
Abstract
Large scale changes to the natural environment, due to both climate change and direct human activities (e.g., urbanization), pose a great risk to biodiversity. As this realization has set in, the number of studies modelling and monitoring the effect of environmental change on species response and survival has increased significantly; however, these studies focus on flagship species, often neglecting the lesser-known, cryptic species, which include the nocturnal primates. This is especially true for the 20 species of galago who, despite their large distribution throughout Africa, have had little to no research conducted on the effect of environmental change on population fitness and long-term survival. With urbanization set to increase throughout their distribution range, along with an increase in climate change parameters (e.g., increased temperature and a decrease in annual rainfall), the most studied species of the family, namely the African lesser bushbaby Galago moholi, provides an ideal model to assess the impact of future environmental change on galago species. In this review, we discuss the possible impact of environmental change on G. moholi and specifically focus on the effect of environmental change on feeding ecology, genetics, thermal biology, and reproduction, all of which contribute to the fitness and survival of the species. We also highlight the need for specific, long-term research to be conducted on the effect of environmental change on the survival likelihood of G. moholi and other galago species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Climate change and habitat destruction pose some of the greatest, long-term risks to species conservation (Thuiller, 2007). An increase in average global temperature will push many species beyond their thermal limits (Somero, 2010; Sunday et al., 2012), and the effects are amplified by other human-induced environmental changes, such as progressing habitat fragmentation and urbanisation. Furthermore, a change in seasonal rainfall and temperature patterns will significantly influence on the timing of reproduction, reproductive output, and sociality (Campos et al., 2017; Mazaris et al., 2008; Visser et al., 2004). Although the effect of climate change on species survivability depends on the distribution, physiology, and adaptability of a species, it is thought that one out of six species will be threatened with extinction due to elevated temperatures and associated environmental changes (Urban, 2015).
Non-human primates are often considered protected from the effects of climate change thanks to specific biological attributes such as behavioural, physiological, and dietary plasticity (Bernard & Marshall, 2020; Korstjens & Hillyer, 2016). Unfortunately, this is erroneous, as recent studies have shown that primates, among all endotherms, are disproportionately at risk to climate change (Pacifici et al., 2017). According to the International Union for the Conservation of Nature (IUCN, 15 February 2023), 62.6% of primates of threatened globally; on the African continent, 52.3% of mainland African species are threatened, with 42% in decline (Estrada et al., 2017). Several reasons are responsible for this: Firstly, most primates inhabit tropical habitats, which are known to have mild, seasonal fluctuations. As a result of this, a slight increase in environmental temperature will reach and exceed the thermal tolerance of these populations (Tewksbury et al., 2008). Next, as primates have evolved to survive in specific, stable habitats, most of these species are incapable of long-distance migration because of biological or anthropogenic factors, which would assist in mitigating any negative change in their present environment (Gnanadesikan et al., 2017; Zhao et al., 2019). Finally, because of the long generation times inherent in primates, the likelihood of accumulating beneficial adaptations fast enough to mitigate the harmful effects of climate change will be limited (Meyer & Pie, 2022).
Found in the strepsirrhine suborder is the family galagidae, which consists of 20 species spread across Africa, making it one of the most widespread primate families (Bearder, 1999; Bearder et al., 1995; Nekaris & Bearder, 2007). Despite their widespread distribution, to date, little to no research has been conducted to determine the effect of climate change and habitat alteration on any African galago species. There is a large size variation between species of the family, ranging from 60 g in the Rondo dwarf galago (Galagoides rondoensis; Honess et al., 2008) to 1500 g in the largest species, the thick-tailed bushbaby (Otolemur crassicaudatus; Nekaris & Bearder, 2007). Despite this difference in size, all species in the family display nocturnal activity patterns (Ellison et al., 2021; Svensson, 2017) and a diet which is generally thought to be specialised, ranging from insectivores, and gummivores to frugivores (Forbanka, 2018; Nekaris & Bearder, 2007). The African lesser bushbaby, G. moholi (Fig. 1), is the most well-known species of galagos (Table I) with an estimated lifespan of 3–5 years in the wild (Dausmann et al., 2012) and a wide distribution range over Southern Africa. Despite the use of the species as bushmeat, traditional medicine or in the pet trade, these activities are not thought to lead to widespread population decreases (Masters et al., 2016). Rather, it is the effects of factors such as climate change and urbanisation which are likely to be the biggest threat to the species. We aim to review the current information available for G. moholi and make predictions on how the species might adapt to environmental and climatic changes in its natural distribution. In this review, we discuss the possible ecological and physiological response of G. moholi to climate change and urbanisation. Specifically, we discuss the effect of habitat quality and fragmented habitats on population density, genetic diversity and population fitness. As habitat alteration will likely lead to a change in resource availability, we discuss the possible change in G. moholi feeding ecology in order for the species to survive. This is followed by a review of the thermoregulatory response of the species to extreme weather events, before discussing the likely change in reproductive activity which could occur when habitat quality and resource availability change considerably.

The African lesser bushbaby, Galago moholi (Photo credit: Polley, 2019).
Climate Change and Habitat Alteration Will Affect the Natural Habitat of Galago Moholi
To determine the correct conservation measures required to ensure species survival, it is crucial to assess species vulnerability in terms of sensitivity, resilience, and potential for adaptation (Huey et al., 2012). While current estimates show an average global temperature increase of 1.09 °C since the start of the twentieth century, the Intergovernmental Panel on Climate Change (2021) suggests this will reach 1.5 °C by 2030. An increase in global temperature along with a predicted decrease in rainfall will have a significant effect on habitat quality and carrying capacity globally (Meadows & Hoffman, 2003; Parnell & Walawege, 2011). Climate change is a significant concern in southern Africa, where the annual, terrestrial temperature increase is 1.1–1.5 times the observed global average of 0.65 °C, (Davis-Reddy & Vincent, 2017; Ziervogel et al., 2014). Unfortunately, it is difficult to monitor and link rainfall patterns to climate change due to its high spatial and year-to-year variability (Engelbrecht et al. 2013). However, there is a clear increase in the number of extreme climate events, for both temperature and rainfall, which includes frequent dry spells and daily rainfall intensity (Davis-Reddy & Vincent, 2017). A permanent increase in annual temperature and extreme weather events, along with the two-fold increase in carbon dioxide concentration, will likely result in the change of terrestrial ecosystems throughout southern Africa, including several vegetation biome shifts (Higgins & Scheiter, 2012). The change in habitat parameters and quality will have several important impacts on wildlife species, including altered distribution limits, population fitness, growth, and survival (Davis-Reddy & Vincent, 2017). Due to the large-scale threat climate change poses in southern Africa, conservation strategies focusing on habitat protection or increasing connectivity between protected land patches seem to be inadequate to assist in species conservation practices (Gillson et al., 2013). Thus, to enhance any conservation event, an adaptive response method is required, which can only be implemented if sufficient species-specific data are available (Prober et al., 2019).
In addition to climate change, countries in southern Africa are experiencing rapid urbanisation, in many cases above the global average (Gambe et al., 2023); however, despite the growth in infrastructure, urban human population growth rates in South Africa are steadily declining and could perhaps indicate a decline in urbanisation rates in the future (Turok & Borel-Saladin, 2014). Population growth in South Africa correlates with infrastructure development and thus urban growth (Marais & Cloete, 2016). Thus, in the period 2001–2011, nearly 60% of the observed population increase in South Africa occurred in metropolitan sections of the country, which led to further urban development (Turok & Borel-Saladin, 2014). One of the hallmarks of urbanisation is the complete transformation, in the form of progressive replacement or fragmentation, of the natural habitat (Antrop, 2004; Güneralp et al., 2017). Such considerable change in the natural habitat will result in several outcomes for wildlife species. Many wildlife populations will be unable to adapt to novel environments and resources, becoming locally extinct (DeVictor et al. 2007; de Andrade et al., 2019; Fenoglio et al., 2021). In contrast to this, several species seem to be able to adapt and survive in the urban landscape (Castillo-Contreras et al., 2021; Tait et al., 2014). These species must contend with several novel stressors and environmental conditions, including linear infrastructure, fragmented habitat, electrocution, and direct human-wildlife interactions, such as vehicle collisions (Cunneyworth & Duke, 2020; Cunneyworth & Slade, 2021; Linden et al., 2022a, b; Tigas et al., 2002).
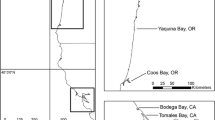
As species unable to adapt to rapidly altered environments will go locally extinct, both climate change and urbanisation poses some of the greatest risk to the survival of wildlife species (Acevedo-Whitehouse & Duffus, 2009; Soulsbury & White, 2015; Taylor-Brown et al., 2019). With a distribution ranging throughout southern Africa, including South Africa, Namibia, Botswana, Angola, and the Democratic Republic of the Congo (Fig. 2; Bearder et al., 2021; Masters et al., 2016), G. moholi is sure to experience the effects of climate change and urbanisation quite drastically. Long-term data from the Lajuma Research Station (23°02′18.0"S 29°26′34.1"E), in Limpopo, South Africa, an area inhabited by G. moholi, already show an upward trend in average annual temperature and a decrease in rainfall between 2012 to 2018 (Fig. 3). Moreover, South Africa experienced a flash drought during the 2015–2016 period (Xuan et al., 2018). Such extreme weather events are known to occur more frequently due to climate change and will likely be an important driver of G. moholi population stability. In addition to this, the change in land use from rural, rural environments to an urban landscape will result in further ecological- and fitness-related changes in the species.

