserves from snowmelts. The occurrence of water deficit in the early development stages of spring plant varieties dramatically reduces crop yield.
Wheat (Triticum aestivum) is one of the major crops grown in most countries, not only in Europe. Wheat starch has unique properties for bread-making that cannot be replaced by other starches, e.g., from corn or potato (Park et al. 2009). The selection procedures for species that are less sensitive to environmental stresses have been based on differences in their agronomic character, i.e. yield, survival, plant height, susceptibility to injury and infection, relative growth rate and relative growth reduction (Ashraf and Ali 2008; Franco et al. 1993; Munns 1993). Better understanding of the physiological mechanisms involved in stresses can be helpful for collecting more tolerant wheat genotypes. Mineral nutrients such as phosphorus, potassium and calcium and organic metabolites such as sugars, proteins, polyamines, water soluble and insoluble antioxidants are essential as distinctive indicators of tolerance at the whole plant and cellular levels (Ashraf and Ali 2008). Under drought and salinity stresses, the lowering of soil water potential reduces the diffusion of nutrients to the absorbing root surface (Hu et al. 2007). Nutrient transport to plants is also restricted by active ion transport and membrane permeability. Cell membrane permeability has long been taken as an indicator of the action of different stresses (Farooq and Azam 2006). Plasma membranes are the first receptors of stress and they can protect the cells through modifications affecting stress perception and rigidity of the cell structure. Both an increase and decrease in membrane fluidity were observed (Guerfel et al. 2008).
The aim of this work was to study the role of membranes and their permeability to micro- and macroelements after the short-term action of drought and salinity stresses in seedlings of spring wheat varieties with different tolerance to stress. The intensity of osmotic stress (−1.5 MPa) applied was chosen on the basis of preliminary tests. This level of stress causes visible effects (wilting and leaf rolling), but does not lead to irreversible changes during a 2-day application. Longer, 3-day treatment results in irreversible damage to plants. Although the short-term effects of water deficit on plant physiology are well documented (Rossi et al. 2009), the level of drought stress that established seedlings can endure and how this stress will affect changes in metabolic events is still unknown. The influence of drought and salinity was determined in leaves where the first symptoms of osmotic stress are particularly evident. Short (days) water stress reduces more strongly the growth of leaves than that of roots (Hsiao 1973). Moreover, our earlier experiments indicated that leaves of wheat can accumulate higher levels of ions than the root system under metal stress conditions (Filek et al. 2010).
It is difficult to quantify differences in water stress tolerance (especially during short-term water deficit) between closely related species on the basis of growth reduction only (Munns 2002). Thus, lipid membrane peroxidation and the leakage of electrolyte from cells were chosen as the main indicators of stress intensity. These investigations can add to the information on possible similarity/differences in membrane changes in drought and the salinity treatments of spring wheat varieties to select genotypes tolerant to both stresses. These studies will form the basis for further study of the mechanism of action of both stressors.
Materials and methods
Plant material and growth conditions
Seeds of ten Polish spring wheat genotypes (cv. Banti, Cytra, Jasna, Katoda, Monsun, Nawra, Parabola, Radunia, Raweta and Zadra) (after surface sterilization with 10% sodium hypochlorite solution followed by washing with sterilized distilled water) were germinated in 9-cm Petri dishes filled with one Whatman No.1 filter paper moistened with sterile half-strength Hoagland nutrient solution (Nagy and Galiba 1995). Twenty-five seeds per dish were used for each cultivar. The seeds were incubated in the dark in a temperature-controlled room at 20°C for 2 days. Seven uniform seedlings (with the same visual physiological features) of each genotype were placed in the holes of polystyrene plates (7 seedlings × 10 genotypes). These plates were suspended on the surface of polyethylene pots (39 cm × 28 cm × 14 cm, 11 dm3) filled with sterile half-strength Hoagland nutrient solution. The whole set was aerated using an aquarium pump. The experiment was carried out in a controlled environment with a 16-h photoperiod (17/20°C night/day) and 1,000 μmol(photon)m−2 s−1 light intensity (SQS, Hansatech Ltd, Kings Lynn, UK) and while the relative humidity in the growth chamber varied from 45 to 50% during the whole experimental period. The solution was renewed every 48 h under sterile conditions. After 2 weeks of culture (seedlings at 3-leaf stage), the nutrient was changed. To part of the pots, osmotic stress was imposed for 2 days by the application of PEG 600 (polyethylene glycol, 26% w/v) or NaCl (0.4 mol dm−3) in half-strength Hoagland nutrient solution. The osmotic potentials of both PEG and NaCl solutions were verified with a vapour pressure osmometer (Wescor Inc., Logan, UT, USA) and stabilized to −1.5 MPa. Another part of the pots not subjected to osmotic stress was taken as the control (0). After this time, 30 uniform seeds (from five pots) of each wheat genotype with 3 replications per salt and PEG treatment, as well as for the control, were used for the analysis of physiological and biochemical parameters.
