
Abstract
Obesity is a weight-related disorder characterized by excessive adipose tissue growth and dysfunction which leads to the onset of a systemic chronic low-grade inflammatory state. Likewise, inflammation is considered a classic cancer hallmark affecting several steps of carcinogenesis and tumor progression. In this regard, novel molecular complexes termed inflammasomes have been identified which are able to react to a wide spectrum of insults, impacting several metabolic-related disorders, but their contribution to cancer biology remains unclear. In this context, prostate cancer (PCa) has a markedly inflammatory component, and patients frequently are elderly individuals who exhibit weight-related disorders, being obesity the most prevalent condition. Therefore, inflammation, and specifically, inflammasome complexes, could be crucial players in the interplay between PCa and metabolic disorders. In this review, we will: 1) discuss the potential role of each inflammasome component (sensor, molecular adaptor, and targets) in PCa pathophysiology, placing special emphasis on IL-1β/NF-kB pathway and ROS and hypoxia influence; 2) explore the association between inflammasomes and obesity, and how these molecular complexes could act as the cornerstone between the obesity and PCa; and, 3) compile current clinical trials regarding inflammasome targeting, providing some insights about their potential use in the clinical practice.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Background
Prostate cancer (PCa) is a heterogeneous, complex, and strongly hormone-dependent disease that represents the second most frequently diagnosed cancer among the male population (14.1%), and the fifth leading cancer-related death in men worldwide among all cancer types (6.8%) [1]. In this context, the development and progression of PCa are controlled by different mechanisms that remain still not fully understood. Established risk factors for PCa development include age, ethnicity, and genetic background, while others like hormone levels, obesity, or prostatic inflammation have not been completely elucidated yet [2,3,4]. Inflammation is frequently associated with carcinogenesis and is also considered a common hallmark and a potential risk factor for the development of diverse malignancies through several mechanisms, including soluble molecules provided by the tumor microenvironment, extracellular matrix remodeling, angiogenesis, invasion, metastasis, and even epithelial to mesenchymal transition (EMT) [5, 6]. In fact, about 20% of all cancers are linked to some form of inflammation [7].
In the case of PCa, chronic inflammation has been proposed as a potential link between environmental stimuli and tumor occurrence. In this sense, the patients with high-grade inflammation surrounding malignant glands in radical prostatectomy specimens had significantly more biochemical PCa recurrence than patients with low-grade inflammation [8]. In fact, different studies have evaluated the relation between the onset of prostate gland abnormalities and some degree of inflammation, showing a high prevalence of mild chronic inflammation in benign prostate hyperplasia (BPH) and PCa samples [9,10,11,12,13].
In addition, genetic polymorphisms association studies highlighted that seven inflammation-related genes (CXCL12, IL4, IL6, IL6ST, PTGS2, STAT3, and TNF) may play a role in the development and aggressiveness of PCa [14]. However, during the last years, subsequent metanalyses have also revealed controversial results [15,16,17,18], suggesting that a positive association between prostate inflammation and PCa could be a consequence of detection bias [19], or that even such relation could be inverse [20]. In addition, reinforcing the idea that inflammation plays a key role in PCa progression, a type of proliferative inflammatory atrophy (PIA) has been associated with increased chronic inflammatory cell infiltration. These lesions are considered a possible precursor of high-grade prostatic intraepithelial neoplasia (PIN) and PCa by some authors and are usually found in the peripheral zone of the prostate, wherein PCa more frequently occurs [15, 21].
Therefore, increasing evidences suggest that inflammation could influence PCa development, and that immune cells could act as primary drivers. In fact, changes in the proportions and ratios of different immune cells populations are observed throughout different stages of PCa compared with healthy tissue [22]. Hence, it has been hypothesized that specific inflammatory infiltrate components are involved in PCa initiation or progression through the recruitment of additional immune cells and subsequent production of cytokines that ultimately creates a tumor microenvironment that promote cell replication, angiogenesis, EMT, migration, and metastasis [3, 23]. In this sense, initiation of prostatic inflammation could be driven by multiple and diverse agents, including uric acid (due to urine reflux), hormonal changes, physical damage, dietary components like heterocyclic amines, or microorganism presence [24].
In this context, a study focusing on the analysis of the urinary microbiome in urine samples suggested a prevalence of pro-inflammatory bacteria uropathogens in the urinary tract of men with PCa [25]. In fact, there is a close relationship between microorganism-originated inflammation, oxidative stress, and genomic instability. Indeed, bacterial prostatic colonization and/or dysbiosis could trigger signaling pathways that lead to immune cell infiltration, which can contribute to chemo-cytokines, growth factors, and ROS production [26]. In this scenario, it has been proposed that a prolonged oxidative stress environment could generate DNA damage (while decreasing DNA repair ability), which, added to the proinflammatory microenvironment, could derive into the above-mentioned PIA, followed by low or high-grade PIN, and ultimately, PCa [27]. Additionally, genetic changes and epigenetic background could drive chronic inflammation in the prostate epithelial cells [28].
Furthermore, weight-related disorders, including overweight or obesity have been postulated to mediate prostate inflammation [29]. Specifically, obesity is a metabolic pathogenic state that courses with systemic chronic low-grade inflammation and high circulating levels of inflammation-related markers (e.g., leptin, IL-6, TNF), which may affect tumor growth [30, 31]. In fact, adipocyte hypertrophy during obesity has been demonstrated to cause dysfunction and inflammation of the adipose tissue (AT) by increasing the secretion of pro-inflammatory cytokines [31, 32]. The relationship between obesity and PCa has been widely explored during the last decade [33,34,35,36]. Different studies reveal somewhat conflicting results, but there seems to be a general trend showing that obesity increases the risk of undergoing aggressive PCa [4]. Notwithstanding both pathological conditions often coexist, the direct molecular mechanisms that connect obesity and PCa are still poorly known.
In this context, inflammasomes have emerged as master regulators of inflammation by enabling the maturation of pro-inflammatory molecules through caspase-1 activation [37]. Evidences links the dysregulation of different types of inflammasomes (or some of their components) with several types of cancer, such as glioblastoma, hepatocellular carcinoma, mesothelioma, colorectal, gastric, lung, breast cancers, head and neck squamous cell carcinoma, oral squamous cell carcinoma or melanoma [38,39,40].
This review aims to summarize the current knowledge on the contribution of the different components of the inflammasome in PCa, incorporating, for the first time, its relationship with obesity and specifically with AT depots, as well as the potential of these inflammatory complexes as therapeutic tools to tackle PCa.
2 The inflammasomes
As the first line of defense, the innate immune system has evolved to detect and react to a wide variety of insults derived from external and self-signals. Those stimuli include conserved structures among microorganisms, which are called pathogen-associated molecular patterns (PAMPs), and endogenous molecules associated with components of host cells that are associated to cell damage, stress, or death, termed damage-associated molecular patterns (DAMPs, also known as alarmins) [41]. These signals include reactive oxygen species (ROS), oxidized and/or methylated DNA, the presence of abnormal proteins in certain cellular compartments, increased levels of ATP, free fatty acids, ceramides, homocysteine, and calcium or cholesterol crystals, among others [42].
To recognize these signals, the innate immune system possesses, among its immunological arsenal, the germline-encoded pattern recognition receptors (PRRs), which can be classified into membrane-bound and cytosolic sensors. Membrane-bound sensors include Toll-like receptors (TLRs) and C-type lectin receptors (CLRs), while cytosolic sensors comprise the nucleotide-binding oligomerization domain-like receptors, or NOD-like receptors (NLRs), and the retinoic acid-inducible gene 1 (RIG-1)-like receptors (RLRs) [43,44,45]. Particularly, the NLR family comprises 22 genes that are characterized by the presence of a central NACHT domain, along with other variables such as PAAD/DAPIN (PYD) and caspase activation and recruitment (CARD) domains, both involved in homotypic protein-protein interactions [46].
Inflammasomes are cytosolic complexes capable of triggering an inflammatory response through the autoactivation of caspase-1, which leads to proteolytic cleavage, maturation, and secretion of pro-inflammatory molecules, including the interleukin-1β (IL-1β) and interleukin-18 (IL-18). These two cytokines are synthesized as the inactive precursors pro-IL-1β (p31) and pro-IL-18 (p24), which are finally processed through inflammasome activation to their mature form IL-1β (p17) and IL-18 (p18), respectively [47, 48]. While pro-IL-18 is constitutively expressed, pro-IL-1β expression is often upregulated after ligand-mediated activation of TLRs or NLRs family members, constituting a previous and limiting step for the activation of certain inflammasomes activation, termed priming [49]. In addition, a third product derived from inflammasome activation, gasdermin D (GSDMD), has been recently described (Fig. 1). Once cleaved, this protein is able to initiate an inflammation-mediated cell death process known as pyroptosis [50].

General NLRP3 inflammasome structure and activation. After priming and danger signal combination, inflammasome monomers composed by sensor molecular (NLRP3), assembly and activity regulator (NEK7), molecular adaptor (PYCARD) and enzymatic subunit (zymogenic CASP1) oligomerize forming a heptameric macromolecular platform that enables proteolytic autoactivation of CASP1 and processing of three proinflammatory molecules: pyroptotic-related membrane pore subunit Gasdermin D (GSDMD), and interleukins-1-β and-18
Inflammasomes usually include 3 types of proteins in a heptameric conformation: 1) a member of the NLR family protein that acts as the stimuli sensor; 2) the molecular adaptor ASC (also known as PYCARD, containing both PYD and CARD domains), which serves as a scaffold; and 3) the inactive zymogen form of the caspase-1, recruited for final autoactivation. According to the specific inflammasome composition, the molecular adaptor ASC may not be necessary since the sensor protein could directly interact with caspase-1 [51]. There are different types of inflammasomes, named according to their sensor component. Of these, NLRP1, NLRP3, NLRC4, AIM2, and pyrin (a PYD domain-containing protein) inflammasomes are considered canonical, while recent studies have identified non-canonical or atypical inflammasomes existence, like NLRP2, NLRP6, NLRP7, NLRP12, and IFI16 inflammasomes [52,53,54,55,56,57]. In addition, a fourth component, NEK7, has been recently discovered to be essential for oligomerization and triggering of some types of inflammasomes [58]. Remarkably, CASP8 activating inflammasomes have been described, and active forms of IL-33 or IL-37 have also been proposed to be produced by certain inflammasomes types [59, 60].
Distinct inflammasomes are activated in response to a plethora of stimuli with different action mechanisms. Among them, the NLRP3 inflammasome is currently the most fully characterized [61, 62]. The proposed NLRP3 inflammasome activation mechanism follows a two-step model (Fig. 1). First, a signal acts as the priming step and licenses inflammasome by upregulation of NLRP3, IL-1β, and IL-18 through downstream activation of TLRs and MyD88, and subsequent NF-κB pathway activation, in an NLRP3/NEK7-dependent mechanism. Second, a hazardous signal is sensed by NLRP3, allowing inflammasome assembly and caspase-1 activation (Fig. 1). The activation of NLRP3 inflammasome across a wide diversity of stimuli allow the convergence of these signals into a common signaling mechanism [63]. The exact convergent process is unclear, but proposed mechanisms include mitochondrial ROS production, ATP-dependent potassium efflux, mitochondrial DNA release, lysosomal cathepsins release, changes in cell volume, and mitochondrial cardiolipin translocation [64,65,66].
Importantly, dysregulation of inflammasome complexes has risen as a critical molecular element in metabolic and cancer pathologies in humans [67,68,69,70]. Specifically, in the context of cancer development and progression, inflammasomes could play opposite roles like tumor-suppressor functions (by inducing cell apoptosis), or tumor-promoting events (by favoring secretion of pro-inflammatory cytokines). Such heterogeneous effects within the tumor microenvironment could be mediated by the tumor cells, the tumor-infiltrating myeloid cells, or both. Furthermore, despite the inflammasomes has been typically associated with myeloid cells such as monocytes, its activation has also been demonstrated in non-myeloid cells [37, 71]. Of special interest, functional inflammasomes have been found in prostate epithelial cells as well as in adipocytes, suggesting that they could be an essential supramolecular mediator in the potential crosstalk between both cell types under normal and tumoral conditions [71, 72].
3 Inflammasomes in prostate cancer
A growing body of evidence suggests inflammation as a potential element in PCa initiation and progression. Thus, the identification of inflammation regulators and associated molecules may provide an opportunity to understand the role of inflammation in PCa. In this context, the inflammasomes has emerged as a crucial player in the inflammatory state. In the next sections, we will discuss the current knowledge of inflammasome components and their potential role as regulators in PCa (listed in Table 1) pathophysiology,as well as their potential association with obesity.
3.1 Toll-like receptors (TLRs)
TLRs are membrane-bound danger-sensing receptors tightly associated with innate immune response. In humans, ten TLRs with differential specificity for distinct types of insults have been identified [43]. Despite TLRs constitute a group of well-characterized pro-inflammatory signaling pathways, recent advances have linked these receptors to inflammasome activation. In fact, previous studies have shown that TLR signaling is not only an enhancer of inflammasome response but also indispensable for certain inflammasomes types [51]. The most explored TLRs in the PCa context are TLR3, TLR4, and TLR9.
3.1.1 TLR3
TLR3 is an intracellular receptor located in endosomal membranes that is involved in the recognition of nucleic acids. TLR3 has been detected at mRNA and protein levels in different PCa cell models (LNCaP, PC-3, and DU145 cell lines). Moreover, TLR3 stimulation has been shown to increase the expression of inflammatory molecules through activation of the NF-κB pathway leading to the recruitment of specific leukocyte populations [73]. In addition, TLR3 has been proved to regulate apoptosis and angiogenesis in LNCaP and PC-3 cells through HIF-1α, a protein highly related to inflammasome activation as we will discuss below [74]. Furthermore, the expression of TLR3 has been corroborated in human samples, finding a significant and positive correlation between TLR3 levels and PCa recurrence [75].
3.1.2 TLR4
TLR4 recognizes lipopolysaccharides and constitutes the most studied TLR in PCa. Several genetic polymorphism studies have been performed correlating the presence of single nucleotide polymorphisms (SNPs) in the TLR4 gene with PCa risk development, showing conflicting results. Some authors concluded that inherited polymorphisms of TLR4 are associated with increased risk of PCa [76,77,78], while others have established that those polymorphisms were not significantly associated [79, 80]. Such differences could be explain due to the different number of patients included in and/or ethnic differences of the population studied. TLR4 has also been detected (at mRNA and protein levels) in distinct PCa cell lines (DU145 and PC-3) and tumor samples, despite at significantly lower levels as compared with BPH tissue [81]. Gatti and co-workers demonstrated that TLR4 levels decreased as the histopathological grade of tumors increased, and that such reduction was directly associated with epithelial, but not stromal, prostate cells. Interestingly, the same authors demonstrated that TLR4 activation in DU145 cells courses through NF-κB and MyD88-dependent pathways, resulting in increased expression of IL-1β mRNA (priming effect) and increased protein levels after TLR4 induction [82]. Additionally, TLR4 activation seems to be mediated by peroxiredoxin 1 (an antioxidant enzyme elevated and secreted by tumor prostate cells) conferring chemoresistance in PC-3 cells [83, 84]. Additionally, a recent work by Zhonghua and collaborators demonstrated that S100A9, classically considered an alarmin, promotes cancer cell invasion in PC-3 and DU145 cells through TLR4 binding in an NF-kB-dependent mechanism [85].
3.1.3 TLR9
On the contrary, TLR9, is a receptor associated with inner cell membranes. TLR9, which recognizes CpG oligonucleotides, has been detected at the protein level in DU145, PC-3, LNCaP, and C4-2B, as well as in human samples of PCa [86]. Indeed, TLR9 levels were associated with poor progression-free survival in PCa patients [87]. In addition, TLR9 up-regulates cyclooxygenase 2 (COX-2) expression in PC-3 cells through NF-κB activation, which has been suggested by some authors to be implicated in tumor invasion and metastasis [88].
Taken together, TLRs activation itself, along with distinct inflammasome licensing and pathway cross-talking, might induce early immune responses in the tumor microenvironment that could influence the fate of PCa progression.
3.2 Sensor component
The essential difference among inflammasome subtypes is the sensor protein, which gives specificity against stimuli that trigger inflammasome assembly. In the next subsections, we compile current information about AIM2 and NLRP3 components, the most explored in the context of carcinogenesis and PCa.
