
Abstract
The equine ascarids, Parascaris spp., are important nematode parasites of juvenile horses and were historically model organisms in the field of cell biology, leading to many important discoveries, and are used for the study of chromatin diminution. In veterinary parasitology, Parascaris spp. are important not only because they can cause clinical disease in young horses but also because they are the only ascarid parasites to have developed widespread anthelmintic resistance. Despite this, much of the general biology and mechanisms of anthelmintic resistance are poorly understood. This review condenses known basic biological information and knowledge on the mechanisms of anthelmintic resistance in Parascaris spp., highlighting the importance of foundational research programs. Although two variants of this parasite were recognized based on the number of chromosomes in the 1870s and suggested to be two species in 1890, one of these, P. univalens, appears to have been largely forgotten in the veterinary scientific literature over the past 100 years. We describe how this omission has had a century-long effect on nomenclature and data analysis in the field, highlighting the importance of proper specimen identification in public repositories. A summary of important basic biology, including life cycle, in vitro maintenance, and immunology, is given, and areas of future research for the improvement of knowledge and development of new systems are given. Finally, the limited knowledge regarding anthelmintic resistance in Parascaris spp. is summarized, along with caution regarding assumptions that resistance mechanisms can be applied across clades.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Helminth parasites have been known to humans for thousands of years. The Egyptian Papyrus Ebers, circa 1550 BCE, refers to intestinal worms; the Greeks knew about helminths infecting other species; and the Romans also clearly described Ascaris parasites, including symptoms of clinical disease (Cox 2002). Carl Linnaeus described and named six helminths in 1758, including Ascaris lumbricoides, which ultimately led to an increasing number of helminths being described and formally named (Linnaeus 1758). Ascarids, also known as parasitic roundworms, encompass a large number of important helminth parasites that have three anterior lips in the superfamily Ascaridoidea, including ascarids of poultry, Ascaridia spp. (Dujardin 1845) and Heterakis gallinarum (Schrank 1788); fish, Anisakis spp. (Rudolphi 1809); canids, Toxocara canis (Werner 1782); felids, Toxocara cati (Schrank 1788); felids and canids, Toxascaris leonina (Linstow 1909); cattle, Toxocara vitulorum (Goeze 1782); mustelids and bears, Baylisascaris spp. (Sprent 1968); pigs, Ascaris suum (Goeze 1782); humans, Ascaris lumbricoides (Linnaeus 1758); and equids, Parascaris equorum (Goeze 1782) and Parascaris univalens (Boveri 1887). Many of these parasites can cause severe clinical diseases, including high mortality in cattle (Borgsteede et al. 1992; Gundran and More 1999; Chelladurai et al. 2015); loss of appetite and weight, anorexia, depression, and increased mortality in chickens (Kaufmann et al. 2011; Thapa et al. 2015; Sharma et al. 2019); and pneumonitis, dyspnea, and coughing in pigs (Yoshihara et al. 1983; Curtis et al. 1987; Stewart and Hale 1988; Holland 2013; Mateus et al. 2015), all of which can lead to millions of dollars in agricultural economic losses. Globally, an estimated 807 million to 1.2 billion humans are infected with A. lumbricoides, many of them children in impoverished countries, causing retardation of physical and mental growth, pneumonia, asthma, abdominal distension, intestinal obstruction, pancreatitis, and death (Bethony et al. 2006). Zoonotic infections can also occur with many ascarid parasites. A. suum can complete its life cycle in humans and cause ascariasis (Nejsum et al. 2005; Volk and Tormey 2017; Avery et al. 2018); however, others cannot, leading to a condition known as visceral larva migrans caused by aberrant larval migration. Cases have been reported with T. cati (Eberhard and Alfano 1998; Zibaei et al. 2014), T. canis (Hill et al. 1985; Xinou et al. 2003; Gakosso et al. 2020), Baylisascaris spp. (Saffra et al. 2010; Kelly et al. 2012), and Anisakis spp. (Kojima et al. 2013; Sohn et al. 2015), resulting in various clinical manifestations including coughing, rash, myalgia, liver lesions, myocarditis, visual impairment, neurological symptoms, and death in rare cases.
The equine ascarids, Parascaris spp., are considered the most pathogenic parasites infecting juvenile horses (Equus caballus, Linnaeus, 1758) globally and can cause coughing, nasal discharge, lethargy, poor appetite, diarrhea, and colic (Reinemeyer 2009; Nielsen 2016). Fibrotic liver lesions (Brown and Clayton 1979), lung lesions, hyperpnea, bronchiolitis, and lobular pneumonia (Clayton and Duncan 1978; Nicholls et al. 1978) have been reported in experimentally infected foals. Poor body condition has been associated with Parascaris spp. infection in working equids (Getachew et al. 2008, 2010); however, foals under parasite management programs have not exhibited these signs in recent studies (Bellaw et al. 2016; Nielsen et al. 2021). Small intestine impaction is one of the largest concerns with this parasite, often requiring hospitalization and surgery, and can ultimately lead to death (Nielsen 2016; Southwood et al. 1996; Cribb et al. 2006; Tatz et al. 2012). In 37 published cases where surgical intervention was necessary for impaction colic due to Parascaris spp., 31 horses survived until discharge, but only 11 survived more than 1 year (Nielsen 2016). While the cause of death was not confirmed in these cases, long-term complications resulting from surgery may have contributed to mortality (Santschi et al. 2000; van Loon et al. 2020). Losing young horses results in a direct financial loss from veterinary care and breeding fees; future losses in sales prices, which can be tens of thousands to millions of dollars; competitive winnings; and stud fees; as well as an emotional loss for the owners and caretakers associated with that horse.
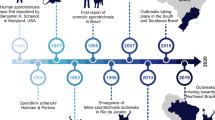
There are three available anthelmintic drug classes for the treatment of Parascaris spp. infection in horses: macrocyclic lactones, benzimidazoles, and tetrahydropyrimidines. Traditionally, foals were treated within their first 30 days of life, and then at either monthly or bimonthly intervals until their first birthday (Drudge and Lyons 1966; Ellingson and Coates-Markle 1996; Robert et al. 2015; Nielsen et al. 2018). Early reports questioning the efficacy of ivermectin against Parascaris spp. (Anderson 1984; Jones 1985) emerged in the mid-1980s, shortly after its introduction to the market, leading to a defense of the drug claiming that the parasite life cycle was misunderstood and errors were made during diagnostic fecal egg counts (Boraski 1987). Formal reports of ivermectin resistance started in the Netherlands in 2002, quickly followed by Canada in 2003 (Boersema et al. 2002; Hearn and Peregrine 2003). This was followed by reports of macrocyclic lactone resistance encompassing the global equine population, and more recently, reports of tetrahydropyrimidine and benzimidazole resistance (Table 1). Current recommendations reduce the overall number of anthelmintic treatments against Parascaris spp. in foals in an attempt to slow down the development of resistance (ESCCAP 2019; Nielsen et al. 2019; Rendle et al. 2019).
Reports of anthelmintic resistance in other ascarid species of veterinary and medical importance are few and far between. Only case reports of resistance are available for a few species, including Ascaris lumbricoides (Krücken et al. 2017), Ascaridia dissimilis (Yazwinksi et al. 2013; Collins et al. 2019), and Heterakis gallinarum (Yazwinksi et al. 2013; Collins et al. 2021), but it is clear from the widespread anthelmintic resistance of many important parasitic nematodes infecting livestock (Rose et al. 2015; Fleming et al. 2006; Sutherland and Leathwick 2011; Kaplan and Vidyashankar 2012; von Samson-Himmelstjerna 2012; von Samson-Himmelstjerna et al. 2021b), including Parascaris spp., that evolution of resistance is a concern. Anthelmintic resistance is an emerging concern in companion animals (Jimenez Castro et al. 2019; Jimenez Castro et al. 2021), and while there have been no reports of resistance in any companion animal ascarid species, frequent monthly treatment intervals necessitate robust anthelmintic resistance monitoring programs (von Samson-Himmelstjerna et al. 2021b). Previous reviews have discussed the need for medical parasitology to learn from veterinary parasitology and identify causes of anthelmintic resistance, modify anthelmintic treatment regimes, and monitor for resistance, in order to, at minimum, slow down the development of resistance (Beech et al. 2010; Vercruysse et al. 2011; Tinkler 2020; von Samson-Himmelstjerna et al. 2021b). This One Health approach and warning to reduce treatment efficacy has been mentioned for nearly two decades (Geerts et al. 1997; Geerts and Gryseels 2000, 2002; Thompson and Roberts 2001), yet little has changed, particularly in human public health (Tinkler 2020).
