
Abstract
The purpose of this study was to evaluate the importance of litterfall as a link in biogeochemical cycling of certain elements (C, N, P, K, Ca, Mg, Fe, Mn, Cu, and Zn) in silver birch stands growing on post-arable soils under temperate climatic conditions (Poland). The study covered three stands on sandy soils (21, 29, and 40 years old) and three on loamy soils (15, 33, and 41 years old). Litterfall was collected during 2020–2021 and analysed using standard procedures. Elemental contents were determined also in green leaves collected from the tree crowns in early summer to determine intensity of retranslocation. The total litterfall production varied from 2576 to 5293 kg ha−1, depending on the stand and year. These values are typical of temperate forests. Litterfall production was positively correlated with stand age. Leaves made up the major fraction, followed by twigs. Silver birch litterfall was generally rich in nutrients compared to many other tree species typical for temperate climatic zone. Nutrient fluxes to the soil were in the upper limits reported in the literature. It may partially be an effect of the post-arable history of these stands. Large fluxes of nutrients to the soil via litterfall are important in the context of the high nutritional demands of the studied tree species. This study also showed that silver birch strongly retranslocates N, P, and K from senescing leaves, whereas Ca, Mg, Fe, Mn, Cu, and Zn are accumulated in these organs.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Silver birch (Betula pendula Roth) is one of the major components of natural and managed forests in Europe, Asia, and North America. This widely distributed tree has adapted to function in a wide spectrum of climatic (Titmuss 1959; Browicz 1979) and site (Bown 1995) conditions. Additionally, its rapid growth, early fertility, and seed dispersal by wind explain its high succession potential (Úradníček et al. 2009; Špulák et al. 2010; Oikonomakis and Ganatsas 2020). Conversely, its low shade tolerance (Hynynen et al. 2010) can be a limiting factor on its growth in lower forest layers. Silver birch easily colonizes open spaces, including abandoned arable land (Špulák et al. 2010; Zasada et al. 2014), and post-mining and post-industrial areas (Ulbrichová et al. 2005; Franiel and Kąpała-Bąba 2021). Moreover, due to climate change and the transformation of forest management concepts, the importance of this species in forest economies has grown in many countries over the last few decades (Oksanen 2021).
Although silver birch plays an important and continuously increasing role in the landscape of the Northern Hemisphere, many aspects related to its ecology remain poorly explored. Among other things, there is still insufficient knowledge concerning the interrelationship between silver birch and the soil, especially its effect on soil properties and processes. Some aspects of this issue have been explored by Mikola et al. (2018), Silfver et al. (2007), and Silfver et al. (2018), who studied effects of silver birch genotype on N use efficiency, litterfall chemistry, its decomposition intensity, and soil microbiome. Possen et al. (2021) highlighted importance of that element in survival and adaptation of silver birch to climate changes. Ostonen et al. (2007) and Rosenvald et al. (2011) studied importance of soil characteristics as factors influencing development of root system. Although certain data are available, a comprehensive assessment of the impact of birch on the soil environment is currently difficult. Major gaps in the knowledge concerning this issue were highlighted by Jonczak et al. (2020) in review paper. According to the authors, the effects of silver birch on the stocks and quality of soil organic matter (SOM) and on nutrient cycling (bioavailability, uptake, distribution in the biomass, and return to the soil) are among the least-explored topics. In addition, the results of published studies are sometimes divergent.
Considering the widespread distribution and rapid growth of silver birch, significant effects on nutrient cycling in terrestrial ecosystems should be expected. Previous studies (e.g. Ferm 1993) have demonstrated the high sensitivity of that tree to soil nitrogen (N), phosphorus (P), and potassium (K) contents. This observation is partially inconsistent with the general opinion that silver birch has a high tolerance for variable environmental conditions. Birch trees intensively uptake N (56 kg ha−1 year−1), P (4.1 kg ha−1 year−1), K (28 kg ha−1 year−1), and calcium (Ca, 44 kg ha−1 year−1) (Ovington and Madgwick 1959), with the process reaching a maximum intensity in the early stages of growth (Miller 1984). Daugaviete et al. (2003) and Aosaar et al. (2016) reported positive effects of birch trees on soil N during the first few years after afforestation. Additionally, Uri et al. (2012) and Gawęda et al. (2014) observed increasing contents of the major nutrients in soils under birch chronosequences. Birch biomass is generally moderately abundant in nutrients; however, their concentrations are strongly controlled by their bioavailability (Daugaviete et al. 2015). Nutrient distribution is uneven in tree organs, usually reaching their highest contents in the leaves, followed by the fine roots or bark, with the lowest in the stemwood (Rustowska 2022). According to Gawęda et al. (2014), the nutrient allocation in a 12-year-old silver birch stand was 543.41 kg ha−1 N, 60.03 kg ha−1 P, 186.42 kg ha−1 K, 220.40 kg ha−1 Ca, and 50.48 kg ha−1 magnesium (Mg) in the aboveground biomass, and 59.67, 13.44, 33.09, 40.55, and 6.39 kg ha−1 in the roots, respectively.
Elements contained in the biomass return to the soil via litterfall, the production and transformation of which is of key importance in the functioning of forest ecosystems, constituting an important link in the biogeochemical cycling of matter and energy (Nordén 1994). Hence, litterfall is considered to be one of the major mechanisms influencing soil cover by forest vegetation. Therefore, it can be applied as a tool in the assessment of the potential effects of various tree species on the soil system (Augusto et al. 2002). Moreover, the Loydi et al. (2014) reported that in forest-field ecotone zone litterfall strongly influence species composition of vegetation, constituting factor of biodiversity. The annual production of litterfall varies strongly across climatic zones and plant communities (Neumann et al. 2018; Shen et al. 2019). The same factors, among many others, also influence its chemistry (Augusto et al. 2002; Jonczak 2011). Unfortunately, studies on birch litterfall have rarely been reported in the literature. Previous works have covered several case studies focused on litter production (Aussenac et al. 1972; Tripathi et al. 2006) and the major nutrient contents (Mork 1942; Berg and Staaf 1987; Perala and Alm 1990; Johansson 1995; Brandtberg et al. 2004; Osono and Takeda 2004; Carnol and Bazgir 2013). Based on the available data, it can be concluded that litterfall production in birch stands has values that are typical of temperate forests (Shen et al. 2019), whereas its abundance in the majority of nutrients falls in the upper limits reported for common tree species in that climatic zone. A broader study providing a detailed characterization of the plant litterfall in pure birch stands has not been undertaken until now.
Considering the ecological and utilitarian importance of the issue, we undertook a broad study on litterfall production and chemistry in silver birch stands growing on post-arable soils in a temperate climatic zone. Three major factors of variability were included in this study––stand age, site fertility (sandy and loamy soils), and time (2-year study period). Moreover, the aspect of nutrients retranslocation from senescing leaves to branches was also studied. Based on the available literature data we expected litterfall production at levels typical for temperate forests and moderate-high abundance in nutrients. We also expected considerable effect of stand age and site fertility as factors influencing litterfall chemistry.
