
Abstract
Potato virus Y (PVY) is considered the most economically damaging virus for seed-potato production. PVY isolates are usually divided into three historical PVY strains, namely PVYC, PVYO and PVYN. More recently, recombinant strains named PVYNTN and PVYN-Wi appeared in Europe and North America, rapidly spread into seed-potato production and gradually replaced historical strains. This progression can be explained by several factors, including the differential susceptibility of cultivars to PVY strains. This article presents the results of a Swiss survey of PVY strains conducted in 2012 in which the progressive increase of the prevalence of PVYN-Wi was observed. The occurrence of PVY strains in the two main potato cultivars grown in Switzerland, cv. Agria and cv. Charlotte, is described through the 2012 survey and one additional PVY strain survey conducted in 2014. Both surveys were completed with a mechanical-inoculation assay. The inoculation assay showed that cv. Agria is more susceptible to PVYN-Wi than to PVYNTN, whereas cv. Charlotte is susceptible to both strains. The inoculation assay also showed that the expression of symptoms on cultivars is strain-dependent. These results stress the major role of the resistance profile of cultivars to explain the balance of the PVY strains in potato crops.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Potato virus Y (PVY) is considered the most economically damaging potato virus worldwide (Gray et al. 2010; Rolland et al. 2008). This Potyvirus (family Potyviridae) affects the yield of susceptible cultivars (Hane and Hamm 1999; Rykbost et al. 1999; Nolte et al. 2004; Whitworth et al. 2006). PVY is transmitted in a nonpersistent manner by aphids of various species (Woodford 1992) and can be spread within the potato field as well as from field to field (Boiteau et al. 1988, 2009; Bagnall 1992; Davis et al. 2009).
As reviewed in Kerlan et al. (2011), Rolland et al. (2008) and Karasev and Gray (2013), PVY potato isolates are divided into three main strains defined according to plant indicators: PVYC, PVYO and PVYN. PVYC induces hypersensitive reactions (HRs) in some potato cultivars (e.g. cv. King Edward), while PVYO induces HRs in other potato cultivars (e.g. cv. Désirée and Pentland Crown). PVYN can be distinguished from PVYC and PVYO by its ability to induce vein necrosis when infecting tobacco (Nicotiana tabacum) but causes milder symptoms in most potato cultivars (Karasev and Gray 2013). Two main variants within the PVYN group, PVYNTN and PVYN-Wi, appeared more recently in Europe and gradually replaced the historical strains (Kerlan et al. 2011). Based on their genome sequences, they were shown to be recombinants of PVYO and PVYN strains. PVYN-Wi and PVYNTN isolates both induce vein necrosis on tobacco (Rolland et al. 2008; Karasev et al. 2011). PVYNTN also induces tuber necrosis in some susceptible cultivars, causing a potato disease called potato tuber necrotic ringspot disease (PTNRD). The molecular distinction between PVYNTN and PVYN-Wi is that most PVYNTN isolates have three or four recombinant junctions, whereas PVYN-Wi isolates have one or two (Rolland et al. 2008; Karasev et al. 2011). Serologically, PVYN-Wi reacts as PVYO, while PVYNTN reacts as PVYN (Karasev et al. 2010).
The PVY recombinant strains, PVYNTN and PVYN-Wi, spread rapidly during the last 30 years into European seed-potato production and almost replaced the historical strains (Kerlan et al. 2011). The spread of PVYN-Wi was more recent than the spread of PVYNTN, as this strain was first reported in Poland in the early 1990s (Chrzanowska 1991) and has since been reported in several countries, including France (Glais et al. 2005; Rolland et al. 2008), the USA, Canada (Gray et al. 2010), Spain (Blanco-Urgoiti et al. 1998), Germany, the UK (Schubert et al. 2007; Lindner et al. 2015), the Netherlands (van der Vlugt et al. 2007), Belgium (Kamangar et al. 2014) and Switzerland (Rigotti et al. 2011). In Switzerland, the prevalence of PVYN-Wi was at about 17% in 2008 (Rigotti et al. 2011). It was comparable/slightly higher in the USA and Canada, with 18% and 33%, respectively, in 2006, but it was much higher in Germany, with 63% in 2011 (Lindner et al. 2015). The prevalence of PVYN-Wi seems to have increased in Switzerland by 12% from 2003 to 2008 (Rigotti et al. 