
Abstract
Dissolved organic matter (DOM) is recognized for its importance in freshwater ecosystems, but historical reliance on DOM quantity rather than indicators of DOM composition has led to an incomplete understanding of DOM and an underestimation of its role and importance in biogeochemical processes. A single sample of DOM can be composed of tens of thousands of distinct molecules. Each of these unique DOM molecules has their own chemical properties and reactivity or role in the environment. Human activities can modify DOM composition and recent research has uncovered distinct DOM pools laced with human markers and footprints. Here we review how land use change, climate change, nutrient pollution, browning, wildfires, and dams can change DOM composition which in turn will affect internal processing of freshwater DOM. We then describe how human-modified DOM can affect biogeochemical processes. Drought, wildfires, cultivated land use, eutrophication, climate change driven permafrost thaw, and other human stressors can shift the composition of DOM in freshwater ecosystems increasing the relative contribution of microbial-like and aliphatic components. In contrast, increases in precipitation may shift DOM towards more relatively humic-rich, allochthonous forms of DOM. These shifts in DOM pools will likely have highly contrasting effects on carbon outgassing and burial, nutrient cycles, ecosystem metabolism, metal toxicity, and the treatments needed to produce clean drinking water. A deeper understanding of the links between the chemical properties of DOM and biogeochemical dynamics can help to address important future environmental issues, such as the transfer of organic contaminants through food webs, alterations to nitrogen cycling, impacts on drinking water quality, and biogeochemical effects of global climate change.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The movement and processing of organic matter within and among terrestrial, freshwater, and oceanic environments has extensive implications for global climate and carbon (C) cycles (Battin et al. 2009; Tranvik et al. 2009). While organic matter pools in freshwater ecosystems are relatively small compared to the marine pool, omission of freshwater organic matter pools leads to > 25% overestimation in terrestrial net ecosystem production (Aufdenkampe et al. 2011; Butman et al. 2016; Cole et al. 2007). One primary pool of organic matter is dissolved organic matter (DOM), which helps control and is shaped by physical, chemical, and biological aspects of freshwater ecosystems. DOM plays an important role in the global C cycle due to its abundance and through its stimulation of microbial metabolism (Battin et al. 2008). The most common analysis to investigate the DOM pool is the measurement of its concentration in units of C (DOC), but DOM contains other elements in abundance as well (e.g., O, H, N, S and P). Recent changes in DOC loading reported in freshwater ecosystems reflect global ecosystem change (e.g., Evans et al. 2006). Here we are proposing that DOM is sensitive to global change because DOM pools integrate aspects of ecosystem energy and nutrient flows, landscapes, and climate. These changes in DOC and DOM can in turn affect primary production, biodiversity, fish production, biogeochemical processes, and drinking water quality.
When the journal Biogeochemistry was founded in 1985 there were only a handful of studies on DOM composition (Fig. 1). Yet, it has long been recognized that not all DOM is chemically equivalent (e.g., Thurman 1985). Over time, this recognition has been solidified by the different means to assess DOM composition including its age (i.e., time since fixation to an organic state), its average molecular weight and size of its molecules, its biochemical composition such as its free amino acid and simple sugar content, and its content of humified terrestrial-derived plant decay products. There are now a wide range of techniques to characterize DOM, varying in complexity such as elemental content, stable isotopic composition, radiocarbon dating, molecular level classification and spectrofluorometric characterization (e.g., Butman et al. 2012; Jaffe et al. 2008; Massicotte et al. 2017; McCallister et al. 2018; Minor et al. 2014). With these approaches, DOM composition has been described using a plethora of terminology (Table 1). The chemical composition, age, color, and molecular size of DOM also varies as it is transformed by biological, chemical, and physical processes (Cory et al. 2007; Kothawala et al. 2006). No matter how you slice it, DOM pools echo the health of ecosystems and provide clear signals of how humans have reshaped the Earth’s biogeochemistry.

Evaluation of the percentage of publications with “dissolved organic matter” (DOM) or “dissolved organic carbon” (DOC) and selected global changes over time, starting in 1985, the year Biogeochemistry began publishing. Data is based on searches performed in Web of Science (August 6, 2020). Raw data and search details are available in the supplemental information. *In 1985, 1/25 (or 4.8%) of publications on DOM or DOC were on eutrophication
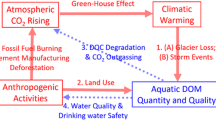
Considerable uncertainty remains about the processing of freshwater C and its net effect on the ecology and biogeochemistry of lakes and rivers. The nature of chemical changes to DOM is further complicated because this processing is proximally controlled both by the environmental context and by the compositional characteristics of the DOM, such that similar sources processed differently can yield disparate end-products (Moran et al. 2000). Consequently, the source of the DOM and the nature of its processing shape the quantity and quality of DOM present in any freshwater ecosystem at a given time. Across spatial and temporal scales, local scale variation in DOM cycles give way to reliable, long-term DOM quality patterns within biomes and aquatic ecosystems. However, both the dominant sources of DOM to freshwater ecosystems and the environmental conditions under which processing occurs (Fig. 2) have been significantly altered by anthropogenic global changes. The aggregate properties of this DOM are an important indicator of the source and past transformation of this C pool and are a key determinant of current and future ecosystem processes. While our knowledge of DOM chemistry has progressed rapidly over the past decade, it has not been well integrated into our understanding of C fluxes across landscapes, the ecological functioning of freshwater, and global change.

Flow chart exemplifying how human disturbances can impact environmental context and the source of DOM to inland waters which can potentially shift DOM composition, biogeochemical processing, ecological functioning, and a feedback back to DOM composition, since altered biogeochemical processing can also result in shifts to DOM composition
In the Anthropocene, freshwater ecosystems are experiencing cumulative stress from climate and land use change, among other human impacts. Through connections of river and lake DOM composition to land use, climate and other human activities, recent research discovered distinct DOM pools laced with “human markers and footprints” (Fig. 3). This “human-modified DOM” can in turn profoundly impact the biogeochemical processing and ecological function of aquatic ecosystems (Figs. 2, 4). The various human and natural drivers can also interact with one another and chronically, acutely, and/or synergistically influence DOM composition. Here, we review global changes that can affect DOM composition and in turn impact the transformation and processing of DOM within freshwater ecosystems. We focus on the following extensive global changes: climate change, land use change, nutrient pollution, browning, forest fires and dams. We then present the possible consequences of this variable, “human-transformed” DOM, on the biogeochemistry of freshwater environments.

Selected global changes and associated selected changes to the composition of the DOM pool leading to human fingerprints. These human DOM fingerprints influence biogeochemical processes in differential ways

DOM composition as a sentinel variable that is sensitive to global change. Changes in the DOM pool may significantly alter important ecosystem functions and ecosystem services. Selected global changes, ecosystem effects and aquatic ecosystem services are shown
There is abundant evidence that DOM pools today have been altered by humans. The human impacts on DOM can be indirect, as in glacial and permafrost thawing and forest fires, or direct as with nutrient pollution, urbanization, agriculture, and dams (Fig. 3). The properties and reactivity of the DOM pool have strong effects on biogeochemical processes, which are mediated by and feedback into the environmental context (physical, chemical, and biological processes; Fig. 2). The environmental impacts of humans are speeding up processes controlling the formation, transformation, burial, and decomposition of DOM. Over the short-term (years to decades), human activities have accelerated terrestrial sourced DOM loading into aquatic ecosystems. Over the long-term (decades to centuries) human activities provide two contrasting sides: (1) human activities are scattering more stable forms of DOM around the globe in the form of black carbon, formed from incomplete fossil fuel and biomass combustion and (2) human activities are shifting aquatic DOM pools either toward simpler, smaller, and potentially more reactive forms than what is observed in reference “natural” systems, or are increasing the amount of larger more aromatic forms in warmer and hydrologically altered ecosystems. Humans are mixing up aquatic DOM pools and leading to new DOM signatures. At the same concentration, different forms and mixtures of DOM can have drastically different effects on C transformations, ecosystem nutrient cycles, ecosystem productivity, metal toxicity, and the treatability of drinking water. Since humans are a major driver of global biogeochemical change, by incorporating human activities into studies of the C cycle and using DOM composition proxies, we can gain a better mechanistic understanding of DOM source, processing, sinks and ultimately the global C-cycle.
Human-dominated landscapes and effects on DOM composition
Across the relatively short-time scale of years, human modified landscapes create novel environmental conditions that alter biological organization and ecosystem functioning in freshwaters (e.g., Fasching et al. 2020), leading to distinct DOM pools and altered fluxes and processing of DOM. Depending on climate, pollution, hydrology, soil properties, the intensity of human activities, and the extent of natural land covers, human altered landscapes produce DOM pools with diverging characteristics. Human altered landscapes can enrich DOM pools with small-sized, aliphatic, and microbial-sourced components and/or load DOM pools with large-sized, aromatic, and terrestrial-sourced components. Hence, DOM pools are being altered by humans, but the local and regional effects are often context dependent.
Several studies have characterized DOM composition over gradients of land use disturbance. The percent cover of forested riparian areas alters temperature and light regimes through shading (Frost et al. 2006), composition of DOM inputs with differing vegetation and soils disturbance, and rate of groundwater input through increased transpiration (Wilson and Xenopoulos 2013), together placing strong constraints on the rates of photochemical degradation and microbial respiration. Most commonly observed across aquatic systems draining cultivated agricultural landscapes are an increase in microbial-derived DOM, protein-like DOM, and a higher fraction of dissolved organic nitrogen (DON) (Baker and Spencer 2004; Graeber et al. 2019; Lu et al. 2014; Pisani et al. 2020; Williams et al. 2010; Wilson and Xenopoulos 2009). In urban aquatic ecosystems and watersheds with high human population densities, a unique anthropogenic DOM signature has been identified with microbial and autochthonous characteristics (Hosen et al. 2014; Lambert et al. 2017; Williams et al. 2013, 2016). At the same time, an increase in the average age of DOC (i.e., fossil C) in streams has been observed in anthropogenic landscapes (Butman et al. 2015) because of changes in primary flow paths and loss of organic matter rich surface soils (Barnes et al. 2018; Ekblad and Bastviken 2019; Drake et al. 2019). In contrast, when agricultural activities result in increasing water yield, soil connectivity, surface flow, and DOC export (Graeber et al. 2012) or are associated with grazing animals rather than cropland (Catalán et al. 2013; Shang et al. 2018; Wilson and Xenopoulos 2009), agricultural activities may increase export of modern C (i.e., C recently fixed circa 1950) and high molecular weight material (Butman et al. 2015; Hosen et al. 2014; Lu et al. 2014).
Anthropogenic DOM in human-dominated landscapes is a symptom of the novel chemical cocktails resulting from soil changes, hydrology, light, temperature, nutrient pollution, organic contaminants (e.g., hydrocarbons, pesticides, pharmaceuticals, personal care products), wastewater inputs, and microbial activity (Hosen et al. 2014; Kaushal et al. 2018, 2020; Lambert et al. 2017; McEnroe et al. 2013). Increase in impervious surfaces, soil degradation, and from by-passing of surface flow with sub-surface artificial drainage, all typically result in a reduction of aromatic DOM. Afforestation, activities that enhance hydrological connection with organic matter rich surface soils, and landscape management that leaves crop/vegetation residuals on the land, all typically result in an increase of aromatic DOM. Over the longer term, as landscapes are further degraded by humans and climate change allows for agricultural expansion and intensification, DOM pools will likely resemble anthropogenic rather than natural qualities. Landscape-mediated changes to the DOM pool should ultimately be super-imposed against large-scale changes to landscapes and hydrology resulting from climate change and nutrient pollution (see sections below).
Climate change effects on DOM composition
While considerable attention has focused on the potential changes in DOC loading resulting from a globally changing climate, less is known about whether and how DOM composition will change. Speculatively, some climate change drivers would likely have strong effects on DOM composition in aquatic ecosystems. In particular, the loss of permafrost, and areas of increased precipitation are associated with greater hydrologic connectivity and may receive more terrestrially derived DOM. In contrast, warmer and drier areas may become hydrologically disconnected and eventually be characterized by more internally derived DOM. Further changes in DOM processing factors (temperature, light, and nutrients) and their response to climate change makes simple predictions about how they will alter DOM chemistry challenging. In addition, climate change effects in terrestrial habitats can easily influence the amounts of terrestrial primary production, the quality of litter, water table level, soil temperature and moisture. All of these factors can lead to changes in the quantity and composition of leachable DOM exported from terrestrial systems to inland waters (e.g., brownification see below). Finally, storms and extreme climate events are expected to mobilize the aforementioned anthropogenic chemical cocktails (sensu Kaushal et al. 2018), but how this will translate into changes in DOM composition is still emerging. What is clear is that climate change is leading to rapid and large pulses of DOM into aquatic ecosystems, which is shifting the composition and reactivity of the DOM pool.
Changes in precipitation and runoff patterns, floods, and droughts
An increase in flooding and discharge could result in more hydrological connections between C-rich terrestrial ecosystems and downstream aquatic ecosystems and increase DOC export. However, whether such increases in DOC will be of a different DOM composition has only recently been considered (e.g., Fasching et al. 2016, 2019). During extended baseflow conditions (e.g., droughts), DOM inputs derived primarily from groundwater have a more protein-like, biolabile composition (Fasching et al. 2016; Inamdar et al. 2011), with summer low flows linked to the production of in-stream DOM from aquatic primary production (Hosen et al. 2020). The mobile pool of DOM in the soils of most small watersheds is large and usually transport limited. In natural/forested headwater systems during high flow/flood events, higher concentration and export of nutrients and C-rich DOM (higher C:N) with higher molecular weight, and more aromatic DOM properties have frequently been observed (Fasching and Battin 2012; Hood et al. 2006; Nguyen et al. 2010; Vidon et al. 2008; Wilson and Xenopoulos 2009; Wilson et al. 2016). Across agricultural landscapes, flooding events modify DOM composition by disproportionally increasing the export of proteinaceous DOM compared to catchments with relatively more natural land cover (Fasching et al. 2019). In landscapes with very little DOM (e.g., glacial melt, a large snowpack, or rain on impervious surfaces) or where conditions favor flow over or around, rather than through organic rich substrates (e.g., the presence of an ice layer at the soil surface during snowmelt, freshet in big Arctic rivers) initially DOM inputs increase but ultimately dilution of DOM occurs with flow increases (Hornberger et al. 1994; Hood and Scott 2008; Wilson and Xenopoulos 2008). Furthermore, increased overland flow from heavy rainfall, can reduce the contact time with the mineral soils decreasing microbial processing and sorption (Inamdar et al. 2012) and consequently increasing the mobile hydrophilic fraction of DOM exported to streams (Kaiser and Zech 1998). Similar to what is observed around land use, these flood-drought patterns then produce divergent DOM pools depending on hydrologic flow paths, water availability, and soil properties.