Materials and methods
Study area
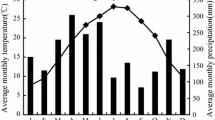
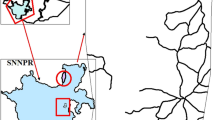
The study was performed in central Poland (Fig. 1), representing a cold climatic zone, with warm summers and a humid continental climate, according to the Köppen–Geiger classification (Peel et al. 2007). The mean annual temperature from the closest meteorological station (Skierniewice) was 8.3 °C and the mean annual sum of precipitation was 541.4 mm for the period 1951–2020. The warmest month was July, while the coldest was January. July was the month with the highest sum of precipitation, while January had the lowest. The year 2020 was characterized by higher average air temperatures––both annual and in the growing season––than the long-term norm (9.9 °C), whereas in the year 2021, the air temperature was close to the multiannual mean value (8.3 °C). The annual sum of precipitation in 2020 and 2021 was 673.3 and 685.4 mm, respectively. This constituted more than 120% of the norm for the period 1951–2020.

Location of the studied silver birch stands
The study covered six stands of silver birch. Three of these (S-21, S-29, and S-40) were located on sandy soils (Dystric Brunic Arenosols), and three (L-15, L-33, and L-41) on loamy soils (Eutric Stagnosols, Eutric Cambisols, and Dystric Cambisols), based on the World Reference Base (WRB) system of the International Union of Soil Sciences Working Group (IUSS 2015) classification. In addition, the stands differed in terms of age, density, and mean diameter. Admixtures of common aspen and grey alder were observed in two stands. The basic characteristics of the stands are included in Table 1.
Soil sampling and analysis
The study was conducted on 20 × 25-m plots established in representative parts of each stand. One soil pit was dug in the central part of each plot. The soils were described using Food and Agriculture Organization (FAO) of the United Nations criteria (FAO 2006), classified according to the WRB system (IUSS 2015), sampled, and prepared for further analysis. In addition, the A horizons were sampled from 20 locations distributed on a regular grid. One disturbed sample and one 100-cm3 undisturbed sample were taken from each location. The soil analyses included the potentiometric determination of pH in a suspension with deionized water (at a 1:2.5 ratio), the total organic carbon (TOC) and N contents by dry combustion (Vario MacroCube, Elementar, Germany), and the total contents of P, K, Ca, Mg, iron (Fe), manganese (Mn), copper (Cu), and zinc (Zn) by inductively coupled plasma–atomic emission spectroscopy (ICP–AES, Avio 200, PerkinElmer, USA) after microwave digestion of the samples (Milestone EthosUp, Italy) in a mixture of 40% hydrofluoric acid (HF), 65% nitric acid (HNO3), and 38% hydrochloric acid (HCl) (5:3:2 by volume).
Litterfall study
The litterfall study was conducted in 2020 and 2021 (except for in Stand L-33, which was covered only in the first year). Litterfall was collected using 10 traps (50 cm in diameter) per stand at quarterly intervals (end of each season). It was dried at 65 °C, divided into fractions (leaves, twigs, flowers, and seeds separately for each tree species, and other components, including bark and small parts difficult to identify), and weighed. Then, an average sample of each fraction was milled into powder. The TOC and N were determined by dry combustion (Vario MacroCube, Elementar, Germany), and the total contents of P, K, Ca, Mg, Fe, Mn, Cu, and Zn by ICP–AES (Avio 200, PerkinElmer, USA) prior microwave digestion (Milestone EthosUp, Italy) in 65% HNO3. Moreover, green silver birch leaves were taken from each stand at the beginning of July 2020 and 2021. Leaves have been collected from three trees per stand, from central parts of their crowns. The collected material was carefully mixed and finally one sample represented each stand. The samples have been dried and analysed using the sample procedures as for litterfall. Only pure per-analysis reagents were used for the analyses. Quality control of the analyses was ensured by using certified reference materials.
Statistical analysis
The statistical analyses included the weighted mean concentrations of the studied elements in the litterfall fractions and the total litterfall, and their annual fluxes to the soil via the litterfall, as a function of the litterfall mass per area unit and element content. In addition, based on the elemental concentrations in the green leaves (collected from the central parts of the crowns of the trees at the beginning of July) and in the fallen leaves during autumn, retranslocation coefficients were calculated according to the formula: ([element content in green leaves – element content in fallen leaves] ∙ 100%)/element content in green leaves. Based on this formula, values above 0 indicated element retranslocation from senescing leaves to the branches, whereas values below 0 represented accumulation in the leaves. Additionally, correlation coefficients between the elemental contents in the litterfall and soils were calculated. The characteristics of silver birch litterfall were compared between the stands using cluster analysis (Ward method, Euclidean distance). Principle-Component Analysis (PCA) was applied to investigate relationships between silver birch litterfall characteristics, stand age, and soil characteristics. All analyses were performed using Statistica 13 software (Dell 2022).
Results
Basic characteristics of the soils
The soils of the studied plots differed in terms of classification and certain characteristics. Their A horizons were acidic, with pH-H2O values ranging from 4.07 to 5.30 (Table 2). The soils contained 9.13–12.97 g kg−1 TOC and 0.62–0.91 g kg−1 N in the A horizons. The contents of the remaining elements were as follows: 0.40–0.83 g kg−1 P; 7.69–11.46 g kg−1 K; 1.21–2.07 g kg−1 Ca; 0.35–0.74 g kg−1 Mg; 4.65–8.18 g kg−1 Fe; 106.35–396.46 mg kg−1 Mn; 1.09–4.50 mg kg−1 Cu, and 13.05–36.21 mg kg−1 Zn. Generally, the soils were moderately abundant in TOC and N, but poor in K, Ca, Mg, and Fe. The Mn, Cu, and Zn contents were typical of the uncontaminated areas of central Poland, showing some variability affected by texture. Based on the obtained results, it can be stated that pH and elemental contents in A horizons were rather a function of stand than groups distinguished based on texture (sandy and loamy soils). Clear differences between those groups can be observed only for K, Mn, and Cu (higher contents in loamy soils).
Litterfall production and botanical composition
The total litterfall production in the studied stands varied from 2576 to 4761 kg ha−1 in 2020 and from 2601 to 5293 kg ha−1 in 2021 (Table 3). Silver birch parts were major components of the litterfall; however, in stands S-29, S-40, L-33, and L-41, there were admixtures of parts originating from other tree species, including pedunculate oak, grey alder, common aspen, and black cherry. The representation of these species in the litterfall resulted from their occurrence in the studied plots (Table 1) or from litterfall influx from external sources (neighbouring stands). Leaves were a major fraction of the silver birch litterfall, constituting 73.7–84.9% of its total mass in 2020 and 69.2–86.9% in 2021, followed by twigs––6.1–14.2% and 11.7–25.5%, respectively (Fig. 2).

Structure of silver birch litterfall in the years 2020 and 2021
Litterfall chemistry
The weighted mean elemental contents in the total litterfall varied among the stands and from year to year (Supplementary material). The least variable was TOC. Relatively low variability was also typical of N, P, Ca, Mg, and Cu, whereas it was greater for K, Fe, Mn, and Zn. The observed elemental contents in the total birch litterfall did not differ considerably from the total litterfall (Table 4). Silver birch litterfall showed large variability year to year, as evidenced by cluster analysis (Fig. 3). The stands do not form groups either in terms of age or soil fertility (sandy, loamy soils).