2011) and in Canada by 9% from 2004 to 2006 (Gray et al. 2010). PVYN-Wi seems to have been relatively stable in Germany, with an increase of 4% from 2009 to 2011 (Lindner et al. 2015) and to have decreased in the USA by 9% from 2004 to 2006 (Gray et al. 2010). Since then, this percentage has significantly increased to reach 47% in 2012 (+ 23% since 2009), PVYN-Wi becoming the most prevalent strain in the USA (Stewart Gray, personal communication). The prevalence of PVYN-Wi seems to increase in the countries where periodic PVY strain monitoring has been performed. The following hypothesis have been proposed to explain the increase of the prevalence of PVYN-Wi strains: climate change and changes in cultural practices (Rolland et al. 2008), a higher transmission rate of PVYN-Wi strains by aphids (Verbeek et al. 2010), a faster translocation of PVYN-Wi than PVYNTN (Dupuis 2017), a faster spread in the field (Dupuis et al. 2014), a lower expression of symptoms by PVYN-Wi-infected plants, which are then less susceptible to being eliminated from fields by roguing (Chrzanowska 1991; Glais et al. 2005), acquisition of an enhanced pathogenicity (Bellstedt and Visser 2016) and finally, a shift in the cultivars that are produced, with the progressive introduction of cultivars susceptible to PVYN-Wi and resistant to other PVY strains (Lindner and Billenkamp 2005; Lindner et al. 2015; Funke et al. 2017). Indeed, it was shown through surveys in Germany and the USA that the susceptibility of the cultivars varies among PVY strains. For example, in surveys conducted in Germany between 2009 and 2011, all PVY-infected plants of cv. Agria (n = 240) were found to be infected by PVYN-Wi, while the prevalence rate of PVYNTN in the PVY-infected plants was 34% in this country over the same period (Lindner et al. 2015). Surveys managed from 2011 to 2015 in the Columbia Basin in the Pacific Northwest of the USA also proved strain-specific resistance exhibited by potato consistency grown in the area (Funke et al. 2017). This resistance resulted in a significant drop of the PVYO strain in favour of the recombinant strains PVYNTN and PVYN-Wi.
This article has two objectives. Firstly, to characterize the incidence of the PVY strains in Switzerland between 2008 (results shown in Rigotti et al. (2011)) and 2012. Secondly, to assess the strain-specific susceptibility of the two main cultivars grown in Switzerland in 2012, namely cv. Agria (22% of the surface) and cv. Charlotte (14% of the surface) (Swisspatat 2012). The results of the survey of 2012 were combined with the results of another survey performed 2 years later to assess the incidence of the strains PVYNTN and PVYN-Wi in these cultivars. In addition to the surveys, a greenhouse trial was performed in which plants of cv. Agria and Charlotte were mechanically inoculated with PVYNTN and PVYN-Wi in order to assess their susceptibility to both strains under controlled conditions and the corresponding development of symptoms.
Materials and Methods
Plant Material
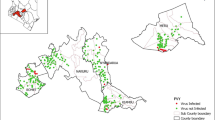
In June 2012, 338 potato leaf samples with severe or mild symptoms were collected from the following post-harvest fields in Switzerland: Grange-Verney (canton of Vaud), Rütti (canton of Bern), Uttewil (canton of Fribourg), Wallierhof (canton of Solothurn), Liebegg (canton of Aarau) and Strickhof (canton of Zürich). A sample of 100 tubers of all Swiss seed potato crops are planted in these fields. Each post-harvest field groups samples coming from crops grown in the same area the previous year. The location of the post-harvest fields in Switzerland is presented in Fig. 1. The samples were collected on 25 different potato cultivars (Agata, Agria, Amandine, Annabelle, Antina, Bintje, Celtiane, Charlotte, Désirée, Ditta, Fontane, Gourmandine, Hermes, Innovator, Jelly, Lady Rosetta, Lady Claire, Lady Jo, Malou, Markies, Nicola, Panda, Pirol, Stella and Victoria). The proportion of Agria samples collected in each location is reported in brackets in Fig. 1. For each sample, three leaflets were selected and stored in plastic bags at − 20 °C until use for serological and molecular analysis (detailed below). In June 2014, 91 additional potato leaf samples of Agria and Charlotte with severe or mild symptoms were collected from the same fields and analysed in the same way.

