
Abstract
Background
As compared to natural forest ecosystems, soils under agroecosystems are more subjected to changes in physical and biochemical attributes. But, tree species that are retained and/or planted on these agroecosystems replenish soil fertility through the influence of leaves litterfall quantity and quality. Tree species-related differences in the quantity and seasonality of leaves litterfall and the associated amounts of annual returns of nutrient elements were assessed from four multipurpose tree/shrub species (Erythrina brucei Schweinf, Cordia africana Lam., Grevillea robusta R. Br. and Malus sylvestris Mill.) in homegarden agroforestry systems. Five replicates of two leaf litterfall traps per species were randomly positioned beneath the subcanopy area for 12 months, totaling 40 litterfall traps distributed among the four species.
Results
The results of this study suggested that total annual leaves litterfall varied significantly among species (ranging from 171.18 to 962 kg ha− 1), with the combined per tree mean monthly (28.13 kg tree− 1), per tree annual (331.10 kg tree− 1 year− 1) and per area basis total annual (9439.33 kg ha− 1) leaves litterfall production. A clear seasonal leaves litterfall pattern with varying peaks was observed for the multipurpose tree/shrub species in the present study, with a bimodal distribution pattern for E. brucei and C. africana and a unimodal distribution pattern for G. robusta and M. sylvestris. Similarly, both the concentrations and inputs of nutrients from the leaves᾽ litterfall showed considerable variations among species (p < 0.05). Among the investigated species, C. africana and E. brucei were found to be the best in terms of carbon (C) and nitrogen (N) return, G. robusta the best in terms of phosphorous (P) return, and C. africana as well best for potassium (K) return. The combined mean annual nutrient inputs recorded for all the species were 3125.32 kg C ha− 1 year− 1, 149.70 kg N ha− 1 year− 1, 54.86 kg K ha− 1 year− 1, and 3.56 kg P ha− 1 year− 1.
Conclusions
Leaves litterfall amount differed among tree species as influenced by tree species characteristics (canopy area, diameter at breast height (DBH), height), but no cause-effect relationship with age and climate (monthly precipitation and temperature).
Similar content being viewed by others
Introduction
Homegarden agroforestry systems are increasingly extended parcels of land surrounding homesteads in which diverse mixtures of plant species are set up in multi-layered configurations, often integrating annual and perennial agricultural crops with multipurpose trees and/or shrubs, and livestock components (Fernandes and Nair 1986; Nair 1993; Abebe et al. 2006; Kehlenbeck et al. 2007; Castro et al. 2018). They are tree-based land use systems that stood the test of time and are more efficient in maintaining soil fertility than annual cropping systems (Schroth et al. 2001; Kumar and Nair 2004). Tropical agroforestry systems, including homegardens, are managed under different ecological and cultural settings where the stocking levels and soil fertility contribution of most indigenous farm trees are limited to farmers᾽ knowledge (Asfaw 2007; Teklay 2007; Yadav et al. 2008; Triadiati et al. 2011).
On account of understanding the ecological rationalities behind the harmony between humans and the homegardens environment (Gajaseni and Gajaseni 1999), it appears crucial to consider the amount and pattern of litterfall, and the associated nutrient fluxes and the implication it had on soil organic matter formation and biogeochemical cycles (Triadiati et al. 2011; Negash and Starr 2021; Wu et al. 2021). However, the scientific selection of farm tree species that possessed efficient nutrient return capabilities remains to be a challenge ahead in these agroecosystems (Hossain et al. 2011; Verma et al. 2021).
Prior experiences did reflect that farm household traditionally improve their homegardens soil fertility by applying litter or green leaves, animal manure, and household refuse (Fernandes and Nair 1986; Kumar and Nair 2004; Kehlenbeck and Mass 2004; Kehlenbeck 2007; Wiehle et al. 2014; Abebe and Bongers 2012) or through removing detritus from the plantation or forest floor for use in their homegardens or field (Kumar 2008). Among those extensively retained and/or planted agroforestry tree species, the combined effect of selected species annual litter inputs and nutrient additions in the homegardens were reported in different parts of the world (Isaac and Nair 2006; Yadav et al. 2008; Das and Das 2010; Hossain et al. 2011), including Ethiopia (Negash and Starr 2013). Some of the most commonly investigated multipurpose tree species by different workers under an agroforestry setting include Mangifera indica L. (Isaac and Nair 2006; Das and Das 2010; Hasanuzzaman and Hossain 2014; Negash and Starr 2021); Artocarpus spp., Ailanthus spp. (Jamaludheen and Kumar 1999; Isaac and Nair 2006); Dalbergia sissoo Roxb., Melia azedarach L. (Isaac and Nair 2006; Hossain et al. 2011; Hasanuzzaman and Hossain 2014), cacao and shade trees (Sari et al. 2022; Dawoe et al. 2010; Asigbaase et al. 2021); and coffee and shade trees (Hairiah et al. 2006; Negash and Starr 2013; López-Rodríguez et al. 2015). In this paper, we focused on four agroforestry tree species of E. brucei, C. africana, G. robusta, and M. sylvestris because of farmers᾽ selection criteria and their dominance in the respective homegarden agroforestry systems. Because there is little generalization at the national or continental level on the effects of litterfall quantity and quality in different tree species and their contribution to the biogeochemical cycles (Hansen et al. 2009).
Litterfall plays a fundamental role in better understanding the tree-soil relationship (Andivia et al. 2010), and the long-term functioning and appropriate management of modified ecosystems (Kunhamu et al. 2009; Zhou et al. 2016). However, quantitative litterfall data as a key parameter in the biogeochemical processes in tropical homegardens are inadequate (Isaac and Nair 2006; Das and Das 2010; Negash and Starr 2013). And, the quantity and quality of litterfall can be influenced by factors such as species composition, density, size and age, climatic and edaphic conditions, season, and management practices (Ashagrie and Zech 2013; Negash and Starr 2013; Negash and Starr 2022). Furthermore, Lebret et al. (2001) demonstrated that age and basal area were closely related to quantities of litterfall in an Atlantic beech. Although annual litterfall rates increase as crown coverage increases (Kumar 2008), to the contrary, no proper relation between the leaf litter amount and DBH or height of the tree was observed by Yang et al. (2021).
It remains unclear how agricultural land use affects nutrient balances and their interrelation with litter quantity, quality, and element cycles in tropical agroecosystems (Becker et al. 2015). Similarly, earlier investigations on the characteristics of litterfall nutrient release, and seasonal dynamics set their primary focus on a few non-native plantation species while paying less attention to those used in traditional farming systems (Teklay 2007). Furthermore, there has also been little assessment of litterfall production as related to tree characteristics (crown area, DBH, height, age) and climate variables (temperature and rainfall) in tropical agroforestry systems (Isaac and Nair 2006; Dawoe et al. 2010; Paudel et al. 2015; Negash and Starr 2021). Therefore, the objectives of this study were to investigate the following three aspects; (1) to analyze tree species-related differences in the quantity and seasonality of leaves litterfall; (2) to quantify the concentration and amounts of annual returns of nutrient elements from leaves litterfall of four multipurpose trees/shrubs species; and, (3) to determine if tree species characteristics (canopy area, DBH, height, age), and climate variables (monthly precipitation and temperature) correlate with leaves litterfall.
Materials and methods
Study areas
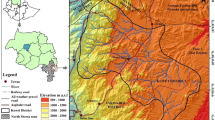
The field research was conducted in the Gamo zone (Chencha woreda), and Yem special woreda, both located in southern and southwestern parts of the country within the Southern Nation, Nationalities, and Peoples᾽ Region (SNNPR) (Fig. 1). All two studies woredas and sites were purposefully selected based on their potential suitability for the study objectives, predominance of enset-based homegarden agroforestry systems, agroclimatic variations, and remoteness and nearness to major roads/markets (Table 1).

A map showing the study sites
Chencha (6°05’ N latitude, 37°30’ E longitude; Coltorti et al. 2019) woreda is confined within the Western Ethiopian Escarpment (WEE) part of the main Ethiopian Rift system [MER] (Wana and Woldu 2005; Abbate et al. 2015), while that of Yem special woreda (7°37’N and 37°40’ E; Kassa et al. 2015) is located in the southern Ethiopian plateau (Abbate et al. 2015), all in southern and southwestern Ethiopia. Chencha is located about ca. 37 km north of Arba Minch, at an elevation ranging from 2000 to 3000 m above sea level (m a.s.l.) (Mazengia et al. 2016). While that Yem special woreda is located ca. 242 km southwest of Addis Ababa, on the Addis Ababa-Jimma tarmac highway, with elevations ranging from 920 to 2939 m a.s.l.