The distribution range of the African lesser bushbaby, Galago moholi (Picture credit: African Geographic)

The annual temperature and rainfall patterns (2012–2018) at the Lajuma Research Centre, Limpopo, South Africa. Temperature and rainfall data were collected by weather stations at the Lajuma Research centre and made available by the South African Environmental Observation Network (SAEON) and Lajuma Research centre
Habitat Quality is Shaping Population Density
The ecology of G. moholi is relatively well studied, with the first documented systematic field study in 1966 (Bearder, 1987; Nekaris & Bearder, 2007). However, despite the species’ wide distribution range (Bearder et al., 2021), most studies have been conducted in north-eastern South Africa, specifically in the Gauteng and Limpopo provinces. As such, population-specific information on behaviour, physiology, and ecology, especially across different environments/climatic conditions, are absent for the species and we will focus our review on South African populations.
Both climate change and anthropogenic activities (deforestation, urbanisation, agriculture) are known to cause large-scale habitat fragmentation, leading to a reduction in habitat area, quality and carrying capacity, while limiting the dispersal ability of primates (see Arroyo-Rodríguez & Mandujano, 2006, 2009; Happold, 1995; Scanes, 2018). Galago moholi is generally found in semi-arid bushveld woodland, where individuals spend most of their time in trees, though foraging at ground-level is frequently observed (see Bearder et al., 2006; Nekaris & Bearder, 2007). Although galagos can survive in less preferred, natural habitats, most species show a clear preference for areas with dense trees due to their arboreal nature (Nekaris & Bearder, 2007). Initial density estimates of G. moholi in the natural environment was determined to be 0.87 animals/ha (Naboomspruit, Gauteng province; Bearder & Doyle, 1974), though later studies at the same study site found the density to be much lower at 0.31 animals/ha (Naboomspruit, Gauteng province; Bearder, 1987; Bearder & Martin, 1980) and 0.01 animals/ha (Loskop Dam Nature Reserve, Mpumalanga province; Ray et al., 2017). The difference observed between study populations could relate to several environmental factors, including the habitat size (dispersal ability), as well as resource availability. High dependence on environmental factors is also suggested by seasonally varying home range sizes of G. moholi. Home range estimates show male territories can range from 5–22 ha, while females maintain a smaller home range of 2–11 ha, but almost double their home range in the winter months when resource availability is drastically reduced (Nekaris & Bearder, 2007; Nowack et al., 2013b). In contrast to this, males seem to maintain a relatively constant home range, which overlaps with that of surrounding females (Nekaris & Bearder, 2007; Nowack et al., 2013a, b).
Due to their arboreal nature and dependence on forest connectivity, G. moholi may be at considerable risk to land transformation, habitat fragmentation and population seclusion (Masters et al., 2016). Furthermore, initial observations of the effect of climate change in the Limpopo province of South Africa, where G. moholi is abundant, shows that there will be a shift in vegetation type, from grasslands to tree-dominated landscapes (Scheiter et al., 2018); this might well be an advantage for galago species should it occur in most grasslands of southern Africa. Habitat fragmentation/alteration is known to cause a decrease in galago population densities, as shown for the dusky galago (G. matschiei; Weisenseel et al., 1993), Demidoff's galago (G. demidovii; Omifolaji et al., 2020), as well as several other nocturnal primate species (Pimley, 2009). This effect is not as simple, and the impact of habitat alteration on galago population dynamics may be driven by a complex relationship between habitat quality and the ability to disperse into and from neighbouring populations (Campos et al., 2012). This should be taken into consideration when predicting the possible effect of habitat alteration on galago densities.
Although no information regarding the effect of habitat fragmentation or alteration on G. moholi densities exists, the differences in range size and population density between different studies and habitats may indicate the importance of habitat quality (e.g., resource availability, climate etc.) and mating partner availability in driving population ecology and fitness. The removal of endemic tree species during urbanisation or tree-harvesting events, such as the gum-producing Vachellia karroo (formerly Acacia karroo), will remove important nesting and food sources, limiting the number of G. moholi which can survive in the environment (Masters et al., 2016). Studies on urban G. moholi populations indicate that the carrying capacity in urban environments are higher than observed in the rural environment (Scheun et al., 2015) in. Groups of up to ten G. moholi were observed in small fragmented suburban gardens (< 1 ha) with anthropogenic feeding sources but limited to no connectivity to adjacent green spaces. It is largely thanks to the constant availability of exotic and anthropogenic food sources that G. moholi can maintain such high population densities in the urban environment.
Fragmented Habitats Can Alter Genetic Diversity and Population Fitness
As high levels of genetic diversity in a population are beneficial in terms of adaptation to environmental change, managers and conservationists often use genetic diversity as a marker of population health and fitness (Frankham, 2005; Grogan et al., 2017; Markert et al., 2010; Toro & Caballero, 2005). High genetic diversity is often highlighted as an important parameter to consider when developing climate change species loss projections. Populations with high genetic diversity generally display higher behavioural and genetic adaptability to climate change and habitat alteration thanks to their genetic make-up (Pauls et al., 2013; Razgour et al., 2019; Willi & Hoffmann, 2009). However, several studies have shown that large-scale fragmentation, a shift in distribution range and habitat conservatism can all lead to the erosion of genetic variability and increased genetic divergence through reduced gene flow and increased genetic drift, especially in smaller wildlife populations (Marsh, 2003; Mhemmed et al., 2008; Razgour et al., 2013; Sousa et al., 2008).
While the ability of an individual to disperse into neighbouring populations can counteract the effect of genetic drift and inbreeding depression, the removal of natural habitat and migration corridors will limit such movement in many arboreal primate species (Ćosić et al., 2013; Jones, 2005). This pattern of genetic diversity loss due to the inability of individuals to disperse is often observed in small, captive primate populations (Charpentier et al., 2008; Neveu et al., 1998; Schulze et al., 1998). In wild, free-ranging primate populations a decrease in genetic diversity, because of habitat fragmentation/alteration, has been found in several primate species, including the samango monkey (Cercopithecus albogularis; Linden et al., 2022a, b), white-headed langur (Trachypithecus leucocephalus; Wang et al., 2017), pied tamarin (Saguinus bicolor; Farias et al., 2015), golden lion tamarin (Leontopithecus rosalia; Dietz et al., 2000) and silvery marmoset (Mico argentatus; Gonçalves et al., 2003). Unfortunately, specific information on the effect of habitat fragmentation on galago genetic diversity is not available. Many species in the family Galagidae are known to be philopatric, with females remaining in or around the home range of their mother; males, however, disperse to distant habitats (Nekaris & Bearder, 2007). As such, it is the males who are essentially responsible for the exchange of genetic material between populations.
In the thick-tailed bushbaby (Otolemur crassicaudatus), genetic diversity was low at various fragmented sites (fixation of mitochondrial haplotypes) likely due to limited migratory patterns observed in females (Phukuntsi et al., 2020); any native genetic material observed in the population was likely due to the movement of males between populations. While genetic diversity was found between several G. moholi populations, differentiation levels among populations were low, likely due to historic migration of males or translocation of individuals by the pet trade industry (Phukuntsi et al., 2021). It is clear from the highlighted studies that movement corridors must remain intact to maintain high levels of genetic variability, and thus population health and fitness. Like most primate species, G. moholi populations trapped in fragmented habitats are at risk of the erosion of genetic variability, and a reduction in the ability to react/adapt to climate change and further habitat alteration. While many other galago species have been observed living in fragmented habitats (Haji & Bakuza, 2022; Hanson et al., 2007; Zungu et al., 2018), no information exists on the effect thereof on the genetic fitness of any of these populations.
Climate Change and Urbanisation Can Lead to a Change in Food Resource Use
Urbanisation and climate change will not only affect habitat connectivity but alter food availability in once natural and now urbanised environments. Galago moholi has historically been described as a food specialist, exclusively feeding on insects, as well as gum from Vachellia karroo trees (Bearder, 1987; Nekaris & Bearder, 2007). Insects are known to be a nutrient dense food resource, consisting of protein, vitamins, mineral and fibre (Belluco et al., 2013; Rothman et al., 2014), making them an ideal food source to meet the high metabolic rates characteristic for small mammals (Clarke et al., 2010). Gum on the other hand seems to be mostly used as a secondary food source. It contains β-linked complex polysaccharides, which are resistant to digestive enzymes as part of the digestive process, and require long gut passage times, fermentation and specific gut microbes (Power, 2010; Caton et al., 2000). Consequently, it has considerably less nutritional value than protein-rich insects, but may provide an important source of calcium and minerals for the species (Bearder & Martin, 1980; Cabana et al., 2018; Jayne, 2020). Furthermore, the nutrient content of gum increases when insect biomass decreases during the cold, dry winter periods (Nowack et al., 2013b; Williams et al., 2015), which may explain why G. moholi shifts its dietary strategy from insects to Vachellia gum during the winter (Bearder & Martin, 1980; Harcourt, 1986; Nash, 1986; Nowack et al., 2013b).
Climate change and habitat modification have led to a permanent, global decrease in insect biomass over the past 25 years (Cardoso et al., 2020; Sánchez-Bayo & Wyckhuys, 2019). The reduction in preferred food sources, as well as the alteration and fragmentation of the natural environment, through both climate change and urbanisation, will pose a considerable threat to G. moholi. In support of this theory, Haji and Bakuza (2023) recently noted that a possible decrease in insect density in the similarly insect-specialised Rondo galago (G. rondoensis) will be one of the greatest threats to the survival of the species throughout its natural distribution. In response to food shortages, primates will need to adapt their movement and foraging behaviours to lower resource availability. One option to adapt to lower food availability could be an increased dependence on secondary foods such as gum to meet metabolic demands as already observed in winter (Nowack et al., 2013b). However, the extended use of gum throughout the year as a primary food source, alongside a decrease in insect consumption, may well lead to developmental constraints, changes in social ecology, elevated physiological stress and a decrease in general health and fitness. This has been noted for other primates using secondary food sources over prolonged periods (Cabana et al., 2018; Chapman et al., 2015; Koenig et al., 1997; Lea et al., 2015).
