
Abstract
Laterality, the tendency of animals to use the structures on one side of the body more frequently or efficiently than the other side, can reflect the evolutionary history of species, their ecological characteristics, and cognitive abilities. Using anurans as a model system, it has been hypothesized that the strength of laterality (measured as the percentage of right-handed individuals) is higher in terrestrial and arboreal species, which exhibit asynchronous locomotion type (e.g., walking, climbing), than in aquatic species where synchronous locomotion type (e.g., swimming) predominates. We tested these predictions, including body size as an additional factor, in 19 Neotropical anuran species. For this, we obtained data on forelimb use preference for individuals of such species. We then combined these data with published data for another 14 anuran species obtained through a systematic review of the literature. The combined dataset was used to perform a phylogenetic comparative analysis to verify the strength of laterality in the species. We found that four out of the 19 tested species show a percentage of right-handed individuals that was higher or lower than expected by chance. Individuals of these four species exhibit asynchronous movements of forelimbs during displacements, foraging behaviors, sexual displays, or agonistic interactions. Results of a phyloANOVA and a phyloANCOVA indicated that the strength of laterality was higher in species with asynchronous movements, but this was unrelated to microhabitat and body size. Asynchronous (i.e., non-simultaneous) movements of limbs imply alternating muscle contractions and a higher neural asymmetry than synchronous (i.e., simultaneous) movements of limbs. Therefore, species with an asynchronous locomotion type could be reflecting such complexity and neural asymmetry in higher levels of forelimb laterality. Our results mostly support the predictions based on models that cover the expected relationship between the strength of laterality and the evolutionary ecology of vertebrates.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Laterality refers to the tendency of animals to use the structures on one side of the body more frequently or efficiently than the other side (Bianki 1988; Bisazza et al. 1998). It relates to the ability of individuals to perform different functions simultaneously, which can influence their fitness (Rogers 2000; Rogers and Andrew 2002; Ferrari et al. 2015), for example, feeding while being attentive to the presence of predators (Rogers et al. 2004), handling food while holding a tool (McGrew and Marchant 1992), and swimming or flying while sleeping (Mascetti 2016). Laterality is associated with a functional difference between the brain’s hemispheres (Estévez-González 1992; Vallortigara et al. 1999). The right hemisphere of the brain is associated with rapid behavioural responses that individuals should have when facing stimuli such as predators or competitors’ attacks (Davidson 1992; Forrester et al. 2018). On the contrary, the left hemisphere of the brain is associated with analytical functions such as tool use, remembering cues in the habitat, and attacking strategies (Robins 2006; Forrester et al. 2018). In general, laterality reflects the evolutionary history of the species and multiple ecological factors that individuals experience during ontogeny (Rogers et al. 2013; Ferrari et al. 2015; Güntürkün et al. 2020), and its study is one of the diverse approaches to understanding the function of the animal mind and its cognitive capacity (Brown and Magat 2011; Rogers 2021).
Due to selective pressure, the predominant use of one side of the body by individuals can be significantly higher or lower than expected by chance (Bisazza et al. 1998; Frasnelli and Vallortigara 2018). The strength of laterality in animals (e.g., the percent of right-handed individuals in a population) could be influenced by the characteristics of their microhabitat and the predominant locomotion movements (Malashichev 2006, 2016). Specifically, the size, slope, and type of substrate that individuals use can affect the kinematics of their movements, thus selecting for specific morphologies, behaviors, and adaptive locomotion patterns (Losos 2009; Herrel et al. 2013; Moen et al. 2016; Fratani et al. 2020). Concerning locomotion type, asynchronous (i.e., non-simultaneous) movements of limbs such as walking and climbing, involve alternating muscle contractions and are associated with higher neural asymmetry compared to synchronous (i.e., simultaneous) movements of limbs, such as swimming and hopping (Syme and Josephson 2002; Malashichev 2006). Therefore, it is expected that species with asynchronous movement exhibit a higher strength of laterality than species with synchronous movements of their limbs (Giljov et al. 2015; Malashichev 2016).
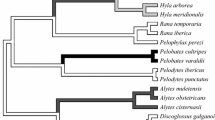
On account of the high diversity of ecological characteristics and life history traits in anurans, Malashichev and Robins (2018) used anuran species as a model to test the expected relationships between the strength of laterality, microhabitat preferences, and locomotion type in animals (Fig. 1). These authors hypothesized that if anurans are aquatic, they tend to exhibit synchronous movements of their limbs during swimming (but see Abourachid and Green 1999 for some exceptions); if anurans are terrestrial, they show both synchronous (hopping) and asynchronous (walking) movements of their limbs; and if anurans are arboreal, they tend to have asynchronous movements of their limbs (walking, climbing) (Abourachid and Green 1999; Malashichev 2006; Sullivan 2018; Fig. 1). Therefore, anurans living in aquatic microhabitat may show less strength of laterality than anurans living in terrestrial and arboreal microhabitats (Malashichev and Robins 2018). Fossorial species exhibit a different pattern, displaying synchronous movements in the forelimbs, but asynchronous movements in the hindlimbs during dragging (Malashichev 2006; Malashichev and Robins 2018). In fact, in the fossorial species Ceratophrys ornata and Pelobates fuscus there is not laterality in the use of forelimbs, but in hindlimbs (Malashichev 2006). Regarding semi-arboreal species, some of them may be predominantly hoppers, while others are walkers or climbers (Duellman 2001, 2005; Vargas-Salinas et al. 2019).