Climatic and soil condition
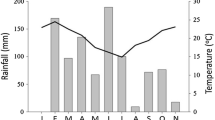
Based on 10 years of data (2007–2016) obtained from the National Meteorological Service Agency (NMSA), the climate diagram of Chencha and Saja Laften (represented by Sokoru the nearest recording station) woredas (Figs. 2 and 3) was computed using R version 4.0.1 (R Development Core Team, 2020). The diagram depicts that Chencha woreda receives a bimodal rainfall pattern from the beginning of mid-March to May and September to October. While a unimodal rainfall pattern extending from April to October was recorded for that of Saja Laften, with minimum rainfall from December to February. The mean annual minimum and maximum temperature records for Chencha range from 9.5–21.6 °C, and 12.3–29.3 °C for Saja Laften. And, the mean annual rainfall recorded for Chencha, and Saja Laften were 1263 and 1365 mm, respectively. The major soil types observed in the Chencha area include Immature Cambisols, Nitisols, Luvisols, Leptosols, Cambisols, and Andosols (Berhanu et al. 2013; Coltorti et al. 2019), while that of Yem include Nitisols, Acrisols, Ferralsols, Vertisols, and Planosols (van Ranst et al. 2011).

(Source: NMSA)
Climate diagram showing the mean monthly rainfall and temperature distribution records of the study area taking 10 years (2007–2016)

(Source: Unpublished Ethiopian National Meteorological Services Agency NMSA)
Climate diagram showing the mean monthly rainfall and temperature distribution records of Saja Laften taking 10 years (2007–2016)
Based on the 2007 census and the 2.9% rate of population growth for SNNPR, in 2021 the human population density of Chencha, and Yem are estimated at 445.57, and 166.1 persons/km2, respectively (FDRE PCC 2008; Debebe and Ayele 2013). Taking into account the method developed by Gerhardt and Hytteborn (1992) and adopted by Bekele (1993), the native vegetations of Chencha are characterized by dry Afromontane forests in the highlands where there appeared to be serious deforestation, with extensive areas being changed into bushlands (Friis et al. 2010; Samberg et al. 2010; Demissew and Nordal 2010). Belonging to the Afromontane vegetation of certain high mountains, the area has been identified as part of the Somalia-Masai Regional Centre of Endemism (White 1983). The ubiquitous existence of individual forest trees such as Juniperus procera Hochst. ex Endl., Olea europaea subsp. cuspidata (Wall. ex G.Don) Cif., and Croton macrostachyus Del., a scattered relict, and few remnant forest patches in most highlands of Yem are indicative of a long history of forest exploitation (Bekele 1993; Senbeta and Denich 2006), and deforestation for thousands of years (Bekele 1994) which can be outcomes of sociocultural activities of the local people (Senbeta and Denich 2006). In the lowlands of Yem, the part of the basin where the Ghibe-Omo lies, wooded grasslands, and bushland of Somalia-Masai type is found (Friis 1992; Bekele 1993).
Leaves litter production
Individual trees and shrubs that were growing inside homegarden agroforestry but isolated from the canopy of other trees were randomly chosen to quantify leaves litterfall deposited. The homegardens were chosen because they are the most important agroforestry systems that support the livelihoods of a large majority of rural society in the southern and southwestern Ethiopia (Abebe 2013; Mellisse et al. 2018). The main reason that aroused our interest to focus on leaves was that chlorophyll-rich parts (leaves) are the essential components and quick sources of organic matter and nutrients to the soil compared to other parts of litter (Lebret et al. 2001; Hasanuzzaman and Hossain 2014; Lalremsang et al. 2022). Selection of the most important and commonly found tree/shrub species for the leaves litterfall study was determined based on farmers᾽ understanding and consensus in their locality using a pairwise ranking technique (Appendix Table 6 and Table 7). Accordingly, the four multipurpose tree/shrub species nominated in the present study were Erythrina brucei Schweinf., Malus sylvestris Mill., Grevillea robusta R.Br., and Cordia africana Lam. E. brucei and C. africana are indigenous popular upper storey trees that have been retained and planted particularly with enset and coffee plants in multistrata agroforestry systems (Asfaw 2007; Negash et al. 2012). M. sylvestris and G. robusta are the most common exotic fast-growing trees grown in association with crops on farmlands or homegardens for fruits and wood production, and could serve as important sources of income (Takaoka 2008; Currey 2009; Kuyah et al. 2020). Leaves litterfall samples were collected from these four selected tree/shrub species under subcanopy sampling area at monthly intervals for 12 months using litter traps (each 1 × 1 m2 wide). The litter traps made up of 1 mm nylon mesh size were mounted on wooden-framed open boxes (Singh et al. 1999). Each trap was raised 15 cm above the ground but modified following Dawoe et al. (2010) and Yang et al. (2004) to avoid any contact with the ground soil. Five replicates of two leaf litterfall traps per species were randomly positioned beneath subcanopy area, totaling 40 litter traps distributed among the four species. The litter traps were placed in opposite directions (north–south and east–west) under the subcanopy area to capture varying wind effects and maintain uniformity on the quantity of leaves litterfall being collected per each sample tree species (Schroth 2003, cited in Dawoe et al. 2010). Some parameters that were recorded before the placement of litter traps include diameter at breast height (DBH), crown area (CA), total height (H), and age (Yadav et al. 2008; Negash and Starr 2013). These characteristics of tree/shrub species selected for leaves litterfall production study in the homegarden agroforestry systems are given in Table 2.
Leaves litterfall sample preparation and chemical analysis
After collection, monthly leaves litterfall from each sample trap was composited by species for each replicated individual to create one sub-sample and was oven-dried at 105 °C for 24 h to constant weights (Andivia et al. 2010; Tangjang et al. 2015). Leaves litterfall was calculated on the unit area of crown (g m− 2) for each month and species, and monthly leaves litterfall data were summed to obtain the annual leaves litterfall yield of each species. Annual leaves litterfall deposition was calculated as the average from traps over one year period. The crown area (ca) in m2 was calculated assuming an elliptical crown shape using the formula ca = π(l/2) × (w/2), where l is the crown diameter at its widest point and w is the perpendicular crown extent at the same height (Kuyah et al. 2012).
The oven-dried leaf litterfall samples were finally subjected to chemical analysis to assess the nutrient contents. Accordingly, a chemical analysis of four sub-samples of nutrients (organic carbon [OC], nitrogen [N], phosphorus [P], and potassium [K]) from leaf litterfalls for each species was undertaken. Organic matter content was determined as loss-on-ignition (ignition at 550 °C for 2 h) (Anderson and Ingram 1993) and values were converted to a C content by considering 50% C in ash-free litter material (Schlesinger 1977; Gallardo and Merino 1993). The N and P contents were analyzed according to the micro-Kjeldahl method. Potassium was also determined by atomic absorption spectrophotometry as outlined by Jackson (1958). Mean annual nutrient input through leaf litterfall by different species was estimated by multiplying the mean annual values of each leaves litterfall mass by its corresponding mean nutrient concentrations (Yang et al. 2004, 2005; Ukonmaanaho et al. 2008; Dawoe et al. 2010).
Statistical analyses
Using Statistical Package for Social Sciences (SPSS, Version 16) and R software, data were subjected to one-way analysis of variance (ANOVA) to analyze variation in monthly and annual leaves litterfall production, and annual leaves litterfall nutrient returns from each species. The relationships between leaves litterfall amounts and tree characteristics (crown area, dbh, height, and age), and climatic variables (such as monthly precipitation and temperature) were carried out using the curve estimation method. Assumptions of normal distribution and homogeneity of variances were checked by using Kolmogorov–Smirnov and Levene᾽s tests (Field 2009). Appropriate post hoc pairwise comparisons were made with Fisher’s Least Significance Difference (LSD) tests at α = 0.05 to isolate group means that show significant differences. Where necessary, data were log-transformed to meet assumptions of normality and homogeneity of variance.
Results
Leaves litterfall input from multipurpose tree/shrub species in the homegarden agroforestry systems
In the studied homegarden agroforestry systems, the combined annual litterfall production for all the species investigated was 9439.33 kg ha− 1. There were significant differences in mean annual leaf litterfall production between species per individual tree (F3, 19 = 6.91; p = 0.003) and total per hectare basis (F3, 19 = 8.842; p = 0.001) (Table 3). Leaves litterfall contributed from C. africana (23.96 kg tree− 1), G. robusta (17.43 kg tree− 1), and E. brucei (23.91 kg tree–1) showed higher mean values than that of M. sylvestris (0.91 kg tree− 1). On an area basis, a comparison of annual leaves litterfall indicated that C. africana (962.00 kg ha− 1) had the highest mean value, varying between 504.32 to 1496.37 kg ha− 1 (Table 3). The sequence of the relative total annual leaves litterfall contribution of multipurpose tree/shrub species with higher mean values was C. africana > E. brucei > G. robusta > M. sylvestris.
Seasonal variation of leaves litterfall
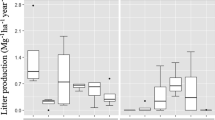
Monthly leaves litterfall values also differed significantly between species (X2 = 27.062, df = 3, p = 0.000), where it varied widely from 9.99 kg tree− 1 in C. africana to a distinctly lower 0.38 kg tree–1 in M. sylvestris (Table 3). In general, the monthly leaf litterfall followed a bimodal distribution pattern for E. brucei and C. africana and a unimodal pattern for G. robusta and M. sylvestris, each with varying peaks (Fig. 4). The quantity of leaf litterfall mass-produced was highest in September to October for most species, and the least coincided with the months between April to May, except for E. brucei litterfall.