In addition to the increased use of gum, G. moholi would likely also need to extend the time spent foraging to meet metabolic requirements. For example, Bearder and Martin (1980) found that in periods of extreme food shortages, G. moholi foraged for additional food sources during the day. This shift in foraging behaviour and movement patterns, to meet metabolic requirements, has also been observed in several other primate species including the colobus monkeys (Colobus guereza; Harris et al., 2010), chimpanzees (Pan troglodytes schweinfurthii; Giuliano, 2021), lowland woolly monkeys (Lagothrix lagotricha poeppigii; Fiore & Rodman, 2001) and Yunnan snub-nosed monkeys (Rhinopithecus bieti; Grueter et al., 2013). Additional foraging activity will not only limit time spent on reproductive, social, and resting activities but increase the number of possible predator–prey encounters. In fact, the nocturnal, solitary foraging and limited social interaction seen in G. moholi are thought to be important antipredator mechanisms (Burnham et al., 2013). In support of the predator avoidance theory, G. senegalensis, of which G. moholi was a subspecies until recently (i.e., G. senegalensis moholi), is considered more gregarious in regions where predators are largely absent (Svensson, 2017). Thus, an increase in nocturnal and diurnal foraging behaviours will likely increase the risk of predator encounters and mortality in G. moholi.
Several studies have shown that feeding specialists, who depend on specific resources and are unable to use additional food sources, are most at risk of extinction should these primary sources be unavailable (Foden et al., 2013; Garnett et al., 2013; Sánchez-Bayo & Wyckhuys, 2021). In contrast, feeding generalists or species able to adapt to rapidly changing environments tend to survive where specialists go locally extinct (Colles et al., 2009; Isabirye-Basuta & Lwanga, 2008). Despite the historic classification of G. moholi as a specialised feeder, several studies have recently shown that the species can act as a generalist feeder under various environmental conditions, highlighting the dietary flexibility of the species. Anecdotal observations during our fieldwork in the urban and natural environment found that galagos can opportunistically supplement their diet with fruits from the jacket plum tree (Pappea capensis) or nectar from the bird of paradise flower (Strelitzia nicolai) (Scheun et al., 2014, 2019). Similarly, wild G. moholi were observed consuming fruit from the sickle bush tree, Dichrostachys cinerea (Ray et al., 2016), as well as feeding on several bird species and their eggs during a drought (Engelbrecht, 2016). As noted by the authors, in the absence of insects, birds and their eggs offer G. moholi a nutrient-rich resource which is easily accessible. Although such a permanent shift in dietary preference will allow the species to survive in altered habitats, it will likely lead to a considerable loss in avian populations that experience elevated nest predation (Kubelka et al., 2022).
Galago moholi populations found in urban, fragmented habitats have been shown to integrate anthropogenic food sources into their diet, which include breadcrumbs and yoghurt (Scheun et al., 2015, 2019). Despite the advantage such dietary plasticity holds for species inhabiting altered habitats, long-term consumption of anthropogenic food sources may have deleterious effects on populations. Several studies have shown that supplementary feeding can have a range of negative effects on wildlife species, including an increase in disease, stress, injury, and malnutrition (Kirby et al., 2019; Murray et al., 2016; Strandin et al., 2018; Wimberger et al., 2017). Furthermore, urban G. moholi populations feeding on anthropogenic food sources also had (1) significantly higher body mass index levels, (2) higher levels of intergroup aggression, (3) as well as a change in group/social dynamics compared to rural populations using natural sources (Scheun et al., 2015). Thus, although food provisioning can assist in short term survival of populations in altered environments, the negative effects of long-term consumption on the general health and reproductive success may well have several deleterious effects on population stability and survivability.
Torpor Use May Facilitate Survival During Drought Conditions
Galagos in South Africa inhabit a highly seasonal habitat with a cold and dry winter and wet and hot summer period where the daily variation can be substantial. Absolute temperatures up to 40°C during daytime have been observed and the daily difference between night and daytime temperature can be up to 35°C (recorded in October 2009 in a study in north-east South Africa; Nowack et al., 2013b). Due to the high seasonality in their habitat, the low food availability during the colder season, and the high energy demands of mammals this size, the use of energy saving torpor (i.e., a controlled reduction in body temperature and energy expenditure) had long been suggested based on anecdotal evidence (Skinner & Smithers, 1990), but was only confirmed in 2010 (Nowack et al., 2010). Earlier studies on the thermoregulation of G. moholi did not find any incidence of torpor use (Dobler, 1976; Knox & Wright, 1989; Mzilikazi et al., 2006), which is likely due to the use of mostly shallow and hard to detect bouts of torpor (body temperature > 30 °C) that are restricted to the rest phase (Nowack et al., 2023). Deeper bouts of torpor (body temperature < 30 °C) are only occasionally used (Nowack et al., 2010, 2013a). Instead, G. moholi does seem to mainly rely on behavioural means, such as changes in nesting sites, increased huddling, and a change in activity pattern to survive the cold and resource-low winter period (Nowack et al., 2013b).
While we have a good idea of how G. moholi cope with cold conditions there are virtually no data on how the species may respond to an increase in heat waves, droughts, or climate change related natural disasters. Exposure to heat is more dangerous than cold-exposure for endothermic mammals and birds and we are already seeing an increase in heat-associated mortality (McKechnie & Wolf, 2019). The upper limit of the thermal neutral zone of G. moholi, i.e., the temperature threshold above which animals must start using evaporative cooling to maintain a stable body temperature, has been determined as 35 °C (Dobler, 1978), but G. moholi are regularly exposed to daily maximum temperatures above this temperature in summer (Nowack et al., 2013b). Nocturnal activity, i.e., inactivity during the hottest time of the day and selection of well-buffered resting sites – if available – likely limits exposure to extreme temperature conditions. Laboratory studies on grey mouse lemurs (Microcebus murinus) suggest that the closely related lemurs cope well with chronic heat exposure up to 34 °C in the laboratory, although aged animals showed greater difficulties coping with heat exposure (Terrien et al., 2011). However, in the wild, heat waves often co-occur with droughts and reduced water availability, which can affect how well animals are able to cope with heat. At ambient temperatures above body temperature, evaporative water loss is the only option for an animal to cool down and limited water availability will thus have a large impact. There are two general physiological adaptations of animals to reduce evaporative water loss when water is scarce: (1) reducing metabolic rate through the use of torpor and thereby reducing water loss through feces, urine as well as respiratory water loss from breathing or (2) thermoconforming behaviour (adaptive or regulated heterothermy), i.e., a voluntary increase in body temperature to reduce the need of evaporative cooling (Nowack et al., 2017). Adaptive heterothermy is well known from many species coping with high ambient temperatures, such as Malagasy bats (Reher & Dausmann, 2021), Australian ring-tailed possums (Turner, 2020) as well as diurnal primates, i.e., baboons (Papio hamadryas; Mitchell et al., 2009). However, so far, an adaptive increase in body temperature to deal with high daytime ambient temperatures has not been observed in any galago species. G. moholi are, however, able to improve their water balance using torpor (Nowack et al., 2010). In addition to water loss through the skin, water is lost through the production of urine and faeces as well as during respiration, all of which are reduced to lower levels during torpor (Cooper et al., 2005). Furthermore, lower levels of metabolisms also lower the need for evaporative cooling as a mechanism of heat dissipation, allowing animals to conserve water for the times when evaporative cooling is crucial (Bondarenco et al., 2013).
While torpor use is mainly considered to be an adaptation to low food availability, a negative water balance can also be a strong trigger for torpor use in species in arid habitats (Cooper et al., 2005) and water availability is also suggested as a main trigger of torpor use in grey-mouse lemurs inhabiting Madagascar´s dry forest (Schmid & Speakman, 2009). Although torpor use in G. moholi has so far only been recorded in the winter period, galagos are able to undergo opportunistic torpor as a last resort strategy and it is thus likely that opportunistic torpor use can also aid survival under extreme temperatures and drought conditions or even climate-change related natural disasters, such as storms and fires, as reported for other species (reviewed in Nowack et al., 2017).
Long-Term Reproductive Change in G. moholi in Response to Climate Change and Urbanisation
Although several species exist that breed throughout the year (non-seasonal breeders), most mammal species outside of the tropical regions tend to breed during specific periods annually (seasonal breeders) (Guh et al., 2019). Seasonal breeding is an adaptive trait and allows an individual to avoid reproductive activity during inappropriate times of the year, while ensuring the survival of offspring who are born during periods of resource abundance and a mild climate (Bronson & Heideman, 1994; Guh et al., 2019; Nakane & Yoshimura, 2014).