Cluster analysis (Ward method, Euclidean distance) of birch litterfall in the years 2020 and 2021 (based on variables: N, P, K, Ca, Mg, Fe, Mn, Cu, and Zn)
Total silver birch litterfall chemistry as a function of stand, soil characteristics, and time was visualized using PCA analysis (Fig. 4). Two major principal components (PC 1 and PC 2) explained 67.9% of the observed variability. PC 1 was affected mainly by soil characteristics, showing very strong positive correlation with majority of them. Moreover, very strong positive correlation was noted with some litterfall components, including P, Ca (first year of the study), and Fe (boith years of the study), and very strong negative correlation with Cu and Mn. PC 2 was very strongly negatively correlated with N, whereas positively with Zn. The analysis paint not fully clear picture on soil-litterfall interrelationships in terms of elemental composition, indicating an important role of time as a factor of litterfall variability.

PCA analysis of the silver birch litterfall based on elemental contents in 2020 (green lines) and 2021 (blue lines), stand age and soil characteristics (grey lines). (Colour figure online)
The elemental contents strongly varied among the silver birch litterfall fractions. Detailed information is included in supplementary material. Typically, the lowest variability was in the TOC. Nitrogen was observed in its highest amounts in flowers, followed by seeds; P usually in leaves, followed by seeds and flowers; K, Ca, and Mg in leaves, with various tendencies in the remaining fractions; and Fe in seeds and then flowers. The highest concentrations of Mn were noted in leaves, usually followed by seeds. The Cu variability was low in the majority of the stands. Its lowest concentrations were usually typical in the leaves, whereas its highest were in the twigs, flowers or seeds, depending on the stand. Similar tendencies were observed for Zn. The elemental contents in the total litterfall, silver birch total litterfall, and silver birch leaf litterfall showed significant correlations with elemental content in the associated soils in some cases (Table 5). The elements that were usually positively correlated were P, Ca, and Fe. The remaining elements showed no clear tendencies––their correlations were poor or reversed during the two years.
Chemistry of green leaves and retranslocation of nutrients
Table 6 shows elemental contents in green leaves of silver birch. The studied elements usually showed considerable variability depending on the stand and year to year. The highest variability showed Mn (CV = 44%), followed by K (CV = 29%), N (CV = 28%), Fe (CV = 26%), Zn (CV = 23%), Ca and Mg (CV = 20%), Cu (CV = 15%), and P (CV = 15%). The intensity of elemental retranslocation from senescing leaves to the branches also varied among the elements and stands, and it changed over time. Nitrogen was retranslocated at rates of 21–48% in 2020 and 59–72% in 2021. In all the stands, K was also retranslocated (9–43% in 2020 and 52–83% in 2021), as well as P, except in one case (9–43% in 2020 and 31–46% in 2021) (Table 7). Iron and Ca were accumulated in the leaves in all the stands, whereas Mg, Mn, Cu, and Zn were accumulated at most of them.
Elements returned to the soil via litterfall
Elemental inputs to the soil in forest ecosystems are strongly controlled by litterfall production and its chemical composition. The highest fluxes are always reported for carbon (C, around 50% of the litterfall mass). In the studied stands, the annual flux of TOC to the soil was estimated at 1311–2715 kg ha−1, depending on the stand and year. The second highest flux was noted for N (33.6–90.7 kg ha−1 year−1) and Ca (26.4–74.4 kg ha−1 year−1), followed by K (5.3–21.0 kg ha−1 year−1), Mg (4.9–13.9 kg ha−1 year−1), and P (2.8–8.2 kg ha−1 year−1). There was also a considerable flux of Mn (2.79–7.93 kg ha−1 year−1), whereas the fluxes of Fe, Cu, and Zn were usually below 1 kg ha−1 year−1 (Table 8).
Discussion
Litterfall production
Litterfall production is controlled by a complex of factors related to vegetation (species composition, age, density, and health condition), climatic conditions (mainly temperature and precipitation), site characteristics (soil fertility, nutrient bioavailability, water regime, contamination, elevation, slope exposition, etc.), and forest management practices (transformation of tree species composition, logging, care treatments, fertilization, etc.). The role of some of these factors as sources of litterfall variability on a global scale has been highlighted in the meta-analyses of Bray and Gorham (1964), Matthews (1997), Berg et al. (1999), Liu et al. (2004), and Shen et al. (2019). According to the latter review, the annual production of aboveground litterfall in worldwide forests ranges from around 0.15 to nearly 27 t ha−1, with values of between 2 and 6 t ha−1 being the most common. The annual litterfall production varied in the present study from 2576 to 5293 kg ha−1, depending on the stand and year (Table 3). Thus, the values are in the range of those most typical of temperate forests. The relatively wide variability observed here should have been expected, considering at least two factors: (1) the varied age and density of the stands; and (2) the varied soil fertility (sandy and loamy soils). The role of stand characteristics was confirmed by statistical analysis. The annual litterfall production was a function of tree age (R2 = 0.497 for 2020 and 0.695 for 2021). This observation confirms the findings of Lebret et al. (2001); however, Berg et al. (1999) observed an opposite tendency. At the same time, the differences resulting from site fertility (sandy and loamy soils) were less clear, and correlations with stand density were weak (R2 = − 0.377 for 2020 and − 0.445 for 2021). Generally, litterfall production in the studied stands was comparable to that reported in the literature for various birch species; however, data are scarce. Aussenac et al. (1972) estimated the annual litterfall production to be 2200 kg ha−1, whereas Tripathi et al. (2006) determined it to be 3770 kg ha−1. The silver birch litterfall consisted mainly of leaves (Fig. 2), which fell predominantly in the early autumn. This is a typical observation for temperate forests and has been reported by several authors (e.g. Parzych and Trojanowski 2009; Jonczak 2011; Staelens et al. 2011a; Jonczak et al. 2016). Binkley (1986) followed by Lebret et al. (2001) proved that the share of this fraction also depends on stand age, showing a negative correlation. This relationship was not confirmed in this study. The share of twigs in litterfall generally oscillates around 10–20% of its total mass (Bray and Gorham 1964; Jonczak 2011; Lebret et al. 2001; Jonczak et al. 2016), being strongly affected by the health condition of the stand and the weather, particularly wind (Staelens et al. 2011b). In this study, twigs constituted 5.7–20.8% of the total litterfall, depending on stand and year. The lowest contribution to the litterfall was from flowers.
The studied litterfall components showed relatively low variability from year to year. Temporal variability in litterfall production is mainly driven by weather conditions (Pedersen and Bille-Hansen 1999; Jonczak and Czarnecki 2008), mast years (Kowalkowski and Jóźwiak 2003; Jonczak 2011), and insect infestations (Dziadowiec and Plichta 1985).