Graphical representation of the hypothesized relationship between the strength of laterality (percentage of right-handed individuals) in the snout-wiping test for anuran species, according to microhabitat (aquatic, semi-aquatic, fossorial, terrestrial, semi-arboreal, arboreal) and predominant locomotion type (synchronous, asynchronous). The figure is adapted from Malashichev and Robins (2018). The grey areas enclose species belonging to each microhabitat type. The colored lines on the right side of the panel indicate the locomotion types that can be present in species using a given microhabitat type. For example, the microhabitat “Terrestrial” can include species that are predominantly walkers or hoppers. The triangle on the left side of the panel represents the hypothesized highest strength of laterality at the base (toward species with asynchronous movement of limbs) and a reduction in the strength of laterality towards the top (species with synchronous movement of limbs). Black triangles indicate the strength of laterality for 14 species according to published literature (Bisazza et al. 1996; Rogers 2002; Malashichev 2006; Robins and Rogers 2006; Sovrano 2007; Sullivan 2018; Robins and Rogers 2021). Open red circles indicate the strength of laterality for 19 Neotropical species (this study)
The model proposed by Malashichev and Robins (2018) offers testable predictions about the strength of laterality in anurans; however, there are limitations to it. First, it is supported by only 12 anuran species, most of them from temperate regions (Fig. 1). Second it does not include body size, which is a pivotal phenotypic trait in many ecological and evolutionary aspects of anurans (Wells 2007; Amado et al. 2019). In the present study, we evaluated the strength of laterality in forelimbs of 20 neotropical anuran species. Consistent with the model by Malashichev and Robins (2018), we expected that terrestrial and arboreal species with asynchronous movements would exhibit greater strength of laterality in forelimbs compared to aquatic species. In addition, we pooled our data with those from previous studies on anurans (e.g., Bisazza et al. 1996; Malashichev 2006; Malashichev and Deckel 2006; Robins and Rogers 2021) to test a possible relationship between strength of laterality in forelimbs, with microhabitat differences, locomotion type and body size. We are aware that the absence of laterality in forelimbs does not preclude laterality in other body parts and behaviours (Rogers 2014; Güntürkün et al. 2020). However, investigating relationships regarding forelimb laterality is an important step for understanding how behaviour and habitat use relates to the cognitive capacity of anurans and other vertebrates.
Materials and methods
Data collection
Between August 2019 and April 2023, we conducted 35 field trips to 10 localities in Colombia (seven in the central Andes, one in the Pacific region, one in the Sierra Nevada of Santa Marta in the Caribbean region, and one in the Valle of Magdalena Medio). These localities are characterized by their humid habitats and high diversity of anurans, including aquatic, semi-aquatic, fossorials, terrestrial, semi-arboreal and arboreal species. We collected data from all available microhabitats. In each locality, we searched randomly for anurans during diurnal (06:30 to 10:30 and 16:00 to 17:30) and nocturnal (18:30 to 02:00) visual encounter surveys without restrictions (Heyer et al. 1994; Angulo et al. 2006). Species identification was based on morphology using local/regional field guides and taxonomic descriptions (e.g., Vargas-Salinas and Bolaños-Lizalda 1999; Acosta-Galvis et al. 2006; Bolívar et al. 2010; Domínguez et al. 2010; Pérez-González et al. 2016; Vanegas-Guerrero et al. 2016; Vargas-Salinas et al. 2019). We followed the taxonomic nomenclature of Frost (2023). We only captured mature individuals for our experiment. They were recognized by their advertisement calls in males, the presence of eggs in the belly of females, or determined by body size. To avoid pseudoreplication, within the same species we distinguish among individuals based on their unique coloration patterns (Donnelly et al. 1994; Ferner 2010). So, during the experiment, each individual was tested only once. For species whose individuals show no difference in coloration pattern, individuals were collected and placed in wet bags until additional individuals were found. After capturing an individual, we recorded its body size (snout-vent length, SVL) using a digital caliper and collected additional information about its microhabitat (aquatic, semi-aquatic, fossorial, terrestrial, semi-arboreal, arboreal). The locomotion type (synchronous, asynchronous) was determined based on literature and personal observations in the field.
The methodology described below was carried out in the field at a pre-selected site prior to capturing the individuals. To assess the strength of laterality in forelimbs of anurans (Fig. 2), we followed the methodology proposed by Bisazza et al. (1996). Prior to the experiment, we placed each individual on a horizontal surface (Fig. 2a), and used a small opaque plastic container to enclose it for five minutes (Fig. 2b). Then, we handled one individual at a time, while being handled, the subject’s limbs were restricted from movement to control for possible biases in the use of their limbs for snout-wiping during the experiments (Malashichev and Nikitina 2002). We placed a wet microfiber cloth covering its head and eyes (Fig. 2c). The size of the cloth was adjusted to the head size of each individual. The experiments began by placing the individual on a horizontal surface and recording the first forelimb it used to remove the cloth from its head (Fig. 2d). The percentage of individuals using the right forelimb to remove the cloth served as a proxy for the strength of laterality (Robins et al. 1998; Malashichev and Nikitina 2002). After the experiments, all individuals were released back into the site of capture. For each species, the experiment was conducted in a single locality to control for possible interpopulation variation in the strength of laterality (Frasnelli and Vallortigara 2018; Vallortigara, personal communication).

