
Abstract
The structure of many aquatic food webs is influenced by tadpole survivorship and duration of seasonal occupancy, which depend on tadpole diet. Pollen has been occasionally reported to occur in tadpole diets, and the available studies show that tadpoles have a strong preference for this supposedly nutritive resource. Thus, we aimed to test whether a diet composed of pollen from varied plant species would boost tadpole growth, development, and agility/mobility, compared to a regular artificial balanced and nutritive diet. Tadpoles in initial developmental stages 25 and 30 grew more when fed with pollen, compared to artificial food and a mixture of both diets. Those initially in stage 30 advanced more developmental stages during one month when fed exclusively with pollen. Tadpoles fed with pollen moved longer distances in response to an aversive stimulus simulating a predatory attack than tadpoles fed with artificial food or a mixed diet. Our results corroborate the hypotheses of pollen being a very important food source to aquatic consumers such as tadpoles, being likely to influence the structure of aquatic food webs through tadpole nutrition and modulation of growth/development speed, as well as tadpole ability to escape predators.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Amphibian larvae are key to many aquatic food webs (e.g., O’Neil & Thorp, 2014; Schmidt et al., 2017) and their patterns of food consumption, growth, and development influence other trophic levels (Costa & Vonesh, 2013; Hite et al., 2018). Predation on anurans in all life stages can trigger a trophic cascade effect leading to increase in populations of their prey (Hite et al., 2018), whereas their seasonal role as aquatic consumers can increase complexity of food webs (Schmidt et al., 2017).
The larval phase is key for growth and development of most amphibians (Heyer, 1979; Wells, 2007) and its duration is affected by many factors such as nutrition, temperature, species interactions, and environmental features (Montaña et al., 2019). Diet composition can be considered a key factor, as rates of food consumption directly influence size and time to metamorphosis (Kupferberg, 1997). Anuran larvae usually adjust their growth to environmental conditions such as resource levels. They can grow until they reach a minimum size to metamorphose and leave an unfavorable aquatic habitat or grow to a maximum size if the aquatic habitat is favorable (Wilbur & Collins, 1973).
Although tadpoles are abundant and diverse in many aquatic ecosystems (Altig & McDiarmid, 1999), their diets, trophic ecology, and trophic level are poorly known (Montaña et al., 2019) compared to other freshwater consumers such as fish (Udoh & Ekpo, 2017). A variety of items are reported to occur in tadpole diets, including algae, fungi, protozoan, and zooplankton (Altig et al., 2007; Jacobson et al., 2017). There are not many records of pollen as a food item for vertebrates in general (Pryor, 2014), but pollen has been recorded in the diets of a few tadpole species (e.g., Wagner, 1986), including several tropical species at the montane meadows of southeastern Brazil (Kloh et al., 2018, 2019, 2021a). Additionally, Scinax machadoi (Bokermann & Sazima, 1973) tadpoles were shown to vertically swim through the water column to actively feed on pollen even when presented with other more easily accessible food items (Kloh et al., 2021b).
In this study, we investigate the effects of diets containing pollen on growth, development, and escape distance of tadpoles of Scinax machadoi, compared to a nutritive and balanced artificial diet likely to contain the nutrients obtained by them in natural habitats. We have previously shown that S. machadoi tadpoles show preference and promptness to consume pollen grains when available, even in the presence of alternative nutritive food (Kloh et al., 2019, 2021b). Thus, we first hypothesized pollen-fed tadpoles to present faster growth and development, corroborating the optimal foraging theory (MacArthur & Pianka, 1966), according to which consumers would be selected to choose the food items that provide the best nutritional gain with the lowest acquisition costs. Considering that tadpole burst swimming ability can be determinant for their escape from predators and survivorship (Álvarez & Nicieza, 2009), we also expected tadpoles fed with pollen to cover greater distances when trying to escape from an aversive stimulus, as a surrogate for higher energy availability.