Location of the post-harvest fields in Switzerland. For each location, the proportion of samples of cv. Agria is reported in brackets
Agria and Charlotte were also challenged with PVY strains in greenhouse trials. The trials were repeated three times, with cv. Charlotte tested in trials 1, 2 and 5, and cv. Agria tested in trials 3, 4 and 5. Virus-free tubers were planted in plastic pots (19 × 19 × 18 cm) filled with compost. The pots were randomized and maintained in an insect-proof greenhouse (20 °C, 70% relative humidity and 12-h photoperiod). Three leaves of each plant were inoculated 7 days after plant emergence using carborundum (Perez 2011). The inoculum was prepared from fresh tobacco leaves showing PVY symptoms crushed in an inoculation buffer (0.02 M phosphate + 0.01 M DIECA). At least 20 plants were inoculated with two PVY isolates (Agroscope collection): PVYN-Wi 1315 isolate (28 in trial 1, 22 in trial 2, 21 in trial 3, 25 in trial 4, and 22 in trial 5) and PVYNTN 1317 isolate (105 in trial 1, 35 in trial 2, 20 in trial 3, 25 in trial 4, and 22 in trial 5). A leaf sample of potentially systemically infected leaves (three leaves per plant) was taken 6 weeks after inoculation for serological analysis. The symptoms were observed at the same time (6 weeks post-inoculation) in trials 3, 4 and 5. As the control for symptomless plants, 6, 15 and 6 plants, respectively, were kept non-inoculated. The symptoms were classified following a four-grade scale: 0 = no visible symptoms, 1 = small necrotic spots, 2 = large necrotic spots and presence of dry leaflets and 3 = completely dry leaves. For trial 4, the tuber yield/plant was measured (g/plant).
Samples Analysis
The serological and molecular methods used for the analysis of the leaf samples were the same as the methods described in Rigotti et al. (2011). In summary, leaf samples were analysed by a double antibody sandwich (DAS)-ELISA using PVY monoclonal antibodies (mAb). The plates were coated with anti-PVY-NOC mAb (210369, Bioreba, Basel, Switzerland), and the PVYN strains were detected using the anti-PVY-N mAb (191073, Bioreba). A sample was considered infected by a PVYO strain if it reacted negatively to anti-PVY-N mAb and positively to anti-PVY-NOC mAb (210369, Bioreba), a mAb detecting both PVYN and PVYO strains. All the collected leaves were ELISA tested. A sample was considered positive if the optical density value was three times higher than the optical density of sap extracted from a healthy potato tuber (control on each ELISA plate).
Part of the leaf samples of the survey was analysed by RT-PCR (numbers in Table 1), as proposed by Rigotti et al. (2011). RNA was extracted from 500 mg of potato leaf tissue, and a one-step triplex RT-PCR was performed using the three primer pairs PVYc3/f, PVY3+/3− and CP2+/1−. This triplex RT-PCR allows for the discrimination of the PVY isolates belonging to the following groups: PVYO, PVYC, PVYN-Wi and PVYNTN.
Monitoring of Aphid Flights
Migration flights of aphids are monitored every year in Changins (Nyon, Switzerland; 6°13′44″ E, 46°23′56″ N; 440 m a.s.l.) using a ‘Rothamsted Insect Survey’ suction trap. This trap samples winged aphids and other airborne organisms at a height of 12.2 m above ground by aspirating 0.75 m3 air per second (Macaulay et al. 1988). This trap is the closest to the main seed potato production basin in Switzerland, which is located 50 to 150 km from Chagins. For this study, the identification of aphid species focused on eight species that are commonly regarded as important vectors of PVY (Verbeek et al. 2010; Fox et al. 2017): Acyrthosiphon pisum (Harris), Aphis fabae (Scopoli), Aphis spp., Brachycaudus helichrysi (Kaltenbach), Macrosiphum euphorbiae (Thomas), Myzus persicae (Sulzer), Phorodon humuli (Schrank) and Rhopalosiphum padi (Linnaeus). The aphid flights were monitored in 2003, 2008 and 2012 during the first 7 weeks after emergence of the main seed potato crops in Switzerland, usually between mid-April and mid-May. The dates of planting and emergence were supplied by the Swiss seed potato certification services (Peter Frei, personal communication).
Data Analysis
The serological diversity of PVY strains in 2012 was compared between post-harvest fields and between cultivars using a chi-squared test (α = 0.05). For this analysis, we considered the post-harvest fields and cultivars for which at least 10 samples were collected. The 2012 distribution for each PVY strain was compared to the corresponding distribution of 2003 and 2008 surveys presented by Rigotti et al. (2011). Mixed infections were counted as positive samples for each of the identified virus strains. The distribution of the strains between years was compared with another chi-squared test (α = 0.05). The distribution of the strains infecting cv. Agria was compared to the distribution of the stains infecting cv. Charlotte also with a chi-squared test (α = 0.05).
For the greenhouse trial, we used the general linear model (α = 0.05) to evaluate the effects of PVY strains on the symptom expressions on the foliage, on the percentage of infected plants and on the tuber yield. The trials were included in the model as the replication factor for the analysis of the symptoms on the foliage and percentage of infected plants. For the analysis of the tuber yield data, the plants were used as the replication factor. The data of each cultivar were analysed separately as they were tested in different trials (except trial 5).
The software Statistica® (StatSoft Inc., Tulsa, USA) was used to perform all analyses.
