
Abstract
Silica-scaled chrysophytes are a unique group of protists representing a major component of numerous aquatic habitats where they can significantly contribute to human well-being in many ways. In the present review, altogether 14 ecosystem services provided by silica-scaled chrysophytes were identified highlighting their importance and roles in aquatic ecosystems. The most important supporting services are related to primary production, nutrient- and biogeochemical cycling, as well as sediment formation. Additionally, they are involved in the regulation of climate, maintenance of air quality, biological control and water purification processes (as regulating services). The most relevant provisioning services include their usefulness in environmental monitoring (paleolimnology, ecological status assessment and climate modelling), production of biochemicals and provision of genetic resources. Cultural services are associated with aesthetic values, inspiration source and knowledge system as well. Although silica-scaled chrysophytes are one of the least studied groups of algae, here they are shown to provide important benefits and to have many further powerful opportunities for humans, thus their detailed research in the future is emphasised as an important key for human well-being.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Chrysophytes (belonging to Eukaryota (superkingdom), Stramenopiles (clade), Ochrophyta (phylum), Chrysophyceae (class), Kristiansen & Škaloud, 2017; Schoch et al., 2020) are known mainly as unicellular or colonial golden-brown algae, which may be flagellated, but there are also some multicellular species (van den Hoek et al., 1995). Chrysophyte algae have many genetically and morphologically distinct lineages, and among this large group there are many species with siliceous coverage. These siliceous structures place them among the silicifiers, a very important group of aquatic organisms (Preisig, 1994). Silicifiers use dissolved Si (dSi) to precipitate biogenic silica (bSi; SiO2) as internal- and/or external structures (Moriceau et al., 2019). Phototrophic silicifiers play an important role in the sequestration of atmospheric CO2 in the ocean, and therefore in long term global climate regulation (web1). Biogeochemically, traditionally and ecologically, diatoms are believed to be the most important silicifiers in modern marine ecosystems, along with radiolarians, silicoflagellates and siliceous sponges (Sutton et al., 2018) while some less-known groups like Bolidophyceae (< 1% in marine phytoplankton) and chrysophytes should also be considered for completeness. The origin, early evolution and key processes for the acquisition of the silica cell wall have not been fully understood and these small groups could play a key role in enhance our understanding of the Si-cycle, silicification processes and the role of silicifiers in ecosystems (Moriceau et al., 2019). For instance, bolidophytes have a hypothetical life cycle that switches between silicified non-flagellated and naked flagellate stages (Kuwata et al., 2018). Despite the hypothetical facultative siliceous skeleton usage in several silifiers, in this review we focus on the scaled chrysophytes, because their silica ultrastructure enabled scientists to elaborate and the best-developed taxonomic system for species differentiation among protists, moreover because their studies are the most consistent and reliable compared to other microalgae within chrysophytes (Kristiansen, 2005; Škaloud et al., 2013a).
Classification of silica-scaled chrysophytes has undergone many changes since 1878 to present (reviewed by Škaloud et al., 2013a). Currently they are a part of the class Chrysophyceae including Chrysosphaerella, Lepidochromonas, Mallomonas, Neotessella, Paraphysomonas, Polylepidomonas, Spiniferomonas and Synura (Kristiansen & Preisig, 2007; Kristiansen & Škaloud, 2016; Kapustin & Guiry, 2019; Guiry & Guiry, 2022) (Fig. 1). As a group, silica-scaled chrysophytes display a great variability in their features: their size varies from nano- to micrometers; they can have unicellar or colonial forms; usually they bear biogenic silica structures (scale, spine, bristle) in different numbers and have flagella. They are nutritionally opportunistic (autotrophic, mixotrophic and heterotrophic, Table 1) and they have no enzymes for the utilisation of bicarbonate and produce only acid phosphatases (Sandgren, 1988; Sandgren et al., 1995; Kristiansen & Škaloud, 2017). Approximately 350 taxa of silica-scaled chrysophytes are described (Guiry & Guiry, 2022), which is highly variably distributed among the genera (Table 1). However, the above number certainly does not reflect their real species richness, which has been, at all probability, underestimated. Despite their best-developed taxonomic system among protists, their differentiation at species level carries difficulties. For instance, the diversity issues based only on morphology are problematic due to their various morphotypes with significant genetic variation that requires extensive molecular support (see also Gusev et al., 2018; Siver et al., 2018), which exist only for the section Petersenianae of Synura (Škaloud et al., 2014; Jo et al., 2016), some clades of Mallomonas (Jo et al., 2013; Čertnerová et al., 2019), and Paraphysomonas (Scoble & Cavalier-Smith, 2014). Next-generation sequencing (NGS) technologies have the potential to reveal their diversity and ecosystem dynamics in a more precise way (e.g. Bock et al., 2014; Škaloud et al., 2020). However, their estimated species number is continuously changing year by year due to the newly described species from both recent and fossil samples (e.g. Barreto, 2001; Wujek et al., 2003; Kim & Soon Kim, 2008; Němcová et al., 2011; Němcová & Kreidlova, 2013; Gusev et al., 2016; Gusev & Siver, 2017; Siver, 2018; Kapustin & Gusev, 2019; Němcová et al., 2022).

Transmission electron microscope (TEM) microphotograph of some of the characteristic silica-scaled chrysophytes found in forest lakes (Hungary). Silica scale of Mallomonas intermedia Kisselev (a), M. insignis Penard (b), M. formosa S. Barreto (c), scale (d) and bristle (e) of M. heterospina J. W. G. Lund, M. tubulosa S. Barreto (f), Chrysosphaerella brevispina Korshikov (g), Synura echinulata Korshikov (h), S. spinosa Korshikov (i), S. uvella Ehrenberg (j), Neotessella lapponica (Skuja) B. Y. Jo, J. I. Kim, W. Shin, P. Škaloud & P. A. Siver (k), Spiniferomonas trioralis E. Takahashi (l), Lepidochromonas eiffelii (H. A. Thomsen) Kapustin & Guiry (m), Paraphysomonas vestita (A. Stokes) De Saedeleer (n). Microscale bar: 5 µm (a-k, n) and 2 µm (l-m)
The acceleration of aquatic ecosystem degradation along with a rapid and continuous decline in the Earth’s biodiversity (Lampert & Sommer, 2007; Irfan & Alatawi, 2019) initiated a cascade of other environmental changes among them reduction in ecosystem services and functions (e.g. Schläpfer & Schmid, 1999; Loreau et al., 2001). Since biodiversity can stabilise ecosystem functions and services (e.g. Worm et al., 2006; Hooper et al., 2012), its investigation and maintenance is a primary and urgent task for our present and future (Irfan & Alatawi, 2019). Despite the fact that many aquatic habitats are among the most endangered ecosystems in the world and play significant role in global context (Tréguer et al., 2018), the study of aquatic organisms and freshwater biodiversity hotspots has received much less attention than that of terrestrial or marine ecosystems (Myers et al., 2000). Regional species richness of silica-scaled chrysophytes varies widely in different habitats from a very few species (< 20 taxa, Karjalainen et al., 1996; Gusev, 2013) through diverse communities (e.g. 80–90 taxa, Němcová et al., 2016; Bessudova et al., 2021) up to extraordinary cases, which are considered as chrysophytes hotspots (Aquitaine, Baikal Region, Northern Vietnam, Scandinavia, Třeboňsko, Řezáčová & Neustupa, 2007; Němcová et al., 2011; Korkonen et al., 2020; Olefeld et al., 2020; Gusev et al., 2021). However, it must be noted that the number of registered species in individual habitats is highly dependent on the intensity of floristic surveys, sampling strategy and the motivation/knowledge of experts.
Silica-scaled chrysophytes represent a pretty much neglected algal group compared to others (e.g. diatoms). Still, they are unique organisms and their contribution to many ecosystem services (supporting, regulating, provisioning, cultural, MEA, 2003, 2005), can be great. These are reviewed and summarised in the present paper (Table 2) emphasising their importance and highlighting the need for their detailed research in the future.
Supporting services
Primary production
Chrysophytes are important components of phytoplankton biomass (usually around 10–75% of the total biomass) in small oligotrophic or dystrophic ecosystems (with low conductivity and nutrient levels, neutral or slightly acidic pH, low to moderate dissolved humic substances, Sandgren, 1988; Kristiansen, 2005; Fig. 2), but many silica-scaled species are able to develop substantial population contributing even more than 85% of the total phytoplankton biomass (e.g. Synura uvella Ehrenberg, Kalinowska & Grabowska, 2016; Mallomonas caudata (Iwanoff) Ivanov, Hoffmann & Wille, 1992). These algae have been also described from mesotrophic and eutrophic lakes (with ~ 5–20% contribution to total biomass Sandgren, 1988) and temporary ditches (Péterfi et al., 1998). Though most of them are freshwater species, some of them are marine (Jeong et al., 2019) or capable of surviving in brackish environments (Němcová et al., 2016). They are not obligatory planktic algae; several taxa are abundant in benthic habitats, especially in mires and bogs (Douglas & Smol, 1995).

Typical habitats of silica-scaled chrysophytes situated in Hungary (a, forest lake in Pilis mountain) and in Romania (b, mountain lakes in South Carpathians)
Many silica-scaled chrysophytes are alternatively photoautotrophic, mixotrophic or heterotrophic (Table 1), whilst only Mallomonas, Neotessella and Synura are considered to be strict photoautotrophs (Andersen, 1987; Bhatti & Colman, 2005). The photosynthetic activity of phytoplankton has significant contribution to the global net primary production and atmospheric oxygen (reviewed by Naselli-Flores & Padisák, 2022). In comparison with the photosynthetic rate of the total phytoplankton communities (920–1866 µmol O2 mg Chl−1 h−1 or 45–1575 µmol C mg Chl–1 h–1, Martínez, 1980; Vincent, 1983; Williams et al., 1983; Guildford et al., 2007), photoautotrophic silica-scaled chrysophytes alone can have significant photosynthetic evolution rate (see examples in Table 3).
A remarkable feature of silica-scaled chrysophytes is their seasonality: some species appear in high amounts in the cold season (e.g. oxbow lakes, Kiss & Kristiansen, 1994; Péterfi et al., 2001) when photosynthesis of other algal groups is quite limited thus maintaining sufficient oxygen supply at ecosystem level. Since global ecosystem models predict a decline in the dissolved oxygen inventory leading to serious consequences (Schmidtko et al., 2017), the autotrophic and mixotrophic silica-scaled species with their photosynthetic activity may also have an important role in supporting ecosystem services.
Nutrient cycle
As primary producers, silica-scaled chrysophytes serve as fuel for food webs. Depending on the trophic state of the aquatic ecosystem (Carney, 1990), the main groups of zooplankton (rotifers, copepods, cladocerans) feeding on chrysophytes, and planktivorous fishes may also indirectly affect their abundance (reviewed by Sandgren et al., 1995). Furthermore, heterotrophic nanoflagellates, such as Paraphysomonas may serve as prey for larger protists and metazoa (e.g. Sieburth & Davies, 1982; Sherr & Sherr, 1994). Despite silica-scaled chrysophytes have some physiological strategies in order diminishing grazing pressure (e.g. large size, colony forming, extruding spines, bristles, recruitment from the sediment; Sandgren et al., 1995; Hansson & Tranvik, 1996), Daphnia, Bosmina, Pseudoboechella, Boeckella especially prefer them (Lehman & Sandgren, 1985; Sandgren et al., 1995; Hansson & Tranvik, 1996; Karjalainen et al., 1996; Arvola & Salonen, 2001; Taipale et al., 2019), which can alter ecosystem properties such as primary productivity and nutrient cycling (Carpenter et al., 1985). For instance, Mallomonas species can be very important primary producers, since they can account for 27% assimilated resources of Daphnia (Taipale et al., 2019). Thus, silica-scaled chrysophytes can contribute to the cycle of micro- and macronutrients through the food web. However, their biomass and composition, consequently their roles in ecosystem services, will be changed by altered top-down control, due to a shift to smaller zooplankton sizes (Moore et al., 1996; Strecker et al., 2004) and altered grazing dynamics induced by global warming.