Climatic changes may further impact the magnitude, composition and timing of DOM exported to aquatic environments. For instance, most DOC export from terrestrial sources and its downstream transport, takes place during floods (Raymond and Saiers 2010) and during spring snowmelt in northern high-latitude and temperate systems (e.g., Laudon et al. 2004; Spencer et al. 2009). Increased frequency and magnitude of floods due to climate change may cause the temporary depletion of the event transportable terrestrial soil organic C (Butman et al. 2015), while the earlier onset of snowmelt alters the timing of DOM fluxes. Additionally, while reductions in subsurface flow paths through mineral soils remove DOM via sorption, increased overland flow from land use change, can lead to increased export of DOM from the C-rich soil surface (e.g., Hood et al. 2006; Kaiser and Kalbitz 2012). This may consequently increase the mobile hydrophilic fraction of DOM exported to streams (Kaiser and Zech 1998). Altogether, this evidence suggests that in order to predict the impact of humans on DOM composition, we need to understand the hydrology and soil–water connectivity of ecosystems.
The melting of glaciers
Ongoing trends in atmospheric warming, and the reduction in snowfall has led to the shrinking of glaciers globally (Milner et al. 2017). Melting glaciers contribute to the DOM pool in glacial streams and downstream ecosystems. DOM derived from melting glaciers in Alaska has been found to be highly biolabile (Hood et al. 2009) and biogeochemically diverse (Singer et al. 2012). Glaciers accumulate organic C from terrestrial sources, atmospheric deposition and in situ primary production (e.g., Stubbins et al. 2012; Hood et al. 2015; Singer et al. 2012; Smith et al. 2017). Atmospheric deposition can take two forms. The first is anthropogenic aerosols and combustion products which have been detected in glaciers using ultrahigh resolution mass spectrometry and act as a source of pre-aged organic matter (Singer et al. 2012; Spencer et al. 2014; Stubbins et al. 2012). The second form is from N atmospheric deposition, which fuels glacier microbes and leads to autochthonous production that is rich in proteinaceous DOM (Dubnick et al. 2010; Pautler et al. 2012). Ongoing loss of glacial mass implies shifts in the quantity and composition (Milner et al. 2017; Behnke et al. 2020) of glacial-derived DOM to downstream ecosystems. Continued net glacial melt may initially increase the deliveries of ancient bioavailable DOM to streams, potentially increasing downstream metabolism (Brighenti et al. 2019). As glacial retreat continues, the export of bioavailable DOM may decrease (Huss and Hock 2018), leading instead to the export of ancient stable C (Bardgett et al. 2007), at least until new soil is formed.
Thawing of permafrost
Permafrost soils are expected to contribute to the DOM pool with climate change. In the Arctic, permafrost contains ~ 1700 Pg of organic C, nearly double the amount in the contemporary atmosphere (Schuur et al. 2013; Tarnocai et al. 2009). These permafrost soils have stored organic C frozen in many regions for millennia, and due to ongoing and amplified Arctic warming are projected to release between 41 and 288 Pg-C by 2100 and up to 616 Pg-C by 2300 due to permafrost thaw (Schaefer et al. 2011; Schuur et al. 2013; Zimov et al. 2006). The fate of this liberated organic matter in Arctic fluvial ecosystems remains unclear but despite its ancient age, upon thaw permafrost derived DOM has been shown to be highly biolabile (Drake et al. 2015; Mann et al. 2015; Spencer et al. 2015). High loss of permafrost DOC has been reported in bottle experiments with typically 30–50% loss of DOC in 14–28-day incubations despite the permafrost DOC having radiocarbon ages > 20,000 ybp (Spencer et al. 2015; Vonk et al. 2013). Microbial utilization of permafrost DOC in headwater streams may be so rapid that little permafrost derived aged DOC is exported from large Arctic rivers where extensive thaw has been shown (Mann et al. 2015; Spencer et al. 2015).
The high biolability of permafrost DOM is from its unique molecular composition exhibiting a high aliphatic content, and relative enrichment in low molecular weight, saturated, and less-oxidized compounds (Drake et al. 2015; Spencer et al. 2015; Ward and Cory 2015). In comparison to other Arctic surface waters a number of unique molecular formulae have been identified by FT-ICR MS in permafrost thaw streams with relatively high H/C CHO and lower H/C N-containing compounds which are energy-rich and thus highly biolabile (Drake et al. 2018a, b; Spencer et al. 2015). Permafrost thaw DOM is rapidly lost via microbial respiration (i.e., Drake et al. 2018a, b) in small stream networks. Concurrent with large inputs of modern DOM, permafrost thaw DOM does not currently persist to be exported by large Arctic rivers, which are therefore dominated by modern DOC export (Raymond et al. 2007; Wild et al. 2019). Moving forward as permafrost thaw continues into deeper layers, studies have suggested even greater biolability as permafrost DOM at depth has been shown to contain even higher relative contributions of aliphatic compounds and low molecular weight organic acids (Ewing et al. 2015; Heslop et al. 2019) ultimately resulting in the transfer of this long-term C store to the atmosphere with ramifications for climate.
Increased nutrient export and deposition impacts on DOM composition
A major aspect of global change involves the alteration to most biologically important elements, which are closely connected to DOM compositional impacts from landscape change and associated human activities. In particular, perturbations to the cycles of N and phosphorus (P) proportionally exceed changes to the C cycle (Rockström et al. 2009). Human land use disturbances through direct fertilizer application, fossil fuel and biomass burning, and/or landscape destabilisation increase N and P loading in freshwaters, potentially leading to eutrophication. These increases in nutrient loading can strongly influence DOM cycling and transformation through altered microbial processing. Increases in algal biomass and resulting hypoxia from eutrophication can also influence DOM composition. Algal-derived DOM has distinct chemistry; it is aliphatic, proteinaceous, rich in polysaccharides, moderately humic with low C:N stoichiometry and low C/H compounds (Cory et al. 2007; Jaffe et al. 2008; McKnight et al. 2001). Hypoxia and anoxia can directly affect the oxidation state of quinones (Fulton et al. 2004) and enhance the re-suspension of aromatic DOM from sediments (Peter et al. 2016). With eutrophication-induced hypoxia, an accumulation of labile DOM derived from surface phytoplankton blooms is expected (Jessen et al. 2017). Hypoxia and anoxia can also lead to changes in DOM composition because of microbial community changes, the inhibition of fauna and microorganisms responsible for remineralization and the increase of anaerobic processes that favor sulfurization of organic matter (Jessen et al. 2017). The role of redox conditions in both the formation and degradation of organic matter is also an active area of research (e.g., Macalady and Walton-Day 2011).
Atmospheric N deposition from fossil fuel combustion and agricultural activities is a chronic input of reactive N to terrestrial ecosystems that can shift terrestrial productivity, soil organic matter pools and thus water quality. Enhanced N deposition can affect DOM exports by either shifting the composition and stability of soil DOM from changing humification (e.g., Aitkenhead-Peterson and Kalbitz 2005; Whittinghill et al. 2012) or altering the production and consumption of soil organic matter. Elevated N deposition is expected to be associated with mobilization of DOM, but this is predicated on the form of N and the subsequent changes in acidity (Evans et al. 2008). In temperate watersheds, soil N pools increase, and stream DOM tends to become more N rich reflecting a shift in terrestrial processing (Brookshire et al. 2007). Similarly, experimental N additions to grassland ecosystems have resulted in greater soil organic matter bioavailability due to increased soil microbial activity and a larger pool of labile organic matter in surface soils (Fang et al. 2014). Enhanced decomposition rates have also been observed in peat bogs (Bragazza et al. 2006) and temperate forests (Magill and Aber 2000) that are receiving elevated N deposition. These shifts in organic matter processing have led to greater losses of CO2 from both systems and DOC from peatlands (Bragazza et al. 2006; Magill and Aber 2000). The fate of this exported DOC remains unclear, in some cases it becomes less reactive over time (Findlay 2005) with continued N loading.
Brownification
DOM loads to, and concentrations in many temperate inland waters have been increasing in recent decades, in response to recovery from anthropogenic acidification, climate change, afforestation, and other drivers (Kritzberg et al. 2020; Monteith et al. 2007; Solomon et al. 2015). This phenomenon is often referred to as “browning”. While most of the work on this topic has focused on DOC quantity, there has been less work documenting long-term changes in DOM composition. Jane et al. (2017) examined trends in both the DOC and DOM composition (as indicated by absorbance-based indices) in seven Wisconsin lakes over 24 years. Their results suggest that trends in DOC and DOM composition may sometimes be decoupled. They observed variable trends in DOC and found that DOM composition varied with a drought severity index, suggesting greater inputs of complex terrestrial molecules during wet periods. In contrast, two other recent studies found more consistent linkages between long-term increases in DOC and a shift towards more complex, terrestrially derived molecular structures (SanClements et al. 2012; Williamson et al. 2015). In general, it is expected that increases in terrestrial DOC loads should shift the DOM pools in inland waters towards a more “terrestrial” composition, including large, humic-like molecules with high molecular weight and aromatic structures (Creed et al. 2018). This shift should be most pronounced where residence time is short and soil organic matter is well connected to hydrologic flow paths. Where residence time is long, biological and photochemical degradation have more opportunity to alter the composition of the DOM pool relative to the DOM inputs and result in the preferential removal of oxidized, aromatic compounds, with greater humic-like fluorescence while more reduced aliphatic, N-containing and protein-like DOM persists (Kellerman et al. 2014; Kothawala et al. 2015).
Forest fires
Although not all forest fires are caused by humans, climate change and agricultural activities have been key factors in increasing the number and extent of wildfires. Changes in vegetative cover and soil properties following fire tend to increase the amount of water exported from a catchment; reflecting decreases in evapotranspiration and increases in surface runoff (e.g., Hallema et al. 2017). These same drivers alter allochthonous C and nutrient inputs and change in-stream conditions and thus organic matter processing. The DOC exported by burned watersheds is a product of C stock combustion during fire and subsequent alterations to organic matter remaining (Rhoades et al. 2019). A large fraction of the charcoal (black carbon) produced by fires is mobilized from soils and transported to the oceans in rivers (Wagner et al. 2018) although decreases in stream DOC from fire (Rodríguez-Cardona et al. 2020) have also been reported, often associated with fire severity and extent (Rhoades et al. 2019) and degree of landscape recovery (Rodríguez-Cardona et al. 2020).
The composition of stream DOM pools also shifts following fire, reflecting new flow paths and sources of organic matter within the landscape. Lower C:N in burned soils is the result of increased mineralization and nitrification of soil organic matter, increasing inorganic N availability (e.g., Knicker 2007). The associated tightening of C and N cycling results in decreased soil C sequestration and in a greater export of N (Bond-Lamberty et al. 2004; Rhoades et al. 2019) from the combustion of N soil pools and the acceleration of N cycling combined with reduced plant uptake (e.g., Rhoades et al. 2011). Reduced aromaticity of DOM pools with increased burned extent in a slowly recovering landscape reflects the reduction in new organic matter inputs to soil and dominance of microbial-derived, more bioavailable soil organic matter (Barnes et al. personal communication). As landscapes recover, the composition of the DOM pools shifts accordingly, reflecting the increased aromaticity of surficial, relatively organic-rich flow paths (Mann et al. 2012) with higher molecular weight and greater amounts of aliphatic compounds (Rodríguez-Cardona et al. 2020).
River flow alterations, dams, and impoundments
Hydropower and storage dams, in addition to the alteration of river channel morphology, impact stream C dynamics, affecting the storage, transformation and export of DOM from inland waters (Wohl et al. 2017). Dams and impoundments alter river hydrodynamics (mixing of DOM source waters and residence times), influencing the composition and timing of DOM transported to downstream ecosystems. The timing of dam filling/releases, determining DOM residence times and upstream source origins, can determine whether dams act as a source or sink of DOM to downstream ecosystems (Knoll et al. 2013; Kraus et al. 2011). For example, longer residence times and increased opportunity for photodegradation may be associated with export of lower molecular weight (Chen et al. 2016; Kraus et al. 2011) and higher levels of autochthonous DOM (Nguyen et al. 2002). Longer residence times, as well as releases during high flows from dams, result in a shift in peak DOM concentration from preceding peak discharge, to coinciding with peak stream discharge (Ulseth and Hall Jr. 2015).