Like most galagids, G. moholi is a seasonal, polygynous breeder (Bearder, 1987); in this regard, mating activity is restricted to two periods annually (Scheun et al., 2016). Several behavioural and physiological studies found that these mating periods occur during May/early June and late September/early October, when resources are abundant and temperatures favourable (Bearder & Martin, 1979; Doyle et al., 1971; Pullen et al. 2000; Scheun et al., 2016). However, the latter mating period is thought to be used as a ‘back-up’ mating period by females who did not become pregnant during the May period (Pullen et al. 2000). Scheun et al. (2017) also found that testosterone levels, testis volume and mating behaviour did not differ between mating seasons; thus, while the September mating period might act as a ‘back-up’ mating period for females, males may attempt to maximise their reproductive output over both mating periods. According to observational data, as determined from intromission to birth, the gestation length in G. moholi ranges from 120–141 days (Doyle et al., 1967; Izard & Nash, 1988; Nekaris & Bearder, 2007). Behavioural and endocrine monitoring in a captive population showed a gestational length of 128 days (Scheun et al., 2016). Taking into consideration the 4-month pregnancy period, most births occur around September and January (Scheun et al., 2016), where the nutritional needs of a lactating female can be met, while ensuring new-borns are not exposed to the extreme colds experienced in winter. In fact, it has been hypothesised that a trade-off with reproduction is one of the major factors explaining why G. moholi does not use torpor on a regular basis during winter (Nowack et al., 2010). Lower body temperatures associated with torpor use would delay parturition and thus shift the second mating period, making it unlikely for juveniles born before winter to reach a sufficient body mass to survive the resource poor period.
Climate change and habitat alteration are set to drastically change both abiotic and biotic parameters in the environment (Boutin & Lane, 2014; Skogen et al., 2018). While an increase in extreme weather events may reduce the mating season length, and lead to lower reproductive success (McNutt et al., 2019; Moss et al., 2001), an increase in favourable conditions (milder winters, early onset of spring, increased resource abundance) would lead to an increase in mating season length and reproductive success (Isaac, 2009; Moyes et al., 2011; Wegge & Rolstad, 2017). For example, late-born pied flycatchers (Ficedula hypoleuca) had an increased rate of survival, albeit at the cost of a reduced fattening phase prior to migration (Tomotani et al., 2016, 2018). Similar patterns will likely be observed in G. moholi in both natural and considerably altered environments, with prolonged or early onset mating and birthing periods occurring. Interestingly, a < 1 month old juvenile has been seen carried around and parked by the mother in July 2022, i.e., about 2 months prior to the normal birthing period, in an urban environment in South Africa (personal observation, JS).
Urban environments may well provide G. moholi populations with not only a mild climate throughout the year, but an abundance of resources which could remove the need for seasonal breeding. Similarly, an increase in insect availability due to warmer temperatures may also assist females who became pregnant in May and carrying the young across winter and into spring. Several studies have shown an increase in insect biomass when environmental temperature increase, providing an important food source for many insectivore species (Cornelissen, 2010; de Sassi & Tylianakis, 2012; Wolf et al., 2008). However, the opposite may be the case during the summer pregnancy period, where an increase in temperature throughout the savannah-like highveld distribution of G. moholi may result in temperatures exceeding the physiological limits of both insect and plant species (Cornelissen, 2010; de Sassi & Tylianakis, 2012). This will drastically decrease the resources available to meet metabolic demands linked to reproduction in the species. Furthermore, extreme fluctuations in environmental temperature may lead to suboptimal development and even genetic deficiency of the embryo (Nord & Giroud, 2020). Thus, although elevated temperature may have little to no direct effect on G. moholi reproduction, aspects such as resource decline and habitat isolation may influence population stability.
Finally, environmental stressors may influence the general and reproductive ecology of a species. The presence of an environmental stressor, be it injury, predator presence, aggressive interactions or low resource availability, will lead to the activation of the physiological stress response (Kersey & Dehnhard, 2014). One component thereof is the hyper-activation of the hypothalamic–pituitary–adrenal axis and an increase in secreted glucocorticoids (GCs; Beehner & Bergman, 2017). While an acute increase in GC concentrations can be adaptive in nature (Crespi et al., 2013), chronic elevated GC levels can have a host of deleterious effects on an organism, including immune and reproductive suppression (Charmandari et al., 2005; Hing et al., 2016). As such, monitoring GCs, or the excreted metabolites thereof in faeces, provides an ideal method for monitoring the perceive physiological stress experienced by an organism within its environment (Kersey & Dehnhard, 2014). Urban G. moholi populations displayed significantly elevated faecal glucocorticoid metabolites (fGCM) concentrations compared to conspecifics inhabiting rural environments (Scheun et al., 2015). The elevated fGCM levels observed in urban G. moholi may likely be because of insufficient resource availability, the use of anthropogenic food sources, altered group dynamics and aggression, as well as interactions with novel stressors. While the activation of the physiological stress response, and an acute increase in glucocorticoid levels, are adaptive in nature and can assist in the survival of an organism (Breuner & Hahn, 2003; Sapolsky et al., 2000; Wingfield et al., 1997), GC increase over extended periods may have several deleterious effects. Aside from the suppression of the immune response, and thus an increase in disease, ‘stressed’ individuals may also experience a suppression of reproductive function (Whirledge & Cidlowski, 2017). While several studies have noted the ability of certain wildlife species to exploit the urban environment (Lyons et al., 2017; Maibeche et al., 2015; Teixeira et al., 2015) and even enhance reproductive success as a result (Kettel et al., 2018; Narango et al., 2018), little to no information exists on the effect of long-term GC elevation on reproduction success in any urban species. Furthermore, as maternal GC levels may alter offspring temperament, fitness, and adult longevity (Hinde et al., 2014; Monaghan & Haussmann, 2015; Petelle et al., 2017; Todd, 2020), elevated GC levels throughout pregnancy may have a significant impact on long-term population growth and fitness.
Conclusion
Primates are currently threatened and nearing extinction due to anthropogenic activities, including factors such as habitat modification (e.g., climate change logging/urbanisation/mining/agriculture), overuse (e.g., bushmeat/traditional medicine), novel predators (e.g., domesticated pets) and disease (Estrada et al., 2017). Because of the number of threats facing primates, they are at risk of local or global extinction due to environmental change (Pacifici et al., 2017). Climate change and human induced environmental change will undoubtedly have drastic effects on G. moholi and other galago species and may affect reproductive timing and success as well as limit gene flow. However, each species of galagos has evolved unique ecological and physiological adaptations to survive in their specific habitat, making general predictions about the impact of future environmental change on galago species difficult. In regard to G. moholi, the species´ nocturnal lifestyle along with high levels of behavioural and physiological plasticity may allow adaptation to changing conditions: Selective torpor use as an energy- and water-saving adaptation may well act as a buffer against elevated temperatures. Furthermore, the ability to use various food sources means that the species is able to adapt to periods of limited naturally occurring food sources due to climate change or urbanisation. Additional research is needed to determine what the effect of long-term consumption of anthropogenic food sources, the possible shift in reproductive timing, along with various novel threats present in the urban environment, such as novel pathogens, predators and linear infrastructure will be, and how these might drive population dynamics and survival rates.
Although this review suggests possible outcomes for the species under current climate change predictions and urbanisation, it also highlights the need to conduct long-term research on galagos to better understand their response to human induced environmental change.
Data Availability
The datasets analyzed during the current study are available from the corresponding author upon reasonable request.
References
Acevedo-Whitehouse, K., & Duffus, A. L. J. (2009). Effects of environmental change on wildlife health. Philosophical Transactions of the Royal Society B: Biological Sciences, 364(1534), 3429–3438.
Antrop, M. (2004). Lanscape change and the urbanisation process in Europe. Landscape and Urban Planning, 67, 9–26.
Arroyo-Rodríguez, V., & Mandujano, S. (2006). Forest fragmentation modifies habitat quality for Alouatta palliata. International Journal of Primatology, 27(4), 1079–1096.
Arroyo-Rodríguez, V., & Mandujano, S. (2009). Conceptualization and measurement of habitat fragmentation from the primates’ perspective. International Journal of Primatology, 30(3), 497–514.
Bearder, S. (1999). Physical and social diversity among nocturnal primates: A new view based on long term research. Primates, 40(1), 267–282.
Bearder, S., Nekaris, K., & Curtis, D. (2006). A re-evaluation of the role of vision in the activity and communication of nocturnal primates. Folia Primatologica, 77(1–2), 50–71.
Bearder, S. K. (1987). Lorises, bushbabies, and tarsiers: Diverse societies in solitary foragers. In B. B. Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham, & T. T. Struhsaker (Eds.), Primate societies (pp. 11–24). University of Chicago Press.
Bearder, S. K. (2008). Lorises, bushbabies, and tarsiers: Diverse societies in solitary foragers. In B. S. Barbara, L. C. Dorothy, M. S. Robert & W. W. Richard (Eds.), Primate Societies (pp. 11–24). University of Chicago Press.
Bearder, S. K., & Doyle, G. A. (1974). Field and laboratory studies of social organization in bushbabies (Galago senegalensis). Journal of Human Evolution, 3(1), 37–50.
Bearder, S. K., Honess, P. E., & Ambrose, L. (1995). Species diversity among galagos with special reference to mate recognition. In L. Alterman, G. A. Doyle, & M. K. Izard (Eds.), Creatures of the dark: The nocturnal prosimians (pp. 331–352). Springer.
Bearder, S. K., & Martin, R. D. (1979). The social organization of a nocturnal primate revealed by radio tracking. In C. J. Amlaner & W. MacDonald (Eds.), A Handbook on biotelemetry and radio tracking (pp. 633–648). Pergamon.
Bearder, S. K., & Martin, R. D. (1980). Acacia gum and its use by bushbabies, Galago senegalensis (Primates: Lorisidae). International Journal of Primatology, 1, 103–128.
Bearder, S. K., Svensson, M., Butynski, T.M. & de Jong, Y.A. (2021). Galago moholi. The IUCN Red List of Threatened Species 2021: e.T8788A206563837.
Beehner, J. C., & Bergman, T. J. (2017). The next step for stress research in primates: To identify relationships between glucocorticoid secretion and fitness. Hormones and Behaviour, 91, 68–83.