Besides the pelagic food web, the microbial loop also has an important role in ecosystem productivity (Sandgren et al., 1995), as well as in the biogeochemical cycles, since under some environmental scenarios the 40–60% of the carbon fixed by the phytoplankton is considered to flux through the heterotrophic bacterial community (Cole et al., 1988). Chrysophytes are also known to be major elements of the microbial loop in many aquatic ecosystems (Scoble & Cavalier-Smith, 2013) and their roles could be multifaceted due to their diverse nutrition types. For instance, extracellular release of dissolved organic carbon compounds by silica-scaled chrysophytes such as Mallomonas caudata can temporarily function as an important nutrient source for the heterotrophic bacterial community (Blaauboer et al., 1982). Apparently, bacteria can utilise algal excretion products, whilst zooplankton feeding on bacteria fuel a considerable amount of carbon flow. Furthermore, the colourless phagotrophic species (Paraphysomonas imperforata Takahashi) can feed on bacteria as well as on other phytoplankton species resulting in the complication of the food web, loss of energy and organic matter (Goldman & Caron, 1985; Goldman & Dennett, 1990). Paraphysomonas may play a key role in transferring essential compounds (such as fatty acids, sterols) upgrading the food quality and improving carbon transfer efficiency as intermediary protists from primary producers to metazoan consumers (Park et al., 2003; Bec et al., 2006, 2010), moreover in the regeneration of nitrogen (Goldman & Caron, 1985; Dolan, 1997) and phosphorus (Andersen et al., 1986; Dolan, 1997). Finally, mixotrophic chrysophytes (e.g. Chrysosphaerella) have a competitive advantage in nutrient-poor and low-light environments (Cen et al., 2020) contributing more than 50% of the total phytoplankton biomass (Olrik, 1998), which is likely to advance under climate change (Vad et al., 2020). Their advantages are based on the fact that they can photosynthesize and also ingest bacteria or elements of phytoplankton for fulfilling their carbon, nitrogen and phosphorus demands (Goldman & Dennett, 1992; Unrein et al., 2007; Charvet et al., 2012; Terrado et al., 2017). Their phagotrophy might be supported by the flagella of the cells to move towards the prey (Sandgren et al., 1995).
As a closely related subject to mixotrophy, food-web studies involving chrysophytes are one of the most inspiring headings, both in freshwater and marine environments (Kristiansen & Škaloud, 2017; Vad et al., 2021; Cormier et al., 2022). Moreover, as an exciting supplement of nutrition mode studies, the molecular basis of nutritional diversification, accompanying pathway reduction and degradation of plastid structures help to understand the evolution of heterotrophy in chrysophytes (Beisser et al., 2017; Graupner et al., 2018; Kim et al., 2020).
Biogeochemical cycles
The most biotically widespread biomineral is silica (SiO2·nH2O), being present in all eukaryotic supergroups (Marron et al., 2016; Hendry et al., 2018). This trait (silicon biomineralization) of algae can be found in Chrysophyceae, Dictyochophyceae, Bacillariophyceae, Bolidophyceae and Xanthophyceae among some others (Hoek et al., 1995; Graham et al., 2000; Kuwata et al., 2018). Their extensive use of Si (Marron et al., 2016) in aquatic ecosystems is a key driver of the global Si-cycle, but each clade can have different contributions to it (e.g. Sandgren et al., 1995). Despite the fact that the ancient gene of silicic acid transport protein in chrysophycean algae have already been developed long before diatoms appeared (Likhoshway et al., 2008), chrysophytes seem to play only a minor part in the global silica cycle, contrary to diatoms (Sandgren et al., 1995). This is attributable to several reasons: (i) silica-scaled chrysophytes have lower silicon content per cell and per unit surface than diatoms (Sandgren et al., 1995), and (ii) macroevolution of their morphology depends on the cost and benefits of silicification since scale formation can be achieved with minimal use of silica, and some silica-scaled chrysophytes (e.g. Synura and Paraphysomonas) can produce even completely scale-less, naked cells (Sandgren et al., 1995). Despite biogenic silica production is provided mainly by diatoms and the contribution of chrysophytes is minimal in the marine environments, they can be locally important sources of biogenic silica procession (Reid et al., 1991; Sandgren et al., 1995; Simpson & Volcani, 2012), and their contribution can be higher in freshwaters than in the sea (Officer & Ryther, 1980; Sandgren et al., 1995; Street-Perrott & Barker, 2008). In the future, climate change is predicted to influence marine phytoplankton (Riebesell, 2004), even to enhance the abundance of chrysophytes (Feng et al., 2009), which consequently may affect their role in the global Si-cycling.
Sediment formation
The Si coverage is characteristic for vegetative cells, but silica-scaled flagellates also form siliceous stomatocysts as resting stages (Sandgren et al., 1995). The silica structures of chrysophytes can dissolve in the water column leading to the re-cycling of silicon. In marine environments, the dissolution of silica structures is almost complete, and only 2% or less can reach the sediment surface where it can be preserved in fossil deposits and may become lithified to form rocks. In freshwaters, this percentage could be higher (Leadbeater, 1986), but it depends on a number of environmental variables (reviewed by Sandgren et al., 1995). Furthermore, silica particles can be grazed by zooplankton which can also accelerate descent to the benthos through the faecal pellets (Sandgren et al., 1995). Chrysophytes remains can be deposited in great abundance and preserve well in certain types of lacustrine sediments where they contribute significantly to sedimentary biosilica content (Fig. 3). Chrysopel is the name of the sediment type that is extremely rich in Chrysophytes microfossils. Furthermore, stomatocysts can act as a microreactor in which the conditions favour fossil pyritization leading to the formation of iron sulphides (Bogush et al., 2013; Zhang et al., 2016).

Chrysopel: lacustrine sediment where chrysophytes were deposited in a great amount (C: cysts, D: diatoms). Lake Gales (South Carpathians, Romania, 178 cm depth in Gales3 core, Buczkó et al., 2018)
Regulating services
Air quality maintenance
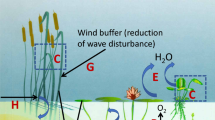
Phytoplankton is known to contribute significantly to the absorption of global annual CO2 emission (about 20–35%, Khatiwala et al., 2009; Naselli-Flores & Padisák, 2022). Autotrophic silica-scaled chrysophytes have special physiological characteristics, such as lacking CO2 concentrating mechanisms (CCM) and external carbonic anhydrase (CA). These features make them successful in from oligo- to slightly eutrophic ecosystems associated with low pH where they can be typical elements of phytoplankton and even may dominate. In such ecosystems, they have an important role in the global absorption process by passive diffusion of CO2 (Saxby-Rouen et al., 1998; Bhatti & Colman, 2005, 2008; Maberly et al., 2009).
Though the oxygen exchange between air and water surface is a slow process and depends on many parameters (e.g. temperature, partial pressures; Wetzel, 2001), silica-scaled chrysophytes can contribute to the oxygen content of the air by the production of significant-amount oxygen as a result of their photosynthetic activity (see details in supporting services).
Climate regulation
Chrysophytes can take part directly in climate regulation via Si-cycle. Silicon is considered as a bioessential element, and the silica cycle plays an important role in the sequestration of carbon by the ocean’s biological pump, and in long term global climate regulation (Tréguer et al., 2021). The global oceanic Si-cycle has evolved through geologic time primarily due to the uptake of dissolved Si and subsequent biomineralization by silicifiers. Biogeochemically, traditionally and ecologically, diatoms are considered to be the most important silicifiers in contemporal marine ecosystems along with radiolarians, silicoflagellates and siliceous sponges (Sutton et al., 2018), whilst some less-known group like Bolidophyceae (< 1% in marine phytoplankton) and chrysophytes must also be taken into consideration.
Biological control
None of the silica-scaled chrysophyte species are considered as toxin-producing or HAB (Harmful Algal Bloom) species, but they can produce and release metabolites affecting many ecosystem services. First to mention are volatile organic compounds (VOCs), which are considered to be beneficial or essential to many food web processes (Arts et al., 2001). VOCs can pose negative effects on grazers resulting in modification of food web dynamics (Jüttner, 2001) and the consequent nutrient cycling. They may have allelopathic effects on other algal species or antimicrobial and/or fungicidal activities (reviewed by Watson et al., 2001b). Further and deeper research of these allelopathic effects may be a critical step in the field of water management, since the natural algicides and biomaterials provide good alternatives of chemical treatments (e.g. Lee et al., 2017). Such allelopathic interaction was shown for Synura, which inhibited Chrysochromulina and prevented its toxic blooms (Sandgren et al., 1995). Synura uvella produces a large number of excreted compounds (e.g. alcohols, ketones, aldehydes, and non-carotenoids, Jüttner, 1981) some of which can influence growth of other algae like Dolichospermum, Synechococcus, Nannochloris and Cyanidium (Jüttner, 1979). Furthermore, heterotrophic nanoflagellates, such as Paraphysomonas, have an important role in controlling the populations of toxic Microcystis aeruginosa (Kützing) Kützing by its effective grazing (Yan et al., 2009).
However, vica versa, there are also a number of known cases that algicides were shown to be effective against silica-scaled chrysophytes. For example, the dinoflagellate Peridinium aciculiferum Lemmermann, barley (Hordeum vulgare Linné) and rice straw extract negatively impacted Synura petersenii (Ferrier et al., 2005; Rengefors & Legrand, 2007; Rajabi Islami, 2010). These interspecific interactions offer a good method for mitigating water supply problems caused by chrysophytes, such as taste and odour events (see more details in the chapter of cultural services).
Water purification
Algae provide powerful techniques for bioremediation purposes (Kaplan et al., 1988; Sen et al., 2013). Among silica-scaled chrysophytes, Synura is shown to produce exopolysaccharide compounds that complex copper and lead thus reducing their availability and toxicity for other aquatic organisms (Lombardi & Vieira, 1998). Paraphysomonas can be effective in the biologically mediated regeneration and recycling process of Fe, Zn, Th and Mn playing a role in their biogeochemical cycles (Hutchins & Bruland, 1994; Barbeau et al., 2001). Synura petersenii and Mallomonas caudata have high capacity for removal of nitrogen and phosphorus, which can be useful in water treatment management (Stevčić et al., 2019).
Seas, oceans and coastal zones are highly susceptible to pollution by crude oil (such as petroleum), which represents a serious threat for the biota (McGenity et al., 2012). The biodegradation of crude oil to carbon dioxide and water is the major process in the remediation of the polluted environments, which can be significantly affected by silica-scaled chrysophytes. The dynamics of phagotrophic chrysophytes, such as Paraphysomonas foraminifera I.A.N. Lucas, should be taken into account, since it can effectively grow and graze on hydrocarbonoclastic bacterial cells in crude-oil polluted environments (Kormas et al., 2007; Dalby et al., 2008). This relation can have an effect on the biodegradation of oil pollution, but future works are highly needed to identify the real role of these eukaryotes in bioremediation processes and their usage in water management practices.
Provisioning services
Environmental monitoring
Though environmental monitoring as ecosystem service is not mentioned by the MEA (2003, 2005), silica-scaled chrysophytes can be directly used to indicate environmental conditions, which could be a very useful and important feature. Therefore, environmental monitoring was additionally considered here as a further possible provisioning service such as it was concluded and used by others, too (e.g. Vaughn, 2018; Thomaz, 2021; B-Béres et al., 2022).