Monthly variations in leaves litterfall for tree/shrub species in homegarden agroforestry systems in southern and southwestern Ethiopia. Symbols represent: “● (filled circle)”—E. brucei; “□ (opened squares)”– C. africana; “∆ (opened triangle)”– G. robusta; and “○ (opened circle)”– M. sylvestris
Leaves litterfall nutrient concentrations and returns
Leaves litterfall macronutrient concentrations did show significant differences between species (F3, 19 = 8.842; p = 0.001), especially for C and N, but for P and K it proved to be non-significant (Table 4). C concentration of leaves litterfall for C. africana (42.65%), varying between 41.70 to 43.75%, was significantly lower than that of the other three tree/shrub species. On the whole, C concentration decreased in the order G. robusta > E. brucei > M. sylvestris > C. africana. Leaves litterfall of E. brucei (3.32%) had significantly higher N concentration compared with the other three tree/shrub species, and its overall concentration declined in the order E. brucei > C. africana > M. sylvestris > G. robusta (Table 4).
The mean annual nutrient input through leaves litterfall by all the species combined contributed 3125.32 kg C ha− 1 year− 1, 149.70 kg N ha− 1 year− 1, 3.56 kg P ha− 1 year− 1, and 54.86 kg K ha− 1 year− 1 (Table 5). Leaves litterfall C, N, and K inputs differed significantly (P < 0.05) between species, with the highest addition by C. africana, followed by E. brucei for each respective nutrient (Table 5). Except for leaves litterfall N input, the lowest return of C, P, and K was associated with M. sylvestris (Table 5). Although the observed differences were not significant between species, the mean annual P input was relatively higher in G. robusta, followed by C. africana, but with much lower P values in the other species.
Effect of tree characteristics (crown area, DBH, height, and age) and climate variables (precipitation and temperature) on leaves litterfall deposition
Among the different factors affecting leaves litterfall amounts, the results of the present study did seem to indicate that annual leaves litterfall deposition (dry matter kg tree− 1 year− 1) corresponds to tree/vegetation variables such as crown area (positive; R2 = 0. 508, F(1, 18) = 18.615, p = 0.000); DBH (positive; R2 = 0. 404, F(1, 18) = 12. 196, p = 0.003); total height (positive; R2 = 0. 480, F(1, 18) = 16.639, p = 0.001), except for the age of trees (R2 = 0.035, F(1, 18) = 0.651, p = 0.430) (Fig. 5). No significant effect of mean monthly precipitation (R2 = 0.010, F(1, 46) = 0.461, p = 0.501) and temperature (R2 = 0.046, F(1, 46) = 2.194, p = 0.145) was also detected on the monthly leaves litterfall production (Fig. 5).

a-f; 5. Relationships of leaves litterfall amounts with a crown area, b DBH, c total heught, d age, e precipitation, and f temperature. Numbers in each graph are coefficients of determination (R2 values), trend lines are shown for significant relationships only. **p < 0.05, **p < 0.001
Discussion
Leaves litterfall input from multipurpose tree/shrub species in homegarden agroforestry systems
Although leaves litterfall-only values were reported from this study, the combined annual leaves litterfall production for all the four species investigated (9439.33 kg ha− 1) was much higher than the mean annual litterfall production (6277.66 kg ha− 1) documented in the forested areas of traditional homegardens at Barak Valley, Assam, northeast India (Das and Das 2010). But comparable litterfall rates have been reported for different tree/shrub species grown in different agroforestry combinations elsewhere around the world, including Ethiopia (Dawoe et al. 2010; Silva et al. 2011; Negash and Starr, 2013). Although generally lower litterfall mean values were documented in agroforestry than in tropical forest ecosystems (George and Kumar 1998), some previous investigations indicated lower annual mean litterfall values for coniferous and Podocarpus-dominated natural forests (2200–4400 kg ha− 1 year− 1; Ashagrie and Zech 2013) and coniferous species (4900–6600 kg ha− 1 year− 1; Demessie et al. 2012) in different parts of Ethiopia as compared to the present study. In general, the values of leaves litterfall reported in the present homegarden agroforestry (9439.33 kg ha− 1 year− 1) are comparable with the mean global estimates of rainforest litterfall that ranged between 6000 and 12000 kg ha− 1 year− 1 (Edwards et al. 2018). Although the full extent of diverse tree species and various litterfall fractions were not considered in the present homegardens, the present paper results also appeared to indicate that litterfall production was somewhat in line with global litterfall data in tropical seasonal forest (9470 kg ha− 1 year− 1; Tang et al. 2010), but remarkably higher than man-made tropical forest (9230 kg ha− 1 year− 1; Tang et al. 2010), evergreen talweg forest (9200 kg ha− 1 year− 1), dry evergreen forest (9100 kg ha− 1 year− 1), Heath forest (9100 kg ha− 1 year− 1; Vitousek 1984), and cool temperate coniferous tree species (2250–6200 kg ha− 1 year− 1; Hansen et al. 2009).
Taking into account monthly litterfall, the mean values of C. africana (9.99 kg tree− 1), E. brucei (9.96 kg tree− 1), and G. robusta (7.26 kg tree− 1) were found to be higher than that reported for D. sissoo (Roxb.) ex DC (4.52 kg tree− 1), Prosopis cineraria (L.) (4.24 kg tree− 1), Acacia leucophloea (Roxb.) Willd. (3.45 kg tree− 1) and Acacia nilotica (L.) Del. (2.98 kg tree− 1) grown under agroecosystems in a semi-arid region of Rajasthan, India (Yadav et al. 2008). A remarkably lower mean value was recorded for M. sylvestris (0.38 kg tree− 1) in this study as compared to other workers᾽ reports (Yadav et al. 2008). Except M. sylvestris, the average annual leaves litterfall amount per tree for the other three tree species was also generally in the higher range of the values (17.43–23.96 kg tree− 1 year− 1) as compared to that reported by Yang et al. (2021) in the main Channel of the Tarim River, China (10.2–14.83 kg tree− 1). On an area basis, C. africana contributed a significantly higher amount of total litter (962 kg ha− 1 year− 1) to the system as compared to that of E. brucei (429.20 kg ha− 1 year− 1), though they had almost equal amount of annual litter 23.96 kg tree− 1 year− 1 and 23.91 kg tree− 1 year− 1, respectively. Despite higher mean annual leaves litter production on a per tree basis (kg tree− 1 year− 1), E. brucei contributed lower total leaves litterfall to the system owing to its low tree density (17.95 trees ha− 1) as compared to that C. africana (40.14 trees ha− 1), which is twice the stocking level of the former species. In general, the amount of leaves litterfall input varies in quantity and quality may be due to different factors such as climate, floristic composition, tree management, the density of different tree species, canopy characteristics, stand age, and soil factors (Jamaludheen and Kumar 1999; Yadav et al. 2008; Negash and Starr 2013; Zhou et al. 2016).
Seasonal variation of leaves litterfall
The leaf litterfall production in this study remarkably evidenced a clear seasonal pattern, as reported by other studies in traditional homegardens, cocoa ecosystems, and lowland exclosures (Das and Das 2010; Dawoe et al. 2010; Tesfay et al. 2020). In addition, there was also discernable variation in leaves litterfall between species in their monthly and total production like several other previous studies done elsewhere (George and Kumar 1998; Isaac and Nair 2006; Yadav et al. 2008; Das and Das 2010). The monthly litterfall in this study followed a bimodal distribution pattern for C. africana and E. brucei, as suggested by Das and Das (2010) and Negash and Starr (2013) who indicated that tree/shrub species investigated yielded two major annual litterfall peaks. However, the two other species that were characterized by a unimodal distribution pattern in this study include G. robusta and M. sylvestris, and this agrees with Jamaludheen and Kumar (1999) and Verma et al. (2021) who reported litterfall to have followed a unimodal distribution pattern for certain multipurpose tree species in Kerala, India, and arid western region of India, respectively. Contrary to previous studies for woodlots, tropical and subtropical forest ecosystems, and traditional homegardens (Jamaludheen and Kumar, 1999; Das and Das, 2010; Paudel et al., 2015), but consistent with the findings of Spain (1984), Yadav et al. (2008), and Jackson (1978), the present study showed that most species followed a major peak season in the late rainy season (September to October), but for that of M. sylvestris a major peak took place during the early wet season (June to July). These variations of seasonal litterfall between species may be due to the differences in tree management, soil, and climatic conditions prevailing at different sites (Isaac and Nair 2006; Negash and Starr 2013). Another possible reason for the litterfall peak distribution pattern during the early and late rainy seasons may be associated with strong wind and precipitation events, which in turn could mechanically promote pulses of nutrient-rich green and senesced leaf tissue accumulation (Wood et al. 2005; Dawoe et al. 2010; Parsons et al. 2014; Londe et al. 2016; Souza et al. 2019). And, the proportion of senescent foliage biomass is closely linked to the quantity of above-ground litterfall which varies from year to year and between species (Ukonmaanaho et al. 2008). It is also remarkable to note that total leaf litter peaks may be linked to the density and phenological behavior of the species and the foliar strategies of dominant populations in the homegarden agroforestry systems (Facelli and Pickett 1991; Das and Das 2010).
