
Abstract
Since 2011, unprecedented pelagic sargassum seaweed blooms have occurred across the tropical North Atlantic, with severe socioeconomic impacts for coastal populations. To investigate the role of physical drivers in post-2010 sargassum blooms in the Central West Atlantic (CWA), conditions are examined across the wider tropical North Atlantic, using ocean and atmospheric re-analyses and satellite-derived datasets. Of particular consequence for the growth and drift of sargassum are patterns and seasonality of winds and currents. Results suggest that in years of exceptionally large sargassum blooms (2015, 2018), the Intertropical Convergence Zone (ITCZ), an area of maximum wind convergence where sargassum naturally accumulates, shifted southward, towards nutrient-rich waters of the Amazon River plume and the equatorial upwelling zone further stimulating sargassum growth. These changes are associated with modes of natural variability in the tropical Atlantic, notably a negative phase of the Atlantic Meridional Mode (AMM) in 2015 and 2018, and a positive phase of the Atlantic Niño in 2018. Negative AMM in these 2 years is also associated with stronger trade winds and enhanced northwest Africa upwelling, probably resulting in stronger southwestward nutrient transport into the eastern part of CWA. Moreover, in contrast with most years, important secondary winter blooms took place in both 2015 and 2018 in the northern part of CWA, associated with excessive wind-driven equatorial upwelling and anomalously strong northwestward nutrient transport.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Since 2011, pelagic sargassum macroalgae have proliferated across the tropical North Atlantic. The extensive longitudinal distribution of sargassum across this region, clearly evidenced by satellite observations, has recently been named “the Great Atlantic sargassum belt” (Wang et al. 2019). As a consequence of recurrent tropical sargassum blooms, Caribbean, Central American, and West African countries have been experiencing excessive stranding events (Franks et al. 2012; Gower et al. 2013; Smetacek and Zingone 2013; Oyesiku and Egunyomi 2014; Schell et al. 2015; Wang and Hu 2017) whereby significant volumes of sargassum seaweed have washed on to beaches and then slowly decompose. These sargassum beaching events have strong implications for coastal populations causing damage to health and livelihoods, negatively affecting fishing, tourism, and nearshore biota (van Tussenbroek et al. 2017; Fidai et al. 2020).
Before 2011, sargassum was mainly observed in the Sargasso Sea of the western subtropical gyre in the North Atlantic (Lapointe 1986, 1995) with minimal presence in the tropical region. However, a pronounced large-scale anomaly of winds and associated ocean currents induced by an exceptionally large negative phase of the North Atlantic Oscillation in late 2010 led to a massive anomalous transport of sargassum biomass from the Sargasso Sea to the central tropical North Atlantic (Johns et al. 2020). Since then, pronounced sargassum blooms have been observed through remote sensing almost every year across this region in spring and summer months, in particular (Gower et al. 2013; Franks et al. 2016; Wang and Hu 2016; Wang et al. 2019; Johns et al. 2020). Sargassum floating across the tropical North Atlantic mainly comprises two species, S. natans and S. fluitans (Schell et al. 2015; Davis et al. 2021) with both species also being typically observed in the Sargasso Sea (Lapointe 1986, 1995).
In contrast with oligotrophic conditions encountered in the Atlantic western subtropical region (Sargasso Sea), the surface layer of the tropical North Atlantic receives relatively large amounts of nutrients from various sources including the Amazon and Orinoco river outflows, the equatorial upwelling, and the Northwest Africa coastal upwelling regions (Wang et al. 2019; Oviatt et al. 2019). Another important physical driver controlling this new sargassum growth in the tropical North Atlantic is the strong convergence of trade (easterly) winds generating the so-called Intertropical Convergence Zone (ITCZ), a narrow zone of deep convection and heavy precipitation. Due to the strong wind convergence, sargassum now accumulates seasonally under the ITCZ, typically between April and September (Johns et al. 2020). Maximum wind convergence and thus maximum accumulation of sargassum occurs in the central equatorial North Atlantic (Fig. 1). A large patch of highly concentrated sargassum is developing there in early spring and moves northward through spring and summer following the ITCZ seasonal meridional migration (Johns et al. 2020). The easterly trade winds also transport Saharan dust, supplying significant amounts of nutrients to the tropical North Atlantic, although its impact on surface nutrient availability in this region is thought to be much less important compared to riverine and upwelling nutrient sources (Wang et al. 2019, supplementary text and Fig. S3).