Methodology used to quantify the percentage of forelimb right-handed anurans (Bisazza et al. 1996). After placing an individual on a horizontal and smooth platform a, he was covered with an opaque plastic container to favor coupling to the experimental arena b. After five minutes, the plastic container was removed, the individual was taken, and a wet microfiber cloth was placed over his head, covering his eyes c. We recorded the first forelimb used by the individual to remove the microfiber cloth d. The red arrow in the last image indicates the movement that would be recorded for the focal individual (in this example: left forelimb). Two videos (Appendix I) shown the use of forelimbs by individuals of Leucostethus brachistriathus (Dendrobatidae) and Atelopus laetissimus (Bufonidae) under snout-wiping testing conditions, as well as Andinobates bombetes (Dendrobatidae) under natural conditions
For expanding our dataset, we performed a systematic review of the literature on the strength of laterality of forelimbs in anurans. The literature search was conducted using Google Scholar, Web of Science, and Scopus platforms using the keywords “lateralization”, “asymmetry”, “locomotion”, “frogs”, and “behavior”. We searched for papers written in English and Spanish languages.
Statistical analysis
To determine whether populations of our target species tend to have a higher or lower percentage of right or left-handed individuals than expected by chance (50%), we performed a two-tailed binomial test (Wagner-Menghin 2005; Whitlock and Schluter 2015). To test for a relationship between the strength of laterality in forelimbs and the microhabitat type, locomotion type, and body size, we transformed the percent of forelimb right-handed individuals per species (strength of laterality) into a “delta value of laterality”. This delta value represents the absolute difference between the recorded percentage of right-handed individuals and the expected percentage of right-handed individuals under the null hypothesis (i.e., 50%). This transformation was necessary because laterality in forelimb can be two-tailed biased (left- or right-handed), and analyzing the statistical data using the percentage of right-handed individuals alone could mask patterns if there are right-handed and left-handed species.
Given that species cannot be considered as independent statistical units (Felsenstein 1985; Revell and Harmon 2022), all of the statistical analysis were corrected for the phylogenetic relationship between species (Jetz and Pyron 2018). These analyses were conducted by pooling delta values of laterality obtained from our experiments and those found in literature. Genetic information was not available for the common toad Rhinella horribilis; therefore, we excluded this species from the following analyses. Due to the small sample size (number of species), which limited our ability to include all potential explanatory variables (body size, microhabitat type, locomotion type, and interactions), we initially tested the relationship between microhabitat type and delta values of laterality using a phylogenetic ANOVA (Revell 2012). In this analysis, the number of levels in the categorical variable “Microhabitat type” was reduced from six (Fig. 1) to three: (i) aquatic, comprising aquatic and semiaquatic species; (ii) terrestrial, referring to both terrestrial and fossorial species, and (iii) arboreal, putting together arboreal and semi-arboreal. The relationship between body size and locomotion type with delta values of laterality in forelimb was tested using a phylogenetic ANCOVA (Fuentes-G et al. 2016). These two comparative phylogenetic analyses were performed using the functions ape (Paradis et al. 2004), geiger (Harmon et al. 2008), phangorn (Schliep 2011), nlme (Pinheiro et al. 2023), and phytools (Revell 2012) with the packages “geiger”, “ape” and “phytools” (Harmon 2018; Revell and Harmon 2022) from R software (v. 3.6.1, R-Development-Core-Team, 2019).
Results
We conducted our laterality experiments on 338 individuals belonging to 19 anuran species: one semi-aquatic, nine terrestrial, one semi-arboreal, and eight arboreal (Table 1). The results of the binomial test indicate that populations of four species (Fig. 3) exhibit a higher proportion of left- or right-handed individuals than expected by chance (Table 1): most individuals of Andinobates bombetes, and Dendropsophus microcephalus used their right hands, while in Leucostethus brachistriatus and Atelopus laetissimus most individuals used their left hands. The other 16 tested species did not show a laterality tendency that significantly differs from that expected by chance (Table 1).

Images of four anuran species that exhibit a statistically significant difference or tendency (at alpha = 0.05) to use the right or left forelimb more frequently in snout-wiping experiments than expected by chance: a Andinobates bombetes, b Leucostethus brachistriatus, c Atelopus laetissimus, and d Dendropsophus mirocephalus
In the systematic review, we found 26 papers (published between 1992 and 2023) in which authors employed a similar methodology to study anurans as in our study (Table 1). In these papers, the strength of laterality was recorded for 14 anuran species, and included information on the type of microhabitat in which they were reported (one aquatic, four semi-aquatic, two fossorial, five terrestrial, and two arboreal, Table 1). The phylogenetic ANOVA revealed that delta values of laterality did not differ significantly between aquatic, terrestrial, and arboreal species (phylANOVA F = 3.09, p = 0.37, N = 32 species). Results of the phylogenetic ANCOVA (Table 2) indicates that species with asynchronous limb movements exhibit higher strength of laterality compared to species with synchronous limb movements, while body size is not related to delta values of laterality. The interaction between body size and locomotion types did not have a significant effect on delta values of laterality (Table 2).
Discussion
We found that four out of the 19 anuran species tested in this study exhibited a higher proportion of left- or right-handed individuals than expected by chance. These four species were Andinobates bombetes, Leucostethus brachistriatus (Dendrobatidae), Atelopus laetissimus (Bufonidae), and Dendropsophus microcephalus (Hylidae). When we pooled our results with those obtained in similar studies (total = 32 species excluding R. horribilis), we found that anurans with asynchronous movements showed a higher strength of laterality in the use of their forelimbs compared to anurans with synchronous movements. The strength of laterality in forelimbs was not related to microhabitat or body size of the species.
In quadrupeds, asynchronous limb movements are associated with greater functional asymmetry in the neural systems compared to organisms with synchronous limb movements (Mikliaeva et al. 1987; Forward et al. 1962; Malashichev 2006). In turn, this would mean that a species with a tendency to use limbs unilaterally will exhibit a higher strength of laterality (Mikliaeva et al. 1987; Malashichev 2006). The results of the phylogenetic ANCOVA performed with the pooled data of 32 anuran species, support this statement. Moreover, the four species in which we detected a significant strength of laterality (D. microcephalus, A. bombetes, L. brachistriatus, A. laetissimus) exhibit asynchronous movements in the use of their forelimbs.