Belluco, S., Losasso, C., Maggioletti, M., Alonzi, C. C., Paoletti, M. G., & Ricci, A. (2013). Edible insects in a food safety and nutritional perspective: A critical review. Comprehensive Reviews in Food Science and Food Safety, 12(3), 296–313.
Bernard, A. B., & Marshall, A. J. (2020). Assessing the state of knowledge of contemporary climate change and primates. Evolutionary Anthropology: Issues, News, and Reviews, 29(6), 317–331.
Bondarenco, A., Körtner, G., & Geiser, F. (2013). Some like it cold: Summer torpor by freetail bats in the Australian arid zone. Journal of Comparative Physiology B, 183(8), 1113–1122.
Boutin, S., & Lane, J. E. (2014). Climate change and mammals: Evolutionary versus plastic responses. Evolutionary Applications, 7(1), 29–41.
Breuner, C. W., & Hahn, T. P. (2003). Integrating stress physiology, environmental change, and behavior in free-living sparrows. Hormones and Behavior, 43(1), 115–123.
Bronson, F. H., & Heideman, P. D. (1994). Seasonal regulation of reproduction in mammals. In E. Knobil & J. D. Neill (Eds.), The physiology of reproduction (541–583). Raven Press.
Burnham, D., Bearder, S., Cheyne, S., Dunbar, R., & Macdonald, D. (2013). Predation by mammalian carnivores on nocturnal primates: Is the lack of evidence support for the effectiveness of nocturnality as an antipredator strategy? Folia Primatologica, 83(3–6), 236–251.
Cabana, F., Jayarajah, P., Oh, P. Y., & Hsu, C. D. (2018). Dietary management of a Hamadryas baboon (Papio hamadryas) troop to improve body and coat condition and reduce parasite burden: Dietary management of baboons. Journal of Zoo and Aquarium Research, 6(1), 16–21.
Campos, F. A., Morris, W. F., Alberts, S. C., Altmann, J., Brockman, D. K., Cords, M., et al. (2017). Does climate variability influence the demography of wild primates? Evidence from long-term life-history data in seven species. Global Change Biology, 23(11), 4907–4921.
Campos, F. A., Wikberg, E. C., & Holmes, T. D. (2012). Population assessment of Demidoff's dwarf galago (Galagoides demidovii) in a Ghanaian forest fragment mosaic. American Journal of Physical Anthropology (147), 642–653.
Cardoso, P., Barton, P. S., Birkhofer, K., Chichorro, F., Deacon, C., Fartmann, T., et al. (2020). Scientists’ warning to humanity on insect extinctions. Biological Conservation, 242, 108426.
Castillo-Contreras, R., Mentaberre, G., Fernandez Aguilar, X., Conejero, C., Colom-Cadena, A., Ráez-Bravo, A., González-Crespo, C., Espunyes, J., Lavín, S., & López-Olvera, J. R. (2021). Wild boar in the city: Phenotpic responses to urbanisation. Science of the Total Environment, 773, 145593.
Caton, J. M., Lawes, M., & Cunningham, C. (2000). Digestive strategy of the south-east African lesser bushbaby, Galago moholi. Comparative Biochemistry and Physiology A, 127, 39–48.
Chapman, C. A., Schoof, V. A., Bonnell, T. R., Gogarten, J. F., & Calmé, S. (2015). Competing pressures on populations: Long-term dynamics of food availability, food quality, disease, stress and animal abundance. Philosophical Transactions of the Royal Society b: Biological Sciences, 370(1669), 20140112.
Charmandari, E., Tsigos, C., & Chrousos, G. (2005). Endocrinology of the stress response. Annual Review of of Physiology, 67, 259–284.
Charpentier, M. J., Williams, C. V., & Drea, C. M. (2008). Inbreeding depression in ring-tailed lemurs (Lemur catta): Genetic diversity predicts parasitism, immunocompetence, and survivorship. Conservation Genetics, 9(6), 1605–1615.
Clarke, A., Rothery, P., & Isaac, N. J. (2010). Scaling of basal metabolic rate with body mass and temperature in mammals. Journal of Animal Ecology, 79(3), 610–619.
Colles, A., Liow, L. H., & Prinzing, A. (2009). Are specialists at risk under environmental change? Neoecological, paleoecological and phylogenetic approaches. Ecology Letters, 12(8), 849–863.
Cooper, C. E., McAllan, B. M., & Geiser, F. (2005). Effect of torpor on the water economy of an arid-zone marsupial, the stripe-faced dunnart (Sminthopsis macroura). Journal of Comparative Physiology B, 175(5), 323–328.
Cornelissen, T. (2010). Climate change and its effects on terrestrial insects and herbivory patterns. Neotropical Entomology, 40(2), 155–163.
Ćosić, N., Říčanová, Š, Bryja, J., Penezić, A., & Ćirović, D. (2013). Do rivers and human-induced habitat fragmentation affect genetic diversity and population structure of the European ground squirrel at the edge of its Pannonian range? Conservation Genetics, 14(2), 345–354.
Crespi, E. J., Williams, T. D., Jessop, T. S., & Delehanty, B. (2013). Life history and the ecology of stress: How do glucocorticoid hormones influence life-history variation in animals? Functional Ecology, 27, 93–106.
Cunneyworth, P. M., & Duke, J. (2020). Vehicle collisions among four species of monkeys between 2000 and 2018 on a suburban road in Diani, Kenya. International Journal of Primatology, 41(1), 45–60.
Cunneyworth, P. M., & Slade, A. M. (2021). Impact of electric shock and electrocution on populations of four monkey species in the suburban town of Diani, Kenya. International Journal of Primatology, 42(2), 171–186.
Dausmann, K. H., Nowack, J., Kobbe, S., & Mzilikazi, N. (2012). Afrotropical heterothermy: A continuum of possibilities. In T. Ruf, C. Bieber, W. Arnold, & E. Millesi (Eds.), Living in a seasonal world: Thermoregulatory and metabolic adaptations (pp. 13–27). Springer.
Davis-Reddy, C. L. & Vincent, K. (2017). Understanding climate risk over southern Africa. In C. L. Davis-Reddy & K. Vincent (Eds.), Climate Risk and vulnerability: A handbook for southern Africa (pp. 6-19). Council for Scientific and Industrial Research
De Andrade, A. C., Delledrate, F. L., & Mesquita, D. O. (2019). Assessing the effect of urbanisation on tropical forest dwelling teiid lizards. Ecological Indicators, 99, 225–229.
de Sassi, C., & Tylianakis, J. M. (2012). Climate change disproportionately increases herbivore over plant or parasitoid biomass. PLoS ONE, 7(7), e40557.
Devictor, V., Julliard, R., Couvet, D., Lee, A. & Jiguet, F. (2007). Functional homozegnization effect of urbanisation on bird communities. Conservation Biology, 21(3), 741–751.
Dietz, J. M., Baker, A. J., & Ballou, J. A. (2000). Demographic evidence of inbreeding depression in wild golden lion tamarins. In A. G. Young & G. M. Clarke (Eds.), Genetics, demography, and the viability of fragmented populations (pp. 203–211). Cambridge University Press.
Dobler, H.-J. (1976). Development of thermoregulation in the newborn lesser bushbaby (Galago senegalensis moholi, Smith 1839). Experientia, 32(2), 194–195.
Dobler, H. J. (1978). Untersuchungen über die Temperatur und Stoffwechselregulation von Galagos (Lorisiformes: Galagidae). Eberhard Karls-Universität.
Doyle, G. A., Andersson, A., & Bearder, S. K. (1971). Reproduction in the lesser bushbaby (Galago senegalensis moholi) under semi-natural conditions. Folia Primatologica, 14(1–2), 15–22.
Doyle, G. A., Pelletier, A., & Bakker, T. (1967). Courtship, mating and parturition in the lesser bushbaby (Galago senegalensis moholi) under semi-natural conditions. Folia Primatologica, 7(3–4), 169–197.
Ellison, G., Jones, M., Cain, B., & Bettridge, C. M. (2021). Taxonomic and geographic bias in 50 years of research on the behaviour and ecology of galagids. PLoS ONE, 16(12), e0261379.
Engelbrecht, D. (2016). Galagos as avian nest predators in South Africa. Primates, 57(4), 455–458.
Engelbrecht, C. J., Engelbrecht, F. A., & Dyson, L. L. (2013). High resolution model-projected changes in mid-tropospheric closed-lows and extreme rainfall events over southern Africa. International Journal of Climatology, 33(1), 173–187.
Estrada, A., Garber, P. A., Rylands, A. B., Roos, C., Fernandez-Duque, E., Di Fiore, A., et al. (2017). Impending extinction crisis of the world’s primates: Why primates matter. Science Advances, 3(1), e1600946.
Farias, I. P., Santos, W. G., Gordo, M., & Hrbek, T. (2015). Effects of Forest Fragmentation on Genetic Diversity of the Critically Endangered Primate, the Pied Tamarin (Saguinus bicolor): Implications for Conservation. Journal of Heredity, 106(S1), 512–521.
Fenoglio, M. S., Calvino, A., Gonzalez, E., Salvo, A., & Videla, M. (2021). Urbanisation drivers and underlying mechanisms of terrestrial insect diversity loss in cities. Ecological Entomology, 46(4), 757–771.
Fiore, A. D., & Rodman, P. S. (2001). Time allocation patterns of lowland woolly monkeys (Lagothrix lagotricha poeppigii) in a neotropical terra firma forest. International Journal of Primatology, 22(3), 449–480.