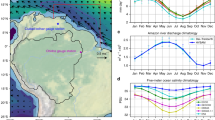
a Climatological mean surface current velocity field (m/s) (from GODAS ocean re-analysis, over 1980–2020) in July. The three gray boxes depict the three sectors partitioning the Central West Atlantic (CWA) where monthly area-integrated alternative floating algae index (AFAI) was calculated: central equatorial North Atlantic (CENA: 38–50°W, 0–10°N), central tropical North Atlantic (CTNA: 38–50°W, 10–20°N), and west tropical North Atlantic (WTNA: 50–63°W, 6–22°N). The red box depicts the North Equatorial Recirculation Region (NERR, 25–50°W, 0–10°N). Dark blue boxes depict areas encompassing the NW Africa coastal upwelling zone (20°W–30°W, 12°N–20°N) and equatorial upwelling zone, respectively, where area-averaged monthly anomalies of various parameters were calculated. Main currents (NEC: North Equatorial Current; NBC: North Brazil Current; nSEC/sSEC: north/south branch of South Equatorial Current, NECC: North Equatorial Counter Current) discussed in the text are depicted (highlighted in yellow). The approximate locations of Amazon and Orinoco river mouths are also depicted. b Climatological mean AFAI (2011–2020) areal fraction (km2) in July
Another important factor regulating nutrient abundance in the tropical North Atlantic is its intense ocean circulation pattern portrayed in Fig. 1a. Equatorial westward currents driven by the prevailing trade (easterly) winds join the North Brazil Current (NBC), a permanent strong northwestward current along the Brazil coast with speeds exceeding 1 m/s. In late spring, the eastward North Equatorial Counter Current (NECC) develops in the approximate zone 5–10°N in the central tropical North Atlantic. The NBC starts to retroflect joining the NECC to form a very efficient re-circulation system in the so-called North Equatorial Recirculation Region (NERR, 0–10°N, 25–50°W) where the largest sargassum biomass peaks are typically observed (Gower et al. 2013; Wang and Hu 2016; Djakoure et al. 2017). Strong re-circulation in this region may act as a nutrient trap in an area where sargassum naturally accumulates within the ITCZ, therefore driving optimal conditions to initiate and sustain a sargassum bloom. Occasionally upwelled nutrients from the Northwest-Africa coastal upwelling region, under enhanced trade winds inducing strong southwestward ocean transport, can reach the central equatorial and tropical North Atlantic to further stimulate sargassum growth (Wang et al. 2019). Moreover, the strong currents in the NERR could further increase sargassum growth, since nutrient uptake by some free floating macroalgae species has been found to be enhanced with increasing current speeds (Gao 1991; Gao and McKinley 1994). Sargassum biomass typically peaks in the NERR in early summer and then spreads both westward and eastward expanding the sargassum belt across the whole tropical North Atlantic (Wang et al. 2019; Johns et al. 2020). During summer and early autumn, the highly concentrated sargassum mats in the NERR spread eastward by the NECC along the ITCZ towards West Africa and westward by the NBC and the North Equatorial Current (NEC). In the Caribbean, the sargassum biomass typically peaks during late summer and sargassum mats there can be traced back to the central tropical and equatorial North Atlantic, several months earlier (Franks et al. 2016; Brooks et al. 2018).
Sargassum blooms in the tropical North Atlantic are highly variable from year to year in terms of total abundance, timing, and spatial extent (Wang et al. 2019). Since the first major bloom event in 2011, the region has experienced extreme interannual variability. A non-bloom year was evidenced in 2013, with very low concentrations across the whole region, while a massive bloom of large spatial extent occurred in 2018 that was sustained throughout most of the year with the total annual abundance being several times the post-2011 long-term mean (Wang et al. 2019). Despite rapid growth in research on sargassum, there remains significant uncertainty with regard to the extent, timing, and relative contribution of physical processes driving major sargassum bloom events in the tropical North Atlantic (Fidai et al. 2020).
Earlier studies of post-2010 sargassum blooms in the tropical Atlantic suggested that increases in riverine, atmospheric deposition and upwelling nutrient fluxes, and higher temperatures could be all factors contributing to sargassum proliferation in this region (Djakoure et al. 2017; Sissini et al. 2017; Oviatt et al. 2019). However, Wang et al. (2019) found a negative correlation between SST and sargassum blooming, suggesting that cool upwelled nutrient-rich waters were associated with increased sargassum biomass. Predominately positive AMO index over the last three decades was linked with weaker trade winds resulting in long-term decreasing Saharan dust transport to the central tropical North Atlantic (Yuan et al. 2016). As a consequence, atmospheric micronutrient fluxes to the ocean surface were reduced, potentially decreasing nutrient availability for sargassum growth. Wang et al. (2019); supplementary text and Fig. S3) showed a low impact of Saharan dust flux interannual variability on sargassum blooms in the tropical North Atlantic over the last decade. More recently, Johns et al. (2020) reported that enhanced mixed layer depth and associated increased upward nutrient fluxes under the sargassum belt were the major factor explaining larger sargassum blooms in recent years.
The very intense tropical Atlantic climate variability results in pronounced seasonal and annual anomalies of wind and ocean circulation patterns that may strongly affect sargassum abundance in the tropical Atlantic. Both winds and currents influence sargassum biomass abundance in two different ways: (a) by concentrating/dispersing or transporting sargassum biomass and (b) by controlling nutrient abundance for sargassum growth. Ocean circulation variability can affect the redistribution and drift of sargassum produced in the central equatorial and tropical North Atlantic (CENA and CTNA) dispersing sargassum mats both westward towards the Caribbean and eastward towards West Africa during summer and early autumn.
In this study, we explore the influence of anomalous winds and ocean currents across the tropical Atlantic associated with regional climate variability in driving and sustaining sargassum blooms in the CWA. Various oceanographic/atmospheric parameters in the wide tropical Atlantic were analyzed to understand their effects and connectivity with the spatiotemporal variability of sargassum biomass found in the CWA. Our main objective is to assess changes in the main physical drivers controlling nutrient availability that could explain the observed excessive interannual variability in sargassum blooms.
The paper is organized as follows. In Section 2, we outline the use of satellite observations, and ocean and atmospheric model re-analysis products. In Section 3, we explore the interannual co-variability of the main physical drivers with the sargassum abundance in the CWA. Particularly we assess how sargassum blooms are regulated by changes in the two major upwelling systems: equatorial upwelling and North-West Africa upwelling. We also focus on the impact of ITCZ location and its interannual modulations on sargassum blooms driven by the major climatic variability modes of the tropical Atlantic. In Section 4, we summarize our main findings, and we underline the uncertainties of our analysis.
2 Data and methods
To determine the spatiotemporal distribution of sargassum abundance over 2010–2020, images of alternative floating algae index (AFAI) were used, provided by the Optical Oceanography Laboratory at the University of South Florida (https://optics.marine.usf.edu), where satellite data products include daily “FA density,” calculated as a mean of the 7 past days (including that day), following the method described in Wang and Hu (2016). Across the Central West Atlantic (CWA), encompassing 38–63°W, 0–22°N, the AFAI data were summed to obtain a daily total area of sargassum since January 2010 (Marsh et al. 2021). Moreover, we separate the CWA into three sectors: central equatorial North Atlantic (CENA: 38–50°W, 0–10°N), central tropical North Atlantic (CTNA: 38–50°W, 10–22°N), and west tropical North Atlantic (WTNA: 50–63°W, 6–22°N) (Fig. 1) in order to reveal areas of initiation and seasonal progression of sargassum blooms and to investigate their interannual variability.
Cloud cover and surface downward shortwave radiation data were used from National Centers for Environmental Prediction-Department of Energy (NCEP-DOE) Re-analysis 2 (https://psl.noaa.gov/data/gridded/data.ncep.reanalysis2.html) spanning 1979–present on a 2.5°× 2.5° grid (Kanamitsu et al. 2002).
Daily observational river discharge data spanning 2010–2019 for the Amazon and Orinoco rivers used here were obtained from the HYBAM (hydro-geochemistry of the Amazonian basin) dataset (https://hybam.obs-mip.fr/data/).
Soil Moisture and Ocean Salinity (SMOS) monthly sea surface salinity (SSS) debiased level 3 fields spanning 2010–2020 at 0.25° × 0.25° resolution (SMOS products dissemination service: https://www.catds.fr/sipad/) were used here to trace changes in the Amazon River outflow and ITCZ location.
Moderate resolution imaging spectroradiometer (MODIS) chlorophyll-a monthly fields (https://oceancolor.gsfc.nasa.gov/atbd/chlor_a/) spanning 2010–2020 at 9 km resolution were used as indicators of changes in nutrient abundance for sargassum growth across the equatorial and tropical North Atlantic, in upwelling regions in particular.
Tropical Rainfall Measuring Mission (TRMM) Rainfall Estimate Level 3 V7 (TRMM Multi-satellite Precipitation Analysis (TMPA)/3B43) monthly fields at 0.25°× 0.25° resolution over 2010–2019 (Greenbelt, MD, Goddard Earth Sciences Data and Information Services Center (GES DISC), Accessed: 15 March 2021, 10.5067/TRMM/TMPA/MONTH/7) were used here as an indicator of changes in ITCZ strength and location.
To assess changes in wind and ocean circulation patterns, NCEP Global Ocean Data Assimilation System (GODAS) re-analysis monthly fields of surface currents, upwelling velocities, mixed layer depth, and wind stress (1980–2020) were used, provided at 1° longitude × 1/3° latitude resolution (https://psl.noaa.gov/data/gridded/data.godas.html). Mixed layer depth is defined in GODAS as the depth where the buoyancy difference with respect to the surface level is equal to 0.03 cm/s2. GODAS is forced by the momentum, heat, and freshwater fluxes from the NCEP-DOE Re-analysis 2.
NOAA Optimum Interpolation SST (v2) monthly fields at 1° × 1° resolution (https://psl.noaa.gov/data/gridded/data.noaa.oisst.v2.html) were used to construct area-averaged monthly timeseries. The above dataset was also used to construct Atlantic Niño (ATL3) index monthly timeseries (monthly area-averaged SST anomalies for the ATL3 region: 3°S–3°N, 0–20°W) over 1982–2020. Monthly timeseries of other two climatic mode indices, the Atlantic Multidecadal Oscillation (AMO) (Trenberth and Shea 2006; https://climatedataguide.ucar.edu/climate-data/atlantic-multi-decadal-oscillation-amo) and the Atlantic Meridional Mode (AMM) (Chiang and Vimont 2004; https://www.aos.wisc.edu/~dvimont/MModes/Data.html) spanning 1980–2020, were used in this study to investigate climate variability impact on sargassum proliferation. Basin-scale warming and cooling of the North Atlantic, and associated environmental changes, have been identified as the Atlantic Multidecadal Oscillation (AMO). The AMO index is calculated as the sea surface temperature anomaly of the North Atlantic (averaged over 0–80°N, 0–60°W), detrended by subtracting the global mean SST anomaly (Trenberth and Shea 2006).
For all datasets, area-averaged monthly anomaly timeseries were constructed by removing the seasonal cycle to investigate co-variability of sargassum abundance with the various physical parameters investigated in this study at seasonal and interannual timescales. In order to assess the physical drivers of sargassum interannual variability, spatial patterns in the CWA and area-averaged property correlations for the three sub-regions considered in this study were calculated between AFAI anomalies and various indicators of nutrient availability, including the ITCZ location relative to nutrient sources, and the intensity of equatorial and West Africa coastal upwelling.
3 Results and discussion
Following an overview of sargassum variability since 2010, we considered contemporaneous variability in nutrient abundance, temperature, and irradiance. With a particular focus on mechanisms influencing the availability of nutrients, we explored the influences of ITCZ shifts and tropical climate modes, the Amazon and Orinoco outflows, northwest Africa coastal upwelling, equatorial upwelling, and ocean circulation intensity in the NERR.
3.1 Sargassum intra-annual and interannual variability over 2010–2020
Recently developed innovative remote sensing based on AFAI images provided strong evidence for the initial growth and establishment of sargassum in the equatorial and tropical North Atlantic and highlighted the large spatiotemporal variability of sargassum blooms since 2011 (Gower et al. 2013; Franks et al. 2016; Wang and Hu 2016; Wang et al. 2019; Johns et al. 2020). Fig. 2a shows monthly averages of sargassum total area across the tropical Central West Atlantic (CWA, encompassing 0–20°N, 38–63°W), integrating AFAI areal fraction, over 2010–2020. After the initial sargassum bloom in 2011, substantial interannual variability unfolds over subsequent years with the largest sargassum blooms developing through 2015, 2018, and 2020 while there was a weak bloom observed in 2016 and no visible bloom in 2013.