Leaves litterfall nutrient concentrations and returns
In the homegarden agroforestry systems investigated, the concentrations of nutrients in the leaves litterfall revealed considerable variations among species as demonstrated by Chaturvedi and Singh (1987) and Krishna and Mohan (2017). This may be because plant species define their litter value (Krishna and Mohan 2017) and the differences in the age and density of trees in turn determine the quantity of litterfall (Chaturvedi and Singh 1987), and soil nutrient status as well (Chaturvedi and Singh 1987; Wood et al. 2006). There is also greater interception capacity of one species over the other (Ukonmaanaho et al. 2008). At the same time, increasing evidence suggests that the presence of tree species influenced ecosystem biogeochemistry because of the differences in their litter quantity and chemistry returned to the soil and the associated impacts on the activities of diverse soil heterotrophic organisms (Reich et al. 2005). The average C concentration of leaf litterfall in the studied species (42.65–45.95%) was found to be slightly higher than values recorded for five species (36.6–39.8%) in indigenous agroforestry systems in the south-eastern Rift Valley escarpment of Ethiopia (Negash and Starr 2013). However, the values of each C concentration in this investigation were consistent with results from other studies (43–44.17%, Semwal et al. 2003; 48.7–49.3%, Yadav et al. 2008) undertaken under different tree species at Banswara, Central Himalaya, and Jaipur district, Rajasthan, India, respectively.
Based on the mean N concentration of leaves litterfall, and taking into consideration Jamaludheen and Kumar’s (1999) classification, the different species in this study can be grouped into high (> 1.5% N, e.g. E. brucei, C. africana), medium (1–1.5% N, e.g. M. sylvestris), and low (< 1.0% N, e.g. G. robusta) detritus nitrogen species. Leaves litterfall N concentrations in this study (0.77–3.32%) were also comparable with those documented for trees/shrub species under agroforestry systems (Jamaludheen and Kumar 1999; Tripathi et al. 2009; Das and Das 2010; Negash and Starr 2013), and plantation and forest species (Demessie et al. 2012), except with that far exceeding value of leguminous species in the present study. The P concentrations in this study had lower mean values as compared to that reported by Yadav et al. (2008) and Tripathi et al. (2009), but it was comparable to the results of Isaac and Nair (2006), as reported for species grown in combination under agroforestry systems. Concerning K concentration, the findings of the present study were consistent with the mean values suggested in the works of Semwal et al. (2003), Yadav et al. (2008), and Isaac and Nair (2006), but it was considerably higher than that reported by George and Kumar (1998). In general, the overall variation in foliage litterfall chemistry among tree species may have a resultant effect on soil properties (Reich et al. 2005).
Taking into account nutrient returns in homegarden agroforestry, C was found to be quite high for C. africana, followed by E. brucei may be due to a combination of high rates of return of leaves litterfall to the soil surface and high concentrations of nutrients in the fallen material (Zhou et al. 2016). The combined leaves litterfall C input from all the multipurpose trees/shrub species (3125.32 kg C ha− 1 year− 1) under investigation in the present study was comparable with the findings of Paudel et al. (2015) who reported 3162.29–3283.81 kg ha− 1 year− 1 in secondary and mature forests along a disturbance gradient in a tropical montane forest, southwest China. Similarly, the mean C inputs in the present study were in the range of 3050–4900 kg C ha− 1 at the forest land use and 1860–5130 kg C ha− 1 at the jelutung (Dyera costulata) plantation reported in Central Kalimantan, Indonesia (Jaya et al. 2022). But this finding indicated remarkably higher C inputs for all the species investigated in the homegarden agroforestry as compared to Chagga homegardens (1454.1 kg C ha− 1 year− 1), Ocotea forest (1635.7 kg C ha− 1 year− 1), and Podocarpus forest (1600.8 kg C ha− 1 year− 1) at Mt. Kilimanjaro, Tanzania (Becker et al. 2015). The combined leaf litterfall N input from all species (149.70 kg N ha− 1 year− 1) in this study was higher than that reported in the native forest and four plantations (ranging from 36.40 to 123.94 kg N ha− 1 year− 1, at the Curuá-Una forest reserve, Brazil; Smith et al. 1998); Chagga homegardens (87 kg N ha− 1 year− 1), shaded coffee plantation (110.3 kg N ha− 1 year− 1), lower montane forest (48.7 kg N ha− 1 year− 1), Ocotea forest (51.9 kg N ha− 1 year− 1), and Podocarpus forest (38.2 kg N ha− 1 year− 1) at Mt. Kilimanjaro, Tanzania (Becker et al. 2015). At the same time, higher mean leaf litterfall N input was reported in the present study as compared to the findings of Jaya et al. (2022) who documented mean values ranging from 21 to 59 kg ha− 1 year− 1 in secondary peat swamp forests in Central Kalimantan, Indonesia. But when considering individual trees species, higher average leaves litterfall N input contributed by C. africana (17.77 kg N ha− 1 year− 1) and E. brucei (14.79 kg N ha− 1 year− 1) reported here were remarkably higher than the mean total leaf litter values of all species (ranging from 0.001 to 9.543 kg N ha− 1 year− 1) documented in traditional homegardens in Barak Valley, Assam, northeast India (Das and Das 2010). Furthermore, the combined total N fluxes (149.70 kg ha− 1 year− 1) contributed by the four species under this study accounted for almost 2.5 times the magnitude of N input (60 kg N ha− 1 year− 1) from biological nitrogen fixation of legume shade trees, as reported by Beer (1988, cited in Schroth et al. 2001). The same author also indicated that the magnitude of 60 KgN ha− 1 year− 1 is a relevant amount in smallholder coffee plantations with little or no fertilization. This combined total N input in the present study was comparatively higher than that reported in secondary and mature forests (94.74–100.64 kg ha− 1 year− 1) along a disturbance gradient in a tropical montane forest, in southwest China (Paudel et al. 2015).
The combined mean values of leaves litterfall P and K fluxes documented for the four species in the present homegarden agroforestry (3.56 kg P ha− 1 year− 1; 54.86 kg K ha− 1 year− 1) were in the range of mean values of previous studies in respect to each P accretion (2.9–6.5 kg P ha− 1 year− 1, Dawoe et al. 2010; 3.01–7.53 kg P ha− 1 year− 1, Celentano et al 2011; 3.06–8.11 kg P ha− 1 year− 1 Paudel et al. 2015; 0.3– 23.1 kg P ha− 1 year− 1, Froufe et al. 2020), but K accretion was higher than that reported for multipurpose trees in Kerala, India and other types of agroforestry combinations and forests in different parts of the world (7.9–31.3 kg K ha− 1 year− 1, Dawoe et al. 2010; 14–23.1 kg K ha− 1 year− 1, Celentano et al. 2011; 10.87–39.25 kg K ha− 1 year− 1, Paudel et al. 2015; 0.8–41.1 kg K ha− 1 year− 1, Froufe et al. 2020). The low values of P may reflect that this is the main limiting element in the systems evaluated and that the plants in the agroforestry systems returned a low litter result for P (Notaro et al. 2014). The combined nutrient inputs from leaf litterfall by tree/shrub species in the studied homegardens followed the sequence, C > N > K > P somewhat similar to that reported in the homegardens by Isaac and Nair (2006) in Southern Kerala, India. The overall litterfall nutrient contents depend on the availability in the soil and/or the uptake capacity of the plants (Dawoe et al. 2010), or may vary due to the differences in litter quantity and biochemical elements of litter (Verma et al. 2021), and seasonal patterns of litterfall production (Paudel et al. 2015).
Effect of tree characteristics (crown area, DBH, height, and age) and climate variables (precipitation and temperature) on leaf litterfall deposition
When considering the effects of different factors on litterfall amounts, the results of the present study established significant relationships between leaf litterfall deposition and tree characteristics as revealed by several other prior studies. For instance, the present paper clearly showed that leaves litterfall amount increased in direct proportion to the crown area (Cizungu et al. 2014; Asigbaase et al. 2021; Yang et al. 2021), DBH (Murovhi et al. 2012; Negash and Starr 2013; Morffi-Mestre et al. 2020), and height (Murovhi et al. 2012; Negash and Starr 2013; Morffi-Mestre et al. 2020) of tree species in Cocoa, sub-tropical fruit trees, and other shade tree dominated agroforestry systems and forests. Yang et al. (2022), on the other hand, reported that higher litterfall production was observed under conditions of lower DBH and tree height in a Cypress plantation. In contrast to previous findings (Chaturvedi and Singh 1987; Lebret et al. 2001; Berg et al. 1999; Tripathi et al. 2009; Becker et al. 2015; Morffi-Mestre et al. 2020), the present study also dictates that no apparent relationship existed between leaves litterfall deposition and age of trees, as revealed by Tang et al. (2010) in different tropical forest ecosystems.
