
Abstract
Production plantation forestry has many economic benefits but can also have negative environmental impacts such as the spreading of invasive pines to native forest habitats. Monitoring forest for the presence of invasive pines helps with the management of this issue. However, detection of vegetation change over a large time period is difficult due to changes in image quality and sensor types, and by the spectral similarity of evergreen species and frequent cloud cover in the study area. The costs of high-resolution images are also prohibitive for routine monitoring in resource-constrained countries. This research investigated the use of remote sensing to identify the spread of Pinus caribaea over a 21-year period (2000 to 2021) in Belihuloya, Sri Lanka, using Landsat images. It applied a range of techniques to produce cloud free images, extract vegetation features, and improve vegetation classification accuracy, followed by the use of Geographical Information System to spatially analyze the spread of invasive pines. The results showed most invading pines were found within 100 m of the pine plantations’ borders where broadleaved forests and grasslands are vulnerable to invasion. However, the extent of invasive pine had an overall decline of 4 ha over the 21 years. The study confirmed that remote sensing combined with spatial analysis are effective tools for monitoring invasive pines in countries with limited resources. This study also provides information to conservationists and forest managers to conduct strategic planning for sustainable forest management and conservation in Sri Lanka.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Forest plantations have been established worldwide to provide timber and firewood and have significantly contributed to local and global economies. However, most of these plantations were established using tree species with fast growth rates, which differ from many naturally regenerated forests in composition and structure, leading to different ecological processes and functional outcomes (Subasinghe et al., 2014). In some environments, these fast-growing exotic tree species have become invasive, replacing existing vegetation (Dash, 2020; Rejmánek & Richardson, 2013; Richardson et al., 2007).
Notably, plantations using pine species are the highest contributor to timber production globally (Burley & Barnes, 2004; McEwan et al., 2020). These species, however, can cause many adverse effects to the environment. For example, pine species native to North America and Europe have been introduced to New Zealand, South Africa, Argentina, Brazil, and Chile as a foundation for exotic forestry enterprises (Pauchard et al., 2016a; Rejmánek, 2014; Singh et al., 2018), but are now invading significant areas such as native forests, grasslands, catchments, and protected areas, causing a potential transformative effect on native ecosystems (Dash, 2020; Richardson et al., 2007). Additionally, Medawatte et al. (2010) study shows that invasive pines can also contribute to an overall decrease in native biodiversity, suppress native plants, contribute to habitat loss for wildlife, decrease streamflow, cause changes in nutrient cycling, and affect groundwater supply levels.
Pinus patula and Pinus caribaea were introduced to Sri Lanka in 1967 to boost timber production and reduce the risk of soil erosion and landslides in the hilly regions due to their faster growth rates compared with indigenous species (Subasinghe, 2007). In particular, Pinus caribaea was used extensively for the reforestation of degraded areas in the country’s wet and intermediate climatic zones with elevations ranging from 100 to 2000 m above mean sea level due to its fast establishment and growth under adverse conditions (Edirisinghe, 2017; Jayawardhane & Gunaratne, 2020; Perera, 2001). However, their invasive behavior in Sri Lanka’s mountainous region has negatively affected the soil biodiversity and regeneration of native flora, and increased the occurrence of wildfires during the dry season (Nissanka et al., 2005; Wijerathna et al., 2016). In 2008, Pinus caribaea was identified as a potentially invasive alien species in Sri Lanka because several areas in the mid-country reported their spread (Wijesundera, 2008). As a result, the Sri Lankan government is planning to replace pine plantations with indigenous plant species (Office of the Cabinet of Ministers-Sri Lanka, 2017).
Previous studies (Bjerreskov et al., 2021; Kaplan, 2021; Liu et al., 2022) have investigated the expansion of broad leaf and conifer forests maintained and managed for commercial timber production in temperate climates where the difference is very significant due to seasonal and phenological changes. Furthermore, it is relatively easy to recognize these two types of temperate forests when coniferous species are evergreen and broad leaf trees are deciduous. Comparatively, studies conducted in tropical environments are limited in their spatial extent and are focused on the simple classification of landscapes into major land cover classes (Petersen et al., 2016). Furthermore, Pauchard et al. (2016b) found that climate-induced invasion, or the latitudinal pattern of invasion, happened all around the world in both tropical and temperate locations showing a growing need to monitor the spread of exotic species, and forecast future distribution of such invasions. Most pinus invasion studies (Pauchard et al., 2016; Weisberg et al., 2007; Xu et al., 2018), however, have been based on temperate region and largely on grasslands and shrublands. Additionally, although there are many records of exotic pines invading the natural vegetation in tropical regions (Afrin et al., 2010; Ayala et al., 2005), few studies have been performed with the use of remote sensing techniques (Amaral et al., 2015; Goncalves et al., 2022; Petersen et al., 2016). For example, only one recent study conducted in Sri Lanka has used Geographic Information System (GIS) analysis (Medawatte et al., 2010). Mapping the expansion of invasions through field surveys is time-consuming and costly; hence, there is a need to develop remote sensing methods that are both reliable and affordable, especially in developing countries.
Remote sensing is a promising tool for mapping, detecting, and monitoring invasive non-native plants across broad geographic extents. The availability of satellite image archives permits mapping non-native invasive plants spread, retrospectively (Gavier-Pizarro et al., 2012). However, most existing studies (Amaral et al., 2015; Andrew & Ustin, 2008; Dash et al., 2019; Khare et al., 2018; Piiroinen et al., 2018) have used high-resolution hyperspectral imagery covering a relatively small area and analyses over a short period of time. From a management perspective, this limits their use for long-term assessments of invasive plant spread, particularly in developing countries where the lack of historical data further compounds management efforts. On the other hand, multispectral sensors with moderate resolutions offer an alternative for low-resourced countries because they enable both change detection and measurements of phenology over multiple years (Petersen et al., 2016; Signori & Ducati, 2019; Xu et al., 2018). Additionally, these data sets are mostly free or low cost (Bradley, 2014). In particular, the Landsat program has been extensively used for forest monitoring, providing a rich dataset to map the invasion of tree species at the landscape scale. The opening of the Landsat archive and its continued free and open access since 2009 has enabled the analysis of both larger areas and more extensive times series (Zhu & Woodcock, 2014).