Monthly timeseries of total area (km2) of sargassum integrating AFAI areal fraction, 2010–2020 a across Central West Atlantic (CWA), b for three sectors of CWA: central equatorial North Atlantic (CENA, 38–50°W, 0–10°N, red line), central tropical North Atlantic (CTNA, 38–50°W, 10–20°N, blue line), and west tropical North Atlantic (WTNA 50–63°W, 6–20°N, green line)
AFAI image analysis over 2010–2020 also showed large spatial and seasonal variability of sargassum abundance and distribution (Figs. 2 and 3). In most years, there was a “unimodal” seasonal cycle with the bloom typically starting in winter and peaking in early summer in the central equatorial North Atlantic (CENA) (Fig. 2b). Maximum sargassum abundance was found every year post-2011 in and around the NERR peaking in June, except for the non-bloom year of 2013 when very low concentrations were obtained across the whole region and in all seasons. The very strong ocean re-circulation system developing during late spring and summer comprising westward equatorial currents and the NBC which retroflects and joins the NECC (Fig. 4) may transport and potentially trap nutrients within NERR originating from the Amazon outflow and the equatorial upwelling zone (Gower et al. 2013; Wang and Hu 2016; Djakoure et al. 2017). Close to the Amazon River mouth, the equatorial westward flow that transports upwelled nutrients all along the equatorial upwelling zone joins the Amazon River nutrient-rich outflow suggesting increased nutrient transport feeding the central equatorial and tropical North Atlantic. In the permanent equatorial upwelling zone (Fig. 5), upwelling intensity increases through spring, typically peaking in June in the eastern part of the equatorial Atlantic, whereas it continues to increase through summer in the western part of the equatorial Atlantic, peaking there in early autumn (Wang and Hu 2017). Amazon outflow is ascending throughout spring and early summer typically peaking in June while Orinoco outflow peaks in late summer. Therefore, seasonal patterns of these nutrient-rich flows roughly coincide with the sargassum bloom developing in spring and summer in the tropical North Atlantic with sargassum biomass typically peaking in June in the NERR, and in late summer in the western tropical Atlantic.

AFAI areal fraction climatological mean (km2) over 2010–2020 for a winter (DJF), b spring (MAM), c summer (JJA), and d autumn (SON) (boreal seasons)

GODAS climatological mean surface current velocity (m/s) over 2010–2020 for a winter (DJF), b spring (MAM), c summer (JJA), and d autumn (SON)

GODAS climatological mean upwelling velocity at 40m depth (m/day) over 2010–2020 for a winter (DJF), b spring (MAM), c summer (JJA), and d autumn (SON) (boreal seasons)
The ITCZ convergent winds accumulate large sargassum biomass in the NERR that spreads both westward to the Caribbean via the NBC along the South American coastline, and via NEC further offshore, and transported via the Caribbean Current as far as the coasts of Central America, and eastward towards West Africa via the NECC (Wang et al. 2019; Johns et al. 2020). In CTNA (north of 10°N), the sargassum bloom was generally much less pronounced peaking approximately a month later compared to CENA (see Fig. 2b). This is probably due to the convergent winds keeping the sargassum mats under the ITCZ, which is located south of 10°N, but also to the presence of the NECC partly blocking northward transport of sargassum originated in the NERR. The strong westward NECC also probably partly blocks nutrient northward transport from the equatorial upwelling region and the Amazon outflow inhibiting sargassum growth in the CTNA. In the western tropical North Atlantic (WTNA), sargassum abundance is peaking with a lag of approximately 2 months with respect to CENA towards the end of summer (see Fig. 2b) whereas sargassum growth is also locally stimulated by nutrient inputs from the Orinoco river outflow which typically peaks in August. After the bloom peak, sargassum concentrations gradually decrease in the whole tropical region throughout autumn reaching very low levels during winter. However, there are years, notably in 2015 and 2018, when large sargassum blooms were sustained almost throughout the whole year and the sargassum belt was extended further north of 10°N in the central tropical North Atlantic with the occurrence there of exceptionally large winter blooms (Fig. 2b). High concentrated sargassum mats are then transported via the NEC to the west tropical North Atlantic with sargassum abundance peaking there, with a lag of ~1 month, in early spring (Fig. 2b). Especially in 2018, this pattern results in two distinct large sargassum biomass peaks in both CTNA and WTNA in February/March and November/December, respectively.
Monthly anomaly timeseries of AFAI area averaged over the three sub-regions considered here showed strong positive linear trends (statistically significant at the 95% confidence interval) over the 2010–2020 period (see Table 1), driven by much larger sargassum blooms since the middle of the decade, in 2015, 2018, and 2020, in particular. Seasonal trends were also statistically significant in CENA and WTNA for all seasons except from autumn (Table 1). These trends suggest that sargassum proliferation is now a well-established annually recurrent feature with increasing intensity in the tropical Atlantic.
In the following sections, we explore the physical drivers of intra-annual and interannual variations of sargassum abundance in CWA with regard to the three major limiting factors of sargassum growth, namely nutrient abundance, temperature, and light.
3.2 Nutrient abundance
Nutrients, especially the macronutrients nitrate and phosphate, constitute an important limiting factor of sargassum growth in the oligotrophic North Atlantic (Lapointe 1986, 1995; Smetacek and Zingone 2013). Recent studies demonstrated that nutrient availability is the major factor controlling interannual variability of sargassum blooms in the central tropical North Atlantic (Wang et al. 2019; Johns et al. 2020). However, nutrients are by far the most under-sampled properties over both time and space compared to the other two limiting factors, i.e., surface temperature and irradiance, which are routinely estimated with high spatiotemporal coverage over the satellite era. Currently, nutrient concentration spatiotemporal coverage is too poor to assess the interannual variability of its distribution in the tropical Atlantic while there are also large uncertainties associated with plankton ecosystem and biogeochemical models providing nutrient data. Moreover, there are no sufficient measurements to properly assess variations in external nutrient inputs, i.e., atmospheric deposition and riverine fluxes. Therefore, there is a strong need for finding suitable indicators of nutrient availability for sargassum growth. In particular, SMOS SSS anomalies are used here as an indicator of the intensity and pathway of the Amazon outflow (see Supplement section S1), a major source of nutrients in the tropical North Atlantic. Also, as in Wang et al. (2019), satellite-derived surface chlorophyll data were used here (in Sections 3.2.2 and 3.2.3) as an indicator of increased nutrient concentration which is closely linked to phytoplankton growth, especially in oligotrophic tropical regions, where under warm and high light availability conditions nutrient abundance is the main limiting factor of primary production. Ocean currents, upwelling velocities, and mixed layer depth from ocean re-analyses are used as indicators of enhanced nutrient availability in the surface layer. Satellite-derived rainfall data were also used together with SMOS SSS to infer meridional shifts of the ITCZ towards or away from the main equatorial nutrient sources, namely the Amazon and Orinoco plumes and the equatorial upwelling zone. On the other hand, the larger source of uncertainty in our analysis arises from the lack of nutrient in situ observations and the use instead of proxies for nutrient variability. The short 10-year timeseries of the sargassum years in the tropical North Atlantic and the use of indicators of nutrient abundance strongly constrain a meaningful statistical analysis to quantify the relative contributions of physical drivers on sargassum blooms. Such analysis is further complicated by the fact that physical drivers act on nutrient distribution and subsequently on sargassum abundance at a highly non-linear fashion, simultaneously or at different periods of the year.
In the following sections, we explore in detail links between the interannual variability of sargassum abundance in CWA and the changes in physical drivers controlling primarily macronutrient availability for sargassum growth in the wider tropical North Atlantic, namely riverine inputs, ocean circulation and mixing, equatorial upwelling, Northwest Africa coastal upwelling, and the ITCZ location.
3.2.1 Riverine nutrient inputs
The Amazon River outflow showed low interannual variability and co-variability with sargassum biomass peaks over the last 10 years, except from the pronounced anomaly of reduced outflow in 2016 which could negatively affect the relatively weak sargassum bloom over that year (see Sup. section S1 and Fig. S1). Orinoco river outflow data revealed a strong positive outflow anomaly during summer 2018 when the largest sargassum abundance was observed in the western tropical Atlantic, whereas a strong negative anomaly was observed in summer 2013 when sargassum abundance was minimal (Fig. S2). However, again the co-variability of the Orinoco outflow with sargassum biomass peaks in WTNA was relatively weak. Our analysis of monthly/seasonal anomaly timeseries of Amazon and Orinoco outflow and their sea surface salinity (SSS) imprints showed low (and not statistically significant) correlations with area-averaged AFAI in CENA, and WTNA, respectively, suggesting that river nutrient input was not a significant factor that could explain the large interannual sargassum variability over the last 10 years. Maximum correlations (but still not statistically significant) were found between spring anomalies of area-averaged AFAI in CENA and Amazon (in-land) outflow (r= 0.24, p>0.1) and SSS close to the Amazon mouth (r= −0.39, p>0.1).
3.2.2 Ocean circulation and mixing
More intense currents may induce stronger transport of nutrient-rich waters in the NERR originating from both the Amazon outflow and the equatorial upwelling zone. Also, higher nutrient uptake has been associated with higher current speeds in some macroalgae species including sargassum (Gao 1991; Gao and McKinley 1994). We explored the impact of NBC and NECC re-circulation system current intensity on sargassum interannual variability (See Sup. section S2 and Fig. S3). Both 2015 and 2018 spring/summer sargassum bloom periods were associated with an enhanced current intensity, consistent with stronger sargassum proliferation in CENA. However, low and not significant correlations were found between seasonal (spring/summer) anomalies of area-averaged AFAI in CENA and current intensity in the re-circulation region over 2010–2020 (spring: r=0.16, p>0.1; summer: r=0.33, p>0.1). Therefore, although enhanced current intensity in NBC and NECC re-circulation system may have contributed to sargassum biomass peaks in some years, it was not a major contributor to sargassum interannual variability in CENA. Moreover, we found no significant link between changes in the recurrent sargassum blooms initiated in CENA with local changes in mixing or upwelling during spring/summer (see Sup. section S3). Monthly and seasonal anomalies of area-averaged AFAI were not correlated with local mixed layer depth or upwelling velocity anomalies in CENA over 2010–2020. This was probably due to low nutrient entrainment in the mixed layer during spring/summer as the upper layer is relatively isolated from the deeper cold and nutrient-rich waters by increased freshwater inputs from the Amazon River and precipitation in the ITCZ (Saha et al. 2021).
3.2.3 ITCZ shifts and tropical climate modes
During April–August, sargassum naturally accumulates under the ITCZ, where the prevailing trade (easterly) winds converge, strongly affecting the spatiotemporal variability of sargassum biomass (Johns et al. 2020). There is a seasonal northward migration of the ITCZ following a sea surface warming pattern that shifts from ~2–3°N in late winter/early spring to ~9°N in late summer/early autumn. Maximum sargassum growth rate in CENA is during spring (Wang et al. 2019), when the ITCZ is relatively closer to nutrient-rich waters originated from the Amazon outflow and the equatorial upwelling region. During summer, the ITCZ migrates further northward, closer to the Orinoco plume, directly affecting sargassum biomass in WTNA, but further away from the nutrient sources of the Amazon River and equatorial upwelling region. This could explain the strong decline of sargassum biomass in CENA after its peak in June.
There is strong interannual variability in the ITCZ strength and meridional location closely linked to the two major climatic modes of natural variability in the tropical Atlantic, the Atlantic Meridional Mode (AMM), and the Atlantic Niño (see Sup. section S4). Both modes are characterized by anomalous trade wind patterns inducing meridional shifts of the ITCZ, especially during spring and early summer, when the sargassum bloom occurs.
In contrast with predominately positive AMM over the last few decades, spring/summer seasons of 2014, 2015, and especially 2018 were characterized by strongly negative AMM index (Fig. 6a). An AMM negative phase typically drives a southward shift of the ITCZ that could indirectly control nutrient availability during the typical sargassum bloom in NERR over these years (Fig. 6c). As the sargassum mats naturally aggregate under the ITCZ, they could move southward towards the nutrient-rich Amazon plume and equatorial upwelling zone, enhancing sargassum growth rate. On the contrary, a positive AMM phase in 2013 could lead to a northward shift of the ITCZ and subsequently transport of the aggregated sargassum mats away from these nutrient sources. The meridional shifts of the ITCZ were deduced here by precipitation and surface salinity anomaly patterns in NERR. During spring 2018, both variables show very strong meridional gradients across the zonal ITCZ band, of increasing rainfall and decreasing surface salinity, respectively, towards the equator, indicating an anomalous southward shift of the ITCZ (Fig. 7). In contrast, these anomaly patterns were reversed over 2013 indicating an anomalous northward shift of the ITCZ.