The monthly leaf litterfall production in the present study did not detect a clear trend in monthly precipitation and temperature, suggesting the need for long-term litterfall studies to confirm seasonal periodicity and establish relations between rainfall and litterfall peaks (Dezzeo and Chacón 2006). Furthermore, Chave et al. (2010) revealed the absence of seasonality dependence of total annual litterfall on total annual rainfall at a global and forest-type level. Contrary to the data obtained, Wang et al. (2013) suggested that there was a positive relationship between precipitation and the quantity of litterfall. Similarly, Fuentes et al. (2018) and Vitousek (1984) also supported Caritat᾽s et al. (2006) findings which dictated a significantly negative relationship between precipitation and the patterns of leaf litter production. Caritat et al. (2006) also reported the strongest correlation between rainfall precipitation and twig litterfall, implying a good concordance of peaks. Similar to this study, other wet season litterfall peaks were also reported by several other workers (Jackson 1978; Meentemeyer et al. 1982; De Moraes et al. 1999). The wet season’s highest litterfall peaks as compared to the dry season might be attributed to other confounding factors such as the presence of diseases and other pathogenic effects (Tesfay 2020), as well as variation in the water holding capacity in soil (De Moraes et al. 1999) and the overlapping individual tree species phenology (Zalamea and Gonzalez 2008; Paudel et al. 2015; Tesfay 2020).
The implication of nutrient return from leaf litterfall of multipurpose trees/shrubs for the sustainability of homegarden agroforestry systems
Litterfall is the main pathway for the return of organic matter and nutrients from plant components to the soil in an ecosystem through the decomposition cycle (Liu et al. 2002; Uriarte et al. 2015). This was proved to be a reality in multistrata agroforestry systems; where they are reported to be more than 50% of all nutrients of leaves that can be accessible to the soil system in less than one year (Froufe et al. 2020). The presence of a large amount of slowly decomposing litter may contribute to the accumulation of a greater quantity of soil organic matter and a larger pool of slow-release nutrients which might sustain crop productivity (Dhanya et al. 2013). The availability of essential nutrients can thus be sustainably maintained by regulating the nutrient cycling processes (Getaneh et al. 2022), which are strongly influenced by, for instance, species composition, tree density, basal area, ages of trees structure (Liu et al. 2002; Tang et al. 2010), and altitude (Liu et al. 2002; Lopes et al. 2015).
Hence, appropriate management strategies that involve the introduction of diverse fast-growing multipurpose woody plants can help provide high-quality and quantity litter resources (Dawoe et al. 2010). The variation in species composition along with the quantity and quality of litter produced can influence nutrient return to the soil, and thus the availability of soil nutrients, which in turn could have a direct effect on ecosystem processes (Parsons et al. 2014; Uriarte et al. 2015). This in turn may augment soil fertility in these systems (Yang et al. 2004; Dawoe et al. 2010; Getaneh et al. 2022). Therefore, litter production can serve as a major pathway by which nutrients are transferred from vegetation to soil (Parsons et al. 2014; Uriarte et al. 2015), and can thus act as an input–output system for nutrients where their variability can determine nutrient cycling, soil fertility and plant productivity in agroecosystems (Dawoe et al. 2010; Parsons et al. 2014). In general, the patterns of litter production and its subsequent decomposition up on which bioelements are released endow sustainability to these homegardens (Kumar and Nair 2004). In addition, past research efforts also revealed that interspecific differences in leaf litter production and quality are expected to have a marked effect on variation in soil physical characteristics and nutrient availability (Uriarte et al. 2015). Together with proper silvicultural management such as pruning, leaf litter is also considered to be a potential source for obtaining organic matter that may highlight the right way for maintaining the long-term productive capacity of soils, and subsequent enhancement of homegarden agroforestry systems sustainability (Froufe et al. 2020; Gerke 2022).
Conclusions
A comparably higher total annual quantity of leaf litterfall was registered in the present study as compared to other agroforestry systems, monocultural tree plantations, and natural forests confined elsewhere in tropical and subtropical ecozones. The sequence of the relative leaf litterfall contribution of multipurpose tree/shrub species with higher mean values was Cordia africana > Erythrina brucei > Grevillea robusta > Malus sylvestris. A clear seasonal leaf litterfall pattern with varying peaks was observed for the multipurpose tree/shrub species in the present study, with a bimodal distribution pattern for E. brucei and C. africana and a unimodal pattern for G. robusta and M. sylvestris. Similarly, both the concentrations and inputs of nutrients from the leaf litter showed considerable variations among species found in the homegarden agroforestry systems. Among the investigated species, C. africana and E. brucei were found to likely contribute better in terms of C and N return, G. robusta relatively the better in terms of phosphorous return, and C. africana as well better for potassium return. However, E. brucei had a higher average concentration of N than the rest. The most important trees/shrubs in the studied sites that reflected differences in the quantity and quality (nutrient content) of leaf litter production imply that there existed variation in foliar nutrient content among each species, and its consequent effect for sustainably enhancing the productive capacity of soils under homegarden agroforestry systems. Quantification of such leaf litterfall and nutrient flux may be essential in providing useful information and making decisions on the potential contributions of multipurpose woody plants for carefully evaluating its consequent effect on nutrient cycling and restoring soil fertility of homegarden agroforestry systems. The amount of leaf litterfall differed among tree species as influenced by tree species characteristics (canopy area, DBH, height), but independent of age and climate variables (monthly precipitation and temperature). Further research on long-term tree litter dynamics, decomposition, and associated nutrient release may be warranted in these homegardens, including the various relevant fractions of litterfall.
Availability of data and materials
Upon a reasonable request, data sets associated with the paper will be made available by contacting the corresponding author.
References
Abbate E, Bruni P, Sagri M (2015) Geology of Ethiopia: A review and geomorphological perspectives. In: Billi P (ed) Landscapes and landforms of Ethiopia, world geomorphological landscapes. Springer, Dordrecht, pp 33–64
Abebe T (2013) Determinants of crop diversity and composition in Enset-coffee agroforestry homegardens of Southern Ethiopia. JARTS 114(1):29–38
Abebe T, Bongers F (2012) Land-use dynamics in enset-based agroforestry homegardens in Ethiopia. In: van Bommel S, Ros-Tonen M, Verschoor G (eds) Arts B. Understanding community forestry and biocultural diversity. Wageningen Academic Publishers The Netherlands, Forest-people interfaces, pp 69–85
Abebe T, Wiersum KF, Bongers F, Sterck F (2006) Diversity and dynamics in homegardens of Southern Ethiopia. In: Kumar BM, Nair PKR (eds) Tropical homegardens: a time-tested example of sustainable agroforestry. Springer, Dordrecht, pp 33–64
Anderson JM, Ingram JSI (1993) Tropical soil biology and fertility: A handbook of methods. CAB International, Oxon
Andivia E, Fernández M, Vázquez-Piqué J, González-Pérez A, Tapias R (2010) Nutrients return from leaves and litterfall in a Mediterranean cork oak (Quercus suber L.) forest in southwestern Spain. Eur J Forest Res 129:5–12. https://doi.org/10.1007/s10342-009-0274-6
Asfaw Z (2007) Ågren IG (2007) Farmers’ local knowledge and topsoil properties of agroforestry practices in Sidama Southern Ethiopia. Agrofor Syst 71:35–48. https://doi.org/10.1007/s10457-007-9087-0
Ashagrie Y, Zech W (2013) Litter production and nutrient cycling in two plantations and a Podocarpus falcatus dominated natural forest ecosystems in south-eastern highlands of Ethiopia. Afr J Agric Res 8(38):4810–4818