Micro-organisms are suggested to have distributional patterns similar to found for higher plants or animals: local/regional, ecological (climatic), historical conditions (Foissner, 2006). Regarding the local/regional patterns, a variety of environmental variables can be driving factors in the development and distribution of silica-scaled chrysophytes, such as geographical distance, anthropogenic pollutants (e.g. petroleum products, surfactants), pH, conductivity, trophic state, temperature, transparency, colour, concentration of carbon dioxide, magnesium ions and total iron (e.g. Sandgren et al., 1995; Kristiansen, 2005; Bessudova et al., 2021). Many silica-scaled chrysophytes have characteristic occurrence ranges or narrow ecological demand and are sensitive to a variety of environmental factors (Siver & Hamer, 1992; Siver & Smol, 1993; Siver, 2012), which make them excellent bioindicators for monitoring ecological status of freshwaters (Sandgren et al., 1995; Sagova-Mareckova et al., 2021). Thus, on the basis of the compositional changes of their assemblages the ecological status and its changes can be deduced (Sandgren et al., 1995; Němcová et al., 2016), but even the appearance/disappearance of one species alone can have indicator value (Hickel & Maass, 1989; Sandgren et al., 1995; Gensemer & Playle, 1999; Kristiansen, 2005). They can also be ideal organisms for evaluating the effects of different environmental stressors, like industrial pollutants (e.g. aluminium or silver ions from everyday consumer products, antibacterial medical applications, herbicides), which enter the aquatic food-webs, effect its architecture by bioaccumulation, and consequently the stability of the ecosystem (Dixit et al., 1989; Bock et al., 2014). For instance, Mallomonas sp. was found to be negatively affected by high concentrations of atrazine and the seasonal bloom of this taxon can be prevented this way (Jüttner et al., 1995), but the effect on biological interactions cannot be ignored in this context since they may lead to controversial results (DeNoyelles et al., 1982).
Geographic distribution of silica-scaled flagellates also provides an option for bioindication due the correct knowledge of their silica ultrastructure (Kristiansen, 2005). As regard of their global distribution, the moderate endemic model was confirmed (Gusev & Martynenko, 2022), and many species have well defined geographic distribution types Kristiansen & Vigna, 1996; Kristiansen, 2001a, b), such as bipolar (e.g. M. transsylvanica Péterfi et Momeu), cosmopolitan (e.g. Synura petersenii), pantropic (e.g. M. bangladeshica (E. Takahashi & T. Hayakawa) Siver & A. P. Wolfe), endemic (app. 1/3 of Mallomonas species, Kristiansen, 2002), etc. However, the distribution type of a species can be changed due to increasing amount of available floristic data (e.g. in Kristiansen, 2005; Ma et al., 2012) or revealing of hidden cryptic diversity by molecular investigations (Kim et al., 2007; Boo et al., 2010; Kynčlová et al., 2010; Škaloud et al., 2013a).
Although the distribution of silica-scaled chrysophytes was shown to be a space-driven pattern (Siver & Lott, 2012) the process of their dispersal remained poorly understood (Boo et al, 2010). Historical conditions (e.g. glaciations, continent movements, Boo et al., 2010; Bessudova et al., 2021), age of the species (Foissner, 2006) are favoured as explanations for dispersal. Cysts are also assumed to serve as dispersal vehicles, but their different properties (such as the ability to production of cysts or their viability, Kristiansen, 2005; Foissner, 2006) and historical conditions (e.g. lack of glaciation) can highly determine the geographical distribution of species. For example, higher frequency of endemics can be found in tropical than temperate regions (Foissner, 2006). Regarding further dispersal vectors suggested for protists (reviewed by Foissner, 2006), wind and animals seemed to be ineffective colonisation vectors in case of some Synura lineage (Boo et al., 2010) and cysts (Kristiansen, 2005). However, chrysophytes can reflect efficient dispersal especially by long-distance avian vectors (Padisák et al., 2016), and the distributional patterns of some species were tried to be explained by migration routes of birds (e.g. Wee et al., 1994; Péterfi & Momeu, 1996; Kristiansen & Vigna, 1996). In the discussion of dispersal mechanisms, the role of human activities have been largely ignored (Foissner, 2006) despite some explored examples (e.g. ballast water, Nicholls, 2001; plankton nets, Wejnerowski et al, 2022).
The changes in the distribution patterns of silica-scaled chrysophyte species may indicate climate change processes. For instance, the tropical influence on the northeast freshwater environments of Argentina has already been confirmed (Vigna & Siver, 2003). As a further example, pantropic species (e.g. M. bangladeshica, M. stellata Croberg, M. peronoides (K. Harris) Momeu & Péterfi) found in sediment cores at higher latitudes allow for reconstruction of community changes during historical climate warming periods (Siver & Wolfe, 2009) since most climate models predict shifts towards higher latitudes for these species. However, such a monitoring would need checking the silica-scaled chrysophyte flora with a frequency of a few years.
Despite being good bioindicators, silica-scaled chrysophytes are rarely used in the routine or sporadic monitoring programs where the light microscopy is the currently most widely applied equipment, whilst their accurate identification requests genetic tools or electron microscopy (transmission [TEM] and scanning electron [SEM]) analyses (Kristiansen, 2005). In contrast, chrysophytes cysts, despite their precise taxonomic affiliations have not been resolved, are among the most important and widely used biological proxies in paleolimnology along with diatoms, ostracods, Cladocera and chironomids. All chrysophytes are believed to produce siliceous resting stages (Duff et al., 1995), which are also known as stomatocysts or simply cysts. The number of their described morphotypes is over 800. Cysts are more resistant to dissolution than chrysophyte scales and spines, preserve well in sediments, therefore, they have high paleolimnological potential in environmental reconstruction (e.g. Duff et al., 1995; Vorobyova et al., 1996; Kamenik et al., 2001; Pla, 2001; Wilkinson et al., 2001; Kamenik & Schmidt, 2005; Pla & Catalan, 2005; Huber et al., 2009; Simmatis et al., 2022). Stomatocyst assemblages provide a sensitive biotic proxy of pH and salinity (Facher & Schmidt, 1996; Pla et al., 2003; Pla & Anderson, 2005), and particularly, they have been used to reconstruct cold-season climate variability (e.g. Huber et al., 2009; de Jong & Kamenik, 2011; Pla˗Rabés & Catalan, 2011; de Jong et al., 2013). Chrysophycean stomatocysts are proven to be a useful and unique proxy for assessing the ice cover changes, lake mixing (Kamenik & Schmidt, 2005) and seasonality linked to lake stratification patterns (Pla˗Rabés & Catalan, 2011). In Alpine lakes, for example, variations in stomatocyst assemblages have been found to co-vary with the time of spring thaw (Kamenik & Schmidt, 2005). In lakes in southwest Greenland, chrysophytes cysts show a strong relationship with some geographic and conductivity gradients (Pla & Anderson, 2005). Their unique potential is that stomatocysts as proxy are among the few biological proxies, which provide a basis for reconstructing former winter temperatures (de Jong & Kamenik, 2011). Moreover, they also appear to be influenced by seasonal changes in lake temperature (Pla-Rabés & Catalan, 2011, 2018). Cysts proved to be a key proxy for monitoring the response of lake systems to the ongoing global warming (e.g. Betts-Piper et al., 2004; Buczkó et al., 2018). The relationships between chrysophyte cysts and changes in meteorological conditions was tested and validated by Szczerba et al. (2021). But we must bear in mind during the evaluation of results that not only silica-scaled chrysophytes produce stomatocytes, but also chrysophytes belonging to non-scaled lineage (Bazhenova & Igoshkina 2020).
Due to their simplicity, an index of chrysophyte stomatocysts relative to diatom frustules is often used as a proxy in paleolimnology. The ratio of cysts and diatoms is expressed as a percentage, C:D% (Smol, 1985; Zeeb & Smol, 2002). A decrease in this ratio has been reported in some studies to accompany increased levels of metal contamination (Cattaneo et al., 2011; Chen et al., 2014), and it is a sensitive climate proxy (Soróczki˗Pintér et al., 2014). For the accuracy it must be mentioned that chrysophytes can be represented in a sedimentary record by not only of their cysts, but also as “disarticulated” scales, bristles and spines produced by members of the Synurales and some Chrysophyceae, but their use as biotic proxy was proved less effective than application of cysts (Duff et al., 1995).
Biochemicals
Microalgae are an important resource of biotechnology and nanotechnology in many ways (e.g. Khalid, 2020) as they are valuable and eco-friendly sources of a variety of bioactive compounds (e.g. vitamins, proteins, lipids, carbohydrates, fatty acids, sterols, photosynthetic pigments, etc.). These compounds have a wide range of applications by humans including food supplements, cosmetics, and pharmaceuticals (Kainz et al., 2009; Sathasivam & Ki, 2018). The practical use of silica-scaled chrysophytes is mainly restricted to the laboratory despite there is plenty of potential industrial opportunities.
Chrysophytes in aquacultures can serve as food for bivalves with high estimated market value (Benemann et al., 1987). Mallomonas spp. and Paraphysomonas spp. produce important bioactive compounds, such as PUFAs (polyunsaturated fatty acids), which increase their nutritional value (Cranwell et al., 1988) and consequently provide a good food source for zooplankton both in quantity and the quality (Bec et al., 2010; Strandberg et al., 2015).
Synura petersenii produces sterols (β-sitosterol and cholesterol, Collins & Kalnins, 1969), as well as Paraphysomonas can also synthesise such compounds de novo (Bec et al. 2006), which can serve as food for grazers, precursors for vitamin D and a multitude of naturally occurring steroid hormones (Kainz et al., 2009). The polysaccharide produced by Synura consists of neutral sugars such as mannose, xylose, and rhamnose (up to 50% of the total polysaccharide composition), and it has a relatively high protein content (Lombardi & Vieira, 1998). Additionally, chrysophyte biomass provides high amounts of hydrocarbons. A significant lipid content was shown in Mallomonas splendens (G.S.West) Playfair (Cooper et al., 2010), hence suitable bio-oil extraction to produce liquid fuels (Yang et al., 2014).
Carotenoids can be used for improving food quality and their demand in the global market increased from ~ 1.24 billion USD in 2016 to ~ 1.53 billion USD in 2021 (web2). In general, the major types of carotenoids used commercially in the global market are astaxanthin, β-carotene, lutein, canthaxanthin, lycopene, and zeaxanthin (Sathasivam & Ki, 2018). Mallomonas sp. can produce fucoxanthin in significant amounts (26.6 mg g−1 dry weight), which have anti-obesity and anti-oxidant property making fucoxanthin to be very promising compound for commercial production (Petrushkina et al., 2017). Furthermore, Mallomonas sp. could also be an important source of β-carotene (Péterfi & Momeu, 2009) with potential use in the human healthcare and cosmetics industry (Bogacz-Radomska & Harasym, 2018).
Paraphysomonas vestita (A.Stokes) De Saedeleer have MGDGs (monogalactosyldiacylglycerols), SQD (sulfoquinovosyldiacylglycerols), whilst Synurales have only MGDG genes. These compounds have important biological properties: MGDGs in anti-inflammatory and anti-cancer activities, SQDGs in immunostimulatory activities and inhibition of the enzyme glutaminyl cyclase involved in Alzheimer disease (Riccio et al., 2020). These species can provide possible biotechnological applications to produce bioactive lipids.
Chrysophytes are among the “cold algae” as most species dominate in cold, even extremely cold regions (Arctic, Antarctic, mountainous regions, Varshney et al., 2015) or they can bloom in low temperature periods (Cen et al., 2020). Their cold resistance could be affiliated with their polyunsaturated fatty acid production (PUFA) such as it was found in case of marine chryptophytes (Peltomaa et al., 2018). PUFA is generally known to have antifreeze properties (Alcaíno et al., 2015) and ensures the fluidity of the cell membranes lowering its melting point (Brunton & Collins, 2007). This special extremophile feature is a further potential in biotechnology (Varshney et al., 2015), e.g. as ingredients for nutraceuticals (Peltomaa et al., 2018).