Despite widespread anthelmintic resistance in Parascaris spp. and the looming specter of resistance in other important ascarid species, little research has been conducted attempting to understand mechanisms of resistance in this species. This review aims to consolidate basic biological information about Parascaris spp. and also highlight the importance of robust foundational research programs.
A tale of two cryptic species
Equine ascarids have an important place in the history of scientific discovery. The German zoologist Johann August Ephraim Goeze was the first to name Ascaris equorum in 1782. In the 1880s, Belgian embryologist Édouard van Beneden used the same parasite—now renamed Ascaris megalocephala—as a model species and showed that fertilization consisted of the union of haploid gametes to form a diploid zygote, contributing greatly to knowledge of both meiosis and mitosis (van Beneden 1883; Hamoir 1992). Over the course of the following 5 years, at least 27 papers—four by van Beneden—were published featuring A. megalocephala studying phenomena such as cell division, chromosome organization, and chromatin diminution (Boveri 1887, 1888).
Two variants of Parascaris were described and distinguished from one another in the late 1800s by counting the number of chromosomes present prior to the first cellular division, a process known as karyotyping (van Beneden 1883; Carnoy 1886/1887; Boveri 1887). Ascaris megalocephala univalens was initially described by van Beneden (van Beneden 1883), and A. meg. bivalens by Jean Baptiste Carnoy (1886/1887) during cytological studies, but it was not until a few years later that Oskar Hertwig recognized them as different species (Hertwig 1890). These species received their currently accepted names in 1978: Parascaris univalens, which has one chromosome pair, and P. equorum which has two pairs of chromosomes (Bullini et al. 1978; Goday and Pimpinelli 1986). Hybrids between these two species have been described (Bullini et al. 1978; Goday and Pimpinelli 1986), although they are sterile (Goday and Pimpinelli 1986). Another species with three pairs of chromosomes, P. trivalens, was described in the 1930s but has not been described in the literature since (Li 1937; Tchou 1937).
The Parascaris species only have a slight morphological difference in their spicula, with Parascaris univalens having a distally truncated spicula and P. equorum having a distally rounded spicula (Biocca et al. 1978). Two methods have been used in the past to distinguish the two species of Parascaris from one another. The first is via karyotyping of primordial germ cells prior to the first cell division, which is an arduous process that requires collecting either parasites with germ cells in the proper stage (Goday and Pimpinelli 1986) or eggs from feces at the first mitotic division (Nielsen et al. 2014; Martin et al. 2018), and the second utilizes electrophoresis of twenty-seven enzyme loci, although this method has only been employed for a single study (Bullini et al. 1978). Karyotyping is a challenging process because it requires either live parasites or eggs that have yet to start developing, and due to this, it is rarely performed for parasitological studies utilizing equine ascarids.
In the nearly 100 years between Hertwig naming the two species and their modern name assignment in 1978, Parascaris univalens and P. equorum were not recognized as separate species and were instead recognized as different variants of P. equorum (Lin, 1954), which may explain why P. equorum was thought to be the only species. This ultimately led to P. equorum being the only species mentioned in veterinary textbooks and research for decades, with the only recognition of two species occurring in cell biology and cytogenetics, highlighting a lack of communication between disciplines. The last positive identification of P. equorum via karyotyping was in 1986 (Goday and Pimpinelli 1986), despite contemporary recognition of two species and an increased effort to karyotype specimens. Ultimately, lack of consensus on species versus variants and naming them led to a decades-long misclassification of specimens that had and continues to have far-reaching consequences.
Phylogenetics
The phylum Nematoda (Cobb, 1932) consists of over 22,000 named species separated into five distinct clades (Blaxter et al. 1998; Blaxter and Koutsovoulos 2015). The ascarid parasites fall under Clade III, which also includes pinworms, filarial nematodes, and parasites of millipedes (Blaxter et al. 1998). Within the Ascaridoidea superfamily, Parascaris spp. belongs to the monophyletic clade of Ascarididae along with Baylisascaris spp., Toxoascaris leonina, and Ascaris spp. (Nadler 1987; Nadler and Hudspeth 2000; Liu et al. 2016; Li et al. 2018). Parasitism in the Ascaridoidea includes prehistoric host-type switches correlated with global changes in sea level (Li et al. 2018), and tissue parasitism within Clade III evolved separately at least three different times (Nadler et al. 2007). Understanding these evolutionary relationships within Clade III and the Ascarididae clade provides important context when comparing these parasites to other groups within the Nematoda.
Some other well-known and heavily studied species, such as the model organism Caenorhabditis elegans (Maupas, 1900), a free-living nematode, and Haemonchus contortus (Rudolphi, 1803), the most pathogenic and economically significant nematode parasite of small ruminants and model organism for parasitic nematodes, fall under Clade V (Blaxter et al. 1998). Anthelmintic resistance is rampant in H. contortus, and it has therefore been heavily studied (Kotze et al. 2014; Kotze and Prichard 2016), along with substantial research on the same topic in C. elegans (Geary and Thompson 2001; Kotze et al. 2014). This means that much of the research conducted regarding anthelmintic resistance that will be described in subsequent sections has been broadly applied to ascarids despite the work being completed in organisms belonging to a completely different clade. The evolutionary distance between Parascaris spp., C. elegans, and H. contortus combined with distinct differences in the life cycle and parasitism suggests that direct comparisons and broadly applying information from one clade to another must be done with caution.
Genetics
Genetic studies opened the door for contemporary recognition of two distinct Parascaris species, although earlier classification as variants named Parascaris equorum and Parascaris equorum univalens led to the majority of specimens in veterinary parasitology being called Parascaris equorum. One study utilizing electrophoresis of enzyme loci showed the opposite, with 93.5% of 2238 specimens collected in an abattoir identified as P. univalens (Bullini et al. 1978). This study, however, was published in Italian and appears to have gone unnoticed for several decades. A more recent population genetics study compared equine ascarid specimens from Sweden, Norway, Germany, Iceland, Brazil, and the USA and found that all of the parasites were genetically homogenous (Tydén et al. 2013b). One of the study populations was later karyotyped and found to consist only of P. univalens (Nielsen et al. 2014). Additional populations in the USA (Nielsen et al. 2014), Sweden (Martin et al. 2018), Iceland (Martin et al. 2021c), and China (Han et al. 2022) were karyotyped and identified as P. univalens, which taken together suggests that the main species present in domestic horses globally is P. univalens and not P. equorum.
A phylogenetic analysis of Parascaris spp. parasites from the mountain zebra (Equus zebra, Linnaeus, 1758), domestic horse, and wild ass (Equus asinus, Gray, 1824) using the mitochondrial genes cox1 and nadh1, demonstrated that the worms from E. asinus formed a distinct clade compared to specimens collected from the other two Equus species (Peng et al. 2019). A recent whole-genome study of specimens from horses, donkeys (Equus africanus asinus, Linnaeus, 1758), and zebras also indicated distinct clades for P. univalens specimens—some of which were confirmed via karyotyping—found within horses and those found in zebras and donkeys (Han et al. 2022). Another recent study completed a phylogenetic analysis for a select group of nuclear and mitochondrial genes using almost all Parascaris spp. Using DNA sequences from GenBank along with karyotyped specimens confirmed to be P. univalens, they found a small group of sequences, all from parasites collected from donkeys on a single farm in China, that formed a cluster (von Samson-Himmelstjerna et al. 2021a). Due to the genetic distance from P. univalens specimens from North America and Europe, the specimens in this cluster may represent another genotype or species of Parascaris (von Samson-Himmelstjerna et al. 2021a). This cluster could represent P. equorum, or even another species such as P. trivalens, which has only been described in a couple of studies using parasites from Chinese horses (Li 1937; Tchou 1937).