Methods
Study site
We collected tadpoles from a third-order stream (sensu Strahler, 1957) called Água Escura (19° 16′ 3.35″ S, 43° 30′ 56.23″ W; 1236 m alt.) within the Doce River basin at the Parque Nacional (National Park) Serra do Cipó (PNSC). The PNSC is inserted in the southern portion of the Espinhaço Mountain Range, an important area for species conservation in Minas Gerais state (Eterovick et al., 2020) due to high levels of endemism, including amphibians (Leite et al., 2008). The climate has marked rainy (October–March) and dry (April–September) seasons, with mean monthly temperatures varying between 13 and 29 °C and mean annual rainfall, between 1460 and 2490 mm (Viveros, 2010). Montane meadows (Campos Rupestres) are the predominant vegetation, with Asteraceae, Fabaceae, Bromeliaceae, Orchidaceae, Eriocaulaceae, Xyridaceae, Melastomataceae, Lentibulariaceae, and Velloziaceae as the most representative families (Giulietti et al., 1997; Machado et al., 2007).
Model species
Scinax machadoi is a common species that breeds year-round in permanent streams of the PNSC. Tadpoles take about five months to metamorphose (Eterovick et al., 2020). They have ovoid bodies, ventral mouth, and dorsal eyes (Pimenta et al., 2014), benthic habits, and are active both during the day and at night (Eterovick et al., 2020). They have a dark brown body with two bright yellow dorsal bars (Eterovick et al., 2018). Their diet includes unicellular and filamentous Zygnematophyceae and Bacillariophyceae, Tecamoeba, Cyanophyceae, and pollen from varied plant families (Kloh et al., 2019; pers. obs.).
Sampling procedures
We used 30 tadpoles in each of the stages 25 and 30 (sensu Gosner, 1960), that were collected by a single person with dipnets at a stream backwater on 14th January 2020. After collection, tadpoles were placed in containers with tap water rested for 24 h to eliminate chlorine. They remained in these containers for 12 h to eliminate previously ingested gut contents. We made previous observations of gut clearance in tadpoles of Scinax machadoi. They can produce a good amount of feces within 35 min and at the end of a 12 h-period they don’t produce any more (i.e., the gut is emptied).
Food manipulation experiment
After tadpoles eliminated previously ingested gut contents, they were placed individually in 8 × 8 × 9.5 cm containers with 300 ml of rested tap water for food treatments. Tadpoles of each stage (25 or 30) were randomly distributed in three treatments (10 tadpoles each), represented by diets composed of (1) pollen, (2) commercial fish food (intended to simulate the variety of nutrients contained in a diversified diet naturally consumed by tadpoles), and (3) a mixture of both pollen and fish food, for the duration of the 30-day experiment.
For treatments containing pollen as food, we used commercial bee pollen (Santa Bárbara®) from Apis mellifera Linnaeus, 1758 produced at the region according to commercial regulations, because not many plants were flowering at the study site at the time of the experiment. This pollen consists of a mixture containing pollen from flowers from different plant families collected by bees, with a very small quantity of nectar and bee salivary secretions. It contains a variety of proteins, lipids, sugars, fibers, mineral salts, aminoacids, and vitamins (Brasil, 2000). We characterized pollen composition as follows. We first dehydrated the whole content of the commercialized container (500 g) at 45 °C for two hours. Posteriorly we mixed the content to homogenize the grains and selected subsamples using the method proposed by Zenebon et al. (2008) for quantification of solid food samples. In order to obtain a random smaller sample, we distributed the whole content on a squared flat surface divided in four equal sized quadrats and separated the contents of two diagonal quadrats. The process was then repeated three times with the separated subsample. The final subsample obtained with this procedure was treated with acids to remove the cytoplasmatic content of pollen grains as described by Erdtman (1960) to make pollen morphology more visible. We then examined the material on slides under an optic microscope (Olympus BX50) with 400 × to 1000 × amplification. The relative frequency of pollen grains from different plants families was determined based on counting of at least 1500 grains (sensu Louveaux et al., 1978) in systematically positioned microscopic fields of view to guarantee a homogeneous examination of the slide. We identified 18 different pollen types belonging to 15 plant families by comparison with reference slides from the collection of Fundação Ezequiel Dias, Belo Horizonte, Brazil, and by consulting specific literature, plus eight non-identified types (Table 1; Fig. 1).