Litterfall chemistry
Litterfall controls the biogeochemical cycling of elements in forest ecosystems (Nordén 1994; Augusto et al. 2002; Parzych and Trojanowski 2009; Staelens et al. 2011a), including their bioavailability, soil pools and forms, and profile distributions. However, its importance varies among elements and habitats, being the greatest for deficient nutrients (particularly N and P) and poor sites, such as those associated with inland dunes (Jasińska et al. 2020). The role of litterfall in the carbon cycle looks somewhat different when compared to the nutrients. First of all, the TOC content in litterfall shows low variability, constituting approximately 50% of the dry mass, regardless of plant species, organ, or season. The C constituents of litterfall cover a wide spectrum of organic structures with varied susceptibility/resistance to decomposition. Susceptible components (i.e. proteins, amino acids, monosaccharides, etc.) are a labile, easily degradable fraction of the SOM, playing an important ecological role as a source of energy for the soil microbiota (Campbell et al. 2022). Additionally, molecular characteristics strongly influence the microbial biomass, its community structure, and functions (Wu et al. 2018; Zhalnina et al. 2018). Decomposition-resistant cellulose, hemicellulose, and lignin play a key role in humus formation, an inherent process of pedogenesis (Fustec et al. 1989).
Although N generally occurs in litterfall in relatively high amounts, it is usually a deficient element in forest ecosystems––one of the major factors limiting biomass production in terrestrial biomes (Cánovas et al. 2018; Manu et al. 2022). The N content varies strongly among litterfall fractions, reflecting the effects of several factors. Generally, it occurs in its lowest amounts in strongly lignified materials, such as wood or seed coats, reaching its highest concentrations in seeds and flowers (Staelens et al. 2011a). The total and silver birch litterfall examined in this study were relatively abundant in N, reaching 12.93–17.15 g kg−1 and 12.57–16.96 g kg−1, respectively, in the two years of the study (Supplementary material, Table 4). Among the birch litter fractions, the highest concentrations were typical in the flowers, followed by seeds, and then leaves (Supplementary material). The N contents in the silver birch litterfall were comparable to those reported by Nordén (1994) for oak, hornbeam, beech, and linden, and by Berg and Meentemeyer (2002) for white birch and silver fir. They were higher compared to beech, pine, and spruce litterfall components in a mixed stand in northern Poland (Jonczak 2011) and in Scots pine, as studied by Berg and Meentemeyer (2002), Ukonmaanaho et al. (2008), and Jasińska et al. (2020). Higher N concentrations have been reported by Berg and Meentemeyer (2002) in grey alder, by Dziadowiec et al. (2007) in black poplar, and by Jonczak et al. (2016) in black alder litterfall. The N contents in the silver birch litterfall in this study were considerably higher than reported by Kasurinen et al. (2006) for young trees of the same species growing under controlled conditions (7.53–9.31 g kg−1). Saerens et al. (2011) reported 17.7–20.9 g kg−1 N in silver birch leaf litterfall, depending on the year. In this study, it was 12.79–17.98 g kg−1. It should be mentioned that the N contents in this study may have been partially affected by the post-arable character of the stands. The importance of land-use history as a factor influencing soil characteristics, including nutrient abundances, and forms has been confirmed by Pulleman et al. (2000) and Latty et al. (2004), among several other authors.
Along with N, P is considered to be limiting factor on biomass production (Elser et al. 2007). Pools and fluxes of this element in natural and semi-natural ecosystems are controlled both by the soil mineral phase (primary source) and organic matter (secondary source). Sohrt et al. (2017) reported that the entanglements between the biotic and abiotic processes that govern P cycling in temperate forests are still poorly understood, and that further and interdisciplinary studies to address this are highly recommended due to the trends observed in forests in some areas. The declining P concentrations in tree leaves in temperate forests over the last few decades (Duquesnay et al. 2000; Ilg et al. 2009) is one such trend. Fresh silver birch biomass is moderately abundant in P, with the highest contents being reported in leaves and the lowest in wood (Novák et al. 2017; Rustowska 2022). A major pool of the element is allocated in the aboveground biomass, in the branches and foliage (Ukonmaanaho et al. 2008; Gawęda et al. 2014; Novák et al. 2017). The silver birch litterfall in the studied stands contained 1.05–1.68 g kg−1 P, and leaves were the richest source of the element among the litterfall fractions, followed by flowers and seeds (Table 4, Supplementary material). This is quite surprising because the highest P concentrations are usually typical of phospholipid-rich seeds (Jonczak 2011). The abundance of the studied silver birch litterfall fell into the average to high range of values reported in the literature. It was generally higher when compared to Scots pine stands in Central and Northern Europe (Ukonmaanaho et al. 2008; Jonczak 2011; Jasińska et al. 2020) and black alder stands in northern Poland (Jonczak et al. 2016), whereas it was comparable to Norway spruce and European beech stands in northern Poland (Jonczak 2011).
The soil pools of K, Ca, and Mg mainly occur in the mineral phases, and the role of SOM is lower in this area when compared to N and P. However, the contents of these elements in litterfall are generally high in relation to other elements, regardless of differences in vegetation species and litter fractions, indicating the considerable role of vegetation in the biogeochemical cycling of these elements. The K contents noted in this study in the silver birch litterfall (2.06–4.41 g kg−1 in 2020 and 2.32–4.11 g kg−1 in 2021) were higher when compared to conifers (Ukonmaanaho et al. 2008; Jonczak 2011), while being comparable or lower than in broadleaved stands (Norden 1994; Dziadowiec et al. 2008; Jonczak 2013b; Jonczak et al. 2016). Typically, the highest contents of the element were noted in leaves, whereas twigs and flowers contained the lowest amounts (Supplementary material). Similar trends were observed for Mg and Ca, their contents in the silver birch litterfall being average for broadleaved species and higher than conifers, based on the authors cited above.
The Fe content in plant biomass rarely exceeds 0.5 g kg−1. This proportion does not reflect its much higher content in soils (usually several per cent), hence the role of vegetation in its biogeochemical cycling seems to be small. On the other hand, its importance is highlighted by certain soil-forming processes (podsolization, brunification), the essence of which is the transformation of Fe compounds, including the weathering of aluminosilicates, the formation of oxides, their linkage with other soil components, and transport. These processes are considerably controlled by vegetation (Egli et al. 2008). The Fe content in the litterfall in the studied fell within the values reported in the literature (Norden 1994; Ukonmaanaho et al. 2008; Jonczak 2011, 2013b; Jonczak et al. 2016), showing relatively wide variability between the stands and over time (Supplementary material, Table 4).
Micronutrients, including Mn, Cu, and Zn, typically occur in litterfall in low concentrations. Among these elements, Mn had the highest content in the studied stands, with the highest amounts occurring in leaves (Supplementary material). The Mn contents fell within the average values reported in the literature, being usually higher than in conifer stands (Ukonmaanaho et al. 2008), but lower than in the stands of common broadleaved species (Norden 1994). The contents of Cu and Zn in the silver birch litterfall were generally much higher than noted by other authors for various tree species (Norden 1994; Dziadowiec et al. 2008; Ukonmaanaho et al. 2008; Jonczak et al. 2016). Elevated concentrations of trace elements in the biomass can indicate environmental pollution, and silver birch has been used as a bioindicator in this field (e.g. Chiarantini et al. 2016; Birke et al. 2018). Considering the observed Cu levels (7.9–12.9 mg kg−1), the environmental contamination was estimated to be low. In the case of Zn, elevated contents were expected due to confirmed opinion about silver birch being a hyperaccumulator of this element (Marguí et al. 2007; Dmuchowski et al. 2014).