Biosilica also provides inspiration in biological materials science (Wysokowski et al., 2018). Despite the silica scales of chrysophytes are made of an identical chemical compound as the frustules of diatoms and have high application potential, their use in nanotechnologies has been in the early stages of research and far behind that of diatoms’ (reviewed by Kratošová et al., 2013). Two studies showed the extensive potential of Mallomonas kalinae Řezáčová for metal nanoparticle biosynthesis with possible utilisation in a multitude of applications (Kratošová et al., 2013; Holišová et al., 2019). Biominerals of Synura petersenii and Mallomonas caudata and their subsequent conversion into elemental silicon via magnesiothermic conversion route (Petrack et al., 2014) is of potential interest for catalytic, biochemical, electronic and thermal applications, as well as the use in lithium batteries (Sandhage et al., 2002; Xing et al., 2013).
Genetic resources
Since the first study focusing on DNA sequences of silica-scaled chrysophytes was published only in the early 1990s (Ariztia et al., 1991), molecular-based research of silica-scaled chrysophytes has started to develop only recently. These investigations are based on nuclear and plastid gene sequencing techniques and are targeted recently on phylogenetic and evolutionary relationships (e.g. Andersen, 2007; Jo et al., 2013; Škaloud et al., 2013a; Bock et al., 2017; Čertnerová et al., 2019). Regarding practical application the small subunit (SSU) rRNA gene was used and proved to be appropriate for measuring abundance of Paraphysomonas with fluorescent oligonucleotide rDNA probes (Lim et al., 1999; Rice et al., 1997). Furthermore, understanding the mechanisms that silica-scaled chrysophytes utilise to produce their silica structures may also provide a good opportunity for their nanotechnological applicability. In this area, Schultz et al. (2001) detected an SP41 protein and its encoding cDNA, which plays a role in morphogenesis and/or silicification of Mallomonas splendens.
Our genetic information about silica-scaled chrysophytes has been poor since many genera remained uncharacterized (e.g. Spiniferomonas). The published sequences are related to only a small portion of species (Škaloud et al., 2013a), and only five species (Mallomonas splendens, Neotessella volvocina, Synura petersenii, S. sphagnicola (Korshikov) Korshikov, S. uvella) have their complete plastid genome sequences explored (Kim et al., 2019) and only one detailed transcriptomic analysis was performed investigating the trophic mode of one Synura species (Beisser et al., 2017).
Cultural services
Aesthetic
Silica-scaled chrysophyte species can develop significant amounts of biomass in many ecosystems, even under ice-cover. These blooms sometimes can manifest in extraordinary appearance (visible coloration) or make unpleasant conditions (Sandgren et al., 1995; Watson et al., 2001a, b), which particular aesthetic features may attract tourists. Synura uvella bloom causes brown, whilst Mallomonas acaroids Perty yellow discolouration (Jüttner, 1981; Kristiansen et al., 1995; Kalinowska & Grabowska, 2016). Besides the visual effects, chrysophytes produce diverse and significant amount of volatile organic compounds (VOCs) causing taste-odour events (Watson et al., 2001b; Watson, 2003; Nicholls & Wujek, 2015; Lee et al., 2017), even causing problems in drinking water supplies and processed food products (Sandgren et al., 1995). Though single species have no known unique chemical fingerprints (Watson, 2003), yet some silica-scaled chrysophyte species can be responsible for some characteristic odour problems:
-
Synura petersenii and Mallomonas papillosa K.Harris & D.E.Bradley for rancid/fishy-smell (Nicholls & Gerrath, 1985; Wee et al., 1994; Rashash et al., 1995; Watson et al., 2001b);
-
Synura petersenii for cucumber odour (Hayes & Burch, 1989; Wee et al., 1994; Rashash et al., 1995);
-
Synura uvella for cod-liver oil- (Jüttner, 1981) and fishy odorants (Liu et al., 2019);
-
Synura ssp. for violets/fruity/ester-like/green/plastic odours (reviewed by Lee et al., 2017). These chemical fingerprints can be useful even for water management (e.g. development or choice of the most effective treatment techniques to minimise or eliminate the aesthetically unpleasant effects, Hayes & Burch, 1989; Watson et al., 2001a, b; Lee et al., 2017).
Inspiration
Silica-scaled chrysophytes (their scales, whole cells, and cysts) have extraordinarily beautiful appearance (e.g. structures of scales, colony form), which can provide inspiration for artists in many areas, whether at an amateur or artistic level. These algae are involved (among many others) in
-
drawings (dragon, covered by Synura scales, web3),
-
paintings (Mallomonas sp., web4),
-
photographs (cysts, Mallomonas sp., Synura sp., web5, web6),
-
designs of face masks (web7),
-
jewellery (web8) and even
-
puzzles (Synura uvella, web9).
Knowledge system
Only one on-line database (https://chrysophytes.eu/) was established in order to collect records dealing with the European silica-scaled chrysophytes (Škaloud et al., 2013b). The database consists of geographical coordinates, distribution map, distribution frequencies (along temperature, pH and conductivity gradients), references, as well as microphotographs (TEM, SEM, light microscope [LM]). At the moment, based on more than 7500 entries, the database provides information on 204 species and intraspecific taxa, which significantly contribute to our knowledge.
No available world-wide database has exist about the distribution of silica-scaled chrysophytes, but the AlgaeBase system (Guiry & Guiry, 2022) can provide information about the whole group of algae, including “Detailed distribution with sources” and “Key references” as their slogan signs: “Listing the World’s Algae”.
Summary and conclusions
Due to the required special identification facilities (high quality LM, SEM, TEM) silica-scaled chrysophytes have been a poorly studied group of algae in many fields of research. Despite these difficulties, the existing information highlights their great ecological value. They are involved altogether in 14 ecosystem services (13 defined by MEA 2003, 2005; and another one), and can also have further powerful opportunities. The deficiency roots in the fact that very little information is available from a major part of the group, which is restricted almost exclusively to floristic data (e.g. Chrysosphaerella, Spiniferomonas, Polylepidomonas). Their detailed research would mainly need physiological tests, but they are difficult to grow in culture due to their trophic requirements or special needs for growth conditions (Bhatti & Colman 2005; Kristiansen & Škaloud, 2017). However, there are a few species that have been better studied and used as model species (e.g. Paraphysomonas vestita, P. imperforata, Synura petersenii). So far, not all of silica-scaled species, even genera, have been investigated in detail at molecular level resulting only in a few available genomic and transcriptomic information. However, silica-scaled chrysophytes can be considered important genetic resources, since they can produce many important bioactive compounds (e.g. silica structures, carotenoids, allelopathic VOCs), thus their detailed investigation will be very useful, for instance, for genetic engineering studies, increasingly evolving nanotechnology and “green” water management technology.
Data availability
Data sharing not applicable to this article as no datasets were generated or analysed during the current study.
References
Alcaíno, J., V. Cifuentes & M. Baeza, 2015. Physiological adaptations of yeasts living in cold environments and their potential applications. World Journal of Microbiology and Biotechnology 31: 1467–1473.
Andersen, R. A., 1987. Synurophyceae classis nov., a new class of algae. American Journal of Botany 74: 337–353.
Andersen, R. A., 2007. Molecular systematics of the Chrysophyceae and Synurophyceae. In Brodie, J. & J. Lewis (eds), Unravelling the algae: the past, present, and future of algal systematics CRC Press: 285–313.
Andersen, K. O., J. C. Goldman, D. A. Caron & M. R. Dennett, 1986. Nutrient cycling in a microflagellate food chain: III phosphorous dynamics. Marine Ecology Progress Series 31: 47–55.
Arber, W., 2004. Biological evolution: lessons to be learned from microbial population biology and genetics. Research in Microbiology 155: 297–300.
Ariztia, E. V., R. A. Andersen & M. L. Sogin, 1991. A new phylogeny for Chromophyte algae using 16S-like RRNA Sequences from Mallomonas papillosa (Synurophyceae) and Tribonema aequale (Xanthophyceae). Journal of Phycology 27: 428–436.
Arts, M. T., R. G. Ackman & B. J. Holub, 2001. “Essential fatty acids” in aquatic ecosystems: a crucial link between diet and human health and evolution. Canadian Journal of Fisheries and Aquatic Sciences 58: 122–137.
Arvola, L. & K. Salonen, 2001. Plankton community of a polyhumic lake with and without Daphnia longispina (Cladocera). Hydrobiologia 445: 141–150.
Barbeau, K., E. Kujawinski & J. Moffett, 2001. Remineralization and recycling of iron, thorium and organic carbon by heterotrophic marine protists in culture. Aquatic Microbial Ecology 24: 69–81.
Barreto, S., 2001. Three new Mallomonas species (Synurophyceae) from a mountain area in North-Hungary. Nordic Journal of Botany 21: 437–448.
Bazhenova, O. P. & I. Y. Igoshkina, 2020. Diversity and peculiarities of the formation of stomatocysts of the Chrysophyceae in a waterbody in the South of Western Siberia. Inland Water Biology 13: 556–565.
B-Béres, V., C. Stenger-Kovács, K. Buczkó, J. Padisák, G. B. Selmeczy, E. Lengyel & K. Tapolczai, 2022. Ecosystem services provided by freshwater and marine diatoms. Hydrobiologia. https://doi.org/10.1007/s10750-022-04984-9.
Bec, A., D. Martin-Creuzburg & E. von Elert, 2006. Trophic upgrading of autotrophic picoplankton by the heterotrophic nanoflagellate Paraphysomonas sp. Limnology and Oceanography 51: 1699–1707.
Bec, A., D. Martin-Creuzburg & E. Von Elert, 2010. Fatty acid composition of the heterotrophic nanoflagellate Paraphysomonas sp.: influence of diet and de novo biosynthesis. Aquatic Biology 9: 107–112.
Beisser, D., N. Graupner, C. Bock, S. Wodniok, L. Grossmann, M. Vos, B. Sures, S. Rahmann & J. Boenigk, 2017. Comprehensive transcriptome analysis provides new insights into nutritional strategies and phylogenetic relationships of chrysophytes. PeerJ 5: e2832.
Benemann, J. R., D. M. Tillett & J. C. Weissman, 1987. Microalgae biotechnology. Trends in Biotechnology 5: 47–53.
Bessudova, A., Y. Bukin & Y. Likhoshway, 2021. Dispersal of silica-scaled chrysophytes in Northern Water bodies. Diversity 13: 284.
Betts-Piper, A. M., B. A. Zeeb & J. P. Smol, 2004. Distribution and autecology of chrysophyte cysts from high Arctic Svalbard lakes: preliminary evidence of recent environmental change. Journal of Paleolimnology 31: 467–481.
Bhatti, S. & B. Colman, 2005. Inorganic carbon acquisition by the chrysophyte alga Mallomonas papillosa. Canadian Journal of Botany 83: 891–897.
Bhatti, S. & B. Colman, 2008. Inorganic carbon acquisition in some synurophyte algae. Physiologia Plantarum 133: 33–40.
Bhatti, S. & B. Colman, 2011. Evidence for the occurrence of photorespiration in synurophyte algae. Photosynthesis Research 109: 251–256.
Blaauboer, M. C., R. Van Keulen & T. E. Cappenberg, 1982. Extracellular release of photosynthetic products by freshwater phytoplankton populations, with special reference to the algal species involved. Freshwater Biology 12: 559–572.
Bock, C., R. Medinger, S. Jost, R. Psenner & J. Boenigk, 2014. Seasonal variation of planktonic chrysophytes with special focus on Dinobryon. Fottea 14: 179–190.
Bock, C., A. Chatzinotas & J. Boenigk, 2017. Genetic diversity in chrysophytes: comparison of different gene markers. Fottea 17: 209–221.
Bock, C., S. Zimmermann, D. Beisser, S.-M. Dinglinger, S. Engelskirchen, P. Giesemann, S. Klink, J. L. Olefeld, S. Rahmann & M. Vos, 2019. Silver stress differentially affects growth of phototrophic and heterotrophic chrysomonad flagellate populations. Environmental Pollution 244: 314–322.
Bogacz-Radomska, L. & J. Harasym, 2018. β-Carotene – properties and production methods. Food Quality and Safety 2: 69–74.