While some studies have found that hydropower and storage dams may not significantly alter the DOM composition between upstream and downstream flows in a river (e.g., Nadon et al. 2015), others highlight a significant reduction in downstream DOM bioavailability and shift in DOM composition in terms of aromaticity and molecular weight (Ulseth and Hall Jr. 2015). Tailwaters, located directly downstream of dams have been found to be sites of increased primary production (Davis et al. 2011), leading to increased production and downstream export of autochthonous, and likely biolabile, DOC (Lieberman and Burke 1993; Ulseth and Hall Jr. 2015). Similarly, the removal of natural structures, like established beaver and debris dams, and gravel bars may impact DOM concentration and composition. These structures are associated with export of autochthonous and more processed DOM of high biodegradability and range of reactivities (Catalán et al. 2017), transforming and removing DOM in streams (Boodoo et al. 2020; Findlay and Sobczak 1996). Changes in river morphology and connectivity to their surrounding environment (channelization, impervious riverbeds and embankments, and beaver dams) can have consequences for DOM source, export and biodegradation dynamics within streams (Wohl et al. 2017; Harvey and Gooseff 2015). Decreased residence times and stream-riparian zone hyporheic mixing via river straightening and channelization, leads to decreased opportunity for DOM degradation via in-stream and hyporheic microbial processing (Griffiths et al. 2012). Furthermore, the burial of streams has been linked to lower levels of humic DOM as well as a reduction in DOM complexity and lability (e.g., Pennino et al. 2014; Arango et al. 2017).
Consequences and effects of human-altered DOM on biogeochemical processes
DOM can be highly reactive and strongly affect the functioning of freshwaters. Humans are changing DOM composition, directly and indirectly, with important ecological and biogeochemical consequences that are not yet well understood. The prospects for addressing this research gap is no longer daunting as the scientific community is increasingly using innovative ways to address biogeochemistry questions with DOM composition proxies. The proxies that can be used vary along a spectrum of complexity between simple optical parameters such as specific UV absorptivity (e.g., SUVA254, ε280) to more detailed molecular and chemical diversity assessments using ultrahigh resolution mass spectrometry (e.g., McCallister et al. 2018). Each methodology used provides unique insights into different aspects of C dynamics and biogeochemical processes (Panel 1). Below we highlight some of the known consequences of human-altered DOM composition on selected biogeochemical processes and ecosystem services.

The dynamic composition of the DOM pool will impact the global C cycle of inland water
There is now a growing recognition that freshwaters are important modifiers of C. They receive large amounts of C despite their small geographic footprint, sequester organic and inorganic C in their sediments and emit large quantities of C to the atmosphere. DOM composition can influence each of these budget compartments but to date no study has fully integrated DOM composition into local, regional, or global C budgets (Hanson et al. 2015).
Large fluxes of C are exported to inland waters from their surrounding terrestrial environment approximately 5.1 Pg C year−1 (Drake et al. 2018a, b; Raymond et al. 2016), which is processed and transported to downstream ecosystems. In general, DOM pools become more autochthonous-like as one moves along the aquatic continuum from land to oceans (e.g., Larson et al. 2014; Xenopoulos et al. 2017). In addition, increases in the export of autochthonous, bioavailable fractions of the DOM pool are expected in human-influenced ecosystems. For glaciers, the amount of DOC exported (1.04 Tg C year−1; Hood et al. 2015) is small compared to non-glaciated catchments (Bhatia et al. 2010) but the relatively higher bioavailability of glacier-derived DOM compared to forested, agricultural and wetland dominated catchments (Fasching et al. 2016; Risse-Buhl et al. 2013) supports heterotrophic metabolism in proglacial streams (Hood et al. 2009; Singer et al. 2012; Spencer et al. 2014). It follows that one consequence of an increase in microbial-like, N-rich, aliphatic DOM from human activities is an increase in microbial metabolism in downstream ecosystems (see below). This increase in microbial metabolism can result into more DOM buried and/or respired and lost into the atmosphere before reaching the ocean. This is complicated by estimates that show that CO2 emissions from small streams are largely from terrestrial DOM whereas the percentage of CO2 emissions from autochthonous metabolism increases in large rivers (Hotchkiss et al. 2015).
Changes in the composition of the DOM will likely further lead to changed rates of carbon uptake, retention, and out-gassing in freshwater ecosystems. In relation to out-gassing, DOM composition can impact the relative production of greenhouse gases (CO2 and CH4) from freshwaters compared to the amount that is buried in sediment. Several studies have related CO2 and CH4 production in freshwaters to DOM composition, with studies linking greater CO2 and CH4 production to more terrestrial, aromatic DOM (e.g., D’Amario and Xenopoulos 2015; West et al. 2012; Zhou et al. 2018). However, DOM composition alone is not enough to determine greenhouse gas production and other factors, such as nutrient levels or temperature (e.g., Jane and Rose 2018), also need to be considered. For example, eutrophic lakes with more autochthonous and less humic DOM were associated with CO2 influx at high phytoplankton biomass and lakes with high amounts of N and more wetland cover were associated with CO2 efflux (Morales-Williams et al. 2020). Nonlinearity may further complicate the relationships between DOM composition and C emissions (D’Amario and Xenopoulos 2015).
Among the primary factors influencing the loss of DOM is water residence time, with a general negative relationship between organic C degradation rates and water residence time across different types of freshwater ecosystems (Catalán et al. 2016). DOM composition plays a role as components with higher molecular weight and longer emission fluorescent wavelengths (i.e., DOM with more oxidised polyphenolic aromatic compounds with low H:C) have been found to degrade faster than DOM fluorescing at shorter emission wavelengths (i.e., DOM with more aliphatic and enriched in N-containing compounds; Kellerman et al. 2015; Köhler et al. 2013; Kothawala et al. 2014). This leads to a progressive loss of color with longer water residence time, which corresponds to losses in the terrestrial-like properties of DOM (Köhler et al. 2013; Weyhenmeyer et al. 2012). With climate change, areas which will experience lower precipitation and lower resulting runoff (thus longer water residence time) could be expected to have lower degradation rates but still degrade a larger portion of incoming terrestrial DOM (Catalán et al. 2016; Jones et al. 2018), potentially resulting in the prevalence of DOM with lower fluorescence and molecular weight, and more N-containing aliphatic compounds. On the other hand, in regions with predicted runoff increase with strong soil–water connectivity (i.e., wetter, thus shorter water residence time) degradation rates will be faster and we will find an apparent increase in terrestrially derived, aromatic DOM.
Sediment burial and storage of organic matter is also affected by DOM composition. Freshwater sediments have accumulated large quantities of organic C during the Holocene (Tranvik et al. 2009; Mendonça et al. 2017a, b) and DOM characteristics change with depth (Xu et al. 2016). The C buried in the sediments is mainly comprised of flocculated DOM compounds of terrestrial origin (Von Wachenfeldt and Tranvik 2008), as autochthonous DOM present in settling particles are preferentially degraded before burial (Guillemette et al. 2017). DOM flocculation may be stimulated by sunlight (von Wachenfeldt et al. 2008) but is also mediated by bacterial respiration (von Wachenfeldt et al. 2009) and in both cases, colored DOM appears as the main precursor of the flocs. Another major contributor to floc formation is the presence of transparent exopolymer particles (Grossart et al. 1998) which are usually related to phytoplankton (de Vicente et al. 2010), but also highly correlated to aromatic and terrestrial DOM (Attermeyer et al. 2019).
Carbon sedimentation is affected by the potential of DOM to adsorb onto inorganic surfaces (Kothawala et al. 2012). Although adsorption also depends on the sorbent and the water chemistry, DOM composition analyzed by different spectroscopic methods show that highly aromatic DOM compounds are preferentially adsorbed, whereas a high degree of saturation reduces DOM's susceptibility to adsorption (Groeneveld et al. 2020; Kalbitz et al. 2005; Kothawala et al. 2012). Finally, once buried, lakes with highly aromatic and terrestrial DOM are negatively correlated with sediment bacterial metabolism, while in lakes with significant autochthonous DOM production, mineralization is higher (Gudasz et al. 2012). An increase of aromatic DOM in aquatic ecosystems (e.g., through soil erosion, floods) may initially stimulate preferential adsorption, flocculation, and preservation of C of terrestrial origin and may therefore result in greater C sequestration initially. However, increased inputs of nutrients into aquatic ecosystems leads to more microbial-like DOM, which while also having high burial rates (e.g., Heathcote and Downing 2012; Mendonça et al. 2017a, b), is more prone to be preferentially metabolised and lost from sediments. Finally, these eutrophic ecosystems, prone to anoxia, lead to an increase in CH4 emissions (Mendonça et al. 2017a, b), effectively resulting in very little of autochthonous freshwater DOM being transported to the oceans (Regnier et al. 2013).
The heterogeneous nature of the DOM pool will influence aquatic metabolism, microbial biogeochemistry and food webs
Heterotroph and autotroph microbes mediate global biogeochemical cycles through their metabolism by using and producing the heterogeneous mixture of compounds that constitute DOM. DOM fuels the majority of in-stream and in-lake microbial processes which is dictated by the timing and composition of DOM fluxes (e.g., Battin et al. 2008; Williams et al. 2010). Typically, biolabile DOM fuels heterotrophic metabolism (e.g., Kaplan and Bott 1989) and high concentrations of protein-like DOM and dissolved free amino acids found in human-dominated catchments are often associated with high biological activity in surface waters (Duan and Bianchi 2007; Williams et al. 2010, 2013; Yamashita and Tanoue 2003). Beyond the microbial food web, DOM composition can also affect primary and secondary producers, and fish (e.g., Creed et al. 2018).
Specific DOM constituents can determine microbial community composition, microbial function and DOM degradation processes (Logue et al. 2016; Osterholz et al. 2016). Distinct microbial functions have been correlated with more biolabile, less complex, protein‐like DOM, and nutrients in agricultural landscapes, likely reflecting functional adaptation to biolabile DOM and higher nutrient inputs (Fasching et al. 2020; Ruiz-González et al. 2015; Wilhelm et al. 2014). Accordingly, functional diversity was found to be reduced in these agricultural landscapes (Fasching et al. 2020), while increased litter inputs to the littoral sediment translated into greater DOM chemical diversity and was associated with higher microbial diversity and function (Tanentzap et al. 2019). However, the links between individual DOM constituents and the metabolic potential of the microbiota remain poorly understood. The identification of such links might improve our understanding of how changes in the concentration of certain compounds in response to anthropogenic and climatic drivers will impact the microbial community composition and function.
DOM composition influences both the primary production and respiration terms of ecosystem metabolism. Primary production in lakes is controlled not only by the magnitude of DOC loads, but also by their specific light absorptivity and nutrient:C stoichiometry (Bergström and Karlsson 2019; Kelly et al. 2018; Olson et al. 2020). DOM composition also interacts with water residence time and lake thermal structure to control rates of ecosystem respiration (e.g., Berggren et al. 2010; Jones et al. 2018; Mostovaya et al. 2016). Slightly different patterns exist for temperate streams dominated by colored, aromatic, terrestrial-derived DOM and higher ecosystem respiration compared to semi-arid streams dominated by protein-like DOM (Catalán et al. 2018). The response of ecosystem respiration to DOM composition is likely complicated by the simultaneous effects of other variables (e.g., temperature) and appears to be nonlinear (Jane and Rose 2018).
The chemical composition of the DOM pool can modify nutrient cycles
The stoichiometry of DOM to nutrients plays a crucial role in regulating nutrient cycling (Berggren et al. 2015; Stutter et al. 2018; Yates et al. 2019). The DON and DOP contributions to the DOM pool are dependent on its source and composition (Pisani et al. 2020; Yates et al. 2019) and have been shown to be positively correlated to DOM bioavailability (Jansson et al. 2012; Pisani et al. 2017; Thompson and Cotner 2018). The amount of bioavailable material can range from 10 to 70% of riverine DON (Pisani et al. 2017) and 0 to 90% of DOP (Li and Brett 2013) with quantities dependant on inputs and sources of terrestrial runoff and photodegradation processes (Moran and Zepp 1997).
Changes in DOM composition will likely affect the bioavailability of N and P in aquatic ecosystems but the factors influencing mineralization and bioavailability of elements associated with DOM (N, P, and S) remains unclear. One important role of DOM in nutrient cycling is its use as a microbial substrate for the removal of reactive N (nitrate) via denitrification. In one stream study, denitrification rates were positively related to protein‐like fluorophores and negatively related to more aromatic and oxidized fractions of the DOM pool (Barnes et al. 2012). In north Florida rivers, terrestrial-derived DOM has been shown to indirectly inhibit denitrification rates via decreased autochthonous production (Fork and Heffernan 2014). In hyporheic zones, biolabile DOC supply limits denitrification (Zarnetske et al. 2011). Although few studies have directly linked DOM composition with denitrification rates, it is expected that a shift in DOM composition towards microbial-derived, low molecular weight material brought about by changes in land use may ultimately increase the microbial processing and utilization of N. Indeed, in catchments with high urban and agricultural land use, increases in DOM concentration and nutrient richness have affected both autotrophic and heterotrophic pathways of nutrient processing (Stutter et al. 2018; Juckers et al. 2013).
DOM plays a role in the transport and toxicity of metals and this varies based on its chemical composition
DOM is a complexing agent in natural waters, plays a role in the transport of metals and hydrophobic compounds (e.g., pesticides) and as such is an important factor mitigating water-borne metal toxicity. Toxicity measures (e.g., LC50) vary substantially among different DOM sources even for similar DOC concentrations (e.g., Haitzer et al. 1998; Conine et al. 2017). DOM properties measured using fluorescent fractions explain considerable variability in the protective effects against metal toxicity (e.g., Al-Reasi et al. 2012). DOM can alleviate metal toxicity differentially and both DOM composition and DOC should be considered in studies of metal toxicity.