The correlation coefficients between the elemental contents in the soil and total litterfall, silver birch litterfall, and silver birch leaf litterfall were calculated as a way of looking for sources of variability in the litterfall chemistry (Table 5). No clear tendency was found based on this analysis. Although the correlations were high in some cases, they were not confirmed over the two subsequent years in the majority of cases. Also, the cluster and PCA analyses did not reveal a clear picture of tendencies. While it did indicate that litterfall chemistry was a function of many factors, none of these had a predominant role.
Retranslocation of elements from senescing leaves
Nutrient retranslocation from senescing leaves is considered to be a strategy for the effective management of deficient nutrients (e.g. Scalon et al. 2017). The intensity of this process strongly varies across forests, reflecting nutrient demand, and their bioavailability in site, and effects of other environmental factors (Richardson et al. 2005; Yuan and Chen 2010). Yuan et al. (2018) found also negative correlation between intensity of that process and tree age, whereas Kobe et al. (2005) with nutrient concentration in green leaves. This study revealed that green leaves of silver birch and fallen leaves considerably differed in terms of chemical composition; however, the observed tendencies varied depending on the element. Generally, all the studied elements, except Mn, occurred in green leaves at concentrations comparable to reported by Rustowska (2022) for stands in Central Poland. Moreover, comparable contents of N, K, Ca and Fe reported Oksanen et al. (2005). Contents of Mn recorded in this study strongly varied, hence it is difficult to compare it with other reports. Our study showed that silver birch strongly retranslocated K, N, and P. The intensity of the P retranslocation was comparable to that observed by Dziadowiec et al. (2008) in poplar stands in central Poland, but lower than in Scots pine and Norway spruce stands in Finland (Ukonmaanaho et al. 2008). Ca, Mg, Fe, Al, Mn, Cu, and Zn were usually accumulated in leaves, although the intensity of that process was not high in most cases. Only Fe was intensively accumulated.
Elements return to the soil with litterfall
The results of this study revealed that post-arable soils under birch stands are supplied with considerable amounts of nutrients via litterfall. Typically, the annual flux of N amounts to a few dozen kilos per hectare, showing great variability among forests. In this study, it fluctuated between 33.6 and 90.7 kg ha−1 (Table 8), mainly being a function of the stand and showing relatively low variability, year to year. However, the roles of tree age and site fertility were not made clear from this study. The recorded annual N fluxes were comparable to, or higher than, those reported by Nordén (1994) for oak (30.6–43.5 kg ha−1), hornbeam (32.5–52.5 kg ha−1), and beech (33.9–42.9 kg ha−1) stands; Tripathi et al. (2006) for a Betula ermanii forest in northern Japan (47 kg ha−1); Hansen et al. (2009) for Norway spruce (44 kg ha−1), Sitka spruce (36 kg ha−1), Douglas fir (35 kg ha−1), beech (39 kg ha−1), and oak (55 kg ha−1) stands; and Astel et al. (2009) for Vaccinio uliginosi–Betuletum pubescentis (36.4–46.8 kg ha−1) and Empetro nigri–Pinetum (19.2–22.1 kg ha−1) forest ecosystems in northern Poland. The N fluxes in Stands S-40 and L-41 were even comparable to those of black alder stands located on fertile soils in a headwater ecosystem, where Jonczak et al. (2016) recorded values from 70.8 to 84.2 kg ha−1.
Also, the annual fluxes of P, K, Ca, and Mg were usually greater than the typical values reported in the literature for common temperate tree species (Nordén 1994; Ukonmaanaho et al. 2008; Astel et al. 2009; Jonczak et al. 2016). The highest values of all elements were noted in the oldest stands, along with the highest litterfall production (Table 3). The annual fluxes of such elements as Fe, Cu, and Zn are usually below 1 kg ha−1 (Nordén 1994; Ukonmaanaho et al. 2008; Jonczak et al. 2016; Jasińska et al. 2020). It was also confirmed that, in this study, the recorded values were usually higher than those reported by the authors cited above. Particularly higher values were noted for Zn, highlighting once again the great ability of silver birch to accumulate this element. Although this study confirmed the great importance of litterfall as a source of soil C and nutrients in silver birch stands growing on post-arable soils, its importance can be still underestimated due to the leaching of nutrients from the litterfall accumulating in litterfall traps, as evidenced by the study by Ukonmaanaho and Starr (2001).
Conclusions
In summary, this study has significantly contributed to expanding the knowledge base on litterfall as a link in the biogeochemical cycling of the chosen elements in silver birch stands growing on post-arable soils in a temperate climatic zone. Litterfall production was found to be mainly a function of stand age, its values fluctuating between those most commonly reported in the literature for temperate forests. Typically, leaves were found to constitute the major litterfall fraction, followed by twigs. The birch litterfall was rich in several of the studied nutrients when compared to common tree species, although considerable differences were found between the stands and from year to year. In addition, the elemental contents of the litterfall did not show any clear interrelationships with their contents in the associated soils, indicating that litterfall chemistry is a function of many factors. Our study showed that silver birch intensively retranslocated N, P, and K from senescing leaves, whereas Ca, Mg, Fe, Mn, Cu, and Zn were usually accumulated. The study highlighted the importance of silver birch litterfall as a source of soil nutrients, their annual fluxes usually being higher than reported in the literature for various deciduous, coniferous, and mixed stands, thus indicating a highly intensive turnover of elements in birch stands growing on post-arable soils. This observation is in-line with previous reports on the high nutritional requirements of this tree species. In light of this study, it can be posited that a significant portion of this demand is likely met by decomposing litter. The obtained results allow the knowledge base to be expanded in the fields of forest ecology, soil science, landscape biogeochemistry, and related scientific disciplines. However, considering the diversity of birch stands, in terms of the environmental determinants of their location, further studies on the litterfall in pure birch forests in natural or semi-natural habitats are highly recommended. Comparative studies on natural and post-arable stands would contribute to the development of a more complete picture of silver birch as a component of temperate landscapes as well as birch–soil interrelationships. This work also had a more utilitarian aspect––our findings can be implemented in the development of more sustainable strategies for the afforestation of abandoned arable lands and the sustainable management of natural resources.