In the tree frog D. microcephalus (right-handed), the individuals inhabit areas with temporary or semi-permanent bodies of water. While individuals jump or climb grasses and thin branches of shrubs, it is possible to observe the asynchronous movement of their limbs. In the case of the two dendrobatid species, A. bombetes (right-handed) and L. brachistriatus (left-handed), they inhabit leaf litter mounds and cavities under trunks or rocks on the forest floor (Lötters et al. 2007; Vargas-Salinas et al. 2014). Due to the numerous physical obstacles in and under such substrates (e.g., small stones, fallen branches, irregularities of the ground, leaf litter) these species predominantly engage in small walks (personal observations). Additionally, as is typical in dendrobatids, these species exhibit active foraging, conducting short walks in search of food (Bourne 2001; Staudt et al. 2010; Duarte-Marín et al. 2020) and engage in physical combat where asynchronous limb movements are evident (Pröhl 2005; Lötters et al. 2007; Wells 2007; Duarte-Marín et al. 2020).
Like dendrobatids, harlequin toads (genus Atelopus, Bufonidae) walk on the ground or climb understory vegetation (Lötters 1996; personal observations). Males of Atelopus also engage in visual displays (semaphoring) to attract mates (Lindquist and Hetherington 1998; Hödl and Amézquita 2001; Rößler et al. 2019). Both walking and visual displays involve asynchronous movements of the limbs. Therefore, in line with the expected relationship between asynchronous movements and the strength of laterality in anurans (Malashichev and Robins 2018; Fig. 1), 18 out of 24 individuals (75%) of A. laetissimus used their left forelimb to remove the microfiber in our experiments. The 80% of A. spurrelli individuals also used their left forelimb for snout-wiping, although this tendency was not statistically significant in the Binomial test, possibly because the small sample size (N = 10 individuals). The natural history characteristics of species in the family Dendrobatidae and the genus Atelopus (Bufonidae) suggest that they frequently exhibit asynchronous movements of their forelimbs, and hence, it is expected that they exhibit a clear tendency to be left- or right-handed. Future studies are required to corroborate this hypothesis.
According to Malashichev and Robins (2018), anurans in aquatic microhabitats exhibit a lower interspecific variability in the strength of forelimb laterality than anurans in terrestrial and arboreal microhabitats (Fig. 1). However, the results of the phylogenetic ANOVA indicate that there is no relationship between the microhabitat type and the strength of forelimb laterality in anurans. This discrepancy with the predictions of the model by Malashichev and Robins (2018) may occur because these authors only had information for 12 species and linked a specific locomotion type to a specific microhabitat type. For instance, they mention that terrestrial species tend to be hoppers while arboreal species are climbers. Since we included a greater number of species to test such a model (i.e., 32 species), a mismatch between locomotion type and microhabitat type arises. That is, within one microhabitat type, there are species with synchronous and species with asynchronous locomotion types (Enríquez-Urzelai et al. 2015).
Body size was unrelated to the strength of laterality, which was unexpected because in vertebrates, body size is known to be related to many physiological, ecological, and life history traits of species (Schmidt-Nielsen 1984; Wells 2007). Malashichev and Robins (2018) mention that Bufotes viridis (a terrestrial anuran), does not exhibit the same strength of laterality in forelimbs as other bufonids because individuals of this species are small compared to other representatives of the family Bufonidae. Hence, B. viridis tend to jump rather than walk. With this statement, those authors suggest that body size could be related to locomotion type in anurans. However, Enríquez-Urzelai et al. (2015) and Citadini et al. (2018) reported for 17 Mediterranean and 64 tropical anuran species, respectively, that there was no relationship between body size and locomotion type. The absence of this relationship can be attributed to the allometry between body size and limb size (Collar et al. 2011; Enríquez-Urzelai et al. 2015). Arboreal and aquatic species have relatively longer fore and hindlimbs than terrestrial and fossorial species regardless of body size differences (Enríquez-Urzelai et al. 2015; Fabrezi et al. 2017; Citadini et al. 2018; Moen 2019). Given that we find a relationship between the strength of laterality in forelimb with locomotion type, but not with body size, our results support the findings by Enríquez-Urzelai et al. (2015) and Citadini et al. (2018). That is, the strength of laterality in anurans may be more related to relative size of limbs or a set of morphological traits that define body architecture of species (i.e., ecomorphs) rather than just body size (Losos 2009; Moen et al. 2016, Moen and Wiens 2017).
We are aware of potential limitations in our study. First, laterality was not assessed at the individual level (e.g., repeated use of the same forelimb for snout wiping by an individual), but rather at the population level. It is possible that each individual has a strong tendency to be right-or left-handed, but if a similar number of individuals in the sample are right- or left-handed, statistical tendences may not be detected in studies like ours. However, it has been found that laterality recorded at the individual level is correlated with laterality at the population level (Frasnelli and Vallortigara 2018). Second, we tested the potential relationship between the strength of laterality in forelimbs and body size (measured as SVL), which has not been quantitatively tested before in a set of anuran species. However, species with similar body size can exhibit divergent body architectures (ecomorphs), which in turn determine performance in various tasks (Losos 2009; Muschick et al. 2012; Moen et al. 2013). Considering that strong selection processes have driven morphological convergences in anurans that occupy similar microhabitats (Moen et al. 2016, 2017), our comparison of forelimb laterality between aquatic, terrestrial and arboreal species serves as a proxy for comparing anuran ecomorphs. Nonetheless, within the same ecomorph there can still be variability in traits; further studies on laterality that directly examine sets of morphological traits of ecomorphs and include a large number of species may reveal new relationships (Moen 2019).