The current study investigates the application of Landsat satellite information to map, quantify and assess spatio-temporal land cover change using a Sri Lankan study area, namely Belihuloya, over a two-decade period (2000 to 2021). The selected study area is one of the few areas that manage pines for both conservation and commercial purposes in Sri Lanka. The Belihuloya region suffers from a shortage of groundwater caused by the water uptake from pine plantations (Starkloff, 1998). This results in seasonal wildfires, so it is therefore important that pine forest expansion is monitored. This study contributes to knowledge development by analyzing the spread of Pinus caribaea to neighboring landscapes over a 20-year period from plantations established and operated for soil conservation in Sri Lanka. Specifically, the paper seeks to answer the following research question: to what extend is the Landsat archive for Sri Lanka suitable for mapping and tracking land cover changes associated with the historical spread and invasion of conifers? To this end, this study uses Landsat archive to distinguish unmanaged pine plantations from native forests in Sri Lanka’s tropical environment.
Materials and methods
Study area
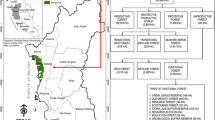
This study used images from the Landsat archive for the years 2000 and 2021 to monitor the historical spread and control of invasive exotic conifers (e.g., Pinus caribaea) in Sri Lanka’s intermediate climatic zone. Sri Lanka has traditionally been generalized into three climatic zones: the wet zone, dry zone, and intermediate zone, according to rainfall, soil, and vegetation types. The intermediate zone separates both the wet and dry zones and receives a mean annual rainfall between 1750 and 2500 mm with a short and less prominent dry season (Punyawardena, 2020). This intermediate climatic zone divides into three further zones based on altitude: low-country, mid-country, and up-country. According to the Sri Lankan climatic classification (Jayawardane & Weerasena, 2000), this study area lies in the intermediate mid-country, where the elevation ranges between 600 and 900 m above mean sea level and contains rugged topography. The study area is located in Southeast Sri Lanka and covers an area of 6234 ha. Administratively, this area belongs to the Balangoda region in the Sabaragamuwa province of Sri Lanka (Fig. 1). The area is densely forested, containing a mixture of forest types, including montane broadleaf forest, pine plantations, and grasslands. This area was selected because it comprises both unmanaged pineland and densely forested native broad leaf species.

Study area
Methods
The following section describes the methodology developed for this research. As shown in Fig. 2, the study comprised five main steps: (1) data acquisition; (2) pre-processing; (3) feature extraction; (4) multitemporal classification; and (5) post-classification.

Overall methodology
Step 1: Data acquisition
Landsat 7 Enhanced Thematic Mapper Plus (ETM +) and Landsat 8 Operational Land Imager (OLI) images from the Google Earth Engine (GEE) Landsat archive (https://developers.google.com/earth-engine/datasets/catalog/landsat) for the years 2000 and 2021 were used to characterize vegetation changes. The advantage of Landsat as a moderate spatial resolution satellite collection is that the entire scene is captured at once with sun illumination, vegetative condition, and atmospheric conditions more likely to be consistent across the scene (Labonté et al., 2020). The study period 2000 and 2021 was chosen as it is difficult to validate the classification prior to 2000 due to the absence of high-resolution images in Google Earth Pro. In 2000, only six images of Landsat 7 that covered the study area were available in the Landsat archive, and in 2021, 21 images of Landsat 8 were available. The GEE cloud-based platform was used for image pre-processing and classification (Setiani et al., 2021), while ArcGIS Pro software was used to produce maps.
The Shuttle Radar Topographic Mission (SRTM) provides digital elevation data (DEM) on a global scale. The SRTM Plus version (spatial resolution of 30 m) was acquired from GEE, and topographic features such as elevation, aspect, and slope were extracted.
Step 2: Pre-processing
As the studied region is frequently cloudy, it is difficult to acquire cloud-free Landsat images. Therefore, atmospheric correction, cloud/shadow masking, and image compositing were applied to construct a clean image time series. A cloud masking procedure was used to identify flagged cloud and cirrus pixels. After cloud removal, gap filling was applied using the median values of each pixel on the images. Then the median composite function in GEE was used to remove anomalous dark pixels (shadows) and bright, saturated pixels (Bjerreskov et al., 2021) in the collection and was used to create composite images for the years 2000 and 2021.
Step 3: Feature extraction
Feature extraction was carried out in GEE using vegetation indices derived from spectral data and tasselled cap transformation, which transforms spectral data into indicators (Dash, 2020).
Vegetation indices
Throughout all wavelengths, coniferous leaves have lower transmittance than broad leaves (Lukeš et al., 2013). Though the difference between visible (VIS) and near-infrared (NIR) spectral reflectance is not hugely evident, some studies (Roberts et al., 2004; Williams, 1991) have reported that coniferous needles have slightly lower VIS reflectance and higher NIR reflectance than broadleaves. It is hard to classify land cover or identify specific species when the vegetation types are heterogeneous and often exhibit both spectral and seasonal similarities (Deng et al., 2020). Therefore, several vegetation indexes were used as shown in Table 1. These were as follows: (1) normalized difference moisture index (NDMI); (2) normalized burn ratio (NBR); (3) enhanced vegetation index (EVI); (4) normalized difference vegetation index (NDVI); and (5) green chlorophyll index (GCI). These indices categorized vegetation changes related to moisture stress, colorization, and needle/leaf structure (Ye et al., 2021).
Tasselled cap transformation
Tasselled cap transformation (TCA) was developed by Kauth and Thomas (1976) as a function for determining a crop’s life cycle. It recognizes the pattern found in the agricultural fields where there is a net increase in near-infrared and decrease in red reflectance based on soil color.
The tasselled cap transformation is a linear transformation of Landsat MSS data that projects soil and vegetation information into a single plane in a multispectral data space. TCA has been broadly engaged in forestry studies of structure, condition, successional state, and change detection in various forest environments (Gómez et al., 2012). It is a special case of principal components analysis which transforms the image data into a new coordinate system with a new set of orthogonal axes. The tasselled cap analysis reduces a multiband data set (4–6) to 3 channels as follows:
-
Tasselled cap greenness (TCG) contrasts the near-infrared and visible bands, conveying information concerning the abundance and vigor of living vegetation.
-
Tasselled cap brightness (TCB) is a weighted sum of all four bands. Brightness is defined in the direction of the principal variation in soil reflectance and is associated with bare areas or partially covered soil.