The first Parascaris spp. draft genome was published in 2017 for P. univalens, followed by a draft genome for P. equorum in 2019, both indicating over 14,000 coding genes present in the parasites (Wang et al. 2017; International Helminth Genomes Consortium 2019). When considering these genomes, however, the Parascaris species conundrum must be taken into consideration. The specimen reported to be P. equorum and used for the 50 Helminth Genomes Project was collected at necropsy from an abattoir, and there is no indication that karyotyping was performed to positively identify the species (International Helminth Genomes Consortium 2019). The previously described phylogenetic study using Parascaris sequences from GenBank along with karyotyped specimens indicated that nearly every sequence for the internal transcribed spacer-1 and spacer-2 and cytochrome oxidase I labeled as P. equorum was clustered within confirmed P. univalens specimens, indicating that they are likely all from P. univalens (von Samson-Himmelstjerna et al. 2021a). This information, combined with the lack of karyotyping, previous research suggesting that P. univalens is the predominant species in domestic horses, and the fact that P. equorum has not been identified via karyotyping since 1986 (Goday and Pimpinelli 1986), suggests that the specimen in WormBase ParaSite, as well as many other data deposits in GenBank labeled as P. equorum, may be P. univalens (Nielsen et al. 2014; International Helminth Genomes Consortium 2019; von Samson-Himmelstjerna et al. 2021a). Currently, there are no GenBank deposits verified as P. equorum via karyotyping.
Incorrectly identified information in public repositories is detrimental to the field and can lead to misinterpretation of results. For example, one study comparing Parascaris mitochondrial genomes utilized fresh specimens that were not karyotyped but assumed to be P. equorum (Gao et al. 2019). These specimens were then compared to mitochondrial genomes from two karyotyped P. univalens isolates and one non-karyotyped isolate assumed to be P. equorum. The subsequent phylogenetic analysis clustered these four specimens into a single clade, and the authors concluded that P. equorum and P. univalens may represent the same species (Gao et al. 2019). This, however, is inaccurate given that no attempt was made to identify the collected specimen. Instead, the clustering of the two identified specimens strongly suggests that the collected specimen was P. univalens. Correctly identifying species is important not only for ensuring accurate research and performing future genome-wide research studies in a variety of disciplines, including the study of anthelmintic resistance, but also for developing molecular techniques, such as PCR, to identify Parascaris specimens to species (Doyle and Cotton 2019; von Samson-Himmelstjerna et al. 2021a).
Chromatin diminution
After the first cell division, Parascaris spp. presomitic cells go through a process called chromatin diminution where chromosomes are fragmented and approximately 85% of the germline genome is eliminated, resulting in the creation of about 35 smaller chromosomes (Boveri 1887; Goday and Pimpinelli 1986; Muller and Tobler 2000; Niedermaier and Moritz 2000). The initial discovery of chromatin diminution was made with Parascaris spp. in 1887 and was later found to occur in other nematodes, including Ascaris suum, A. lumbricoides, and Toxocara spp., copepods, ciliates, hagfish, lamprey, and rat fish (Boveri 1887; Wang and Davis 2014). The P. univalens germline genome has an estimated 2500 megabases (Mb), whereas the somatic genome has an estimated 250 Mb, indicating a large loss of genetic information in an organism with only a single chromosome (Wang et al. 2017). Comparisons between Parascaris spp. and Ascaris spp. indicate that the mechanism for chromatin diminution is evolutionarily conserved between the two species and likely present in a common ancestor (Bachmann-Waldmann et al. 2004). Comparative analysis of Parascaris, Ascaris, and Toxocara genomes shows that somewhere between 1000 and 2000 genes are eliminated, with 35% of those being expressed during spermatogenesis, and it is hypothesized that this allows for rapid adaptation and evolutionary change in the testes without causing deleterious effects because those genes are silenced and eliminated (Bachmann-Waldmann et al. 2004; Wang et al. 2017; Wang 2021).
Ultimately, the process of chromatic diminution, even though it causes a large loss of genetic information, is likely evolutionarily advantageous for Parascaris spp in some way. It is possible that chromatin diminution helps prevent events such as population bottlenecks due to the ability of the parasites to undergo rapid evolutionary changes in the germ line and may even have played a role in the evolution of parasitism (Bachmann-Waldmann et al. 2004). Evidence from cytogenetic studies suggests that there are differences in chromosome and heterochromatin organization between the two species (Goday and Pimpinelli 1984, 1986; Goday et al. 1985). It remains to be seen how this process differs molecularly and on a whole-genome level between the two species, particularly since the last karyotyped P. equorum specimen was identified in 1986, 2 years before the method for PCR was first published and 6 years before the first-ever whole genome sequence was completed (Goday and Pimpinelli 1986; Mullis and Faloona 1987; Fleischmann et al. 1995). If P. equorum was out-competed by P. univalens due to a fitness disadvantage as anthelmintic use became more prevalent, understanding chromatin diminution and comparing the two species could be an important key to understanding anthelmintic resistance development in ascarid parasites (von Samson-Himmelstjerna et al. 2021a).
Life cycle and immunology
Life cycle
Parascaris spp. are robust, cream-colored nematode parasites with a direct life cycle whose adult stages are found primarily within the small intestine of equids. Females are typically 10–20 cm in length with a diameter of 5 mm, whereas their smaller male counterparts are only 10–15 cm in length with a 3-mm diameter (Wells 1924; Clayton and Duncan 1979a). Males can be distinguished from females of a similar size by a curl in the posterior end and a lack of visible ovaries through the cuticle (Wells 1924).
Adult Parascaris spp. reproduce sexually in the small intestine via genital pores, and females lay their 40–70 μm eggs in the small intestinal content, from where they are excreted into the environment (Wells 1924). In the pasture, the fertilized parasite eggs embryonate and larvae develop within the eggs; it is this egg containing a second-stage larva that is infective (Wells 1924; Clayton and Duncan 1979a). Once ingested by a foal, the eggs hatch in the small intestine, and the larvae penetrate the intestinal wall, where they subsequently migrate to the liver within a week of initial infection (Clayton and Duncan 1979a). Within 2 weeks after the initial infection, the larvae enter the lungs via the pulmonary circulation, where they emerge from arterioles and capillaries. The larvae are coughed up and then swallowed by the foal, making their way back to the small intestine 2 to 3 weeks after initial infection (Clayton and Duncan 1979a). At this stage, Parascaris spp. larvae are approximately 2–4 mm in length and, over the next 4.5 months, will grow 70–80 × in size as they feed on intestinal content and mature into adults (Clayton and Duncan 1979a). While the general life cycle of Parascaris spp. has been described, the biological reasoning for larval migration is poorly understood. It has been suggested that tissue migration may be linked to increased body size and faster growth (Read and Skorping 1995) and may also play a role in immune evasion (Mulcahy et al. 2005; Deslyper et al. 2016, 2019). This, however, has not been directly studied in Parascaris spp., and no biological signals that may be required for parasite maturation have been identified. Understanding these signals would provide valuable insight into parasite biology, possible control mechanisms, and conditions necessary for in vitro culture from egg to adult.