Monthly timeseries over 2010–2020 of a AMM index, b ATL3 index (SST anomaly averaged over 3°S–3°N, 0–20°W), c AFAI (% coverage) averaged over central equatorial North Atlantic (CENA), and d SST anomaly averaged over CENA

Monthly anomalies of TRMM precipitation (mm/day) (a, c), SMOS SSS (psu) (b, d), AFAI areal fraction (e, f) over May 2018 (a, c, e) and May 2013 (b, d, f), respectively
The spatial seasonal correlation pattern between local AFAI and AMM index during April–August (when sargassum mats are concentrated by the convergent winds under the ITCZ in this region) shows coherent strong (negative) correlations (r~ −0.6 to −0.9, p<0.05) throughout the CENA and WTNA regions (Fig. 8a). Annual timeseries (2010–2020) of AMM index and area-averaged AFAI in CENA over April–August are highly (negatively) correlated (r= −0.78, p<0.01), suggesting a strong control of AMM on sargassum proliferation in CENA during the typical spring/summer bloom period (Fig. 8b).

a Correlation spatial pattern in CWA between April–August mean AFAI areal fraction and AMM index during 2010–2020. Regions where the correlation is not significant at the 95 % confidence interval are stippled. b April–August mean AFAI areal fraction averaged over the central equatorial North Atlantic (CENA) and AMM index during 2010–2020
Although correlation of area-averaged AFAI over April–August with positive Atlantic Niño index was much less and not statistically significant over 2010–2020 (r=0.23, p>0.1), the combination of a positive phase of Atlantic Niño with a negative phase of AMM could further exacerbate sargassum blooms. In particular, the negative AMM was combined with an Atlantic Niño in early 2018 (see Fig. 6b) which may have further enhanced the southward migration of the ITCZ stimulating the sargassum bloom in CENA during spring and early summer. Moreover, with anomalous high current intensity within NERR during summer 2018 (see Fig. S3b) and as NBC is retroflected northward, nutrient-rich waters originated from the Amazon River plume and the equatorial upwelling zone could more easily reach an anomalously southward shifted ITCZ location even later in the season.
The following 2 years (2019, 2020) were also characterized by a positive phase of Atlantic Niño and a negative phase of AMM. However, AMM amplitudes were relatively low and therefore southward migration of ITCZ and subsequent stimulation of the sargassum spring blooms were probably less important. A quite large but short-lived Atlantic Niño developed earlier than usual in late winter 2020, the largest positive winter anomaly over the 2010–2020 record, and may have contributed to the relatively large spring/summer bloom of 2020 in CENA.
AMM index and Atlantic Niño (averaged over April–August) annual timeseries over 2010–2020 show statistically significant linear trends (AMM: ~0.4/year, r=0.53, p<0.1; Atl. Niño: ~0.04/year, r=0.62, p<0.1) towards negative and positive phases, respectively, over the last decade, both associated with a southward displacement of the ITCZ (see Sup. Section 4). Therefore, increasing intensity of sargassum blooms over 2010–2020 could be partially driven by increasing nutrient availability due to decadal natural tropical climate variability which controls the wind pattern and associated ITCZ location and upwelling intensity. This trend could be also superimposed on an increasing trend of anthropogenic river nutrient inputs over the last decade (Wang et al. 2019), further exacerbating sargassum blooms. However, both processes are difficult to be established here as drivers of the long-term increasing sargassum bloom intensity as nutrient spatiotemporal observational coverage is very poor in this region and the 10-year period of “tropical sargassum years” is too short to study the impact of decadal variations in tropical climate modes on sargassum abundance.
3.2.4 Northwest Africa coastal upwelling
A remote nutrient source for the tropical North Atlantic is the Northwest Africa coastal upwelling zone (Wang et al. 2019). The tropical region is mainly affected by waters originated in the southern part of the upwelling region, the Mauritania-Senegalese upwelling zone, located approximately between 12°N and 19°N (Cropper et al. 2014). In contrast with the permanent Northwest Africa upwelling zone located further north (~21°N–26°N) where upwelling intensity peaks in summer, upwelling in the Mauritania-Senegalese zone occurs in winter months and vanishes during the summer months following the seasonal migration of the trade winds. During late winter, chlorophyll-a maps (Fig. 9) suggest a longitudinal extension of the influence of the upwelling zone roughly varying between 25°W and 30°W. Under stronger trade winds, upwelling is intensified and upwelled nutrients can reach the central tropical and equatorial North Atlantic west of 30° W via a stronger NEC, favorably pre-conditioning the spring/summer sargassum blooms (Wang et al. 2019).