-
Tasselled cap wetness (TCW) is related to the canopy and soil moisture which differences the sum of the visible and near-infrared bands with the longer infrared bands.
Step 4: Multitemporal classification
The supervised classification of both maps (the years 2000 and 2021) was carried out using the Random Forest (RF) classifier because of its relatively high accuracy and computational efficiency (Zhu & Woodcock, 2014). Random Forest is an integrated learning method that has become increasingly common in remote sensing applications due to its nonparametric nature and ability to limit overfitting (Cheng & Wang, 2019). The Smile Random Forest function on the GEE platform (Liu et al., 2022) was used, and the number of decision trees was set to 500. A score was calculated using the RF algorithm for each combination of parameters. The combination of parameters with the highest score was used for developing the vegetation classification maps.
Training and validation
Stratified random sampling was used to obtain a sample population that best represents each landcover class. In these samples, 20 polygons for each class were randomly identified by the visual interpretation of the Landsat images with the help of Google Earth (Mohamed & El-Raey, 2019; Zhu & Liu, 2014). Then, five hundred training samples per class were generated to train the classification for each year.
The goal of the validation was to assess the overall accuracy of the vegetation change map compared to available reference data. Validation was based on existing datasets and a combination of ancillary sources (Google earth, field plots) assembled using a human interpreter approach (Weng, 2018). High spatial resolution images from Google Earth Pro were used to manually interpret the land cover classes and also help determine land cover change at longer intervals (Kamga et al., 2020; Mohamed & El-Raey, 2019; Zhu & Woodcock, 2014). This research used high-resolution images of 2021 and 2000 in Google Earth pro as reference data. Specifically, about 120 samples were used as ground reference data for each landcover class to train the random forest classifier except for the settlement class, which was smaller in 2000. The classification accuracy was tested using the overall agreement with reference data. In addition, the user and producer agreements for the five landcover types were calculated. The producer accuracy measures the percentage of given reference data correctly classified; in contrast, the user accuracy measures the percentage of the trained data correctly classified (April et al., 2015).
Step 5: Post-classification
Following the classification of imagery for the two individual years, a post-classification approach of subtracting the classification maps was applied and used to produce a detailed change detection map using the ArcGIS Pro software. This map detects more subtle classes of change and determines the invasive pine and its spread over the selected landscape. An important aspect of change detection is determining what is changing to what category of land use type (Abebe et al., 2021). Therefore, a vegetation conversion matrix was calculated to demonstrate the direction of change and the land use type that remains at the end of the study period. Quantitative data analysis of the overall land cover change as well as gains and losses in each category between 2000 and 2021 were compiled (Haregeweyn et al., 2013; Kamga et al., 2020) and presented as a percentage of the area. In this manner, maps were created for the entire study area and provided insight into the areas affected by invasive conifer encroachment.
Buffer analysis: identification of invasive pine
In order to investigate the progressive spread of Pinus caribaea outside the original plantations, the distance from the perimeter of parental pine plantations was measured using six 50-m intervals between 0 and 300 m demarcated. The buffer distance was based on the previous study by Medawatte et al. (2010) in Sri Lanka. The area of invading pines was measured in relation to the distance from the plantation edge.
Results
The vegetation maps for 2000 and 2021 are shown in Fig. 3 and Fig. 4, respectively. These maps collectively show places with stable and changed vegetation areas appearing in the study area. Five major vegetation types were classified for 2000 and 2021: pine plantation, broadleaf forest, bare land, grassland, and invasive pines (settlements are also identified). The most prominent vegetation type found in both maps (2000 and 2021) is the broadleaf forest which is mainly distributed on the gentle slopes and the flat regions of the study area. In contrast, pine plantations and grasslands are located on ridges and steep areas with a slope higher than 18%. The vegetation classification for the year 2000 image shows that the majority of the study area was under broadleaf forest, covering about 2690 ha (43%). Grassland and bare land cover an area of 1627 ha (26%) and 969 ha (16%), respectively. Broadleaf forest (3336 ha) still covered the largest area in 2021, which reflects the conversion of other classes to the natural forest for conservation purposes, while grassland, pine plantation, and bare land cover an area of 1284 ha, 1013 ha, and 244 ha, respectively. The land areas contain pine species outside the plantation boundaries, with an absence of planting patterns, and areas smaller than 0.5 ha are identified as invasive pines.

Vegetation classification for the year 2000

Vegetation classification for the year 2021
Overall agreement with reference data was computed for both vegetation maps. The year 2000 vegetation map produced a 72% overall accuracy, and the year 2021 vegetation map yielded an 81% overall accuracy. Vegetation groups were classified with producer accuracy ranging from 20–84 and 55–100 for the year 2000 and 2021, respectively. In the year 2000 map, the settlement had the highest user accuracy (Appendix 1). On the other hand, pine plantation had the highest user accuracy for the year 2021, whereas all other vegetation classes have more than 70% user accuracy except the grassland class (Appendix 2).
The cover and rate of changes in each vegetation type for the 21 years are summarized in Table 2. From 2000 to 2021, the broadleaf forest and pine plantation area increased by 24% and 19%, respectively. In contrast, bare land and grassland showed a reverse trend, reduced by 75% and 21%, respectively, during the same period. This indicates that the increasing broadleaf forest area mainly emerged in the steep slope areas, meaning some grassland and bare land areas were converted to natural forests without disturbance.
The vegetation conversion matrix depicts the direction of change and the land use type that remains at the end of the period. Thus, the change matrix for each period was analyzed to understand the source and destination of significant vegetation changes. Over 21 years, 3476 ha out of 6234 ha of the total area has not changed, accounting for 56% of the study area (Table 3). In 2021, there was a significant settlement build up activity, with an area of 269 ha or 4% of the region’s total area (Table 2).
Significantly, from 2000 to 2020, approximately 705 ha of bare land and 663 ha of grassland have converted to other vegetated areas. The increase in settlement surfaces is coupled with the decline in these grasslands and bare land areas. Furthermore, approximately 80% of the area covered with broadleaf forest in 2000 was still the same in 2021. The remaining 20% (536 ha) was transformed to other uses in 2021. Of the total pine plantation cover in 2000, 53% remained unchanged, while the remaining portion vastly changed to grassland (100 ha) and broadleaf forest (262 ha). Overall, there was a significant increase in the broadleaf forest from 2691 to 3336 ha. This gradual increase is attributed to the replanting programs carried out in the western part of the study area (Kumara, 2010).