Culturing in vitro
Parasites can be difficult to maintain and grow in vitro because of their sometimes-complicated life cycles that are reliant on the correct host. This makes research complicated, particularly in the case of species whose hosts are either unethical to use as research subjects, such as humans, or too difficult—whether because of size, expense, or husbandry needs—to maintain as a research population. In the case of Parascaris spp., equine research herds are expensive to maintain, require a large amount of land, and obtaining adult parasites requires euthanasia of healthy foals that require eleven months gestation and another approximate five months before adult parasites can be harvested. There is one research herd known globally that is regularly used for this purpose (Lyons et al. 1990), and many specimens obtained elsewhere are from abattoirs (Janssen et al. 2013; Martin et al. 2020, 2021a; Trailovic et al. 2021) or collected opportunistically at diagnostic necropsies (Burk et al. 2014, 2016; Rakhshandehroo et al. 2016; Malekpour et al. 2019). Adult Parascaris spp. can be maintained in vitro for up to a week (Janssen et al. 2013; Scare et al. 2019; Martin et al. 2021a) but show transcriptional stress responses to culture conditions within the first 24 h compared to non-cultured worms via an increase in differentially expressed genes (Martin et al. 2020). Adult Parascaris in general do not maintain fitness well in culture, as evidenced by their short survival time and stress responses, compared to their Ascaris suum counterparts that are able to be kept alive for at least 2 weeks (Islam et al. 2004). Despite these challenges, meaningful gene expression data have been obtained from current in vitro systems (Janssen et al. 2013; Scare et al. 2020; Martin et al. 2021a), as well as the development of a Parascaris spp. fitness scoring system (Scare et al. 2019).
L2/L3 larvae can be hatched from eggs (Burk et al. 2014; Rakhshandehroo et al. 2017; Martin et al. 2021a), which does not require sacrificing a horse, although they cannot be grown into L4 and L5 larvae or adults. The longevity of these larvae in culture is unknown, but they have been kept alive for at least 48 h (Martin et al. 2021a) and have been used in drug exposure (Rakhshandehroo et al. 2017; Martin et al. 2021a) and immunology (Burk et al. 2014, 2016) studies. Differences in protein transport gene expression between adults and larvae hatched from eggs in vitro (Martin et al. 2021a) must be considered when interpreting data and comparing results between life stages, but the larval culturing system is a promising path forward in Parascaris spp. research because it allows studies to take place without sacrificing young horses.
Immunology
With a few exceptions, such as adult horses in tropical regions and donkeys (Vercruysse et al. 1986; Getachew et al. 2008, 2010; Lem et al. 2012), Parascaris spp. are generally found in juvenile horses up until the age of 6 to 8 months when an age-dependent immunity develops (Clayton and Duncan 1979b; Fabiani et al. 2016). Fecal egg shedding and worm counts in juvenile horses occur in an age-dependent manner, and in older foals, fewer larvae reach the small intestine, patent infections are less likely to develop, and fecal egg counts are lower (Clayton and Duncan 1979b; Donoghue et al. 2015; Fabiani et al. 2016). In a study where eight worm-free foals and two yearlings were experimentally infected with Parascaris spp., yearlings had a more severe respiratory response but maintained their body condition, whereas foals had a mild respiratory response and lost body condition, suggesting an age-dependent immune response, although the sample size was small (Clayton and Duncan 1978). Increased titers of antibodies to whole worm antigen have been shown to correlate with foal age and subsequent reduction in parasite prevalence (Bello 1985), and immune responses to migrating larvae in the lungs (Nicholls et al. 1978) and liver (Brown and Clayton 1979) have also been illustrated. There are no studies showing direct parasite death or fitness loss as a result of equine immune responses, and molecular evidence of immune response has yet to be demonstrated in horses, despite evidence of an age-dependent response. Understanding the equine immune response to Parascaris spp. at an in-depth molecular level would provide invaluable information regarding host-parasite dynamics and open the door for possible vaccine development.
Helminth excretory–secretory products, including microribonucleic acids (RNAs) (Sotillo et al. 2020) and extracellular vesicles (Zakeri et al. 2021), are thought to play a role in immune evasion, elicit host immune response, and may allow for the development of vaccines and/or diagnostic tests (Lightowlers and Rickard 1988). In Ascaris suum, extracellular vesicles contain immunomodulatory proteins (Hansen et al. 2019), and microRNAs may be important for parasite development (Xu et al. 2013). An in vitro analysis of larval Parascaris spp. excretory–secretory products identified 19 kDa, 22 kDa, 26 kDa, and 34 kDa products that elicited an antibody response in sera of previously infected foals (Burk et al. 2014). Mares were shown to have antibodies against these products and passed them to foals via colostrum during the first suckling (Burk et al. 2016). These antibodies are not useful for diagnosis because the foals acquire them shortly after birth, and they are likely not useful for vaccination because, despite their presence, foals still become infected with Parascaris spp. There have been no studies to date examining Parascaris spp. extracellular vesicles or microRNAs, and ultimately, more research is necessary to determine the nature of equine immunity against Parascaris spp.
The biology of resistance
In the early twentieth century, John D. Rockefeller committed over US$1 million to hookworm control and research, and Epsom salts, thymol, carbon tetrachloride, oil of chenopodium, tetrachlorethylene, and hexylresorcinol were all either used or investigated for use as anthelmintics (Horton 2003). Continuing into the twentieth century, various dyes and synthetic compounds were used to treat helminth infections, but many were ineffective, difficult to use, and/or toxic, causing a plethora of issues such as deafness, blindness, skin irritation, diarrhea, vomiting, organ damage, and death (Faust 1937; Horton 2003). The first safe, modern anthelmintics were phenothiazine, introduced in the 1940s, and piperazine, introduced in the 1950s. This was followed by the major anthelmintics currently used in horses, starting in 1961 with benzimidazoles and ending with the introduction of the macrocyclic lactones in 1981 (Brown et al. 1961; Campbell et al. 1983; Laing et al. 2016).
Anthelmintic resistance is rampant in veterinary parasitology (Rose et al. 2015; Fleming et al. 2006; Sutherland and Leathwick 2011; Kaplan and Vidyashankar 2012), and understanding how it developed in order to slow down the progression and preserve current anthelmintics for as long as possible, as well as preserve any new anthelmintic for as long as possible, is important. Husbandry practices and anthelmintic treatment strategies as methods to slow down the development of resistance have been discussed in detail elsewhere (Reinemeyer 2009; von Samson-Himmelstjerna 2012; Matthews 2014; Nielsen 2016; Reinemeyer and Nielsen 2017; von Samson-Himmelstjerna et al. 2021b), although the effect of these practices is not always apparent. Anthelmintic mechanisms of action as well as mechanisms of resistance in parasitic nematodes have also been thoroughly reviewed in the past (Prichard 1994; Kotze et al. 2014; Whittaker et al. 2017; Kaplan 2020). This section will briefly describe drug mechanisms of action and resistance in general, with a focus on relevant research conducted using Parascaris spp.
Benzimidazoles
Benzimidazole is a heterocyclic aromatic organic compound, and various modifications to this structure have resulted in the development of anthelmintic drugs (Townsend and Wise 1990). The first was introduced in 1961, followed by numerous formulations in the 1960s and 1970s (Brown et al. 1961; Harder 2002). Benzimidazole is a microtubule inhibitor that interacts with the colchicine-binding domain of β-tubulin and interrupts polymerization, disrupting vital cellular processes and causing parasite death (Friedman and Platzer 1978; Lacey 1988, 1990). Microtubules are polymers made of tubulin dimers consisting of α- and β-tubulin and are essential for cellular structure and processes such as intracellular transport and cell division (Lacey 1988, 1990). Benzimidazoles developed as anthelmintics have a higher binding affinity for nematode β-tubulin than mammalian β-tubulin, making them safe for use in horses and other mammalian species (Lacey 1988).
Benzimidazole resistance mechanisms are the most well-studied of the anthelmintic classes because of their rapid development in Clade V nematodes, in particular Haemonchus contortus, just 3 years after its introduction to the market (Drudge et al. 1964; Kotze and Prichard 2016). Benzimidazole resistance is associated with mutations in isotype-1 and isotype-2 β-tubulin genes that decrease the binding affinity of the drug for its target (Lubega and Prichard 1990; Lacey and Gill 1994; von Samson-Himmelstjerna et al. 2007a). There are a few single nucleotide polymorphisms (SNPs) that are associated with benzimidazole resistance in H. contortus, with phenylalanine to tyrosine substitution at codon 200 (F200Y) in the isotype-1 β-tubulin gene being the most common in wild type parasite populations (Kotze and Prichard 2016). Other mutations in isotype-1 β-tubulin linked to benzimidazole resistance include F167Y and E198A, with the former being quite rare and the latter conferring the highest level of resistance of the three (Ghisi et al. 2007; Kotze et al. 2012). Limited research regarding isotype-2 β-tubulin genes has been performed, but some resistant populations of H. contortus show loss or decreased levels of the gene (Beech et al. 1994; Lubega et al. 1994).