Foden, W. B., Butchart, S. H. M., Stuart, S. N., Vié, J.-C., Akçakaya, H. R., Angulo, A., et al. (2013). Identifying the world’s most climate change vulnerable species: A systematic trait-based assessment of all birds, amphibians and corals. PLoS ONE, 8(6), e65427.
Forbanka, D. N. (2018). Population surveys of fork-marked dwarf lemurs and needle-clawed galagos. Primates, 59(4), 355–360.
Frankham, R. (2005). Genetics and extinction. Biological Conservation, 126(2), 131–140.
Garnett, S., Franklin, D., Ehmke, G., VanderWal, J., Hodgson, L., Pavey, C., et al. (2013). Climate change adaptation strategies for Australian birds (Report). Charles Darwin University.
Gillson, L., Dawson, T. P., & Jack, S. (2013). Accommodating climate change contingencies in conservation strategy. Trend in Ecology and Evolution, 28(3), 135–142.
Giuliano, C. (2021). Chimpanzee (Pan troglodytes schweinfurthii) behavioral responses to resource scarcity in the savanna-woodland environment of Issa valley, Tanzania. Unpublished Doctor of Philosophy, Liverpool John Moores University, Liverpool, UK.
Gnanadesikan, G. E., Pearse, W. D., & Shaw, A. K. (2017). Evolution of mammalian migrations for refuge, breeding, and food. Ecology and Evolution, 7(15), 5891–5900.
Gonçalves, E. C., Ferrari, S. F., Silva, A., Coutinho, P. E. G., Menezes, E. V., & Schneider, M. P. C. (2003). Effects of habitat fragmentation on the genetic variability of silvery marmosets, Mico argentatus. In L. K. Marsh (Ed.), Primates in fragments: Ecology and conservation (pp. 17–28). Springer.
Grogan, K. E., Sauther, M. L., Cuozzo, F. P., & Drea, C. M. (2017). Genetic wealth, population health: Major histocompatibility complex variation in captive and wild ring-tailed lemurs (Lemur catta). Ecology and Evolution, 7(19), 7638–7649.
Grueter, C. C., Li, D., Ren, B., & Li, M. (2013). Overwintering strategy of Yunnan snub-nosed monkeys: Adjustments in activity scheduling and foraging patterns. Primates, 54(2), 125–135.
Guh, Y.-J., Tamai, T. K., & Yoshimura, T. (2019). The underlying mechanisms of vertebrate seasonal reproduction. Proceedings of the Japan Academy, Series B, 95(7), 343–357.
Güneralp, B., Lwasa, S., Masundire, H., Parnell, S., & Seto, K. C. (2017). Urbanization in Africa: Challenges and opportunities for conservation. Environmental Research Letters, 13(1), 015002.
Haji, M., & Bakuza, J. S. (2023). Climate change vulnerability assessment for the Rondo dwarf galago in coastal forests, Tanzania. Environmental Management, 71(1), 145-158.
Happold, D. (1995). The interactions between humans and mammals in Africa in relation to conservation: A review. Biodiversity and Conservation, 4(4), 395–414.
Harcourt, C. (1986). Seasonal variation in the diet of South African galagos. International Journal of Primatology, 7, 491–506.
Harcourt, C., & Bearder, S. (1989). A comparison of Galago moholi in South Africa with Galago zanzibaricus in Kenya. International Journal of Primatology, 10(1), 35–45.
Harris, T. R., Chapman, C. A., & Monfort, S. L. (2010). Small folivorous primate groups exhibit behavioral and physiological effects of food scarcity. Behavioral Ecology, 21(1), 46–56.
Higgins, S. I., & Scheiter, S. (2012). Atmospheric CO2 forces abrupt vegetation shifts locally, but not globally. Nature, 488(7410), 209–212.
Hinde, K., Skibiel, A. L., Foster, A. B., Del Rosso, L., Mendoza, S. P., & Capitanio, J. P. (2014). Cortisol in mother’s milk across lactation reflects maternal life history and predicts infant temperament. Behavioral Ecology, 26(1), 269–281.
Hing, S., Narayan, E. J., Thompson, R. C. A., & Godfrey, S. S. (2016). The relationship between physiological stress and wildlife disease: Consequences for health and conservation. Wildlife Research, 43, 51–60.
Honess, P. E., Perkin, A., & Bearder, S. K. (2008). Rondo Dwarf Galago. Folia Primatologica, 74, 337–354.
Huey, R. B., Kearney, M. R., Krockenberger, A., Holtum, J. A., Jess, M., & Williams, S. E. (2012). Predicting organismal vulnerability to climate warming: Roles of behaviour, physiology and adaptation. Philosophical Transactions of the Royal Society B: Biological Sciences, 367(1596), 1665–1679.
Isaac, J. L. (2009). Effects of climate change on life history: Implications for extinction risk in mammals. Endangered Species Research, 7(2), 115–123.
Isabirye-Basuta, G. M., & Lwanga, J. S. (2008). Primate populations and their interactions with changing habitats. International Journal of Primatology, 29(1), 35–48.
Izard, M. K., & Nash, L. T. (1988). Contrasting reproductive parameters in Galago senegalensis en G. s. moholi. International Journal of Primatology, 9(6), 519–527.
Izard, M. K., & Simons, E. L. (1987). Lactation and interbirth interval in the senegal galago (Galago senegalensis moholi). Journal of Medical Primatology, 16(5), 323–332.
Jayne, R. (2020). Gum. It’s what’s for dinner: the dietary significance and nutritional content of acacia gum for five sympatric primate species living in a high-altitude temperate environment. Unpublished Masterthesis, University of Colorado, Boulder, Colorado.
Jones, C. B. (2005). The costs and benefits of behavioral flexibility to inclusive fitness: Dispersal as an option in heterogeneous regimes. In C. B. Jones (Ed.), Behavioral flexibility in primates: Causes and consequences (pp. 17–29). Springer.
Kersey, D. C., & Dehnhard, M. (2014). The use of noninvasive and minimally invasive methods in endocrinology for threatened mammalian species conservation. General and Comparative Endocrinology, 203, 296–306.
Kettel, E. F., Gentle, L. K., Quinn, J. L., & Yarnell, R. W. (2018). Thebreeding performance of raptors in urban landscapes: A review and meta-analysis. Journal of Ornithology, 159(1), 1–18.
Kirby, R., Johnson, H. E., Alldredge, M. W., & Pauli, J. N. (2019). The cascading effects of human food on hibernation and cellular aging in free-ranging black bears. Scientific Reports, 9(1), 1–7.
Knox, C., & Wright, P. (1989). Thermoregulation and energy metabolism in the lesser bushbaby, Galago senegalensis moholi. African Zoology, 24(2), 89–94.
Koenig, A., Borries, C., Chalise, M. K., & Winkler, P. (1997). Ecology, nutrition, and timing of reproductive events in an Asian primate, the Hanuman langur (Presbytis entellus). Journal of Zoology, 243(2), 215–235.
Korstjens, A. H., & Hillyer, A. P. (2016). Primates and climate change: A review of current knowledge. In S. A. Wich & A. J. Marshall (Eds.), An introduction to primate conservation (pp. 175–192). Oxford University Press.
Kubelka, V., Sandercock, B. K., Székely, T., & Freckleton, R. P. (2022). Animal migration to northern latitudes: Environmental changes and increasing threats. Trends in Ecology and Evolution, 37(1), 30–41.
Lea, A. J., Altmann, J., Alberts, S. C., Tung, J., Associate Editor: Robert, D., & Editor: Judith, L. B. (2015). Developmental constraints in a wild primate. The American Naturalist, 185(6), 809–821.
Linden, B., Cuozzo, F. P., Sauther, M. L., & Jonker, W. C. (2022a). Impact of linear infrastructure on South Africa’s primate fauna: The need for mitigation. Folia Primatologica, 1, 1–19.
Linden, B., Dalton, D. L., Van Wyk, A., de Jager, D., Moodley, Y., & Taylor, P. J. (2022b). Potential drivers of samango monkey (Cercopithecus albogularis) population subdivision in a highly fragmented mountain landscape in northern South Africa. Primates, 63(3), 245–260.
Lyons, J., Mastromonaco, G., Edwards, D. B., & Schulte-Hostedde, A. I. (2017). Fat and happy in the city: Eastern chipmunks in urban environments. Behavioral Ecology, 28(6), 1464–1471.
Maibeche, Y., Moali, A., Yahi, N., & Menard, N. (2015). Is diet flexibility an adaptive life trait for relictual and peri-urban populations of the endangered primate Macaca sylvanus? PLoS ONE, 10(2), e0118596.
Markert, J. A., Champlin, D. M., Gutjahr-Gobell, R., Grear, J. S., Kuhn, A., McGreevy, T. J., et al. (2010). Population genetic diversity and fitness in multiple environments. BMC Evolutionary Biology, 10(1), 1–13.
Marsh, L. K. (2003). Genetics and population dynamics. In L. K. Marsh (Ed.), Primates in fragments (pp. 11–16). Springer.
Masters, J., Génin, F., Bearder, S. (2016). A conservation assessment of Galago moholi. In M. F. Child, L. Roxburgh, S. E. Do Linh, D. Raimondo, H. T. Davies-Mostert (Eds.) The red list of mammals of South Africa, Swaziland and Lesotho (pp. 1-6). South African National Biodiversity Institute and Endangered Wildlife Trust.
Mazaris, A. D., Kallimanis, A. S., Sgardelis, S. P., & Pantis, J. D. (2008). Do long-term changes in sea surface temperature at the breeding areas affect the breeding dates and reproduction performance of Mediterranean loggerhead turtles? Implications for climate change. Journal of Experimental Marine Biology and Ecology, 367(2), 219–226.