Pollen grain spectra of the commercial bee pollen from Apis mellifera showing A Melastomataceae pollen type and B Eucalyptus, Myrtaceae pollen type (arrows). Pictures made under microscope at ×400 magnification, during pollen quantification procedures
For the treatments with artificial food, we used commercial fish food (Probetta Show®), that has a rich nutrient content, aiming to simulate a combination of nutrients likely to be supplied by the mixture of natural items consumed by tadpoles (Kloh et al., 2019). Probetta Show® contains mollusk, crustacean compounds, meat and fish derivatives, vegetal subproducts, yeast, oils, minerals, vitamins, and antioxidants, according to manufacturer specifications. Although the real nutritional composition of Scinax machadoi tadpoles’ diet is not known, we conservatively aimed for a nutritive commercial food, as colleagues have successfully used this and similar fish commercial foods for maintenance of tadpoles of varied species until metamorphosis (C. Vinicius, C. Lisboa, I. F. Machado, T. Kohlsdorf, pers. comm.). Even if this food has a higher nutritional value than the mixture of items in the natural diet of tadpoles, it would then underestimate the difference among treatments, reinforcing possible significant results.
Each tadpole received standardized 36 mg portions (weighed to the nearest 0.1 mg) of macerated food (pollen, fish food, or mixture of both at equal proportions) every three days. The water of the containers was replaced immediately before food delivery to keep them clean. Tadpoles had their total length (TL), body length (BL), body width (BW), tail length (TaL), and tail height (TH; sensu Altig & McDiarmid, 1999) measured with digital calipers (to the nearest 0.1 mm) at the beginning and at the end of the experiment to obtain their increase in each measurement (given by the division of the final measurement by the initial one). We also recorded developmental stages of all tadpoles at the end of the experiment, to access through how many stages they developed toward metamorphosis starting from their initial stages (25 or 30). During the experiment, the containers were kept in a chamber under a 14 h light/10 h dark regime at 21 °C. Water temperature in the containers varied between 16 and 18 °C. We chose values within the mean temperature range at the study site to avoid high temperatures that could increase activity and energy expenditure, as well as very low temperatures, that could make tadpoles lethargic and less prone to eat (Maciel & Juncá, 2009).
Escape tests
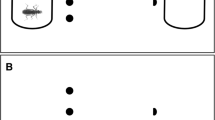
By the end of the experiment, we randomly picked five tadpoles of each stage/diet treatment to perform escape tests. These tests were based on tadpole response to an artificial disturbance simulating a predator attack, as in Eterovick et al. (2018). Tadpoles randomly assigned to escape tests were placed individually at 20 × 25 × 7 cm containers with a grid of 20 5-cm-cells marked and identified with letters at the bottom. Each container was filled with 500 ml rested tap water. Tadpoles were left in the container for two minutes and then subject to an aversive stimulus represented by an approaching stick, simulating a predator attack (Eterovick et al., 2018; Fig. 2). The stick was inserted by the same person (J. S. Kloh) slowly and at a constant speed toward the head of the tadpole until it moved or until the stick touched its head. As fleeing tadpoles moved short distances with a single, direct swimming movement, the initial and final points of the trajectory could be easily identified in the marked grid and the covered distance was immediately measured with a ruler. We are aware that this procedure can include some bias in the exact position of the tadpole within a grid cell, for this reason all measurements were conducted by the same person to avoid observer effects. The order of the tadpoles to be tested was also shuffled and tests were made blind. The observer had previous experience with this type of test, being trained to record the results with precision. The side to where the tadpole escaped was also recorded to detect any laterality and its potential influence on fleeing distance.