Results
Survey 2012
In 2012, 176 of the collected samples were tested positive to PVY by ELISA, among which 130 were PVYN positive (74%) and 46 PVYO positive (26%). The chi-squared test detected a significant difference (p < 0.01) between the balance of the prevalence of the PVY strains in Uttewil compared to the average balance of strains in the other fields (Table 1), with a lower proportion of PVYN strain in Uttewil. Nevertheless, this difference is low as there is an important overlap between the lower and higher bounds of the confidence intervals of this site and the average of all experimental sites (Table 1). Most of the consistency were mainly infected by PVYN (Table 2) except cv. Agria for which most of the samples were infected by PVYO strain (Table 2); this strain distribution was significantly different from the average of the other cultivars (p < 0.001).
We further characterized 103 isolates from the 2012 survey by molecular testing, with 57 isolates serologically positive for PVYN and 46 isolates serologically positive for PVYO. In 2012, only one mixed infection was found between PVYN and PVYN-Wi.
The data and distribution of each PVY group are presented in Table 3 and compared with data from the 2003 and 2008 surveys published by Rigotti et al. (2011). The chi-squared test shows that the distribution of the PVY strains significantly changed from 2003 to 2008 (p < 0.001), from 2008 to 2012 (p < 0.05) and from 2003 to 2012 (p < 0.01). Table 3 presents these changes in details, as well as their significance. A change in prevalence of a virus strain from one survey to the other can be considered as significant when there is no overlap between the confidence intervals of both surveys (see Table 3). The PVYNTN group remains the most prevalent in Switzerland but has decreased from 78.9% of prevalence in 2003, to 70.4% in 2008, and 71.0% in 2012. This important decrease (7.9% between 2003 and 2012) is marginally significant as there is only 3.9% of overlap between the higher bound of the confidence interval of 2003 and the lower bound of the confidence interval of 2012. This decrease of the prevalence of the PVYNTN strain is compensated by an important increase in the prevalence of the PVYN-Wi strain. The prevalence of this strain has been increasing from 5.5% in 2003 to 17.3% in 2008 and 18.8% in 2012. This increase of 13.3% from 2003 to 2012 also compensates the decrease of the PVYO strain observed during the same period (5.9%). The prevalence of this strain has importantly decreased between the first two surveys (8.4%) with 13.3% in 2003 and 4.9% in 2008. Then, the prevalence of PVYO remained relatively stable with 7.4% prevalence in 2012. Finally, PVYN populations have been increasing from 2003 to 2008 with 2.3% of prevalence in 2003 and 7.4% in 2008. This increase of 5.1% was almost compensated by a decrease of 4.8% between 2008 and 2012, with 2.6% of occurrence in the last survey.
Agria and Charlotte Surveys
The data from the 2014 survey were combined with the data from the 2012 survey in Table 4 to assess the prevalence of the main PVY strains in the cultivars Agria and Charlotte. Results of both years are presented together in Table 4, as there were not enough positive samples for each single year (n = 26 in 2012 and n = 85 in 2014) to be able to determine tendencies by year. In both surveys, 58 cv. Agria plants and 53 cv. Charlotte plants were found to be infected by PVY. The chi-squared test showed that the distribution of the PVY strains was significantly different in cv. Agria compared to cv. Charlotte (p < 0,001). Cultivar Agria was mainly infected by PVYN-Wi (81%) and PVYO (16%) strains, while only eight plants were found to be infected by PVYNTN strains (3%) and no plants by PVYN. In contrast, cv. Charlotte was mostly infected by PVYNTN strains (90%), whereas only 6% of the plants were infected by PVYN-Wi. One and two plants, respectively, of cv. Charlotte were found to be infected by PVYN and PVYO strains.
Monitoring of Aphid Flights
The results of the aphids monitoring performed in Changins in 2003, 2008 and 2012 are presented in Fig. 2. We had a very low amount of captures of Aphis fabae and M. euphorbiae during the 3 years of monitoring with less than 20 individuals captured every week. During the 3 years of monitoring, the captures of M. persicae were also at a low level for the first 6 weeks of measurement, but increased in the last week, particularly in 2003 with 39 captures in week 7. The captures of B. helichrysi were also at a low level in 2003 and 2012 but relatively high in the beginning of the growing season of 2008, with 57 individuals captured the first week after emergence. The captures of Aphis spp. were low the first 2 weeks of 2008 and 2012 (less than 20), and then increased to reach respectively 51 and 27 captures the seventh week. In 2003, the captures remained below 20 the first 5 weeks of the growing season and then increased sharply to reach 44 captures the seventh week. The captures of R. padi remained under 20 captures per week in 2003 and 2012, while the captures were very high in 2008 with two peaks of 41 individuals captured 2 weeks after emergence and up to 154 individuals 7 weeks after emergence. The captures of A. pisum and P. humuli were fluctuating in 2008 with peaks of 30 individuals the third and fifth weeks. The captures of A. pisum remained between 0 and 10 in 2012, and the captures of P. humuli were in the same range in 2003. A late increase in the captures was observed with A. pisum (35 individuals the seventh week) and P. humuli (43 individuals the sixth week), respectively, in 2003 and 2012.