The physiological parameters (fresh and dry weight) were measured in the upper part of seedlings after removing the root system. The fresh weight was measured immediately after separation of leaves from roots, whereas the dry weight was measured in samples oven-dried at 70°C for 48 h. The water content in leaves was calculated as the difference between fresh and dry matter (Romero-Aranda et al. 2006). Biochemical parameters were determined in the second leaf of the collected plants.
Electrolyte leakage determination
Sample disks (ϕ = 5 mm) were cut from fresh leaves, washed in sterile water (1 g of fresh weight in total) and introduced to test tubes containing 10 ml of deionised water. After gently shaking in a rotary shaker (120×g) at 20°C for 24 h in the dark, the electrical conductivity of the bathing solution (EL1) was measured with an Elmenton CX 505 conductometer (Zabrze, Poland). Then, the solution with leaf disks was boiled (90°C) for 10 min, until the destruction of membrane integrity, leading to the leakage of all the electrolyte from cells. After cooling to a temperature of 20°C and centrifugation to remove the plant material, electrical conductivity of the bathing solution was measured (EL2). Relative electrolyte leakage was calculated as (EL1/EL2) × 100%.
Protoplast isolation and analysis of fatty acids in plasma membrane phospholipids
About 1 g of fresh mass of leaves was cut into small pieces (about 1 mm long) and placed in a water solution containing 0.2 mmol dm−3 KH2PO4, 1 mmol dm−3 KNO3, 1 mmol dm−3 MgSO4, 10 mmol dm−3 CaCl2, 1 μmol dm−3 KI, 0.01 μmol dm−3 CuSO4, 0.6 mol dm−3 mannitol, 1% Celululase Onozuka R-10 (Yakult Honsa) and 0.8% Macerase R-10 (Yakult Honsha). The solution was stabilized at pH 5.6 (Liang et al. 2002; Zembala et al. 2010). Thus, prepared solutions were shaken (Heidolph Rotamax 120, Germany) at 10 g for 16 h in the dark (25°C) and then free protoplasts were collected by gentle pipetting. The protoplast solution was filtered through a 70-μm nylon mesh, centrifuged at 100g for 5 min and resuspended in a mixture of 0.6 mol dm−3 mannitol and 10 mmol dm−3 CaCl2. Plasmalemma was isolated in a two-phase system containing 6.5% Dextran T-500: 6.5% PEG 4 000 with 0.25 mmol dm−3 sucrose, 4 mmol dm−3 KCl and 5 mmol dm−3 buffer (KH2PO4 + K2HPO4, pH 7.8) at 4°C.
For lipid extraction, a modified method of Bligh and Dyer (1959) was used. Plasma membranes were first homogenized with boiling isopropanol, and then with chloroform:methanol (1:1, v:v) and chloroform. The fraction of phospholipids was separated on galactolipid and neural lipids using adsorption and distribution column chromatography on silica acid under low nitrogen pressure, and then purified by thin-layer chromatography (Block et al. 1983). Fatty acids were analyzed after transmethylation with 14% BF3 (in methanol solution) by gas chromatography (Hewlett Packard, USA) with a capillary column (30 m × 0.25 mm) at 170°C with 17:0 acid as an internal standard. Quantitative determination of fatty acids was performed using appropriate standards for each acid.
Determination of lipid peroxidation
Lipid peroxidation was determined according to Dhindsa et al. (1981). About 1 g of leaves was homogenized with 5 ml of 0.5% trichloroacetic acid (TCA) and after centrifugation at 1,000×g, 1 ml of the supernatant was mixed with 4 ml of 0.5% thiobarbituric acid (TBA) in 20% TCA. The mixture was boiled for 30 min and malondialdehyde (MDA) concentration, as the indicator of lipid peroxidation, was analyzed spectrophotometrically at room temperature at λ = 532 nm and non-specific background λ = 600 nm. The molar extinction coefficient was equal to 155 × 105 mmol−1 cm−1.