The role of DOM in metal toxicity is best known for how it regulates mercury (Hg) dynamics. DOM composition plays an important role on the regulation of Hg bioavailability and the formation of the toxic methylmercury (MeHg) (Kim et al. 2011; Schartup et al. 2013). Organic compounds act as an electron donor during bacteria-mediated methylation and therefore DOM composition strongly affects the process (Bravo and Cosio 2020). Accordingly, in sediments from boreal lakes, phytoplankton-derived organic compounds enhanced methylation rates mediated by an increase of bacterial activity (Bravo et al. 2017). Similar results have been found in ponds, where higher methylation rates have been related to newly exported humic DOM substances and an increased production of algal-derived organic matter triggered by enhanced nutrient availability (Herrero Ortega et al. 2018) and in rivers, where autochthonous DOM was linked to higher total % of MeHg in water (Bravo et al. 2018). However, methylation rates and concentrations of MeHg are not necessarily coupled, thus in lakes dominated by terrigenous organic matter inputs the main source and driver of MeHg concentrations are in catchment sources (Bravo et al. 2017). In rivers, total Hg concentrations have been linked to streams with highly terrestrial soil-derived DOM (Bravo et al. 2017). Taken altogether, while increases in terrestrial DOM might translate into inputs of MeHg in freshwaters, increases in autochthonous and freshly exported DOM might lead to increases in methylation rates in freshwaters.
DOM composition is related to ease of treatability for drinking water utilities
The combined effects of land use change, human activities, and climate change on DOM pools pose a challenge to the production of affordable, safe, and reliable drinking water (e.g., Gagliano et al. 2020; Solomon et al. 2015). The quantity and composition of DOM in raw and finished drinking water can contribute to poor taste, odor and color, as well as the formation of potentially carcinogenic halogenated by-products (Lavonen et al. 2013, 2015; Williams et al. 2019). To avoid these problems a variety of methods are used to remove DOM from drinking water (Matilainen et al. 2010), and new methods are being developed such as granulated or powdered active carbon, anion exchange resins, and membranes of different pores sizes to remove DOM with more hydrophobic and aliphatic properties.
The efficiency with which drinking water plants can remove DOM from water supplies is variable, and depends not only on the methods employed, but also on the composition of the DOM and on other chemical and biochemical characteristics of the water. Thus, understanding the composition of the DOM can be important for supplying quality drinking water. The level of complexity at which the DOM composition needs to be characterized seems to be related to the kinds of human impacts that influence the water supply. When the human impact is browning, simple measurements of SUVA may be sufficient, with SUVA > 4 suggesting high removal efficiency and SUVA < 2 indicating DOM difficult to remove by coagulation and flocculation (Edzwald and Tobiason 1999). When the human impact is something like eutrophication or urbanization, more complex characterization of the DOM may be necessary. Typically, the most difficult to remove DOM is colorless and hydrophilic from algal and microbially-derived DOM produced within aquatic ecosystems (Liao et al. 2017), rather than the darker DOM that is of terrestrial origin (Golea et al. 2017; Lavonen et al. 2013; Yang et al. 2015). During algal blooms, in systems with higher levels of autochthonous DOM (i.e., low SUVA), and/or with chloramine treatments, N-containing and regulated by-products can be produced above what are considered safe levels even when ambient DOC are low (e.g., Golea et al. 2017; Lavonen et al. 2013; Liao et al. 2017; Yang et al. 2015).
From a societal perspective, there is a growing need to continuously monitor DOM quality in source water with techniques such as absorbance and fluorescence. To effectively remove organic contaminants from drinking water an increased knowledge of the complexity and composition of the DOM pool is needed. In eutrophic systems a molecular level understanding of the source and treated-water DOM pool are necessary to effectively remove both aliphatic DOM and cyanotoxins (Merel et al. 2013; Westrick et al. 2010). Detailed DOM characterization is also useful for identifying compounds in source water with the potential to form harmful disinfection by-products (Beggs and Summers 2011; Lavonen et al. 2013). Finally, a more thorough examination of DOM characteristics may be useful for the development of early warning indicators able to detect the contamination of drinking water with wastewater (Stedmon et al. 2011).
Concluding thoughts
There is a growing opportunity to use advanced chemical techniques to study DOM composition and gain a better mechanistic understanding of DOM sources and its processing in the environment. Striking a balance between studies that consider simple C fluxes to those that use easy-to-measure DOM composition analyses, with greater application of advanced molecular methods to better recognize underlying mechanisms will revolutionize our knowledge and understanding of the sources, transformations, and composition of DOM, its role in the global C cycle and its effects on the environment (Panel 1).
Using information on detailed DOM composition will allow us to predict and better prepare for future global changes. For example, with prolonged drought and increases in the amount and severity of forest fires, post-fire aquatic DOM will be more N-rich and higher in carbohydrate content. Similarly, in regions that become too arid and experience desertification an increase in microbial-like DOM is expected due to the loss of hydrological connections. In both scenarios and based on current observations and short-term trends, the evidence suggests DOM pools of the future will not look like what they look like today. This human derived DOM cocktail will be rich in aliphatic, polysaccharide, and N-containing compounds, but poor in the humic, aromatic, terrestrial produced compounds found in more natural DOM pools. This will increase DOM photoreactivity in aquatic ecosystems, reduce the amount of carbon buried in aquatic ecosystems, and shift how DOM is processed from the land to the ocean.
Climate change, alterations of the landscape and other global changes can impact biological, physical, and chemical properties of ecosystems. Together, these changes interact, often in nonlinear ways (e.g., D’Amario et al. 2019), to alter DOC and DOM composition, trigger significant changes to freshwater ecosystem function and, relatedly, the services they provide (e.g., nutrient cycling, primary production, provision of clean water). Advances in aquatic biogeochemistry have now made it possible to tackle questions on the effects of human population growth using DOM composition proxies so that the underlying principles in biogeochemistry can be applied to emerging global changes. But with human population growth also comes a greater detection of a human-laced DOM signature. We may be quickly losing opportunities to understand the dynamics of DOM in pristine environments. There are also very few studies examining long-term changes in DOM composition, an important undertaking to accurately document the effects of global change (Xenopoulos 2019).
DOM characterization is changing rapidly and being applied to a greater range of biogeochemical questions. New approaches to better examine DOM composition are continuously being developed and rapidly changing our view of its chemistry and reactivity in the environment. Signals of human-induced changes in the composition of DOM using multiple proxies can be used to gain a better mechanistic understanding of DOM sources, processing and sinks in the environment. The results emerging from the advancing field of DOM characterization are provoking a reconsideration of the conceptual approach to DOM in biogeochemistry.
Data availability
Data for Fig. 1 is available in the supplementary information.
References
Aitkenhead-Peterson JA, Kalbitz K (2005) Short-term response on the quantity and quality of rhizo-deposited carbon from Norway spruce exposed to low and high N inputs. J Plant Nutr Soil Sci 168:687–693. https://doi.org/10.1002/jpln.200420468
Al-Reasi HA, Smith DS, Wood CM (2012) Evaluating the ameliorative effect of natural dissolved organic matter (DOM) quality on copper toxicity to Daphnia magna: improving the BLM. Ecotoxicology 21:524–537. https://doi.org/10.1007/s10646-011-0813-z
Arango CP, Beaulieu JJ, Fritz KM, Hill BH, Elonen CM, Pennino MJ, Pennino MJ, Mayer PM, Kaushal SS, Balz AD (2017) Urban infrastructure influences dissolved organic matter quality and bacterial metabolism in an urban stream network. Freshw Biol 62:1917–1928. https://doi.org/10.1111/fwb.13035
Attermeyer KS, Andersson S, Catalán N, Einarsdottir K, Groeneveld M, Székely AJ, Tranvik LJ (2019) Potential terrestrial influence on transparent exopolymer particle concentrations in boreal freshwaters. Limnol Oceanogr 64:2455–2466. https://doi.org/10.1002/lno.11197
Aufdenkampe AK, Mayorga E, Raymond PA, Melack JM, Doney SC, Alin SR, Aalto RE, Yoo K (2011) Riverine coupling of biogeochemical cycles between land, oceans, and atmosphere. Front Ecol Environ 9:53–60. https://doi.org/10.1890/100014
Baker A, Spencer RG (2004) Characterization of dissolved organic matter from source to sea using fluorescence and absorbance spectroscopy. Sci Total Environ 333:217–232. https://doi.org/10.1016/j.scitotenv.2004.04.013
Bardgett RD, Richter A, Bol R, Garnett MH, Bäumler R, Xu X, Lopex-Capel E, Manning DAC, Hobbs PJ, Hartley IR, Wanek W (2007) Heterotrophic microbial communities use ancient carbon following glacial retreat. Biol Lett 3:487–490. https://doi.org/10.1098/rsbl.2007.0242
Barnes RT, Smith RL, Aiken GR (2012) Linkages between denitrification and dissolved organic matter quality, Boulder Creek watershed, Colorado. J Geophys Res Biogeosci. https://doi.org/10.1029/2011JG001749
Barnes RT, Butman DE, Wilson H, Raymond PA (2018) Riverine export of aged carbon driven by flow path depth and residence time. Environ Sci Technol 52:1028–1035. https://doi.org/10.1021/acs.est.7b04717
Battin TJ, Kaplan LA, Findlay S, Hopkinson CS, Marti E, Packman AI, Newbold JD, Sabater F (2008) Biophysical controls on organic carbon fluxes in fluvial networks. Nat Geosci 1:95–100. https://doi.org/10.1038/ngeo101
Battin TJ, Luyssaert S, Kaplan LA, Aufdenkampe AK, Richter A, Tranvik LJ (2009) The boundless carbon cycle. Nat Geosci 2:598–600. https://doi.org/10.1038/ngeo618
Beggs KM, Summers RS (2011) Character and chlorine reactivity of dissolved organic matter from a mountain pine beetle impacted watershed. Environ Sci Technol 45:5717–5724. https://doi.org/10.1021/es1042436
Behnke MI, Stubbins A, Fellman JB, Hood E, Dittmar T, Spencer RGM (2020) Dissolved organic matter sources in glacierized watersheds delineated through compositional and carbon isotopic modeling. Limnol Oceanogr. https://doi.org/10.1002/lno.11615
Berggren M, Laudon H, Haei M, Strom L, Jansson M (2010) Efficient aquatic bacterial metabolism of dissolved low-molecular-weight compounds from terrestrial sources. ISME J 4:408–416. https://doi.org/10.1038/ismej.2009.120
Berggren M, Sponseller RA, Alves Soares AR, Bergström AK (2015) Toward an ecologically meaningful view of resource stoichiometry in DOM-dominated aquatic systems. J Plankton Res 37:489–499. https://doi.org/10.1093/plankt/fbv018
Bergström AK, Karlsson J (2019) Light and nutrient control phytoplankton biomass responses to global change in northern lakes. Glob Chang Biol 25:2021–2029. https://doi.org/10.1111/gcb.14623
Bhatia MP, Das SM, Longnecker K, Charette MA, Kujawinski EB (2010) Molecular characterization of dissolved organic matter associated with the Greenland ice sheet. Geochim Cosmochim Acta 74:3768–3784. https://doi.org/10.1016/j.gca.2010.03.035
Bond-Lamberty B, Wang C, Gower ST (2004) Net primary production and net ecosystem production of a boreal black spruce wildfire chronosequence. Glob Chang Biol 10:473–487. https://doi.org/10.1111/j.1529-8817.2003.0742.x
Boodoo KS, Fasching C, Battin T (2020) Sources, transformation and fate of dissolved organic matter in the gravel bar of a pre-alpine stream. J Geophys Res Biogeosci. https://doi.org/10.1029/2019JG005604