Number of aphids weekly captured during the potato growing season by the ‘Rothamsted Insect Survey’ suction trap of Changins (VD, Switzerland). The following species were considered: Acyrthosiphon pisum (Harris), Aphis fabae (Scopoli), Aphis spp., Brachycaudus helichrysi (Kaltenbach), Macrosiphum euphorbiae (Thomas), Myzus persicae (Sulzer), Phorodon humuli (Schrank) and Rhopalosiphum padi (Linnaeus). The black dotted line shows the captures for the year 2003, the grey dotted line shows the captures for 2008 and the black line shows the captures for 2012
Inoculation Trials
In the greenhouse assays, the infection rate of cv. Charlotte was similar (p > 0.05) for PVYN-Wi (57%) and PVYNTN (69%) strains, while the inoculation rate of cv. Agria was significantly higher for PVYN-Wi (p = 0.029), with 65% compared to PVYNTN with 29% (Fig. 3). The plants of cv. Agria infected with PVYNTN showed more severe symptoms (p < 0.001) compared to plants infected with PVYN-Wi, with necrosis (local and systemic necrosis) on nearly all leaflets and a high number of totally dry leaflets and possible systemic reactions (Figs. 4 and 5). The plants inoculated with PVYN-Wi mainly showed small necrotic spots on the foliage (Figs. 4 and 5). The plants of cv. Charlotte showed severe symptoms regardless of the strain used for the inoculation (Fig. 4). No significant difference of tuber yield was observed between the cv. Agria plants inoculated with different PVY strains (p = 0.607) with an average of 49 g of tubers for the control plants and 59 g and 62 g, respectively, of tubers for the plants inoculated with PVYNTN and PVYN-Wi.

Percentage of infected plants of cv. Agria and Charlotte after mechanical inoculation with PVYN-Wi 1315 and PVYNTN 1317 strains. The number in brackets in the legend gives the p value of the analysis of variance

Severity of symptoms (0 = non-visible symptoms; 1 = small necrotic spots; 2 = big necrotic spots and presence of dry leaflets and 3 = leafs completely dry). The number in brackets in the legend gives the p value of the analysis of variance

Symptoms of PVY strains on leaflets 60 days after mechanical inoculation of plants of cv. Agria. a Uninoculated plants, b plants inoculated with PVYN-Wi and c plants inoculated with PVYNTN
Discussion
The surveys for PVY revealed a slow yet progressive increase of the prevalence of PVYN-Wi strains in Switzerland and, simultaneously, a gradual decrease of the prevalence of PVYNTN and PVYO strains in Swiss seed-potato crops. The PVYN strain is still present albeit at low levels. This result is consistent with the increase of the prevalence of PVYN-Wi strains observed in Germany, Canada, and the USA (Gray et al. 2010; Lindner et al. 2015). Such increase in the prevalence of PVYN-Wi might be associated with factors specific to each country.
Several main factors may explain the increase in the prevalence of PVYN-Wi in Switzerland: (i) the imports of potato seed infected by PVYN-Wi, (ii) the share of each vector species in the aphid flights during the growing season, (iii) the resistance profile of the potato cultivars grown in the country (iv) and the fitness of translocation of the PVY strains from the leaves to the progeny tubers for these cultivars.
In Switzerland, the role of importation of infected seed potatoes can be excluded, as they represent only 15% of the seed potatoes with low PVY infection rates (0.1 to 0.2%—Peter Frei, personal communication).
Verbeek et al. (2010) showed that the efficacy of transmission of PVYN-Wi, compared to PVYNTN, was higher for six aphid species: A. pisum, A. fabae, A. nasturtii, Aphis spp., P. humuli and R. padi. All these species were monitored during the years of our PVY strain surveys except A. nasturtii which was included in the captures of Aphis spp. (Fig. 2). It is difficult to draw conclusions regarding the impact of the aphid flights on the evolution of PVY strain prevalence in Switzerland, but we can highlight aphid flight events that could have played a role in increasing the prevalence of PVYN-Wi. In 2008 and 2012, we observed important flights of Aphis spp. and P. humuli compared to 2003. For these species, the main flights occurred 3 weeks after the emergence in 2008. At this time, the young plants are very susceptible to PVY infections due to the low expression of the phenomenon of mature plant resistance. This phenomenon implies an increase of the resistance of the plants to PVY infections with the age of the plant (Sigvald 1985). In 2012, the main flights of the same species occurred later (6 weeks after emergence), but with a higher intensity compared to 2008. Therefore, the flights of Aphis spp. and P. humuli may have played a role in facilitating the spread of PVYN-Wi in both 2008 and 2012 compared to 2003. The flights of A. pisum came early in 2008 but with a low intensity and were higher in 2003 later in the season. Therefore, the flights of this species may not explain the increase of PVYN-Wi in Switzerland. Massive flights of R. padi were monitored in 2008 both early and late in the growing season with up to 154 individuals captured the seventh week of sampling. Nevertheless, the flights of R. padi were at very low level in 2012, and hence, the increase of the prevalence of PVYN-Wi cannot be attributed solely to this aphid species.