3.2.1 AIM2
AIM2 encodes a cytosolic member of the IFI20X/IFI16 (PYHIN) protein family. This protein associates with cytoplasmic double-stranded DNA (dsDNA) via a HIN200 domain, while its PYD domain interacts with the adapter molecule ASC to activate caspase-1, conforming the AIM2 inflammasome [89,90,91]. AIM2 inflammasome has been postulated to be activated in response to exogen and self-derived dsDNA, enabling pro-inflammatory cytokines production [71]. Moreover, AIM2 has been associated with different tumors subtypes wherein classically has been described as a tumor suppressor. For instance, Aim2−/− models showed a higher susceptibility to develop colon cancer compared to control mice, although cytokine production and inflammasome activation is not compromised. In this sense, it has been postulated that AIM2 could be exerting this tumor suppressor role through the regulation of PTEN on the ATK pathway, and by the activation of the proto-oncogene c-Myc, in a very specific context where gut microbiota and the AIM2 dsDNA sensing capacity could be crucial, although mechanistic details are lacking [92]. However, it should be mentioned that a prooncogenic role for AIM2 has also been described in squamous cell carcinoma, and endometrial cancers [93].
AIM2 is expressed in human normal prostate epithelial cells and its expression, along with that of other key inflammasome components (i.e., pro-caspase-1 and pro-IL-1β), is potentiated when prostate cells are primed with INFα, β, and γ [71]. Furthermore, steady-state levels of AIM2 are significantly lower in young epithelial prostate cells than in senescent cells. Accordingly, AIM2 levels were also significantly lower in normal prostate epithelial cells compared with the BPH-1 cell line, suggesting that the accumulation of prostate cells with a senescent secretory phenotype (also refer as SASP) may play an important role in BPH initiation [71]. Surprisingly, AIM2 mRNA levels were significantly lower in various PCa cell lines (DU145, PC-3, and LNCaP) as well as in prostate adenocarcinoma samples compared with normal prostate epithelial cells [71, 94]. Notably, it has been proven that primed AIM2 inflammasome is activated in the presence of cytosolic dsDNA in normal epithelial prostate cells and PC-3 cells [71].
Panchanathan and cols. recently demonstrated that hypoxic conditions often associate with inflammation in solid tumors, and, specifically with tumor promotion, resistance to therapy, and malignant progression in PCa [95]. Thus, hypoxic conditions prime and activate AIM2 inflammasome in human normal epithelial cells, besides promoting the activity of those previously primed by cytosolic dsDNA [95]. In line with this, hypoxia also increases caspase-1 activity levels in normal prostate epithelial cells, BPH-1 and in PC-3 cell line via NF-κB pathway activation. Similar results were obtained in the myeloid cell line TPH-1 [95]. Furthermore, hypoxia and inflammation keep a feed-forward loop relation in prostatic tumors, enabling tumor promotion, malignant progression, resistance to cancer therapy, and consequently poor outcome for PCa patients [96, 97]. With all this evidences, we could conclude that AIM2 may represent a protein with an important role in oncogenesis in a tissue and context-dependent manner. Therefore, a special effort should be made to clarify its potential consequences in PCa development.
3.2.2 NLRP3
NLRP3 inflammasome is characterized by the ability to respond to a wide spectrum of stimuli. However, this inflammasome subtype rarely responds to a single signal. NLRP3 inflammasome possesses a double-step initiation model that requires previous priming, resulting in NLRP3 and IL1B genes upregulation [63]. This dual signal requirement may act as a checkpoint mechanism to avoid accidental or uncontrolled NLRP3 activation. In cancer research, NLRP3 inflammasome has been also widely explored, appearing to be relevant in such diverse tumors as glioblastoma, and colorectal or liver cancers. Moossavi and colleagues recently summarized the known role of NLRP3 inflammasome in cancer [39]. However, scarce, and conflicting information is available in PCa. Some studies reveal that this sensor component can be detected in several prostate-derived cell lines (RWPE-1, PZ-HPV-7, LNCaP, Ln3, 22Rv1, PC-3, and DU145) at mRNA and protein levels, while other studies indicate that NLRP3 protein and transcripts were undetectable or extremely low in PC-3 cells [98]. A recent study showed that NLRP3 expression level was significantly high (at mRNA and protein levels) in various PCa cell lines (i.e., DU145, 22Rv1, PC-3, LNCaP) compared with the healthy cell line WPMY-1. In humans, PCa samples have been shown to exhibit heterogeneous immunostaining for NLRP3 compared with normal adjacent tissue [99], while other studies indicate that NLRP3 expression levels are significantly high in PCa tissues, wherein this overexpression is correlated with clinical parameters such as TNM stage and lymph node invasion [98].
Panchanathan and cols. also demonstrated a post-transcriptional stabilization of NLRP3 by hypoxia. Apart from acting as a priming agent in human normal prostate cells, benign BPH-1, and tumor PC-3 cell lines, hypoxia also resulted in enhanced activation of NLRP3 inflammasome [95]. Unlike that observed in the case of AIM2 inflammasome, hypoxia was not sufficient to prime or activate NLRP3 inflammasome in the myeloid cell line TPH-1 [95]. Finally, recent works have demonstrated that blocking NLRP3 pathway in PC-3 cells and PCa-derived xenografts have a significant impact on tumor progression by disrupting cell proliferation, migration, and/or inducing apoptosis [98, 100, 101].
Considering these results, it might be suggested that among the complex tumor microenvironment, distinct stimuli could trigger specific inflammasomes in a cell type-specific manner, which could finally affect the development of prostate inflammatory-related pathologies, including cancer.
3.3 Molecular adaptor: PYCARD (ASC)
Apoptosis-associated Speck-like protein containing a CARD (ASC), also referred as PYCARD, is a molecular adaptor necessary for assembling different types of inflammasomes. Through PYD and CARD domains, PYCARD participates in canonical (NLRP3, AIM2) and non-canonial (NLRP6, NLRP7, NLRP12, IFI16) inflammasomes [51]. Even though some sensor components like NLRP1 harbor domains that enable direct interaction with caspase-1, the presence of the PYCARD adaptor seems to be crucial to reach the maximum capacity of inflammasome [102].
PYCARD has been one of the most studied inflammasome components as it was first described as a methylation-associated silenced gene in human breast cancer cells [103]. Furthermore, PYCARD silencing and methylation status has been reported in several other neoplastic diseases such as colorectal, lung and ovarian cancer, melanoma, neuroblastoma, and glioblastoma [104,105,106,107,108,109], and has typically been supposed to act as a tumor suppressor implicated in caspase-mediated apoptosis (involving predominantly CASP8 and CASP9), and mitochondrial translocation of BAX coupled to activation of CASP9, CASP2, and CASP3 [110, 111]. However, since inflammasomes were discovered, ASC has been proposed to be implicated in tumor development/progression through different indirect mechanisms. Thus, in gastric cancer, it has been suggested the existence of potential signaling crosstalk between the activation of ASC inflammasomes (through NF-κB) and CASP8 apoptotic machinery in an intricate mechanism [112]. This likely dual role of PYCARD [apoptotic/tumor-suppressor vs. pro-inflammatory/oncogene (as happened with the above-mentioned AIM2)] in different cancer types has been recently reviewed [113].
In PCa, methylation of PYCARD has also been demonstrated in several tumor cell lines, including LNCaP, LAPC-4, MDAPCa2b, DU145, and PC-3 [114, 115], but not in the benign prostate epithelial cell line BPH-1, or normal prostate epithelial cells. Furthermore, Collard and cols. showed similar methylation trends of PYCARD with 64% of methylation in high-grade PIN samples examined, 65% in cancer specimens, and 28% in the adjacent normal tissue, while all normal donor prostate samples presented unmethylated PYCARD promoter [114]. However, further investigation is needed to unveil the intricate role of this multifunctional protein within this tumor pathology.
3.4 Caspase 1
The CASP1 gene encodes for caspase-1, a member of the cysteine-aspartic acid protease family that plays a central role in the execution-phase of cell apoptosis and the proteolytical activation of the inactive precursor of pro-inflammatory molecules as it is the enzymatic component of inflammasomes. Caspase-1 has recently gained attention as it may play a crucial role in human carcinogenesis [116]. In fact, several studies demonstrated a significant downregulation of CASP1 expression in different types of cancers, including renal, breast, colon, or ovarian cancer, while upregulation has been reported in pancreatic tumors [117,118,119,120,121].
In the case of PCa, immunohistochemical analysis showed significantly decreased levels of the active form of CASP1 protein compared with the normal gland [122]. A constitutive expression level of the proenzyme form of CASP1 in human PCa cell lines (PC-3, DU145, and LNCaP) has been also reported [122]. The loss of CASP1 has also been proposed as a required step in the dysregulation of apoptotic control during PCa development [122, 123]. Recently, Chang and cols. demonstrated that reduced in vivo levels of CASP1 may be due to a negative modulation effect of cytochrome P450-1B1 (CYP1B1) [124]. In line with this, the authors also found an inverse correlation between CYP1B1 and caspase-1 in human tissue specimens [124].
3.5 Inflammasome targets
3.5.1 IL-1β
IL-1β is a major upstream cytokine that controls the local pro-inflammatory status and can be produced by monocytes, fibroblast, and epithelial cells as a paracrine or autocrine signal. IL-1β is one of the seven ligands included in the IL-1 family, which require inflammasome action to reach the mature and active form that binds to its heterodimeric IL-1RI/IL-1RAcP receptor on target cells to initiate IL-1β signaling [125]. IL-1β recognition trough IL1R leads to different associated kinases family (IRAK) assembly around Myd88, forming other types of macromolecular complexes (myddosomes) that may play an important role in myeloid malignancies and in the adaptative resistance to various forms of cancer therapy [126].
Due to IL-1β key role in the inflammatory response, its production affects the balance established between protective immunity and destructive inflammation. However, the role of IL-1β in tumor biology remains controversial. Some authors have proposed that IL-1β is able to induce tumor angiogenesis and metastatic spread, while an antitumor role has also been suggested [127,128,129]. In this sense, the role of IL-1β in PCa is still unclear. Different IL-1β polymorphisms have been associated with either increased or decreased PCa risk, providing additional evidence that this cytokine may play an important role in PCa etiology [130, 131]. Notably, Ricote and cols. demonstrated the immunohistochemical presence of IL-1β in 75% of normal prostate samples, in 42.86% of BPH samples, and in 31.25% of PCa samples with low Gleason grade, while no positive immunoreactions were observed in PCa samples with high Gleason grade, suggesting that a progressive loss of IL-1β is associated with more aggressive pathologies [132]. In line with these results, external reestablishment of IL-1β provokes growth inhibition in LNCaP and androgen-independent PC-3 and DU145 cell lines [133]. Furthermore, in LNCaP cells, IL-1β exposure is also able to decrease PSA production, Androgen Receptor (AR) mRNA/protein levels, and causes antiandrogen bicalutamide resistance after a few passages, linking the cytokine with castration resistant prostate cancer (CRPC) development [134]. Chang and cols. identified IL-1β as the HS-5 bone marrow stromal cell-secreted paracrine factor responsible for AR mRNA downregulation in C4-2 PCa cell line (LNCaP derived subcutaneous xenograft tumor of castrated mouse) through upregulation of p62, a multifunctional adaptor needed for NF-κB pathway activation [135].
A body of evidence points out that IL-1β exerts its effects on PCa through the NF-κB pathway [133, 135]. NF-κB is generally associated with antiapoptotic and pro-inflammatory effects, and recent evidence suggests that NF-κB may have anti-inflammatory roles and promote apoptosis during tumor progression [136, 137]. NF-κB is constitutively activated in diverse PCa cell lines and human prostate adenocarcinoma, while trans-localization into the nucleus of PCa cells positively correlates with a high Gleason score, biochemical recurrence, and metastasis due to its transcriptional regulation activity [138, 139]. A detailed list of proposed targeted genes regulated by NF-κB in PCa is available [138]. Of note, interleukin-6 (IL-6), a cytokine widely studied in PCa, is upregulated by NF-κB action and it has been proposed as a key factor in the transition from hormone-dependent to CRPC [140]. In fact, NF-κB pathway activation itself seems to be sufficient to allow prostate and PCa to grow in an androgen-independent manner through AR activity regulation, confer protection against apoptosis, and participate in the induction of metastatic phenotype, thus being considered the most important effector of pro-inflammatory processes involved in PCa pathogenesis by various authors [138, 141, 142].
Notably, NF-κB activation also produces upregulation of IL-1β, therefore causing a positive feedback loop [138]. Hypoxia regulator HIF-1α induced by Prx1 also promoted NF-κB activity, which has the potential to perpetuate Prx1 induction of angiogenesis in PCa. In addition, this mechanism occurs through TLR4-dependent activation, which has been related to the inflammasome priming phenomenon [83]. In this regard, it has been suggested that hypoxia can induce AIM2 and NLRP3 inflammasome activation and consequent IL-1β production through stimulation of the NF-κB pathway in human normal prostate, PC-3, and myeloid THP-1 cell line [95]. Notably, the expression of the master regulator of hypoxia HIF-1α increases in response to IL-1β through NF-κB activation, and IL-1β itself can stabilize HIF-1α even under normoxia conditions, through a ROS-dependent mechanism [143, 144]. Moreover, hypoxia strongly induced the expression of PD-L1 in 22Rv1, DU145 and PC3 cell lines, while HIF1-α knockdown and chrysin treatment (an inhibitor of HIF1-α stability) decreases PD-L1 protein levels (except for 22Rv1) [145]. Interestingly, it seems to be a mutual crosstalk, between the exploited PD-1/PD-L immunosuppressive system, that drives immune scape and tumor growth, and inflammasomes. For example, it is known that AIM2 can regulate PD-L1, and that IL-1β is able to increase PD-L1 at the membrane of cancer cells and CAFs. A review has recently been published addressing this connection [60].
In a recent work, Tran and colleagues explored the link between HIF1-α and oncogenic AR signaling in CRPC development through transcriptomic profiles, concluding that both pathways occur in an independent manner [146]. However, it is feasible that hypoxia and therefore increased HIF1-α levels could alter AR signaling and promote CRPC development through a complex network of inflammasome-derived cytokines such as IL-1β. Interestingly, ROS, which are also capable to stabilize HIF-1α, have been proposed to be necessary, but not sufficient, for triggering inflammasome activation [51, 144]. Indeed, a double role for ROS has been suggested concerning inflammasomes functioning: first, ROS are thought to be able to direct priming and triggering some types of inflammasomes; additionally, ROS can also exert an indirect inductor effect through cytoplasmic proteins that control inflammasomes activity. Moreover, ROS has been proposed as a likely common signal throughout different stimuli that can converge in inflammasome activation [147].
In a hypoxic-ROS enriched microenvironment, DNA damage and repair mechanisms disfunction, could contribute significantly to the establishment of tumor origin. For example, Rar50 (a major constituent of the MRN-DSBs [Mre11-Rad50-Nbs1 Double Strand Breaks] complex), is needed for the NF-kB activation and pro-IL1-β production. Moreover, Rad50 is also implied in viral and bacterial dsDNA recognition, elements that trigger certain inflammasomes types [148].
In addition, IFN signaling (released upon viral infection), which is able to prime and activate AIM2 inflammasome, interacts with the NF-κB pathway providing an additional link between inflammasomes and NF-κB and PCa [71, 137]. Of note, NF-κB signaling is a major inducer of SASP, which is related to inflammasome component AIM2 accumulation and BPH initiation [149]. Taken together, current knowledge places inflammasomes in a central position within a complex crossover of pathways orbiting around danger or stress signals, IL-1β and NF-κB (Fig. 2).

The central role of inflammasomes and NF-kB signaling in prostate cancer. Different types of inflammasomes are capable of activating and responding to a wide variety of stimuli such as pathogens (PAMPs) or stress situations intrinsic to the cell itself (DAMPs). The activation of the inflammasome is enhanced and shares some signaling in common with the hypoxia signaling pathway, which in turn exacerbates inflammasome activation, establishing multiple positive feedback loops. As a result, it exists a convergence in terms of signaling through NF-kB, which acts as a transcription factor of essential genes in the development and progression of prostate cancer toward more aggressive phenotypes
3.5.2 IL-18
IL-18 is a pleiotropic cytokine that plays a central role in inflammation and immune response by exerting several functions, mostly related to immunomodulation in macrophages, T-helper type I (Th1), and natural killer (NK) cells [150]. Like IL-1β, IL-18 reaches its active form after caspase-1 processing activity. IL-18 has been widely associated with antitumor effects in different types of cancer by immunosurveillance modulation through different immune cell types [151]. Despite its antitumor effects, IL-18 is expressed in PC-3 and DU145 cell lines after IFN-γ induction [152]. In line with this, PCa samples also exhibit IL-18 overexpression, being its levels associated with a better clinical outcome [152, 153]. However, IL-18 binding protein (IL18BP), a glycoprotein with a high affinity to IL-18 able to neutralize it, is also overexpressed in PCa human samples and some PCa cell lines under IFN-γ stimulation. Noteworthy, IL-18BP serum levels correlate with the Gleason score [151]. Taking all these evidences together, it could be suggested that these processes might be involved in a regulation mechanism of inflammasome assembly and caspase-1 activation affecting prostate and contributing to PCa development, which needs to be further investigated.