Bragazza L, Freeman C, Jones T, Rydin H, Limpens J, Fenner N, Ellis T, Gerdol R, Hájek M, Hájek T, Iacumin P, Kutnar L, Tahvanainen T, Toberman H (2006) Atmospheric nitrogen deposition promotes carbon loss from peat bogs. PNAS 103:19386–19389. https://doi.org/10.1073/pnas.0606629104
Bravo AG, Cosio C (2020) Biotic formation of methylmercury: a bio–physico–chemical conundrum. Limnol Oceanogr 65:1010–1027. https://doi.org/10.1002/lno.11366
Bravo AG, Bouchet S, Tolu J, Björn E, Mateos-Rivera A, Bertilsson S (2017) Molecular composition of organic matter controls methylmercury formation in boreal lakes. Nat Commun 8:1–9. https://doi.org/10.1038/ncomms14255
Bravo A, Kothawala DN, Attermeyer K, Tessier E, Bodmer P, Ledesma JLJ, Audet J, Casas-Ruíz JP, Catalán N, Cauvy-Fraunié S, Colls M, Deininger A, Evtimova VV, Fonvielle JA, Fuß T, Gilbert P, Herrero Ortega S, Liu L, Mendoza-Lera C, Monteiro J, Mor JR, Nagler M, Niedrist GH, Nydahl AC, Pastor A, Pegg J, Gutmann Roberts C, Pilotto F, Portela AP, González-Quijano CR, Romero F, Rulík M, Amouroux D (2018) The interplay between total mercury, methylmercury and dissolved organic matter in fluvial systems: a latitudinal study across Europe. Water Res 144:172–182. https://doi.org/10.1016/j.watres.2018.06.064
Brighenti S, Tolotti M, Bruno MC, Wharton G, Pusch MT, Bertoldi W (2019) Ecosystem shifts in Alpine streams under glacier retreat and rock glacier thaw: a review. Sci Total Environ 675:542–559. https://doi.org/10.1016/j.scitotenv.2019.04.221
Brookshire ENJ, Valett HM, Thomas SA, Webster JR (2007) Atmospheric N deposition increases organic N loss from temperate forests. Ecosystems 10:252–262. https://doi.org/10.1007/s10021-007-9019-x
Butman D, Raymond PA, Butler K, Aiken GR (2012) Relationships between D14C, and the molecular quality of dissolved organic carbon in rivers draining to the coast from the conterminous United States. Glob Biogeochem Cycles. https://doi.org/10.1029/2012GB004361
Butman DE, Wilson HF, Barnes RT, Xenopoulos MA, Raymond PA (2015) Increased mobilization of aged carbon to rivers by human disturbance. Nat Geosci 8:112–116. https://doi.org/10.1038/NGEO2322
Butman D, Stackpoole S, Stets E, McDonald CP, Clow DW, Striegl RG (2016) Aquatic carbon cycling in the conterminous United States and implications for terrestrial carbon accounting. Proc Natl Acad Sci USA 113:58–63. https://doi.org/10.1073/pnas.1512651112
Catalán N, Obrador B, Alomar C, Pretus JL (2013) Seasonality and landscape factors drive dissolved organic matter properties in Mediterranean ephemeral washes. Biogeochemistry 112:261–274. https://doi.org/10.1007/s10533-012-9723-2
Catalán N, Marcé R, Kothawala DN, Tranvik LJ (2016) Organic carbon decomposition rates controlled by water retention time across inland waters. Nat Geosci 9:501–504. https://doi.org/10.1038/ngeo2720
Catalán N, Ortega SH, Gröntoft H, Hilmarsson TG, Bertilsson S, Wu P, Levanoni O, Bishop K, Bravo AG (2017) Effects of beaver impoundments on dissolved organic matter quality and biodegradability in boreal riverine systems. Hydrobiologia 793:135–148. https://doi.org/10.1007/s10750-016-2766-y
Catalán N, Casas-Ruiz JP, Arce MI, Abril M, Bravo AG, Del Campo R, Gómez-Gener L (2018) Behind the scenes: mechanisms regulating climatic patterns of dissolved organic carbon uptake in headwater streams. Glob Biogeochem Cycles 32:1528–1541. https://doi.org/10.1029/2018GB005919
Chen M, He W, Choi I, Hur J (2016) Tracking the monthly changes of dissolved organic matter composition in a newly constructed reservoir and its tributaries during the initial impounding period. Environ Sci Pollut Res 23:1274–1283. https://doi.org/10.1007/s11356-015-5350-5
Cole JJ, Prairie YT, Caraco NF, McDowell WH, Tranvik LJ, Striegl RG, Duarte CM, Kortelainen P, Downing JA, Middelburg JJ, Melack J (2007) Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget. Ecosystems 10:172–185. https://doi.org/10.1007/s10021-006-9013-8
Conine AL, Rearick DC, Xenopoulos MA, Frost PC (2017) Variable silver nanoparticle toxicity to Daphnia in boreal lake. Aquat Toxicol 192:1–6
Cory RM, McKnight DM, Chin Y-P, Miller P, Jaros CL (2007) Chemical characteristics of fulvic acids from Arctic surface waters: microbial contributions and photochemical transformations. J Geophys Res Biogeosci 112:G04S51. https://doi.org/10.1029/2006JG000343
Creed IF, Bergström AK, Trick CG, Grimm NB, Hessen DO, Karlsson J, Kidd KA, Kritzberg E, McKnight DM, Freeman EC, Senar OE, Andersson A, Ask J, Berggren M, Cherif M, Giesler R, Hotchkiss ER, Kortelainen P, Palta MM, Vrede T, Weyhenmeyer GA (2018) Global change-driven effects on dissolved organic matter composition: implications for food webs of northern lakes. Glob Chang Biol 24:3692–3714. https://doi.org/10.1111/gcb.14129
D’Amario SC, Xenopoulos MA (2015) Linking dissolved carbon dioxide to dissolved organic matter quality in streams. Biogeochemistry 126:99–114. https://doi.org/10.1007/s10533-015-0143-y
D’Amario SC, Rearick DC, Fasching C, Kembel SW, Porter-Goff E, Spooner DE, Williams CJ, Xenopoulos MA (2019) The prevalence of nonlinearity and detection of ecological breakpoints across a land use gradient in streams. Sci Rep 9:1–11. https://doi.org/10.1038/s41598-019-40349-4
Davis CJ, Fritsen CH, Wirthlin ED, Memmott JC (2011) High rates of primary productivity in a semi-arid tailwater: implications for self-regulated production. River Res Appl 28:1820–1829. https://doi.org/10.1002/rra.1573
De Vicente I, Ortega-Retuerta E, Mazuecos IP, Pace ML, Cole JJ, Reche I (2010) Variation in transparent exopolymer particles in relation to biological and chemical factors in two contrasting lake districts. Aquat Sci 72:443–453. https://doi.org/10.1007/s00027-010-0147-6
Drake TW, Wickland KP, Spencer RG, McKnight DM, Striegl RG (2015) Ancient low–molecular-weight organic acids in permafrost fuel rapid carbon dioxide production upon thaw. PNAS 112:13946–13951. https://doi.org/10.1073/pnas.1511705112
Drake TW, Guillemette F, Hemingway JD, Chanton JP, Podgorski DC, Zimov NS, Spencer RG (2018a) The ephemeral signature of permafrost carbon in an Arctic fluvial network. J Geophys Res Biogeosci 123:1475–1485. https://doi.org/10.1029/2017JG004311
Drake TW, Raymond PA, Spencer RG (2018b) Terrestrial carbon inputs to inland waters: a current synthesis of estimates and uncertainty. Limnol Oceanogr Lett 3:132–142. https://doi.org/10.1002/lol2.10055
Drake TW, Van Oost K, Barthel M, Bauters M, Hoyt AM, Podgoski DC, Six J, Boeckx P, Trumbore SE, Cizungu Ntaboba L, Spencer RGM (2019) Mobilization of aged and biolabile soil carbon by tropical deforestation. Nat Geosci 12:541–546. https://doi.org/10.1038/s41561-019-0384-9
Duan S, Bianchi TS (2007) Particulate and dissolved amino acids in the lower Mississippi and Pearl Rivers (USA). Mar Chem 107:214–229. https://doi.org/10.1016/j.marchem.2007.07.003
Dubnick A, Barker J, Sharp M, Wadham J, Lis G, Telling J, Fitzsimons S, Jackson M (2010) Characterization of dissolved organic matter (DOM) from glacial environments using total fluorescence spectroscopy and parallel factor analysis. Ann Glaciol 51:111–122. https://doi.org/10.3189/172756411795931912
Edzwald JK, Tobiason JE (1999) Enhanced coagulation: US requirements and a broader view. Water Sci Technol 40:63–70. https://doi.org/10.1016/S0273-1223(99)00641-1
Ekblad A, Bastviken D (2019) Deforestation releases old carbon. Nat Geosci 12:499–500. https://doi.org/10.1038/s41561-019-0394-7
Evans CD, Chapman PJ, Clark JM, Monteith DT, Cresser MS (2006) Alternative explanations for rising dissolved organic carbon export from organic soils. Glob Chang Biol 12:2044–2053. https://doi.org/10.1111/j.1365-2486.2006.01241.x
Evans C, Goodale CL, Caporn SJM, Emmett BA, Fernandez IJ, Field CD, Findlay SEG, Lovett GM, Meesenburg H, Moldan F, Sheppard LJ (2008) Does elevated nitrogen deposition or ecosystem recovery from acidification drive increased dissolved organic carbon loss from upland soil? A review of evidence from field nitrogen addition experiments. Biogeochemistry 91:13–35. https://doi.org/10.1007/s10533-008-9256-x
Ewing SA, O’Donnell JA, Aiken GR, Butler K, Butman D, Windham-Myers L, Kanevskiy MZ (2015) Long-term anoxia and release of ancient, labile carbon upon thaw of Pleistocene permafrost. Geophys Res Lett 42:10730–10738. https://doi.org/10.1002/2015GL066296
Fang H, Cheng S, Yu G, Xu M, Wang Y, Li L, Dang X, Wang L, Li Y (2014) Experimental nitrogen deposition alters the quantity and quality of soil dissolved organic carbon in an alpine meadow on the Qinghai-Tibetan Plateau. Appl Soil Ecol 81:1–11. https://doi.org/10.1016/j.apsoil.2014.04.007
Fasching C, Battin TJ (2012) Exposure of dissolved organic matter to UV-radiation increases bacterial growth efficiency in a clear-water Alpine stream and its adjacent groundwater. Aquat Sci 74:143–153. https://doi.org/10.1007/s00027-011-0205-8
Fasching C, Ulseth AJ, Schelker J, Steniczka G, Battin TJ (2016) Hydrology controls dissolved organic matter export and composition in an Alpine stream and its hyporheic zone. Limnol Oceanogr 61:558–571. https://doi.org/10.1002/lno.10232
Fasching C, Wilson HF, D’Amario SC, Xenopoulos MA (2019) Natural land cover in agricultural catchments alters flood effects on DOM composition and decreases nutrient levels in streams. Ecosystems 22:1530–1545. https://doi.org/10.1007/s10021-019-00354-0
Fasching C, Akotoye C, Bižić M, Fonvielle J, Ionescu D, Mathavarajah S, Zoccarato L, Walsh D, Grossart HP, Xenopoulos MA (2020) Linking stream microbial community functional genes to dissolved organic matter and inorganic nutrients. Limnol Oceanogr. https://doi.org/10.1002/lno.11356
Findlay SE (2005) Increased carbon transport in the Hudson River: unexpected consequence of nitrogen deposition? Front Ecol Environ 3:133–137
Findlay S, Sobczak WV (1996) Variability in removal of dissolved organic carbon in hyporheic sediments. J N Am Benthol Soc 15:35–41. https://doi.org/10.2307/1467431
Fork M, Heffernan JB (2014) Direct and indirect effects of dissolved organic matter source and concentration on denitrification in northern Florida rivers. Ecosystems 17:14–28
Frost PC, Larson JH, Johnston CA, Young KC, Maurice PA, Lamberti GA, Bridgham SD (2006) Landscape predictors of stream dissolved organic matter concentration and physicochemistry in a Lake Superior river watershed. Aquat Sci 68:40–51. https://doi.org/10.1007/s00027-005-0802-5
Fulton JR, McKnight DM, Cory RM, Stedmon C, Blunt E, Foreman CM (2004) Changes in fulvic acid redox state through the oxycline of a permanently ice-covered Antarctic lake. Aquat Sci 66:27–46. https://doi.org/10.1007/s00027-003-0691-4
Gagliano E, Sgroi M, Falciglia PP, Vagliasindi FG, Roccaro P (2020) Removal of poly-and perfluoroalkyl substances (PFAS) from water by adsorption: role of PFAS chain length, effect of organic matter and challenges in adsorbent regeneration. Water Res. https://doi.org/10.1016/j.watres.2019.115381
Golea DM, Upton A, Jarvis P, Moore G, Sutherland K, Parsons SA, Judd SJ (2017) THM and HAA formation from NOM in raw and treated surface waters. Water Res 112:226–235. https://doi.org/10.1016/j.watres.2017.01.051
Graeber D, Gelbrecht J, Pusch MT, Anlanger C, von Schiller D (2012) Agriculture has changed the amount and composition of dissolved organic matter in Central European headwater streams. Sci Total Environ 438:435–446. https://doi.org/10.1016/j.scitotenv.2012.08.087
Graeber D, Gücker B, Wild R et al (2019) Biofilm-specific uptake does not explain differences in whole-stream DOC tracer uptake between a forest and an agricultural stream. Biogeochemistry 144:85–101. https://doi.org/10.1007/s10533-019-00573-6
Griffiths NA, Tank JL, Royer TV, Warrner TJ, Frauendorf TC, Rosi-Marshall EJ, Whiles MR (2012) Temporal variation in organic carbon spiraling in Midwestern agricultural streams. Biogeochemistry 108:149–169. https://doi.org/10.1007/s10533-011-9585-z
Groeneveld M, Catalán N, Attermeyer K, Hawkes JA, Einarsdóttir K, Kothawala DN, Bergquist J, Tranvik L (2020) Selective adsorption of terrestrial dissolved organic matter to inorganic surfaces along a boreal inland water continuum. J Geophys Res Biogeosci. https://doi.org/10.1029/2019JG005236
Grossart HP, Berman T, Simon M, Pohlmann K (1998) Occurrence and microbial dynamics of macroscopic organic aggregates (lake snow) in Lake Kinneret, Israel, in fall. Aquat Microb Ecol 14:59–67. https://doi.org/10.3354/ame014059