The invasive pine areas during this period showed a 4% slight decline. Concurrently, 45 ha of invasive pine areas was converted to broadleaf forest, and 15 ha was cleared. Furthermore, 26 ha of invasive pine in 2000 was identified as pine plantations in 2021, whereas 19 ha of pine plantations in 2000 was recognized as invasive pine in 2021. This misperception may be due to less aerial visibility of continuous clumps of pine cover over the area in 2000, with an identifiable canopy only appearing later.
As this study focuses on the dispersal of invasive pine around the study area, the following section emphasizes the extent of escaped pines from the pine plantations into the adjacent habitats. Figure 5 observes the conversion of pine plantations and broadleaf forests from 2000 to 2021. Moreover, an expansion of pine plantations was detected primarily around the existing plantations of 2000. Broadleaf forests in more remote valley areas tended to retain extensive forest in 2021, and broadleaf forest expansion or regeneration of the forest can be observed primarily on grassland. During the 21 years, pine plantations have increased by 159 ha, whereas invasive pine declined by 4 ha. Thus, invasive pines are more visible along with broad leaf forests managing the same borders as the pine plantations.

Pine and broadleaf forest change map 2000–2021
The extent of invading pines was measured with respect to the distance from the plantation edge. For the buffer analysis, three different vegetation types were selected: broadleaf forest, grassland, and bare land, which are adjacent to the pine plantations. The results showed that pines invaded broadleaf forests to a greater extent than the other two habitat types (Fig. 6). Furthermore, analysis showed that the highest scattering of pines was within 100-m buffer zone of the plantation, while encroachment of pines reduced beyond the 100-m buffer zone.

Invasive pine expansion into the different vegetation types adjacent to pine plantations
Discussion
This research demonstrated that freely available Landsat images combined with topographical data can be used for mapping vegetation change over a twenty-year time period, confirming its applicability for mapping and understanding the extent of pine invasion. The results have revealed that pine invasion is at a moderate level but is decreasing might be due to anthropogenic activities.
The research has found that a combination of spectral, textural, and topographical bands provides an effective solution for classifying the vegetation cover, similar to the findings of Yu et al. (2020) on forest classification in subtropical regions, and the discriminating of urban forest types in a semi-humid monsoon region in China by Zhou et al. (2019). Earlier works by Wang et al. (2006) and Zhu and Liu (2014) had specified that using topographical data with spectral data improved the discrimination between conifers and broadleaf forests. Correspondingly, studies by Peterson (2005) and April et al. (2015) had found that model accuracy was slightly better using topographical parameters. In contrast, Bjerreskov et al. (2021) found that the elevation did not improve accuracy when classifying broadleaf and conifer forest in the Danish landscape. However, it appears that elevation plays a big role in forest classification in mountain terrain or when the landscape is complex, as demonstrated by this research.
This study obtained accuracies for the vegetation cover classification similar to those obtained by other studies using multi-year Landsat images (Coyle et al., 2020; Gómez et al., 2012), and the accuracies obtained for land classification using other higher resolution satellite images (Hantson et al., 2012; Perera, 2001). The classification accuracy of forest types obtained in this study is higher that obtained studies by Akumu et al. (2021), Senf et al. (2013), and an investigation by Sá et al. (2017), which focuses on the invasion of Acacia longifolia in a pine forest. However, studies by Andrew and Ustin (2008); Bjerreskov et al. (2021); Fagan et al. (2018); Hestir et al. (2008); and Underwood et al. (2003) obtained higher classification accuracies than the present study by using satellite imagery with a higher resolution.
Even though better results can be obtained with higher resolution images (Goncalves et al., 2022), Landsat has been shown to provide long-term assessments of invasive plant spread (Gavier-Pizarro et al., 2012; Labonté et al., 2020; Sá et al., 2017). Furthermore, the freely available Landsat series of satellites is one of the most important data sources for developing countries that have budget constraints (Gavier-Pizarro et al., 2012; Haregeweyn et al., 2013). Liu et al. (2022) concluded that Landsat is better for detecting long-term changes in Chinese pine tree species because of its availability over a long time period. For developing countries such as Sri Lanka, we conclude that Landsat images provide a vital low-cost solution for the frequent monitoring of vegetation change.
According to this study’s findings, broadleaved forests and grasslands are vulnerable to invasion. Previous researchers (Dash et al., 2019; Higgins & Richardson, 1998; Medawatte et al., 2010) found that grasslands were more prone to invasion than other areas. However, Goncalves et al. (2022), Förster et al. (2017), and Ayala et al. (2005) identified forest habitats as the most vulnerable to the pine invasion. Forest habitats become vulnerable to pine invasion when exposed to disturbances such as wildfire (Medawatte et al., 2010). Moreover, most invading pines were found within a 100-m buffer zone for all types of habitats, while encroachment declined sharply beyond the 50–100-m buffer zone. These results are in line with those of Ayala et al. (2005).
An important finding in this research was the decline in invasive pine in the Belihuloya region. Wijesundera (2008) had stated that Pinus caribaea had spread rapidly in several mid country areas and predicted that it could be a potential invasive alien in the near future. Due to social pressure from local communities, pine planting was stopped (Edirisinghe, 2017), and the country has identified pines an invasive species (Medawatte et al., 2010). In 2017, the Sri Lankan government implemented a project to plant indigenous plant species to develop an under-storey in the existing pine plantations (Office of the Cabinet of Ministers-Sri Lanka, 2017). This research gives insight into the invasion of pines in the intermediate zone and helps inform management strategies to maintain this downward trend.
While the study could have obtained better classification accuracy using higher spatial or spectral resolution images, these are not free and are only available for recent years. Extending the study period to before 2000 could also have reduced its limitation, but it would have been difficult to validate a classification prior to 2000 due to the absence of high-resolution images in Google Earth Pro.
Future research could seek to understand the spatial variation in vegetation change and the reasons the invasive pines spread to certain areas. For example, the relationship between pine invasion and disturbances such as forest fires could be explored. Areas impacted by forest fire could be remotely sensed using satellite images. Furthermore, mapping and monitoring invasive pines in other climatic regions would also be relevant for forest management and conservation planning.