This study documents some of the highest strengths of laterality recorded to date in forelimb use for snout-wiping in anurans (Robins et al. 1998, Malashichev 2002, 2006), and to the best of our knowledge, is the first one for neotropical anuran species (but see Anderson et al. 2021). Our results mostly align with the predictions of the model hypothesized by Malashichev and Robins (2018) and demonstrate that despite advances in our understanding of the interplay between ecology and laterality in animals (Rogers and Andrew 2002; Csermely and Regolin 2013; Rogers et al. 2013), there are still many gaps that need to be addressed. For instance, the right hemisphere and the left hemisphere of the brain in vertebrates control different types of activities and abilities (Robins 2006; Csermely and Regolin 2013). Therefore, the fact that some species, even within the same family (e.g., Dendrobatidae), tend to be right-handed while others tend to be left-handed is intriguing. Testing possible causes of these contrasting results is beyond the scope of the current study, but an interaction between aposematism, predation risk, and tendencies in forelimb laterality cannot be ruled out (Anderson et al. 2021). Exploring the relationship between the strength of laterality and the asynchronous use of limbs for capturing and manipulating prey (Gray et al. 1997; Csermely and Regolin 2013) is also an aspect that has been overlooked in anurans and requires further studies (but see Bizzasa et al. 1996, 1997). Overall, the strength of laterality in animals can be influenced by numerous factors (Rogers et al. 2013; Malashichev and Robins 2018), and with this study, we hope to encourage future research on laterality and its relationship with the evolutionary ecology of neotropical anurans.
Code availability
Script used in this study is available at the FigShare data repository https://doi.org/10.6084/m9.figshare.22237471.
References
Abourachid A, Green DM (1999) Origins of the frog-kick? Alternate-leg swimming in primitive frogs, families Leiopelmatidae and Ascaphidae. J Herpetol 33:657–663
Acosta-Galvis AR, Huertas-Salgado C, Rada M (2006) Aproximación al conocimiento de los anfibios en una localidad del Magdalena medio (Departamento de Caldas, Colombia). Rev Acad Colomb Cienc 30:291–303
Amado TF, Bidau CJ, Olalla-Tárraga MÁ (2019) Geographic variation of body size in new world Anurans: energy and water in a balance. Ecography 42:456–466
Amphibiaweb, (2023) Amphibiaweb: an online referencie. University of California, Berkeley, CA, USA. Available from: https://amphibiaweb.org/index.html. Accessed september 2023.
Andreone F, Piazza R (1990) A bioacoustic study on Pelobates fuscus insubricus (Amphibia, Pelobatidae). Bolletino Zool 57:341–349
Anderson HM, Fisher DN, McEwen BL et al (2021) Episodic correlations in behavioural lateralization differ between a poison frog and its mimic. Anim Behav 174:207–215
Angulo A, Rueda-Almonacid JV, Rodríguez-Mahecha JV et al (2006) Técnicas de inventario y monitoreo para los anfibios de la región tropical andina. Conservación Internacional. Serie Manuales de Campo 2. Panamericana Formas e Impresos SA, Colombia
Bianki VL (1988) The right and left hemispheres of the animal brain: cerebral lateralization of function. Gordon and Breach, New York
Bisazza A, Cantalupo C, Robins A et al (1996) Right-pawedness in toads. Nature 379:408
Bisazza A, Cantalupo C, Robins A et al (1997) Pawedness and motor asymmetries in toads. Laterality 2:49–64
Bisazza A, Rogers LJ, Vallortigara G (1998) The origins of cerebral asymmetry in fishes, amphibians and reptiles. Neurosci Biobehav Rev 22:411–426
Bolívar GW, Eusse-González D, Castro-Herrera F et al (2010) Aves y herpetos de la reserva forestal protectora regional de Bitaco. Corporación Autónoma Regional del Valle del Cauca CVC, Fundación Calidris, y Universidad del Valle, Santiago de Cali
Bourne GR (2001) Color pattern, predator avoidance, and foraging behavior in the golden frog Colestethus beebei (Anura: Dendrobatidae). Herpetol Rev 32:225–228
Brown C, Magat M (2011) The evolution of lateralized foot use in parrots: a phylogenetic approach. Behav Ecol 22:1201–1208
Bülbül U, Eroğlu Aİ, Kurnaz M et al (2018) Age structure and some growth parameters in a population of Bombina bombina (L., 1761) from Turkey. Russ J Herpetol 25:221–226
Citadini JM, Brandt R, Williams CR, Gomes FR (2018) Evolution of morphology and locomotor performance in anurans: relationships with microhabitat diversification. J Evol Biol 31:371–381
Collar DC, Schulte IIJA, Losos JB (2011) Evolution of extreme body size disparity in monitor lizards (Varanus). Evolution 65:2664–2680
Csermely D, Regolin L (eds) (2013) Behavioral lateralization in vertebrates: two sides of the same coin. Springer, Berlin
Daugherty CH, Sheldon AL (1982) Age-determination, growth, and life history of a montana population of the tailed frog (Ascaphus truei). Herpetol 38:461–468
Davidson RJ (1992) Emotion and affective style: Hemispheric Substrates. Psychol Sci 3:39–43
Dittrich C, Rödel MO (2020) Description of female release calls of the european common frog, Rana temporaria (Anura: Ranidae). Salamandra 56:91–94
Dodd CK (2013) Frogs of the United States and Canada. Johns Hopkins University Press, Maryland