McKechnie, A. E., & Wolf, B. O. (2019). The physiology of heat tolerance in small endotherms. Physiology, 34(5), 302–313.
McNutt, J., Groom, R., & Woodroffe, R. (2019). Ambient temperature provides an adaptive explanation for seasonal reproduction in a tropical mammal. Journal of Zoology, 309(3), 153–160.
Meadows, M. E., & Hoffman, T. M. (2003). Land degradation and climate change in South Africa. Geographical Journal, 169(2), 168–177.
Meyer, A. L. S., & Pie, M. R. (2022). Climate change estimates surpass rates of climatic niche evolution in primates. International Journal of Primatology, 43(1), 40–56.
Mhemmed, G., Kamel, H., & Chedly, A. (2008). Does habitat fragmentation reduce genetic diversity and subpopulation connectivity? Ecography, 31(6), 751–756.
Mitchell, D., Fuller, A., & Maloney, S. K. (2009). Homeothermy and primate bipedalism: Is water shortage or solar radiation the main threat to baboon (Papio hamadryas) homeothermy? Journal of Human Evolution, 56(5), 439–446.
Monaghan, P., & Haussmann, M. F. (2015). The positive and negative consequences of stressors during early life. Early Human Development, 91(11), 643–647.
Moss, R., Oswald, J., & Baines, D. (2001). Climate change and breeding success: decline of the capercaillie in Scotland. Journal of Animal Ecology, 70(1), 47–61.
Moyes, K., NUSSEY, D. H., Clements, M. N., guinness, F. E., Morris, A., MORRIS, S., et al. (2011). Advancing breeding phenology in response to environmental change in a wild red deer population. Global Change Biology, 17(7), 2455–12469.
Murray, M. H., Becker, D. J., Hall, R. J., & Hernandez, S. M. (2016). Wildlife health and supplemental feeding: A review and management recommendations. Biological Conservation, 204, 163–174.
Mzilikazi, N., Masters, J. C., & Lovegrove, B. G. (2006). Lack of torpor in free-ranging southern lesser galagos, Galago moholi: Ecological and physiological considerations. Folia Primatologica, 77(6), 465–476.
Nakane, Y., & Yoshimura, T. (2014). Universality and diversity in the signal transduction pathway that regulates seasonal reproduction in vertebrates. Frontiers in Neuroscience, 8, 115.
Narango, D. L., Tallamy, D. W., & Marra, P. P. (2018). Nonnative plants reduce population growth of an insectivorous bird. Proceedings of the National Academy of Sciences, 115(45), 11549–11554.
Nash, L. T. (1986). Dietary, behavioral, and morphological aspects of gummivory in primates. American Journal of Physical Anthropology, 29, 113–137.
Nekaris, A., & Bearder, S. K. (2007). The lorisiform primates of Asia and mainland Africa: Diversity shrouded in darkness. In C. J. Campbell, A. Fuentes, K. C. Mackinnon, M. Panger, & S. K. Bearder (Eds.), Primates in perspective (pp. 24–45). Oxford University Press.
Neveu, H., Hafen, T., Zimmermann, E., & Rumpler, Y. (1998). Comparison of the genetic diversity of wild and captive groups of Microcebus murinus using the random amplified polymorphic DNA method. Folia Primatologica, 69, 127–135.
Nord, A., & Giroud, S. (2020). Lifelong effects of thermal challenges during development in birds and mammals. Frontiers in Physiology, 11, 1–9.
Nowack, J., Mzilikazi, N., & Dausmann, K. (2013a). Torpor as an emergency solution in Galago moholi: Heterothermy is triggered by different constraints. Journal of Comparative Physiology B, 183(4), 547–556.
Nowack, J., Mzilikazi, N., & Dausmann, K. H. (2023). Saving energy via shot and shallow torpor bouts. Journal of Thermal Biology, 114, 103572.
Nowack, J., Mzilikazi, N., & Dausmann, K. H. (2010). Torpor on demand: Heterothermy in the non-lemur primate Galago moholi. PLoS ONE, 5(5), e10797.
Nowack, J., Stawski, C., & Geiser, F. (2017). More functions of torpor and their roles in a changing world. Journal of Comparative Physiology B, 187(5), 889–897.
Nowack, J., Wippich, M., Mzilikazi, N., & Dausmann, K. H. (2013b). Surviving the cold, dry period in Africa: Behavioral adjustments as an alternative to heterothermy in the African lesser bushbaby (Galago moholi). International Journal of Primatology, 34, 49–64.
Omifolaji, J. K., Ikyaagba, E. T., Alarape, A. A., Ojo, V. A., Modu, M., Lateef, L. F., et al. (2020). Estimates of Demidoff's galago (Galagoides demidovii) density and abundance in a changing landscape in the Oban hills, Nigeria. Hystrix: The Italian Journal of Mammalogy, 31(2), 117–122.
Pacifici, M., Visconti, P., Butchart, S. H. M., Watson, J. E. M., Cassola, F. M., & Rondinini, C. (2017). Species’ traits influenced their response to recent climate change. Nature Climate Change, 7(3), 205–208.
Parnell, S., & Walawege, R. (2011). Sub-Saharan African urbanisation and global environmental change. Global Environmental Change, 21, 12–20.
Pauls, S. U., Nowak, C., Bálint, M., & Pfenninger, M. (2013). The impact of global climate change on genetic diversity in populations and species. Molecular Ecology, 22(4), 925–946.
Petelle, M. B., Dang, B. N., & Blumstein, D. T. (2017). The effect of maternal glucocorticoid levels on juvenile docility in yellow-bellied marmots. Hormones and Behavior, 89, 86–91.
Phukuntsi, M. A., Du Plessis, M., Dalton, D. L., Jansen, R., Cuozzo, F. P., Sauther, M. L., et al. (2020). Population genetic structure of the thick-tailed bushbaby (Otolemur crassicaudatus) from the Soutpansberg Mountain range, northern South Africa, based on four mitochondrial DNA regions. Mitochondrial DNA Part A, 31(1), 1–10.
Phukuntsi, M. A., Du Plessis, M., Dalton, D. L., Jansen, R., Sauther, M. L., Cuozzo, F. P., et al. (2021). Population and genetic structure of a male-dispersing strepsirrhine, Galago moholi (Primates, Galagidae), from northern South Africa, inferred from mitochondrial DNA. Primates, 62(4), 667–675.
Pimley, E. (2009). A survey of nocturnal primates (Strepsirrhini: Galaginae, Perodictinae) in southern Nigeria. African Journal of Ecology, 47(4), 784–787.
Power, M. L. (2010). Nutritional and digestive challenges to being a gum-feeding primate. In A. M. Burrows & L. T. Nash (Eds.), The Evolution of Exudativory in Primates (pp. 25–44). Springer New York.
Prober, S. M., Doerr, V. A. J., Broadhurst, L. M., Williams, K. J., & Dickson, F. (2019). Shifting the conservation paradigm: a synthesis of options for renovating nature under climate change. Ecological Monographs, 89(1), e01333.
Pullen, S. L., Bearder, S. K., & Dixson, A. F. (2000). Preliminary observations on sexual behavior and the mating system in free-ranging lesser galagos (Galago moholi). American Journal of Primatology, 51, 79–88.
Ray, I. S., Wren, B. T., & Bowers, E. (2017). Species density of Galago moholi at Loskop Dam Nature Reserve, South Africa. African Primates, 12, 1–8.
Ray, I. S., Wren, B. T., & Bowers, E. J. (2016). Documentation of plant consumption by Galago moholi in South Africa. African Primates, 11(1), 45–48.
Razgour, O., Forester, B., Taggart, J. B., Bekaert, M., Juste, J., Ibáñez, C., et al. (2019). Considering adaptive genetic variation in climate change vulnerability assessment reduces species range loss projections. Proceedings of the National Academy of Sciences, 116(21), 10418–10423.
Razgour, O., Juste, J., Ibáñez, C., Kiefer, A., Rebelo, H., Puechmaille, S. J., et al. (2013). The shaping of genetic variation in edge-of-range populations under past and future climate change. Ecology Letters, 16(10), 1258–1266.
Reher, S., & Dausmann, K. H. (2021). Tropical bats counter heat by combining torpor with adaptive hyperthermia. Proceedings of the Royal Society b: Biological Sciences, 288(1942), 20202059.
Rothman, J. M., Raubenheimer, D., Bryer, M. A., Takahashi, M., & Gilbert, C. C. (2014). Nutritional contributions of insects to primate diets: Implications for primate evolution. Journal of Human Evolution, 71, 59–69.
Sánchez-Bayo, F., & Wyckhuys, K. A. (2019). Worldwide decline of the entomofauna: A review of its drivers. Biological Conservation, 232, 8–27.
Sánchez-Bayo, F., & Wyckhuys, K. A. (2021). Further evidence for a global decline of the entomofauna. Austral Entomology, 60(1), 9–26.
Sapolsky, R. M., Romero, L. M., & Munck, A. U. (2000). How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocrine Reviews, 21(1), 55–89.
Scanes, C. G. (2018). Human activity and habitat loss: Destruction, fragmentation, and degradation. In C. G. Scanes & S. R. Toukhsati (Eds.), Animals and Human Society (pp. 451–482). Elsevier.
Scheiter, S., Gaillard, C., Martens, C., & Erasmus, B. F. N. (2018). How vulnerable are ecosystems in the Limpopo province to climate change. South African Journal of Botany, 116, 86–95.
Scheun, J., Bennett, N., Ganswindt, A., & Nowack, J. (2014). Spicing up the menu: Evidence of fruit feeding in Galago moholi. Primates, 55, 359–363.