Bogush, A. A., G. A. Leonova, S. K. Krivonogov, V. A. Bobrov, V. D. Tikhova, L. M. Kondratyeva, A. E. Kuzmina & A. E. Maltsev, 2013. Diagenetic transformation of sapropel from Lake Dukhovoe (East Baikal region, Russia). Procedia Earth and Planetary Science 7: 81–84.
Boo, S. M., H. S. Kim, W. Shin, G. H. Boo, S. M. Cho, B. Y. Jo, J.-H. Kim, J. H. Kim, E. C. Yang, P. A. Siver, A. P. Wolfe, D. Bhattacharya, R. A. Andersen & H. S. Yoon, 2010. Complex phylogeographic patterns in the freshwater alga Synura provide new insights into ubiquity vs. endemism in microbial eukaryotes. Molecular Ecology 19: 4328–4338.
Brunton, S. & N. Collins, 2007. Differentiating prescription omega-3-acid ethyl esters (P-OM3) from dietary-supplement omega-3 fatty acids. Current Medical Research and Opinion 23: 1139–1145.
Buczkó, K., E. Szurdoki, M. Braun & E. Magyari, 2018. Reconciling diverse diatom-based lake responses to climate change in four mountain lakes in the South-Carpathian Mountains during the last 17 kyrs. Quaternary International 477: 117–137.
Carney, H. J., 1990. A general hypothesis for the strength of food web interactions in relation to trophic state. Internationale Vereinigung für theoretische und angewandte Limnologie, Verhandlungen 24: 487–492.
Carpenter, S. R., J. F. Kitchell & J. R. Hodgson, 1985. Cascading trophic interactions and lake productivity. BioScience 35: 634–639.
Cattaneo, A., Y. Couillard, S. Wunsam & C. Fortin, 2011. Littoral diatoms as indicators of recent water and sediment contamination by metals in lakes. Journal of Environmental Monitoring 13: 572–582.
Cen, C., K. Zhang, T. Zhang, X. Zhou & R. Pan, 2020. Algae-induced taste and odour problems at low temperatures and the cold stress response hypothesis. Applied Microbiology and Biotechnology 104: 9079–9093.
Čertnerová, D., M. Čertner & P. Škaloud, 2019. Molecular phylogeny and evolution of phenotype in silica-scaled chrysophyte genus Mallomonas. Journal of Phycology 55: 912–923.
Charvet, S., W. F. Vincent & C. Lovejoy, 2012. Chrysophytes and other protists in high Arctic lakes: molecular gene surveys, pigment signatures and microscopy. Polar Biology 35: 733–748.
Chen, X., C. Li, S. McGowan & X. Yang, 2014. Diatom response to heavy metal pollution and nutrient enrichment in an urban lake: evidence from paleolimnology. Annales de Limnologie – International Journal of Limnology 50: 121–130.
Cole, J. J., S. Findlay & M. L. Pace, 1988. Bacterial production in fresh and saltwater ecosystems: a cross-system overview. Marine Ecology Progress Series Oldendorf 43: 1–10.
Collins, R. & K. Kalnins, 1969. Sterols produced by Synura petersenii (Chrysophyta). Comparative Biochemistry and Physiology 30: 779–782.
Cooper, M. S., W. R. Hardin, T. W. Petersen & R. A. Cattolico, 2010. Visualizing “green oil” in live algal cells. Journal of Bioscience and Bioengineering 109: 198–201.
Cormier, M. A., J. B. Berard, G. Bougaran, C. N. Trueman, D. J. Mayor, R. S. Lampitt, N. J. Kruger, K. J. Flynn & R. E. Rickaby, 2022. Deuterium in marine organic biomarkers: toward a new tool for quantifying aquatic mixotrophy. New Phytologist 234: 776–782.
Cranwell, P. A., M. E. Creighton & G. H. Jaworski, 1988. Lipids of four species of freshwater chrysophytes. Phytochemistry 27: 1053–1059.
Dalby, A. P., K. A. Kormas, U. Christaki & H. Karayanni, 2008. Cosmopolitan heterotrophic microeukaryotes are active bacterial grazers in experimental oil-polluted systems. Environmental Microbiology 10: 47–56.
de Jong, K. & C. Kamenik, 2011. Validation of chrysophyte stomatocysts-based cold-season climate reconstruction from high-Alpine Lake Silvaplana, Switzerland. Journal of Quaternary Science 26: 268–275.
de Jong, K., C. Kamenik, K. Westover & M. Grosjean, 2013. A chrysophyte stomatocyst-based reconstruction of cold-season air temperature from alpine Lake Silvaplana (AD 1500–2003); methods and concepts for quantitative inferences. Journal of Paleolimnology 50: 519–533.
DeNoyelles, F., W. D. Kettle & D. E. Sinn, 1982. The responses of plankton communities in experimental ponds to atrazine, the most heavily used pesticide in the United States. Ecology 63: 1285–1293.
Dixit, S. S., A. S. Dixit & R. D. Evans, 1989. Paleolimnological evidence for trace-metal sensitivity in scaled chrysophytes. Environmental Science & Technology 23: 110–115.
Dolan, J., 1997. Phosphorus and ammonia excretion by planktonic protists. Marine Geology 139: 109–122.
Douglas, M. S. & J. P. Smol, 1995. Paleolimnological significance of observed distribution patterns of chrysophyte cysts in arctic pond environments. Journal of Paleolimnology 13: 79–83.
Duff, K. E., B. A. Zeeb & J. P. Smol, 1995. Atlas of chrysophycean cysts, Kluwer Academic Press, Dordrecht.
Facher, E. & R. Schmidt, 1996. A siliceous chrysophycean cyst˗based pH transfer function for Central European lakes. Journal of Paleolimnology 16: 275–321.
Feng, Y., C. E. Hare, K. Leblanc, J. M. Rose, Y. Zhang, G. R. DiTullio, P. A. Lee, S. W. Wilhelm, J. M. Rowe & J. Sun, 2009. Effects of increased pCO2 and temperature on the North Atlantic spring bloom. I: the phytoplankton community and biogeochemical response. Marine Ecology Progress Series 388: 13–25.
Ferrier, M., B. Butler Sr., D. Terlizzi & R. Lacouture, 2005. The effects of barley straw (Hordeum vulgare) on the growth of freshwater algae. Bioresource Technology 96: 1788–1795.
Foissner, W., 2006. Biogeography and dispersal of micro-organisms: a review emphasizing protists. Acta Protozoologica 45: 111–136.
Gensemer, R. W. & R. C. Playle, 1999. The bioavailability and toxicity of aluminum in aquatic environments. Critical Reviews in Environmental Science and Technology 29: 315–450.
Goldman, J. C. & M. R. Dennett, 1990. Dynamics of prey selection by an omnivorous flagellate. Marine Ecology Progress Series 59: 183–194.
Goldman, J. C. & M. R. Dennett, 1992. Phagotrophy and NH4+ regeneration in a three-member microbial food loop. Journal of Plankton Research 14: 649–663.
Graham, L. E., J. M. Graham, L. W. Wilcox & M. E. Cook, 2000. Algae, Prentice Hall, Upper Saddle River.
Graupner, N., M. Jensen, C. Bock, S. Marks, S. Rahmann, D. Beisser & J. Boenigk, 2018. Evolution of heterotrophy in chrysophytes as reflected by comparative transcriptomics. FEMS Microbiology Ecology 94: fiy039.
Guildford, S. J., H. A. Bootsma, W. D. Taylor & R. E. Hecky, 2007. High variability of phytoplankton photosynthesis in response to environmental forcing in oligotrophic Lake Malawi/Nyasa. Journal of Great Lakes Research 33: 170–185.
Guiry, M. & G. Guiry, 2022. AlgaeBase. Worldwide electronic publication, National University of Ireland, Galway. https://www.algaebase.org; searched on 2022. március 13.
Gusev, E. S., 2013. Studies on synurophycean algae from mangrove wetlands (Vietnam). Nova Hedwigia Beiheft 142: 87–95.
Gusev, E. & N. Martynenko, 2022. Diversity of silica-scaled chrysophytes in Central Vietnam. Water 14: 65.
Gusev, E. & P. A. Siver, 2017. Mallomonas neoampla sp. nov. from Vietnam, a new species that bridges the gap between fossil and modern taxa. Nova Hedwigia 104: 521–528.
Gusev, E. S., H. Doan-Nhu, L. Nguyen-Ngoc & D. A. Kapustin, 2016. Two new species of the genus Mallomonas from the Cat Tien National Park (Vietnam): Mallomonas distinguenda and Mallomonas skvortsovii. Phytotaxa 273: 59–64.
Gusev, E. S., D. Čertnerová, M. Škaloudová & P. Škaloud, 2018. Exploring cryptic diversity and distribution patterns in the Mallomonas kalinae/rasilis species complex with a description of a dew taxon – Mallomonas furtiva sp. nov. Journal of Eukaryotic Microbiology 65: 38–47.
Gusev, E., N. Martynenko & H. Tran, 2021. Studies on algae from the order Synurales (Chrysophyceae) in Northern Vietnam. Diversity 13: 602.
Hansson, L.-A. & L. J. Tranvik, 1996. Quantification of invertebrate predation and herbivory in food chains of low complexity. Oecologia 108: 542–551.
Hayes, K. P. & M. D. Burch, 1989. Odorous compounds associated with algal blooms in South Australian waters. Water Research 23: 115–121.
Hendry, K. R., A. O. Marron, F. Vincent, D. J. Conley, M. Gehlen, F. M. Ibarbalz, B. Quéguiner & C. Bowler, 2018. Competition between silicifiers and non-silicifiers in the past and present ocean and its evolutionary impacts. Frontiers in Marine Science 5: 22.
Hickel, B. & I. Maass, 1989. Scaled chrysophytes, including heterotrophic nanoflagellates from the lake district in Holstein, northern Germany. Beihefte Zur Nova Hedwigia 95: 233–257.
Hoek, C., D. Mann, H. M. Jahns & M. Jahns, 1995. Algae: an introduction to phycology, Cambridge University Press, New York.
Hoffmann, L. & E. Wille, 1992. Occurrence of a metalimnetic summer peak of Mallomonas caudata (Synurophyceae). Nordic Journal of Botany 12: 465–469.
Holišová, V., M. Urban, M. Kolenčík, Y. Němcová, A. Schröfel, P. Peikertová, J. Slabotinský & G. Kratošová, 2019. Biosilica-nanogold composite: Easy-to-prepare catalyst for soman degradation. Arabian Journal of Chemistry 12: 262–271.
Hooper, D. U., E. C. Adair, B. J. Cardinale, J. E. Byrnes, B. A. Hungate, K. L. Matulich, A. Gonzalez, J. E. Duffy, L. Gamfeldt & M. I. O’Connor, 2012. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 486: 105–108.
Huber, K., C. Kamenik, K. Weckström & R. Schmidt, 2009. Taxonomy, stratigraphy, and palaeoecology of chrysophyte cysts from late glacial sediment core section of Längsee, Austria. Nova Hedwigia 89: 245–261.
Hutchins, D. & K. Bruland, 1994. Grazer-mediated regeneration and assimilation of Fe, Zn and Mn from planktonic prey. Marine Ecology Progress Series 110: 259–259.
Irfan, S. & A. M. M. Alatawi, 2019. Aquatic ecosystem and biodiversity: a review. Open Journal of Ecology 9: 1–13.
Jeong, M., J. I. Kim, B. Y. Jo, H. S. Kim, P. A. Siver & W. Shin, 2019. Surviving the marine environment: two new species of Mallomonas (Synurophyceae). Phycologia 58: 276–286.
Jo, B. Y., W. Shin, H. S. Kim, P. A. Siver & R. A. Andersen, 2013. Phylogeny of the genus Mallomonas (Synurophyceae) and descriptions of five new species on the basis of morphological evidence. Phycologia 52: 266–278.