Micro- and macroelement analysis
The leaves were lyophilized (Freeze Dry System/Freezone 4.5, Labconco, USA) and samples of 0.02 or 0.11 g (for micro- and macroelements, respectively) were mineralized in ultrapure concentrated nitric acid (Merck, Damstad, Germany) in a closed microwave system (Uni Clever, Plazmatronika, Poland). The microelements were analyzed by ICP-MS spectrometry (Elan DRC-e, Perkin Elmer, Shelton, USA) in relation to standard isotopes: B(11), Mn(55), Fe(57), Cu(63), Zn(66), Mo(98). The determination of macroelements was performed by an ICP-AES spectrometer (Optima 2100, Perkin Elmer) at the following wavelengths: for K, λ = 766.490 nm; Ca, λ= 317.933 nm; Mg, λ = 285.213 nm; P, λ = 213.617 nm; S, λ = 181.975 nm and Na, λ = 589.592 nm (for details see Zembala et al. 2010).
Statistical analysis
All biochemical analyses were repeated three times and averaged (±SE). All the data were subjected to analysis of variance. The data were analyzed by Duncan’s Multiple Range test and t test at P < 0.05 using PC SAS 8.0. The significances between the means were assessed using the SAS ANOVA procedure. In addition, Pearson’ correlation coefficients were calculated for relationship between the weight of leaves of seedlings under stresses (NaCl and PEG) and those of control plants, and water uptake in leaves under both of water stresses, and between the fatty acid unsaturation (U/S ratio) and lipid peroxidation (MDA content) under both stresses, for 56 pairs of data. The statistical tests were run using STATISTICA 5.5. software.
Results
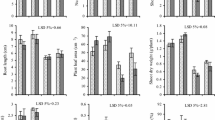
For the 10 spring wheat genotypes used in the experiments, the fresh weight of leaves varied between 0.47 and 0.32 g plant−1 in the control (non-water stress conditions, 0) applications (Fig. 1). The short-term (2 days) NaCl stress affected the leaves’ fresh weight negatively, but the effective changes differed among genotypes from about 13 to 24% in comparison to the control. Drought stress, initiated by PEG, influenced the plants more significantly than NaCl treatment, and caused around a 50% decrease in the fresh weight of seedlings as compared to the seedlings of the plants in the control media. The smallest decreases in leaf fresh weight under the action of both studied stresses were noted for the Katoda, Monsun and Nawra varieties, which were characterized by the lowest weight of control seedlings. Consequently, the greatest change was noted in the Radunia, Zadra and Cytra varieties with a relatively high weight of leaves of control seedlings, as compared to other varieties studied.

The fresh weight of leaves of 2-week seedlings (mg plant−1) from 10 spring wheat varieties after a culture on Hoagland (1:1 with water) media (0) and on media supplemented with NaCl (ψ = −1.5 MPa) and PEG (ψ = −1.5 MPa) for 2 days. Values are means of three independent experiments (n = 90, ±SE). Different letters indicate significant (P < 0.05) differences between treatments
Plants with less fresh weight generally accumulated less water (the difference between fresh and dry weight) (Fig. 2). In seedlings grown on control media, the differences between genotypes ranged from about 0.14 to 0.1 g plant−1. Water deficit caused by the application of PEG to the culture resulted in more than a twofold reduction in the water content in leaves of wheat seedlings, while the introduction of NaCl to the medium resulted in significantly smaller changes in this parameter. As a result of both these stress factors, the water level in the leaves of wheat seedlings ranged between 64 and 90% of that of the control, and between 5 and 21% of that of the control for plants cultured in NaCl and PEG treated media, respectively. The smallest changes occurred in Katoda, Monsun, Nawra and Parabola genotypes, whereas the greatest ones in Radunia, Banti and Cytra.