3.5.3 Gasdermin D
While most cytokines include an N-terminal sequence related to its secretion, members of the IL-1 family, including IL-1β and IL-18, do not possess any known sequence associated with extracellular release [154]. Recently, a novel cytokine releasing mechanism involving a third inflammasome product, Gasdermin D (GSDMD) has been discovered. GSDMD belongs to the gasdermin family, together with GSDMA, GSDMB, and GSDMC. This protein remains inactive until cleavage activation by several caspases (including inflammasome component caspase-1); once cleaved, GSDMD oligomerize on membranes through specific phospholipid interactions, forming pores. It has been suggested that low levels of GSDMD form functional pores, which is associated with long-term IL-1β release by still-living cells [155]. However, it has been proposed that at high GSDMD levels, cell membrane repair activity would not be sufficient to avoid cellular death, provoking the releasing of intracellular content causing a highly inflammatory uncontrolled death known as pyroptosis. Ultimately, GSDMD pores have been proposed to enable potassium efflux alteration, leading to NLPR3 inflammasome activation and to the potentiation of a positive feedback loop [156]. Such inflammatory cell death may play a dual role in promoting or inhibiting tumor cell growth in different cancer types, showing controversial results [157]. Particularly, GSDMD contribution to cancer development is poorly explored and has only been reported by Wang and cols. in gastric cancer, wherein GSDMD is downregulated among different gastric cancer cell lines by inhibiting the cyclinA2/CDK2 complex and arresting into the S/G2 phase transition [158]. This study also proposes GSDMD as a tumor suppressor acting through ERK, STAT3, and PI3k pathway inactivation. Although to date, GSDMD role in PCa is completely unexplored, Zhang and cols. recently demonstrated GSDME ability to induce pyroptosis in PC-3 cells through PKCδ/JNK signal activation [123].
4 Obesity, adipose tissue, and inflammasomes in prostate cancer
According to World Health Organization (WHO), obesity is defined as a condition of excessive fat accumulation in adipose tissue (AT) due to a positive energy balance as a consequence of an increased energy intake and/or decreased energy expenditure [159]. Obesity has risen in the last decades as a major global issue, reaching pandemic proportions and becoming a real challenge for healthcare systems [160, 161]. Obesity is characterized by AT disfunction; this tissue constitutes a complex, heterogeneous, and metabolically active organ that has been classically proposed to comprise mature adipocytes and the stromal-vascular fraction (SVF), composed of preadipocytes, fibroblasts, vascular endothelial cells, and a variety of immune cells including macrophages, lymphocytes, mast cells, neutrophils, eosinophils, and foam cells [162,163,164]. Pathophysiologically, obesity triggers an inflammatory response orchestrated by adipocytes, then, milieu composition modification occurs (including extracellular remodeling and immune cell infiltration and activation) and, finally, a characteristic maintained/chronic low-inflammatory systemic state is established, a condition termed “metaflammation” by some authors [165]. Once established, this condition associates with comorbidities including type 2 diabetes (T2D), cardiovascular disease, atherosclerosis, hepatic steatosis, central nervous system dysfunction, dementia, and certain cancers, including PCa. In this context, AT is now considered an important mediator of systemic and local inflammatory responses, thus emerging as a key immunometabolic organ connecting lipid metabolism dysfunction and altered adipokine release, and distinct types of pathologies.
In that respect, inflammasomes exert a great impact on obesity and weight-related comorbidities. Specifically, Vandanmagsar and collaborators described increased mRNA expression of NLRP3 and IL-1β (but not ASC) in AT from patients with obesity and T2D, and these levels correlated with glycemic state [166]. In addition, the presence of NLRP3 inflammasome components in a mouse model of diet-induced obesity was higher in AT macrophages (ATMs) and SVF than in mature adipocytes of epididymal fat tissue, which suggested a theoretical model based on positive feed-forward loops among adipocytes, macrophages, and T cells [166]. Increased inflammation levels due to increased caspase-1 activity and IL-1β production by ATMs have been also described in visceral adipose tissue from metabolically unhealthy patients [167].
NLRP3 inflammasome is activated by ceramides sensing, inducing IL-1β production through caspase-1 activation, contributing to obesity-induced inflammation and insulin resistance. In line with this, cholesterol crystals and palmitate (the most increased serum fatty acid in obesity) have also been categorized as sterile inflammation instigators through NLRP3 inflammasome activation [166, 168]. In contrast, oleate, linoleate, and ω-3 fatty acids (docosahexaenoic and eicosapentaenoic acids) showed inhibitory effects [169]. Moreover, increased glucose levels also contribute to a low-chronic inflammatory state and insulin resistance partly through NLRP3 inflammasome activation and IL-1β production in subcutaneous adipose tissue (SAT) [170]. NLRP3 inflammasome has also been reported to be activated under obesity conditions through a hypoxia-dependent mechanism [171]. NLRP3, but not NLRP1 inflammasomes, are regulated by hypoxia conditions in visceral adipocytes [172]. Moreover, a recent report identified HIF-1α activation in ATMs responsible for local and systemic IL-1β maintained production under obesity conditions through ceramides sensing [173]. Moreover, it has been demonstrated that High Fat diet induces insulin resistance through NLRP3 inflammasome activation, and by a subsequent IL-1β secretion and ROS production in mouse AT [174]. Furthermore, in a T2D-induced mouse model, fenofibrate (a PPARα agonist) was able to inhibit NLRP3 inflammasome and the subsequent IL1-β production through a ROS-dependent mechanism that implies thioredoxin-interacting protein (TXNIP), improving the angiogenic epithelial precursor cells ability [175]. Altogether, these results indicate that the NLRP3 inflammasome can be regulated by distinct molecules under different metabolic states, and that these effects are mainly achieved through IL-1β production.
The role of IL-18 (also produced upon inflammasome activation) in obesity, remains controversial [176]. In this sense, serum levels of IL-18 correlate with insulin resistance and metabolic syndrome components. Strikingly, IL-18 has marked anti-obesity effects as interleukin administration prevents weight gain, while IL-18 loss is associated with increased adiposity and insulin resistance in mice models [177, 178]. Murphy and cols. demonstrated that IL-18 production relies on NLRP1 inflammasome. Using mice models, they demonstrated that loss of Nlrp1 decreased IL-18 levels, lipolysis, and led to obesity and metabolic syndrome [179]. Noteworthy, Raut and coworkers demonstrated that leptin, one of the major adipokines produced in obese conditions and related to tumor growth, is able to activate NLRP3 inflammasomes in MCF-7 breast cancer cells [180]. This induction, which courses with IL-1β production, and overexpression of NLRP3 and ASC component, was ROS dependent (in a mechanism involving NADPH oxidase) and was further corroborated in xenograft models. Raut and coworkers also demonstrated that adiponectin, an adipokine severely diminished in obesity that exerts anti-inflammatory and antitumor actions, completely suppressed leptin-induced NLRP3 inflammasome activation [181]. Interestingly, leptin has been shown to activate also NLRP3 inflammasome in macrophages increasing IL-18 secretion [182]. Altogether, these results unveil the complex relationship between adipokines and inflammasomes, which could be responsible, at least in part, for the inflammatory responses triggered in the AT under obesity conditions.
Investigation about other inflammasomes types in the obesity field is limited. Aim2 deficient mice exhibit impaired glucose homeostasis, upregulation of pro-inflammatory pathways, augmented adipogenesis, and inflammation in the AT with an increased rate of monocyte infiltration, highlighting the role of AIM2 inflammasomes in energy metabolism homeostasis [183]. In a novel study, NLRC4 inflammasome has been associated to breast cancer progression in obesity conditions. Specifically, NLRC4 mRNA is elevated in normal breast tissues from patients with obesity and positively correlates with BMI. Moreover, it has also been reported that obesity-induced NLRC4 inflammasome in tumor-infiltrating myeloid cells promotes angiogenesis in diet-induced obese mice through IL-1β, and that such activation would finally lead to increased Vegfa expression in surrounding adipocytes [184]. In fact, angiogenesis has also been suggested to be disturbed through NLRP3-related mechanisms in cardiovascular diseases influenced by short fatty chain acids [185]. In line with these results, it has been shown that, in an obese state, breast tissue-associated adipocytes recruit and activate macrophages through an IL-1β dependent mechanism [186]. Furthermore, a protective role has been suggested for NLRP12 inflammasome whose expression is reduced in the AT of humans with increased BMI. Also, Nlrp12 deficient mice showed increased weight gain, macrophages infiltration, and adipose tissue inflammation [187].
Furthermore, in this specific context, certain coenzymes have demonstrate to exert advantageous effects. Specifically, coenzyme Q10, which is considered to be a potential anti-cancer agent, shown to be beneficial when administered daily to overweight and obese patients with diabetes, improving glucose and lipid homeostasis [188]. However, weak associations have been found in PCa plasma patients [189]. Another coenzyme, Q0 was able to decrease HIF-1α expression, to ameliorate NLRP3-mediated inflammation, and suppress the EMT/metastasis and the metabolic rewiring in breast cancer [190].
Along with increased systemic inflammation levels, specific organs have also been related to inflammatory responses under obesogenic conditions. For example, inflammation in the liver and pancreatic islets is also evident in individuals with obesity [165]. These ‘from systemic to localized’ inflammatory responses could also affect the prostate under obesogenic conditions, leading, in the last instance, to prostate alterations, and malignancies. In this context, the relationship between obesity and some typical hallmarks of cancer, such as angiogenesis, EMT, genomic instability, and inflammation has been already described [191]. However, the casual link between obesity and PCa is more controversial. In fact, unlike other tumor pathologies, PCa is energetically more dependent on lipid β-oxidation than on glucose oxidation, thus exhibiting a unique profile far from the "Warburg" glycolytic phenotype compared with that found in the majority of the tumors [192]. Concordantly, PCa cells show a high rate of de novo synthesis of fatty acids (partially driven by AR activation), and active lipid uptake, which make even lipid accumulation visible intracellularly [193]. In fact, various key lipidic enzymes (i.e., ACLY, ACC, and FASN) are markedly overexpressed in PCa [194]. Although this peculiarity could be understood as a point to exploit, a recent review article concludes that lipid-targeted drugs are unlikely to replace current highly effective treatments for metastatic PCa but can be used in combination to improve response rates and longevity of cancer control [195].
Despite this, the causal relationship between obesity—whose origin lies in adipose tissue which could be a direct source of metabolic fuel—and PCa is still a matter of debate. A study affirms that the higher BMI, the lower risk of PCa diagnosis, but the risk of PCa mortality increases in such cases [196]. In consonance, another work suggest that the median diagnostic age decreases with increasing BMI, and that overweight and obese patients are more likely to have and advanced or metastatic PCa at diagnosis [197]. Parallelly, they also demonstrated that PSA levels tend to become lower in the higher BMI groups, a possible explanation for the late diagnosis [197]. In this regard, it has also been proved that a further increase in the BMI (above 28) is inversely associated with the likelihood of surgical treatment compared with radiotherapy, although no evidence of worse functional outcomes was found in obese patients undergoing definitive treatment nor remained on active surveillance [198]. However, through genetic profiling-based approaches, it has been also concluded that adiposity is unlikely to influence PCa via metabolic factors [199]. In vitro, a recent study demonstrated that conditioned media derived from the 3T3-L1 adipocyte line was able to elicit profound phenotypic changes in DU145 and PC-3 models, i.e., elevated stemness markers, increased EMT, clonogenic, prostatosphere formation, and invasiveness capacity, as well as showing evidence of chemoresistance to docetaxel and cabazitaxel [200].
In this scenario, during the last years, a specific fat tissue depot, the periprostatic adipose tissue (PPAT) has attracted attention in this research field. PPAT directly surrounds the prostate gland and could act as an energy reservoir for growing tumors, as well as a local source of factors [201]. Remarkably, a significant positive correlation has been found between BMI and PPAT thickness [202]. Thus, this tissue could represent a crucial player in the connection between systemic low-chronic inflammation associated with obesity and localized inflammation of the prostate, which could contribute to PCa development [203]. A detailed transcriptomic PPAT profiling study suggests a hypoxic state that courses with HIF2-α overexpression [204]. Concordantly, various inflammatory-related pathways are also overrepresented, finally leading to a drastically altered PPAT extracellular matrix remodeling, deposition, and composition [204].
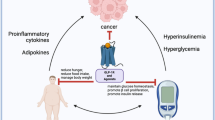
Another example is concerning IL-6, which is associated with the most aggressive phenotypes of PCa and may be involved in metastatic process. In addition, IL-6 is involved in CRPC development through accessory activation of AR [140]. It is also known that M1 macrophages aggregate around necrotic adipocytes in inflamed tissue, forming ‘crown-like structures’ and producing IL-6 [205]. High IL-6 levels have been found in PPAT and may be involved in a malignant environment that favors the development of more aggressive PCa [206]. Therefore, for all the information stated above, it is possible that cytokine signaling between the PPAT, and the prostate may be controlled, at least in part, by inflammasomes activation (Fig. 3).

Interaction between periprostatic adipose tissue, prostate gland, and inflammasomes. Different types of functional inflammasomes have been detected in multiple cell types in periprostatic adipose tissue (PPAT) and the prostate gland. Crosstalk between both tissues could be mediated reciprocally by surrounding proinflammatory molecules produced by inflammasomes, and miRNAs included in adipose tissue derived exosomes
4.1 The influence of miRNAs in adipose tissue, prostate, and inflammatory response
In the context of AT and prostate, and how the metabolic status could influence the potential tumor malignancy through inflammasome-related mechanisms, the exchange of information via exosomes and their cargo, may be also of great importance (Fig. 3). In fact, it has been shown that the number and the specific content of exosomes released from AT significantly change in patients with obesity [207]. Specifically, it has been demonstrated that different miRNAs may play a key role in this scenario. For example, leptin levels have been shown to influence PCa through the downregulation of miRNA-628, which is able to inhibit proliferative, migratory, invasive, and spheroid-forming capacity in vitro [208]. A comprehensive revision that compiled the miRNAs controlled by three of the main adipokines (i.e., adiponectin, leptin, and resistin) that influence certain tumor diseases has been published [209]. Notably, it has been reported that miR-7a-2 and miR-1977 are two of the critical transcriptomic factors (many of them related to the immune and inflammatory response) that contribute to a specific and discriminating PPAT expression signature between obese and overweight patients with PCa [201].
Simultaneously, those secreted factors (adipokines and/or miRNAs) released by AT, whose levels are influenced by metabolic status, could also affect the endogenous levels of miRNAs produced and released by the prostate, constituting an additional complexity layer between AT, obesity, and PCa [209, 210]. In this context, the uptake of tumor-secreted exosomes by stromal cells induces a reprogramming that contributes to the formation of new tumor cells [211]. Some examples include: miR-205, which has been reported to be regulated by the activation of the NLRP1-inflammasome locus snpRNAs and the subsequent control of 8q24-locus expression, establishing a complex epigenetic regulatory network, which ultimately modulate the expression of a footprint of genes that includes the tumor suppressor PTEN [212]; miR-17/92, whose levels are modulated by testosterone and 1,25-dihydroxyvitamin D3, and affect PPARα transcript stability promoting lipid synthesis and energy storage in PCa cells [213]; miR-21, which is significantly associated with PCa recurrence after radical prostatectomy in BMI-dependent manner [214]; or miR-139-5p, which is directly regulated by the fat mass and obesity-associated (FTO) protein, and exerts tumors modulating functions through ZNF217 [215].
Besides acting as potential messengers, miRNAs have also become a potent diagnostic and prognostic tool in PCa, becoming a highly exploited source of biomarkers in recent years. For example, it has been proved that plasma levels of let-7c, let-7e, let-7i, miR-26a-5p, miR-26b-5p, and, especially, miR-18b-5p and miR-25-3p showed a clear discriminatory capacity between PCa with elevated PSA levels among patients with BPH [216]. In urine, the combined measurement of miR-21, miR-19a, and miR19b also showed a higher prediction value that overcomes the PSA-based detection method [216]. Recently, the analysis of the whole plasma miRNome from PCa patients pointed out miR-107 as the best discriminatory miRNA between PCa and control patients, showing even better discrimination capacity when only patients with obesity were considered [33]; likewise, miR-4454 levels are highly altered and associated with metabolic disorders such as obesity and insulin metabolism in PCa [217].