Gudasz C, Bastviken D, Premke K, Steger K, Tranvik LJ (2012) Constrained microbial processing of allochthonous organic carbon in boreal lake sediments. Limnol Oceanogr 57:163–175. https://doi.org/10.4319/lo.2012.57.1.0163
Guillemette F, von Wachenfeldt E, Kothawala DN, Bastviken D, Tranvik LJ (2017) Preferential sequestration of terrestrial organic matter in boreal lake sediments. J Geophys Res Biogeosci 122:863–874. https://doi.org/10.1002/2016JG003735
Haitzer M, Höss S, Traunspurger W, Steinberg C (1998) Effects of dissolved organic matter (DOM) on the bioconcentration of organic chemicals in aquatic organisms: a review. Chemosphere 37:1335–1362. https://doi.org/10.1016/S0045-6535(98)00117-9
Hallema DW, Sun G, Bladon KD, Norman SP, Caldwell PV, Liu Y, McNulty SG (2017) Regional patterns of postwildfire streamflow response in the Western United States: the importance of scale-specific connectivity. Hydrol Process 31:2582–2598. https://doi.org/10.1002/hyp.11208
Hanson PC, Pace ML, Carpenter SR, Cole JJ, Stanley EH (2015) Integrating landscape carbon cycling: research needs for resolving organic carbon budgets of lakes. Ecosystems 18:363–375. https://doi.org/10.1007/s10021-014-9826-9
Harvey J, Gooseff M (2015) River corridor science: hydrologic exchange and ecological consequences from bedforms to basins. Water Resour Res 51:6893–6922. https://doi.org/10.1002/2015WR017617
Heathcote AJ, Downing JA (2012) Impacts of eutrophication on carbon burial in freshwater lakes in an intensively agricultural landscape. Ecosystems 15:60–70. https://doi.org/10.1007/s10021-011-9488-9
Herrero Ortega S, Catalán N, Björn E, Gröntoft H, Hilmarsson TG, Bertilsson S, Wu P, Bishop K, Levanoni O, Bravo AG (2018) High methylmercury formation in ponds fueled by fresh humic and algal derived organic matter. Limnol Oceanogr. https://doi.org/10.1002/lno.10722
Heslop JK, Winkel M, Walter Anthony KM, Spencer RGM, Podgorski DC, Zito P, Kholodov A, Zhang M, Liebner S (2019) Increasing organic carbon biolability with depth in yedoma permafrost: ramifications for future climate change. J Geophys Res Biogeosci 124:2021–2038. https://doi.org/10.1029/2018JG004712
Hood E, Scott D (2008) Riverine organic matter and nutrients in southeast Alaska affected by glacial coverage. Nat Geosci 1:583–587. https://doi.org/10.1038/ngeo280
Hood E, Gooseff MN, Johnson SL (2006) Changes in the character of stream water dissolved organic carbon during flushing in three small watersheds, Oregon. J Geophys Res Biogeosci. https://doi.org/10.1029/2005JG000082
Hood E, Fellman J, Spencer RGM, Hernes PJ, Edwards R, D’Amore D, Scott D (2009) Glaciers as a source of ancient and labile organic matter to the marine environment. Nature 462:1044–1048. https://doi.org/10.1038/nature08580
Hood E, Battin TJ, Fellman J, O’neel S, Spencer RG (2015) Storage and release of organic carbon from glaciers and ice sheets. Nat Geosci 8:91–96. https://doi.org/10.1038/ngeo2331
Hornberger GM, Bencala KE, McKnight DM (1994) Hydrological controls on dissolved organic carbon during snowmelt in the Snake River near Montezuma, Colarado. Biogeochemistry 25:147–165. https://doi.org/10.1007/BF00024390
Hosen JD, McDonough OT, Febria CM, Palmer MA (2014) Dissolved organic matter quality and bioavailability changes across an urbanization gradient in headwater streams. Environ Sci Technol 48:7817–7824. https://doi.org/10.1021/es501422z
Hosen JD, Aho KS, Fair JH, Kyzivat ED, Matt S, Morrison J, Stubbins A, Weber LC, Yoon B, Raymond PA (2020) Source switching maintains dissolved organic matter chemostasis across discharge levels in a large temperate river network. Ecosystems. https://doi.org/10.1007/s10021-020-00514-7
Hotchkiss ER, Hall RO Jr, Sponseller RA, Butman D, Klaminder J, Laudon H, Rosvall M, Karlsson J (2015) Sources of and processes controlling CO 2 emissions change with the size of streams and rivers. Nat Geosci 8:696–699. https://doi.org/10.1038/ngeo2507
Huss M, Hock R (2018) Global-scale hydrological response to future glacier mass loss. Nat Clim Chang 8:135–140. https://doi.org/10.1038/s41558-017-0049-x
Inamdar S, Singh S, Dutta S, Levia D, Mitchell M, Scott D, Bais H, McHale P (2011) Fluorescence characteristics and sources of dissolved organic matter for stream water during storm events in a forested mid-Atlantic watershed. J Geophys Res Biogeosci 116:G3. https://doi.org/10.1029/2011JG001735
Inamdar S, Finger N, Singh S, Mitchell M, Levia D, Bais H, Scott D, McHale P (2012) Dissolved organic matter (DOM) concentration and quality in a forested mid-Atlantic watershed, USA. Biogeochemistry 108:55–76. https://doi.org/10.1007/s10533-011-9572-4
Jaffe R, McKnight DM, Maie N, Cory RM, McDowell WH, Campbell JL (2008) Spatial and temporal variations in DOM composition in ecosystems: the importance of long-term monitoring of optical properties. J Geophys Res 113:G4. https://doi.org/10.1029/2008JG000683
Jane SF, Rose KC (2018) Carbon quality regulates the temperature dependence of aquatic ecosystem respiration. Freshw Biol 63:1407–1419. https://doi.org/10.1111/fwb.13168
Jane SF, Winslow LA, Remucal CK, Rose KC (2017) Long-term trends and synchrony in dissolved organic matter characteristics in Wisconsin, USA, lakes: quality, not quantity, is highly sensitive to climate. J Geophys Res Biogeosci 122:546–561. https://doi.org/10.1002/2016JG003630
Jansson M, Berggren M, Laudon H, Jonsson A (2012) Bioavailable phosphorus in humic headwater streams in boreal Sweden. Limnol Oceanogr 57:1161–1170. https://doi.org/10.4319/lo.2012.57.4.1161
Jessen GL, Lichtschlag A, Ramette A, Pantoja S, Rossel PE, Schubert CJ, Struck U, Boetius A (2017) Hypoxia causes preservation of labile organic matter and changes seafloor microbial community composition (Black Sea). Sci Adv 3:e1601897. https://doi.org/10.1126/sciadv.1601897
Jones SE, Zwart JA, Kelly PT, Solomon CT (2018) Hydrologic setting constrains lake heterotrophy and terrestrial carbon fate. Limnol Oceanogr Lett 3:256–264. https://doi.org/10.1002/lol2.10054
Juckers M, Williams CJ, Xenopoulos MA (2013) Land-use effects on resource net flux rates and oxygen demand in stream sediments. Freshw Biol 58:1405–1415. https://doi.org/10.1111/fwb.12136
Kaiser K, Zech W (1998) Soil dissolved organic matter sorption as influenced by organic and sesquioxide coatings and sorbed sulfate. Soil Sci Soc Am J 62:129–136. https://doi.org/10.2136/sssaj1998.03615995006200010017x
Kaiser K, Kalbitz K (2012) Cycling downwards–dissolved organic matter in soils. Soil Biol Biochem 52:29–32. https://doi.org/10.1016/j.soilbio.2012.04.002
Kalbitz K, Schwesig D, Rethemeyer J, Matzner E (2005) Stabilization of dissolved organic matter by sorption to the mineral soil. Soil Biol Biochem 37:1319–1331. https://doi.org/10.1016/j.soilbio.2004.11.028
Kaplan LA, Bott TL (1989) Diel fluctuations in bacterial activity on streambed substrata during vernal algal blooms: effects of temperature, water chemistry, and habitat. Limnol Oceanogr 34:718–733. https://doi.org/10.4319/lo.1989.34.4.0718
Kaushal SS, Gold AJ, Bernal S, Newcomer Johnson TA, Addy K, Burgin A, Burns D, Coble A, Hood E, Lu Y, Mayer P, Minor EC, Schroth AW, Vidon P, Wilson H, Xenopoulos MA, Doody T, Galella JG, Goodling P, Haviland K, Haq S, Wessel B, Wood KL, Jaworski N, Belt KT (2018) Watershed ‘chemical cocktails’: forming novel elemental combinations in Anthropocene fresh waters. Biogeochemistry 141:281–305. https://doi.org/10.1007/s10533-018-0502-6
Kaushal SS, Wood KL, Galella JG, Gion AM, Haq S, Goodling PJ, Haviland KA, Reimer JE, Morel CJ, Wessel B, Nguyen W, Hollingsworth JW, Mei K, Leal J, Widmer J, Sharif R, Mayer PM, Newcomer Johnson TA, Newcomb KD, Smith E, Belt KT (2020) Making ‘chemical cocktails’–evolution of urban geochemical processes across the Periodic Table of elements. Appl Geochem. https://doi.org/10.1016/j.apgeochem.2020.104632
Kellerman AM, Dittmar T, Kothawala DN, Tranvik LJ (2014) Chemodiversity of dissolved organic matter in lakes driven by climate and hydrology. Nat Commun 5:1–8. https://doi.org/10.1038/ncomms4804
Kellerman AM, Kothawala DN, Dittmar T, Tranvik LJ (2015) Persistence of dissolved organic matter in lakes related to its molecular characteristics. Nat Geosci 8:454–457. https://doi.org/10.1038/ngeo2440
Kellerman AM, Guillemette F, Podgorski DC et al (2018) Unifying concepts linking dissolved organic matter composition to persistence in aquatic ecosystems. Environ Sci Technol 52:2538–2548. https://doi.org/10.1021/acs.est.7b05513
Kelly PT, Solomon CT, Zwart JA, Jones SE (2018) A framework for understanding variation in pelagic gross primary production of lake ecosystems. Ecosystems 21:1364–1376. https://doi.org/10.1007/s10021-018-0226-4
Kim M, Han S, Gieskes J, Deheyn DD (2011) Importance of organic matter lability for monomethylmercury production in sulfate-rich marine sediments. Sci Total Environ 409:778–784. https://doi.org/10.1016/j.scitotenv.2010.10.050
Knicker H (2007) How does fire affect the nature and stability of soil organic nitrogen and carbon? A review. Biogeochemistry 85:91–118. https://doi.org/10.1007/s10533-007-9104-4
Knoll LB, Vanni MJ, Renwick WH, Dittman EK, Gephart JA (2013) Temperate reservoirs are large carbon sinks and small CO2 sources: results from high resolution carbon budgets. Glob Biogeochem Cycles 27:52–64. https://doi.org/10.1002/gbc.20020
Köhler SJ, Kothawala D, Futter MN, Liungman O, Tranvik L (2013) In-lake processes offset increased terrestrial inputs of dissolved organic carbon and color to lakes. PLoS One. https://doi.org/10.1371/journal.pone.0070598
Kothawala DN, Evans RD, Dillon PJ (2006) Changes in the molecular weight distribution of dissolved organic carbon within a Precambrian shield stream. Water Resour Res. https://doi.org/10.1029/2005WR004441
Kothawala DN, Roehm C, Blodau C, Moore TR (2012) Selective adsorption of dissolved organic matter to mineral soils. Geoderma 189:334–342. https://doi.org/10.1016/j.geoderma.2012.07.001
Kothawala DN, Stedmon CA, Müller RA, Weyhenmeyer GA, Köhler SJ, Tranvik LJ (2014) Controls of dissolved organic matter quality: evidence from a large-scale boreal lake survey. Glob Chang Biol 20:1101–1114. https://doi.org/10.1111/gcb.12488
Kothawala DN, Ji X, Laudon H, Ågren AM, Futter MN, Köhler SJ, Tranvik LJ (2015) The relative influence of land cover, hydrology, and in-stream processing on the composition of dissolved organic matter in boreal streams. J Geophys Res Biogeosci 120:1491–1505. https://doi.org/10.1002/2015JG002946
Kraus TEC, Bergamaschi BA, Hernes PJ, Doctor D, Kendall C, Downing BD, Losee RF (2011) How reservoirs alter drinking water quality: organic matter sources, sinks, and transformations. Lake Reserv Manag 27:205–219. https://doi.org/10.1080/07438141.2011.597283
Kritzberg ES, Hasselquist EM, Škerlep M, Löfgren S, Olsson O, Stadmark J, Valinia S, Hansson LA, Laudon H (2020) Browning of freshwaters: consequences to ecosystem services, underlying drivers, and potential mitigation measures. Ambio 49:375–390. https://doi.org/10.1007/s13280-019-01227-5
Lambert T, Bouillon S, Darchambeau F, Morana C, Roland FA, Descy JP, Borges AV (2017) Effects of human land use on the terrestrial and aquatic sources of fluvial organic matter in a temperate river basin (The Meuse River, Belgium). Biogeochemistry 136:191–211. https://doi.org/10.1007/s10533-017-0387-9
Larson JH, Frost PC, Xenopoulos MA, Williams CJ, Morales-Williams AM, Vallazza J, Nelson JC, Richardson JA (2014) Relationships between land cover and dissolved organic matter change along the river to lake transition. Ecosystems 17:1413–1425. https://doi.org/10.1007/s10021-014-9804-2
Laudon H, Köhler S, Buffam I (2004) Seasonal TOC export from seven boreal catchments in northern Sweden. Aquat Sci 66:223–230. https://doi.org/10.1007/s00027-004-0700-2
Lavonen EE, Gonsior M, Tranvik LJ, Schmitt-Kopplin P, Köhler SJ (2013) Selective chlorination of natural organic matter: identification of previously unknown disinfection byproducts. Environ Sci Technol 47:2264–2271. https://doi.org/10.1021/es304669p
Lavonen EE, Kothawala DN, Tranvik LJ, Gonsior M, Schmitt-Kopplin P, Köhler SJ (2015) Tracking changes in the optical properties and molecular composition of dissolved organic matter during drinking water production. Water Res 85:286–294. https://doi.org/10.1016/j.watres.2015.08.024
Li B, Brett MT (2013) The influence of dissolved phosphorus molecular form on recalcitrance and bioavailability. Environ Pollut 182:37–44. https://doi.org/10.1016/j.envpol.2013.06.024