Schematic representation of agility experiments conducted with tadpoles of Scinax machadoi in stages 25 and 30 subject to three different diets (pollen, fish food, mixture of pollen and fish food)
Statistical analyses
To compare measurement gains of tadpoles in stages 25 and 30 subject to three different diets (1) pollen, (2) fish food, and (3) a mixture of both, we used Non-Metric Multidimensional Scaling (NMDS). NMDS allowed us to represent measurement gains (given by the final measurement divided by the initial measurement) in a single multidimensional space to show not only growth but also potential shape variations under different diets. We used metaMDS function in the package MASS (Venables & Ripley, 2002) for R (R Core Team, 2020). We used the same data to conduct a PERMANOVA in the package Vegan (Oksanen et al., 2016) using diet as explanatory variable for growth/shape changes. We performed separated analyses for tadpoles that started the experiment in stage 25 and in stage 30.
We tested whether initial developmental stage (25 or 30), diet, and their interactions explained number of developmental stages advanced by tadpoles during the experiment (starting from either stage 25 or 30) and whether initial developmental stage, diet, or laterality and their interactions explained distance covered by fleeing tadpoles using Generalized Linear Models (GLMs). We used the package MuMIn (Bartón, 2022) to select the best model to explain fleeing distance based on AICc values and the package emmeans (Lenth, 2022) for post hoc tests.
Results
Tadpoles at both initial stages (25 or 30) grew more when fed exclusively with pollen than tadpoles fed a mixed diet or fish food (Fig. 3; Table 2). The contributions of all measurements were important to distribute experimental groups in the multidimensional space (Fig. 3; Table 3). The spatial distribution of the groups indicated that the variation among treatments was mainly in length measurements (Fig. 3).

Distribution of tadpoles of Scinax machadoi with initial stages 25 (A) and 30 (B) from different experimental groups (tadpoles fed with pollen, fish food, or both) in a multidimensional space according to NMDS (Non-Metric Multidimensional Scaling) representing gain in body measurements (vectors). The contribution of each vector to the two axes is seen in Table 3
We used the complete model (and did not perform model selection) to explain number of developmental stages advanced by tadpoles during the experiment because both variables, diet (Deviance = 129.0; df = 2, 56; P < 0.001) and initial developmental stage (Deviance = 123.3; df = 1, 58; P < 0.001), as well as their interaction (Deviance = 41.0; df = 2, 54; P = 0.006) were significant. Tadpoles beginning the experiment at stage 30 developed faster when fed with pollen than the other diets, whereas diet did not influence the developmental rate of tadpoles that started the experiment at stage 25 (Fig. 4; Table 4).

Number of developmental stages advanced by tadpoles with initial stages 25 and 30 (sensu Gosner, 1960) after three different diet treatments (pollen, fish food, mixture of pollen and fish food). Different letters represent treatments/initial stages that differed significantly (P < 0.05)
The best model to explain fleeing distance included developmental stage at the beginning of the trials (Deviance = 101.2; df = 1, 28; P = 0.001) and diet (Deviance = 137.6; df = 2, 26; P < 0.001), without interactions (df = 5, logLik = − 74.32, AICc = 161.15, weight = 0.88). The next best model included the same variables (developmental stage and diet) but also their interaction (df = 7, logLik = − 73.03, AICc = 165.14, delta = 4, weight = 0.12). Tadpoles that started the trials in stage 25 always covered longer distances than tadpoles that started the trials in stage 30 within the same diet treatment. Tadpoles that started the trials in either stage 25 or 30 covered significantly longer distances when fed with pollen than tadpoles fed only with fish food. Tadpoles fed with mixed diets had intermediate values (Fig. 5; Table 4). Most tadpoles moved to the left, but laterality did not influence tadpole performance regarding fleeing distance.