Considering that functional inflammasomes have been found in visceral fat tissue and prostate epithelial cells, and that a wide variety of immune infiltrate cells are present in both types of tissues, inflammasome activation could represent a chicken-and-egg situation, enabling to establish a bidirectional crosstalk and self-amplifying signaling driving PCa development in patients with obesity.
5 Future perspectives
The compiled evidences suggest that inflammasome may be considered a key target in inflammation-related diseases. In fact, several initiatives focused on the blockade of inflammasome have been recently implemented in diverse areas. Thus, although the possibilities are limited and more selective drugs are still under development, various treatments have been already identified to target inflammasomes components [218].
Among them, the compound termed MCC950, a daily sulfonylurea-containing molecule, is considered the most potent and selective NLRP3 inflammasome inhibitor [219]. MCC950 treatment improved hematopoiesis in a myeloid leukemia mouse model and human-derived samples from myelodysplastic syndrome patients [220, 221], and to reduce tumor burden in squamous cell head and neck carcinoma mouse model [222], though research in this field is still limited.
A special mention of caspase-1 inhibitors should be made. Molecules such as VX-765 (an oral pro-drug) have been demonstrated to notably diminish the release of IL-1β and IL-18, yet its clinical use is still under investigation [223]. Another group of strategies comprises those based on exploring the modulation of the up and down-stream pathways of the inflammasome, by targeting priming and activating signals or by directly blocking the downstream IL-1β pathway. We have summarized in Table 2 the current information regarding inhibitors that target structural NLRP3 inflammasome components.
Remarkably, some clinical trials have been performed concerning inflammasomes and obesity. The NCT05071391 trial aimed to determine the relationship between autophagy and inflammasomes regulation in the pathogenesis of obesity and related comorbidities; however, no published results are available yet. On the other hand, the NCT04315376 trial evaluated the effect of a hypocaloric diet on inflammasome adaptor ASC (22), alone or in combination with physical exercise-based intervention (15) in two different groups of patients with obesity showing its potential as a follow-up biomarker in such types of interventions [224]. Additionally, three other inflammasome-related clinical trials are in the recruiting phase. The clinical trial NCT04814147 intends to study the inflammasomes after sleeve gastrectomy intervention in women with obesity; the trial NCT04275674 aims to study the macrophages inflammasome in a cohort composed of 200 women with obesity and the likely connection with miscarriages; and finally, the global aim of NCT04485871 is to explore the pivotal role of NLRP3 inflammasome in T2D-associated inflammation through an ω-3 dietetical supplementation.
However, to the best of our knowledge, no studies have been made to explore the modulation of inflammasomes activities in the context of cancer and especially PCa. Although the therapeutic possibilities of inflammasome cascade inhibition are promising, the available results should be taken carefully, as blocking central inflammatory mediators could be a double-edged sword. In this regard, a thoroughly investigation about possible risks/benefits should be accomplished before this type of therapies could be used in the future.
6 Conclusions
Inflammasomes are macromolecular structures able to regulate the inflammatory response through the production of pro-inflammatory molecules. Although inflammasomes were initially associated with immune cell populations, recent studies have demonstrated that functional inflammasomes could be assembled under certain conditions (e.g., hypoxia, lesions, or infections) in human prostate cells. However, in the case of prostate cancer, the available information is limited and only a few types of inflammasomes have been explored. On the other hand, evidence connecting the role of inflammasomes and weight-related disorders, such as obesity, is more solid due to the marked inflammatory nature of these conditions, being also active in adipocytes and in the immune cells infiltrating the adipose tissue. In both pathological states, current information points that IL-1β and NF-kB are central players orchestrating an inflammatory response, although the investigation in this regard is in the initial steps and much more effort is needed to unveil the connection between prostate cancer, obesity, and inflammation. Therefore, understanding the complex role that inflammasomes could be playing, and considering the availability of drugs that can specifically modulate them, may represent a novel and valuable source of therapeutic tools and strategies of action.
Abbreviations
- AT:
-
Adipose Tissue
- ATMs:
-
Adipose Tissue Macrophages
- AR:
-
Androgen Receptor
- BPH:
-
Benign Prostate Hyperplasia
- BMI:
-
Body Mass Index
- CARD:
-
Caspase Activation and Recruitment Domain
- CRPC:
-
Castration Resistant Prostate Cancer
- CLRs:
-
C-type lectin receptors
- DAMPs:
-
Damage-Associated Molecular Patterns
- EMT:
-
Epithelial to Mesenchymal Transition
- HIF-1α:
-
Hypoxia-inducible factor 1-alpha
- IL-18:
-
Interleukin-18
- IL18BP:
-
Interleukin-18 Binding Protein
- IL-1β:
-
Interleukin-1β
- NLRs:
-
Nucleotide-binding Oligomerization Domain-like receptors
- PYD:
-
PAAD/DAPIN Domain
- PAMPs:
-
Pathogen-Associated Molecular Patterns
- PRRs:
-
Pattern Recognition Receptors
- PPAT:
-
Periprostatic Adipose Tissue
- PIA:
-
Proliferative Inflammatory Atrophy
- PCa:
-
Prostate Cancer
- PIN:
-
Prostatic Intraepithelial Neoplasia
- ROS:
-
Reactive Oxygen Species
- RLRs:
-
Retinoic acid-Inducible Gene 1
- SNPs:
-
Single Nucleotide Polymorphisms
- SVF:
-
Stromal-Vascular Fraction
- TLRs:
-
Toll-like receptors
- T2D:
-
Type 2 Diabetes
References
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–49. https://doi.org/10.3322/caac.21660.
Eeles RA, Dearnaley DP, ArdernJones A, Shearer RJ, Easton DF, Ford D, Edwards S, Dowe A, Anderson J, Archibold J, Bailey M, Barker C, Bellringer J, Bishop DT, Bishop M, Bolger J, Boyd J, Budd D, Butler M, Brookstein R, Charig C, Conn I, Cranston C, Crundwell M, Das G, Doble A, Duncan W, Duchesne J, Eccles D, Fawcett D, Fisher C, Fletcher M, Fowler JW, Gallegos C, Ghaznavi A, Gilbert H, Glaholm J, Gordon E, Hampson S, Hanbury DC, Hargreave T, Harland S, Harrison GS, Harrison NW, Hart JL, Hehir M, Hendry W, Higgins A, Hopper J, Horwich A, Hughes M, Irving S, James N, Jenkins IL, Jones C, Key T, Kaisary A, Kirby R, Kirk D, Lee J, Lemberger R, Lloyd S, Lynch M, Malone PR, Mansi J, Mason M, McEwan AB, McNicholas TA, Moffat LEF, Morgan RJ, Murday V, Muir G, Munson KW, Murray K, Murrell S, Newman H, Oboyle PJ, Odonoghue E, Notley RG, Pengelly A, Philip T, Plail R, Powell C, Russell J, Read G, Reddy PJ, Richmond W, Roberts T, Rowley K, Rouse AD, Russell RM, Ryan P, Senanayake L, Sandhu D, Shridhar P, Shweitzer R, Shearer R, Smith J, Smith P, Stockdale A, Stower M, Thomas P, Terry T, Thurston A, Tullock D, Turner G, Wallace M, Weston P, Whelan P, Whillis D, Wilson R, Williams G, Woodhouse C. Familial prostate cancer: The evidence and the Cancer Research Campaign British Prostate Group (CRC/BPG) UK Familial Prostate Cancer Study. Br J Urol. 1997;79:8–14. https://doi.org/10.1111/j.1464-410X.1997.tb00795.x.
De Nunzio C, Kramer G, Marberger M, Montironi R, Nelson W, Schroder F, Sciarra A, Tubaro A. The controversial relationship between benign prostatic hyperplasia and prostate cancer: The role of inflammation. Eur Urol. 2011;60(1):106–17. https://doi.org/10.1016/j.eururo.2011.03.055.
Allott EH, Masko EM, Freedland SJ. Obesity and prostate cancer: Weighing the evidence. Eur Urol. 2013;63(5):800–9. https://doi.org/10.1016/j.eururo.2012.11.013.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74. https://doi.org/10.1016/j.cell.2011.02.013.
Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature. 2008;454(7203):436–44. https://doi.org/10.1038/nature07205.
Coussens LM, Werb Z. Inflammation and cancer. Nature. 2002;420(6917):860–7. https://doi.org/10.1038/nature01322.
Irani J, Goujon JM, Ragni E, Peyrat L, Hubert J, Saint F, Mottet N, Pathologist G. Multi Center Study, High-grade inflammation in prostate cancer as a prognostic factor for biochemical recurrence after radical prostatectomy. Urology. 1999;54(3):467–72. https://doi.org/10.1016/s0090-4295(99)00152-1.
Di Silverio F, Gentile V, De Matteis A, Mariotti G, Giuseppe V, Luigi PA, Sciarra A. Distribution of inflammation, pre-malignant lesions, incidental carcinoma in histologically confirmed benign prostatic hyperplasia: A retrospective analysis. Eur Urol. 2003;43(2):164–75. https://doi.org/10.1016/s0302-2838(02)00548-1.
Nickel JC, Roehrborn CG, O’Leary MP, Bostwick DG, Somerville MC, Rittmaster RS. Examination of the relationship between symptoms of prostatitis and histological inflammation: Baseline data from the REDUCE chemoprevention trial. J Urol. 2007;178(3):896–900. https://doi.org/10.1016/j.juro.2007.05.041.
Nickel JC, Roehrborn CG, O’Leary MP, Bostwick DG, Somerville MC, Rittmaster RS. The relationship between prostate inflammation and lower urinary tract symptoms: Examination of baseline data from the REDUCE trial. Eur Urol. 2008;54(6):1379–84. https://doi.org/10.1016/j.eururo.2007.11.026.
Murtola TJ, Gurel B, Umbehr M, Lucia MS, Thompson IM, Goodman PJ, Kristal AR, Parnes HL, Lippman SM, Sutcliffe S, Peskoe SB, Barber JR, Drake CG, Nelson WG, De Marzo AM, Platz EA. Inflammation in benign prostate tissue and prostate cancer in the finasteride arm of the prostate cancer prevention trial. Cancer Epidemiol Biomark Prev. 2016;25(3):463–9. https://doi.org/10.1158/1055-9965.epi-15-0987.
Gurel B, Lucia MS, Thompson IM Jr, Goodman PJ, Tangen CM, Kristal AR, Parnes HL, Hoque A, Lippman SM, Sutcliffe S, Peskoe SB, Drake CG, Nelson WG, De Marzo AM, Platz EA. Chronic inflammation in benign prostate tissue is associated with high-grade prostate cancer in the placebo arm of the prostate cancer prevention trial. Cancer Epidemiol Biomark Prev. 2014;23(5):847–56. https://doi.org/10.1158/1055-9965.epi-13-1126.
Kwon EM, Salinas CA, Kolb S, Fu R, Feng Z, Stanford JL, Ostrander EA. Genetic polymorphisms in inflammation pathway genes and prostate cancer risk. Cancer Epidemiol Biomarkers Prev. 2011;20(5):923–33. https://doi.org/10.1158/1055-9965.epi-10-0994.
Davidsson S, Fiorentino M, Andren O, Fang F, Mucci LA, Varenhorst E, Fall K, Rider JR. Inflammation, focal atrophic lesions, and prostatic intraepithelial neoplasia with respect to risk of lethal prostate cancer. Cancer Epidemiol Biomark Prev. 2011;20(10):2280–7. https://doi.org/10.1158/1055-9965.epi-11-0373.
Moreira DM, Nickel JC, Gerber L, Muller RL, Andriole GL, Castro-Santamaria R, Freedland SJ. Baseline prostate inflammation is associated with a reduced risk of prostate cancer in men undergoing repeat prostate biopsy. Cancer. 2014;120(2):190–6. https://doi.org/10.1002/cncr.28349.
Pepe P, Aragona F. Does an inflammatory pattern at primary biopsy suggest a lower risk for prostate cancer at repeated saturation prostate biopsy? Urol Int. 2011;87(2):171–4. https://doi.org/10.1159/000328043.
Yli-Hemminki TH, Laurila M, Auvinen A, Maattanen L, Huhtala H, Tammela TLJ, Kujala PM. Histological inflammation and risk of subsequent prostate cancer among men with initially elevated serum prostate-specific antigen (PSA) concentration in the Finnish prostate cancer screening trial. BJU Int. 2013;112(6):735–41. https://doi.org/10.1111/bju.12153.
Langston ME, Horn M, Khan S, Pakpahan R, Doering M, Dennis LK, Sutcliffe S. A systematic review and meta-analysis of associations between clinical prostatitis and prostate cancer: New estimates accounting for detection bias. Cancer Epidemiol Biomark Prev. 2019;28(10):1594–603. https://doi.org/10.1158/1055-9965.epi-19-0387.
Vasavada SR, Dobbs RW, Kajdacsy-Balla AA, Abern MR, Moreira DM. Inflammation on prostate needle biopsy is associated with lower prostate cancer risk: A meta-analysis. J Urol. 2018;199(5):1174–81. https://doi.org/10.1016/j.juro.2017.11.120.
De Marzo AM, Platz EA, Sutcliffe S, Xu JF, Gronberg H, Drake CG, Nakai Y, Isaacs WB, Nelson WG. Inflammation in prostate carcinogenesis. Nat Rev Cancer. 2007;7(4):256–69. https://doi.org/10.1038/nrc2090.
Solinas C, Chanzá NM, Awada A, Scartozzi M. The immune infiltrate in prostate, bladder and testicular tumors: An old friend for new challenges. Cancer Treat Rev. 2017;53:138–45. https://doi.org/10.1016/j.ctrv.2016.12.004.
Sfanos KS, De Marzo AM. Prostate cancer and inflammation: the evidence. Histopathology. 2012;60(1):199–215. https://doi.org/10.1111/j.1365-2559.2011.04033.x.
Patel AR, Klein EA. Risk factors for prostate cancer. Nat Clin Pract Urol. 2009;6(2):87–95. https://doi.org/10.1038/ncpuro1290.
Shrestha E, White JR, Yu SH, Kulac I, Ertunc O, De Marzo AM, Yegnasubramanian S, Mangold LA, Partin AW, Sfanos KS. Profiling the urinary microbiome in men with positive versus negative biopsies for prostate cancer. J Urol. 2018;199(1):161–71. https://doi.org/10.1016/j.juro.2017.08.001.
Karan D, Dubey S. From inflammation to prostate cancer: The role of inflammasomes. Adv Urol. 2016;2016:3140372. https://doi.org/10.1155/2016/3140372.
Brüggemann H, Al-Zeer MA. Bacterial signatures and their inflammatory potentials associated with prostate cancer. APMIS. 2020;128(2):80–91. https://doi.org/10.1111/apm.13021.
Oseni SO, Naar C, Pavlović M, Asghar W, Hartmann JX, Fields GB, Esiobu N, Kumi-Diaka J. The molecular basis and clinical consequences of chronic inflammation in prostatic diseases: prostatitis, benign prostatic hyperplasia, and prostate cancer. Cancers (Basel). 2023;15(12). https://doi.org/10.3390/cancers15123110.
Fujita K, Hayashi T, Matsushita M, Uemura M, Nonomura N. Obesity, inflammation, and prostate cancer. J Clin Med. 2019;8(2). https://doi.org/10.3390/jcm8020201.
Gilbert CA, Slingerland JM. Cytokines, obesity, and cancer: new insights on mechanisms linking obesity to cancer risk and progression. Ann Rev Med. 2013;64:45–47. https://doi.org/10.1146/annurev-med-121211-091527.
Fuster JJ, Ouchi N, Gokce N, Walsh K. Obesity-induced changes in adipose tissue microenvironment and their impact on cardiovascular disease. Circ Res. 2016;118(11):1786–807. https://doi.org/10.1161/circresaha.115.306885.
Hsing AW, Sakoda LC, Chua SC. Obesity, metabolic syndrome, and prostate cancer. Am J Clin Nutr. 2007;86(3):843S–857S. https://doi.org/10.1093/ajcn/86.3.843S.
Herrero-Aguayo V, Sáez-Martínez P, Jiménez-Vacas JM, Moreno-Montilla MT, Montero-Hidalgo AJ, Pérez-Gómez JM, López-Canovas JL, Porcel-Pastrana F, Carrasco-Valiente J, Anglada FJ, Gómez-Gómez E, Yubero-Serrano EM, Ibañez-Costa A, Herrera-Martínez AD, Sarmento-Cabral A, Gahete MD, Luque RM. Dysregulation of the miRNome unveils a crosstalk between obesity and prostate cancer: miR-107 asa personalized diagnostic and therapeutic tool. Mol Ther Nucleic Acids. 2022;27:1164–78. https://doi.org/10.1016/j.omtn.2022.02.010.