MODIS chlorophyll-a (mg/m3) monthly mean maps of the tropical North Atlantic (a, b) and AFAI areal fraction (km2) over Central West Atlantic (CWA) (c, d) for February 2013 (a, c) and February 2018 (b, d)
Strongly negative AMM phases over 2015 and 2018 were associated with stronger trade (easterly) winds in the tropical and subtropical eastern North Atlantic enhancing both the coastal upwelling and NEC, thus driving stronger southwestward nutrient transport. As a result, more nutrient-rich waters could reach the central tropical North Atlantic sustaining larger sargassum blooms there. Moreover, part of the remnant sargassum population (from December to February, Johns et al. 2020) is located close to the area of higher nutrient input due to the winter Mauritania-Senegalese upwelling. Figure 10a shows monthly mean anomalies of chlorophyll-a averaged over an area encompassing the upwelling region and extended to 30°W. Strong positive chlorophyll monthly anomalies were obtained in 2015 and 2018 sustained through the spring and summer months of sargassum proliferation. In contrast, negative anomalies were obtained during the non-bloom year of 2013 and the weak bloom of 2016. In particular, ocean color images indicate much larger chlorophyll concentrations extending from the Northwest Africa upwelling zone to the central tropical North Atlantic during late winter 2018 (see Fig. 9). This was in contrast with the no-bloom year of 2013 where chlorophyll levels were lower and the chlorophyll high-concentration patch was confined close to the upwelling region, being much less extended to the west compared to 2018. A robust lag-correlation was found here between January and March mean chlorophyll-a averaged in the extended upwelling region considered here (20°W–30°W, 12°N–20°N) with averaged AFAI in both CENA and WTNA regions during the spring/summer bloom period over 2010–2020 (CENA: r=0.65, p<0.05; WTNA: r=0.61, p<0.05) (Fig. 10b), suggesting that West Africa coastal upwelling could be a key process controlling variability of sargassum blooms in accordance with the studies of Wang et al. (2019) and Johns et al. (2020). The spatial correlation pattern shows coherent broad-scale strong correlations (r>0.6, p<0.05) within the eastern part of CENA and large parts of CTNA as well as northern parts of WTNA (north of 15°N) (Fig. 10c). It is interesting to note that during typical spring/early summer bloom conditions sargassum biomass peaks are much less pronounced in these regions which are further away from the typical equatorial nutrient sources (i.e., Amazon plume and equatorial upwelling region) as well as away from the ITCZ where sargassum aggregates. Therefore, our results suggest that anomalously strong West Africa upwelling and southeastward nutrient transport via the NEC, both related with enhanced trade (easterly) winds, can further stimulate sargassum blooms in these areas.

a MODIS chlorophyll-a monthly anomalies (mg/m3) for an area (20°W–30°W, 12°N–20°N) encompassing (but extended well offshore of) the Mauritania-Senegalese coastal upwelling zone. b Spring–summer mean AFAI areal fraction averaged over the central equatorial North Atlantic (CENA) and January–March mean anomalies of chlorophyll-a (mg/m3) averaged over the extended upwelling region (20°W–30°W, 12°N–20°N). c Correlation spatial pattern in CWA between April–August mean AFAI areal fraction and January–March mean anomaly of chlorophyll-a (mg/m3) averaged over the extended upwelling region during 2010–2020. Regions where the correlation is not significant at the 95% confidence interval are stippled
3.2.5 Equatorial upwelling and winter blooms
We focus here on precursors of the sargassum winter blooms in 2015 and 2018. In contrast with other post-2011 sargassum bloom seasons, important secondary blooms occurred, starting late autumn 2014 and 2017 and typically peaking in late winter of (early) 2015 and 2018 (see Fig. 2b). These blooms take place in the central tropical North Atlantic at ~10°N–15°N, that is north of NERR where the recurrent spring/summer sargassum blooms are typically observed. During late autumn and winter, sargassum abundance is much less affected by the ITCZ location relative to nutrient sources as sargassum accumulation under the ITCZ is minimal over this period. The ITCZ and its seasonal meridional migration typically affect sargassum accumulation between roughly April and August (Johns et al. 2020). Also, during late autumn and winter, the Amazon outflow is much lower, i.e., it is ~60% less in December with respect to its peak in June (Wang et al. 2019), and together with weak NBC retroflection this may result in reduced nutrient availability for sargassum in the NERR. The NECC develops in late spring and strengthens during summer forming a strong zonal westward flow that blocks northward transport of nutrient-rich waters from the equatorial upwelling zone. In autumn, the NECC moves together with the ITCZ further north and becomes more unstable facilitating exchange with the northern part of the central tropical Atlantic region (see Fig. 4). In winter, the NECC gradually weakens and finally vanishes resulting in a further enhancement of northward transport from the nutrient-rich equatorial upwelling region to the central tropical North Atlantic where these secondary winter blooms occur. The winter blooms peaking late winter 2015 and 2018 seem to be linked to exceptionally strong wind-driven equatorial upwelling and associated northwestward transport of nutrient-rich equatorial waters feeding the central tropical North Atlantic in the preceding autumn and early winter. Figure 11 shows monthly anomalies of equatorial integrated westward transport, mixed layer depth (3°S–3°N, 0°–40°W), and upwelling velocity (3°S–3°N, 0°–20°W) over 2011–2020. The largest positive anomalies of westward transport and upwelling velocity were observed during late autumn and winter 2014 and 2017 preceding the large winter blooms of 2015 and 2018. Increased wind-induced mixed layer depth was also obtained along the pathway of the enhanced westward current during these periods (Fig. 11b). Especially in autumn 2017, there was a very strong increase in the intensity of westward equatorial currents along the whole equatorial upwelling zone, i.e., exceeding 80% increase with respect to the long-term seasonal mean (Fig. 12a) driven by stronger easterly winds along the Equator (Fig. 12b). Increased wind-induced mixed layer depth was also obtained along the pathway of the enhanced westward currents (not shown) whereas a large increase in upwelling velocities was observed in the eastern part of the equatorial upwelling zone (Fig. 12c). This pattern was also supported by satellite ocean color observations indicating much higher concentration chlorophyll-a patches during autumn 2017 and winter 2018 that are anomalously extended towards the northwest part of the equatorial region where nutrient-rich waters can more easily reach the central tropical North Atlantic via the intense ocean circulation pattern (Fig. 12d). Very strong correlations were obtained here between winter mean AFAI integrated over CTNA and equatorial mean westward current intensity (r=0.83, p<0.01) and upwelling velocity (r=0.78, p<0.01) during autumn and early winter over the 2010–2020 period (Fig. 13a, b). The spatial correlation patterns show statistically significant strong positive local correlations between both physical parameters and AFAI roughly between 10°N and 15°N within CTNA but very low and not significant correlations in most parts of CENA (Fig. 13c, d). It is interesting to note that upwelling velocity anomalies in the western part of the equatorial upwelling and the NERR were small (see Fig. 12c) indicating that upwelled nutrients reaching the central tropical North Atlantic were probably mainly originated from the eastern equatorial upwelling region. High sargassum concentrations from these exceptional winter blooms were transported by the NEC to the western tropical Atlantic with a lag of ~ 1 month (see Fig. 2b) but they probably affect much less the sargassum bloom evolution in the NERR and the equatorial region south of 10°N as currents move northwestward away from this region (see Fig. 4).