Fleeing distances covered by tadpoles with initial stages 25 and 30 after three different diet treatments (pollen, fish food, mixture of pollen and fish food). Different letters represent treatments/initial stages that differed significantly (P < 0.05)
Discussion
Our experiments corroborated the high nutritional value of pollen, considered to be rich in vitamins, mineral salts, lipids, and proteins (Roulston & Cane, 2000), for tadpole growth and development. Tadpoles grew more when fed with only pollen compared to a nutritive and balanced commercial food and showed an intermediate response to a mixture of both diets (pollen and commercial food), no matter whether they started the experiment at developmental stage 25 or 30. Tadpoles starting the experiment at stage 25 did not differ in development among food treatments. However, tadpoles that started in stage 30 reached more advanced developmental stages when fed with pollen compared to the other diets, reinforcing the value of pollen not only for growth, but also for development in the later stages before metamorphosis. Diet composition is extremely important for tadpole metamorphosis (Montaña et al., 2019), which is influenced by diversity of consumed food items and their protein, carbohydrate, and lipidic proportions (Kupferberg, 1997; Montaña et al., 2019). A well-balanced nutrition is important to trigger hormonal changes that favor larval development (Kupferberg, 1997), like the regulation of thyroid hormones, responsible for a broad body structure remodeling (Brown & Cai, 2007).
Tadpoles that started the experiment in stage 25 always covered longer distances while fleeing compared to tadpoles that started in stage 30 under the same diet treatment. It is possible that a more advanced developmental stage of hind limbs and larger size reached by tadpoles that started the experiment in stage 30 would have hampered mobility. A greater susceptibility to predation could also explain a more pronounced fleeing response in vulnerable (smaller) tadpoles (Bateman & Fleming, 2015). The main predators at the study site are aquatic insects such as water bugs and Odonata naiads (Espanha et al., 2015). At least in some instances, smaller tadpoles were shown to present lower survivorship when exposed to such predators (Jara & Perotti, 2010), what remains to be tested specifically for S. machadoi. However, when tadpoles were compared among diet treatments, they covered longer distances when fed with pollen compared to the commercial food, likely also indicating higher energy available due to better quality food. The behavior of Scinax machadoi tadpoles corroborates the high energetic content of pollen, as they change their feeding behavior, swimming more actively to reach pollen grains at the water surface, despite their usual benthic habits (Kloh et al., 2021b). According to the optimal foraging theory (MacArthur & Pianka, 1966), only a high benefit provided by pollen would justify this energy investment for its acquisition. Even with no influence in fleeing distance, the laterality observed in this study, with most of the tadpoles turning to the left to escape, is an interesting record. Laterality consists in a tendency to use preferentially one side of the body due to structural asymmetries of the nervous system (Stancher et al., 2018) and has been recorded for some vertebrate groups (Stancher et al., 2018; Goree & Wassersug, 2001), including adult amphibians (Wassersug et al., 1999; Goree & Wassersug, 2001). For tadpoles, however, there is still little information available about laterality (Wassersug et al., 1999). The preferential use of the left side by most Scinax machadoi tadpoles had already been recorded for feeding behavior (Kloh et al., 2021b) and is now reinforced by escape behavior, with no signs of influence in tadpole performance.
Our study corroborated a superior performance of tadpoles fed with pollen regarding growth, development, and agility/mobility. In this scenario, pollen input could be considered an important nutrient source for aquatic food webs, reinforcing the importance of preserving riparian vegetation for amphibian conservation. Such habitats not only offer shelter and favorable conditions for amphibian movement among breeding and resting sites (Semlitsch & Bodie, 2003), but also the flowering plants that provide tadpoles and likely other consumers with pollen showed here to be a valuable food item. Our results open many interesting questions to be addressed in the future, like how tadpoles adjust pollen consumption to seasonal variations in pollen availability, how efficient are tadpoles of different species regarding pollen consumption and assimilation, how different species are benefited (or not) by increased pollen consumption, to mention just a few. For example, we have already demonstrated that different species vary in pollen consumption and digestion capacity under natural conditions (Kloh et al., 2023).