References
Aosaar J, Mander Ü, Varik M, Becker H, Morozov G, Maddison M, Uri V (2016) Biomass production and nitrogen balance of naturally afforested silver birch (Betula pendula Roth.) stand in Estonia. https://doi.org/10.14214/sf.1628
Astel A, Parzych A, Trojanowski J (2009) Comparison of litterfall and nutrient return in a Vaccinio uliginosi–Betuletum pubescentis and an Empetro nigri–Pinetum forest ecosystem in northern Poland. For Ecol Manag 257:2331–2341. https://doi.org/10.1016/j.foreco.2009.03.026
Augusto L, Ranger J, Binkley D, Rothe A (2002) Impact of several common tree species of European temperate forests on soil fertility. Ann for Sci 59:233–253. https://doi.org/10.1051/forest:2002020
Aussenac G, Bonneau M, Le Tacon F (1972) Restitution des éléments minéraux au sol par l’intermédiaire de la litière et des précipitations dans quatre peuplements forestiers de l’est de la France. Oecol Plant 7:1–21
Berg B, Meentemeyer V (2002) Litter quality in a north European transect versus carbon storage potential. Plant Soil 242:83–92. https://doi.org/10.1023/A:1019637807021
Berg B, Staaf H (1987) Release of nutrients from decomposing white birch and Scots pine needle litter. Pedobiologia 30:55–63
Berg B, Albrektson A, Berg MP, Cortina J, Johansson M-B, Gallardo A, Mgadeira M, Pausas J, Kiratz W, Vallejo R, McClaugherty C (1999) Amounts of litter fall in some pine forests in a European transect, in particular Scots pine. Ann for Sci 56:625–639. https://doi.org/10.1051/forest:19990801
Binkley D (1986) Forest nutrition management. Wiley, New York
Birke M, Rauch U, Hofmann F (2018) Tree bark as a bioindicator of air pollution in the city of Stassfurt, Saxony-Anhalt, Germany. J Geochem Explor 187:97–117. https://doi.org/10.1016/j.gexplo.2017.09.007
Bown D (1995) The Royal horticultural society (RHS) encyclopedia of herbs and their uses. Dorling Kindersley Publishers
Brandtberg PO, Bengtsson J, Lundkvist H (2004) Distributions of the capacity to take up nutrients by Betula spp and Picea abies in mixed stands. For Ecol Manag 198(1–3):193–208. https://doi.org/10.1016/j.foreco.2004.04.012
Bray JR, Gorham E (1964) Litter production in forests of the world. https://doi.org/10.1016/S0065-2504(08)60331-1
Browicz K (1979) Geograficzne rozmieszczenie brzóz. In: Brzozy. PWN. Warszawa-Poznań
Campbell TP, Ulrich DEM, Toyoda J, Thompson J, Munsky B, Albright MBN, Bailey VL, Tfaily MM, Dunbar J (2022) Microbial communities influence soil dissolved organic carbon concentration by altering metabolite composition. Front Microbiol 12:4345. https://doi.org/10.3389/fmicb.2021.799014
Cánovas FM, Cañas RA, de la Torre FN, Pascual MB, Castro-Rodríguez V, Avila C (2018) Nitrogen metabolism and biomass production in forest trees. Front Plant Sci 9:1449. https://doi.org/10.3389/fpls.2018.01449
Carnol M, Bazgir M (2013) Nutrient return to the forest floor through litter and throughfall under 7 forest species after conversion from Norway spruce. For Ecol Manag 309:66–75. https://doi.org/10.1016/j.foreco.2013.04.008
Chiarantini L, Rimondi V, Benvenuti M, Beutel MW, Costagliola P, Gonnelli C, Lattanzi C, Paolieri M (2016) Black pine (Pinus nigra) barks as biomonitors of airborne mercury pollution. Sci Total Environ 569:105–113. https://doi.org/10.1016/j.scitotenv.2016.06.029
Daugaviete M, Krŭmina M, Kǎpopstos V, Lazdinš A (2003) Farmland afforestation: the plantations of birch Betula pendula Roth. on different soils. Baltic for 9:9–21
Daugaviete M, Korica AM, Silins I, Barsevskis A, Bardulis A, Bardule A, Spalvis K, Daugavietis M (2015) The use of mineral nutrients for biomass production by young birch stands and stands vitality in different forest growing conditions. J Environ Sci Eng 4:177–189. https://doi.org/10.17265/2162-5263/2015.04.002
Dmuchowski W, Gazdowski D, Brągoszewska P, Baczewska AH, Suwara I (2014) Phytoremediation of zinc contaminated soils using silver birch (Betula pendula Roth). Ecol Eng 71:32–35. https://doi.org/10.1016/j.ecoleng.2014.07.053
Duquesnay A, Dupouey J, Clement A, Ulrich E, Le Tacon F (2000) Spatial and temporal variability of foliar mineral concentration in beech (Fagus sylvatica) stands in northeastern France. Tree Physiol 20:13–22. https://doi.org/10.1093/treephys/20.1.13
Dziadowiec H, Plichta W (1985) The effect of nun month (Lymantria monacha L.) outbreak on characteristics of litterfall in the pine forest. Ekol Pol 33:715–728
Dziadowiec H, Jonczak J, Czarnecki A, Kacprowicz K (2007) Masa, dynamika i skład chemiczny opadu roślinnego w różnowiekowych plantacjach odmiany uprawnej topoli czarnej Hybryda 275. Rocz Glebozn 58(3/4):68–77
Dziadowiec H, Jonczak J, Czarnecki A, Kacprowicz K (2008) Comparision of plant litterfall in two poplar plantations of Hybryda 275 and Robusta. Soil Sci Ann 59:76–84
Egli M, Mirabella A, Sartori G (2008) The role of climate and vegetation in weathering and clay mineral formation in late Quaternary soils of the Swiss and Italian Alps. Geomorphology 102(3–4):307–324. https://doi.org/10.1016/j.geomorph.2008.04.001
Elser JJ, Bracken ME, Cleland EE, Gruner DS, Harpole WS, Hillebrand H, Ngai JT, Seabloom EW, Shurin JB, Smith JE (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett 10:1135–1142. https://doi.org/10.1111/j.1461-0248.2007.01113.x
FAO (2006) Guidelines for Soil Description. FAO, Rome
Ferm A (1993) Birch production and utilization for energy. Biomass Bioenerg 4:391–404. https://doi.org/10.1016/0961-9534(93)90061-8
Franiel I, Kompała-Bąba A (2021) Reproduction strategies of the silver birch (Betula pendula Roth) at post-industrial sites. Sci Rep 11:11969. https://doi.org/10.1038/s41598-021-91383-0
Fustec E, Chauvet E, Gas G (1989) Lignin degradation and humus formation in alluvial soils and sediments. Appl Environ Microbiol 55:922–926. https://doi.org/10.1128/aem.55.4.922-926.1989
Gawęda T, Małek S, Zasada M, Jagodziński AM (2014) Allocation of elements in a chronosequence of silver birch afforested on former agricultural lands. Drewno 57:107–117
Hansen K, Vesterdal L, Schmidt KI, Gundersen P, Sevel L, Bastrup-Birk A, Pedersen LB, Bille-Hansen J (2009) Litterfall and nutrient return in five tree species in a common garden experiment. For Ecol Manag 257:2133–2144. https://doi.org/10.1016/j.foreco.2009.02.021
Hynynen J, Niemistö P, Viherä-Aarnio A, Brunner A, Hein S, Velling P (2010) Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 83:103–119. https://doi.org/10.1093/forestry/cpp035
Ilg K, Wellbrock N, Lux W (2009) Phosphorus supply and cycling at long-term forest monitoring sites in Germany. Eur J for Res 128:483–492. https://doi.org/10.1007/s10342-009-0297-z
Dell Inc. (2016) Dell statistica data analysis software system. Version 13
IUSS Working Group WRB (2015) World reference base for soil resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. FAO, Rome