Scheun, J., Bennett, N., Ganswindt, A., & Nowack, J. (2015). The hustle and bustle of city life: Monitoring the effects of urbanisation in the African lesser bushbaby. The Science of Nature, 102(9–10), 1–11.
Scheun, J., Bennett, N. C., Nowack, J., & Ganswindt, A. (2017). Reproductive behaviour, testis size and faecal androgen metabolite concentrations in the African lesser bushbaby. Journal of Zoology, 301(4), 263–270.
Scheun, J., Greeff, D., & Nowack, J. (2019). Urbanisation as an important driver of nocturnal primate sociality. Primates, 60(4), 375–381.
Scheun, J., Nowack, J., Bennett, N., & Ganswindt, A. (2016). Female reproductive activity and its endocrine correlates in the African lesser bushbaby, Galago moholi. Journal of Comparative Physiology B, 186(2), 255–264.
Schmid, J., & Speakman, J. R. (2009). Torpor and energetic consequences in free-ranging grey mouse lemurs (Microcebus murinus): A comparison of dry and wet forests. Naturwissenschaften, 96(5), 609–620.
Schulze, H., Doyle, G., Johann, A., Meier, B., Wirth, R., & Zimmermann, E. (1998). A ‘checklist’ of possible items for prosimian husbandry manuals and research. Folia Primatologica, 69, 152–170.
Skinner, J. D., & Smithers, R. H. N. (1990). Mammals of the Southern African Subregion: University of Pretoria.
Skogen, K., Helland, H., & Kaltenborn, B. (2018). Concern about climate change, biodiversity loss, habitat degradation and landscape change: Embedded in different packages of environmental concern? Journal for Nature Conservation, 44, 12–20.
Somero, G. (2010). The physiology of climate change: How potentials for acclimatization and genetic adaptation will determine ‘winners’ and ‘losers.’ Journal of Experimental Biology, 213(6), 912–920.
Soulsbury, C. D., & White, C. L. (2015). Human–wildlife interactions in urban areas: A review of conflicts, benefits and opportunities. Wildlife Research, 42(7), 541–553.
Sousa, C., Casanova, C., & Fernandes, C. (2008). Contribution of molecular data to the study of non-human primates. Advances in Molecular Biology and Its Applications to Biological Anthropology, 115, 115–166.
Strandin, T., Babayan, S. A., & Forbes, K. M. (2018). Reviewing the effects of food provisioning on wildlife immunity. Philosophical Transactions of the Royal Society B: Biological Sciences, 373(1745), 20170088.
Sunday, J. M., Bates, A. E., & Dulvy, N. K. (2012). Thermal tolerance and the global redistribution of animals. Nature Climate Change, 2(9), 686–690.
Svensson, M. S. (2017). Conservation and ecology of nocturnal primates: night monkeys, galagos, pottos and Angwantibos as case studies. Oxford Brookes University.
Thatcher, H. R., Downs, C. T. & Koyama, N. C. (2023). Primates in the urban mosaic: terminology, flexibility and management. In T. McKinney, Waters, S. & Rodrigues, M. A. (Eds.), Primates in anthropogenic landscapes (pp. 121–137). Springer.
Tigas, L. A., Van Vuren, D. H., & Sauvajot, R. M. (2002). Behavioral responses of bobcats and coyotes to habitat fragmentation and corridors in an urban environment. Biological Conservation, 108(2), 299–306.
Tait, J., Perotto-Baldivieso, H. L., McKeown, A., & Westcott, D. A. (2014). Are flying-foxes coming to town? Urbanisation of the spectacled flycing-fox (Pteropus conspicillatus) in Australia. PLoS ONE, 9(10), e109810.
Taylor-Brown, A., Booth, R., Gillet, A., Mealy, E., Ogbourne, S. M., & Polkinghorne, A. (2019). The impact of human activities on Australian wildlife. PLoS ONE, 14(1), e0206958.
Teixeira, B., Hirsch, A., Goulart, V. D. L. R., Passos, L., Teixeira, C. P., James, P., et al. (2015). Good neighbours: Distribution of black-tufted marmoset (Callithrix penicillata) in an urban environment. Wildlife Research, 42(7), 579–589.
Terrien, J., Blanc, S., Zizzari, P., Epelbaum, J., & Aujard, F. (2011). Physiological responses to chronic heat exposure in an aging non-human primate species, the gray mouse lemur (Microcebus murinus). Experimental Gerontology, 46(9), 747–754.
Tewksbury, J. J., Huey, R. B., & Deutsch, C. A. (2008). Putting the heat on tropical animals. Science, 320(5881), 1296–1297.
Thuiller, W. (2007). Climate change and the ecologist. Nature, 448(7153), 550–552.
Todd, K. R. (2020). Influence of maternal aggression and stress response on offspring quality in tree swallows. Unpublished Bachelor of Science Appalachian State University, Boone, North Carolina, USA.
Tomotani, B. M., Gienapp, P., Beersma, D. G. M., & Visser, M. E. (2016). Climate change relaxes the time constraints for late-born offspring in a long-distance migrant. Proceedings of the Royal Society b: Biological Sciences, 283(1839), 20161366.
Tomotani, B. M., van der Jeugd, H., Gienapp, P., de la Hera, I., Pilzecker, J., Teichmann, C., et al. (2018). Climate change leads to differential shifts in the timing of annual cycle stages in a migratory bird. Global Change Biology, 24(2), 823–835.
Toro, M., & Caballero, A. (2005). Characterization and conservation of genetic diversity in subdivided populations. Philosophical Transactions of the Royal Society B: Biological Sciences, 360(1459), 1367–1378.
Turner, J. M. (2020). Facultative hyperthermia during a heatwave delays injurious dehydration of an arboreal marsupial. Journal of Experimental Biology, 223(5), jeb219378.
Turok, I., & Borel-Saladin, J. (2014). Is urbanisation in South Africa on a sustainable trajectory? Development Southern Africa, 31(5), 675–691.
Urban, M. C. (2015). Accelerating extinction risk from climate change. Science, 348(6234), 571–573.
Visser, M. E., Both, C., & Lambrechts, M. M. (2004). Global climate change leads to mistimed avian reproduction. Advances in Ecological Research, 35, 89–110.
Wang, W., Qiao, Y., Li, S., Pan, W., & Yao, M. (2017). Low genetic diversity and strong population structure shaped by anthropogenic habitat fragmentation in a critically endangered primate, Trachypithecus leucocephalus. Heredity, 118(6), 542–553.
Wegge, P., & Rolstad, J. (2017). Climate change and bird reproduction: Warmer springs benefit breeding success in boreal forest grouse. Proceedings of the Royal Society B: Biological Sciences, 284(1866), 20171528.
Weisenseel, K., Chapman, C. A., & Chapman, L. J. (1993). Nocturnal primates of Kibale Forest: Effects of selective logging on prosimian densities. Primates, 34(4), 445–450.
Whirledge, S., & Cidlowski, J. A. (2017). Glucocorticoids and reproduction: Traffic control on the road to reproduction. Trends in Endocrinology & Metabolism, 28(6), 399–415.
Willi, Y., & Hoffmann, A. A. (2009). Demographic factors and genetic variation influence population persistence under environmental change. Journal of Evolutionary Biology, 22(1), 124–133.
Williams, C. M., Henry, H. A., & Sinclair, B. J. (2015). Cold truths: How winter drives responses of terrestrial organisms to climate change. Biological Reviews, 90(1), 214–235.
Wingfield, J. C., Hunt, K., Breuner, C., Dunlap, K., Fowler, G. S., Freed, L., et al. (1997). Environmental stress, field endocrinology, and conservation biology. In J. R. Clemmons & R. Bucholz (Eds.), Behavioral approaches to conservation in the wild (pp. 95–131). Cambridge University Press.
Wimberger, K., Nowak, K., & Hill, R. A. (2017). Reliance on exotic plants by two groups of threatened samango monkeys, Cercopithecus albogularis labiatus, at their southern range limit. International Journal of Primatology, 38, 151–171.
Wolf, A., Kozlov, M. V., & Callaghan, T. V. (2008). Impact of non-outbreak insect damage on vegetation in northern Europe will be greater than expected during a changing climate. Climatic Change, 87(1), 91–106.
Zhao, X., Ren, B., Li, D., Garber, P. A., Zhu, P., Xiang, Z., et al. (2019). Climate change, grazing, and collecting accelerate habitat contraction in an endangered primate. Biological Conservation, 231, 88–97.
Ziervogel, G., New, M., Archer van Garderen, E., Midgley, G., Taylor, A., Hamann, R., et al. (2014). Climate change impacts and adaptation in South Africa. Wiley Interdisciplinary Reviews: Climate Change, 5(5), 605–620.
Acknowledgements
We would like to thank the Lajuma Research Centre and the South African Environmental Observation Network (SAEON) for providing the climate data used in this review.
Funding
Open access funding provided by Tshwane University of Technology.
Author information
Authors and Affiliations
Contributions
Both JS and JN designed, researched, and wrote the review manuscript.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Inclusion and Diversity Statement
While citing references scientifically relevant for this work, we also actively worked to promote gender balance in our reference list.
The author list includes contributors from the location where the research was conducted, who participated in study conception, study design, data collection, analysis, and/or interpretation of the findings.
Additional information
Handling Editor: Michelle Sauther
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Scheun, J., Nowack, J. Looking Ahead: Predicting the Possible Ecological and Physiological Response of Galago Moholi to Environmental Change. Int J Primatol (2023). https://doi.org/10.1007/s10764-023-00373-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10764-023-00373-8