Jiménez-Vacas JM, Montero-Hidalgo AJ, Gómez-Gómez E, Fuentes-Fayos AC, Ruiz-Pino F, Guler I, Camargo A, Anglada FJ, Carrasco-Valiente J, Tena-Sempere M, Sarmento-Cabral A, Castaño JP, Gahete MD, Luque RM. In1-ghrelin splicing variant as a key element in the pathophysiological association between obesity and prostate cancer. J Clin Endocrinol Metab. 2021;106(12):e4956–68. https://doi.org/10.1210/clinem/dgab516.
Gómez-Gómez E, Carrasco-Valiente J, Campos-Hernández JP, Blanca-Pedregosa AM, Jiménez-Vacas JM, Ruiz-García J, Valero-Rosa J, Luque RM, Requena-Tapia MJ. Clinical association of metabolic syndrome, C-reactive protein and testosterone levels with clinically significant prostate cancer. J Cell Mol Med. 2019;23(2):934–42. https://doi.org/10.1111/jcmm.13994.
L‐López F, Sarmento-Cabral A, Herrero-Aguayo V, Gahete MD, Castaño JP, Luque RM. Obesity and metabolic dysfunction severely influence prostate cell function: role of insulin and IGF1. J Cell Mol Med. 2017;21(9):1893–904. https://doi.org/10.1111/jcmm.13109.
Martinon F, Burns K, Tschopp J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell. 2002;10(2):417–26. https://doi.org/10.1016/s1097-2765(02)00599-3.
Man SM, Karki R, Kanneganti TD. AIM2 inflammasome in infection, cancer, and autoimmunity: Role in DNA sensing, inflammation, and innate immunity. Eur J Immunol. 2016;46(2):269–80. https://doi.org/10.1002/eji.201545839.
Moossavi M, Parsamanesh N, Bahrami A, Atkin SL, Sahebkar A. Role of the NLRP3 inflammasome in cancer. Mol Cancer. 2018;17. https://doi.org/10.1186/s12943-018-0900-3.
Terlizzi M, Casolaro V, Pinto A, Sorrentino R. Inflammasome: Cancer’s friend or foe? Pharmacol Ther. 2014;143(1):24–33. https://doi.org/10.1016/j.pharmthera.2014.02.002.
Pelka K, De Nardo D. Emerging concepts in innate immunity. Methods Mol Biol. 2018;1714:1–18. https://doi.org/10.1007/978-1-4939-7519-8_1.
Rider P, Carmi Y, Guttman O, Braiman A, Cohen I, Voronov E, White MR, Dinarello CA, Apte RN. IL-1α and IL-1β recruit different myeloid cells and promote different stages of sterile inflammation. J Immunol. 2011;187(9):4835–43. https://doi.org/10.4049/jimmunol.1102048.
Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–20. https://doi.org/10.1016/j.cell.2010.01.022.
Iwasaki A, Medzhitov R. Toll-like receptor control of the adaptive immune responses. Nat Immunol. 2004;5(10):987–95. https://doi.org/10.1038/ni1112.
Medzhitov R. Recognition of microorganisms and activation of the immune response. Nature. 2007;449(7164):819–26. https://doi.org/10.1038/nature06246.
Chen G, Shaw MH, Kim YG, Nunez G. NOD-Like receptors: Role in innate immunity and inflammatory disease. Ann Rev Pathol-Mech Dis. 2009;4:365–98. https://doi.org/10.1146/annurev.pathol.4.110807.092239.
Sahoo M, Ceballos-Olvera I, del Barrio L, Re F. Role of the inflammasome, IL-1 beta, and IL-18 in bacterial infections. Sci World J. 2011;11:2037–50. https://doi.org/10.1100/2011/212680.
Martinon F, Mayor A, Tschopp J. The inflammasomes: Guardians of the body. Annu Rev Immunol. 2009;27:229–65. https://doi.org/10.1146/annurev.immunol.021908.132715.
Deswaerte V, Nguyen P, Jenkins B, Putoczki T. The inflammasome adaptor ASC suppresses tumor cell apoptosis, independent of inflammation, via IL18 in gastric cancer. Cytokine. 2017;100:93–93. https://doi.org/10.1158/0008-5472.CAN-17-1887.
Kayagaki N, Stowe IB, Lee BL, O’Rourke K, Anderson K, Warming S, Cuellar T, Haley B, Roose-Girma M, Phung QT, Liu PS, Lill JR, Li H, Wu JS, Kummerfeld S, Zhang J, Lee WP, Snipas SJ, Salvesen GS, Morris LX, Fitzgerald L, Zhang YF, Bertram EM, Goodnow CC, Dixit VM. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature. 2015;526(7575):666–71. https://doi.org/10.1038/nature15541.
Schroder K, Tschopp J. The inflammasomes. Cell. 2010;140(6):821–32. https://doi.org/10.1016/j.cell.2010.01.040.
Janowski AM, Sutterwala FS. Atypical Inflammasomes. NLR Proteins: Methods and Protocols, F. Di Virgilio and P. Pelegrín, Editors. Springer New York: New York, NY. 2016;45–62. https://doi.org/10.1007/978-1-4939-3566-6_2.
Khare S, Dorfleutner A, Bryan NB, Yun C, Radian AD, de Almeida L, Rojanasakul Y, Stehlik C. An NLRP7-containing inflammasome mediates recognition of microbial lipopeptides in human macrophages. Immunity. 2012;36(3):464–76. https://doi.org/10.1016/j.immuni.2012.02.001.
Minkiewicz J, Vaccari JPD, Keane RW. Human astrocytes express a novel NLRP2 inflammasome. Glia. 2013;61(7):1113–21. https://doi.org/10.1002/glia.22499.
Elinav E, Strowig T, Kau AL, Henao-Mejia J, Thaiss CA, Booth CJ, Peaper DR, Bertin J, Eisenbarth SC, Gordon JI, Flavell RA. NLRP6 inflammasome regulates colonic microbial ecology and risk for colitis. Cell. 2011;145(5):745–57. https://doi.org/10.1016/j.cell.2011.04.022.
Vladimer GI, Weng D, Paquette SWM, Vanaja SK, Rathinam VAK, Aune MH, Conlon JE, Burbage JJ, Proulx MK, Liu Q, Reed G, Mecsas JC, Iwakura Y, Bertin J, Goguen JD, Fitzgerald KA, Lien E. The NLRP12 inflammasome recognizes Yersinia pestis. Immunity. 2012;37(1):96–107. https://doi.org/10.1016/j.immuni.2012.07.006.
Kerur N, Veettil MV, Sharma-Walia N, Bottero V, Sadagopan S, Otageri P, Chandran B. IFI16 acts as a nuclear pathogen sensor to induce the inflammasome in response to kaposi sarcoma-associated herpesvirus infection. Cell Host Microbe. 2011;9(5):363–75. https://doi.org/10.1016/j.chom.2011.04.008.
Shi H, Wang Y, Li X, Zhan X, Tang M, Fina M, Su L, Pratt D, Bu CH, Hildebrand S, Lyon S, Scott L, Quan J, Sun Q, Russell J, Arnett S, Jurek P, Chen D, Kravchenko VV, Mathison JC, Moresco EM, Monson NL, Ulevitch RJ, Beutler B. NLRP3 activation and mitosis are mutually exclusive events coordinated by NEK7, a new inflammasome component. Nat Immunol. 2016;17(3):250–8. https://doi.org/10.1038/ni.3333.
Sagulenko V, Thygesen SJ, Sester DP, Idris A, Cridland JA, Vajjhala PR, Roberts TL, Schroder K, Vince JE, Hill JM, Silke J, Stacey KJ. AIM2 and NLRP3 inflammasomes activate both apoptotic and pyroptotic death pathways via ASC. Cell Death Differ. 2013;20(9):1149–60. https://doi.org/10.1038/cdd.2013.37.
Perrichet A, Ghiringhelli F, Rébé C. Understanding inflammasomes and PD-1/PD-L1 crosstalk to improve cancer treatment efficiency. Cancers (Basel). 2020;12(12). https://doi.org/10.3390/cancers12123550.
Bauernfeind F, Bartok E, Rieger A, Franchi L, Nunez G, Hornung V. Cutting edge: Reactive oxygen species inhibitors block priming, but not activation, of the NLRP3 inflammasome. J Immunol. 2011;187(2):613–7. https://doi.org/10.4049/jimmunol.1100613.
Bauernfeind FG, Horvath G, Stutz A, Alnemri ES, MacDonald K, Speert D, Fernandes-Alnemri T, Wu JH, Monks BG, Fitzgerald KA, Hornung V, Latz E. Cutting edge: NF-kappa B activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol. 2009;183(2):787–91. https://doi.org/10.4049/jimmunol.0901363.
He Y, Hara H, Núñez G. Mechanism and regulation of NLRP3 inflammasome activation. Trends Biochem Sci. 2016;41(12):1012–21. https://doi.org/10.1016/j.tibs.2016.09.002.
Lamkanfi M, Dixit VM. Mechanisms and functions of inflammasomes. Cell. 2014;157(5):1013–22. https://doi.org/10.1016/j.cell.2014.04.007.
Vanaja SK, Rathinam VAK, Fitzgerald KA. Mechanisms of inflammasome activation: recent advances and novel insights. Trends Cell Biol. 2015;25(5):308–15. https://doi.org/10.1016/j.tcb.2014.12.009.
Place DE, Kanneganti TD. Recent advances in inflammasome biology. Curr Opin Immunol. 2018;50:32–8. https://doi.org/10.1016/j.coi.2017.10.011.
Herrera-Martínez AD, Herrero-Aguayo V, Pérez-Gómez JM, Gahete MD, Luque RM. Inflammasomes: Cause or consequence of obesity-associated comorbidities in humans. Obesity (Silver Spring). 2022;30(12):2351–62. https://doi.org/10.1002/oby.23581.
Herrero-Aguayo V, Sáez-Martínez P, López-Cánovas JL, Prados-Carmona JJ, Alcántara-Laguna MD, López FL, Molina-Puerta MJ, Calañas-Continente A, Membrives A, Castilla J, Ruiz-Ravelo J, Alonso-Echague R, Yubero-Serrano EM, Castaño JP, Gahete MD, Gálvez-Moreno MA, Luque RM, Herrera-Martínez AD. Dysregulation of components of the inflammasome machinery after bariatric surgery: Novel targets for a chronic disease. J Clin Endocrinol Metab. 2021;106(12):e4917–34. https://doi.org/10.1210/clinem/dgab586.
Karki R, Man SM, Kanneganti TD. Inflammasomes and cancer. Cancer Immunol Res. 2017;5(2):94–9. https://doi.org/10.1158/2326-6066.cir-16-0269.
Karan D. Inflammasomes: Emerging central players in cancer immunology and immunotherapy. Front Immunol. 2018;9:3028. https://doi.org/10.3389/fimmu.2018.03028.
Ponomareva L, Liu H, Duan X, Dickerson E, Shen H, Panchanathan R, Choubey D. AIM2, an IFN-inducible cytosolic DNA sensor, in the development of benign prostate hyperplasia and prostate cancer. Mol Cancer Res. 2013;11(10):1193–202. https://doi.org/10.1158/1541-7786.mcr-13-0145.
Vinaik R, Barayan D, Abdullahi A, Jeschke MG. NLRP3 inflammasome mediates white adipose tissue browning after burn. Am J Physiol-Endocrinol Metab. 2019;317(5):E751–9. https://doi.org/10.1152/ajpendo.00180.2019.
Galli R, Starace D, Busà R, Angelini DF, Paone A, De Cesaris P, Filippini A, Sette C, Battistini L, Ziparo E, Riccioli A. TLR stimulation of prostate tumor cells induces chemokine-mediated recruitment of specific immune cell types. J Immunol. 2010;184(12):6658–69. https://doi.org/10.4049/jimmunol.0902401.
Paone A, Galli R, Gabellini C, Lukashev D, Starace D, Gorlach A, De Cesaris P, Ziparo E, Del Bufalo D, Sitkovsky MV, Filippini A, Riccioli A. Toll-like receptor 3 regulates angiogenesis and apoptosis in prostate cancer cell lines through hypoxia-inducible factor 1 alpha. Neoplasia. 2010;12(7):539–49. https://doi.org/10.1593/neo.92106.
González-Reyes S, Fernández JM, González LO, Aguirre A, Suárez A, González JM, Escaff S, Vizoso FJ. Study of TLR3, TLR4, and TLR9 in prostate carcinomas and their association with biochemical recurrence. Cancer Immunol Immunother. 2011;60(2):217–26. https://doi.org/10.1007/s00262-010-0931-0.
Chen YC, Giovannucci E, Lazarus R, Kraft P, Ketkar S, Hunter DJ. Sequence variants of toll-like receptor 4 and susceptibility to prostate cancer. Can Res. 2005;65(24):11771–8. https://doi.org/10.1158/0008-5472.can-05-2078.
Cheng I, Plummer SJ, Casey G, Witte JS. Toll-like receptor 4 genetic variation and advanced prostate cancer risk. Cancer Epidemiol Biomark Prev. 2007;16(2):352–5. https://doi.org/10.1158/1055-9965.epi-06-0429.
Kim HJ, Bae JS, Chang IH, Kim KD, Lee J, Shin HD, Lee JY, Kim WJ, Kim W, Myung SC. Sequence variants of toll-like receptor 4 (TLR4) and the risk of prostate cancer in Korean men. World J Urol. 2012;30(2):225–32. https://doi.org/10.1007/s00345-011-0690-3.
Weng PH, Huang YL, Page JH, Chen JH, Xu J, Koutros S, Berndt S, Chanock S, Yeager M, Witte JS, Eeles RA, Easton DF, Neal DE, Donovan J, Hamdy FC, Muir KR, Giles G, Severi G, Smith JR, Balistreri CR, Shui IM, Chen YC. Polymorphisms of an innate immune gene, toll-like receptor 4, and aggressive prostate cancer risk: A systematic review and meta-analysis. PLoS ONE. 2014;9(10):12. https://doi.org/10.1371/journal.pone.0110569.
Shui IM, Stark JR, Penney KL, Schumacher FR, Epstein MM, Pitt MJ, Stampfer MJ, Tamimi RM, Lindstrom S, Sesso HD, Fall K, Ma J, Kraft P, Giovannucci E, Mucci LA. Genetic variation in the toll-like receptor 4 and prostate cancer incidence and mortality. Prostate. 2012;72(2):209–16. https://doi.org/10.1002/pros.21423.
König JE, Senge T, Allhoff EP, König W. Analysis of the inflammatory network in benign prostate hyperplasia and prostate cancer. Prostate. 2004;58(2):121–9. https://doi.org/10.1002/pros.10317.
Gatti G, Quintar AA, Andreani V, Nicola JP, Maldonado CA, Masini-Reviso AM, Rivero VE, Maccioni M. Expression of toll-like receptor 4 in the prostate gland and its association with the severity of prostate cancer. Prostate. 2009;69(13):1387–97. https://doi.org/10.1002/pros.20984.
Riddell JR, Bshara W, Moser MT, Spernyak JA, Foster BA, Gollnick SO. Peroxiredoxin 1 controls prostate cancer growth through toll-like receptor 4-dependent regulation of tumor vasculature. Can Res. 2011;71(5):1637–46. https://doi.org/10.1158/0008-5472.can-10-3674.
Zhang YT, Wang Y, Yuan JL, Qin WJ, Liu F, Wang FL, Zhang G, Yang XJ. Toll-like receptor 4 ligation confers chemoresistance to docetaxel on PC-3 human prostate cancer cells. Cell Biol Toxicol. 2012;28(4):269–77. https://doi.org/10.1007/s10565-012-9221-2.
Lv Z, Li W, Wei X. S100A9 promotes prostate cancer cell invasion by activating TLR4/NF-κB/integrin β1/FAK signaling. Onco Targets Ther. 2020;13:6443–52. https://doi.org/10.2147/ott.s192250.
Ilvesaro JM, Merrell MA, Swain TM, Davidson J, Zayzafoon M, Harris KW, Selander KS. Toll like receptor-9 agonists stimulate prostate cancer invasion in vitro. Prostate. 2007;67(7):774–81. https://doi.org/10.1002/pros.20562.
Vaisanen MR, Jukkola-Vuorinen A, Vuopala KS, Selander KS, Vaarala MH. Expression of Toll-like receptor-9 is associated with poor progression-free survival in prostate cancer. Oncol Lett. 2013;5(5):1659–63. https://doi.org/10.3892/ol.2013.1204.