An impoverished diet can result in slower tadpole development (Kupferberg, 1997) with implications for the food web due to longer tadpole permanence in the aquatic habitat (Hite et al., 2018). Tadpole agility to escape predators can also influence survivorship (Van Buskirk & McCollum, 2000) and reflect in the food web (Hite et al., 2018), as predation is an important mortality source controlling tadpole abundance (Wells, 2007). Our results indicate that pollen should not be neglected as an important resource in the study of aquatic food webs involving tadpoles, as well as the importance of the riparian vegetation for nutrition of aquatic consumers.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Altig, R. & R. W. McDiarmid, 1999. Diversity: familial and generic characterizations. In McDiarmid, R. W. & R. Altig (eds), Tadpoles – The Biology of Anuran Larvae University of Chicago Press, Chicago, IL: 295–337.
Altig, R., R. Matt & C. L. Taylor, 2007. What do tadpoles really eat? Assessing the trophic status of an understudied and imperiled group of consumers in freshwater habitats. Freshwater Biology 52: 386–395.
Álvarez, D. & A. G. Nicieza, 2009. Differential success of prey escaping predators: tadpole vulnerability or predator selection? Copeia 2009: 453–457.
Bartoń, K., 2022. MuMIn: Multi-Model Inference. R package version 1.46.0. https://CRAN.R-project.org/package=MuMIn
Bateman, P. W. & P. A. Fleming, 2015. Body size and group size of Cuban tree frog (Osteopilus septentrionalis) tadpoles influence their escape behaviour. Acta Ethologica 18: 161–166.
Bokermann, W. C. A. & I. Sazima, 1973. Anfíbios da Serra do Cipó, Minas Gerais, Brasil. II: Duas espécies novas de Hyla (Anura, Hylidae). Revista Brasileira De Biologia 33: 521–528.
Brasil, 2000. Instrução normativa n° 11, de 20 de outubro de 2000. Regulamento Técnico de identidade e qualidade do mel. Diário Oficial [da] República Federativa do Brasil.
Brown, D. D. & L. Cai, 2007. Amphibian metamorphosis. Developmental Biology 306: 20–33.
Costa, Z. J. & J. R. Vonesh, 2013. Interspecific differences in the direct and indirect effects of two neotropical hylid tadpoles on primary producers and zooplankton. Biotropica 45: 503–510.
Erdtman, G., 1960. The acetolysis method-a revised description. Svensk Botanisk Tidskrift 54: 516–564.
Espanha, J., M. F. Vasconcelos & P. C. Eterovick, 2015. The role of tadpole coloration against visually oriented predators. Behavioral Ecology and Sociobiology 70: 255–267. https://doi.org/10.1007/s00265-015-2044-4.
Eterovick, P. C., I. S. Mendes, J. S. Kloh, L. T. Pinheiro, A. B. H. P. Václav, T. Santos & A. S. B. Gontijo, 2018. Tadpoles respond to background colour under threat. Scientific Reports 8: 4085. https://doi.org/10.1038/s41598-018-22315-8.
Eterovick, P. C., A. M. Souza & I. Sazima, 2020. Anuran Amphibians from Serra do Cipó. Gráfion Estúdio Editorial, Belo Horizonte, Brazil.
Giulietti, A. M., J. R. Pirani & R. M. Harley, 1997. Espinhaço Range region, eastern Brazil. In Davis, S. D., V. H. Heywood, O. Herrera-MacBryde, J. Villa-lobos & A. C. Hamilton (eds), Centres of Plant Diversity: a Guide and Strategy for their Conservation, Vol. 3. WWF/IUCN, Cambridge: 397–404. The Americas.
Goree, B. D. & R. J. Wassersug, 2001. Are archeobatrachian anurans ambidextrous? Assessing handedness in Bombina orientalis. Journal of Herpetology 35: 538–541.
Gosner, K. L., 1960. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 16: 183–190.
Heyer, W. R., 1979. Annual variation in larval amphibian populations within a temperate pond. Journal of the Washington Academy of Sciences 69: 65–74.