Water content (calculated as a difference between dry and fresh weight) in leaves of 2-week seedlings (mg plant−1) of 10 spring wheat varieties after a culture on Hoagland (1:1 with water) media (0) and on media supplemented with NaCl (ψ = −1.5 MPa) and PEG (ψ = −1.5 MPa) for 2 days. Values are means of three independent experiments (n = 90, ±SE). Different letters indicate significant (P < 0.05) differences between treatments
The leakage of electrolyte from the leaves of seedlings of the control genotypes, calculated relative to the total ion content in these tissues, was similar (about 7–9%) (Fig. 3). In the presence of NaCl, the leakage of electrolyte increased from two to three times compared to the controls, whereas in the presence of PEG in the medium, only by about 28–42%. The smallest changes were noted for Katoda, Monsun, Nawra and Parabola genotypes. Other genotypes were characterized by a significant leakage of electrolytes, however, the greatest effects were observed in Banti, Cytra, Raweta and Radunia genotypes under the action of both stress factors.

Relative electrolyte leakage (%) from leaves of 2-week seedlings of 10 spring wheat varieties after a culture on Hoagland (1:1 with water) media (0) and on media supplemented with NaCl (ψ = −1.5 MPa) and PEG (ψ = −1.5 MPa) for 2 days (for more details see “Materials and methods”). Values are means of three independent experiments (n = 30, ±SE). Different letters indicate significant (P < 0.05) differences between treatments
The values of MDA—the indicator of lipid peroxidation—were at similar levels in seedlings of the control (about 1.43–1.84 μmol g−1); the highest MDA contents were observed for Banti, Cytra and Jasna genotypes (Fig. 4). In cultures grown on media containing NaCl, the levels of MDA increased by 20–40%, while on media containing PEG, it increased about twice compared to the control. The smallest changes were observed in Katoda, Monsun, Nawra and Parabola genotypes, whereas the greatest ones in Radunia, Banti and Zadra genotypes.

Content of malondialdehyde (MDA), in μg per mg of fresh weight (FW), in leaves of 10 spring wheat varieties after a culture on Hoagland (1:1 with water) media (0) and on media supplemented with NaCl (ψ = −1.5 MPa) and PEG (ψ = −1.5 MPa) for 2 days. Values are means of three independent experiments (n = 30, ±SE). Different letters indicate significant (P < 0.05) differences between treatments
The phosholipid fraction of plasma membrane lipids of wheat seedlings contained mainly palmitic (16:0), strearic (18:0), oleic (18:1), linolic (18:2) and linoleic (18:3) acids (Table 1), but trace amounts of acids with less than 16 and more than 18 carbon atoms were also detected. In control conditions, the studied genotypes differed in the proportions of these acids. The lipid unsaturation index, calculated as the ratio of the sum of unsaturated (U) to the sum of saturated acids (S), ranged from 2.14 to 2.9 (Fig. 5). During NaCl treatment, the level of unsaturated fatty acids generally increased, whereas it decreased during PEG treatment. The U/S ratio (Fig. 5) indicates that only such genotypes as Monsun, Nawra and Parabola showed the opposite direction of changes, in comparison to other studied plants, in response to both stresses, i.e. a decrease in unsaturation in NaCl- and an increase in PEG-treated plants.

Ratio of unsaturated (U) to saturated (S) fatty acids of phospholipids in plasmalemma isolated from spring wheat leaves (2nd leaf) of seedlings cultured on media with NaCl (ψ = −1.5 MPa) and PEG (ψ = −1.5 MPa) and control (0, without NaCl and PEG). Values are means of three independent experiments (n = 30, ±SE). Different letters indicate significant (P < 0.05) differences between treatments
The content of macroelements in leaves of seedlings of the wheat genotypes studied that were grown on the control media was similar (Table 2). The total macroelement content was about 35–42 mg g−1 of dry weight (DW). In plants cultured in media containing NaCl, the smallest accumulation of this macroelement was observed for Katoda, Monsun, Nawra and Parabola genotypes and the highest for Radunia and Raweta. Both stress agents (NaCl and PEG) have decreased the Mg, Ca and S content. The concentration of K and P in the leaves of wheat seedlings decreased only in cultures grown on media containing NaCl, whereas it generally increased in cultures grown on media with PEG, or was close to the level observed for the control conditions.
The microelement content in the control seedlings ranged from about 280 to 390 μg g−1 DW (Table 3) and was more diverse than that of the macroelements. The greatest accumulation of these elements was observed in the Jasna and Raweta cultivars. Both stresses caused a decrease in the accumulation of these elements: the smallest changes were observed in the Nawra and Katoda cultivars. Generally, a greater reduction in the accumulation of microelements relative to the control occurred in seedlings grown on media containing PEG than in the cultures grown on media containing NaCl.