Conclusion
This study demonstrated the extent of pine invasion in the Belihuloya region, Sri Lanka. Remote sensing has enabled the mapping of vegetation change in the study area between 2000 and 2021 using freely available Landsat data. A combination of spectral, textural, and topographical bands provided an effective solution for detecting the spread of invasive Pinus caribaea. This study provided evidence that pines have invaded the natural habitat in the intermediate climate areas of Sri Lanka, especially native forests, and grasslands but the extent of this invasion has declined between 2000 and 2021.
It is clear that remote sensing is a valuable tool for monitoring and understanding forest dynamics, and the use of freely available satellite images, such as Landsat, is particularly important for developing countries such as Sri Lanka. The availability of the Google Earth Engine platform and its extensive range of analysis functions and free processing power is also very beneficial for countries with limited financial resources.
Data availability
The data that support the findings of this study are openly available in GEE at https://code.earthengine.google.com/5a840aa5fb12eba70dc0802153319af9.
Change history
05 March 2023
Missing Open Access funding information has been added in the Funding Note.
References
Abebe, G., Getachew, D., & Ewunetu, A. (2021). Analysing land use/land cover changes and its dynamics using remote sensing and GIS in Gubalafito district. Northeastern Ethiopia. SN Applied Sciences, 4(1), 30. https://doi.org/10.1007/s42452-021-04915-8
Afrin, S., Sharmin, S., & Mowla, Q. A. (2010). The environmental impact of alien invasive plant species in Bangladesh. International Conference on Environmental Aspects of Bangladesh (ICEAB10), Japan.
Akumu, C., Smith, R., & Haile, S. (2021). Mapping and monitoring the canopy cover and greenness of southern yellow pines (loblolly, shortleaf, and Virginia pines) in Central-Eastern Tennessee using multi-temporal landsat satellite data. Forests, 12(4). https://doi.org/10.3390/f12040499
Amaral, C. H., Roberts, D. A., Almeida, T. I. R., & Souza Filho, C. R. (2015). Mapping invasive species and spectral mixture relationships with neotropical woody formations in southeastern Brazil. ISPRS Journal of Photogrammetry and Remote Sensing, 108, 80–93. https://doi.org/10.1016/j.isprsjprs.2015.06.009
Andrew, M. E., & Ustin, S. L. (2008). The role of environmental context in mapping invasive plants with hyperspectral image data. Remote Sensing of Environment, 112(12), 4301–4317. https://doi.org/10.1016/j.rse.2008.07.016
April, S. E., Sarr, D. A., Van Kirk, R. W., & Jules, E. S. (2015). Quantifying habitat loss: Assessing tree encroachment into a serpentine savanna using dendroecology and remote sensing. Forest Ecology and Management, 340, 9–21. https://doi.org/10.1016/j.foreco.2014.12.019
Ayala, L., Perevolotsky, A., Kigel, J., & Noy-Meir, I. (2005). Invasion of Pinus halepensis from plantations into adjacent natural habitats. Applied Vegetation Science, 8(1), 85–92. http://www.jstor.org/stable/4620435
Bjerreskov, K. S., Nord-Larsen, T., & Fensholt, R. (2021). Classification of nemoral forests with fusion of multi-temporal Sentinel-1 and 2 data. Remote Sensing, 13(5). https://doi.org/10.3390/rs13050950
Bradley, B. A. (2014). Remote detection of invasive plants: A review of spectral, textural and phenological approaches. Biological Invasions, 16(7), 1411–1425. https://doi.org/10.1007/s10530-013-0578-9
Burley, J., & Barnes, R. D. (2004). Tropical ecosystems | Tropical pine ecosystems and genetic resources. In J. Burley (Ed.), Encyclopedia of forest sciences (pp. 1728–1740). Elsevier. https://doi.org/10.1016/B0-12-145160-7/00089-2
Cheng, K., & Wang, J. (2019). Forest type classification based on integrated spectral-spatial-temporal features and random forest algorithm—A case study in the Qinling Mountains. Forests, 10(7), 559. https://www.mdpi.com/1999-4907/10/7/559
Coyle, D. R., Barnes, B. F., Klepzig, K. D., Koch, F. H., Morris, L. A., Nowak, J. T., Otrosina, W. J., Smith, W. D., & Gandhi, K. J. K. (2020). Abiotic and Biotic Factors Affecting Loblolly Pine Health in the Southeastern United States. Forest Science, 66(2), 145–156. https://doi.org/10.1093/forsci/fxz054
Dash, J. P. (2020). On the detection and monitoring of invasive exotic conifers in New Zealand using remote sensing. University of Canterbury. https://ir.canterbury.ac.nz/bitstream/handle/10092/101329/Dash%2C%20Jonathan_Final%20PhD%20Thesis.pdf?sequence=1&isAllowed=y
Dash, J. P., Watt, M. S., Paul, T. S. H., Morgenroth, J., & Pearse, G. D. (2019). Early detection of invasive exotic trees using UAV and manned aircraft multispectral and LiDAR data. Remote Sensing, 11(15). https://doi.org/10.3390/rs11151812
de Sá, N. C., Carvalho, S., Castro, P., Marchante, E., & Marchante, H. (2017). Using Landsat time series to understand how management and disturbances influence the expansion of an invasive tree. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing, 10(7), 3243–3253. https://doi.org/10.1109/JSTARS.2017.2673761
Deng, X., Guo, S., Sun, L., & Chen, J. (2020). Identification of short-rotation eucalyptus plantation at large scale using multi-satellite imageries and cloud computing platform. Remote Sensing, 12(13). https://doi.org/10.3390/rs12132153
Edirisinghe, N. (2017). Enrichment of pine plantations of Sri Lanka with native species. https://www.apfnet.cn/uploads/file/20170425/1493111411376528.pdf
Fagan, M. E., Morton, D. C., Cook, B. D., Masek, J., Zhao, F., Nelson, R. F., & Huang, C. (2018). Mapping pine plantations in the southeastern U.S. using structural, spectral, and temporal remote sensing data. Remote Sensing of Environment, 216, 415–426. https://doi.org/10.1016/j.rse.2018.07.007