Benzimidazoles are still an effective anthelmintic for the treatment of Parascaris spp. infections and resistance has only been reported in three studies beginning in 2014 (Table 1). Due to this limited emerging anthelmintic resistance, only a few studies have examined the resistance-related SNPs or transcriptional responses to benzimidazoles in Parascaris spp. Five studies have sequenced Parascaris spp. β-tubulin genes, including one using a known benzimidazole-resistant isolate, and none found any known resistance-related SNPs, suggesting a potentially different mechanism of resistance in ascarid parasites to this anthelmintic class (Tydén et al. 2013a, 2014; Malekpour et al. 2019; Martin et al. 2021b; Özben et al. 2022). Interestingly, it has been shown that isotype-1 and isotype-2 β-tubulin genes are expressed at higher levels in Parascaris spp. eggs, and while isotype-1 remains at similar levels of expression in larvae and adults, isotype-2 gene expression is very low in adults, suggesting differing functions throughout the life cycle (Tydén et al. 2016). Additionally, in vitro exposure to benzimidazoles significantly increased gene expression of isotype-1 β-tubulin genes in one study using eggs (Tydén et al. 2016), whereas in vitro studies using adult parasites showed either downregulation of isotype-2 β-tubulin (Martin et al. 2020) or no differential expression of β-tubulin genes (Scare et al. 2020). Previously discussed enzymes aiding in the removal of xenobiotic compounds, as well as genes related to detoxification, microtubule polymerization, regulation of membrane potential, and muscle contraction were also differentially expressed following in vitro exposure to benzimidazoles (Martin et al. 2020; Scare et al. 2020).
The β-tubulin genes targeted by benzimidazoles are different even within Clade V nematodes (Saunders et al. 2013), and resistance-related β-tubulin SNPs have a low frequency in benzimidazole-resistant equine cyathostomins, another Clade V parasite group, suggesting that they may not fully describe benzimidazole resistance even within the clade (Pape et al. 2003; von Samson-Himmelstjerna et al. 2003, 2007a; James et al. 2009). Considering these dissimilarities within Clade V and the lack of identification of known resistance-related SNPs in benzimidazole-resistant Parascaris spp., it is possible that the mechanism of resistance in Clade III ascarid-type nematodes is different and thus using these SNPs for anthelmintic resistance surveillance is inadvisable (Diawara et al. 2009, 2013; Rashwan et al. 2017; Zuccherato et al. 2018; Palma et al. 2020).
Tetrahydropyrimidines
The tetrahydropyrimidines include two formulations of pyrantel using different salts: pyrantel pamoate and pyrantel tartrate. Drugs in this class act as agonists of acetylcholine receptors (AChRs) and cause them to stay open, leading to prolonged muscle contraction and paralysis in the parasites (Harrow and Gration 1985; Robertson et al. 1994). Nicotinic acetylcholine receptors are ligand-gated ion channels activated by acetylcholine, a neurotransmitter, and are made up of five subunits surrounding a central pore (Beech and Neveu, 2015). The AChR repertoire of parasitic nematodes is not widely studied, with only a few subtypes having been described in nematodes in general, and even fewer when the scope is narrowed to Parascaris. Recently, a Parascaris ACR-16 receptor subunit was described, and it was found that parasitic nematodes have two AChR subunits, ACR-26 and ACR-27, that are not found in free-living nematodes (Courtot et al. 2015; Charvet et al. 2018). In Parascaris spp., these two subunits have a higher affinity for pyrantel than acetylcholine (Courtot et al. 2015).
Parascaris spp. resistance to pyrantel has only been reported in seven studies globally starting in 2007, and thus has limited research due to the lack of resistant parasite populations (Table 1). There has, however, been one in vitro study investigating transcriptional responses in Parascaris spp. when exposed to pyrantel, ivermectin, and thiabendazole (Martin et al. 2020). Expression of eight transcripts orthologous to AChR was differentially expressed, but with no clear pattern between drug classes (Martin et al. 2020). Differential expression was also found in genes coding for enzymes that aid in the removal of xenobiotic compounds including short-chain dehydrogenases/reductases and flavin-containing monooxygenases, but these enzymes have not been characterized in parasitic nematodes and thus more research must be completed to understand their possible involvement in anthelmintic resistance (Martin et al. 2020).
Macrocyclic lactones
Macrocyclic lactones are a group of drugs derived from avermectin produced by Streptomyces avermitilis (Campbell et al. 1983; Kim and Goodfellow 2002) or milbemycins produced by S. hygroscopicus (Takiguchi et al. 1980) or S. cyaneogriseus (Carter et al. 1988) and consist of some of the most well-known and widely used anthelmintics in the world. The avermectin derivatives—particularly ivermectin—have had a large impact in both veterinary and human medicine and the 2015 Nobel Prize in Physiology or Medicine was awarded to William C. Campbell and Satoshi Ōmura for its discovery (Nobel Prize 2015). Ivermectin was first introduced in 1981, and by the end of the decade, it was the best-selling animal health product in the world (Laing et al. 2016). Moxidectin, a milbemycin derivative, was introduced in the mid-1990s and has a longer half-life and higher potency than ivermectin (Lyons et al. 1992; Afzal et al. 1997). Macrocyclic lactones irreversibly activate glutamate-gated chloride channels (GluCls) that are present in nematode neuron and muscle cells, inhibiting neuronal and muscle activity and ultimately causing paralysis and death (Wolstenholme 2012; Laing et al. 2016). Within nematodes, even those that are within the same clade such as Caenorhabditis elegans and Haemonchus contortus, GluCls are highly divergent, making comparisons between species, let alone clades, difficult when studying both the mechanism of action and development of resistance (Laing et al. 2016).
Macrocyclic lactone resistance was first reported in Haemonchus contortus in 1987—just 6 years after ivermectin hit the market—and continued to spread globally (Carmichael et al. 1987; Van Wyk et al. 1987; Prichard 1994). Despite widespread anthelmintic resistance to ivermectin in some species of nematode parasites, the mechanism for resistance remains poorly understood. Similar to benzimidazole resistance, ivermectin resistance has been studied in H. contortus, as well as in Caenorhabditis elegans (reviewed in Lespine et al. 2011; Doyle and Cotton 2019), but little research has been conducted in ascarid parasites. P-glycoproteins (Pgp) are cell membrane efflux proteins that pump foreign substances out of cells and were first associated with ivermectin resistance in parasitic nematodes in the late 1990s (Xu et al. 1998). Subsequently, they are one of the most widely studied putative mechanisms for macrocyclic lactone resistance and the only one that has been studied in Parascaris spp. Similar to benzimidazole resistance, the bigger picture is complex. Macrocyclic lactone resistance is likely multigenic (Choi et al. 2017; Khan et al. 2020) and the molecular mechanism is not fully understood (Laing et al. 2016; Rezansoff et al. 2016).