Liao X, Chen C, Yuan B, Wang J, Zhang X (2017) Control of nitrosamines, THMs, and HAAs in heavily impacted water with O3-BAC. J Am Water Works Assoc 109:E215–E225. https://doi.org/10.5942/jawwa.2017.109.0057
Lieberman DM, Burke TA (1993) Particulate organic matter transport in the lower Colorado River, South-Western USA. Regul River 8:323–334. https://doi.org/10.1002/rrr.3450080403
Logue JB, Stedmon CA, Kellerman AM, Nielsen NJ, Andersson AF, Laudon H, Lindström ES, Kritzberg ES (2016) Experimental insights into the importance of aquatic bacterial community composition to the degradation of dissolved organic matter. ISME J 10:533–545. https://doi.org/10.1038/ismej.2015.131
Lu YH, Bauer JE, Canuel EA, Chambers RM, Yamashita Y, Jaffé R, Barrett A (2014) Effects of land use on sources and ages of inorganic and organic carbon in temperate headwater streams. Biogeochemistry 119:275–292. https://doi.org/10.1007/s10533-014-9965-2
Macalady DL, Walton-Day K (2011) Redox chemistry and natural organic matter (NOM): geochemists’ dream, analytical chemists’ nightmare. In: Tratnyek PG, Grundl TJ, Haderlein SB (eds) Aquatic redox chemistry. American Chemical Society, Washington, pp 85–111
Magill AH, Aber JD (2000) Dissolved organic carbon and nitrogen relationships in forest litter as affected by nitrogen deposition. Soil Biol Biochem 32:603–613. https://doi.org/10.1016/S0038-0717(99)00187-X
Mann PJ, Davydova A, Zimov N, Spencer RGM, Davydov S, Zimov S, Holmes RM (2012) Controls on the composition and lability of dissolved organic matter in Siberia’s Kolyma River basin. J Geophys Res Biogeosci 117:1–15. https://doi.org/10.1029/2011JG001798
Mann PJ, Eglinton TI, McIntyre CP, Zimov N, Davydova A, Vonk JE, Holmes RM, Spencer RGM (2015) Utilization of ancient permafrost carbon in headwaters of Arctic fluvial networks. Nat Commun 6:1–7. https://doi.org/10.1038/ncomms8856
Massicotte P, Asmala E, Stedmon C, Markager S (2017) Global distribution of dissolved organic matter along the aquatic continuum: across rivers, lakes and oceans. Sci Total Environ 609:180–191. https://doi.org/10.1016/j.scitotenv.2017.07.076
Matilainen A, Vepsäläinen M, Sillanpää M (2010) Natural organic matter removal by coagulation during drinking water treatment: a review. Adv Colloid Interface 159:189–197. https://doi.org/10.1016/j.cis.2010.06.007
McCallister SL, Ishikawa NF, Kothawala DN (2018) Biogeochemical tools for characterizing organic carbon in inland aquatic ecosystems. Limnol Oceanogr Lett 3:444–457. https://doi.org/10.1002/lol2.10097
McEnroe NA, Williams CJ, Xenopoulos MA, Porcal P, Frost PC (2013) Distinct optical chemistry of dissolved organic matter in urban pond ecosystems. PLoS One. https://doi.org/10.1371/journal.pone.0080334
McKnight DM, Boyer EW, Westerhoff PK, Doran PT, Kulbe T, Andersen DT (2001) Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity. Limnol Oceanogr 46:38–48. https://doi.org/10.4319/lo.2001.46.1.0038
Mendonça R, Müller RA, Clow D, Verpoorter C, Raymond P, Tranvik LJ, Sobek S (2017a) Organic carbon burial in global lakes and reservoirs. Nat Commun 8:1–7. https://doi.org/10.1038/s41467-017-01789-6
Mendonça RF, Müller RA, Tranvik LJ, Sobek S, Clow D, Verpoorter C, Raymond PA (2017b) Organic carbon burial in global lakes and reservoirs. Nat Commun 8:1–7. https://doi.org/10.1038/s41467-017-01789-6
Merel S, Walker D, Chicana R, Snyder S, Baurès E, Thomas O (2013) State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ Int 59:303–327. https://doi.org/10.1016/j.envint.2013.06.013
Milner AM, Khamis K, Battin TJ, Brittain JE, Barrand NE, Füreder L, Cauvy-Fraunié S, Gíslason GM, Jacobsen D, Hannah DM, Hodson AJ, Hood E, Lencioni V, Ólafsson JS, Robinson CT, Tranter M, Brown LE (2017) Glacier shrinkage driving global changes in downstream systems. PNAS 114:9770–9778. https://doi.org/10.1073/pnas.1619807114
Minor EC, Swenson MM, Mattson BM, Oyler AR (2014) Structural characterization of dissolved organic matter: a review of current techniques for isolation and analysis. Environ Sci Process Impacts 16:2064–2079. https://doi.org/10.1039/C4EM00062E
Monteith DT, Stoddard JL, Evans CD, De Wit HA, Forsius M, Høgåsen T, Wilander A, Skjelkvåle BL, Jeffries DS, Vuorenmaa J, Keller B, Kopácek J, Vesely J (2007) Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 450:537–540. https://doi.org/10.1038/nature06316
Morales-Williams AM, Wanamaker AD, Williams CJ, Downing JA (2020) Eutrophication drives extreme seasonal CO2 flux in lake ecosystems. Ecosystems. https://doi.org/10.1007/s10021-020-00527-2
Moran MA, Zepp RG (1997) Role of photoreactions in the formation of biologically labile compounds from dissolved organic matter. Limnol Oceanogr 42:1307–1316. https://doi.org/10.4319/lo.1997.42.6.1307
Moran MA, Sheldon WM Jr, Zepp RG (2000) Carbon loss and optical property changes during long-term photochemical and biological degradation of estuarine dissolved organic matter. Limnol Oceanogr 45:1254–1264. https://doi.org/10.4319/lo.2000.45.6.1254
Mostovaya A, Koehler B, Guillemette F, Brunberg AK, Tranvik LJ (2016) Effects of compositional changes on reactivity continuum and decomposition kinetics of lake dissolved organic matter. J Geophys Res Biogeosci 121:1733–1746. https://doi.org/10.1002/2016JG003359
Nadon MJ, Metcalfe RA, Williams CJ, Somers KM, Xenopoulos MA (2015) Assessing the effects of dams and waterpower facilities on riverine dissolved organic matter composition. Hydrobiologia 744:145–164. https://doi.org/10.1007/s10750-014-2069-0
Nguyen ML, Baker LA, Westerhoff P (2002) DOC and DBP precursors in western US watersheds and reservoirs. J Am Water Works Assoc 94:98–112. https://doi.org/10.1002/j.1551-8833.2002.tb09474.x
Nguyen HVM, Hur J, Shin HS (2010) Changes in spectroscopic and molecular weight characteristics of dissolved organic matter in a river during a storm event. Water Air Soil Pollut 212:395–406. https://doi.org/10.1007/s11270-010-0353-9
Olson CR, Solomon CT, Jones SE (2020) Shifting limitation of primary production: experimental support for a new model in lake ecosystems. Ecol Lett 23:1800–1808
Osterholz H, Singer G, Wemheuer B, Daniel R, Simon M, Niggemann J, Dittmar T (2016) Deciphering associations between dissolved organic molecules and bacterial communities in a pelagic marine system. ISME J 10:1717–1730. https://doi.org/10.1038/ismej.2015.231
Pautler BG, Woods GC, Dubnick A, Simpson AJ, Sharp MJ, Fitzsimons SJ, Simpson MJ (2012) Molecular characterization of dissolved organic matter in glacial ice: coupling natural abundance 1H NMR and fluorescence spectroscopy. Environ Sci Technol 46:3753–3761. https://doi.org/10.1021/es203942y
Pennino MJ, Kaushal SS, Beaulieu JJ, Mayer PM, Arango CP (2014) Effects of urban stream burial on nitrogen uptake and ecosystem metabolism: implications for watershed nitrogen and carbon fluxes. Biogeochemistry 121:247–269. https://doi.org/10.1007/s10533-014-9958-1
Peter S, Isidorova A, Sobek S (2016) Enhanced carbon loss from anoxic lake sediment through diffusion of dissolved organic carbon. J Geophys Res Biogeosci 121:1959–2197. https://doi.org/10.1002/2016JG003425
Pisani O, Boyer JN, Podgorski DC, Thomas CR, Coley T, Jaffé R (2017) Molecular composition and bioavailability of dissolved organic nitrogen in a lake flow-influenced river in south Florida, USA. Aquat Sci 79:891–908. https://doi.org/10.1007/s00027-017-0540-5
Pisani O, Bosch DD, Coffin AW, Endale DM, Liebert D, Strickland TC (2020) Riparian land cover and hydrology influence stream dissolved organic matter composition in an agricultural watershed. Sci Total Environ. https://doi.org/10.1016/j.scitotenv.2020.137165
Raymond PA, Saiers JE (2010) Event controlled DOC export from forested watershed. Biogeochemistry 100:197–209. https://doi.org/10.1007/s10533-010-9416-7
Raymond PA, McClelland JW, Holmes RM, Zhulidov AV, Mull K, Peterson BJ, Striegel RG, Aiken GR, Gurtovaya TY (2007) Flux and age of dissolved organic carbon exported to the Arctic Ocean: a carbon isotopic study of the five largest arctic rivers. Glob Biogeochem Cycles. https://doi.org/10.1029/2007GB002934
Raymond PA, Saiers JE, Sobczak WV (2016) Hydrological and biogeochemical controls on watershed dissolved organic matter transport: pulse-shunt concept. Ecology 97:5–16. https://doi.org/10.1890/14-1684.1
Regnier P, Friedlingstein P, Ciais P, Mackenzie FT, Gruber N, Janssens IA, Laruelle GG, Lauerwald R, Luyssaert S, Andersson AJ, Arndt S, Arnosti C, Borges AV, Dale AW, Gallego-Sala A, Goddéris Y, Goossens N, Hartmann J, Heinze C, Ilyina T, Joos F, LaRowe DE, Leifeld J, Meysman FJR, Munhoven G, Raymond PA, Spahni R, Suntharalingam P, Thullner M (2013) Anthropogenic perturbation of the carbon fluxes from land to ocean. Nat Geosci 6:597–607. https://doi.org/10.1038/ngeo1830
Rhoades CC, Entwistle D, Butler D (2011) The influence of wildfire extent and severity on streamwater chemistry, sediment and temperature following the Hayman Fire, Colorado. Int J Wildland Fire 20:430–442. https://doi.org/10.1071/WF09086
Rhoades CC, Chow AT, Covino TP, Fegel TS, Pierson DN, Rhea AE (2019) The legacy of a severe wildfire on stream nitrogen and carbon in headwater catchments. Ecosystems 22:643–657. https://doi.org/10.1007/s10021-018-0293-6
Risse-Buhl U, Hagedorn F, Dümig A, Gessner MO, Schaaf W, Nii-Annang S, Gerull L, Mutz M (2013) Dynamics, chemical properties and bioavailability of DOC in an early successional catchment. Biogeosciences 10:4751–4765. https://doi.org/10.5194/bgd-10-1011-2013
Rockström J, Steffen W, Noone K, Persson Å, Chapin FS, Lambin EF, Lenton TM, Scheffer M, Folke C, Schellnhuber HJ, Nykvist B, de Wit CA, Hughes T, van der Leeuw S, Rodhe H, Sörlin S, Snyder PK, Costanza R, Svedin U, Falkenmark M, Karlberg L, Corell RW, Fabry VJ, Hansen J, Walker B, Liverman D, Richardson K, Crutzen P, Foley JA (2009) A safe operating space for humanity. Nature 461:472–475. https://doi.org/10.1038/461472a
Rodríguez-Cardona BM, Coble AA, Wymore AS, Kolosov R, Podgorski DC, Zito P, Spencer RGM, Prokushkin AS, McDowell WH (2020) Wildfires lead to decreased carbon and increased nitrogen concentrations in upland arctic streams. Sci Rep 10:1–9. https://doi.org/10.1038/s41598-020-65520-0
Ruiz-González C, Niño-García JP, Lapierre JF, del Giorgio PA (2015) The quality of organic matter shapes the functional biogeography of bacterioplankton across boreal freshwater ecosystems. Glob Ecol Biogeogr 24:1487–1498. https://doi.org/10.1111/geb.12356
SanClements MD, Oelsner GP, McKnight DM, Stoddard JL, Nelson SJ (2012) New insights into the source of decadal increases of dissolved organic matter in acid-sensitive lakes of the Northeastern United States. Environ Sci Technol 46:3212–3219. https://doi.org/10.1021/es204321x
Schaefer K, Zhang T, Bruhwiler L, Barrett AP (2011) Amount and timing of permafrost carbon release in response to climate warming. Tellus B Chem Phys Meteorol 63:168–180. https://doi.org/10.1111/j.1600-0889.2010.00527.x
Schartup AT, Mason RP, Balcom PH, Hollweg TA, Chen CY (2013) Methylmercury production in estuarine sediments: role of organic matter. Environ Sci Technol 47:695–700. https://doi.org/10.1021/es302566w
Schuur EA, Abbott BW, Bowden WB, Brovkin V, Camill P, Canadell JG, Chanton JP, Chapin FS III, Christensen TR, Ciais P, Crosby BT, Czimczik CI, Grosse G, Harden J, Hayes DJ, Hugelius G, Jastrow JD, Jones JB, Kleinen T, Koven CD, Krinner G, Kuhry P, Lawrence DM, McGuire AD, Natali SM, O’Donnell JA, Ping CL, Riley WJ, Rinke A, Romanovsky VE, Sannel ABK, Schädel C, Schaefer K, Sky J, Subin M, Tarnocai C, Turetsky MR, Waldrop MP, Walter Anthony KM, Wickland KP, Wilson CJ, Zimov SA (2013) Expert assessment of vulnerability of permafrost carbon to climate change. Clim Chang 119:359–374. https://doi.org/10.1007/s10584-013-0730-7
Shang P, Lu Y, Du Y, Jaffé R, Findlay RH, Wynn A (2018) Climatic and watershed controls of dissolved organic matter variation in streams across a gradient of agricultural land use. Sci Total Environ 612:1442–1453. https://doi.org/10.1016/j.scitotenv.2017.08.322
Singer GA, Fasching C, Wilhelm L, Niggemann J, Steier P, Dittmar T, Battin TJ (2012) Biogeochemically diverse organic matter in Alpine glaciers and its downstream fate. Nat Geosci 5:710–714. https://doi.org/10.1038/ngeo1581
Smith HJ, Foster RA, McKnight DM, Lisle JT, Littmann S, Kuypers MM, Foreman CM (2017) Microbial formation of labile organic carbon in Antarctic glacial environments. Nat Geosci 10:356–359. https://doi.org/10.1038/ngeo2925