Förster, M., Gränzig geb. Schmidt, T., Wolf, R., Kleinschmit, B., Fassnacht, F., Cabezas Peña, J., & Kattenborn, T. (2017). Detecting the spread of invasive species in central Chile with a Sentinel-2 time-series 9th International Workshop on the Analysis of Multitemporal Remote Sensing Images (MultiTemp), Brugge, Belgium. https://ieeexplore.ieee.org/document/8035216
Gavier-Pizarro, G. I., Kuemmerle, T., Hoyos, L. E., Stewart, S. I., Huebner, C. D., Keuler, N. S., & Radeloff, V. C. (2012). Monitoring the invasion of an exotic tree (Ligustrum lucidum) from 1983 to 2006 with Landsat TM/ETM+ satellite data and support vector machines in Córdoba, Argentina. Remote Sensing of Environment, 122, 134–145. https://doi.org/10.1016/j.rse.2011.09.023
Gómez, C., Wulder, M. A., White, J. C., Montes, F., & Delgado, J. A. (2012). Characterizing 25 years of change in the area, distribution, and carbon stock of Mediterranean pines in Central Spain. International Journal of Remote Sensing, 33(17), 5546–5573. https://doi.org/10.1080/01431161.2012.663115
Goncalves, V. P., Ribeiro, E. A., & Imai, N. N. (2022). Mapping areas invaded by Pinus sp. from geographic object-based image analysis (GEOBIA) applied on RPAS (drone) color images. Remote Sensing, 14(12). https://doi.org/10.3390/rs14122805
Hantson, W., Kooistra, L., & Slim, P. A. (2012). Mapping invasive woody species in coastal dunes in the Netherlands: a remote sensing approach using LIDAR and high-resolution aerial photographs. Applied Vegetation Science, 15(4), 536–547. https://doi.org/10.1111/j.1654-109X.2012.01194.x
Haregeweyn, N., Tsunekawa, A., Tsubo, M., Meshesha, D., & Melkie, A. (2013). Analysis of the invasion rate, impacts and control measures of Prosopis juliflora: A case study of Amibara District Eastern Ethiopia. Environmental Monitoring and Assessment, 185(9), 7527–7542. https://doi.org/10.1007/s10661-013-3117-3
Hestir, E. L., Khanna, S., Andrew, M. E., Santos, M. J., Viers, J. H., Greenberg, J. A., Rajapakse, S. S., & Ustin, S. L. (2008). Identification of invasive vegetation using hyperspectral remote sensing in the California Delta ecosystem. Remote Sensing of Environment, 112(11), 4034–4047. https://doi.org/10.1016/j.rse.2008.01.022
Higgins, S. I., & Richardson, D. M. (1998). Pine invasions in the southern hemisphere: Modelling interactions between organism, environment and disturbance. Plant Ecology, 135(1), 79–93. http://www.jstor.org/stable/20050598
Jayawardane, S. S. B. D. G., & Weerasena, L. A. (2000). Crop diversification in Sri Lanka. Crop diversification in the Asia-Pacific region.
Jayawardhane, J., & Gunaratne, A. (2020). Restoration success evaluation of a thinned and enriched pine plantation in Sri Lanka. Journal of Tropical Forest Science, 32(4), 402–413. https://www.jstor.org/stable/26940730
Kamga, M. A., Nguemhe Fils, S. C., Ayodele, M. O., Olatubara, C. O., Nzali, S., Adenikinju, A., & Khalifa, M. (2020). Evaluation of land use/land cover changes due to gold mining activities from 1987 to 2017 using Landsat imagery East Cameroon. Geojournal, 85(4), 1097–1114. https://doi.org/10.1007/s10708-019-10002-8
Kaplan, G. (2021). Broad-leaved and coniferous forest classification in Google Earth Engine using Sentinel imagery. Environmental Sciences Proceedings, 3(1), 64. https://www.mdpi.com/2673-4931/3/1/64
Kauth, R. J., & Thomas, G. S. P. (1976). The tasselled cap -- A graphic description of the spectral-temporal development of agricultural crops as seen by Landsat. Symposium on Machine Processing of Remotely Sensed Data, West Lafayette, Indiana.
Khare, S., Latifi, H., & Ghosh, S. K. (2018). Multi-scale assessment of invasive plant species diversity using Pléiades 1A, RapidEye and Landsat-8 data. Geocarto International, 33(7), 681–698. https://doi.org/10.1080/10106049.2017.1289562
Kumar, V., Sharma, A., Bhardwaj, R., & Thukral, A. K. (2018). Comparison of different reflectance indices for vegetation analysis using Landsat-TM data. Remote Sensing Applications: Society and Environment, 12, 70–77. https://doi.org/10.1016/j.rsase.2018.10.013
Kumara, C. (2010). Effectiveness of establishing forest buffer zones as a community forest management approach: A case study from the Sripada Tropical Peak Wilderness Sanctuary in Sri Lanka. University of Agder.
Labonté, J., Drolet, G., Sylvain, J. -D., Thiffault, N., Hébert, F., & Girard, F. (2020). Phenology-based mapping of an alien invasive species using time series of multispectral satellite data: A case-study with glossy buckthorn in Québec, Canada. Remote Sensing, 12(6), 922. https://www.mdpi.com/2072-4292/12/6/922
Liu, L., Guo, Y., Li, Y., Zhang, Q., Li, Z., Chen, E., Yang, L., & Mu, X. (2022). Comparison of machine learning methods applied on multi-source medium-resolution satellite images for Chinese pine (Pinus tabulaeformis) extraction on Google Earth Engine. Forests, 13(5). https://doi.org/10.3390/f13050677
Lukeš, P., Stenberg, P., Rautiainen, M., Mõttus, M., & Vanhatalo, K. M. (2013). Optical properties of leaves and needles for boreal tree species in Europe. Remote Sensing Letters, 4(7), 667–676. https://doi.org/10.1080/2150704X.2013.782112
McEwan, A., Marchi, E., Spinelli, R., & Brink, M. (2020). Past, present and future of industrial plantation forestry and implication on future timber harvesting technology. Journal of Forestry Research, 31(2), 339–351. https://doi.org/10.1007/s11676-019-01019-3
Medawatte, W., Tennakoon, K., Hulme, P., Gunatilleke, C., & Gunatilleke, I. (2010). Is Caribbean pine invading grasslands in the Knuckles range. Invasive alien species–Strengthening capacity to control introduction and spread in Sri Lanka, 131–140.