Di JM, Pang J, Sun QP, Zhang Y, Fang YQ, Liu XP, Zhou JH, Ruan XX, Gao X. Toll-like receptor 9 agonists up-regulates the expression of cyclooxygenase-2 via activation of NF-kappa B in prostate cancer cells. Mol Biol Rep. 2010;37(4):1849–55. https://doi.org/10.1007/s11033-009-9620-5.
Rathinam VAK, Jiang Z, Waggoner SN, Sharma S, Cole LE, Waggoner L, Vanaja SK, Monks BG, Ganesan S, Latz E, Hornung V, Vogel SN, Szomolanyi-Tsuda E, Fitzgerald KA. The AIM2 inflammasome is essential for host defense against cytosolic bacteria and DNA viruses. Nat Immunol. 2010;11(5):395–403. https://doi.org/10.1038/ni.1864.
DeYoung KL, Ray ME, Su YA, Anzick SL, Johnstone RW, Trapani JA, Meltzer PS, Trent JM. Cloning a novel member of the human interferon-inducible gene family associated with control of tumorigenicity in a model of human melanoma. Oncogene. 1997;15(4):453–7. https://doi.org/10.1038/sj.onc.1201206.
Adamczak SE, de Rivero Vaccari JP, Dale G, Brand FJ III, Nonner D, Bullock MR, Dahl GP, Dietrich WD, Keane RW. Pyroptotic neuronal cell death mediated by the AIM2 inflammasome. J Cereb Blood Flow Metab. 2014;34(4):621–9. https://doi.org/10.1038/jcbfm.2013.236.
Man SM, Zhu Q, Zhu L, Liu Z, Karki R, Malik A, Sharma D, Li L, Malireddi RK, Gurung P, Neale G, Olsen SR, Carter RA, McGoldrick DJ, Wu G, Finkelstein D, Vogel P, Gilbertson RJ, Kanneganti TD. Critical role for the DNA sensor AIM2 in stem cell proliferation and cancer. Cell. 2015;162(1):45–58. https://doi.org/10.1016/j.cell.2015.06.001.
Qin Y, Pan L, Qin T, Ruan H, Zhang Y, Zhang Y, Li J, Yang J, Li W. Pan-cancer analysis of AIM2 inflammasomes with potential implications for immunotherapy in human cancer: A bulk omics research and single cell sequencing validation. Front Immunol. 2022;13:998266. https://doi.org/10.3389/fimmu.2022.998266.
Hasegawa S, Furukawa Y, Li M, Satoh S, Kato T, Watanabe T, Katagiri T, Tsunoda T, Yamaoka Y, Nakamura Y. Genome-wide analysis of gene expression in intestinal-type gastric cancers using a complementary DNA microarray representing 23,040 genes. Cancer Res. 2002;62(23):7012–7. PMID: 12460921.
Panchanathan R, Liu H, Choubey D. Hypoxia primes human normal prostate epithelial cells and cancer cell lines for the NLRP3 and AIM2 inflammasome activation. Oncotarget. 2016;7(19):28183–94. https://doi.org/10.18632/oncotarget.8594.
Ammirante M, Shalapour S, Kang Y, Jamieson CAM, Karin M. Tissue injury and hypoxia promote malignant progression of prostate cancer by inducing CXCL13 expression in tumor myofibroblasts. Proc Natl Acad Sci USA. 2014;111(41):14776–81. https://doi.org/10.1073/pnas.1416498111.
Bao B, Ahmad A, Kong DJ, Ali S, Azmi AS, Li YW, Banerjee S, Padhye S, Sarkar FH. Hypoxia induced aggressiveness of prostate cancer cells is linked with deregulated expression of VEGF, IL-6 and miRNAs that are attenuated by CDF. Plos One. 2012;7(8). https://doi.org/10.1371/journal.pone.0043726.
Xu Z, Wang H, Qin Z, Zhao F, Zhou L, Xu L, Jia R. NLRP3 inflammasome promoted the malignant progression of prostate cancer via the activation of caspase-1. Cell Death Discov. 2021;7(1):399. https://doi.org/10.1038/s41420-021-00766-9.
Karan D, Tawfik O, Dubey S. Expression analysis of inflammasome sensors and implication of NLRP12 inflammasome in prostate cancer. Sci Rep. 2017;7(1):4378. https://doi.org/10.1038/s41598-017-04286-4.
Liu Z, Zhu X, Xu C, Min F, Yu G, Chen C. Ulinastatin ameliorates the malignant progression of prostate cancer cells by blocking the RhoA/ROCK/NLRP3 pathway. Drug Dev Res. 2022. https://doi.org/10.1002/ddr.22010.
Zhao AN, Yang Z, Wang DD, Shi B, Zhang H, Bai Y, Yan BW, Zhang Y, Wen JK, Wang XL, Qu CB. Disturbing NLRP3 acetylation and inflammasome assembly inhibits androgen receptor-promoted inflammatory responses and prostate cancer progression. Faseb j. 2022;36(11):e22602. https://doi.org/10.1096/fj.202200673RRR.
Faustin B, Chen Y, Zhai DY, Le Negrate G, Lartigue L, Satterthwait A, Reed JC. Mechanism of Bcl-2 and Bcl-X-L inhibition of NLRP1 inflammasome: Loop domain-dependent suppression of ATP binding and oligomerization. Proc Natl Acad Sci USA. 2009;106(10):3935–40. https://doi.org/10.1073/pnas.0809414106.
Conway KE, McConnell BB, Bowring CE, Donald CD, Warren ST, Vertino PM. TMS1, a novel proapoptotic caspase recruitment domain protein, is a target of methylation-induced gene silencing in human breast cancers. Cancer Res. 2000;60(22):6236–42. PMID: 11103776.
Akahira J, Sugihashi Y, Ito K, Niikura H, Okamura K, Yaegashi N. Promoter methylation status and expression of TMS1 gene in human epithelial ovarian cancer. Cancer Sci. 2004;95(1):40–3. https://doi.org/10.1111/j.1349-7006.2004.tb03168.x.
Alaminos M, Davalos V, Cheung NKV, Gerald WL, Esteller M. Clustering of gene hypermethylation associated with clinical risk groups in neuroblastoma. Jnci-J Natl Cancer Inst. 2004;96(16):1208–19. https://doi.org/10.1093/jnci/djh224.
Guan X, Sagara J, Yokoyama T, Koganehira Y, Oguchi M, Saida T, Taniguchi S. ASC/TMS1, A caspase-1 activating adaptor, is downregulated by aberrant methylation in human melanoma. Int J Cancer. 2003;107(2):202–8. https://doi.org/10.1002/ijc.11376.
Stone AR, Bobo W, Brat DJ, Devi NS, Van Meir EG, Vertino PM. Aberrant methylation and down-regulation of TMS1/ASC in human glioblastoma. Am J Pathol. 2004;165(4):1151–61. https://doi.org/10.1016/s0002-9440(10)63376-7.
Virmani A, Rathi A, Sugio K, Sathyanarayana UG, Toyooka S, Kischel FC, Tonk V, Padar A, Takahashi T, Roth JA, Euhus DM, Minna JD, Gazdar AF. Aberrant methylation of TMS1 in small cell, non small cell lung cancer and breast cancer. Int J Cancer. 2003;106(2):198–204. https://doi.org/10.1002/ijc.11206.
Yokoyama T, Sagara J, Guan X, Masumoto J, Takeoka M, Komiyama Y, Miyata K, Higuchi K, Taniguchi S. Methylation of ASC/TMS1, a proapoptotic gene responsible for activating procaspase-1, in human colorectal cancer. Cancer Lett. 2003;202(1):101–8. https://doi.org/10.1016/j.canlet.2003.08.027.
Masumoto J, Dowds TA, Schaner P, Chen FF, Ogura Y, Li M, Zhu L, Katsuyama T, Sagara J, Taniguchi S, Gumucio DL, Nunez G, Inohara N. ASC is an activating adaptor for NF-kappa B and caspase-8-dependent apoptosis. Biochem Biophys Res Commun. 2003;303(1):69–73. https://doi.org/10.1016/s0006-291x(03)00309-7.
Richards N, Schaner P, Diaz A, Stuckey J, Shelden E, Wadhwa A, Gumucio DL. Interaction between pyrin and the apoptotic speck protein (ASC) modulates ASC-induced apoptosis. J Biol Chem. 2001;276(42):39320–9. https://doi.org/10.1074/jbc.M104730200.
Deswaerte V, Nguyen P, West A, Browning AF, Yu L, Ruwanpura SM, Balic J, Livis T, Girard C, Preaudet A, Oshima H, Fung KY, Tye H, Najdovska M, Ernst M, Oshima M, Gabay C, Putoczki T, Jenkins BJ. Inflammasome adaptor ASC suppresses apoptosis of gastric cancer cells by an IL18-mediated inflammation-independent mechanism. Can Res. 2018;78(5):1293–307. https://doi.org/10.1158/0008-5472.can-17-1887.
Protti MP, De Monte L. Dual role of inflammasome adaptor ASC in cancer. Front Cell Dev Biol. 2020;8:40. https://doi.org/10.3389/fcell.2020.00040.
Collard RL, Harya NS, Monzon FA, Maier CE, O’Keefe DS. Methylation of the ASC gene promoter is associated with aggressive prostate cancer. Prostate. 2006;66(7):687–95. https://doi.org/10.1002/pros.20371.
Das PM, Ramachandran K, VanWert J, Ferdinand L, Gopisetty G, Reis IM, Singal R. Methylation mediated silencing of TMS1/ASC gene in prostate cancer. Mol Cancer. 2006;5. https://doi.org/10.1186/1476-4598-5-28.
Franchi L, Eigenbrod T, Muñoz-Planillo R, Nuñez G. The inflammasome: a caspase-1-activation platform that regulates immune responses and disease pathogenesis. Nat Immunol. 2009;10(3):241–7. https://doi.org/10.1038/ni.1703.
Ueki T, Takeuchi T, Nishimatsu H, Kajiwara T, Moriyama N, Narita Y, Kawabe K, Ueki K, Kitamura T. Silencing of the caspase-1 gene occurs in murine and human renal cancer cells and causes solid tumor growth in vivo. Int J Cancer. 2001;91(5):673–9. https://doi.org/10.1002/1097-0215(200002)9999:9999<::aid-ijc1113>3.0.co;2-u.
Sun Y, Guo Y. Expression of Caspase-1 in breast cancer tissues and its effects on cell proliferation, apoptosis and invasion. Oncol Lett. 2018;15(5):6431–5. https://doi.org/10.3892/ol.2018.8176.
Jarry A, Vallette G, Cassagnau E, Moreau A, Bou-Hanna C, Lemarre P, Letessier E, Le Neel JC, Galmiche JP, Laboisse CL. Interleukin 1 and interleukin 1beta converting enzyme (caspase 1) expression in the human colonic epithelial barrier. Caspase 1 downregulation in colon cancer. Gut. 1999;45(2):246–51. https://doi.org/10.1136/gut.45.2.246.
Feng Q, Li P, Salamanca C, Huntsman D, Leung PC, Auersperg N. Caspase-1alpha is down-regulated in human ovarian cancer cells and the overexpression of caspase-1alpha induces apoptosis. Cancer Res. 2005;65(19):8591–6. https://doi.org/10.1158/0008-5472.can-05-0239.
Gansauge S, Gansauge F, Yang Y, Müller J, Seufferlein T, Ramadani M, Beger HG. Interleukin 1beta-converting enzyme (caspase-1) is overexpressed in adenocarcinoma of the pancreas. Cancer Res. 1998;58(13):2703–6.
Winter RN, Kramer A, Borkowski A, Kyprianou N. Loss of caspase-1 and caspase-3 protein expression in human prostate cancer. Cancer Res. 2001;61(3):1227–32. ID: 11221855.
Zhang YQ, Yang J, Wen ZH, Chen XY, Yu J, Yuan DB, Xu BX, Luo H, Zhu JG. A novel 3 ’,5 ’-diprenylated chalcone induces concurrent apoptosis and GSDME-dependent pyroptosis through activating PKC delta/JNK signal in prostate cancer. Aging-Us. 2020;12(10):9103–24. https://doi.org/10.18632/aging.103178.
Chang I, Mitsui Y, Kim SK, Sun JS, Jeon HS, Kang JY, Kang NJ, Fukuhara S, Gill A, Shahryari V, Tabatabai ZL, Greene KL, Dahiya R, Shin DM, Tanaka Y. Cytochrome P450 1B1 inhibition suppresses tumorigenicity of prostate cancer via caspase-1 activation. Oncotarget. 2017;8(24):39087–100. https://doi.org/10.18632/oncotarget.16598.
Garlanda C, Dinarello CA, Mantovani A. The interleukin-1 family: Back to the future. Immunity. 2013;39(6):1003–18. https://doi.org/10.1016/j.immuni.2013.11.010.
Bennett J, Starczynowski DT. IRAK1 and IRAK4 as emerging therapeutic targets in hematologic malignancies. Curr Opin Hematol. 2022;29(1):8–19. https://doi.org/10.1097/moh.0000000000000693.
Liu LM, Gudas LJ. Retinoic acid induces expression of the interleukin-1 beta gene in cultured normal human mammary epithelial cells and in human breast carcinoma lines. J Cell Physiol. 2002;193(2):244–52. https://doi.org/10.1002/jcp.10173.
Matanic D, Beg-Zec Z, Stojanovic D, Matakoric N, Flego V, Milevoj-Ribic F. Cytokines in patients with lung cancer. Scand J Immunol. 2003;57(2):173–8. https://doi.org/10.1046/j.1365-3083.2003.01205.x.
Carmi Y, Dotan S, Rider P, Kaplanov I, White MR, Baron R, Abutbul S, Huszar M, Dinarello CA, Apte RN, Voronov E. The role of IL-1 beta in the early tumor cell-induced angiogenic response. J Immunol. 2013;190(7):3500–9. https://doi.org/10.4049/jimmunol.1202769.
Yencilek F, Yildirim A, Yilmaz SG, Altinkilic EM, Dalan AB, Bastug Y, Isbir T. Investigation of interleukin-1 beta polymorphisms in prostate cancer. Anticancer Res. 2015;35(11):6057–61. PMID: 26504029.
Xu H, Ding Q, Jiang HW. Genetic polymorphism of interleukin-1A (IL-1A), IL-1B, and IL-1 receptor antagonist (IL-1RN) and prostate cancer risk. Asian Pac J Cancer Prev. 2014;15(20):8741–7. https://doi.org/10.7314/apjcp.2014.15.20.8741.
Ricote M, Garcia-Tunon I, Bethencourt FR, Fraile B, Paniagua R, Royuela M. Interleukin-1 (IL-1 alpha and IL-1 beta) and its receptors (IL-1RI, IL-1RII, and IL-1Ra) in prostate carcinoma. Cancer. 2004;100(7):1388–96. https://doi.org/10.1002/cncr.20142.
Bouraouj Y, Ben Rais N, Culig Z, Oueslati R. Involvement of interleukin-1 beta mediated nuclear factor kappa B signalling pathways to down-regulate prostate-specific antigen and cell proliferation in LNCaP prostate cancer cells. Cell Biol Int. 2012;36(5):449–54. https://doi.org/10.1042/cbi20100922.
Staverosky JA, Zhu XH, Ha S, Logan SK. Anti-androgen resistance in prostate cancer cells chronically induced by interleukin-1 beta. Am J Clin Exp Urol. 2013;1(1):53–65. PMID: 25374900.
Chang MA, Patel V, Gwede M, Morgado M, Tomasevich K, Fong EL, Farach-Carson MC, Delk NA. IL-1 beta Induces p62/SQSTM1 and Represses Androgen Receptor Expression in Prostate Cancer Cells. J Cell Biochem. 2014;115(12):2188–97. https://doi.org/10.1002/jcb.24897.
Lawrence T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb Perspect Biol. 2009;1(6):a001651. https://doi.org/10.1101/cshperspect.a001651.
Royuela M, Rodriguez-Berriguete G, Fraile B, Paniagua R. TNF-alpha/IL-1/NF-kappa B transduction pathway in human cancer prostate. Histol Histopathol. 2008;23(10):1279–90. https://doi.org/10.14670/HH-23.1279.
Nguyen DP, Li JY, Yadav SS, Tewari AK. Recent insights into NF-kappa B signalling pathways and the link between inflammation and prostate cancer. BJU Int. 2014;114(2):168–76. https://doi.org/10.1111/bju.12488.
Grosset AA, Ouellet V, Caron C, Fragoso G, Barrès V, Delvoye N, Latour M, Aprikian A, Bergeron A, Chevalier S, Fazli L, Fleshner N, Gleave M, Karakiewicz P, Lacombe L, Lattouf JB, van der Kwast T, Trudel D, Mes-Masson AM, Saad F. Validation of the prognostic value of NF-κB p65 in prostate cancer: A retrospective study using a large multi-institutional cohort of the Canadian Prostate Cancer Biomarker Network. PLoS Med. 2019;16(7):e1002847. https://doi.org/10.1371/journal.pmed.1002847.