Solomon CT, Jones SE, Weidel BC, Buffam I, Fork ML, Karlsson J, Larsen S, Lennon JT, Read JS, Sadro S, Saros JE (2015) Ecosystem consequences of changing inputs of terrestrial dissolved organic matter to lakes: current knowledge and future challenges. Ecosystems 18:376–389. https://doi.org/10.1007/s10021-015-9848-y
Spencer RGM, Aiken GR, Butler KD, Dornblaser MM, Striegl RG, Hernes PJ (2009) Utilizing chromophoric dissolved organic mattermeasurements to derive export and reactivity of dissolved organic carbon exported to the Arctic ocean: a case study of the Yukon River. Alaska. Geophys Res Lett 36:L06401. https://doi.org/10.1029/2008GL036831
Spencer RG, Guo W, Raymond PA, Dittmar T, Hood E, Fellman J, Stubbins A (2014) Source and biolability of ancient dissolved organic matter in glacier and lake ecosystems on the Tibetan Plateau. Geochim Cosmochim Acta 142:64–74. https://doi.org/10.1016/j.gca.2014.08.006
Spencer RG, Mann PJ, Dittmar T, Eglinton TI, McIntyre C, Holmes RM, Zimov N, Stubbins A (2015) Detecting the signature of permafrost thaw in Arctic rivers. Geophys Res Lett 42:2830–2835. https://doi.org/10.1002/2015GL063498
Spencer RGM, Kellerman AM, Podgorski DC, Macedo MN, Jankowski KJ, Nunes D, Neill C (2019) Identifying the molecular signatures of agricultural expansion in Amazonian headwater streams. J Geophys Res Biogeosci 124:1637–1650. https://doi.org/10.1029/2018JG004910
Stedmon CA, Bro R (2008) Characterizing dissolved organic matter fluorescence with parallel factor analysis: a tutorial. Limnol Oceanogr Methods 6:572–579. https://doi.org/10.4319/lom.2008.6.572
Stedmon CA, Thomas DN, Papadimitriou S et al (2011) Using fluorescence to characterize dissolved organic matter in Antarctic sea ice brines. J GeophysRes Biogeosciences 116:1–9. https://doi.org/10.1029/2011JG001716
Stubbins A, Hood E, Raymond PA, Aiken GR, Sleighter RL, Hernes PJ, Butman D, Hatcher PG, Striegl RG, Schuster P, Abdulla HA, Vermilyea AW, Scott DT, Spencer RGM (2012) Anthropogenic aerosols as a source of ancient dissolved organic matter in glaciers. Nat Geosci 5:198–201. https://doi.org/10.1038/ngeo1403
Stutter MI, Graeber D, Evans CD, Wade AJ, Withers PJA (2018) Balancing macronutrient stoichiometry to alleviate eutrophication. Sci Total Environ 634:439–447. https://doi.org/10.1016/j.scitotenv.2018.03.298
Tanentzap AJ, Fitch A, Orland C, Emilson EJ, Yakimovich KM, Osterholz H, Dittmar T (2019) Chemical and microbial diversity covary in fresh water to influence ecosystem functioning. PNAS 116:24689–24695. https://doi.org/10.1073/pnas.1904896116
Tarnocai C, Canadell JG, Schuur EAG, Kuhry P, Mazhitova G, Zimov S (2009) Soil organic carbon pools in the northern circumpolar permafrost region. Glob Biogeochem Cycles. https://doi.org/10.1029/2008GB003327
Thompson SK, Cotner JB (2018) Bioavailability of dissolved organic phosphorus in temperate lakes. Front Environ Sci. https://doi.org/10.3389/fenvs.2018.00062
Thurman EM (1985) Aquatic humic substances. Organic geochemistry of natural waters. Developments in biogeochemistry, vol 2. Springer, Dordrecht, pp 273–361. https://doi.org/10.1007/978-94-009-5095-5_11
Tranvik LJ, Downing JA, Cotner JB, Loiselle SA, Striegl RG, Ballatore TJ, Dillon P, Finlay K, Fortino K, Knoll LB, Kortelainen PL, Kutser T, Larsen S, Laurion I, Leech DM, McCallister SL, McKnight DM, Melack JM, Overholt E, Porter JA, Prairie Y, Renwick WH, Roland F, Sherman BS, Schindler DW, Sobek S, Tremblay A, Vanni MJ, Verschoor AM, von Wachenfeldt E, Weyhenmeyer GA (2009) Lakes and reservoirs as regulators of carbon cycling and climate. Limnol Oceanogr 54:2298–2314. https://doi.org/10.4319/lo.2009.54.6_part_2.2298
Ulseth AJ, Hall RO Jr (2015) Dam tailwaters compound the effects of reservoirs on the longitudinal transport of organic carbon in an arid river. Biogeosciences 12:4345–4359. https://doi.org/10.5194/bg-12-4345-2015
Vidon P, Wagner LE, Soyeux E (2008) Changes in the character of DOC in streams during storms in two Midwestern watersheds with contrasting land uses. Biogeochemistry 88:257–270. https://doi.org/10.1007/s10533-008-9207-6
von Wachenfeldt E, Tranvik LJ (2008) Sedimentation in boreal lakes—the role of flocculation of allochthonous dissolved organic matter in the water column. Ecosystems 11:803–814. https://doi.org/10.1007/s10021-008-9162-z
von Wachenfeldt E, Sobek S, Bastviken D, Tranvik LJ (2008) Linking allochthonous dissolved organic matter and boreal lake sediment carbon sequestration: the role of light-mediated flocculation. Limnol Oceanogr 53:2416–2426. https://doi.org/10.4319/lo.2008.53.6.2416
von Wachenfeldt E, Bastviken D, Tranvik LJ (2009) Microbially induced flocculation of allochthonous dissolved organic carbon in lakes. Limnol Oceanogr 54:1811–1818. https://doi.org/10.4319/lo.2009.54.5.1811
Vonk JE, Mann PJ, Davydov S, Davydova A, Spencer RG, Schade J, Sobczak WV, Zimov N, Zimov S, Bulygina E, Eglinton T, Holmes RM (2013) High biolability of ancient permafrost carbon upon thaw. Geophys Res Lett 40:2689–2693. https://doi.org/10.1002/grl.50348
Wagner S, Jaffe R, Stubbins A (2018) Dissolved black carbon in aquatic ecosystems. Limnol Oceanogr Lett 3:168–175. https://doi.org/10.1002/lol2.10076
Ward CP, Cory RM (2015) Chemical composition of dissolved organic matter draining permafrost soils. Geochim Cosmochim Acta 167:63–79. https://doi.org/10.1016/j.gca.2015.07.001
West WE, Coloso JJ, Jones SE (2012) Effects of algal and terrestrial carbon on methane production rates and methanogen community structure in a temperate lake sediment. Freshw Biol 57:949–955. https://doi.org/10.1111/j.1365-2427.2012.02755.x
Westrick JA, Szlag DC, Southwell BJ, Sinclair J (2010) A review of cyanobacteria and cyanotoxins removal/inactivation in drinking water treatment. Anal Bioanal Chem 397:1705–1714. https://doi.org/10.1007/s00216-010-3709-5
Weyhenmeyer GA, Fröberg M, Karltun E, Khalili M, Kothawala D, Temnerud J, Tranvik LJ (2012) Selective decay of terrestrial organic carbon during transport from land to sea. Glob Chang Biol 18:349–355. https://doi.org/10.1111/j.1365-2486.2011.02544.x
Whittinghill KA, Currie WS, Zak DR, Burton AJ, Pregitzer KS (2012) Anthropogenic N deposition increases soil C storage by decreasing the extent of litter decay: analysis of field observations with an ecosystem model. Ecosystems 15:450–461. https://doi.org/10.1007/s10021-012-9521-7
Wild B, Andersson A, Bröder L, Vonk J, Hugelius G, McClelland JW, Song W, Raymond PA, Gustafsson Ö (2019) Rivers across the Siberian Arctic unearth the patterns of carbon release from thawing permafrost. PNAS 116:10280–10285. https://doi.org/10.1073/pnas.1811797116
Wilhelm L, Besemer K, Fasching C, Urich T, Singer GA, Quince C, Battin TJ (2014) Rare but active taxa contribute to community dynamics of benthic biofilms in glacier-fed streams. Environ Microbiol 16:2514–2524. https://doi.org/10.1111/1462-2920.12392
Williams CJ, Yamashita Y, Wilson HF, Jaffé R, Xenopoulos MA (2010) Unraveling the role of land use and microbial activity in shaping dissolved organic matter characteristics in stream ecosystems. Limnol Oceanogr 55:1159–1171. https://doi.org/10.4319/lo.2010.55.3.1159
Williams CJ, Frost PC, Xenopoulos MA (2013) Beyond best management practices: pelagic biogeochemical dynamics in urban stormwater ponds. Ecol Appl 23:1384–1395. https://doi.org/10.1890/12-0825.1
Williams CJ, Frost PC, Morales-Williams AM, Larson JH, Richardson WB, Chiandet AS, Xenopoulos MA (2016) Human activities cause distinct dissolved organic matter composition across freshwater ecosystems. Glob Chang Biol 22:613–626. https://doi.org/10.1111/gcb.13094
Williams CJ, Conrad D, Kothawala DN, Baulch HM (2019) Selective removal of dissolved organic matter affects the production and speciation of disinfection byproducts. Sci Total Environ 652:75–84. https://doi.org/10.1016/j.scitotenv.2018.10.184
Williamson CE, Overholt EP, Pilla RM, Leach TH, Brentrup JA, Knoll LB, Mette EM, Moeller RE (2015) Ecological consequences of long-term browning in lakes. Sci Rep. https://doi.org/10.1038/srep18666
Wilson HF, Xenopoulos MA (2008) Ecosystem and seasonal control of stream dissolved organic carbon along a gradient of land use. Ecosystems 11:555–568. https://doi.org/10.1007/s10021-008-9142-3
Wilson HF, Xenopoulos MA (2009) Effects of agricultural land use on the composition of fluvial dissolved organic matter. Nat Geosci 2:37–41. https://doi.org/10.1038/ngeo391
Wilson HF, Xenopoulos MA (2013) Diel changes of dissolved organic matter in streams of varying watershed land use. River Res Appl 29:1330–1339. https://doi.org/10.1002/rra.2606
Wilson HF, Raymond PA, Saiers JE, Sobczak WV, Xu N (2016) Increases in humic and bioavailable dissolved organic matter in a forested New England headwater stream with increasing discharge. Mar Freshw Res 67:1279–1292. https://doi.org/10.1071/MF15286
Wohl E, Hall RO Jr, Lininger KB, Sutfin NA, Walters DM (2017) Carbon dynamics of river corridors and the effects of human alterations. Ecol Monogr 87:379–409. https://doi.org/10.1002/ecm.1261
Xenopoulos MA (2019) Long-term studies in limnology and oceanography. Limnol Oceanogr 64:S1. https://doi.org/10.1002/lno.11112
Xenopoulos MA, Downing JA, Kumar MD, Menden-Deuer S, Voss M (2017) Headwaters to oceans: ecological and biogeochemical contrasts across the aquatic continuum. Limnol Oceanogr 62:S3–S14. https://doi.org/10.1002/lno.10721
Xu H, Guo L, Jiang H (2016) Depth-dependent variations of sedimentary dissolved organic matter composition in a eutrophic lake: implications for lake restoration. Chemosphere 145:551–559. https://doi.org/10.1016/j.chemosphere.2015.09.015
Yamashita Y, Tanoue E (2003) Chemical characterization of protein-like fluorophores in DOM in relation to aromatic amino acids. Mar Chem 82:255–271. https://doi.org/10.1016/S0304-4203(03)00073-2
Yang L, Kim D, Uzun H, Karanfil T, Hur J (2015) Assessing trihalomethanes (THMs) and N-nitrosodimethylamine (NDMA) formation potentials in drinking water treatment plants using fluorescence spectroscopy and parallel factor analysis. Chemosphere 121:84–91. https://doi.org/10.1016/j.chemosphere.2014.11.033
Yates CA, Johnes PJ, Owen AT, Brailsford FL, Glanville HC, Evans CD, Marshall MR, Jones DL, Lloyd CEM, Jickells T, Evershed RP (2019) Variation in dissolved organic matter (DOM) stoichiometry in UK freshwaters: assessing the influence of land cover and soil C: N ratio on DOM composition. Limnol Oceanogr 64:2328–2340. https://doi.org/10.1002/lno.11186
Zarnetske JP, Haggerty R, Wondzell SM, Baker MA (2011) Labile dissolved organic carbon supply limits hyporheic denitrification. J Geophys Res Biogeosci. https://doi.org/10.1029/2011JG001730
Zhou Y, Xiao Q, Yao X, Zhang Y, Zhang M, Shi K, Lee X, Podgorski DC, Spencer RGM, Jeppesen E (2018) Accumulation of terrestrial dissolved organic matter potentially enhances dissolved methane levels in eutrophic Lake Taihu, China. Environ Sci Technol 52:10297–10306. https://doi.org/10.1021/acs.est.8b02163
Zimov SA, Davydov SP, Zimova GM, Davydova AI, Schuur EAG, Dutta K, Chapin FS III (2006) Permafrost carbon: stock and decomposability of a globally significant carbon pool. Geophys Res Lett. https://doi.org/10.1029/2006GL027484
Acknowledgements
Thank you to Elizabeth Rohde for help with the reference list. Funding by Canada’s Natural Sciences and Engineering Council (NSERC) Discovery Grant to M.A.X, NSERC Undergraduate Student Research Award to E.R. and U.S. National Science Foundation Grant #1754363 to CTS.
Author information
Authors and Affiliations
Contributions
MAX coordinated the team and drafted the first outline with initial input by CF, SD, CJW and HFW, then all authors contributed intellectual content. All authors participated in writing and editing. Figures completed by CTS, KSB, DNK and MAX. Panel completed by RGMS, OP, CJW, MAX and RTB.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Responsible Editor: Robert W. Howarth.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This paper is an invited contribution to the 35th Anniversary Special Issue, edited by Sujay Kaushal, Robert Howarth, and Kate Lajtha.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xenopoulos, M.A., Barnes, R.T., Boodoo, K.S. et al. How humans alter dissolved organic matter composition in freshwater: relevance for the Earth’s biogeochemistry. Biogeochemistry 154, 323–348 (2021). https://doi.org/10.1007/s10533-021-00753-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-021-00753-3