Ten Parascaris spp. Pgps have been identified to date, along with their tissue-specific expression levels and some evidence for interaction with ivermectin: Pun-Pgp-2, Pun-Pgp-3, Pun-Pgp-9, Pun-Pgp-10, Pun-Pgp-11.1, Pun-Pgp-11.2, Pun-Pgp-12, Pun-Pgp-16.1, Pun-Pgp-16.2, and Pun-Pgp-18 (Janssen et al. 2013; Chelladurai and Brewer 2019; Gerhard et al. 2020; Martin et al. 2021a). Their role in anthelmintic resistance, however, is unclear. Transgenic expression of P. univalens Pun-Pgp-9 and Pun-Pgp-11 in Caenorhabditis elegans decrease susceptibility to ivermectin (Janssen et al. 2015), and Pun-Pgp-9 does so in a tissue-specific manner, with intestinal expression conferring a protective effect, and depends on active ingestion via pharyngeal pumping (Gerhard et al. 2021). Comparisons between ivermectin resistant and susceptible Parascaris spp. populations revealed the presence of SNPs in Pun-Pgp-11 and increased Pun-Pgp-11 mRNA levels correlating to decreased macrocyclic lactone susceptibility (Janssen et al. 2013), but drug exposure assays showed no change in Pgp expression in response to ivermectin exposure (Gerhard et al. 2020; Scare et al. 2020; Martin et al. 2021a). Differentially expressed genes for enzymes aiding in the removal of xenobiotic compounds and other cellular processes were similar to those previously described for other drugs after ivermectin exposure, with the only exception being the upregulation of a gamma-aminobutyric acid subunit (Martin et al. 2020; Scare et al. 2020). While these studies suggest some potential candidates for ivermectin resistance mechanisms, it is similar to the other two previously discussed drug classes in that mechanisms for resistance are unclear.
Novel anthelmintics
Several novel anthelmintic candidates have been tested for Parascaris spp., many of them involving plant extracts. Wild tarragon (Artemisia dracunculus, Linnaeus, 1753), pennyrile (Mentha pulegium, Linnaeus, 1753), Zataria multiflora (Boiss), cinnamon (Cinnamomum zeylanicum, Blume), pomegranate flower (Punica granatum, Linnaeus, 1753), and pepper (Capsicum annuum, Linnaeus, 1753) extracts were all lethal to L2/L3 larval Parascaris in vitro (Rakhshandehroo et al. 2016, 2017). Zinc oxide nanoparticles showed in vitro anthelmintic efficacy against Parascaris spp., including changes to morphological appearance (Morsy et al. 2019). The monoterpenic phenol isomer carvacrol, isolated from herbs, also showed in vitro anthelmintic activity against Parascaris spp. by inhibiting acetylcholine-induced currents and stopping muscle contractions, suggesting that it is an antagonist of AChRs similar to pyrantel (Trailovic et al. 2021). The Bacillus thuringiensis (Berliner, 1915) crystal protein Cry5B has shown efficacy against Parascaris spp. when administered to foals via nasogastric tube, dropping fecal egg counts to zero, and is the only in vivo experimental drug efficacy study that has been completed recently (Urban et al. 2021). While these treatments have shown some efficacy, there is little information regarding the mechanisms of action, which will be an essential piece of information if they make it to the commercial market in order to help prevent the development of resistance.
Conclusions
Anthelmintic resistance mechanisms are poorly understood in helminths, particularly ascarid parasites due to Parascaris spp. being the only ascarid exhibiting widespread anthelmintic resistance. Limited research suggests that mechanisms of resistance in Clade III ascarids may be different from those in Clade V members such as strongylid parasites and C. elegans, and applying information learned from these Clade V nematodes to ascarids must be done cautiously. Continued research on resistance mechanisms in ascarid parasites using Parascaris spp. is important for the understanding of genetic and molecular mechanisms of resistance in order to preserve anthelmintics currently used in other ascarid species and develop new treatment options for the future.
The recent development of egg-hatching larval culture methods for Parascaris spp. makes it a prime candidate for research regarding anthelmintic resistance in ascarid-type parasites, although in vitro systems have limitations such as short Parascaris spp. lifespan, differences in gene expression between life stages due to culture conditions, and specimen acquisition. Continued improvement of in vitro systems for both adults and hatched L2/L3 larvae, particularly increasing lifespan in culture and decreasing stress by optimizing culture conditions, would not only increase the quality of data collected but also allow for a larger number of studies to be completed.
Open access data repositories such as GenBank and WormBase Parasite are used by scientists globally for various research applications, and it is important that the information is updated as necessary. Parascaris spp. is an example of the issues that incorrect submissions can cause due to mislabeled accessions. Samples must be properly identified prior to genetic analysis and submission, as illustrated by the confusion between P. univalens and P. equorum. This mislabeling and lack of identification of species is not only something that needs to be rectified in date repositories, but also serves as a warning regarding proper specimen identification and labeling. Additionally, the Parascaris species discrepancy highlights the importance of developing a molecular method for Parascaris species identification.
Data availability
N/A.
Code availability
N/A.
References
Afzal J, Burke AB, Batten PL, DeLay RL, Miller P (1997) Moxidectin: metabolic fate and blood pharmacokinetics of 14C-labeled moxidectin in horses. J Agric Food Chem 45(9):3627–3633
Alanazi AD, Mukbel RM, Alyousif MS, AlShehri ZS, Alanazi IO, Al-Mohammed HI (2017) A field study on the anthelmintic resistance of Parascaris spp. in Arab foals in the Riyadh region, Saudi Arabia. Vet Q 37(1):200–205
Anderson RR (1984) The use of ivermectin in horses: research and clinical observations. Compend Contin Educ Vet 6(9):s516–s521
Armstrong SK, Woodgate RG, Gough S, Heller J, Sangster NC, Hughes KJ (2014) The efficacy of ivermectin, pyrantel and fenbendazole against Parascaris equorum infection in foals on farms in Australia. Vet Parasitol 205(3–4):575–580
Avery RH, Wall LA, Verhoeve VI, Gipson KS, Malone JB (2018) Molecular Confirmation of Ascaris suum: further investigation into the Zoonotic origin of infection in an 8-year-old boy with Loeffler syndrome. Vector Borne Zoonotic Dis 18(11):638–640
Bachmann-Waldmann C, Jentsch S, Tobler H, Müller F (2004) Chromatin diminution leads to rapid evolutionary changes in the organization of the germ line genomes of the parasitic nematodes A. suum and P. univalens. Mol Biochem 134(1):53–64
Beasley A, Coleman G, Kotze AC (2015) Suspected ivermectin resistance in a south-east Queensland Parascaris equorum population. Aust Vet J 93(9):305–307
Beech RN, Neveu C (2015) The evolution of pentameric ligand-gated ion-channels and the changing family of anthelmintic drug targets. Parasitol 142(2):303–317
Beech RN, Prichard RK, Scott ME (1994) Genetic variability of the beta-tubulin genes in benzimidazole-susceptible and -resistant strains of Haemonchus contortus. Genetics 138(1):103–110
Beech RN, Skuce P, Bartley DJ, Martin RJ, Prichard RK, Gilleard JS (2010) Anthelmintic resistance: markers for resistance, or susceptibility? Parasitol 138(2):160–174
Bellaw JB, Pagan J, Cadell S, Phethean E, Donecker JM, Nielsen MK (2016) Objective evaluation of two deworming regimens in young thoroughbreds using parasitological and performance parameters. Vet Parasitol 221:69–75
Bello TR (1985) The insidious invasive verminous antigens of the horse. J Equine Vet Sci 5(3):163–167
Bethony J, Brooker S, Albonico M, Geiger SM, Loukas A, Diemert D, Hotez PJ (2006) Soil-transmitted heliminth infections: ascariasis, trichuriasis, and hookworm. Lancet 367(9521):1521–1532
Biocca E, Nascetti G, Iori A, Constantini R, Bullini L (1978) Descrizione di Parascaris univalens, parassita degli equini, e suo differenziamento da Parascaris equorum. Atti della Accademia Nazionale dei Lincei: Classe di Scienze Fisiche, Matematiche e Naturali 65(3–4):133–140