Jasińska J, Sewerniak P, Puchałka R (2020) Litterfall in a Scots pine forest on inland dunes in Central Europe: mass, seasonal dynamics and chemistry. Forests 11:678. https://doi.org/10.3390/f11060678
Johansson MB (1995) The chemical composition of needle and leaf litter from Scots pine, Norway spruce and white birch in Scandinavian forests. For Int J for Res 68:49–62. https://doi.org/10.1093/forestry/68.1.49
Jonczak J (2011) Structure, dynamics and properties of litterfall in a 110-year-old beech stand with admixture of pine and spruce. Sylwan 155:760–768
Jonczak J (2013a) Soil organic matter properties in Stagnic Luvisols under different land use types. Acta Agrophys 20:565–576
Jonczak J (2013b) Dynamics, structure and properties of plant litterfall in a 120-year old beech stand in Middle Pomerania between 2007–2010. Soil Sci Ann 64:8–13
Jonczak J, Czarnecki A (2008) Risk assessment for biomass plantations planning on marginal soils as an effect of increasing frequency of weather extreme events. Ecol Quest 9:113–118
Jonczak J, Olejniczak M, Parzych A, Sobisz Z (2016) Dynamics, structure and chemistry of litterfall in headwater riparian forest on the area of Middle Pomerania. J Elementol. https://doi.org/10.5601/jelem.2015.20.3.938
Jonczak J, Jankiewicz U, Kondras M, Kruczkowska B, Oktaba L, Oktaba J, Olejniczak I, Pawłowicz E, Polláková N, Raab T, Regulska E, Słowińska S, Sut-Lohmann M (2020) The influence of birch trees (Betula spp) on soil environment—a review. For Ecol Manag 477:118486. https://doi.org/10.1016/j.foreco.2020.118486
Kasurinen A, Riikonen J, Oksanen E, Vapaavuori E, Holopainen T (2006) Chemical composition and decomposition of silver birch leaf litter produced under elevated CO2 and O3. Plant Soil 282:261–280. https://doi.org/10.1007/s11104-005-6026-6
Kobe RK, Lepczyk CA, Iyer M (2005) Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology 86:2780–2792. https://doi.org/10.1890/04-1830
Kowalkowski A, Jóźwiak M (2003) Dynamika masy opadu organicznego w latach 1994–2002 w dwóch drzewostanach górskiej kwaoenej buczyny na głównym masywie Łysogór, Regionalny Monitoring OErodowiska Przyrodniczego 4, Kieleckie Towarzystwo Naukowe Kielce, pp 79–98
Latty EF, Canham CD, Marks PL (2004) The effects of land-use history on soil properties and nutrient dynamics in northern hardwood forests of the Adirondack Mountains. Ecosystems 7:193–207. https://doi.org/10.1007/s10021-003-0157-5
Lebret M, Nys C, Forgeard F (2001) Litter production in an Atlantic beech (Fagus sylvatica L.) time sequence. Ann for Sci 58:755–768. https://doi.org/10.1051/forest:2001161
Liu C, Westman CJ, Berg B, Kutsch W, Wang GZ, Man R, Ilvesniemi H (2004) Variation in litterfall-climate relationships between coniferous and broadleaf forests in Eurasia. Glob Ecol Biogeogr 13:105–114. https://doi.org/10.1111/j.1466-882X.2004.00072.x
Loydi A, Lohse K, Otte A, Donath TW, Eckstein RL (2014) Distribution and effects of tree leaf litter on vegetation composition and biomass in a forest–grassland ecotone. J Plant Ecol 7:264–275. https://doi.org/10.1093/jpe/rtt027
Manu R, Corre MD, Aleeje A, Mwanjalolo MJG, Babweteera F, Veldkamp F, van Straaten O (2022) Responses of tree growth and biomass production to nutrient addition in a semi-deciduous tropical forest in Africa. Ecology 103:e3659. https://doi.org/10.1002/ecy.3659
Marguí E, Queralt I, Carvalho ML, Hidalgo M (2007) Assessment of metal availability to vegetation (Betula pendula) in Pb-Zn ore concentrate residues with different features. Environ Pollut 145:179–184. https://doi.org/10.1016/j.envpol.2006.03.028
Matthews E (1997) Global litter production, pools, and turnover times: estimates from measurement data and regression models. J Geophys Res 102(D15):18771–18800
Mikola J, Silfver T, Paaso U, Possen BJMH, Rousi M (2018) Leaf N resorption efficiency and litter N mineralization rate have a genotypic tradeoff in a silver birch population. Ecology 99:1227–1235. https://doi.org/10.1002/ecy.2176
Miller HG (1984) Nutrient cycles in birch woods. Proc R Soc Edinb 853:83–96
Mork E (1942) Om strofallet i vare skoger. Medd Fr Det Norske Skogforsoksvesen 29:297–357
Neumann M, Ukonmaanaho L, Johnson J, Benham S, Vesterdal L, Novotný R, Verstraeten A, Lundin L, Thimonier A, Michopoulos P, Hasenauer H (2018) Quantifying carbon and nutrient input from litterfall in European forests using field observations and modeling. Glob Biogeochem Cycles 32:784–798. https://doi.org/10.1029/2017GB005825
Nordén U (1994) Leaf litterfall concentrations and fluxes of elements in deciduous tree species. Scand J for Res 9(1–4):9–16. https://doi.org/10.1080/02827589409382807
Novák J, Dušek D, Kacálek D, Slodičák M (2017) Nutrient content in silver birch biomass on nutrient-poor, gleyic sites. Zpravy Lesnickeho Vyzkumu 62:135–141
Oikonomakis NG, Ganatsas P (2020) Secondary forest succession in Silver birch (Betula pendula Roth) and Scots pine (Pinus sylvestris L.) southern limits in Europe, in a site of Natura 2000 network—an ecogeographical approach. For Syst 29:81–96. https://doi.org/10.5424/fs/2020292-15680
Oksanen E (2021) Birch as model system for acclimation and adaptation of northern forest ecosystem to changing environment. Front for Glob Change 4:682512. https://doi.org/10.3389/ffgc.2021.682512
Oksanen E, Riikonen J, Kaakinen S, Holopainen T, Vapaavuori E (2005) Structural characteristics and chemical composition of birch (Betula pendula) leaves are modified by increasing CO2 and ozone. Glob Change Biol 11:732–748. https://doi.org/10.1111/j.1365-2486.2005.00938.x
Osono T, Takeda H (2004) Accumulation and release of nitrogen and phosphorus in relation to lignin decomposition in leaf litter of 14 tree species. Ecol Res 19:593–602. https://doi.org/10.1111/j.1440-1703.2004.00675.x
Ostonen I, Lõhmus K, Helmisaari H-S, Truu J, Meel S (2007) Fine root morphological adaptations in Scots pine, Norway spruce and silver birch along a latitudinal gradient in boreal forests. Tree Physiol 27:1627–1634. https://doi.org/10.1093/treephys/27.11.1627
Ovington JD, Madgwick HAI (1959) The growth and composition of natural stands of birch. 2. The uptake of mineral nutrients. Plant Soil 10:389–400. https://doi.org/10.1007/BF01666212
Parzych A, Trojanowski J (2009) The structure and dynamics of litterfall in forest stands in the Słowiński National Park in 2003–2005. For Res Papers 70:41–48
Pedersen LB, Bille-Hansen J (1999) A comparison of litterfall and element fluxes in even aged Norway spruce, Sitka spruce and beech stands in Denmark. For Ecol Manag 114:55–70. https://doi.org/10.1016/S0378-1127(98)00381-8
Peel MC, Finlayson BL, McMahon TA (2007) Updated world map of the Köppen–Geiger climate classification. Hydrol Earth Syst Sci 11:1633–1644. https://doi.org/10.1127/0941-2948/2006/0130