Discussion
The decrease in the fresh weight of plants after both salinity- and drought-induced water stress have been reported for many species as one of the physiological symptoms of stress (Passioura and Munns 2000; Sucre and Suárez 2011). Our results confirmed these observations of growth reduction due to stress factors. However, in spring wheat varieties, in spite of the application of the same level of water stress induced by NaCl and PEG, the smaller decrease in the fresh weight caused by salinity can indicate a higher tolerance to this kind of stress in comparison to water stress induced by PEG. It is interesting because high salinity causes not only osmotic, but also ionic stress. High concentrations of Na+ ions disturb osmotic balance and result in “physiological drought” preventing plant water uptake (Türkan and Demiral 2009). If changes in leaf fresh weight (relative to the control) are taken as an indicator of stress tolerance to NaCl treatment, the spring varieties can be arranged in the following order of increasing sensitivity to stress: Katoda = Monsun = Nawra < Parabola < Raweta = Jasna = Banti = Cytra < Zadra < Radunia. Using the same parameter of the relative change in the weight of seedlings, these varieties can be arranged in a similar order of resistance to stress caused by the presence of PEG in the culture medium. The correlation between the mass of leaves of seedlings under stress and those of control plants was r = 0.738 and 0.602 (P < 0.005) for NaCl and PEG, respectively, which shows that varieties with a lower weight of leaves are more resistant to both stress factors studied.
At short-term exposure to high levels of osmotic stress, leaf growth is chiefly the result of a reduction in water uptake into the growing zone and an increased number of dead cells (Sucre and Suárez 2011). Changes in water uptake of leaves in spring wheat varieties were correlated with a decrease in fresh weight (r = 0.824 and r = 0.668 for NaCl and PEG induces stress, respectively). The higher reduction of fresh weight and water uptake after PEG treatment, in comparison to NaCl-treated seedlings, could be associated with the death of part of the leaf cells and membrane destruction under such stress. The increase in electrolyte leakage has usually been considered to be one of the major causes of increased cell membrane permeability of plants growing under different stresses (Tabaei-Aghdaei et al. 2000). About 50% relative electrolyte leakage (in comparison to total ion level) is regarded as an indicator of the damage to cell membranes. This study has not shown such a significant electrolyte leakage under the influence of both these stressors, which may indicate that under these conditions, despite a significant decrease in the weight of seedlings, no damage to the plasmalemma occurred. In addition, the leakage of electrolyte from plant cells after treatment with NaCl was higher than in plants treated with PEG, which may indicate a greater permeability of the membrane under the influence of NaCl treatment.
The overall stability of the membrane depends on the polar lipid content and fatty acid saturation. An increase in fatty acid unsaturation could cause an increase in membrane fluidity and thus its permeability (Quartacci et al. 2002). This study was focused on the determination of fatty acids in the phospholipid fraction, which represents about 60% of the total amount of the plasmalemma lipids in spring wheat seedling (data not shown). A short-term exposure to both stress factors resulted in no statistically significant changes in the level of this fraction.
Other authors have found a meaningful reorganization of polar lipid membranes during prolonged (more than 7 days) water stress (Guerfel et al. 2008; Tuomi et al. 2008). Moreover, the data for long-term drought stress showed an increase in the unsaturation level of the leaf polar lipids (Guerfel et al. 2008; Tuomi et al. 2008). In this study, water deficit induced by NaCl and PEG differentiated the direction of changes in unsaturation in the plasma membrane of spring varieties.