Mohamed, S. A., & El-Raey, M. E. (2019). Land cover classification and change detection analysis of Qaroun and Wadi El-Rayyan lakes using multi-temporal remotely sensed imagery. Environmental Monitoring and Assessment, 191(4), 229. https://doi.org/10.1007/s10661-019-7339-x
Nissanka, S. P., Mohotti, K., & Wijetunga, A. (2005). Alleopathic influences of Pinus caribea on vegetation regeneration and soil biodiversity. Fourth World Congress on Allelopathy, Wagga Wagga, NSW, Australia.
Office of the Cabinet of Ministers-Sri Lanka. (2017). Conversion of the Pinus plantation in Sri Lanka into a plantation of indigenous plant species. Office of the Cabinet of Ministers-Sri Lanka Retrieved from http://www.cabinetoffice.gov.lk/cab/index.php?option=com_content&view=article&id=16&Itemid=49&lang=en&dID=8386
Pauchard, A., Escudero, A., García, R. A., de la Cruz, M., Langdon, B., Cavieres, L. A., & Esquivel, J. (2016a). Pine invasions in treeless environments: Dispersal overruns microsite heterogeneity. Ecology and Evolution, 6(2), 447–459. https://doi.org/10.1002/ece3.1877
Pauchard, A., García, R., Zalba, S., Sarasola, M., Zenni, R., Ziller, S., & Nuñez, M. A. (2016b). Pine invasions in South America: Reducing their ecological impacts through active management. In J. Canning-Clode (Ed.), Biological invasions in changing ecosystems: Vectors, ecological impacts, management and predictions (pp. 318–342). De Gruyter Open Poland. https://doi.org/10.1515/9783110438666-020
Perera, G. A. D. (2001). Secondary forest situation in Sri Lanka: A review. Journal of Tropical Forest Science, 13(4), 768–785.
Petersen, R., Goldman, E. D., Harris, N. L., Sargent, S. N., Aksenov, D., Manisha, A., Esipova, E., Shevade, V. S., Loboda, T. V., Kuksina, N., & Kurakina, I. (2016). Mapping tree plantations with multispectral imagery: Preliminary results for seven tropical countries.
Peterson, E. B. (2005). Estimating cover of an invasive grass (Bromus tectorum) using tobit regression and phenology derived from two dates of Landsat ETM+ data. International Journal of Remote Sensing, 26(12), 2491–2507. https://doi.org/10.1080/01431160500127815
Piiroinen, R., Fassnacht, F. E., Heiskanen, J., Maeda, E., Mack, B., & Pellikka, P. (2018). Invasive tree species detection in the Eastern Arc Mountains biodiversity hotspot using one class classification. Remote Sensing of Environment, 218, 119–131. https://doi.org/10.1016/j.rse.2018.09.018
Pu, R., Gong, P., Tian, Y., Miao, X., Carruthers, R. I., & Anderson, G. L. (2008). Invasive species change detection using artificial neural networks and CASI hyperspectral imagery. Environmental Monitoring and Assessment, 140(1), 15–32. https://doi.org/10.1007/s10661-007-9843-7
Punyawardena, B. V. R. (2020). Climate. In R. B. Mapa (Ed.), The soils of Sri Lanka (pp. 13–22). Springer International Publishing. https://doi.org/10.1007/978-3-030-44144-9_2
Rejmánek, M. (2014). Invasive trees and shrubs: Where do they come from and what we should expect in the future? Biological Invasions, 16(3), 483–498. https://doi.org/10.1007/s10530-013-0603-z
Rejmánek, M., & Richardson, D. M. (2013). Trees and shrubs as invasive alien species – 2013 update of the global database. Diversity and Distributions, 19(8), 1093–1094. https://doi.org/10.1111/ddi.12075
Richardson, D., Rundel, P., Jackson, S., Teskey, R., Aronson, J., Bytnerowicz, A., Wingfield, M., & Procheş, Ş. (2007). Human impacts in pine forests: Past, present, and future. Annual Review of Ecology, Evolution and Systematics, 38, 275–97297. https://doi.org/10.1146/annurev.ecolsys.38.091206.095650
Roberts, D. A., Ustin, S. L., Ogunjemiyo, S., Greenberg, J., Dobrowski, S. Z., Chen, J., & Hinckley, T. M. (2004). Spectral and structural measures of northwest forest vegetation at leaf to landscape scales. Ecosystems, 7(5), 545–562. https://doi.org/10.1007/s10021-004-0144-5
Senf, C., Pflugmacher, D., Van Der Linden, S., & Hostert, P. (2013). Mapping rubber plantations and natural forests in Xishuangbanna (Southwest China) using multi-spectral phenological metrics from MODIS time series. Remote Sensing, 5(6), 2795–2812. https://doi.org/10.3390/rs5062795
Setiani, P., Devianto, L. A., & Ramdani, F. (2021). Rapid estimation of CO2 emissions from forest fire events using cloud-based computation of Google Earth Engine. Environmental Monitoring and Assessment, 193(10), 669. https://doi.org/10.1007/s10661-021-09460-w
Shahfahad, T. S., Ali, R., Nguyen, K. -A., Naikoo, M. W., Liou, Y. -A., Islam, A. R. M. T., Mallick, J., & Rahman, A. (2022). Monitoring drought pattern for pre- and post-monsoon seasons in a semi-arid region of western part of India. Environmental Monitoring and Assessment, 194(6), 396. https://doi.org/10.1007/s10661-022-10028-5
Signori, L., & Ducati, J. (2019). Spatio-temporal mapping of the invasive alien species Pinus sp in the northern area of the Lagoa Do Peixe National Park-Rs. Revista Árvore, 43. https://doi.org/10.1590/1806-90882019000100007
Singh, S. P., Inderjit, S. J. S., Majumdar, S., Moyano, J., Nuñez, M. A., & Richardson, D. M. (2018). Insights on the persistence of pines (Pinus species) in the Late Cretaceous and their increasing dominance in the Anthropocene. Ecology and Evolution, 8(20), 10345–10359. https://doi.org/10.1002/ece3.4499
Starkloff, R. (1998). Social and environmental aspects of watershed vulnerability in the Sri Lankan hill country. Paper 135 of Session 13.
Subasinghe, K., Sumanapala, A. P., & Weerawardhena, S. R. (2014). The impact of forest conversion on bird communities in the northern flank of the Knuckles Mountain Forest Range, Sri Lanka. Journal of Asia-Pacific Biodiversity, 7(4), 367–373. https://doi.org/10.1016/j.japb.2014.07.004
Subasinghe, S. (2007). Plantation forestry in Sri Lanka challenges and constraints Annual Sessions of the Institute of Biology, Sri Lanka.