Hite, J. L., M. C. Hughey, K. M. Warkentin & J. R. Vonesh, 2018. Cross-ecosystem effects of terrestrial predators link treefrogs, zooplankton, and aquatic primary production. Ecosphere 9(9): e02377. https://doi.org/10.1002/ecs2.2377.
Jacobson, B. J., A. Cervantes-Martínez & M. A. Gutierrez-Aguirre, 2017. Selectivity of Incilius valliceps (Anura: Bufonidae) tadpoles on freshwater zooplankton. Hidrobiológica 27: 211–217.
Jara, F. G. & M. G. Perotti, 2010. Risk of predation and behavioural response in three anuran species: influence of tadpole size and predator type. Hydrobiologia 644: 313–324.
Kloh, J. S., C. C. Figueredo & P. C. Eterovick, 2018. You are what, where, and when you eat: seasonal and ontogenetic changes in a tropical tadpole’s diet. Amphibia-Reptilia 39: 445–456.
Kloh, J. S., C. C. Figueredo & P. C. Eterovick, 2019. How close is microhabitat and diet association in aquatic ecomorphotypes? A test with tadpoles of syntopic species. Hydrobiologia 828: 271–285.
Kloh, J. S., C. C. Figueredo & P. C. Eterovick, 2021a. No choice is not a choice: food selectivity occurs in different ontogenetic stages of Phasmahyla jandaia (Anura: Phyllomedusidae). Salamandra 57: 513–519.
Kloh, J. S., C. C. Figueredo & P. C. Eterovick, 2021b. Tadpole dance: the gymnastics of Ololygon machadoi larvae to feed on pollen. Salamandra 57: 89–95.
Kloh, J. S., C. C. Figueredo, D. B. Provete & P. C. Eterovick, 2023. Taste for pollen comes in different shapes: Consumption by tadpoles from three divergent ecomorphotypes. Journal of Zoology 320: 42–52.
Kupferberg, S. J., 1997. The role of larval diet in anuran metamorphosis. American Zoologist 37: 146–159.
Leite, F. S. F., F. A. Juncá & P. C. Eterovick, 2008. Status do conhecimento, endemismo e conservação de anfíbios anuros da Cadeia do Espinhaço, Brasil. Megadiversidade 4: 182–200.
Lenth, R. V., 2022. emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.7.2. https://CRAN.R-project.org/package=emmeans
Louveaux, J., A. Maurizio & G. Vorwohl, 1978. Methods of melissopalynology. Bee World 59: 139–157.
MacArthur, R. H. & E. R. Pianka, 1966. On optimal use of a patchy environment. American Naturalist 100: 603–609.
Machado, C. G., A. G. Coelho, C. S. Santana & M. Rodrigues, 2007. Beija-flores e seus recursos florais em uma área de campo rupestre da Chapada Diamantina, Bahia. Revista Brasileira De Ornitologia 15: 215–227.
Maciel, T. A. & F. A. Juncá, 2009. Effects of temperature and volume of water on the growth and development of tadpoles of Pleurodema diplolister and Rhinella granulosa (Amphibia: Anura). Zoologia 26: 413–418.
Montaña, C. G., S. D. G. T. M. Silva, D. Hagyari, J. Wager, L. Tiegs, C. Sadeghian, T. A. Schriever & C. M. Schalk, 2019. Revisiting “what do tadpoles really eat?” A 10-year perspective. Freshwater Biology 64: 2269–2282.
O’Neill, B. J. & J. H. Thorp, 2014. Untangling food-web structure in an ephemeral ecosystem. Freshwater Biology 59: 1462–1473.
Oksanen J., F. G. Blanchet, M. Friendly, R. Kindt, P. Legendre, D. McGlinn, P. R. Minchin, R. B. O’Hara, G. L. Simpson, P. Solymos, M. H. H. Stevens, E. Szoecs & H. Wagner, 2016. vegan: Community Ecology Package. R package version 2.4-1. https://CRAN.R-project.org/package=vegan.