Nguyen DP, Li JY, Tewari AK. Inflammation and prostate cancer: the role of interleukin 6 (IL-6). BJU Int. 2014;113(6):986–92. https://doi.org/10.1111/bju.12452.
Jin RJ, Lho YS, Connelly L, Wang YQ, Yu XP, Saint Jean L, Case TC, Ellwood-Yen K, Sawyers CL, Bhowmick NA, Blackwell TS, Yull FE, Matusik RJ. The nuclear factor-kappa B pathway controls the progression of prostate cancer to androgen-independent growth. Cancer Res. 2008;68(16):6762–9. https://doi.org/10.1158/0008-5472.can-08-0107.
Lessard L, Karakievvicz PI, Bellon-Gagnon P, Alam-Fahmy M, Ismail HA, Mes-Masson AM, Saad F. Nuclear localization of nuclear factor-kappa B p65 in primary prostate tumors is highly predictive of pelvic lymph node metastases. Clin Cancer Res. 2006;12(19):5741–5. https://doi.org/10.1158/1078-0432.ccr-06-0330.
Jung YJ, Isaacs JS, Lee SM, Trepel J, Neckers L. IL-1 beta mediated up-regulation of HIF-1 alpha via an NFkB/COX-2 pathway identifies HIF-1 as a critical link between inflammation and oncogenesis. Faseb J. 2003;17(12):2115–+. https://doi.org/10.1096/fj.03-0329fje.
Sartori-Cintra AR, Mara CS, Argolo DL, Coimbra IB. Regulation of hypoxia-inducible factor-1α (HIF-1α) expression by interleukin-1β (IL-1 β), insulin-like growth factors I (IGF-I) and II (IGF-II) in human osteoarthritic chondrocytes. Clinics (Sao Paulo). 2012;67(1):35–40. https://doi.org/10.6061/clinics/2012(01)06.
Xanthopoulou ET, Kakouratos C, Nanos C, Gkegka AG, Kalaitzis C, Giatromanolaki A, Koukourakis MI. HIF1α-dependent and independent pathways regulate the expression of PD-L1 in prostate cancer. Med Oncol. 2023;40(5):151. https://doi.org/10.1007/s12032-023-02017-6.
Tran MGB, Bibby BAS, Yang LJ, Lo F, Warren AY, Shukla D, Osborne M, Hadfield J, Carroll T, Stark R, Scott H, Ramos-Montoya A, Massie C, Maxwell P, West CML, Mills IG, Neal DE. Independence of HIF1a and androgen signaling pathways in prostate cancer. BMC Cancer. 2020;20(1):12. https://doi.org/10.1186/s12885-020-06890-6.
Martinon F. Signaling by ROS drives inflammasome activation. Eur J Immunol. 2010;40(3):616–9. https://doi.org/10.1002/eji.200940168.
Anuja K, Roy S, Ghosh C, Gupta P, Bhattacharjee S, Banerjee B. Prolonged inflammatory microenvironment is crucial for pro-neoplastic growth and genome instability: a detailed review. Inflamm Res. 2017;66(2):119–28. https://doi.org/10.1007/s00011-016-0985-3.
Salminen A, Kauppinen A, Kaarniranta K. Emerging role of NF-kappa B signaling in the induction of senescence-associated secretory phenotype (SASP). Cell Signal. 2012;24(4):835–45. https://doi.org/10.1016/j.cellsig.2011.12.006.
Gracie JA, Robertson SE, McInnes IB. Interleukin-18. J Leukoc Biol. 2003;73(2):213–24. https://doi.org/10.1189/jlb.0602313.
Fujita K, Ewing CM, Isaacs WB, Pavlovich CP. Immunomodulatory IL-18 binding protein is produced by prostate cancer cells and its levels in urine and serum correlate with tumor status. Int J Cancer. 2011;129(2):424–32. https://doi.org/10.1002/ijc.25705.
Lebel-Binay S, Thiounn N, De Pinieux G, Vieillefond A, Debre B, Bonnefoy JY, Fridman WH, Pages F. IL-18 is produced by prostate cancer cells and secreted in response to interferons. Int J Cancer. 2003;106(6):827–35. https://doi.org/10.1002/ijc.11285.
Tse BWC, Russell PJ, Lochner M, Forster I, Power CA. IL-18 inhibits growth of murine orthotopic prostate carcinomas via both adaptive and innate immune mechanisms. PLoS ONE. 2011;6(9):12. https://doi.org/10.1371/journal.pone.0024241.
Lieberman J, Wu H, Kagan JC. Gasdermin D activity in inflammation and host defense. Sci Immunol. 2019;4(39). https://doi.org/10.1126/sciimmunol.aav1447.
Evavold CL, Ruan J, Tan Y, Xia S, Wu H, Kagan JC. The pore-forming protein gasdermin D regulates interleukin-1 secretion from living macrophages. Immunity. 2018;48(1):35–44.e6. https://doi.org/10.1016/j.immuni.2017.11.013.
Ruan J, Xia S, Liu X, Lieberman J, Wu H. Cryo-EM structure of the gasdermin A3 membrane pore. Nature. 2018;557(7703):62–7. https://doi.org/10.1038/s41586-018-0058-6.
Liu X, Zhang Z, Ruan J, Pan Y, Magupalli VG, Wu H, Lieberman J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature. 2016;535(7610):153–8. https://doi.org/10.1038/nature18629.
Wang WJ, Chen D, Jiang MZ, Xu B, Li XW, Chu Y, Zhang YJ, Mao R, Liang J, Fan DM. Downregulation of gasdermin D promotes gastric cancer proliferation by regulating cell cycle-related proteins. J Dig Dis. 2018;19(2):74–83. https://doi.org/10.1111/1751-2980.12576.
Obesity: preventing and managing the global epidemic. Report of a WHO consultation. World Health Organ Tech Rep Ser. 2000;894:i-xii, 1–253. PMID: 11234459.
Chung A, Backholer K, Wong E, Palermo C, Keating C, Peeters A. Trends in child and adolescent obesity prevalence in economically advanced countries according to socioeconomic position: a systematic review. Obes Rev. 2016;17(3):276–95. https://doi.org/10.1111/obr.12360.
World Health Organization. Obesity and poverty: a new public health challenge. Pan American Health Organization. 2000. Available at: https://iris.paho.org/handle/10665.2/4007.
Kershaw EE, Flier JS. Adipose tissue as an endocrine organ. J Clin Endocrinol Metab. 2004;89(6):2548–56. https://doi.org/10.1210/jc.2004-0395.
Rosen ED, Spiegelman BM. What we talk about when we talk about fat. Cell. 2014;156(1–2):20–44. https://doi.org/10.1016/j.cell.2013.12.012.
Bourin P, Bunnell BA, Casteilla L, Dominici M, Katz AJ, March KL, Redl H, Rubin JP, Yoshimura K, Gimble JM. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: a joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy. 2013;15(6):641–8. https://doi.org/10.1016/j.jcyt.2013.02.006.
Gregor MF, Hotamisligil GS. Inflammatory mechanisms in obesity. Annu Rev Immunol. 2011;29:415–45. https://doi.org/10.1146/annurev-immunol-031210-101322.
Vandanmagsar B, Youm YH, Ravussin A, Galgani JE, Stadler K, Mynatt RL, Ravussin E, Stephens JM, Dixit VD. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat Med. 2011;17(2):179–U214. https://doi.org/10.1038/nm.2279.
Esser N, L’Homme L, De Roover A, Kohnen L, Scheen AJ, Moutschen M, Piette J, Legrand-Poels S, Paquot N. Obesity phenotype is related to NLRP3 inflammasome activity and immunological profile of visceral adipose tissue. Diabetologia. 2013;56(11):2487–97. https://doi.org/10.1007/s00125-013-3023-9.
Duewell P, Kono H, Rayner KJ, Sirois CM, Vladimer G, Bauernfeind FG, Abela GS, Franchi L, Nuñez G, Schnurr M, Espevik T, Lien E, Fitzgerald KA, Rock KL, Moore KJ, Wright SD, Hornung V, Latz E. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature. 2010;464(7293):1357–61. https://doi.org/10.1038/nature08938.
Legrand-Poels S, Esser N, L’Homme L, Scheen A, Paquot N, Piette J. Free fatty acids as modulators of the NLRP3 inflammasome in obesity/type 2 diabetes. Biochem Pharmacol. 2014;92(1):131–41. https://doi.org/10.1016/j.bcp.2014.08.013.
Koenen TB, Stienstra R, van Tits LJ, de Graaf J, Stalenhoef AF, Joosten LA, Tack CJ, Netea MG. Hyperglycemia activates caspase-1 and TXNIP-mediated IL-1beta transcription in human adipose tissue. Diabetes. 2011;60(2):517–24. https://doi.org/10.2337/db10-0266.
Xia QS, Lu FE, Wu F, Huang ZY, Dong H, Xu LJ, Gong J. New role for ceramide in hypoxia and insulin resistance. World J Gastroenterol. 2020;26(18):2177–86. https://doi.org/10.3748/wjg.v26.i18.2177.
Unamuno X, Gómez-Ambrosi J, Ramírez B, Rodríguez A, Becerril S, Valentí V, Moncada R, Silva C, Salvador J, Frühbeck G, Catalán V. NLRP3 inflammasome blockade reduces adipose tissue inflammation and extracellular matrix remodeling. Cell Mol Immunol. 2019. https://doi.org/10.1038/s41423-019-0296-z.
Sharma M, Boytard L, Hadi T, Koelwyn G, Simon R, Ouimet M, Seifert L, Spiro W, Yan B, Hutchison S, Fisher EA, Ramasamy R, Ramkhelawon B, Moore KJ. Enhanced glycolysis and HIF-1α activation in adipose tissue macrophages sustains local and systemic interleukin-1β production in obesity. Sci Rep. 2020;10(1):5555. https://doi.org/10.1038/s41598-020-62272-9.
Zhu P, Zhang JJ, Cen Y, Yang Y, Wang F, Gu KP, Yang HT, Wang YZ, Zou ZQ. High endogenously synthesized N-3 polyunsaturated fatty acids in Fat-1 Mice attenuate high-fat diet-induced insulin resistance by inhibiting NLRP3 inflammasome activation via Akt/GSK-3β/TXNIP pathway. Molecules. 2022;27(19). https://doi.org/10.3390/molecules27196384.
Deng Y, Han X, Yao Z, Sun Y, Yu J, Cai J, Ren G, Jiang G, Han F. PPARα agonist stimulated angiogenesis by improving endothelial precursor cell function via a NLRP3 inflammasome pathway. Cell Physiol Biochem. 2017;42(6):2255–66. https://doi.org/10.1159/000479999.
Trøseid M, Seljeflot I, Arnesen H. The role of interleukin-18 in the metabolic syndrome. Cardiovasc Diabetol. 2010;9:11. https://doi.org/10.1186/1475-2840-9-11.
Hung J, McQuillan BM, Chapman CM, Thompson PL, Beilby JP. Elevated interleukin-18 levels are associated with the metabolic syndrome independent of obesity and insulin resistance. Arterioscler Thromb Vasc Biol. 2005;25(6):1268–73. https://doi.org/10.1161/01.atv.0000163843.70369.12.
Esposito K, Nappo F, Giugliano F, Di Palo C, Ciotola M, Barbieri M, Paolisso G, Giugliano D. Meal modulation of circulating interleukin 18 and adiponectin concentrations in healthy subjects and in patients with type 2 diabetes mellitus. Am J Clin Nutr. 2003;78(6):1135–40. https://doi.org/10.1093/ajcn/78.6.1135.
Murphy AJ, Kraakman MJ, Kammoun HL, Dragoljevic D, Lee MK, Lawlor KE, Wentworth JM, Vasanthakumar A, Gerlic M, Whitehead LW, DiRago L, Cengia L, Lane RM, Metcalf D, Vince JE, Harrison LC, Kallies A, Kile BT, Croker BA, Febbraio MA, Masters SL. IL-18 production from the NLRP1 inflammasome prevents obesity and metabolic syndrome. Cell Metab. 2016;23(1):155–64. https://doi.org/10.1016/j.cmet.2015.09.024.
Raut PK, Kim SH, Choi DY, Jeong GS, Park PH. Growth of breast cancer cells by leptin is mediated via activation of the inflammasome: Critical roles of estrogen receptor signaling and reactive oxygen species production. Biochem Pharmacol. 2019;161:73–88. https://doi.org/10.1016/j.bcp.2019.01.006.
Raut PK, Park PH. Globular adiponectin antagonizes leptin-induced growth of cancer cells by modulating inflammasomes activation: Critical role of HO-1 signaling. Biochem Pharmacol. 2020;180:114186. https://doi.org/10.1016/j.bcp.2020.114186.
Fu S, Liu L, Han L, Yu Y. Leptin promotes IL-18 secretion by activating the NLRP3 inflammasome in RAW 264.7 cells. Mol Med Rep. 2017;16(6):9770–6. https://doi.org/10.3892/mmr.2017.7797.
Gong Z, Zhang X, Su K, Jiang R, Sun Z, Chen W, Forno E, Goetzman ES, Wang J, Dong HH, Dutta P, Muzumdar R. Deficiency in AIM2 induces inflammation and adipogenesis in white adipose tissue leading to obesity and insulin resistance. Diabetologia. 2019;62(12):2325–39. https://doi.org/10.1007/s00125-019-04983-x.
Kolb R, Phan L, Borcherding N, Liu Y, Yuan F, Janowski AM, Xie Q, Markan KR, Li W, Potthoff MJ, Fuentes-Mattei E, Ellies LG, Knudson CM, Lee MH, Yeung SJ, Cassel SL, Sutterwala FS, Zhang W. Obesity-associated NLRC4 inflammasome activation drives breast cancer progression. Nat Commun. 2016;7:13007. https://doi.org/10.1038/ncomms13007.
Yao J, Chen Y, Xu M. The critical role of short-chain fatty acids in health and disease: A subtle focus on cardiovascular disease-NLRP3 inflammasome-angiogenesis axis. Clin Immunol. 2022;238:109013. https://doi.org/10.1016/j.clim.2022.109013.
Arendt LM, McCready J, Keller PJ, Baker DD, Naber SP, Seewaldt V, Kuperwasser C. Obesity promotes breast cancer by CCL2-mediated macrophage recruitment and angiogenesis. Cancer Res. 2013;73(19):6080–93. https://doi.org/10.1158/0008-5472.can-13-0926.
Truax AD, Chen L, Tam JW, Cheng N, Guo H, Koblansky AA, Chou WC, Wilson JE, Brickey WJ, Petrucelli A, Liu R, Cooper DE, Koenigsknecht MJ, Young VB, Netea MG, Stienstra R, Sartor RB, Montgomery SA, Coleman RA, Ting JP. The inhibitory innate immune sensor NLRP12 maintains a threshold against obesity by regulating gut microbiota homeostasis. Cell Host Microbe. 2018;24(3):364–378.e6. https://doi.org/10.1016/j.chom.2018.08.009.
Huang H, Chi H, Liao D, Zou Y. Effects of coenzyme Q(10) on cardiovascular and metabolic biomarkers in overweight and obese patients with type 2 diabetes mellitus: a pooled analysis. Diabetes Metab Syndr Obes. 2018;11:875–86. https://doi.org/10.2147/dmso.s184301.
Chai W, Cooney RV, Franke AA, Caberto CP, Wilkens LR, Le Marchand L, Goodman MT, Henderson BE, Kolonel LN. Plasma coenzyme Q10 levels and prostate cancer risk: the multiethnic cohort study. Cancer Epidemiol Biomarkers Prev. 2011;20(4):708–10. https://doi.org/10.1158/1055-9965.epi-10-1309.
Yang HL, Lin PY, Vadivalagan C, Lin YA, Lin KY, Hseu YC. Coenzyme Q(0) defeats NLRP3-mediated inflammation, EMT/metastasis, and Warburg effects by inhibiting HIF-1α expression in human triple-negative breast cancer cells. Arch Toxicol. 2023;97(4):1047–68. https://doi.org/10.1007/s00204-023-03456-w.
Donohoe CL, Lysaght J, O’Sullivan J, Reynolds JV. Emerging concepts linking obesity with the hallmarks of cancer. Trends Endocrinol Metab. 2017;28(1):46–62. https://doi.org/10.1016/j.tem.2016.08.004.
Poulose N, Amoroso F, Steele RE, Singh R, Ong CW, Mills IG. Genetics of lipid metabolism in prostate cancer. Nat Genet. 2018;50(2):169–71. https://doi.org/10.1038/s41588-017-0037-0.