Domínguez TDM, Cortés GMA, Valencia AA, García CLM et al (2010) Guía de los anfibios y reptiles del área en conservación de la microcuenca quebrada Pericos. Corporación Autónoma del Valle del Cauca, CVC, Santiago de Cali
Donnelly MA, Guyer C, Juterbock JE et al (1994) Techniques for marking amphibians. In: Heyer R, Donnelly MA, Foster M et al (eds) Measuring and monitoring biological diversity: standard methods for amphibians. Smithsonian Institution Press, Washington, pp 277–284
Duarte-Marín S, González-Acosta CC, Santos-Dias PH et al (2020) Advertisement call, tadpole morphology, and other natural history aspects of the threatened poison frog Andinobates daleswansoni (Dendrobatidae). J Nat Hist 54:3005–3030
Duellman WE (2001) Hylid frogs of middle America. Natural History Museum of the University of Kansas, Ithaca
Duellman WE (2005) Cusco Amazónico: the lives of amphibians and reptiles in an amazonian rainforest. Natural History Museum of the University of Kansas, Lawrence
Enríquez-Urzelai U, Montori A, Llorente GA et al (2015) Locomotor mode and the evolution of the hindlimb in western Mediterrean anurans. Evol Biol 42:199–209
Estévez-González A (1992) Lateralidad y asimetría funcional cerebral: fundamentos. Rev Logop Fon Audiol 12:138–151
Fabrezi M, Goldberg J, Chuliver-Pereyra M (2017) Morphological variation in anuran limbs: constraints and novelties. J Exp Zool Mol Dev Evol 00B:1–29
Felsenstein J (1985) Phylogenies and the comparative method. Am Nat 125:1–15
Ferner JW (2010) Measuring and marking post-metamorphic amphibians. In: Dodd CK (ed) Amphibian ecology and conservation. A handbook of techniques. Oxford University Press, New York, pp 123–141
Ferrari M, McCormick MI, Allan BJ et al (2015) The effects of background risk on behavioural lateralization in a coral reef fish. Funct Ecol 29:1553–1559
Forrester G, Hudry K, Lindell A et al (eds) (2018) Cerebral lateralization and cognition: evolutionary and developmental investigations of behavioral biases. Academic Press, Cambridge
Forward E, Warren JM, Hara K (1962) The effects of unilateral lesions in sensorimotor cortex on manipulation by cats. J Comp Physiol Psychol 55:1130–1135
Frasnelli E, Vallortigara G (2018) Individual-level and population-level lateralization: two sides of the same coin. Symmetry 10:739
Fratani J, Ponssa ML, Rada M, Abdala V (2020) The influence of locomotion and habitat use on tendo-muscular units of an anuran clade (Anura, Diphyabatrachia). Zool Anz 284:66–77
Frost DR (2023) Amphibian species of the world: an online reference. Version 6.1 electronic database. American Museum of Natural History, New York. Available from https://amphibiansoftheworld.amnh.org. Accessed May 2023
Fuentes-GJA, Housworth EA, Weber A, Martins EP (2016) Phylogenetic ANCOVA: estimating changes in evolutionary rates as well as relationships between traits. Am Nat 188:615–627
Giljov A, Karenina K, Ingram J, Malashichev Y (2015) Parallel emergence of true handedness in the evolution of marsupials and placentals. Curr Biol 25:1878–1884
Goree BD, Wassersug RJ (2001) Are archeobatrachian anurans ambidextrous? Assessing handedness in Bombina orientalis. J Herpetol 35:538–541
Gray LA, O’Reilly JC, Nishikawa KC (1997) Evolution of forelimb movement patterns for prey manipulation in anurans. J Exp Zool 277:417–424
Green DM (2003) Rocky Mountain tail frog, Ascaphusmontanus. In: Hutchins M, Duellman WE, Schlager N (eds) Grzimek’s Animal Life Encyclopedia. Farmington Hills, Michigan, p 500
Güntürkün O, Ströckens F, Ocklenburg S (2020) Brain lateralization: a comparative perspective. Physiol Rev 100:1019–1063
Harmon L (2018) Phylogenetic comparative methods: learning from trees. CreateSpace Independent Publishing Platform, Scotts Valley
Harmon LJ, Weir JT, Brock CD, Glor RE, Challenger W (2008) GEIGER: investigating evolutionary radiations. Bioinform 24:129–131
Herrel A, Perrenoud M, Decamps T et al (2013) The effect of substrate diameter and incline on locomotion in an arboreal frog. J Exp Biol 216:3599–3605
Heyer R, Donnelly MA, Foster M et al (eds) (1994) Measuring and monitoring Biological Diversity: standard methods for amphibians. Smithsonian Institution Press, Washington
Hödl W, Amézquita A (2001) Visual signaling in anuran amphibians. In: Ryan MJ (ed) Anuran communication. Smithsonian Institution Press, Washington, pp 121–141
Hoser RT (1989) Australian reptiles and frogs. Pierson and Company, Sydney
Jetz W, Pyron RA (2018) The interplay of past diversification and evolutionary isolation with present imperilment across the amphibian tree of life. Nat Ecol Evol 2:850–858
Kostylev MA, Malashichev YB (2007) Correlation of the shoulder girdle asymmetry with the limb skeleton asymmetry in Xenopus laevis. Dokl Biol Sci 416:374–376
Lang M (1988) Notes on the genus Bombina oken (Anura, Bombinatoridae). Br Herpetol Soc Bull 26:3–13
Lindquist ED, Hetherington TE (1998) Semaphoring in an earless frog: the origin of a novel visual signal. Anim Cogn 1:83–87
Losos J (2009) Lizards in an evolutionary tree: ecology and adaptive radiation of anoles: organisms and environments. University of California Press, California
Lötters S (1996) The neotropical toad genus Atelopus: checklist, biology, distribution. Vences F and Glaw Verlags, Cologne
Lötters S, Jungfer KH, Henkel FW et al (2007) Poison frogs. Biology, species and captive care. Edition Chimaira, Frankfurt
Malashichev YB (2002) Asymmetries in amphibians: a review of morphology and behaviour. Laterality 7:197–217