Jo, B. Y., J. I. Kim, P. Škaloud, P. A. Siver & W. Shin, 2016. Multigene phylogeny of Synura (Synurophyceae) and descriptions of four new species based on morphological and DNA evidence. European Journal of Phycology 51: 413–430.
Jüttner, F., 1979. Nor-carotenoids as the major volatile excretion products of Cyanidium. Zeitschrift für Naturforschung C 34: 186–191.
Jüttner, F., 1981. Detection of lipid degradation products in the water of a reservoir during a bloom of Synura uvella. Applied and Environmental Microbiology 41: 100–106.
Jüttner, F., 2001. Liberation of 5, 8, 11, 14, 17-eicosapentaenoic acid and other polyunsaturated fatty acids from lipids as a grazer defense reaction in epilithic diatom biofilms. Journal of Phycology 37: 744–755.
Jüttner, I., A. Peither, J. Lay, A. Kettrup & S. Ormerod, 1995. An outdoor mesocosm study to assess ecotoxicological effects of atrazine on a natural plankton community. Archives of Environmental Contamination and Toxicology 29: 435–441.
Kainz, M., M. T. Brett & M. T. Arts, 2009. Lipids in aquatic ecosystems, Springer, New York.
Kalinowska, K. & M. Grabowska, 2016. Autotrophic and heterotrophic plankton under ice in a eutrophic temperate lake. Hydrobiologia 777: 111–118.
Kamenik, C. & R. Schmidt, 2005. Chrysophyte resting stages: a tool for reconstructing winter/spring climate from Alpine lake sediments. Boreas 34: 477–489.
Kamenik, C., R. Schmidt, K. A. Koinig, A. Agusti-Panareda, R. Thompson & R. Psenner, 2001. The chrysophyte stomatocyst composition in high alpine lake (Gossenköllesee, Tyrol) in relation to seasonality, temperature and land-use. Nova Hedwigia, Beiheft 122: 1–22.
Kaplan, D., D. Christiaen & S. Arad, 1988. Binding of heavy metals by algal polysaccharides. In Stadler, T., J. Mollion, M. C. Verdus, Y. Karamanos, H. Morvan & D. Christiaen (eds), Algal biotechnology Elsevier Applied Science, London: 179–187.
Kapustin, D. A. & M. D. Guiry, 2019. Reinstatement of Lepidochromonas kristiansen (Lepidochromonadaceae fam. Nov., Chrysophyceae). Phytotaxa 413: 049.
Kapustin, D. A. & E. S. Gusev, 2019. Silica-scaled chrysophytes from West Java (Indonesia) including description of a new Chrysosphaerella species. Nova Hedwigia 148: 11–20.
Karjalainen, J., A.-L. Holopainen & P. Huttunen, 1996. Spatial patterns and relationships between phytoplankton, zooplankton and water quality in the Saimaa lake system, Finland. In Simola, H., M. Viljanen, T. Slepukhina & R. Murthy (eds), The first international lake ladoga symposium: development in hydrobiology Springer, Dordrecht: 267–276.
Khalid, M., 2020. Nanotechnology and chemical engineering as a tool to bioprocess microalgae for its applications in therapeutics and bioresource management. Critical Reviews in Biotechnology 40: 46–63.
Khatiwala, S., F. Primeau & T. Hall, 2009. Reconstruction of the history of anthropogenic CO2 concentrations in the Ocean. Nature 462: 346–349.
Kim, H. & J. Soon Kim, 2008. Mallomonas koreana sp. nov. (Synurophyceae), a new species from South Korea. Nova Hedwigia 86: 469–476.
Kim, H. S., W. G. Shin & S. M. Boo, 2007. Nuclear SSU and plastid rbcL genes and ultrastructure of Mallomonas caudata (Synurophyceae) from Korea. Korean Journal of Ecology and Environment 40: 387–394.
Kim, J. I., H. Shin, P. Škaloud, J. Jung, H. S. Yoon, J. M. Archibald & W. Shin, 2019. Comparative plastid genomics of Synurophyceae: inverted repeat dynamics and gene content variation. BMC Evolutionary Biology 19: 1–13.
Kim, J. I., M. Jeong, J. M. Archibald & W. Shin, 2020. Comparative plastid genomics of non-photosynthetic chrysophytes: genome reduction and compaction. Frontiers in Plant Science 11: 572703.
Kiss, K. T. & J. Kristiansen, 1994. Silica-scaled chrysophytes (Synurophyceae) from some rivers and shallow lakes in Hungary. Hydrobiologia 289: 157–162.
Kormas, K. A., U. Christaki & A. Dalby, 2007. Microscopic eukaryotes are alive and well in marine oil-polluted environments. Rapp. Comm. Int. Mer Médit. 38: 377.
Kratošová, G., I. Vávra, K. Horská, O. Zˇivotský, Y. Nemcová, M. Bohunická, J. Slabotinský, K. Rosenbergová, A. Kadilak & A. Schröfel, 2013. Synthesis of metallic nanoparticles by diatoms and chrysophytes-prospects and applications. In Rai, M. & C. Posten (eds), Green biosynthesis of nanoparticles: mechanisms and applications CABI, Boston: 61–78.
Kristiansen, J., 2001a. Biogeography of silica-scaled chrysophytes. Proceeding of the fifth international chrysophyte symposium. Nova Hedwigia 122: 23–39.
Kristiansen, J., 2001b. Cosmopolitan chrysophytes. Systematics and Geography of Plants 70: 291–300.
Kristiansen, J., 2002. The genus Mallomonas (Synurophyceae): a taxonomic survey based on structure of silica scales and bristles. Opera Botanica 139: 1–218.
Kristiansen, J., 2005. Golden algae: a biology of chrysophytes, Koeltz Botanical Books.
Kristiansen, J. & H. Preisig, 2007. Chrysophyte und Haptophyte algae: 2. Teil: Synurophyceae. Spektrum Akademischer Verlag, Springer.
Kristiansen, J., L. R. Wilken & T. Jürgensen, 1995. A bloom of Mallomonas acaroides, a silica-scaled chrysophyte, in the crater pond of a pingo, Northwest Greenland. Polar Biology 15: 319–324.
Kristiansen, J. & P. Škaloud, 2017. Chrysophyta. In Archibald, J. M., et al., (eds), Handbook of the protists Springer, Dordrecht: 331–366.
Kristiansen, J. & M. S. Vigna, 1996. Bipolarity in the distribution of silica-scaled flagellates. Hydrobiologia 336: 121–126.
Kuwata, A., K. Yamada, M. Ichinomiya, S. Yoshikawa, M. Tragin, D. Vaulot & A. Lopes dos Santos, 2018. Bolidophyceae, a sister picoplanktonic group of diatoms – a review. Frontiers in Marine Science 5: 370.
Kynčlová, A., P. Škaloud & M. Škaloudová, 2010. Unveiling hidden diversity in the Synura petersenii species complex (Synurophyceae, Heterokontophyta). Nova Hedwigia, Beihefte 136: 283–298.
Lampert, W. & U. Sommer, 2007. Limnoecology: the ecology of lakes and streams, Oxford University Press, New York.
Leadbeater, B. S., 1986. Biomineralization in lower plants and animals, Clarendon Press, Oxford.
Lee, J., P. K. Rai, Y. J. Jeon, K.-H. Kim & E. E. Kwon, 2017. The role of algae and cyanobacteria in the production and release of odorants in water. Environmental Pollution 227: 252–262.
Lehman, J. T. & C. D. Sandgren, 1985. Species-specific rates of growth and grazing loss among freshwater algae. Limnology and Oceanography 30: 34–46.
Likhoshway, Y. V., E. Sorokovikova, O. Belykh, O. Kaluzhnaya, S. Belikov, Y. D. Bedoshvili, O. Kaluzhnaya, J. A. Masyukova & T. Sherbakova, 2008. Visualization of the silicon biomineralization in cyanobacteria, sponges and diatoms. In Dobretsov, N., N. Kolchanov, A. Rozanov & G. Zavarzin (eds), Biosphere origin and evolution Springer, Boston: 219–230.
Lim, E. L., M. R. Dennett & D. A. Caron, 1999. The ecology of Paraphysomonas imperforata based on studies employing oligonucleotide probe identification in coastal water samples and enrichment cultures. Limnology and Oceanography 44: 37–51.
Liu, T., J. Yu, M. Su, Z. Jia, C. Wang, Y. Zhang, C. Dou, M. Burch & M. Yang, 2019. Production and fate of fishy odorants produced by two freshwater chrysophyte species under different temperature and light conditions. Water Research 157: 529–534.
Lombardi, A. & A. Vieira, 1998. Copper and lead complexation by high molecular weight compounds produced by Synura sp. (Chrysophyceae). Phycologia 37: 34–39.
Loreau, M., S. Naeem, P. Inchausti, J. Bengtsson, J. P. Grime, A. Hector, D. Hooper, M. Huston, D. Raffaelli & B. Schmid, 2001. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294: 804–808.
Ma, C. X., J. Zhang, H. X. Yu, Y. X. Wei & X. P. Yuan, 2012. Silica-scaled chrysophytes of cold region: a case study Zhenbao Island Wetland Preserve, Northeast China. Applied Mechanics and Materials 178: 922–925.
Maberly, S. C., L. A. Ball, J. A. Raven & D. Sültemeyer, 2009. Inorganic carbon acquisition by chrysophytes. Journal of Phycology 45: 1052–1061.
Marron, A. O., S. Ratcliffe, G. L. Wheeler, R. E. Goldstein, N. King, F. Not, C. De Vargas & D. J. Richter, 2016. The evolution of silicon transport in eukaryotes. Molecular Biology and Evolution 33: 3226–3248.
Martínez, R., 1980. Seasonal variation of phytoplankton biomass and photosynthesis in the High-Mountain Lake La Caldera (Sierra Nevada, Spain). In Dokulil, M., H. Metz & D. Jewson (eds), Shallow lakes contributions to their limnology: developments in hydrobiology Springer, Dordrecht: 111–119.
McGenity, T. J., B. D. Folwell, B. A. McKew & G. O. Sanni, 2012. Marine crude-oil biodegradation: a central role for interspecies interactions. Aquatic Biosystems 8: 1–19.
MEA, 2003. Ecosystems and human well–being: a framework for assessment, Island Press, Washington, DC.
MEA, 2005. Ecosystems and human well–being, Island press, Washington, DC.
Moore, M. V., C. F. Folt & R. S. Stemberger, 1996. Consequences of elevated temperatures for zooplankton assemblages in temperate lakes. Archiv für Hydrobiologie 135: 289–319.
Moriceau, B., M. Gehlen, P. Tréguer, S. Baines, J. Livage & L. André, 2019. Editorial: Biogeochemistry and genomics of silicification and silicifiers. Frontiers in Marine Science. 6: 57.
Myers, N., R. A. Mittermeier, C. G. Mittermeier, G. A. Da Fonseca & J. Kent, 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853–858.
Naselli-Flores, L. & J. Padisák, 2022. Ecosystem services provided by marine and freshwater phytoplankton. Hydrobiologia. https://doi.org/10.1007/s10750-022-04795-y.
Němcová, Y. & J. Kreidlova, 2013. Two new species of Mallomonas (Chrysophyceae: Synurales): Mallomonas temonis and Mallomonas divida. Phytotaxa 87: 11–18.
Němcová, Y., P. Bulant & J. Kristiansen, 2011. Mallomonas solea–ferrea and Mallomonas siveri (Chrysophyceae/Synurophyceae): two new taxa from the Western Cape (South Africa). Nova Hedwigia 93: 375–384.
Němcová, Y., M. Pusztai, M. Škaloudová & J. Neustupa, 2016. Silica-scaled chrysophytes (Stramenopiles, Ochrophyta) along a salinity gradient: a case study from the Gulf of Bothnia western shore (northern Europe). Hydrobiologia 764: 187–197.