Bishop RM, Scott I, Gee EK, Rogers CW, Pomroy WE, Mayhew IG (2014) Sub-optimal efficacy of ivermectin against Parascaris equorum in foals on three Thoroughbred stud farms in the Manawatu region of New Zealand. N Z Vet J 62(2):91–95
Blaxter M, Koutsovoulos G (2015) The evolution of parasitism in Nematoda. Parasitol 142(Suppl 1):S26–S39
Blaxter ML, De Ley P, Garey JR, Liu LX, Scheldeman P, Vierstraete A, Vanfleteren JR, Mackey LY, Dorris M, Frisse LM, Vida JT, Thomas WK (1998) A molecular evolutionary framework for the phylum Nematoda. Nature 392(6671):71–75
Boersema JH, Eysker M, Nas JWM (2002) Apparent resistance of Parascaris equorum to macrocylic lactones. Vet Rec 150(9):279–281
Boraski EA (1987) Efficacy of oral ivermectin against ascarids in horses. Mod Vet Prac 68:486–488
Borgsteede FHM, Deleeuw WA, Dijikstra T, Alsma G, de Vries W (1992) Ascaris suum infections causing clinical problems in cattle. Tijdschr Diergeneeskd 117(10):296–298
Boveri T (1887) Über Differenzierung der Zellkerne während der Furchung des Eies von Ascaris megalocephala. Anat Anz 2:688–693
Boveri T (1888) Zellen-Studien: Die Befruchtung und Teilung des Eies von Ascaris megalocephala. Zeitschrift Für Naturwissenschaften 22:685–882
Brown PJ, Clayton HM (1979) Hepatic pathology of experimental Parascaris equorum infection in worm-free foals. J Comp Path 89(1):115–123
Brown HD, Matzuk AR, Ilves IR, Peterson LH, Harris SA, Sarett LH, Egerton JR, Yakstis JJ, Campbell WC, Cuckler AC (1961) Antiparasitic drugs. IV. 2-(4′-Thiazolyl)-benzimidazole, a new anthelmintic. JACS 83(7):1764–1765
Bullini L, Nascetti G, Carrè S, Rumore F, Biocca E (1978) Ricerche cariologiche ed elettroforetiche su Parascaris univalens e Parascaris equorum. Atti della Accademia Nazionale dei Lincei. Classe di Scienze Fisiche, Matematiche e Naturali. Rendiconti Serie 8 65:151–156
Burk SV, Dangoudoubiyam S, Brewster-Barnes T, Bryant UK, Howe DK, Carter CN, Vanzant ES, Harmon RJ, Kazacos KR, Rossano MG (2014) In vitro culture of Parascaris equorum larvae and initial investigation of parasite excretory-secretory products. Parasitol Res 113(11):4217–4224
Burk SV, Dangoudoubiyam S, Brewster-Barnes T, Howe DK, Carter CN, Bryant UK, Rossano MG (2016) Equine antibody response to larval Parascaris equorum excretory-secretory products. Vet Parasitol 226:83–87
Campbell WC, Fisher MH, Stapley O, Albers-Schönberg G, Jacob TA (1983) Ivermectin: A potent new antiparasitic agent. Science 221(4613):823–828
Carmichael I, Visser R, Schneider D, Stoll M (1987) Haemonchus contortus resistance to ivermectin. J S Afr Vet Assoc 58(2):93
Carnoy JB (1886/1887) La cellule, recueil de cytologie et d’histologie générale. T. III. I. La vésicule germinative et les globules polaires chez divers nématodes. IL La segmentation chez les nématodes. Heft 1. 1886. Conférence donnée à la société belge de microscopic Les globules polaires de l’Ascaris clavata. Heft II. 1887
Carter GT, Neitsche JA, Hertz MR, Williams DR, Siegel MM, Morton GO, James JC, Borders JD (1988) LL-F28249 antibiotic complex: a new family of antiparasitic macrocyclic lactones. J Antibiot 41(4):519–529
Charvet CL, Guégnard F, Courtot E, Cortet J, Neveu C (2018) Nicotine-sensitive acetylcholine receptors are relevant pharmacological targets for the control of multidrug resistant parasitic nematodes. Int J Parasitol Drugs Drug Resist 8(3):540–549
Chelladurai JJ, Brewer MT (2019) Detection and quantification of Parascaris P-glycoprotein drug transporter expression with a novel mRNA hybridization technique. Vet Parasitol 267:75–83
Chelladurai JJ, Bader C, Snobl T, Magstadt D, Cooper V, Brewer MT (2015) Toxocara vitulorum infection in a cohort of beef calves in Iowa. Vet Parasitol 214(1–2):96–99
Choi YJ, Bisset SA, Doyle SR, Hallsworth-Pepin K, Martin J, Grant WN, Mitreva M (2017) Genomic introgression mapping of field-derived multiple-anthelmintic resistance in Teladorsagia circumcincta. PLoS Genet 13(6):e1006857
Cirak VY, Kar S, Girişgin O (2010) A survey on anthelmintic resistance in strongyles to ivermectin and pyrantel and macrocyclic lactone-resistance in Parascaris equorum. Turkiye Paraziol Derg 34(1):35–39
Clayton HM, Duncan JL (1978) Clinical signs associated with Parascaris equorum infection in worm-free pony foals and yearlings. Vet Parasitol 4(1):69–78
Clayton HM, Duncan JL (1979a) The migration and development of Parascaris equorum in the horse. Int J Parasitol 9(4):285–292
Clayton HM, Duncan JL (1979b) The development of immunity to Parascaris equorum infection in the foal. Res Vet Sci 26(3):383–384
Collins JB, Jordan B, Baldwin L, Hebron C, Paras K, Vidyaskankar AN, Kaplan RM (2019) Resistance to fenbendazole in Ascaridia dissimilis, an important nematode parasite of turkeys. Poult Sci 98(11):5412–5415
Collins JB, Jordan B, Bishop A, Kaplan RM (2021) Fenbendazole resistance in Heterakis gallinarum, the vector of Histomonas meleagridis, the causative agent of blackhead disease in poultry. bioRxiv Preprint. 03.30.437780
Cooper LG, Caffe G, Cerutti J, Nielsen MK, Anziani OS (2020) Reduced efficacy of ivermectin and moxidectin against Parascaris spp. in foals from Argentina. Vet Parasitol Reg Stud Rep 20:100388
Courtot E, Charvet CL, Beech RN, Harmache A, Wolstenholme AJ, Holden-Dye L, O’Connor V, Peineau N, Woods DJ, Neveu C (2015) Functional characterization of a novel class of morantel-sensitive acetylcholine receptors in nematodes. PLoS Path 11(12):e1005267
Cox FEG (2002) History of human parasitology. Clin Microbiol Rev 15(4):595–612
Craig TM, Diamond PL, Ferwerda NS, Thompson JA (2007) Evidence of ivermectin resistance by Parascaris equorum on a Texas horse farm. J Equine Vet Sci 27(2):67–71
Cribb NC, Cote NM, Boure LP, Peregrine AS (2006) Acute small intestinal obstruction associated with Parascaris equorum infection in young horses: 25 cases (1985–2004). N Z Vet J 54(6):338–343
Curtis SE, Tisch DA, Todd KS, Simon J (1987) Pulmonary bacterial deposition and clearance during ascarid larval migration in weanling pigs. Can J Vet Res 51(4):525–527
Deslyper G, Colgan TJ, Cooper AJR, Holland CV, Carolan JC (2016) A proteomic investigation of hepatic resistance to Ascaris in a murine model. PLoS Negl Trop Dis 10(8):e0004837
Deslyper G, Holland CV, Colgan TJ, Carolan JC (2019) The liver proteome in a mouse model for Ascaris suum resistance and susceptibility: evidence for an altered innate immune response. Parasit Vectors 12(1):402
Diawara A, Drake LJ, Suswillo RR, Kihara J, Bundy DAP, Scott ME, Halpenny C, Stothard JR, Prichard RK (2009) Assays to detect beta-tubulin codon 200 polymorphism in Trichuris trichiura and Ascaris lumbricoides. PLoS Negl Trop Dis 3(3):e397
Diawara A, Schwenkenbecher JM, Kaplan RM, Prichard RK (2013) Molecular and biological diagnostic test for monitoring benzimidazole resistance in human soil-transmitted helminths. Am J Top Med Hyg 88(6):1052–1061
Donoghue EM, Lyons ET, Bellaw JL, Nielsen MK (2015) Biphasic appearance of corticated and decorticated ascarid egg shedding in untreated horse foals. Vet Parasitol 214(1–2):114–117
Doyle SR, Cotton JA (2019) Genome-wide approaches to investigate anthelmintic resistance. Trends Parasitol 35(4):289–301
Drudge JH, Lyons ET (1966) Control of internal parasites of the horse. J Am Vet Med Assoc 148(4):378–383
Drudge JH, Szanto J, Wyant ZN, Elam G (1964) Field studies on parasite control in sheep: comparison of thiabendazole, ruelene, and phenothiazine. A J Vet Res 25:1512–1518
Eberhard ML, Alfano E (1998) Adult Toxocara cati infections in U.S. children: report of four cases. Am J Trop Med Hyg 59(3):404–406
Ellingson J, Coates-Markle L (1996) Managing your pregnant mare and her foal. Oregon State University Extension Service
ESCCAP (2019) A guide to the treatment and control of equine gastrointestinal parasite infections. In: European Scientific Counsel Companion Animal Parasites, 2nd edn. https://www.esccap.org/uploads/docs/rtjqmu6t_0796_ESCCAP_Guideline_GL8_v7_1p.pdf. Accessed July-December 2021.