Malashichev YB (2006) One-sided limb preference is linked to alternating-limb locomotion in anuran amphibians. J Comp Psychol 120:401–410
Malashichev YB (2016) Asymmetry of righting reflexes in sea turtles and its behavioral correlates. Physiol Behav 157:1–8
Malashichev YB, Deckel AW (eds) (2006) Behavioral and morphological asymmetries in vertebrates. Landes Bioscience, Georgetown
Malashichev YB, Nikitina NG (2002) Preferential limb use in relation to epicoracoid overlap in the shoulder girdle of toads. Laterality 7:1–18
Malashichev YB, Robins A (2018) Lateralized motor responses in anuran amphibians-an overview of methods and perspectives of studies. Biol Commun 63:210–242
Mascetti GG (2016) Unihemispheric sleep and asymmetrical sleep: behavioral, neurophysiological, and functional perspectives. Nat Sci Sleep 8:221–238
Mayer M, Hawlitschek O, Zahn A, Glaw F (2013) Composition of twenty green frog populations (Pelophylax) across Bavaria, Germany. Salamandra 49:31–44
McGrew WC, Marchant LF (1992) Chimpanzees, tools, and termites: Hand preference or handedness? Curr Anthropol 33:114–119
Mikliaeva EI, Kulikov MA, Ioffe ME (1987) Motor asymmetry of the forelimbs of the rat. Zh Vyssh Nerv Deiat Im I P Pavlova 37:254–264
Moen DS (2019) What determines the distinct morphology of species with a particular ecology? The roles of many-to-one mapping and trade-offs in the evolution of frog ecomorphology and performance. Am Nat 194:E81–E95
Moen DS, Wiens JJ (2017) Microhabitat and climatic niche change explain patterns of diversification among frog families. Am Nat 190:29–44
Moen DS, Irschick DJ, Wiens JJ (2013) Evolutionary conservatism and convergence both lead to striking similarity in ecology, morphology and performance across continents in frogs. Proc R Soc B 280:20132156
Moen DS, Morlon H, Wiens JJ (2016) Testing convergence versus history: convergence dominates phenotypic evolution for over 150 million years in frogs. Syst Biol 65:146–160
Muschick M, Indermaur A, Salzburger W (2012) Convergent evolution within an adaptive radiation of cichlid fishes. Curr Biol 22:2362–2368
Paradis E, Claude J, Strimmer K (2004) APE: analyses of phylogenetics and evolution in R language. Bioinform 20:289–290
Pérez-González JL, Mejía-Quintero LR, Jiménez-López LC et al (2016) Anfibios y reptiles de Santa Marta y sus alrededores. Universidad del Magdalena, Santa Marta
Pincheira-Donoso D, Harvey LP, Cotter SC et al (2021) The global macroecology of brood size in amphibians reveals a predisposition of low-fecundity species to extinction. Glob Ecol Biogeogr 30:1299–1310
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Development Core Team (2023) NLME: linear and nonlinear mixed effects models (R Package Version: 3.1-163, 2023). https://cran.r-project.org/web/packages/nlme/nlme.pdf. Accessed August 2023
Pröhl H (2005) Territorial behavior in dendrobatid frogs. J Herpetol 39:354–365
R Core Team (2019) R: a language and environment for statistical computing. R Found Stat Comput, Vienna
Reilly S, Essner R, Wren S et al (2015) Movement patterns in leiopelmatid frogs: insights into the locomotor repertoire of basal anurans. Behav Process 121:43–53
Revell LJ (2012) Phytools:an R package for phylogenetic comparative biology (and other things). Methods Ecol Evol 3:217–223
Revell LJ, Harmon LJ (2022) Phylogenetic comparative methods in R. Princeton University Press, New Jersey
Robins LJ (2006) Lateralized visual processing in anurans: new vistas through ancient eyes. In: Malashichev YB, Deckel AW (eds) (2006) Behavioral and morphological asymmetries in vertebrates. Landes Bioscience, Georgetown, pp 86–106
Robins A, Rogers LJ (2002) Limb preference and skeletal asymmetry in the cane toad, Bufo marinus (Anura: Bufonidae). Laterality 7:261–275
Robins A, Rogers LJ (2006) Lateralized visual and motor responses in the green tree frog, Litoria caerulea. Anim Behav 72:843–852
Robins A, Rogers LJ (2021) Lateralized motor behaviour in the righting responses of the cane toad (Rhinella marina). Laterality 27:129–171
Robins A, Rogers LJ, Vallortigara G, Rogers LJ (1998) Lateralized agonistic responses and hindlimb use in toads. Anim Behav 56:875–881
Rogers LJ (2000) Evolution of hemispheric specialization: advantages and disadvantages. Brain Lang 73:236–253
Rogers LJ (2002) Lateralized brain function in anurans: comparison to lateralization in other vertebrates. Laterality 7:219–239
Rogers LJ (2014) Asymmetry of brain and behavior in animals: its development, function, and human relevance. Genesis 52:555–571
Rogers LJ (2021) Brain lateralization and cognitive capacity. Animals 11:1996
Rogers LJ, Andrew RJ (eds) (2002) Comparative vertebrate lateralization. Cambridge University Press, Cambridge, pp 94–125
Rogers LJ, Zucca P, Vallortigara G (2004) Advantages of having a lateralized brain. Proc R Soc Lond B Biol Sci 271:S420–S422
Rogers LJ, Vallortigara G, Andrew RJ (2013) Divided brains: the biology and behaviour of brain asymmetries. Cambridge University Press, New York
Rößler DC, Lötters S, Mappes J et al (2019) Sole coloration as an unusual aposematic signal in a neotropical toad. Sci Rep 9:1128
Ron SR, Merino-Viteri A, Ortiz DA (2023) Anfibios del Ecuador. Version 2022.0. Museo de Zoología, Pontificia Universidad Católica del Ecuador. Available from: https://bioweb.bio/faunaweb/amphibiaweb. Accessed August 2023.