Němcová, Y., J. Faturova & J. Kreidlova, 2022. The new species of Mallomonas (Stramenopiles, Chrysophyceae, Synurales): Mallomonas decora sp. nov. Nova Hedwigia 114: 1–8.
Nicholls, K. H., 2001. New and little-known Mallomonas (Synurophyceae) taxa from Ontario, Canada. Nordic Journal of Botany 21: 551–560.
Nicholls, K. & J. Gerrath, 1985. The taxonomy of Synura (Chrysophyceae) in Ontario with special reference to taste and odour in water supplies. Canadian Journal of Botany 63: 1482–1493.
Nicholls, K. H. & D. E. Wujek, 2015. Chrysophyceae and Phaeothamniophyceae. In Wehr, J. D., R. G. Sheath & P. J. Kociolek (eds), Freshwater algae of North America – ecology and classification Academic Press, San Diego: 537–586.
Officer, C. & J. Ryther, 1980. The possible importance of silicon in marine eutrophication. Marine Ecology Progress Series 3: 83–91.
Olrik, K., 1998. Ecology of mixotrophic flagellates with special reference to Chrysophyceae in Danish lakes. Hydrobiologia 370: 329–338.
Olsen, N. E., L. K. Poulsen, N. Reuss & S. S. Steinarsdottir, 1999. A new subspecies of Paraphysomonas punctata (Paraphysomonadaceae, Chrysophyceae). Nordic Journal of Botany 19: 635–640.
Padisák, J., G. Vasas & G. Borics, 2016. Phycogeography of freshwater phytoplankton – traditional knowledge and new molecular tools. Hydrobiologia 764: 3–27.
Park, S., M. T. Brett, D. C. Müller-Navarra, S.-C. Shin, A. M. Liston & C. R. Goldman, 2003. Heterotrophic nanoflagellates and increased essential fatty acids during Microcystis decay. Aquatic Microbial Ecology 33: 201–205.
Peltomaa, E., M. D. Johnson & S. J. Taipale, 2018. Marine cryptophytes are great sources of EPA and DHA. Marine Drugs 16: 3.
Péterfi, L. Ş & L. Momeu, 1996. Three Mallomonas species (Synurophyceae), with special distribution patterns. Hydrobiologia 336: 143–150.
Péterfi, L. Ş & L. Momeu, 2009. Ecology and biodiversity conservation of the Romanian Synurophyceae. Contributii Botanice 44: 107–117.
Péterfi, L. S., L. Momeu, J. Padisák & V. Varga, 1998. Silica-scaled chrysophytes from permanent and temporary waters of Hortobágy, eastern Hungary. Hydrobiologia 369: 339–351.
Péterfi, S. L., L. Momeu & J. Padisák, 2001. Mallomonas acaroides Perty emend: Ivanov bloom in the Calcescu subalpine lake of the Parang Mountains. Romania. Contributii Botanice 36: 73–77.
Petrack, J., S. Jost, J. Boenigk & M. Epple, 2014. Magnesiothermic conversion of the silica-mineralizing golden algae Mallomonas caudata and Synura petersenii to elemental silicon with high geometric precision. Beilstein Journal of Nanotechnology 5: 554–560.
Petrushkina, M., E. Gusev, B. Sorokin, N. Zotko, A. Mamaeva, A. Filimonova, M. Kulikovskiy, Y. Maltsev, I. Yampolsky & E. Guglya, 2017. Fucoxanthin production by heterokont microalgae. Algal Research 24: 387–393.
Pla, S., 2001. Chrysophycean cysts from the pyrenees. Bibliotheca Phycologica Band 109, Cramer, Berlin.
Pla, S. & N. J. Anderson, 2005. Environmental factors correlated with chrysophyte cyst assemblages in low arctic lakes of south-west Greenland. Journal of Phycology 41: 957–974.
Pla, S. & J. Catalan, 2005. Chrysophyte cysts from lake sediments reveal the submillennial winter/spring climate variability in the northwestern Mediterranean region throughout the Holocene. Climate Dynamics 24: 263–278.
Pla, S., L. Camarero & J. Catalan, 2003. Chrysophyte cyst relationships to water chemistry in Pyrenean lakes (NE Spain) and their potential for environmental reconstruction. Journal of Paleolimnology 30: 21–34.
Pla-Rabés, S. & J. Catalan, 2011. Deciphering chrysophyte responses to climate seasonality. Journal of Paleolimnology 46: 139–150.
Pla-Rabés, S. & J. Catalan, 2018. Diatom species variation between lake habitats: implications for interpretation of paleolimnological records. Journal of Paleolimnology 60: 169–187.
Preisig, H. R., 1994. Siliceous structures and silicification in flagellated protists. In Wetherbee, R., Pickett-Heaps, J.D., Andersen, R.A. (eds), The Protistan Cell Surface. Springer, Vienna: 29–42.
Rajabi Islami, H., 2010. The comparison of inhibitory effects of barley and rice straws to control of algae and aquatic weeds in fish ponds, Thesis, Islamic Azad University, Iran:
Rashash, D., A. Dietrich, R. Hoehn & B. Parker, 1995. The influence of growth conditions on odor-compound production by two chrysophytes and two cyanobacteria. Water Science and Technology 31: 165–172.
Reid, P. C., C. M. Turley & P. H. Burkill, 1991. Protozoa and their role in marine processes, Springer-Verlag, Berlin.
Rengefors, K. & C. Legrand, 2007. Broad allelopathic activity in Peridinium aciculiferum (Dinophyceae). European Journal of Phycology 42: 341–349.
Řezáčová, M. & J. Neustupa, 2007. Distribution of the genus Mallomonas (Synurophyceae)–ubiquitous dispersal in microorganisms evaluated. Protist 158: 29–37.
Riccio, G., D. De Luca & C. Lauritano, 2020. Monogalactosyldiacylglycerol and sulfolipid synthesis in microalgae. Marine Drugs 18: 237.
Rice, J., C. D. O’Connor, M. A. Sleigh, P. H. Burkill, I. G. Giles & M. V. Zubkov, 1997. Fluorescent oligonucleotide rDNA probes that specifically bind to a common nanoflagellate, Paraphysomonas vestita. Microbiology 143: 1717–1727.
Riebesell, U., 2004. Effects of CO2 enrichment on marine phytoplankton. Journal of Oceanography 60: 719–729.
Sandgren, C. D., J. P. Smol & J. Kristiansen, 1995. Chrysophyte algae: ecology, phylogeny and development, Cambridge University Press, New York.
Sagova-Mareckova, M., J. Boenigk, A. Bouchez, K. Cermakova, T. Chonova, T. Cordier, U. Eisendle, T. Elersek, S. Fazi & T. Fleituch, 2021. Expanding ecological assessment by integrating microorganisms into routine freshwater biomonitoring. Water Research 191: 116767.
Sandgren, C. D., 1988. Growth and reproductive strategies of freshwater phytoplankton, Cambridge University Press, New York.
Sandhage, K. H., M. B. Dickerson, P. M. Huseman, M. A. Caranna, J. D. Clifton, T. A. Bull, T. J. Heibel, W. R. Overton & M. E. Schoenwaelder, 2002. Novel, bioclastic route to self-assembled, 3D, chemically tailored meso/nanostructures: Shape-preserving reactive conversion of biosilica (diatom) microshells. Advanced Materials 14: 429–433.
Sathasivam, R. & J.-S. Ki, 2018. A review of the biological activities of microalgal carotenoids and their potential use in healthcare and cosmetic industries. Marine Drugs 16: 26.
Saxby-Rouen, K., B. Leadbeater & C. Reynolds, 1998. The relationship between the growth of Synura petersenii (Synurophyceae) and components of the dissolved inorganic carbon system. Phycologia 37: 467–477.
Schläpfer, F. & B. Schmid, 1999. Ecosystem effects of biodiversity: a classification of hypotheses and exploration of empirical results. Ecological Applications 9: 893–912.
Schmidtko, S., L. Stramma & M. Visbeck, 2017. Decline in global oceanic oxygen content during the past five decades. Nature 542: 335–339.
Schoch, C. L., S. Ciufo, M. Domrachev, C. L. Hotton, S. Kannan, R. Khovanskaya, D. Leipe, R. Mcveigh, K. O’Neill, B. Robbertse & S. Sharma, 2020. NCBI taxonomy: a comprehensive update on curation, resources and tools. Database. https://doi.org/10.1093/database/baaa062.
Schultz, T. F., L. Egerton-Warburton, S. A. Crawford & R. Wetherbee, 2001. Identification of a 41 kDa protein embedded in the biosilica of scales and bristles isolated from Mallomonas splendens (Synurophyceae, Ochrophyta). Protist 152: 315–327.
Scoble, J. M. & T. Cavalier-Smith, 2013. Molecular and morphological diversity of Paraphysomonas (Chrysophyceae, order Paraphysomonadida); a short review. Nova Hedwigia 142: 117–126.
Scoble, J. M. & T. Cavalier-Smith, 2014. Scale evolution in Paraphysomonadida (Chrysophyceae): sequence phylogeny and revised taxonomy of Paraphysomonas, new genus Clathromonas, and 25 new species. European Journal of Protistology 50: 551–592.
Sen, B., M. T. Alp, F. Sonmez, M. A. T. Kocer & O. Canpolat, 2013. Relationship of algae to water pollution and waste water treatment. In Elshorbagy, W. & R. K. Chowdhury (eds), Water treatment InTech, Rijeka: 335–354.
Sherr, E. & B. Sherr, 1994. Bacterivory and herbivory: key roles of phagotrophic protists in pelagic food webs. Microbial Ecology 28: 223–235.
Sieburth, J. M. & P. G. Davies, 1982. The role of heterotrophic nanoplankton in the grazing and nurturing of planktonic bacteria in the Sargasso and Caribbean Sea. Annales de l’Institut océanographique 58: 285–296.
Simmatis, B., K. M. Rühland, M. Evans, C. Meyer-Jacob, J. Kirk, D. C. G. Muir & J. P. Smol, 2022. Metal contamination in alkaline Phantom Lake (Flin Flon, Manitoba, Canada) generates strong responses in multiple paleolimnological proxies. Science of the Total Environment 811: 152299.
Simpson, T. L. & B. E. Volcani, 2012. Silicon and siliceous structures in biological systems, Springer-Verlag, Berlin.
Siver, P. A., 2012. The biology of Mallomonas: morphology, taxonomy and ecology, Springer Science & Business Media, Dordrecht.
Siver, P. A., 2018. Mallomonas skogstadii sp. nov. and M. bakeri sp. nov.: two new fossil species from the middle Eocene representing extinct members of the section Heterospinae? Cryptogamie, Algologie 39: 511–524.
Siver, P. A. & J. S. Chock, 1986. Phytoplankton dynamics in a chrysophycean lake. In Kristiansen, J. & R. A. Andersen (eds), Chrysophytes: aspects and problems Cambridge University Press, Cambridge: 165–183.
Siver, P. A. & J. S. Hamer, 1992. Seasonal periodicity of Chrysopyceae and Synurphyceae in a small New England lake: implications for paleolimnological research. Journal of Phycology 28: 186–198.
Siver, P. A. & A. M. Lott, 2012. Biogeographic patterns in scaled chrysophytes from the east coast of North America. Freshwater Biology 57: 451–466.
Siver, P. A. & J. P. Smol, 1993. The use of scaled chrysophytes in long term monitoring programs for the detection of changes in lakewater acidity. Water, Air, and Soil Pollution 71: 357–376.
Siver, P. A. & A. P. Wolfe, 2009. Tropical ochrophyte algae from the Eocene of northern Canada: a biogeographic response to past global warming. Palaios 24: 192–198.
Siver, P. A., D. Kapustin & E. Gusev, 2018. Investigations of two-celled colonies of Synura formerly described as Chrysodidymus with descriptions of two new species. European Journal of Phycology 53: 245–255.
Škaloud, P., J. Kristiansen & M. Škaloudová, 2013a. Developments in the taxonomy of silica-scaled chrysophytes – from morphological and ultrastructural to molecular approaches. Nordic Journal of Botany 31: 385–402.