A change in the cultivars grown in the fields is an important factor impacting the distribution of PVY strains. It has been shown by Funke et al. (2017) in the context of seed-potato production in the Columbia Basin (USA), where the prevalence of PVYO dropped down sharply due to the introduction of potato cultivars resistant to this strain. Consistent with surveys performed in Germany (Lindner and Billenkamp 2005; Lindner et al. 2015), our surveys revealed that Agria is more frequently infected by PVYN-Wi (81% prevalence). In our greenhouse trial, this cultivar resisted PVYNTN better than it resisted the PVYN-Wi inoculation. This cultivar’s specific resistance to PVYNTN leads to a higher detection of PVYN-Wi in the symptomatic plants, as observed in our surveys. It is important to remember that our trials were done through mechanical inoculation, meaning that a large amount of virus particles was introduced into the plant tissues, and much more than the few (1 to 3) virus particles usually inoculated by aphids (Moury et al. 2007). Nevertheless, the inoculation was done on a single leaf, while the detection of the virus post-inoculation was done on three uninoculated leaves. This means that the virus had to replicate and migrate systemically into the plant to be detected. Therefore, our technique is reliable to detect phenomenon of resistance but may give different results than trials performed through inoculation by aphids. We observed in the inoculation trials that Agria displays less severe symptoms in response to PVYN-Wi infection. Given that symptom expressions strongly influence the efficacy of elimination of symptomatic plants from fields by roguing (Chrzanowska 1991; Glais et al. 2005), the resistance of Agria to PVYNTN and the lower expression of symptoms of Agria infected by PVYN-Wi might both contribute to PVYN-Wi prevalence compared to PVYNTN. The increase of PVYN-Wi prevalence may then be amplified where roguing is used routinely such as in Switzerland. The cultivar-specific property of developing milder symptoms when infected by PVYN-Wi was also reported in cv. Elipsa by Chrzanowska (1991). In our greenhouse trial, Agria plants infected by PVYNTN displayed local necrosis in inoculated leaves or systemic necrosis with total desiccation of the infected stems followed by leaf-drop syndrome. Such symptoms in potato plants can be associated with hypersensitive resistance (HR) response in spite of the fact that the virus can still spread systemically (Valkonen et al. 1998; Funke et al. 2017). This HR response of cv. Agria has been previously described by Petrov et al. (2015), in a trial with mechanically inoculated plants with PVYNTN. This trial revealed the blocking of the replication of PVYNTN in the newly grown leaves. The inoculated leaves remained infected and collapsed after desiccation, while the virus was absent from the newly grown parts of potato plants. By contrast, no difference in symptom expressions or cultivar-specific resistance is observed in other cultivars, such as cv. Charlotte inoculated with the PVYNTN or PVYN-Wi strains. In the field, however, the observed prevalence of PVYNTN in this cultivar is close to 90%, to the detriment of the plants being infected by PVYN-Wi. This level exceeds the average prevalence of PVYNTN in the country by 20%. This shows that other factors are probably playing a role in the higher prevalence of PVYNTN in cv. Charlotte. In Switzerland, the increase in the prevalence of PVYN-Wi cannot only be attributed to the prevalence of Agria in the seed potato crops for two main reasons. Firstly, the prevalence of Agria in Switzerland remained relatively stable between 2003 and 2012. In 2003, Agria accounted for 22% of the acreage, in 2008 for 20%, and in 2012 for 22% (Christine Heller, personal communication). Secondly, we observed a lower proportion of PVYNTN strain in the post-harvest field of Uttewil. We would then expect to find a higher proportion of Agria samples coming from this field. However, the proportion of Agria samples collected in this location was lower (9.7%) than in Rütti (11.1%) and Liebegg (13.3%), as shown in Fig. 1.
Finally, the increase of the prevalence of PVYN-Wi in Switzerland may also be due to a faster translocation of this strain in the progeny tubers compared to PVYNTN. This phenomenon has been shown by Dupuis (2017) with potato plants infected with these strains.