An increase in fatty acid unsaturation was observed after NaCl treatment in those wheat genotypes that were considered to be more sensitive to water stress, taking changes in fresh weight and uptake of water in leaves as indicators of stress tolerance (compare Figs. 1 and 2 with Fig. 5). An increase in the amounts of polyunsaturated fatty acids in plant membranes during salinity stress has already been reported by Allakhverdiev et al. (2001). It has been suggested that the high level of unsaturation of membrane lipids may be required to maintain the degree of fluidity needed for diffusion of lipophilic compounds and may confer a suitable geometry to the lipid molecules (Guerfel et al. 2008). However, a decrease in unsaturation in response to metal stress has also been reported (Ben Ammar et al. 2007; Filek et al. 2010). The increase in plasmalemma unsaturation in less tolerant plants under NaCl treatment may indicate an increase in the fluidity of the membrane surface, which could facilitate the deep penetration of toxic active oxygen species into the cell membranes where receptive fatty acid double bonds are located (Guerfel et al. 2008). The correlation (r = 0.912) between the U/S ratio and lipid peroxidation (MDA content), which is an indicator of the action of active oxygen species on membrane lipids under NaCl stress, suggests such a mechanism. However, under the influence of water stress caused by the presence of PEG, there is a negative correlation (r = −0.750) between the U/S ratio and the MDA content. This may indicate that under drought stress induced by a 2-day action of PEG, which causes major changes in physiological parameters (leaf weight and water content) an increased production of active oxygen species takes place, which penetrate the membranes and thus cause lipid peroxidation and an increase in the saturation degree of lipid fatty acids. However, in genotypes selected as tolerant, the increase in fatty acid saturation, as a result of NaCl stress, and a decrease after stress caused by the presence of PEG in the medium, indicate that these genotypes react differently to both stressful situations. Also, the relatively small changes in either the U/S parameter and the MDA content, as compared to the controls, are signs of higher stability of the membrane bilayer conformation, which can protect membranes from degradation. With the increasing lipid peroxidation in the stressed plants, the structural stability of the membranes seems to be the key factor in protecting leaves against osmotic stress.
Membrane permeability determined by differences in the unsaturation of lipid fatty acids (U/S) correlated with the accumulation (total amount) of macroelements in the leaves of seedlings grown on control media and under NaCl and PEG stresses. Under saline stress conditions, the higher content of macroelements in plants, as compared to PEG conditions, was a consequence of Na ion uptake. Recently, it has been hypothesised that Na ions may assume a positive function in the response to water stress, since plants specifically increase the absorption of this ion when submitted to drought in non-saline conditions (Sucre and Suárez 2011). In the halophyte species, Na ion concentration in leaves was significantly increased by simultaneous exposure to drought and salinity, compared to plants exposed only to saline stress, suggesting that Na ions are involved in the resistance to PEG-stimulated water stress. This also suggests that Na ions may directly or indirectly exert a positive influence on the accumulation of other compounds involved in osmotic adaptation (Slama et al. 2008; Sucre and Suárez 2011). Such an assumption could explain the greater tolerance to osmotic stress applied in wheat seedlings grown on media containing NaCl as opposed to that grown on media with PEG. However, in this study the Na ion content in wheat seedlings grown in Hoagland nutrient (control) was at a similar level to seedlings grown on media also containing PEG. In addition, the accumulation of other macroelements, important for the functioning of cells, was found to be lower under the stress induced by PEG than that caused by NaCl treatment. In particular, the effect of water stress on calcium (Ca), magnesium (Mg), phosphorus (P), sulphur (S) and potassium (K) concentration has been extensively investigated (Han et al. 2011; Hu et al. 2007). The K+/Na+ parameter has been suggested as a selection criterion for salt tolerance (Ashraf and Ali 2008). In the experiments presented here, no significant correlation between accumulated macroelements was found. These results are in agreement with suggestions made by other authors, that osmotic stress tolerance mechanisms may show difference depending on cultivars (Han et al. 2011; Tunçtürk et al. 2011).
The observed decrease in the accumulation of microelements (total content) in terms of both stress factors might suggest that the inhibition of the transport of these ions to the leaves of wheat seedlings is less selective (less dependent on the agent causing osmotic stress) than that of macroelements.
The function of microelements in cells is mainly associated with the action of antioxidative enzymes. In stress conditions, a decrease in manganese (Mn), iron (Fe), cooper (Cu) and zinc (Zn) levels may suggest a reduction in the synthesis of enzymes, especially of superoxide dismutases (SODs), which contain these ions. The smaller changes in the accumulation of microelements under the influence of both stressors, that were observed in seedlings of genotypes selected as more resistant (as in the case of macroelements), may be related to the smaller differences in the membrane permeability of these genotypes (in comparison to control).