Schliep KP (2011) Phangorn: phylogenetic analysis in R. Bioinformatics 27:592–593
Schmidt-Nielsen K (1984) Scaling: why is animal size so important? Cambridge University Press, Cambridge
Sovrano VA (2007) A note on asymmetric use of the forelimbs during feeding in the european green toad (Bufo viridis). Laterality 12:458–463
Staudt K, Mebs D, Ospina SM, Pröhl H (2010) Foraging behaviour and territoriality of the strawberry poison frog (Oophaga pumilio) in dependence of the presence of ants. Amphib-Reptil 31:217–227
Sullivan RA (2018) Handedness is not linked to locomotion in a basal anuran. University Southern Illinois, Illinois
Syme AD, Josephson RK (2002) How to build fast muscles: synchronous and asynchronous designs. Integr Comp Biol 42:762–770
Vallortigara G, Rogers LJ, Bisazza A et al (1998) Complementary right and left hemifield use for predatory and agonistic behaviour in toads. NeuroReport 9:3341–3344
Vallortigara G, Rogers LJ, Bisazza A (1999) Possible evolutionary origins of cognitive brain lateralization. Brain Res Rev 30:164–175
Vanegas-Guerrero J, Buitrago-González W, Fernández C, Vargas-Salinas F (2016) Urban remnant forests: are they important for herpetofaunal conservation in the Central Andes of Colombia? Herpetol Rev 47:180–185
Vargas-Salinas F, Bolaños-Lizalda ME (1999) Anfibios y reptiles en hábitats perturbados de selva lluviosa tropical en el Bajo Anchicayá, Pacifico colombiano. Rev Acad Colomb Cienc 23:499–511
Vargas-Salinas F, Rodríguez-Collazos LA, Suárez-Mayorga MA (2014) Andinobates bombetes. Catálogo de Anfibios y Reptiles de Colombia 2:13–18
Vargas-Salinas F, Muñoz-Ávila JA, Morales-Puentes ME (2019) Biología en anfibios y reptiles en el bosque seco tropical del norte de Colombia. Editorial UPTC y Ecopetrol, Tunja
Wagner-Menghin MM (2005) Binomial test. In: Everitt BS, Howell DC (eds) Encyclopedia of statistics in behavioral science. Wiley, Chichester, pp 158–163
Wells KD (2007) The ecology and behavior of amphibians. University of Chicago Press, Chicago
Whitlock M, Schluter D (2015) The analysis of biological data. Roberts Publishers, Colorado
Acknowledgements
We are grateful with JD Carvajal-Castro, SM Rojas-Henao, AV Rojas-Henao, AM Mopán-Chilito, E Ochoa-Berrío, JS Salazar-Contreras, C González-Acosta, S Muñoz-Acevedo, P Navarro-Salcedo, N Vergara-Herrera, JD Barros-Castañeda, A Barros-Granados, D Gómez-Murcia, Y Aguirre-López and students of the ECCO and GHUQ research groups, and specially to Fundación Atelopus for their collaboration in field work. We are also grateful with AM Jaramillo and CA Arango-Rojas for allowing us to work with species in the Natural Reserve Rio Manso and to PH Carvajal, M Gallo, N Rivillas, D Martínez, S Canal, and J Marulanda (owners of some study areas). Thanks to S Muñoz-Acevedo and C Echeverry for sharing videos where behavior of two species with laterality is observed. We also thank to JF Aristizabal, LA Rueda-Solano, OA Obando-Aguirre, and two anonymous reviewers for comments that greatly improved previous versions of this manuscript. We extend our gratitude to G Vallortigara and the A Robins for kindly sharing studies and their expertise about laterality in animals and providing suggestion for the discussion of results. We would like to thank P Böning for improving the English of the manuscript. Finally, we thank the Biology Program at the Universidad del Quindío (Colombia) for their logistic support and research permits.
Funding
Open Access funding provided by Colombia Consortium. No funding source was received for this work.
Author information
Authors and Affiliations
Contributions
Conceptualization: RMM, VSF; Data collection: RMM; Data analysis: RMM, VSF; Writing and review manuscript drafting: RMM, VSF.
Corresponding author
Ethics declarations
Conflict of interest
The authors have not conflict of interest.
Ethical approval
All international, national, and regional guidelines for the care and manipulation of animals were followed. This study was conducted under the research permit #04601 of the Corporación Autónoma Regional del Quindío CRQ, resolution #01789 issued on October 11, 2018 by National Environmental Licensing Authority ANLA, Colombia, to Professor Fernando Vargas-Salinas, and resolution #00250 issued on February 23, 2018 by National Environmental Licensing Authority ANLA, Colombia, to Professor Luis Alberto Rueda-Solano.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1 (MP4 45648 kb)
Supplementary Material 2 (MP4 19235 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rojas-Montoya, M., Vargas-Salinas, F. Right or left-handed: are locomotion type, body size, and microhabitat related to forelimb laterality in anurans?. Evol Ecol 37, 961–980 (2023). https://doi.org/10.1007/s10682-023-10271-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10682-023-10271-3