Yue S, Li J, Lee SY, Lee HJ, Shao T, Song B, Cheng L, Masterson TA, Liu X, Ratliff TL, Cheng JX. Cholesteryl ester accumulation induced by PTEN loss and PI3K/AKT activation underlies human prostate cancer aggressiveness. Cell Metab. 2014;19(3):393–406. https://doi.org/10.1016/j.cmet.2014.01.019.
Sena LA, Denmeade SR. Fatty acid synthesis in prostate cancer: Vulnerability or epiphenomenon? Cancer Res. 2021;81(17):4385–93. https://doi.org/10.1158/0008-5472.can-21-1392.
Scheinberg T, Mak B, Butler L, Selth L, Horvath LG. Targeting lipid metabolism in metastatic prostate cancer. Ther Adv Med Oncol. 2023;15:17588359231152840. https://doi.org/10.1177/17588359231152839.
Hurwitz LM, Dogbe N, Hughes Barry K, Koutros S, Berndt SI. Obesity and prostate cancer screening, incidence, and mortality in the PLCO cancer screening trial. J Natl Cancer Inst. 2023. https://doi.org/10.1093/jnci/djad113.
Popovici D, Stanisav C, Pricop M, Dragomir R, Saftescu S, Ciurescu D. Associations between body mass index and prostate cancer: The impact on progression-free survival. Medicina (Kaunas). 2023;59(2). https://doi.org/10.3390/medicina59020289.
Samora NL, Wallis CJD, Huang LC, Tallman JE, Zhao Z, Hoffman K, Morgans A, Cooperberg M, Goodman M, Greenfield S, Hamilton AS, Hashibe M, Kaplan S, O’Neil B, Paddock LE, Stroup A, Wu XC, Koyama T, Penson DF, Barocas DA. Association between body mass index and localized prostate cancer management and disease-specific quality of life. BJUI Compass. 2023;4(2):223–33. https://doi.org/10.1002/bco2.197.
Perez-Cornago A, Smith-Byrne K, Hazelwood E, Watling CZ, Martin S, Frayling T, Lewis S, Martin RM, Yaghootkar H, Travis RC, Key TJ. Genetic predisposition to metabolically unfavourable adiposity and prostate cancer risk: A Mendelian randomization analysis. Cancer Med. 2023. https://doi.org/10.1002/cam4.6220.
Fontana F, Anselmi M, Limonta P. Adipocytes reprogram prostate cancer stem cell machinery. J Cell Commun Signal. 2023;17(3):915–24. https://doi.org/10.1007/s12079-023-00738-x.
Ribeiro R, Monteiro C, Catalán V, Hu P, Cunha V, Rodríguez A, Gómez-Ambrosi J, Fraga A, Príncipe P, Lobato C, Lobo F, Morais A, Silva V, Sanches-Magalhães J, Oliveira J, Pina F, Lopes C, Medeiros R, Frühbeck G. Obesity and prostate cancer: gene expression signature of human periprostatic adipose tissue. BMC Med. 2012;10:108. https://doi.org/10.1186/1741-7015-10-108.
Woo S, Cho JY, Kim SY, Kim SH. Periprostatic fat thickness on MRI: correlation with Gleason score in prostate cancer. AJR Am J Roentgenol. 2015;204(1):W43–7. https://doi.org/10.2214/ajr.14.12689.
Taylor RA, Lo J, Ascui N, Watt MJ. Linking obesogenic dysregulation to prostate cancer progression. Endocr Connect. 2015;4(4):R68–80. https://doi.org/10.1530/ec-15-0080.
Roumiguié M, Estève D, Manceau C, Toulet A, Gilleron J, Belles C, Jia Y, Houël C, Pericart S, LeGonidec S, Valet P, Cormont M, Tanti JF, Malavaud B, Bouloumié A, Milhas D, Muller C. Periprostatic adipose tissue displays a chronic hypoxic state that limits its expandability. Am J Pathol. 2022;192(6):926–42. https://doi.org/10.1016/j.ajpath.2022.03.008.
Exley MA, Hand L, O’Shea D, Lynch L. Interplay between the immune system and adipose tissue in obesity. J Endocrinol. 2014;223(2):R41–8. https://doi.org/10.1530/joe-13-0516.
Toren P, Venkateswaran V. Periprostatic adipose tissue and prostate cancer progression: New insights into the tumor microenvironment. Clin Genitourin Cancer. 2014;12(1):21–6. https://doi.org/10.1016/j.clgc.2013.07.013.
Kwan HY, Chen M, Xu K, Chen B. The impact of obesity on adipocyte-derived extracellular vesicles. Cell Mol Life Sci. 2021;78(23):7275–88. https://doi.org/10.1007/s00018-021-03973-w.
Rios-Colon L, Chijioke J, Niture S, Afzal Z, Qi Q, Srivastava A, Ramalinga M, Kedir H, Cagle P, Arthur E, Sharma M, Moore J, Deep G, Suy S, Collins SP, Kumar D. Leptin modulated microRNA-628-5p targets Jagged-1 and inhibits prostate cancer hallmarks. Sci Rep. 2022;12(1):10073. https://doi.org/10.1038/s41598-022-13279-x.
Jasinski-Bergner S, Kielstein H. Adipokines regulate the expression of tumor-relevant MicroRNAs. Obes Facts. 2019;12(2):211–25. https://doi.org/10.1159/000496625.
Zorena K, Jachimowicz-Duda O, Ślęzak D, Robakowska M, Mrugacz M. Adipokines and obesity. Potential link to metabolic disorders and chronic complications. Int J Mol Sci. 2020;21(10). https://doi.org/10.3390/ijms21103570.
Jin Y, Xing J, Xu K, Liu D, Zhuo Y. Exosomes in the tumor microenvironment: Promoting cancer progression. Front Immunol. 2022;13:1025218. https://doi.org/10.3389/fimmu.2022.1025218.
Glinskii AB, Ma S, Ma J, Grant D, Lim CU, Guest I, Sell S, Buttyan R, Glinsky GV. Networks of intergenic long-range enhancers and snpRNAs drive castration-resistant phenotype of prostate cancer and contribute to pathogenesis of multiple common human disorders. Cell Cycle. 2011;10(20):3571–97. https://doi.org/10.4161/cc.10.20.17842.
Wang WL, Welsh J, Tenniswood M. 1,25-Dihydroxyvitamin D3 modulates lipid metabolism in prostate cancer cells through miRNA mediated regulation of PPARA. J Steroid Biochem Mol Biol. 2013;136:247–51. https://doi.org/10.1016/j.jsbmb.2012.09.033.
Amankwah EK, Anegbe E, Park H, Pow-Sang J, Hakam A, Park JY. miR-21, miR-221 and miR-222 expression and prostate cancer recurrence among obese and non-obese cases. Asian J Androl. 2013;15(2):226–30. https://doi.org/10.1038/aja.2012.160.
Azhati B, Reheman A, Dilixiati D, Rexiati M. FTO-stabilized miR-139-5p targets ZNF217 to suppress prostate cancer cell malignancies by inactivating the PI3K/Akt/mTOR signal pathway. Arch Biochem Biophys. 2023;741:109604. https://doi.org/10.1016/j.abb.2023.109604.
Lakshmanan VK, Ojha S, Jung YD. A modern era of personalized medicine in the diagnosis, prognosis, and treatment of prostate cancer. Comput Biol Med. 2020;126:104020. https://doi.org/10.1016/j.compbiomed.2020.104020.
Herrero-Aguayo V, Jiménez-Vacas JM, Sáez-Martínez P, Gómez-Gómez E, López-Cánovas JL, Garrido-Sánchez L, Herrera-Martínez AD, García-Bermejo L, Macías-González M, López-Miranda J, Castaño JP, Gahete MD, Luque RM. Influence of obesity in the miRNome: miR-4454, a key regulator of insulin response via splicing modulation in prostate. J Clin Endocrinol Metab. 2021;106(2):e469–84. https://doi.org/10.1210/clinem/dgaa580.
López-Castejón G, Pelegrín P. Current status of inflammasome blockers as anti-inflammatory drugs. Expert Opin Investig Drugs. 2012;21(7):995–1007. https://doi.org/10.1517/13543784.2012.690032.
Perregaux DG, McNiff P, Laliberte R, Hawryluk N, Peurano H, Stam E, Eggler J, Griffiths R, Dombroski MA, Gabel CA. Identification and characterization of a novel class of interleukin-1 post-translational processing inhibitors. J Pharmacol Exp Ther. 2001;299(1):187–97. ID: 11561079.
Basiorka AA, McGraw KL, Eksioglu EA, Chen X, Johnson J, Zhang L, Zhang Q, Irvine BA, Cluzeau T, Sallman DA, Padron E, Komrokji R, Sokol L, Coll RC, Robertson AA, Cooper MA, Cleveland JL, O’Neill LA, Wei S, List AF. The NLRP3 inflammasome functions as a driver of the myelodysplastic syndrome phenotype. Blood. 2016;128(25):2960–75. https://doi.org/10.1182/blood-2016-07-730556.
Hamarsheh S, Osswald L, Saller BS, Unger S, De Feo D, Vinnakota JM, Konantz M, Uhl FM, Becker H, Lübbert M, Shoumariyeh K, Schürch C, Andrieux G, Venhoff N, Schmitt-Graeff A, Duquesne S, Pfeifer D, Cooper MA, Lengerke C, Boerries M, Duyster J, Niemeyer CM, Erlacher M, Blazar BR, Becher B, Groß O, Brummer T, Zeiser R. Oncogenic Kras(G12D) causes myeloproliferation via NLRP3 inflammasome activation. Nat Commun. 2020;11(1):1659. https://doi.org/10.1038/s41467-020-15497-1.
Chen L, Huang CF, Li YC, Deng WW, Mao L, Wu L, Zhang WF, Zhang L, Sun ZJ. Blockage of the NLRP3 inflammasome by MCC950 improves anti-tumor immune responses in head and neck squamous cell carcinoma. Cell Mol Life Sci. 2018;75(11):2045–58. https://doi.org/10.1007/s00018-017-2720-9.
Wannamaker W, Davies R, Namchuk M, Pollard J, Ford P, Ku G, Decker C, Charifson P, Weber P, Germann UA, Kuida K, Randle JC. (S)-1-((S)-2-{[1-(4-amino-3-chloro-phenyl)-methanoyl]-amino}-3,3-dimethyl-butanoyl)-pyrrolidine-2-carboxylic acid ((2R,3S)-2-ethoxy-5-oxo-tetrahydro-furan-3-yl)-amide (VX-765), an orally available selective interleukin (IL)-converting enzyme/caspase-1 inhibitor, exhibits potent anti-inflammatory activities by inhibiting the release of IL-1beta and IL-18. J Pharmacol Exp Ther. 2007;321(2):509–16. https://doi.org/10.1124/jpet.106.111344.
Barrón-Cabrera E, González-Becerra K, Rosales-Chávez G, Mora-Jiménez A, Hernández-Cañaveral I, Martínez-López E. Low-grade chronic inflammation is attenuated by exercise training in obese adults through down-regulation of ASC gene in peripheral blood: a pilot study. Genes Nutr. 2020;15(1):15. https://doi.org/10.1186/s12263-020-00674-0.
Juliana C, Fernandes-Alnemri T, Wu J, Datta P, Solorzano L, Yu JW, Meng R, Quong AA, Latz E, Scott CP, Alnemri ES. Anti-inflammatory compounds parthenolide and Bay 11–7082 are direct inhibitors of the inflammasome. J Biol Chem. 2010;285(13):9792–802. https://doi.org/10.1074/jbc.M109.082305.
Lamkanfi M, Dixit VM. A new lead to NLRP3 inhibition. J Exp Med. 2017;214(11):3147–9. https://doi.org/10.1084/jem.20171848.
Marchetti C, Swartzwelter B, Gamboni F, Neff CP, Richter K, Azam T, Carta S, Tengesdal I, Nemkov T, D’Alessandro A, Henry C, Jones GS, Goodrich SA, St Laurent JP, Jones TM, Scribner CL, Barrow RB, Altman RD, Skouras DB, Gattorno M, Grau V, Janciauskiene S, Rubartelli A, Joosten LAB, Dinarello CA. OLT1177, a β-sulfonyl nitrile compound, safe in humans, inhibits the NLRP3 inflammasome and reverses the metabolic cost of inflammation. Proc Natl Acad Sci U S A. 2018;115(7):E1530–39. https://doi.org/10.1073/pnas.1716095115.
Gastaldi S, Boscaro V, Gianquinto E, Sandall CF, Giorgis M, Marini E, Blua F, Gallicchio M, Spyrakis F, MacDonald JA, Bertinaria M. Chemical modulation of the 1-(Piperidin-4-yl)-1,3-dihydro-2H-benzo[d]imidazole-2-one scaffold as a novel NLRP3 inhibitor. Molecules. 2021;26(13). https://doi.org/10.3390/molecules26133975.
He Y, Varadarajan S, Muñoz-Planillo R, Burberry A, Nakamura Y, Núñez G. 3,4-methylenedioxy-β-nitrostyrene inhibits NLRP3 inflammasome activation by blocking assembly of the inflammasome. J Biol Chem. 2014;289(2):1142–50. https://doi.org/10.1074/jbc.M113.515080.
He H, Jiang H, Chen Y, Ye J, Wang A, Wang C, Liu Q, Liang G, Deng X, Jiang W, Zhou R. Oridonin is a covalent NLRP3 inhibitor with strong anti-inflammasome activity. Nat Commun. 2018;9(1):2550. https://doi.org/10.1038/s41467-018-04947-6.
Huang Y, Jiang H, Chen Y, Wang X, Yang Y, Tao J, Deng X, Liang G, Zhang H, Jiang W, Zhou R. Tranilast directly targets NLRP3 to treat inflammasome-driven diseases. EMBO Mol Med. 2018;10(4). https://doi.org/10.15252/emmm.201708689.
Leu WJ, Chu JC, Hsu JL, Du CM, Jiang YH, Hsu LC, Huang WJ, Guh JH. Chalcones display anti-NLRP3 inflammasome activity in macrophages through inhibition of both priming and activation steps-structure-activity-relationship and mechanism studies. Molecules. 2020;25(24). https://doi.org/10.3390/molecules25245960.
Shao BZ, Xu ZQ, Han BZ, Su DF, Liu C. NLRP3 inflammasome and its inhibitors: a review. Front Pharmacol. 2015;6:262. https://doi.org/10.3389/fphar.2015.00262.
Chen S, Yao L, Huang P, He Q, Guan H, Luo Y, Zou Z, Wei S, Peng G, Yan J, Chen R, Zhang Q, Tao A. Blockade of the NLRP3/Caspase-1 axis ameliorates airway neutrophilic inflammation in a toluene diisocyanate-induced murine asthma model. Toxicol Sci. 2019;170(2):462–75. https://doi.org/10.1093/toxsci/kfz099.
Rudolphi K, Gerwin N, Verzijl N, van der Kraan P, van den Berg W. Pralnacasan, an inhibitor of interleukin-1beta converting enzyme, reduces joint damage in two murine models of osteoarthritis. Osteoarthritis Cartilage. 2003;11(10):738–46. https://doi.org/10.1016/s1063-4584(03)00153-5.
Keller M, Sollberger G, Beer HD. Thalidomide inhibits activation of caspase-1. J Immunol. 2009;183(9):5593–9. https://doi.org/10.4049/jimmunol.0900476.
Funding
This research was supported by the Spanish Ministry of Science, Innovation, and Universities (Research-Grant: PID2022-1381850B-I00, PID2019‐105564RB‐I00; Predoctoral contracts: FPU18-06009, FPU16-05059, FPU18-02485); Junta de Andalucía (BIO-0139); CIBERobn (CIBER is an initiative of Instituto de Salud Carlos III, Ministerio de Sanidad, Servicios Sociales e Igualdad, Spain).
Author information
Authors and Affiliations
Contributions
R. M. L. had the idea for the review and coordinated the work. J. M. P. G performed the literature search and redacted the main parts of every section. A. J. M. H, A. C. F. F, A. S. C, R. G. R, and A. D. H provided critical insights during the writing process and provided help for the redaction in every section. All the authors critically revised the work, and collectively reviewed and approved the final version of this review.
Corresponding author
Ethics declarations
Competing interests
The authors have no financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pérez-Gómez, J.M., Montero-Hidalgo, A.J., Fuentes-Fayos, A.C. et al. Exploring the role of the inflammasomes on prostate cancer: Interplay with obesity. Rev Endocr Metab Disord 24, 1165–1187 (2023). https://doi.org/10.1007/s11154-023-09838-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11154-023-09838-w