In conclusion, short-term drought and salinity, in spite of the same level of water potential in the culture medium, cause stress reactions of varying intensity in leaves of spring wheat varieties. The smaller changes in physiological parameters that occurred after treatment with NaCl (than with PEG), as compared to the controls, suggest that the genotypes studied are more tolerant to this type of osmotic stress. They also suggest that the higher tolerance to NaCl treatment was related to lower peroxidation (MDA content) of lipids, but with greater membrane permeability, determined by the leakage of the electrolyte and the U/S ratio, as well as by an increased accumulation of macroelements, mainly Na ions. The greater decrease in fresh weight and water content in the leaves, which occurred under stress caused by the presence of PEG in growing media, was connected with the higher lipid peroxidation as well as the lower permeability of membranes, compared to the stress induced by NaCl. It was found that more tolerant genotypes were characterized by a higher stability of membrane lipid fatty acid content and membrane permeability compared to the more sensitive ones. Thus, cell membrane permeability can be an effective determinant of osmotic stress tolerance in the varieties examined in this study. The relationship between changes in the activation of antioxidant enzymes and the production of organic osmoprotectants will be examined in further studies to compare the differences in effect of osmotic stress induced by NaCl and PEG on agronomically important genotypes of spring wheat.
References
Allakhverdiev SI, Kinoshita M, Inaba M, Suzuki I, Murata N (2001) Unsaturated fatty acids in membrane lipids protect the photosynthetic machinery against salt-induced damage in Synechoccus. Plant Physiol 125:1842–1853
Asada K (1999) The water–water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Ann Rev Plant Physiol Plant Mol Biol 50:601–639
Ashraf M, Ali Q (2008) Relative membrane permeability and activities of some antioxidant enzymes as the key determinants of salt tolerance in canola (Brassica napus L.). Environ Exp Bot 63:266–273
Ben Ammar W, Nouairi I, Zarrouk M, Jemal F (2007) Cadmium stress induces changes in the lipid composition and biosynthesis in tomato (Lycopersicon esculentum Mill.) leaves. Plant Growt Regul 53:75–85
Bligh EG, Dyer WJ (1959) A rapid method of total lipids extraction and purification. Can J Biochem 37:911–915
Block MA, Dorne AJ, Joyard R, Douce R (1983) Preparation and characterization of membrane fraction enriched in outer and inner envelope membranes from spinach chloroplasts. J Biol Chem 258:13281–13286
Dhindsa RS, Plumb-Dhindsa P, Thorpe TA (1981) Leaf senescence correlation with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J Exp Bot 32:96–101
Farooq S, Azam F (2006) The use of cell membrane stability (CMS) technique to stress for salt tolerant wheat varieties. J Plant Physiol 163:629–637
Filek M, Zembala M, Kornaś A, Walas S, Mrowiec H, Hartikainen H (2010) The uptake and translocation of macro- and microelements in rape and wheat seedlings as affected by selenium supply level. Plant Soil 336:303–312
Franco JA, Esteban C, Rodriguez C (1993) Effects of salinity on various growth stages of muskmelon cv. Revigal. J Hortic Sci 68:899–904
Guerfel M, Baccouri O, Boujnah D, Zarrouk M (2008) Changes in lipid composition, water relations and gas exchange in leaves of two young ‘Chemlali’ and ‘Chetoui’ olive trees in response to water stress. Plant Soil 311:121–129
Han LP, Steinberger Y, Zhao YL, Xie GH (2011) Accumulation and partitioning of nitrogen, phosphorus and potassium in different varieties of sweet sorghum. Field Crop Res 120:230–240
Hsiao T (1973) Plant responses to water stress. Ann Rev Plant Physiol 24:519–570
Hu Y, Burucs Z, von Tucher S, Schmidhalter U (2007) Short-term effects of drought and salinity on mineral nutrient distribution along growing leaves of maize seedlings. Environ Exp Bot 60:268–275
Kramer PJ (1988) Measurement of plant water status: historical perspectives and current concerns. Irrig Sci 9:275–287
Liang XZ, Ding SW, Wong SM (2002) Development of a kenaf (Hibiscus cannabinus L.) protoplast system for a replication study of Hibiscus chlorotic rings pot virus. Plant Cell Rep 20:982–986