Asigbaase M, Dawoe E, Lomax BH (2021) Sjogersten S (2021) Temporal changes in litterfall and potential nutrient return in cocoa agroforestry systems under organic and conventional management. Ghana Heliyon 7:e08051. https://doi.org/10.1016/j.heliyon.2021.e08051
Becker J, Pabst H, Mnyonga J, Kuzyakov Y (2015) Annual litterfall dynamics and nutrient deposition depending on elevation and land use at Mt Kilimanjaro. Biogeosciences 12:5635–5646. https://doi.org/10.5194/bg-12-5635-2015
Bekele T (1993) Vegetation ecology of remnant Afromontane forests on the Central Plateau of Shewa, Ethiopia. Acta Phytogeogr Suec 79, Opulus Press AB, Uppsala, Sweden, pp. 61. ISBN 91-72 1 0- 079-6. (91-7210-479-1)
Bekele T (1994) Phytosociology and ecology of a humid Afromontane forest on the central plateau of Ethiopia. J Veget Sci 5:87–98
Berg B, Albrektson A, Berg MP, Cortina J, Johansson MB, Gallardo A, Madeira M, Pausas G, Kratzi W, Vallejo R, McClaugherty C (1999) Amounts of litter fall in some pine forests in a European transect, in particular Scots pine. Ann for Sci 56(8):625–639
Berhanu B, Melesse AM, Seleshi Y (2013) GIS-based hydrological zones and soil geo-database of Ethiopia. Catena 104:21–31. https://doi.org/10.1016/j.catena.2012.12.007
Caritat A, Garcia-Borthou E, Lapenã R, Vilar L (2006) Litter production in a Quercus suber forest of Montseny (NE Spain) and its relationship to meteorological conditions. Ann for Sci 63:791–800. https://doi.org/10.1051/forest:2006061
Castro A, Lascurain-Rangel M, Gomez-Díaz JA, Sosa V (2018) Mayan homegardens in decline: the case of the Pitahaya (Hylocereus undatus), a vine cactus with edible fruit. Trop Conserv Sci 11:1–10
Celentano D, Zahawi RA, Finegan B, Ostertag R, Cole RJ, Holl KD (2011) Litterfall dynamics under different tropical forest restoration strategies in Costa Rica. Biotropica 43:279–287. https://doi.org/10.1111/j.1744-7429.2010.00688.x
Chaturvedi OP, Singh JS (1987) A quantitative study of the forest floor biomass, litter fall and nutrient return in a Pinus roxburghii forest in Kumaun Himalaya. Vegetatio 71:97–106
Chave J, Navarrete D, Almeida S, Álvarez E, Aragão LE, Bonal D, Châtelet P, Silva-Espejo JE, Goret JY, Hildebrand PV, Jiménez E, Patiño S, Penuela MC, Phillips OL, Stevenson P, Malhi Y (2010) Regional and seasonal patterns of litterfall in tropical South America. Biogeosciences 7:43–55
Cizungu L, Staelens J, Huygens D, Walangululu J, Muhindo D, Van Cleemput O, Boeckx P (2014) Litterfall and leaf litter decomposition in a central African tropical mountain forest and Eucalyptus plantation. For Ecol Manage 326:109–116. https://doi.org/10.1016/j.foreco.2014.04.015
Coltorti M, Pieruccini P, Arthur KJW, Arthur J, Curtis M (2019) Geomorphology, soils and palaeosols of the Chencha area (Gamo Gofa, south western Ethiopian Highlands). J Afr Earth Sci 151:225–240
Currey R (2009) Diversity of hymenoptera, cultivated plants and management practices in home garden agroecosystems, Kyrgyz Republic. Florida International University Electronic Theses and Dissertations. 124. https://doi.org/10.25148/etd.FI09120805
Das T, Das AK (2010) Litter production and decomposition in the forested areas of traditional homegardens: a case study from Barak Valley, Assam, Northeast India. Agrofor Syst 79:157–170. https://doi.org/10.1007/s10457-010-9284-0
Das T, Das AK (2010) Litter production and decomposition in the forested areas of traditional homegardens: a case study from Barak Valley, Assam, Northeast India. Agrofor Syst 79:157–170. https://doi.org/10.1007/s10457-010-9284-0
Das T, Das AK (2010) Litter production and decomposition in the forested areas of traditional homegardens: a case study from Barak Valley, Assam, Northeast India. Agrofor Syst 79:157–170. https://doi.org/10.1007/s10457-010-9284-0
Dawoe EK, Isaac ME, Quashie-Sam J (2010) Litterfall and litter nutrient dynamics under cocoa ecosystems in lowland humid Ghana. Plant Soil 330:55–64. https://doi.org/10.1007/s11104-009-0173-0
De Moraes RM, Delitti WBC, Vuono YS (1999) Litterfall and litter nutrient content in two Brazilian tropical forests. Rev Bras Bot 22(1):9–16
Debebe D, Ayele T (2013) Land use patterns and its implication for climate change: the case of Gamo Gofa, southern Ethiopia. Int J Sci Res Rev 2(3):155–173
Demessie A, Singh BR, Lal R, Strand LT (2012) Leaf litter fall and litter decomposition under Eucalyptus and coniferous plantations in Gambo District, southern Ethiopia. Acta Agric Scand-B Soil Plant Sci 62(5):467–476. https://doi.org/10.1080/09064710.2011.645497
Demissew S, Nordal I (2010) Aloes and Lilies of Ethiopia and Eritrea.Shama Books, Addis Ababa, Ethiopia, pp. 351
Dezzeo N, Chacón N (2006) Litterfall and nutrient input in undisturbed and adjacent fire disturbed forests of the gran sabana, southern Venezuela. Interciencia 31(21):894–899
Dhanya B, Viswanath S, Purushothaman S (2013) Decomposition and nutrient release dynamics of Ficus benghalensis L litter in traditional agroforestry systems of Karnataka, Southern India. ISRN Forestry. https://doi.org/10.1155/2013/524679
Edwards W, Liddell MJ, Franks P, Nichols C, Laurance SGW (2018) Seasonal patterns in rainforest litterfall: detecting endogenous and environmental influences from long-term sampling. Austral Ecol 43(2):225–235. https://doi.org/10.1111/aec.12559
Facelli JM, Pickett STA (1991) Plant litter: its dynamics and effects on plant community structure. Bot Rev 57:1–32
FDRE PCC (Federal Democratic Republic of Ethiopia Population Census Commission) (2008) Summary and Statistical Report of the 2007 Population and Housing Census: Population size and sex. United Nations Population Fund(UNFPA), Addis Ababa, Ethiopia. http://www.ethiopianreview.com/pdf/001/Cen2007_firstdraft(1).pdf
Fernandes ECM, Nair PKR (1986) An evaluation of the structure and function of tropical homegardens. Agric Syst 21:279–310
Field A (2009) Discovering statistics using SPSS. SAGE Publications Ltd., London
Friis I, Demissew S, van Breugel P (2010) Atlas of the potential vegetation of Ethiopia the royal danish academy of sciences and letters. Copenhagen, Denmark, pp. 307
Friis I (1992). Forests and forest trees of northeast tropical africa: their natural habitats and distribution patterns in Ethiopia, Djibouti and Somalia. Royal Botanical Gardens, Kew Bulletin, Additional Series XV, Her Majesty's Stationery Office (HMSO), London, pp. 396
Froufe LCM, Schwiderke DK, Castilhano AC, Cezar RM, Steenbock W, Seoane CES, Bognola IA, Vezzani FM (2020) Nutrient cycling from leaf litter in multistrata successional agroforestry systems and natural regeneration at Brazilian Atlantic Rainforest Biome. Agrof Syst. https://doi.org/10.1007/s10457-019-00377-5
Fuentes N, Vizcaino LY, Britto TS (2018) Leaf litter production of a tropical forest of bank Santa Marta Colombia. Contemp Eng Sci 11(87):4301–4310. https://doi.org/10.12988/ces.2018.88444
Gajaseni J, Gajaseni N (1999) Ecological rationalities of the traditional homegarden system in the Chao Phraya Basin, Thailand. Agrofor Syst 46:3–23
Gallardo A, Merino J (1993) Leaf decomposition in two mediterranean ecosystems of Southwest Spain: influence of substrate quality. Ecology 74:152–161
George SJ, Kumar BM (1998) Litter dynamics and cumulative soil fertility changes in silvopastoral systems of a humid tropical region in central Kerala, India. Int Tree Crops J 9:267–282
Gerhardt K, Hytteborn H (1992) Natural dynamics and regeneration methods in tropical dry forests: an introduction. J Veget Sci 3(3):361–364. https://doi.org/10.2307/3235761
Gerke J (2022) The central role of soil organic matter in soil fertility and carbon storage. Soil Syst 6(33):1–14. https://doi.org/10.3390/soilsystems6020033
Getaneh S, Honnay O, Desie E, Helsen K, Couck L, Shibru S, Muys B (2022) Impact of tree litter identity, litter diversity and habitat quality on litter decomposition rates in tropical moist evergreen forest. For Ecosyst 9:100023. https://doi.org/10.1016/j.fecs.2022.100023
Hairiah K, Sulistyani H, Suprayogo D, Widianto PP, Widodo RH, Van Noordwijk M (2006) Litter layer residence time in forest and coffee agroforestry systems in Sumberjaya, West Lampung. For Ecol Manag 224:45–57. https://doi.org/10.1016/j.foreco.2005.12.007
Hansen K, Veterdal L, Schmidt IK, Gundersen P, Sevel L, Bastrup-Birk A, Pedersen LB, Bille-Hansen J (2009) Litterfall and nutrient return in five tree species in a common garden experiment. For Ecol Manag 257:2133–2144. https://doi.org/10.1016/j.foreco.2009.02.021