Škaloud, P., M. Škaloudová, M. Pichrtová, Y. Němcová, J. Kreidlová & M. Pusztai, 2013b. www.chrysophytes: EU–a database on distribution and ecology of silica-scaled chrysophytes in Europe. Nova Hedwigia 142: 141–146.
Škaloud, P., M. Škaloudová, A. Procházková & Y. Němcová, 2014. Morphological delineation and distribution patterns of four newly described species within the Synura petersenii species complex (Chrysophyceae, Stramenopiles). European Journal of Phycology 49: 213–229.
Škaloud, P., M. Škaloudová, I. Jadrná, H. Bestová, M. Pusztai, D. Kapustin & P. A. Siver, 2020. Comparing morphological and molecular estimates of species diversity in the freshwater genus Synura (Stramenopiles): a model for understanding diversity of eukaryotic microorganisms. Journal of Phycology 56: 574–591.
Smol, J. P., 1985. The ratio of diatom frustules to chrysophycean statospores: a useful paleolimnological index. Hydrobiologia 123: 199–208.
Soróczki-Pintér, É., S. Pla-Rabes, E. K. Magyari, C. Stenger-Kovács & K. Buczkó, 2014. Late quaternary chrysophycean stomatocysts in a Southern Carpathian mountain lake, including the description of new forms (Romania). Phytotaxa 170: 169–186.
Stevčić, Č, K. Pulkkinen & J. Pirhonen, 2019. Screening of microalgae and LED grow light spectra for effective removal of dissolved nutrients from cold-water recirculating aquaculture system (RAS) wastewater. Algal Research 44: 101681.
Strandberg, U., S. Taipale, M. Hiltunen, A. Galloway, M. Brett & P. Kankaala, 2015. Inferring phytoplankton community composition with a fatty acid mixing model. Ecosphere 6: 1–18.
Strecker, A. L., T. P. Cobb & R. D. Vinebrooke, 2004. Effects of experimental greenhouse warming on phytoplankton and zooplankton communities in fishless alpine ponds. Limnology and Oceanography 49: 1182–1190.
Street-Perrott, F. A. & P. A. Barker, 2008. Biogenic silica: a neglected component of the coupled global continental biogeochemical cycles of carbon and silicon. Earth Surface Processes and Landforms: The Journal of the British Geomorphological Research Group 33: 1436–1457.
Sutton, J. N., L. André, D. Cardinal, D. J. Conley, G. F. De Souza, J. Dean, J. Dodd, C. Ehlert, M. J. Ellwood, P. J. Frings & P. Grasse, 2018. A review of the stable isotope bio-geochemistry of the global silicon cycle and its associated trace elements. Frontiers in Earth Science 5: 112.
Szczerba, A., S. Pla-Rabés, M. Żarczyński & W. Tylmann, 2021. The relationship between chrysophyte cyst assemblages and meteorological conditions: evidence from a sediment-trap study in northeast Poland. Ecological Indicators 133: 108395.
Taipale, S. J., S. L. Aalto, A. W. Galloway, K. Kuoppamäki, P. Nzobeuh & E. Peltomaa, 2019. Eutrophication and browning influence Daphnia nutritional ecology. Inland Waters 9: 374–394.
Terrado, R., A. L. Pasulka, A. A. Lie, V. J. Orphan, K. B. Heidelberg & D. A. Caron, 2017. Autotrophic and heterotrophic acquisition of carbon and nitrogen by a mixotrophic chrysophyte established through stable isotope analysis. The ISME Journal 11: 2022–2034.
Thomaz, S. M., 2021. Ecosystem services provided by freshwater macrophytes. Hydrobiologia. https://doi.org/10.1007/s10750-021-04739-y.
Tréguer, P., C. Bowler, B. Moriceau, S. Dutkiewicz, M. Gehlen, O. Aumont, L. Bittner, R. Dugdale, Z. Finkel & D. Iudicone, 2018. Influence of diatom diversity on the ocean biological carbon pump. Nature Geoscience 11: 27.
Tréguer, P. J., J. N. Sutton, M. Brzezinski, M. A. Charette, T. Devries, S. Dutkiewicz, C. Ehlert, J. Hawkings, A. Leynaert, S. M. Liu, L. N. Monferrer, M. López-Acosta, M. Maldonado, S. Rahman, L. Ran & O. Rouxel, 2021. Reviews and syntheses: the biogeochemical cycle of silicon in the modern ocean. Biogeosciences 18: 1269–1289.
Unrein, F., R. Massana, L. Alonso-Sáez & J. M. Gasol, 2007. Significant year-round effect of small mixotrophic flagellates on bacterioplankton in an oligotrophic coastal system. Limnology and Oceanography 52: 456–469.
Vad, C. F., C. Schneider, D. Lukić, Z. Horváth, M. J. Kainz, H. Stibor & R. Ptacnik, 2020. Grazing resistance and poor food quality of a widespread mixotroph impair zooplankton secondary production. Oecologia 193: 489–502.
Vad, C. F., C. Schneider, R. Fischer, M. J. Kainz & R. Ptacnik, 2021. From adverse to beneficial: Contrasting dietary effects of freshwater mixotrophs on zooplankton. Freshwater Biology 66: 2272–2282.
Van den Hoek, C., D. G. Mann & H. M. Jahns, 1995. Algae: an introduction to phycology, Cambridge University Press, Cambridge.
Varshney, P., P. Mikulic, A. Vonshak, J. Beardall & P. P. Wangikar, 2015. Extremophilic micro-algae and their potential contribution in biotechnology. Bioresource Technology 184: 363–372.
Vaughn, C., 2018. Ecosystem services provided by freshwater mussels. Hydrobiologia 810: 15–27.
Vigna, M. S. & P.A. Siver, 2003. Biodiversity and biogeographical implications of silica-scaled chrysophytes (Chrysophyceae and Synurophyceae) of the northeast wetlands of Argentina. Archiv für Hydrobiologie 158: 359–372.
Vincent, W. F., 1983. Phytoplankton production and winter mixing: contrasting effects in two oligotrophic lakes. Journal of Ecology 71: 1–20.
Vorobyova, S. S., G. V. Pomazkina, E. Y. Baranova, Y. V. Likhoshway & C. D. Sandgren, 1996. Chrysophycean cysts (stomatocysts) from Lake Baikal and Irkutsk reservoir, Siberia. Journal of Paleolimnology 15: 271–277.
web2. www.bccresearch.com
web6. https://sciencephoto.printstoreonline.com/synura-golden-brown-algae-6295467.html?prodid=80876
web7. https://fineartamerica.com/featured/golden-algae-dinobryon-lm-frank-fox.html?product=face-mask-flat
web8. https://lottadjossou.com/products/collier-algue-doree?lang=en
Wee, J. L., S. A. Harris, J. P. Smith, C. P. Dionigi & D. F. Millie, 1994. Production of the taste/odor-causing compound, trans-2, cis-6-nonadienal, within the Synurophyceae. Journal of Applied Phycology 6: 365–369.
Wejnerowski, L., T. O. Aykut, A. Pelechata, M. Rybak, T. Dulic, J. Meriluoto & M. K. Dziuba, 2022. Plankton hitch-hikers on naturalists’ instruments as silent intruders of aquatic ecosystems: current risks and possible prevention. NeoBiota 73: 193–219.
Wetzel, R. G., 2001. Limnology: lake and river ecosystems, Academic Press, New York:
Wilkinson, A. N., B. Zeeb & J. P. Smol, 2001. Atlas of chrysophycean cysts, vol. II. Kluwer Academic Publishers, Dordrecht.
Williams, P., K. Heinemann, J. Marra & D. A. Purdie, 1983. Comparison of 14C and O2 measurements of phytoplankton production in oligotrophic waters. Nature 305: 49–50.
Worm, B., E. B. Barbier, N. Beaumont, J. E. Duffy, C. Folke, B. S. Halpern, J. B. Jackson, H. K. Lotze, F. Micheli & S. R. Palumbi, 2006. Impacts of biodiversity loss on ocean ecosystem services. Science 314: 787–790.
Watson, S. B., 2003. Cyanobacterial and eukaryotic algal odour compounds: signals or by products? A review of their biological activity. Phycologia 42: 332–350.
Watson, S., T. Satchwill, E. Dixon & E. McCauley, 2001a. Under-ice blooms and source-water odour in a nutrient-poor reservoir: biological, ecological and applied perspectives. Freshwater Biology 46: 1553–1567.
Watson, S., T. Satchwill & E. McCauley, 2001b. Drinking water taste and odour: a chrysophyte perspective. Nova Hedwigia Beiheft 122: 119–146.
Wujek, D. E., E. M. Wright & G. L. Williams, 2003. Mallomonas weei (Synurophyceae), a new species from North Carolina, USA. Nordic Journal of Botany 23: 261–264.
Wysokowski, M., T. Jesionowski & H. Ehrlich, 2018. Biosilica as a source for inspiration in biological materials science. American Mineralogist: Journal of Earth and Planetary Materials 103: 665–691.
Xing, A., J. Zhang, Z. Bao, Y. Mei, A. S. Gordin & K. H. Sandhage, 2013. A magnesiothermic reaction process for the scalable production of mesoporous silicon for rechargeable lithium batteries. Chemical Communications 49: 6743–6745.
Yan, C., J.-H. Li, J.-J. Li, J. Wang & Y.-P. Weng, 2009. A heterotrophic nanoflagellate grazing on the toxic Cyanobacterium Microcystis aeruginosa. Annales de Limnologie – International Journal of Limnology 45: 23–28.
Yang, X., X. Wang, B. Zhao & Y. Li, 2014. Simulation model of pyrolysis biofuel yield based on algal components and pyrolysis kinetics. BioEnergy Research 7: 1293–1304.
Zeeb, B. A. & J. P. Smol, 2002. Chrysophyte scales and cysts. In Smol, J. P., H. J. B. Birks, W. M. Last, R. S. Bradley & K. Alverson (eds), Tracking environmental change using lake sediments: developments in paleoenvironmental research Springer, Dordrecht: 203–223.
Zhang, W., H. Yang, X. Xia, L. Xie & G. Xie, 2016. Triassic chrysophyte cyst fossils discovered in the Ordos Basin, China. Geology 44: 1031–1034.
Acknowledgements
TEM studies were mainly performed at Nanolab, the electron microscopy laboratory of the University of Pannonia. We thank the National Research, Development and Innovation Office and Széchenyi Plan Plus program for the supports.
Funding
Open access funding was provided by University of Pannonia. This study and its authors were supported by the National Research, Development and Innovation Office through the following NKFIH projects: K137950 (CSK), K119208 (KB) and KKP 144068 (JP, CSK, EL). The research presented in the article was carried out within the framework of the Széchenyi Plan Plus program with the support of the RRF 2.3.1 21 2022 00008 project (JP, CSK, DL, EL).
Author information
Authors and Affiliations
Contributions
EL and KB designed the structure of the paper, wrote parts and organised the cooperation among the other co-authors. SB, DL, JP and CSK wrote sections of this paper and participated actively in its modifications, corrections, and improvements.
Corresponding author
Ethics declarations
Conflict of interest
The co-author Judit Padisák is an associate editor of the Hydrobiologia, moreover she is a guest editor for this special volume (“Ecosystem services”). Therefore, she cannot participate in processing this paper.
Ethical approval
N.A.
Additional information
Handling editor: Sidinei M. Thomaz
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Guest editors: Verónica Ferreira, Luis Mauricio Bini, Katya E. Kovalenko, Andre A. Padial, Judit Padisák & María de los Ángeles González Sagrario / Aquatic Ecosystem Services
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lengyel, E., Barreto, S., Padisák, J. et al. Contribution of silica-scaled chrysophytes to ecosystems services: a review. Hydrobiologia 850, 2735–2756 (2023). https://doi.org/10.1007/s10750-022-05075-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-022-05075-5