Overall, the increase of the prevalence of the PVYN-Wi strain in the seed potato crops of Switzerland may be due to the interactions of several factors such as the evolution of the populations of vectors, the fitness of the PVY strains in the plant and the resistance profile of the potato cultivars grown in the country.
References
Bagnall RH (1992) Epidemiology of potato virus Y in New-Brunswick, Canada—a biennial rhythm and 9-plus year cycle. Can J Plant Pathol 14:137–146
Bellstedt DU, Visser JC (2016) Why are some PVY recombinant strains replacing the older non-recombinant strains? In: EAPR (ed) 16th triennial meeting of the Virology Section of the European Association of Potato Research (EAPR), Ljubljana, Slovenia. NIB, p 17
Blanco-Urgoiti B, Tribodet M, Leclere S, Ponz F, de San Roman CP, Legorburu FJ, Kerlan C (1998) Characterization of potato potyvirus Y (PVY) isolates from seed potato batches. Situation of the NTN, Wilga and Z isolates. Eur J Plant Pathol 104:811–819
Boiteau G, Singh RP, Parry RH, Pelletier Y (1988) The spread of PVYo in New-Brunswick potato fields: timing and vectors. Am Potato J 65:639–649
Boiteau G, Singh M, Lavoie J (2009) Crop border and mineral oil sprays used in combination as physical control methods of the aphid-transmitted potato virus Y in potato. Pest Manag Sci 65:255–259
Chrzanowska M (1991) New isolates of the necrotic strain of potato virus Y (PVYN) found recently in Poland. Potato Res 34:179–182
Davis JA, Radcliffe EB, Ragsdale DW (2009) Planter skips and impaired stand favors potato virus Y spread in potato. Am J Potato Res 86:203–208. https://doi.org/10.1007/s12230-009-9073-3
Dupuis B (2017) The movement of potato virus Y (PVY) in the vascular system of potato plants. Eur J Plant Pathol 147:365–373. https://doi.org/10.1007/s10658-016-1008-5
Dupuis B, Balmelli C, Schumpp O, Steinger T (2014) Within-field spread of two strains of potato virus Y (PVY) In: Research EAfP (ed) 19th Triennial Conference of the European Association for Potato Research, Brussels, vol 1. European Association for Potato Research, p 65
Fox A, Collins LE, Macarthur R, Blackburn LF, Northing P (2017) New aphid vectors and efficiency of transmission of potato virus a and strains of potato virus Y in the UK. Plant Pathol 66:325–335
Funke CN, Nikolaeva OV, Green KJ, Tran LT, Chikh-Ali M, Quintero-Ferrer A, Cating RA, Frost KE, Hamm PB, Olsen N, Pavek MJ, Gray SM, Crosslin JM, Karasev AV (2017) Strain-specific resistance to potato virus Y (PVY) in potato and its effect on the relative abundance of PVY strains in commercial potato fields. Plant Dis 101:20–28
Glais L, Tribodet M, Kerlan C (2005) Specific detection of the PVYN-W variant of potato virus Y. J Virol Methods 125:131–136. https://doi.org/10.1016/j.jviromet.2005.01.007
Gray S, de Boer S, Lorenzen J, Karasev A, Whitworth J, Nolte P, Singh R, Boucher A, Xu H (2010) Potato virus Y: an evolving concern for potato crops in the United States and Canada. Plant Dis 94:1384–1397. https://doi.org/10.1094/pdis-02-10-0124
Hane DC, Hamm PB (1999) Effects of seedborne potato virus Y infection in two potato cultivars expressing mild disease symptoms. Plant Dis 83:43–45
Kamangar SB, Smagghe G, Maes M, De Jonghe K (2014) Potato virus Y (PVY) strains in Belgian seed potatoes and first molecular detection of the N-Wi strain. J Plant Dis Prot 121:10–19
Karasev AV, Gray SM (2013) Genetic diversity of potato virus Y complex. Am J Potato Res 90:7–13. https://doi.org/10.1007/s12230-012-9287-7
Karasev AV, Nikolaeva OV, Hu XJ, Sielaff Z, Whitworth J, Lorenzen JH, Gray SM (2010) Serological properties of ordinary and necrotic isolates of potato virus Y: a case study of PVYN misidentification. 87. https://doi.org/10.1007/s12230-009-9110-2
Karasev AV, Hu X, Brown CJ, Kerlan C, Nikolaeva OV, Crosslin JM, Gray SM (2011) Genetic diversity of potato virus Y-O and origin of recombinant PVY strains. Phytopathology 101:778–785
Kerlan C, Nikolaeva OV, Hu XJ, Meacham T, Gray SM, Karasev AV (2011) Identification of the molecular make-up of the potato virus Y strain PVYZ: genetic typing of PVYZ-NTN. Phytopathology 101:1052–1060. https://doi.org/10.1094/phyto-11-10-0317