Hasanuzzaman Md, Hossain M (2014) Litter production and nutrient return through leaf litter of selected cropland agroforest tree species in southwestern Bangladesh. Agric Forestry 60(1):221–233
Hossain M, Siddique MRH, Hossain MdSRMdZ, Hasan MdM (2011) Nutrient dynamics associated with leaf litter decomposition of three agroforestry tree species (Azadirachta indica, Dalbergia sissoo, and Melia azedarach) of Bangladesh. J Forestry Res 22(4):577–582. https://doi.org/10.1007/s11676-011-0175-7
Isaac SR, Nair MA (2006) Litter dynamics of six multipurpose trees in a homegarden in Southern Kerala, India. Agrofor Syst 67:203–213. https://doi.org/10.1007/s10457-005-1107-3
Jackson ML (1958) Soil chemical analyses. Prentice Hall, New Delhi
Jackson JF (1978) Seasonality of flowering and leaf fall in a Brazilian subtropical lower montane moist forest. Biotropica 10:38–42
Jamaludheen V, Kumar BM (1999) Litter of multipurpose trees in Kerala, India: variation in the amounts of quality, decay rates and release of nutrients. For Ecol Manage 115:1–11
Jaya A, Sancho JJ, Page S, Sjorgensen S, Evans C, Sulistiyanto Y (2022) Litter production and leaf litter decomposition rate in secondary peat swamp forests in Central Kalimantan, Indonesia. IOP Conf Ser Earth Environ Sci 1025:012024. https://doi.org/10.1088/1755-1315/1025/1/012024
Kassa G, Abebe T, Ewnetu Z (2015) Diversity, density and management of trees in different agroforestry practices of Yem special district, southern Ethiopia. SINET Ethiop J Sci 38(1):01–16
Kehlenbeck K, Mass BL (2004) Crop diversity and classification of homegardens in central Sulawesi, Indonesia. Agrofor Syst 63:53–62
Kehlenbeck K, Arifin H, Maass B (2007) Plant diversity in homegardens in a socio-economic and agro-ecological context. In: Tscharntke T, Leuschner C, Zeller M, Guhardja E, Bidin A (eds) The stability of tropical rainforest margins: Linking ecological, economic and social constraints of land use and conservation. Springer, Berlin, pp 297–317
Kehlenbeck K (2007) Rural Homegardens in Central Sulawesi, Indonesia: An Example for a Sustainable Agro-Ecosystem? Ph.D. dissertation. Georg-August-Universität, Göttingen
Krishna MP, Mohan M (2017) Litter decomposition in forest ecosystems: a review. Energ Ecol Environ 2(4):236–249. https://doi.org/10.1007/s40974-017-0064-9
Kumar BM (2008) Litter dynamics in plantation and agroforestry systems of the tropics: a review of observations and methods. In: Batish DR, Kohli RK, Jose S, Singh HP (eds) Ecological basis of agroforestry. CRC Press, Boca Raton, pp 181–216
Kumar BM, Nair PKR (2004) The enigma of tropical homegardens. Agrofor Syst 61:135–152
Kunhamu TK, Kumar BM, Viswanath S (2009) Does thinning affect litterfall, litter decomposition, and associated nutrient release in Acacia mangium stands of Kerala in peninsular India? Can J for Res 39:792–802. https://doi.org/10.1139/X09-008
Kuyah S, Muthuri C, Jamnadass R, Mwangi P, Neufeldt H, Dietz J (2012) Crown area allometries for estimation of aboveground tree biomass in agricultural landscapes of western Kenya. Agrofor Syst 86:267–277
Kuyah S, Sileshi GW, Luedeling E, Akinnifesi FK, Whitney CW, Bayala J, Kuntashula E, Dimobe K, Mafongoya PL (2020) Potential of agroforestry to enhance livelihood security in Africa. In: Dagar JC, Gupta SR, Teketay D (eds) Agroforestry for degraded landscapes: recent advances and emerging challenges. Springer, Singapore, pp 135–167
Lalremsang P, Gopichand P, Upadhaya K, Singson L, Sahoo LS (2022) Leaf litter decomposition and nutrient release dynamics of flemingia semialata roxb—a potential woody perennial species for mountain agroforestry. J Sust Forestry. https://doi.org/10.1080/10549811.2022.2045504
Lebret M, Nys C, Forgeard F (2001) Litter production in an Atlantic beech (Fagus sylvatica L.) time sequence. Ann For Sci 58:755–768
Liu W, Fox JED, Xu Z (2002) Litterfall and nutrient dynamics in a montane moist evergreen broad-leaved forest in Ailao Mountains, SW China. Plant Ecol 164:157–170
Londe V, De Sousa HC, Kozovits SR (2016) Litterfall as an indicator of productivity and recovery of ecological functions in a rehabilitated riparian forest at Das Velhas River, southeast Brazil. Trop Ecol 57(2):355–360
Lopes MCA, Araújo VFP, Vasconcellos A (2015) The effects of rainfall and vegetation on litterfall production in the semiarid region of northeastern Brazil. Braz J Biol 75(3):703–708. https://doi.org/10.1590/1519-6984.21613
López-Rodríguez G, Sotomayor-Ramírez D, Amador JA, Schröder EC (2015) Contribution of nitrogen from litter and soil mineralization to shade and sun coffee (Coffea arabica L.) agroecosystems. Tropical Ecol 56(2):155–167
Mazengia W, Tadesse Y, Schulte RPO, Struik PC (2016) The analysis of potato farming systems in Chencha, Ethiopia: Input, output and constraints. Am J Potato Res 93:436–447. https://doi.org/10.1007/s12230-016-9521-9
Meentemeyer V, Box EO, Thompson R (1982) World patterns and amounts of terrestrial plant litter production. BioScience 32(2):125–128
Mellisse BT, van de Ven GWJ, Giller KE, Descheemaeker K (2018) Home garden system dynamics in southern Ethiopia. Agrofor Syst 92:1579–1595. https://doi.org/10.1007/s10457-017-0106-5
Morffi-Mestre H, Ángeles-Pérez G, Powers JS, Andrade JL, Ruiz AHH, May-Pat F, Dupuy JM (2020) Multiple factors influence seasonal and interannual litterfall production in a tropical dry forest in Mexico. Forests 11:1241. https://doi.org/10.3390/f11121241
Murovhi NR, Materechera SA, Mulugeta SD (2012) Seasonal changes in litter fall and its quality from three sub-tropical fruit tree species at Nelspruit South Africa. Agrofor Syst. https://doi.org/10.1007/s10457-012-9508-6
Nair PKR (1993) An introduction to agroforestry. Kluwer Academic Publishers, Dordrecht
Negash M, Starr M (2013) Litterfall production and associated carbon and nitrogen fluxes of seven woody species grown in indigenous agroforestry systems in the south-eastern Rift Valley escarpment of Ethiopia. Nutr Cycl Agroecosyst 97:29–41. https://doi.org/10.1007/s10705-013-9590-9
Negash M, Starr M (2021) Litter decomposition of six tree species on indigenous agroforestry farms in south-eastern Ethiopia in relation to litterfall carbon inputs and modelled soil respiration. Agrofor Syst 95:755–766. https://doi.org/10.1007/s10457-021-00630-w
Negash M, Yirdaw E, Luukkanen O (2012) Potential of indigenous multistrata agroforests for maintaining native floristic diversity in the south-eastern Rift Valley escarpment, Ethiopia. Agrofor Syst 85:9–28. https://doi.org/10.1007/s10457-011-9408-1
Notaro KA, Erika VM, Gustavo PD, Aline OS, Patrícia MM (2014) Agroforestry systems, nutrients in litter and microbial activity in soils cultivated with coffee at high altitude. Sci Agric 71(2):87–95. https://doi.org/10.1590/S0103-90162014000200001
Parsons SA, Congdon RA, Shoo LP, Valdez-Ramirez V, Williams, (2014) Spatial variability in litterfall, litter standing crop and litter quality in a tropical rain forest region. Biotropica. https://doi.org/10.1111/btp.12113
Paudel E, Dossa GGO, Xu J, Harrison RD (2015) Litterfall and nutrient return along a disturbance gradient in a tropical montane forest. For Ecol Manage 353:97–106. https://doi.org/10.1016/j.foreco.2015.05.028
R Development Core Team (2020) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Reich PB, Oleksyn J, Modrzynski J, Mrozinski P, Hobbie SA, Eissenstat DM, Chorover J, Chadwick OA, Hale CM, Tjoelker MG (2005) Linking litter calcium, earthworms and soil properties: a common garden test with 14 tree species. Ecol Lett 8:811–818. https://doi.org/10.1111/j.1461-0248.2005.00779.x
Samberg LH, Shennan C, Zavaleta ES (2010) Human and environmental factors affect patterns of crop diversity in an Ethiopian highland agroecosystem. Prof Geogr 62(3):395–408
Sari RR, Rozendaal DMA, Saputra DW, Hairiah K, Roshetko JM, van Noordwijk M (2022) Balancing litterfall and decomposition in cacao agroforestry systems. Plant Soil 473:251–271. https://doi.org/10.1007/s11104-021-05279-z
Schlesinger W (1977) Carbon balance in terrestrial detritus. Annu Rev Ecol Evol Syst 8:51–81
Schroth G (2003) Decomposition and nutrient supply from biomass. In: Schroth G, Sinclair FL (eds) Trees, crops and soil fertility: concepts and research methods. CABI Publishing, Wallingford, UK, pp 131–150
Schroth G, Lehmann J, Rodrigues MRL, Barros E, Macêdo JLV (2001) Plant-soil interactions in multistrata agroforestry in the humid tropics. Agrofor Syst 53:85–102
Semwal RL, Maikhuri RK, Rao KS, Sen KK, Saxena KG (2003) Leaf litter decomposition and nutrient release patterns of six multipurpose tree species of central Himalaya, India. Biomass Bioenergy 24:3–11