GODAS monthly anomalies (2010–2020) averaged over parts of the equatorial upwelling area of a westward current velocity (0°–40°W, 3°S–3°N), b mixed layer depth (0°–40°W, 3°S–3°N), and c vertical (upwelling) velocity at 80m depth (0°–20°W, 3°S–3°N)

Anomaly maps for autumn (Sep–Nov) 2017: a GODAS surface current velocity (m/s), b GODAS surface wind stress anomaly (N/m2), c GODAS upwelling velocity at 80m depth, d MODIS Chl-a concentration (mg/m3), e AFAI areal fraction

Timeseries of winter mean AFAI areal fraction averaged over the central tropical North Atlantic (CTNA) and equatorial (a) zonal (westward) surface current velocity (m/s) (averaged over 0°–40°W, 3°S–3°N), and upwelling velocity (0°–20°W, 3°S–3°N) (b), over the preceding Autumn, during 2010–2020. Associated correlation spatial patterns in CWA over 2010–2020 between AFAI areal fraction and surface zonal current velocity (c) and upwelling velocity during 2010–2020 (d). Regions where the correlation is not significant at the 95% confidence interval are stippled
Sargassum remnant concentrations from the previous proliferation season are also found to be an important factor by maintaining sufficient amounts of sargassum to set favorable initial conditions for the next bloom season (Wang et al. 2019; Johns et al. 2020). After the massive winter and spring/summer sargassum blooms in 2018, remnant concentrations remained at relatively high levels across large parts of the central and Northwest tropical Atlantic throughout late autumn and winter facilitating the spring/summer bloom of 2019 (see Fig. 2b).
Wind-driven westward transport along the equatorial upwelling region was also significantly enhanced through spring and summer of 2020, indicating increased nutrient availability that probably contributed to the relatively large sargassum bloom of that year. However, as this occurs much later in the season, when NBC retroflection is strong and the NECC is developed, nutrient-rich water transport was partially blocked at approximately 10°N mainly affecting the typical recurrent sargassum spring/summer bloom in the NERR (see Fig. 2b).
3.3 Temperature
SST has been increasing over the last 40 years in the tropical North Atlantic, at a rate of ~0.018 °C/year in the CWA (Fig. 14a). Natural climate variability seems to have accentuated the effects of global warming on the ocean surface in this region. As expected, high statistically significant correlations were found between CWA SST anomaly timeseries and both SST-derived AMO (r ~ 0.8, p<0.01) and AMM (r ~ 0.7, p<0.01) indices (Fig. 14b, c). AMO and AMM are also correlated (r~0.6, p<0.05) as they are both associated with similar wind and SST anomaly patterns in the tropical North Atlantic. Surface warming anomalies after the mid-2000s were consistent with continuously positive AMO phase and predominantly positive AMM phase. During positive AMO and AMM phases, the trade winds become weaker, resulting in reduced (southwestward) Saharan dust transport and cloud fraction in the tropical North Atlantic. Both these responses result in increasing downwelling shortwave radiation contributing to surface warming in this region (Yuan et al. 2016). However, the large sargassum growth events in 2015 and especially 2018 were mainly associated with negative phases of the AMM, while the non-bloom year of 2013 and the relatively weak bloom of 2016 both coincided with positive phases of the AMM and positive SST anomalies.

Monthly timeseries (1982–2020) of a SST anomaly averaged over Central West Atlantic (CWA) (from NOAA OISST). The linear trend (0.018 °C/year, p<0.01) is also depicted, b AMO index, c AMM index
These results indicate that warming is not driving the large sargassum growth peaks over the last decade. Laboratory temperature sensitivity experiments investigating the growth of S. natans, under nutrient-rich conditions (Hanisak and Samuel 1987), showed that the growth rate is increased with a step change from 18 to 24°C and then slightly decreased (i.e., by less than 20%) when temperature increased with a step change from 24 to 30°C indicating that the optimal temperature for growth is between these two values. This range almost coincides with the range of seasonal variation of SST in the tropical North Atlantic which makes it difficult to assess the optimal temperature and more generally the temperature dependence for sargassum growth. Typical high SST values (>28°C) encountered during summer and early autumn in the central tropical North Atlantic probably slightly inhibit sargassum growth (i.e., according to Hanisak and Samuel (1987) study), although sargassum growth dependency on temperature could be stronger under oligotrophic conditions. Maximum monthly local SST anomalies in the NERR over 2010–2020 (not shown) are relatively small (~1–2°C) (see also Wang et al. 2019) considering the low sargassum growth dependency on temperature, and certainly cannot explain the very strong interannual variability in sargassum abundance, and especially the extreme difference between the no-bloom year of 2013 and the massive sargassum blooms of 2015 and 2018. Wang et al. (2019) found a significant negative correlation between SST and sargassum growth rate in central-west tropical North Atlantic during November–December but suggested that surface cooling could be associated with higher nutrient concentrations from upwelling or river inputs enhancing sargassum growth.
The large sargassum growth events of 2015 and 2018 were associated with a strongly negative AMM phase which induces both a surface cooling and a southward displacement of the ITCZ, but our results suggest that it is the latter and not the former that drives the sargassum proliferation in the central tropical North Atlantic by transporting/concentrating sargassum in an area with higher nutrient availability mainly related to large nutrient fluxes from the Amazon River and equatorial upwelling region. Negative AMM may also increase wind-induced turbulence that deepens the surface mixed layer (that can be seen as cooler SST), which bring nutrients towards the mixed layer by the turbulent vertical entrainment of the thermocline. Moreover, large growth rates especially during the exceptionally large sargassum proliferation through 2018 were associated with stronger both equatorial and Northwest Africa upwelling that were also associated with surface cooling.
3.4 Irradiance
Shortwave radiation is increasing with predominantly positive AMO over the last few decades (Fig. 15), partially driving the SST warming trend (Yuan et al. 2016) and providing higher light availability for sargassum. Our analysis of NCEP/NCAR re-analysis outputs shows a robust positive trend in shortwave radiation over the last 40 years (2.5 W/m2/decade, p<0.01) driven by a negative trend in cloud cover (−1.2%/decade, p<0.01) with the two timeseries being highly anti-correlated (r~ −0.9, p<0.01). Post-2010, over the tropical sargassum proliferation era, positive monthly anomalies of shortwave radiation were observed almost throughout this period with relatively small variations from year to year (Fig. 15). This is consistent with the study of Wang et al. (2019) showing no significant changes in light availability (photosynthetically active radiation) across the tropical North Atlantic during recent years.