Martyniak L, Dabrowska-Zielnska K, Szymczyk R, Gruszczynska M (2007) Validation of satellite-derived soil-vegetation indices for prognosis of spring cereals yield reduction under drought conditions—case study from central-western Poland. Adv Space Res 39:67–72
Munns R (1993) Physiological processes limiting plant growth in saline soils: some dogmas and hypothesis. Plant Cell Environ 16:201–218
Munns R (2002) Comparative physiology of salt and water stress. Plant Cell Environ 25:239–250
Nagy Z, Galiba G (1995) Drought and salt tolerance are not necessarily linked: a study on wheat varieties differing in drought resistance under consecutive water salinity stresses. J Plant Physiol 145:168–174
Parida AK, Das AB (2005) Salt tolerance and salinity effects on plants: a review. Ecotoxicol Environ Saf 60:324–349
Park S-H, Wilson JD, Seabourn BW (2009) Starch granule size distribution of hard red winter and hard red spring wheat: its effects on mixing and breadmaking quality. J Cereal Sci 49:98–105
Passioura JB, Munns R (2000) Rapid environmental changes that affect leaf water status induce transient surges or pauses in leaf expansion rate. Aust J Plant Physiol 27:941–948
Quartacci MF, Glisic O, Steranovic B, Navarri-Izzo F (2002) Plasma membrane lipids in the resurrection plants Ramonda serbica following dehydratation and rehydratation. J Exp Bot 53:2159–2166
Romero-Aranda MR, Jurado O, Cuartero J (2006) Silicon alleviates the deleterious salt effect on tomato plant growth by improving plant water status. J Plant Physiol 163:847–855
Rossi S, Simard S, Rathgeber CBK, Deslauriers A, Zan C (2009) Effects of a 20-day-long dry period on cambial and apical meristem growth in Abies balsamea seedlings. Trees 23:85–93
Schar C, Vidale PL, Luthi D, Frei C, Haberli C, Liniger MA, Appenenzeller C (2004) The role of increasing temperature variability in European summer heatwaves. Nature 427:332–336
Slama I, Ghaya T, Savouré A, Abdelly C (2008) Combined effects of long-term salinity and soil drying on growth, water relations, nutrient status and proline accumulation of Sesuvium portulacastrum. C R Biol 331:442–451
Silva EN, Ferreira-Silva SL, Fontenelea AV, Ribeirob RV, Viégasc RA, Silveira JAG (2010) Photosynthetic changes and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in Jatropha curcas plants. J Plant Physiol 167:1157–1164
Sucre B, Suárez N (2011) Effect of salinity and PEG-induced water stress on water status, gas exchange, solute accumulation, and leaf growth in Ipomoea pes-caprae. Environ Exp Bot 70:192–203
Tabaei-Aghdaei S, Harrison P, Pearee RS (2000) Expression of dehydration-stress related genes in crown of wheat, grass species having contrasting acclimation to salt, cold and drought. Plant Cell Environ 23:561–571
Tuomi I, Gargouri M, Nouairi I, Moschou PN, Salem-Fnayou AB, Mliki A, Zarrouk M, Ghorbel A (2008) Water stress induced changes in the leaf lipid composition of four grapevine genotypes with different drought tolerance. Biol Plant 52:161–164
Tunçtürk M, Tunçtürk R, Yildirim B, Çiftçi V (2011) Effect of salinity stress on plant fresh weight and nutrient composition of some Canola (Brassica napus L.) cultivars. Afr J Biotechnol 10:1827–1832
Türkan I, Demiral T (2009) Recent development in understanding salinity tolerance. Environ Exp Bot 67:2–9
Walter J, Nagy L, Hein R, Rascher U, Beierkuhnlein C, Willner E, Jentsch A (2011) Do plants remember drought? Hints towards a drought-memory in grasses. Environ Exp Bot 71:34–40
Wang W-B, Kim Y-H, Lee H-S, Kim K-Y, Deng X-P, Kwak S-S (2009) Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses. Plant Physiol Biochem 47:570–577
Zembala M, Filek M, Walas S, Mrowiec H, Kornaś A, Mieszalski Z, Hartikainen H (2010) Effect of selenium on macro- and microelement distribution and physiological parameters of rape and wheat seedlings exposed to cadmium stress. Plant Soil 329:457–468
Acknowledgments
This work was partly supported by COST project No.556/COST/2009.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Zwiazek.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Filek, M., Walas, S., Mrowiec, H. et al. Membrane permeability and micro- and macroelement accumulation in spring wheat cultivars during the short-term effect of salinity- and PEG-induced water stress. Acta Physiol Plant 34, 985–995 (2012). https://doi.org/10.1007/s11738-011-0895-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-011-0895-5