Senbeta F, Denich M (2006) Effects of wild coffee management on species diversity in the Afromontane rainforests of Ethiopia. For Ecol Manage 232:68–74
Silva AKL, Vasconcelos SS, de Carvalho CJR, Cordeiro IMCC (2011) Litter dynamics and fine root production in Schizolobium parahyba var. amazonicum plantations and regrowth forest in Eastern Amazon. Plant Soil 347:377–386. https://doi.org/10.1007/s11104-011-0857-0
Singh KP, Singh PK, Tripathi SK (1999) Litterfall, litter decomposition and nutrient release patterns in four native tree species raised on coal mine spoil at Singrauli, India. Biol Fertil Soils 29:371–378
Smith K, Gholz HL, Oliveira FA (1998) Litterfall and nitrogenuse efficiency of plantations and primary forest in the eastern Brazilian Amazon. Forest Ecol Manage 109:209–220
Souza SR, Veloso MDM, Espírito-Santo MM, Silva JO, Sánchez-Azofeifa A, Brito e BGS, Fernandes GW (2019) Litterfall dynamics along a successional gradient in a Brazilian tropical dry forest. For Ecosyst 6(35):1–12
Spain AV (1984) Litterfall and the standing crop of litter in three tropical Australian rainforests. J Ecol 72:947–961
Takaoka S (2008) Long-term growth performance of Cordia africana and Grevillea robusta trees in the Mount Kenya region. Agrofor Syst 72:169–172. https://doi.org/10.1007/s10457-007-9056-7
Tang J-W, Cao M, Zhang J-H, Li M-H (2010) Litterfall production, decomposition and nutrient use efficiency varies with tropical forest types in Xishuangbanna, SW China: a 10-year study. Plant Soil 335:271–288. https://doi.org/10.1007/s11104-010-0414-2
Tangjang S, Arunachalam A, Arunachalam K, Deb S (2015) Litterfall, decomposition and nutrient dynamics in traditional agroforestry systems of Northeast India. Int J Ecol Environ Sci 41(1–2):43–53
Teklay T (2007) Decomposition and nutrient release from pruning residues of two indigenous agroforestry species during the wet and dry seasons. Nutr Cycl Agroecosyst 77:115–126. https://doi.org/10.1007/s10705-006-9048-4
Tesfay F, Kibret K, Gebrekirstos A, Hadgu KM (2020) Litterfall production and associated carbon and nitrogen fux along exclosure chronosequence at Kewet district, central lowland of Ethiopia. Environ Syst Res 9(11):1–12. https://doi.org/10.1186/s40068-020-00172-7
Tesfay HM (2020) Ecosystem services of the indigenous agroforestry systems in the south-eastern Rift-valley landscapes, Ethiopia: Plant diversity, carbon pools, soil fertility and local livelihoods’ support PhD dissertation. University of Natural Resources and Life Sciences, Austria
Triadiati TS, Guhardja E, Sudarsono QI, Leuschner C (2011) Litterfall production and leaf-litter decomposition at natural forest and Cacao agroforestry in central Sulawesi Indonesia Asian. J Biol Sci 4(3):221–234. https://doi.org/10.3923/ajbs.2011.221.234
Tripathi OP, Pandey HN, Tripathi RS (2009) Litter production, decomposition and physico-chemical properties of soil in 3 developed agroforestry systems of Meghalaya Northeast India. Afr J Plant Sci 3(8):160–167
Ukonmaanaho L, Merilä P, Nöjd P, Nieminen TM (2008) Litterfall production and nutrient return to the forest floor in Scots pine and Norway spruce stands in Finland. Boreal Env Res 13(suppl B):67–91
Uriarte M, Turner BL, Thompson J, Zimmerman JK (2015) Linking spatial patterns of leaf litterfall and soil nutrients in a tropical forest: a neighborhood approach. Ecol Appl 25(7):2022–2034
van Ranst E, Dumon M, Tolossa AR, Cornelis J-T, Stoops G, Vandenberghe RE, Deckers J (2011) Revisiting ferrolysis processes in the formation of Planosols for rationalizing the soils with stagnic properties in WRB. Geoderma 163:265–274. https://doi.org/10.1016/j.geoderma.2011.05.002
Verma A, Kumar P, Soni ML, Pawar N, Pradhan U, Tanwar SPS, Kumar S (2021) Litter production and litter dynamics in different agroforestry systems in the arid western region of India. Biol Agric Hortic. https://doi.org/10.1080/01448765.2021.1971110
Vitousek PM (1984) Litterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 65(1):285–298
Wana D, Woldu Z (2005) Vegetation of Chencha highlands in southern Ethiopia. SINET Ethiop J Sci 28(2):109–118
Wang H-C, Wang S-F, Lin K-C, Shaner P-JL, Lin T-C (2013) Litterfall and element fluxes in a natural hardwood forest and a Chinese-fir plantation experiencing frequent typhoon disturbance in Central Taiwan. Biotropica 45(5):541–548. https://doi.org/10.1111/btp.12048
White F (1983) The vegetation of Africa: A descriptive memoir to accompany the UNESCO/AETFAT/UNESCO vegetation map of Africa. Natural Resource Research 20, UNESCO, Paris, pp 354
Wiehle M, Goenster S, Gebauer J, Mohamed SA, Andreas B, Katja K (2014) Effects of transformation processes on plant species richness and diversity in homegardens of the Nuba Mountains, Sudan. Agrofor Syst 88:539–562. https://doi.org/10.1007/s10457-014-9717-2
Wood TE, Lawrence D, Clark DA (2005) Variation in leaf litter nutrients of a Costa Rican rain forest is related to precipitation. Biogeochemistry 73:417–437. https://doi.org/10.1007/s10533-004-0563-6
Wood TE, Lawrence D, Clark DA (2006) Determinants of leaf litter nutrient cycling in a tropical rain forest: soil fertility versus topography. Ecosystems 9:700–710. https://doi.org/10.1007/s10021-005-0016-7
Wu W, Zhang Y, Wang L, Zhou Y, Chen Y, He S, Zhang J, Liu Y (2021) Litterfall and element return in an Abies faxoniana forest in Tibet—a five-year study. Forests 12:1577. https://doi.org/10.3390/f12111577
Yadav RS, Yadav BL, Chhipa BR (2008) Litter dynamics and soil properties under different tree species in a semi-arid region of Rajasthan, India. Agrofor Syst 73:1–12
Yang YS, Guo JF, Chen GS, Xie JS, Cai LP, Lin P (2004) Litterfall, nutrient return, and leaf-litter decomposition in four plantations compared with a natural forest in subtropical China. Ann for Sci 61:465–476
Yang YS, Guo JF, Chen GS, Xie GS, Gao R, Li Z, Jin Z (2005) Litter production, seasonal pattern and nutrient return in seven natural forests compared with a plantation in southern China. Forestry 78(4):403–415. https://doi.org/10.1093/forestry/cpi044
Yang Y, Zhou H, Ye Z, Zhu C (2021) Estimation of Populus euphratica forest leaf litterfall and time variation of nutrient in leaf litter during decomposition along the main channel of the Tarim River. China Water 13:2514. https://doi.org/10.3390/w13182514
Yang Y, Yang H, Wang Q, Dong Q, Yang J, Wu L, You C, Hu J, Wu Q (2022) Effects of two management practices on monthly litterfall in a Cypress plantation. Forests 13:1581. https://doi.org/10.3390/f13101581
Zalamea M, Gonzalez G (2008) Leaffall Phenology in a Subtropical Wet Forest in Puerto Rico: From Species to Community Patterns. Biotropica 40(3):295–304. https://doi.org/10.1111/j.1744-7429.2007.00389.x
Zhou J, Lang X, Du B, Zhang H, Liu H, Zhang Y, Shang L (2016) Litterfall and nutrient return in moist evergreen broad-leaved primary forest and mixed subtropical secondary deciduous broadleaved forest in China. Eur J Forest Res 135:77–86. https://doi.org/10.1007/s10342-015-0918-7
Acknowledgements
The authors would like to gratefully acknowledge the laboratory support from the staff of the Department of Plant Biology and Biodiversity Management, especially those of Plant Physiology/Ecophysiology at Addis Ababa University and the Chemistry Department at Arba Minch University. Our thanks also go to these two Universities for covering some portions of this research. We further thank the homegarden agroforestry owners for granting permission to carry out the research work, key informants, and local agricultural experts who kindly collaborated in sharing useful information and for their assistance during the fieldwork.
Funding
The research was supported by Addis Ababa and Arba Minch Universities, Ethiopia.
Author information
Authors and Affiliations
Contributions
GK conceived and performed the field research and lab analysis. GK and TA designed the methodology. GK analyzed the data and wrote the original draft preparation with input from TB, SD, and TA. Funding acquisition was facilitated through TB and SD. TB, SD, and TA supervised the research project, and reviewed and edited the paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix
Appendix
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kassa, G., Bekele, T., Demissew, S. et al. Leaves litterfall and nutrient inputs from four multipurpose tree/shrub species of homegarden agroforestry systems. Environ Syst Res 11, 29 (2022). https://doi.org/10.1186/s40068-022-00278-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40068-022-00278-0