Pimenta, B. V. S, D. Costa, R. Murta-Fonseca & T. Pezutti, 2014. Anfíbios: Alvorada de Minas, Conceição do Mato Dentro, Dom Joaquim - Minas Gerais. Bicho do Mato, Belo Horizonte, Brazil.
Pryor, G., 2014. Tadpole nutritional ecology and digestive physiology: Implications for captive rearing of larval anurans. Zoo Biology 33: 502–507.
R Core Team, 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Version 3.6.3, Vienna, Austria. https://cran.r-project.org/
Roulston, T. H. & J. H. Cane, 2000. Pollen nutritional content and digestibility for animals. Plant Systematics and Evolution 222: 187–209.
Schmidt, K., M. L. Blanchette, R. G. Pearson, R. A. Alford & A. M. Davis, 2017. Trophic roles of tadpoles in tropical Australian streams. Freshwater Biology 62: 1929–1941.
Semlitsch, R. D. & J. R. Bodie, 2003. Biological criteria for buffer zones around wetlands and riparian habitats for amphibians and reptiles. Conservation Biology 17: 1219–1228.
Stancher, G., A. V. Sourano & G. Vallortigara, 2018. Chapter 2 – Motor asymmetries in fisher, amphibians, and reptiles. Progress in Brain Research 238: 33–56.
Strahler, A. N., 1957. Quantitative analysis of watershed geomorphology. Transactions of the American Geophysical Union 38: 913–920.
Udoh, J. P. & I. E. Ekpo, 2017. Diet-morphology relationships of some fish species in the cross-river estuary, Southeast Nigeria. International Journal of Fisheries and Aquaculture Research 3: 10–29.
Van Buskirk, J. V. & S. A. McCollum, 2000. Functional mechanisms of an inducible defence in tadpoles: morphology and behaviour influence mortality risk from predation. Journal of Evolutionary Biology 13: 336–347. https://doi.org/10.1046/j.1420-9101.2000.00173.x.
Venables, W. N. & B. D. Ripley, 2002. Modern Applied Statistics with S, 4th ed. Springer, New York:
Viveros, R. S., 2010. Pteridófitas da Serra do Caraça, Minas Gerais, Brasil. Master Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil.
Wagner, W. E., 1986. Tadpoles and pollen: observations on the feeding behavior of Hyla regilla larvae. Copeia 1986: 802–804.
Wassersug, R. J., T. Naitoh & M. Yamashita, 1999. Turning bias in tadpoles. Journal of Herpetology 33: 543–548.
Wells, K. D., 2007. The Ecology and Behavior of Amphibians, The University of Chicago Press, Chicago, IL:
Wilbur, H. M. & J. P. Collins, 1973. Ecological aspects of amphibian metamorphosis. Science 182: 1305–1314.
Zenebon, O., N. S. Pascuet & P. Tiglea (ccords.), 2008. Normas Analíticas do Instituto Adolfo Lutz. Métodos físico-químicos para análise de alimentos. Instituto Adolfo Lutz, São Paulo, Brazil.
Acknowledgements
We are thankful to Fundação Ezequiel Dias (FUNED) for technical support in pollen analyses, the Parque Nacional da Serra do Cipó for support during the field trip, the Universidade Federal de Minas Gerais for infra-structure. JSK and PCE are thankful to Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for a Scholarship and a Research Productivity grant, respectively. Collection permits were provided by Sisbio/ICMBio (64500).
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
JSK was responsible for the study conception and design. Material preparation, data collection, and analysis were performed by all authors. The first draft of the manuscript was written by JSK and PCE, and the other authors commented on all versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Additional information
Handling editor: Lee B. Kats
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kloh, J.S., Figueredo, C.C., Calaça, P. et al. Pollen as food: effects of consumption on tadpole growth, development, and mobility. Hydrobiologia 851, 2071–2080 (2024). https://doi.org/10.1007/s10750-023-05439-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-023-05439-5