Monthly anomaly timeseries (1979–2020) of a surface downward shortwave radiation and b total cloud cover (%), averaged over Central West Atlantic. Linear trends (statistically significant at the 99% confidence interval) are also depicted. From NCEP/DOE Re-analysis 2
Based on laboratory experiments, the growth light saturation level for sargassum natans (at 24°C and under nutrient-rich conditions) is found to be in the range of ~43–65 Wm-2 (Hanisak and Samuel 1987). Although again growth dependency on light availability could be stronger under oligotrophic conditions, this range is well below the typical values of irradiance in the tropical Atlantic almost through the whole year (Thandlam and Rahaman 2019). Therefore, increased irradiance does not seem to have a significant impact on sargassum growth in the tropical North Atlantic. Our analysis suggests that variations in surface temperature or irradiance directly affecting the physiology of sargassum play a minor role in the interannual variability of sargassum abundance in the CWA.
4 Concluding remarks
Our results suggest that changes in the intensity of sargassum blooms from one year to another are mainly driven by anomaly patterns of regional winds and currents controlling nutrient abundance and concentrating/dispersing or transporting sargassum biomass, rather than changes in surface warming, light, or river outflows. Anomalies in the trade wind and ocean circulation patterns affecting sargassum proliferation are associated with AMM and Atlantic Niño, the two major ocean/atmosphere coupled climate variability modes of the tropical Atlantic.
In particular, in 2018 when the largest sargassum bloom was observed, a strong negative phase of AMM and a positive Atlantic Niño event were probably combined to produce a pronounced southward displacement of the ITCZ as inferred by satellite-derived rainfall and surface salinity data. During spring when the ITCZ is typically located between 3 and 5° N, a southward shift of the ITCZ can move aggregated sargassum mats much closer to the nutrient-rich waters originated from the Amazon River plume and the equatorial upwelling zone, further stimulating the sargassum bloom in CENA. During summer, the ITCZ typically migrates north of 7°N, away from the nutrient sources of the Amazon River and equatorial upwelling region, and closer to the Orinoco plume, directly affecting the sargassum biomass in WTNA. However, with more intense re-circulation and NBC retroflection in NERR during summer 2018, nutrient-rich waters originated from the Amazon River plume and the equatorial upwelling zone could also potentially reach an anomalously southward shifted ITCZ location within CENA even later in the season. This pattern probably strongly contributed in driving and sustaining the exceptionally large sargassum bloom over 2018.
Larger sargassum blooms were also associated with negative AMM phases due to highly anomalous stronger trade winds which drive enhanced Northwest Africa upwelling and stronger southwestward nutrient transport to the central tropical North Atlantic, as inferred by satellite ocean color observations. Moreover, in contrast with most years where typical spring/summer blooms are initiated in the NERR and then spread through the whole tropical region, important secondary winter blooms took place early in 2015 and 2018. These blooms occurred further north of NERR, associated with anomalously strong wind-driven equatorial upwelling and westward equatorial flow in the previous autumn and early winter, an important precursor for nutrient influx to the central tropical North Atlantic.
Despite the relatively short 10-year period of sargassum proliferation in the tropical North Atlantic, very robust strong correlations at interannual scales were obtained here between area-averaged sargassum abundance and indicators of the three main physical drivers found here, namely the West Africa coastal upwelling and ITCZ shifts driven by tropical climate variability controlling the typical spring/summer sargassum blooms, and the equatorial upwelling controlling sporadic winter sargassum blooms. However, we need to stress again here the large uncertainties in our analysis mainly associated with the lack of nutrient observations and the use instead of indicators of nutrient availability. Moreover, the large number of physical drivers affecting the sargassum accumulation, drift, and growth in a highly non-linear fashion and at different spatiotemporal scales makes it particularly difficult to quantify the relative contributions of the various drivers of the interannual variations of sargassum abundance.
More research is also needed to determine the growth dependency on the various limiting factors (nutrient uptake, temperature, and irradiance) of the sargassum species currently proliferating in the tropical North Atlantic, as well as the sargassum seaweed competition with phytoplankton for nutrient uptake, especially under oligotrophic conditions.
Although AFAI is commonly used to assess spatiotemporal variations of sargassum (Wang et al. 2019; Johns et al. 2020), more studies are needed to properly validate this dataset with in situ observations. There is still large uncertainty in determining sargassum abundance as AFAI may only detect it after surface aggregations become large enough, while it is difficult to distinguish it from other floating materials such as Trichodesmium mats (Wang and Hu 2016). Also, better satellite-derived data of sargassum biomass/coverage is needed, especially over near-perennial cloudy regions such as the eastern equatorial Atlantic, where additional sargassum bloom distribution and drift are still poorly defined for a large part of the year (Johnson et al. 2020). Another source of uncertainty concerns the use of a relatively coarse ocean model re-analysis (GODAS) to assess anomalies in ocean physical processes. GODAS spatial resolution is not able to capture fine-scale mesoscale dynamics that could affect nutrient distribution and its impact on sargassum growth.
AFAI data clearly show an increasing intensity of sargassum blooms throughout the CWA over 2010–2020, probably at least partially due to decadal variations of the tropical climate modes. The occurrence of recurrent sargassum blooms in the tropical North Atlantic over the last decade with increasing intensity over recent years indicates that sargassum is “here to stay.” If deforestation and fertilizer consumption in Brazil continue at the present accelerating rate, continuous increase of Amazon River nutrient inputs will likely exacerbate sargassum blooms in the tropical North Atlantic in the near future with strong negative implications for coastal populations in West Africa and across the Caribbean. A considerable effort has been made in the last few years to predict sargassum drift in the tropical Atlantic at seasonal timescales using various methods such as statistical analysis of satellite observations (Wang and Hu 2017), climatological drifter data (Johnson et al. 2020), coupled physical/biogeochemical ocean models (Jouanno et al. 2021), and ensembles of ocean model hindcasts (Marsh et al. 2021). With improved understanding of physical processes controlling sargassum growth and drift in the region, we will be better able to predict long-term changes in the prevalence and distribution of sargassum. Our results underline the importance of tropical Atlantic climatic modes in controlling sargassum blooms via changes in the trade winds and associated ocean circulation patterns. Therefore, based on the seasonal predictability of tropical Atlantic climate modes, we may improve and refine our models to establish a reliable seasonal forecast system of sargassum growth and drift across the region which is of pivotal importance for coastal populations in order to tackle the ongoing challenges of seasonal sargassum inundation events.
Data Availability
All data and materials used in this manuscript are freely available and comply with field standards.
Code Availability
Not applicable
References
Brooks MT, Coles VJ, Hood RR, Gower JFR (2018) Factors controlling the seasonal distribution of pelagic Sargassum. Mar Ecol Prog Ser 599:1–18. https://doi.org/10.3354/meps12646
Chiang JCH, Vimont DJ (2004) Analagous meridional modes of atmosphere-ocean variability in the tropical Pacific and tropical Atlantic. J Clim 17(21):4143–4158
Cropper TE, Hanna E, Bigg GR (2014) Spatial and temporal seasonal trends in coastal upwelling off Northwest Africa, 1981–2012. Deep-Sea Res I Oceanogr Res Pap 86:94–111. https://doi.org/10.1016/j.dsr.2014.01.007
Davis D, Simister R, Campbell S, Marston M, Bose S, McQueen-Mason SJ, Gomez LD, Gallimore WA, Tonon T (2021) Biomass composition of the golden tide pelagic seaweeds Sargassum fluitans and S. natans (morphotypes I and VIII) to inform valorisation pathways. Sci Total Environ 762:143134
Djakoure S, Araujo M, Hounsou-Gbo A, Noriega C, Bourles B (2017) On the potential causes of the recent Pelagic Sargassum blooms events in the tropical North Atlantic Ocean. Biogeosci Discuss. https://doi.org/10.5194/bg-2017-346
Fidai YA, Dash J, Tompkins EL, Tonon T (2020) A systematic review of floating and beach landing records of Sargassum beyond the Sargasso Sea. Environ Res Commun 2:122001
Franks JS, Johnson DR, Ko D-S, Sanchez-Rubio G, Hendon R, Lay M (2012) Unprecedented influx of pelagic Sargassum along Caribbean island coastlines during summer 2011. Proc Gulf Caribb Fish Inst 64:6–8
Franks JS, Johnson DR, Ko DS (2016) Pelagic Sargassum in the tropical North Atlantic. Gulf Caribb Res 27:SC6–S11. https://doi.org/10.18785/gcr.2701.08
Gao K (1991) Effects of seawater current speed on the photosynthetic oxygen evolution of Sargassum thunbergii (Phaeophyta). Jpn J Phycol 39:291–293
Gao K, McKinley KR (1994) Use of macroalgae for marine biomass production and CO2 remediation: a review. J Appl Phycol 6:45–60. https://doi.org/10.1007/BF02185904
Gower J, Young E, King S (2013) Satellite images suggest a new Sargassum source region in 2011. Remote Sens Lett 4:764–773
Hanisak MD, Samuel MA (1987) Growth rates in culture of several species of Sargassum from Florida, USA. Hydrobiologia 151(152):399–404 In “Twelfth International Seaweed Symposium”, MA Ragan & CJ Bird (eds), Springer, Dordrecht
Johns EM, Lumpkin R, Putman NF, Smith RH, Muller-Karger FE, Rueda-Roa DT, Hu C, Wang M, Brooks MT, Gramer LJ, Werner FE (2020) The establishment of a pelagic Sargassum population in the tropical Atlantic: biological consequences of a basin-scale long distance dispersal event. Prog Oceanogr 182:102269. https://doi.org/10.1016/j.pocean.2020.102269
Johnson DR, Franks JS, Oxenford HA, Cox SL (2020) Pelagic Sargassum prediction and marine connectivity in the tropical Atlantic. Gulf Caribb Res 31(1):GCFI20–GCFI30. https://doi.org/10.18785/gcr.3101.15
Jouanno J, Benshila R, Berline L et al (2021) A NEMO-based model of Sargassum distribution in the tropical Atlantic: description of the model and sensitivity analysis (NEMO-Sarg1.0). Geosci Model Dev 14:4069–4086. https://doi.org/10.5194/gmd-14-4069-2021
Kanamitsu M, Ebisuzaki W, Woollen J, Yang S-K, Hnilo JJ, Fiorino M, Potter GL (2002) DOE AMIP-II Reanalysis (R-2). Bull Am Meteor Soc 83:1631–1643
Lapointe BE (1986) Phosphorus-limited photosynthesis and growth of Sargassum natans and Sargassum fluitans (Phaeophyceae) in the Northwest North Atlantic. Deep-Sea Res 33:391–399
Lapointe BE (1995) A comparison of nutrient-limited productivity in Sargassum natans from neritic vs. oceanic waters of the Northwest North Atlantic Ocean. Limnol Oceanogr 40(3):625–633
Lübbecke JF, Rodríguez-Fonseca B, Richter I, Martín-Rey M, Losada T, Polo I, Keenlyside NS (2018) Equatorial Atlantic variability - modes, mechanisms, and global teleconnections. WIREs Clim Change 9:e527. https://doi.org/10.1002/wcc.527
Marsh R, Addo KA, Jayson-Quashigah P-N, Oxenford HA, Maxam A, Anderson R, Skliris N, Dash J, Tompkins EL (2021) Seasonal predictions of holopelagic sargassum across the tropical Atlantic accounting for uncertainty in drivers and processes: the SARTRAC Ensemble Forecast System. Front Mar Sci 8:722524. https://doi.org/10.3389/fmars.2021.722524
Oviatt CA, Huizenga K, Rogers CS, Miller WJ (2019) What nutrient sources support anomalous growth and the recent Sargassum mass stranding on Caribbean beaches? A review. Mar Pollut Bull 145:517–525. https://doi.org/10.1016/j.marpolbul.2019.06.049
Oyesiku OO, Egunyomi A (2014) Identification and chemical studies of pelagic masses of Sargassum natans (Linnaeus) Gaillon and S. fluitans (Borgessen) Borgesen (brown algae) found offshore in Ondo State Nigeria. Afr J Biotechnol 13(10):1188–1193. https://doi.org/10.5897/AJB2013.12335
Saha A, Serra N, Stammer D (2021) Growth and decay of northwestern tropical Atlantic barrier layers. J Geophys Res Oceans 126:e2020JC016956. https://doi.org/10.1029/2020JC016956
Schell JM, Goodwin DS, Siuda ANS (2015) Recent Sargassum inundation events in the Caribbean: shipboard observations reveal dominance of a previously rare form. Oceanography 28(3):8–10. https://doi.org/10.5670/oceanog.2015.70
Sissini MN, de Barros Barreto MBB, Szechy MTM et al (2017) The floating Sargassum (Phaeophyceae) of the South Atlantic Ocean – likely scenarios. Phycologia 56(3):321–328
Smetacek V, Zingone A (2013) Green and golden seaweed tides on the rise. Nature 504:84–88
Thandlam V, Rahaman H (2019) Evaluation of surface shortwave and longwave downwelling radiations over the global tropical oceans. SN Appl Sci 1:1171. https://doi.org/10.1007/s42452-019-1172-2
Trenberth KE, Shea DJ (2006) Atlantic hurricanes and natural variability in 2005. Geophys Res Lett 33:L12704. https://doi.org/10.1029/2006GL026894
van Tussenbroek BI, Hernández Arana HA, Rodríguez-Martínez RE, Espinoza-Avalos J, Canizales-Flores HM, González-Godoy CE, Barba-Santos MG, Vega-Zepeda A, Collado-Vides L (2017) Severe impacts of brown tides caused by Sargassum spp. on near-shore Caribbean seagrass communities. Mar Pollut Bull 122(1-2):272–281. https://doi.org/10.1016/j.marpolbul.2017.06.057
Vallès-Casanova I, Lee S-K, Foltz GR, Pelegrí JL (2020) On the spatiotemporal diversity of Atlantic Niño and associated rainfall variability over West Africa and South America. Geophys Res Lett 47:e2020GL087108
Wang M, Hu C (2016) Mapping and quantifying Sargassum distribution and coverage in the Central West Atlantic using MODIS observations. Remote Sens Environ 183:356–367
Wang MQ, Hu CM (2017) Predicting Sargassum blooms in the Caribbean Sea from MODIS observations. Geophys Res Lett 44(7):3265–3273
Wang MQ, Hu CM, Barnes BB, Mitchum G, Lapointe B, Montoya JP (2019) The great Atlantic Sargassum belt. Science 365:83–87. https://doi.org/10.1126/science.aaw7912
Xie S-P, Carton JA (2004) Tropical Atlantic variability: patterns, mechanisms, and impacts. In: Wang C, Xie SP, Carton JA (eds) Earth’s climate. American Geophysical Union, Washington D.C.
Yuan TL, Oreopoulos L, Zelinka M, Yu H, Norris JR, Chin M, Platnick S, Meyer K (2016) Positive low cloud and dust feedbacks amplify tropical North Atlantic Multidecadal Oscillation. Geophys Res Lett 43:1349–1356
Zebiak SE (1993) Air–sea interaction in the equatorial Atlantic region. J Clim 6:1567–1586
Acknowledgements
This publication is supported by the UK Economic and Social Research Council through the Global Challenges Research Fund (GCRF) project, Teleconnected sargassum risks across the Atlantic: building capacity for TRansformational Adaptation in the Caribbean and West Africa (SARTRAC), grant number ES/T002964/1. We acknowledge the Optical Oceanography Laboratory at the University of South Florida, for freely providing AFAI images via https://optics.marine.usf.edu.
Funding
The research was funded by the UK’s Economic and Social Research Council through the Global Challenges Research Fund (GCRF) project, Teleconnected sargassum risks across the Atlantic: building capacity for TRansformational Adaptation in the Caribbean and West Africa (SARTRAC), grant number ES/T002964/1.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Responsible Editor: Ricardo de Camargo.
Supplementary information
ESM 1
(DOCX 21 kb)