Lindner K, Billenkamp N (2005) Changes in the spectrum of PVY strain group: could this be a cause for the increase of virus susceptibility of potato and tobacco varieties. Nac De Pfla 57:245–253
Lindner K, Trautwein F, Kellermann A, Bauch G (2015) Potato virus Y (PVY) in seed potato certification. J Plant Dis Prot 122:109–119
Macaulay EDM, Tatchell GM, Taylor LR (1988) The Rothamsted insect survey ‘12-metre’ suction trap. Bull Entomol Res 78:121–128
Moury B, Fabre F, Senoussi R (2007) Estimation of the number of virus particles transmitted by an insect vector. Proc Natl Acad Sci USA 104(45):17891–17896. https://doi.org/10.1073/pnas.0702739104
Nolte P, Whitworth JL, Thornton MK, McIntosh CS (2004) Effect of seedborne potato virus Y on performance of Russet Burbank, Russet Norkotah, and Shepody potato. Plant Dis 88:248–252
Petrov N, Stoyanova M, Andonova R, Teneva A (2015) Induction of resistance to potato virus Y strain NTN in potato plants through RNAi. Biotechnol Biotechnol Equip 29:21–26
Perez I (2011) Virus protocol. In CIP (ed) Standard evaluation trials. https://research.cip.cgiar.org/confluence/display/SET/Virus+Protocol
Rigotti S, Balmelli C, Gugerli P (2011) Census report of the potato virus Y (PVY) population in Swiss seed potato production in 2003 and 2008. Potato Res 54:105–117. https://doi.org/10.1007/s11540-010-9183-9
Rolland M, Lacroix C, Blanchard A, Baldwin T, Kerlan C, Jacquot E (2008) Potato virus Y (PVY): from its discovery to the latest outbreaks. Virologie 12:261–273. https://doi.org/10.1684/vir.2008.0190
Rykbost KA, Hane DC, Hamm PB, Voss R, Kirby D (1999) Effects of seedborne potato virus Y on russet Norkotah performance. Am J Potato Res 76:91–96
Schubert J, Fomitcheva V, Sztangret-Wisniewska J (2007) Differentiation of potato virus Y strains using improved sets of diagnostic PCR-primers. J Virol Methods 140:66–74. https://doi.org/10.1016/j.jviromet.2006.10.017
Sigvald R (1985) Mature-plant resistance of potato plants against potato-virus Yo (PVYo). Potato Res 28:135–143
Swisspatat (2012) Données statistiques 2012 sur la mise en valeur des pommes de terre. Swisspatat, Bern
Valkonen JPT, Rokka VM, Watanabe KN (1998) Examination of the leaf-drop symptom of virus-infected potato using anther culture-derived haploids. Phytopathology 88:1073–1077. https://doi.org/10.1094/phyto.1998.88.10.1073
van der Vlugt RAA, Verbeek M, Cuperus C, Piron PGM (2007) Strains of potato virus Y in Dutch seed potato culture, vol 51. EAPR, Coylumbridge, Scotland
Verbeek M, Piron PGM, Dullemans AM, Cuperus C, van der Vlugt RAA (2010) Determination of aphid transmission efficiencies for N, NTN and Wilga strains of potato virus Y. Ann Appl Biol 156:39–49. https://doi.org/10.1111/j.1744-7348.2009.00359.x
Whitworth JL, Nolte P, McIntosh C, Davidson R (2006) Effect of potato virus Y on yield of three potato cultivars grown under different nitrogen levels. Plant Dis 90:73–76. https://doi.org/10.1094/pd-90-0073
Woodford JAT (1992) Virus transmission by aphids in potato crops. Neth J Plant Pathol 98:47–54
Acknowledgments
The authors thank Aurelie Buchwalder, Emile Schaer, Maud Tallant, Nathalie Dubuis, Justine Brodard, Henri Gilliand, Werner Wild and Peter Frei for their technical support. The authors also thank Thomas Steinger, Jacques Derron, Gabriel Goy and Floriane Bussereau for supplying the data on aphid flights.
Funding
This work was undertaken within the framework of the programme of activities of Agroscope.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Human and Animal Rights
This research does not include any animal and/or human trials.
Ethical Approval
The authors bear all the ethical responsibilities of this manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Dupuis, B., Bragard, C. & Schumpp, O. Resistance of Potato Cultivars as a Determinant Factor of Potato virus Y (PVY) Epidemiology. Potato Res. 62, 123–138 (2019). https://doi.org/10.1007/s11540-018-9401-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11540-018-9401-4