Underwood, E., Ustin, S., & DiPietro, D. (2003). Mapping nonnative plants using hyperspectral imagery. Remote Sensing of Environment, 86(2), 150–161. https://doi.org/10.1016/S0034-4257(03)00096-8
Wang, L., Zhang, X., Luo, Y., Shi, J., & Huang, H. (2006). A study on the changes of Pinus massoniana spatial pattern by pine wood nematode invasion based on remote sensing and GIS. 2006 IEEE International Symposium on Geoscience and Remote Sensing, Denver, CO, USA.
Weisberg, P. J., Lingua, E., & Pillai, R. B. (2007). Spatial patterns of pinyon-juniper woodland expansion in central Nevada. In (Vol. 60, pp. 115–124): Society for Range Management.
Weng, Q. (2018). Remote sensing time series image processing. CRC Press, Taylor & Francis Group.
Wijerathna, N., Hettiarachchi, C., & Wickramagamage, P. (2016). Effect of Pinus caribaea plantation on vegetation diversity and distribution by reforestation in lower Hantana. International Conference on the Humanities and the Social Sciences (ICHSS) 2016, Peradeniya, Sri Lanka.
Wijesundera, S. (2008). Major invasive plant species in different climatic zones of Sri Lanka. Proceedings of the National Symposium on Invasive Alien Species (IAS 2008), Sri Lanka Foundation Institute.
Williams, D. L. (1991). A comparison of spectral reflectance properties at the needle, branch, and canopy level for selected Conifer species. Remote Sensing of Environment, 35(2), 79–93. https://doi.org/10.1016/0034-4257(91)90002-N
Xu, D., Chen, B., Yan, Y., Sun, X., & Xin, X. (2018). Spatial-temporal dynamic monitoring of Mongolian pine (Pinus sylvestris var. mongolica) based on remote sensing data. Remote Sensing Letters, 9(11), 1079–1088. https://doi.org/10.1080/2150704X.2018.1508908
Ye, S., Rogan, J., Zhu, Z., Hawbaker, T. J., Hart, S. J., Andrus, R. A., Meddens, A. J. H., Hicke, J. A., Eastman, J. R., & Kulakowski, D. (2021). Detecting subtle change from dense Landsat time series: Case studies of mountain pine beetle and spruce beetle disturbance. Remote Sensing of Environment, 263, 112560. https://doi.org/10.1016/j.rse.2021.112560
Yu, X., Lu, D., Jiang, X., Li, G., Chen, Y., Li, D., & Chen, E. (2020). Examining the roles of spectral, spatial, and topographic features in improving land-cover and forest classifications in a subtropical region. Remote Sensing, 12(18). https://doi.org/10.3390/rs12182907
Zhou, X., Li, L., Chen, L., Liu, Y., Cui, Y., Zhang, Y., & Zhang, T. (2019). Discriminating urban forest types from Sentinel-2A image data through linear spectral mixture analysis: A case study of Xuzhou, East China. Forests, 10(6), 478. https://www.mdpi.com/1999-4907/10/6/478
Zhu, X., & Liu, D. (2014). Accurate mapping of forest types using dense seasonal Landsat time-series. ISPRS Journal of Photogrammetry and Remote Sensing, 96, 1–11. https://doi.org/10.1016/j.isprsjprs.2014.06.012
Zhu, Z., & Woodcock, C. E. (2014). Continuous change detection and classification of land cover using all available Landsat data. Remote Sensing of Environment, 144, 152–171. https://doi.org/10.1016/j.rse.2014.01.011
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study’s conception and design. Workflow preparation, data collection, data analysis, and the draft of the manuscript were performed by the corresponding author, Mrs. W.D.K.V.Nandasena. The co-authors, Dr. Lars Brabyn and Associate Professor Silvia Serrao-Neumann, have contributed to enriching the previous versions through editing, guiding research procedures, and overall work supervisor.
Corresponding author
Ethics declarations
Ethic and informed consent
This is an observational study. This study did not involve research on humans or involve animals. The University of Waikato Research Ethics Committee has confirmed that no ethical approval or consent is required.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendices
Appendix 1 Confusion matrix for the year 2000
Classification | |||||||
|---|---|---|---|---|---|---|---|
Land cover | Pine plantation | Broadleaf forest | Grassland | Bare land | Settlement | Total | |
Reference | Pine plantation | 90 | 14 | 12 | 4 | 0 | 120 |
Broadleaf forest | 0 | 96 | 24 | 0 | 0 | 120 | |
Grassland | 13 | 3 | 87 | 17 | 0 | 120 | |
Bare land | 6 | 5 | 30 | 79 | 0 | 120 | |
Settlement | 0 | 4 | 4 | 8 | 4 | 20 | |
Total | 109 | 122 | 157 | 108 | 4 | 500 | |
User accuracy (%) | 82 | 79 | 58 | 73 | 100 | ||
Producer accuracy (%) | 75 | 84 | 72 | 66 | 20 | ||
Overall accuracy (%) | 72 | ||||||
Appendix 2 Confusion matrix for the year 2021
Classification | |||||||
|---|---|---|---|---|---|---|---|
Land cover | Pine plantation | Broadleaf forest | Grassland | Bare land | Settlement | Total | |
Reference | Pine plantation | 116 | 4 | 0 | 0 | 0 | 120 |
Broadleaf forest | 0 | 120 | 0 | 0 | 0 | 120 | |
Grassland | 0 | 4 | 104 | 12 | 0 | 120 | |
Bare land | 0 | 0 | 30 | 84 | 6 | 120 | |
Settlement | 0 | 18 | 18 | 18 | 66 | 120 | |
Total | 116 | 146 | 152 | 114 | 72 | 600 | |
User accuracy (%) | 100 | 82 | 68 | 74 | 92 | ||
Producer accuracy (%) | 96 | 100 | 86 | 70 | 55 | ||
Overall accuracy (%) | 81 | ||||||
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nandasena, W., Brabyn, L. & Serrao-Neumann, S. Monitoring invasive pines using remote sensing: a case study from Sri Lanka. Environ Monit Assess 195, 347 (2023). https://doi.org/10.1007/s10661-023-10919-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10661-023-10919-1