Fig. S1
Monthly anomaly timeseries of (a) Amazon River discharge (m3 /s) measured at Obidos Gauge (Brazil) over 2010-2019, (b) SMOS SSS averaged over the area 46°W-50°W, 2°N-6°N, typically encompassing the Amazon river outflow close to the river mouth. (PNG 371 kb)

Fig. S2
Monthly anomaly timeseries of (a) Orinoco River discharge (m3 /s) measured at Ciudad Bolivar Gauge (Venezuela) over 2010-2019, (b) SMOS SSS averaged over the area 59°W-63°W, 9°N-12°N, typically encompassing the Orinoco river outflow close to the river mouth. (PNG 382 kb)

Fig. S3
Surface current intensity anomaly (m/s) in spring (Mar-May) (a) and summer (Jun-Aug) (b) 2018 (from GODAS re-analysis). Mean seasonal current velocity vectors for spring and summer 2018, respectively, are also depicted. (c) Monthly surface current intensity anomalies (m/s) over 2010-2020, averaged over 38°W-50°W, 3°S-5°N. (PNG 1892 kb)

Fig. S4
Monthly anomaly timeseries over 2010-2020 of (a) Mixed Layer Depth (m) and (b) Vertical (upwelling) velocity at 40m depth area-averaged over Central Equatorial North Atlantic (CENA). From GODAS ocean re-analysis. (PNG 446 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Skliris, N., Marsh, R., Appeaning Addo, K. et al. Physical drivers of pelagic sargassum bloom interannual variability in the Central West Atlantic over 2010–2020. Ocean Dynamics 72, 383–404 (2022). https://doi.org/10.1007/s10236-022-01511-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10236-022-01511-1