Perala DA, Alm AA (1990) Reproductive ecology of birch: a review. For Ecol Manag 32:1–38. https://doi.org/10.1016/0378-1127(90)90104-J
Possen BJHM, Rousi M, Keski-Saari S, Silfver T, Kontunen-Soppela S, Oksanen E, Mikola J (2021) New evidence for the importance of soil nitrogen on the survival and adaptation of silver birch to climate warming. Ecosphere 12:e03520. https://doi.org/10.1002/ecs2.3520
Pulleman MM, Bouma J, van Essen EA, Meijles EW (2000) Soil organic matter content as a function of different land use history. Soil Sci Soc Am J 64:689–693
Richardson SJ, Peltzer DA, Allen RB, McGlone MS (2005) Resorption proficiency along a chronosequence: responses among communities and within species. Ecology 86:20–25. https://doi.org/10.1890/04-0524
Rosenvald K, Ostonen I, Truu M, Truu J, Uri V, Vares A, Lõhmus K (2011) Fine-root rhizosphere and morphological adaptations to site conditions in interaction with tree mineral nutrition in young silver birch (Betula pendula Roth.) stands. Eur J for Res 130:1055–1066. https://doi.org/10.1007/s10342-011-0492-6
Rustowska B (2022) Long-term wildfire effect on nutrient distribution in silver birch (Betula pendula Roth) biomass. Soil Sci Annu 73:149943. https://doi.org/10.37501/soilsa/149943
Scalon MC, Wright IJ, Franco AC (2017) To recycle or steal? Nutrient resorption in Australian and Brazilian mistletoes from three lowphosphorus sites. Oikos 126:32–39. https://doi.org/10.1111/oik.03455
Shen G, Chen D, Wu Y, Liu L, Liu C (2019) Spatial patterns and estimates of global forest litterfall. Ecosphere 10:e02587. https://doi.org/10.1002/ecs2.2587
Silfver T, Mikola J, Rousi M, Roininen H, Oksanen E (2007) Leaf litter decomposition differs among genotypes in a local Betula pendula population. Oecologia 152:707–714. https://doi.org/10.1007/s00442-007-0695-1
Silfver T, Kontro M, Paaso U, Karvinen H, Keski-Saari S, Keinanen M, Rousi M, Mikola J (2018) Intrapopulation genotypic variation in leaf litter chemistry does not control microbial abundance and litter mass loss in silver birch, Betula pendula. Plant Soil 426(2018):253–266. https://doi.org/10.1007/s11104-018-3631-8
Sohrt J, Lang F, Weiler M (2017) Quantifying components of the phosphorus cycle in temperate forests. Wiley Interdiscip Rev Water 4:e1243. https://doi.org/10.1002/wat2.1243
Špulák O, Souček J, Bartoš J, Kacálek D (2010) Potential of young stands with birch dominance established by succession on abandoned agricultural land. For Res Rep 55:165–170
Staelens J, Nachtergale J, De Schrijver A, Vanhellemont M, Wuyts K, Verheyen K (2011a) Spatio-temporal litterfall dynamics in a 60-year-old mixed deciduous forest. Ann for Sci 68(2011):89–98. https://doi.org/10.1007/s13595-011-0010-5
Staelens J, Nachtergale L, Luyssaert S, Lust N (2011b) A model of wind-influenced leaf litterfall in a mixed hardwood forest. Can J for Res 33:201–209. https://doi.org/10.1139/x02-174
Titmuss FH (1959) A concise encyclopedia of wood timbers. The Technical Press Ltd., London
Tripathi SK, Sumida A, Shibata H, Ono K, Uemura S, Kodama Y, Hara T (2006) Leaf litterfall and decomposition of different above- and belowground parts of birch (Betula ermanii) trees and dwarf bamboo (Sasa kurilensis) shrubs in a young secondary forest in Northern Japan. Biol Fertil Soils 43:237–246. https://doi.org/10.1007/s00374-006-0100-y
Ukonmaanaho L, Starr M (2001) The importance of leaching from litter collected in litterfall traps. Environ Monit Assess 66(2001):129–146. https://doi.org/10.1023/A:1026432315707
Ukonmaanaho L, Merillä P, Nöjd P, Nieminen TM (2008) Litterfall production and nutrient return to the forest floor in Scots pine and Norway spruce stands in Finland. Boreal Environ Res 13(suppl. B):67–91
Ulbrichová I, Podrázský V, Slodičák M (2005) Soil forming role of birch in the ore mts. J for Sci 51:54–58. https://doi.org/10.17221/11850-JFS
Úradníček L, Maděra P, Tichá S, Koblížek J (2009) Dřeviny České Republiky (Woody plants of Czech Republic), 2nd edn. Lesnická práce, Kostelec nad Černými lesy (in Czech)
Uri V, Varik M, Aosaar J, Kanal A, Kukumägi M, Lõhmus K (2012) Biomass production and carbon sequestration in a fertile silver birch forest chronosequence. For Ecol Manag 267:117–126. https://doi.org/10.1016/j.foreco.2011.11.033
Wu X, Wu L, Liu Y, Zhang P, Li Q, Zhou J, Hess NJ, Hazen TC, Yang W, Chakraborty R (2018) Microbial interactions with dissolved organic matter drive carbon dynamics and community succession. Front Microbiol 9:1234. https://doi.org/10.3389/fmicb.2018.01234
Yuan ZY, Chen HYH (2010) Changes in nitrogen resorption of trembling aspen (Populus tremuloides) with stand development. Plant Soil 327:121–129. https://doi.org/10.1007/s11104-009-0036-8
Yuan Z, Shi X, Jiao F, Han F (2018) N and P resorption as functions of the needle age class in two conifer trees. J Plant Ecol 11:780–788. https://doi.org/10.1093/jpe/rtx055
Zasada M, Bijak S, Bronisz K, Bronisz A, Gawęda T (2014) Biomass dynamics in young silver birch stands on post-agricultural lands in central Poland. Drewno Prace Naukowe, Doniesienia, Komunikaty. https://doi.org/10.12841/wood.1644-3985.S07.02
Zhalnina K, Louie KB, Hao Z, Mansoori N, da Rocha UN, Shi S, Cho H, Karaoz U, Loqué D, Bowen BP, Firestone MK, Northen TR, Brodie EL (2018) Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat Microbiol 3:470–480. https://doi.org/10.1038/s41564-018-0129-3
Funding
The research was supported by the National Science Centre, Poland, Project No. 2018/31/B/NZ9/01994, title “Transformation of post-arable soils due to afforestation with birch”.
Author information
Authors and Affiliations
Contributions
JJ contributed to planning and general management of the project, laboratory analyses, concept of the manuscript, and writing manuscript. LO contributed to field studies and writing of the manuscript. AC contributed to field studies and writing of the manuscript. EP contributed to determination of basic soil properties. BK contributed to laboratory analysis and writing of the manuscript. JO contributed to stands characterization. SS contributed to characterization of the study area and writing of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper. The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Jerzy Jonczak reports financial support was provided by National Science Centre Poland.
Additional information
Communicated by Agustin Merino.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jonczak, J., Oktaba, L., Chojnacka, A. et al. Nutrient fluxes via litterfall in silver birch (Betula pendula Roth) stands growing on post-arable soils. Eur J Forest Res 142, 981–996 (2023). https://doi.org/10.1007/s10342-023-01569-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-023-01569-6