Fabiani JV, Lyons ET, Nielsen MK (2016) Dynamics of Parascaris and Strongylus spp. parasites in untreated juvenile horses. Vet Parasitol 230:62–66
Faust EC (1937) The use of anthelmintics. JAMA 108(5):386–392
Fleischmann RD, Adams MD, White O, Clayton RA, Kirkness EF, Kerlavage AR, Bult CJ, Tomb JF, Dougherty BA, Merrick JM, McKenney K, Sutton G, Fitzhugh W, Fields C, Gocayne JD et al (1995) Whole-genome random sequencing and assembly of Haemophilus influenzae Rd. Science 269(5223):496–512
Fleming SA, Craig T, Kaplan RM, Miller JE, Navarre C, Rings M (2006) Anthelmintic resistance of gastrointestinal parasites in small ruminants. J Vet Intern Med 20(2):435–444
Friedman PA, Platzer EG (1978) Interaction of anthelmintic benzimidazoles and benzimidazole derivatives with bovine brain tubulin. Biochim Biophys Acta 544(3):605–614
Gakosso LGC, Baadi F, Abakka FZ, Basraoui D (2020) The visceral larva migrans caused by Toxocara canis: a case report. Pan Afr Med J 36:150
Gao JF, Zhang XX, Wang XX, Li Q, Li Y, Xu WW, Gao Y, Wang CR (2019) According to mitochondrial DNA evidence, Parascaris equorum and Parascaris univalens may represent the same species. J Helminthol 93(3):383–388
Geary TG, Thompson DP (2001) Caenorhabditis elegans: how good a model for veterinary parasites? Vet Parasitol 101(3–4):371–386
Geerts S, Gryseels B (2000) Drug resistance in human helminths: current situation and lessons from livestock. Clin Microbiol Rev 13(2):207–222
Geerts S, Gryseels B (2002) Anthelmintic resistance in human helminths: a review. Trop Med Int Health 6(11):915–921
Geerts S, Coles GC, Gryseels B (1997) Anthelmintic resistance in human helminths: learning from the problems with worm control in livestock. Parasitol Today 13(4):149–151
Gerhard AP, Krücken J, Heitlinger E, Janssen IJI, Besiaga M, Kornaś S, Beier C, Nielsen MK, Davis RE, Wang J, von Samson-Himmelstjerna G (2020) The P-glycoprotein repertoire of the equine parasitic nematode Parascaris univalens. Sci Rep 10(1):13586
Gerhard AP, Krücken J, Neveu C, Charvet CL, Harmache A, von Samson-Himmelstjerna G (2021) Pharyngeal pumping and tissue-specific transgenic P-glycoprotein expression influence macrocyclic lactone susceptibility in Caenorhabditis elegans. Pharmaceuticals 14(2):153
Getachew AM, Innocent GT, Trawford AF, Feseha G, Reid SJW, Love S (2008) Equine parascarosis under the tropical weather conditions of Ethiopia: a coprological and postmortem study. Vet Rec 162(6):177–180
Getachew M, Trawford A, Feseha G, Reid SWJ (2010) Gastrointestinal parasites of working donkeys in Ethiopia. Trop Anim Health Prod 42(1):27–33
Geurden T, Betsch JM, Maillard K, Vanimisetti B, D’Espois M, Besognet B (2013) Determination of anthelmintic efficacy against equine cyathostomins and Parascaris equorum in France. Equine Vet Ed 25:304–307
Ghisi M, Kaminsky R, Mäser P (2007) Phenotyping and genotyping of Haemonchus contortus isolates reveals a new putative candidate mutation for benzimidazole resistance in nematodes. Vet Parasitol 144(3–4):313–320
Goday C, Pimpinelli S (1984) Chromosome organization and heterochromatin elimination in Parascaris. Science 224(4647):411–413
Goday C, Pimpinelli S (1986) Cytological analysis of chromosomes in the two species Parascaris univalens and P. equorum. Chromosoma 94:1–10
Goday C, Ciofi-Luzzatto A, Pimpinelli S (1985) Centromere ultrastructure in germ-line chromosomes of Parascaris. Chromosoma 91(2):121–125
Goeze JAE (1782) Versuch einer Naturgeschichte der Eingeweidewürmer thierischer Körper., Ed. Papen, Blankenburg
Gundran RS, More SJ (1999) Health and growth of water-buffalo calves in Nueva Ecija, the Philippines. Prev Vet Med 40(2):87–100
Hamoir G (1992) The discovery of meiosis by E. Van Beneden, a breakthrough in the morphological phase of heredity. Int J Dev Biol 36(1):9–15
Han L, Lan T, Lu Y, Zhou M, Li H, Lu H, Wang Q, Li X, Du S, Guan C, Zhang Y, Sahu SK, Qian P, Zhang S, Zhou H et al (2022) Equus roundworms (Parascaris univalens) are undergoing rapid divergence while genes involved in metabolic as well as anthelminic resistance are under positive selection. BMC Genomics 23(1):489
Hansen EP, Fromm B, Andersen SD, Marcilla A, Andersen KL, Borup A, Williams AR, Jex AR, Gasser RB, Young ND, Hall RS, Stensballe A, Ovchinnikov V, Yan Y, Fredholm M et al (2019) Exploration of extracelluar vesicles from Ascaris suum provides evidence of parasite-host cross talk. J Extracell Vesicles 8(1):1578116
Harder A (2002) Chemotherapeutic approaches to nematodes: current knowledge and outlook. Parasitol Res 88(3):272–277
Harrow ID, Gration AF (1985) Mode of action of the anthelmintics morantel, pyrantel and levamisole on muscle cell membrane of the nematode Ascaris suum. Pestic Sci 16(6):662–672
Hautala K, Näreaho A, Kauppinen O, Nielsen MK, Sukura A, Rajala-Schultz PJ (2019) Risk factors for equine intestinal parasite infections and reduced efficacy of pyrantel embonate against Parascaris sp. Vet Parasitol 273:52–59
Hearn FPD, Peregrine AS (2003) Identification of foals infected with Parascaris equorum apparently resistant to ivermectin. J Am Vet Med Assoc 223(4):482–485
Hertwig O (1890) Vergleich der Ei- und Samenbildung bei Nematoden. Arch Mikrosk Anat 36:1–138
Hill IR, Denham DA, Scholtz CL (1985) Toxocara canis larvae in the brain of a British child. Trans R Soc Trop Med Hyg 79(3):351–354
Holland C (ed) (2013) Impact of Ascaris suum in livestock. In: Ascaris: the neglected parasite. Elsevier, Amsterdam, pp 363–382
Horton J (2003) Global anthelmintic chemotherapy programs: learning from history. Trend Parasitol 19(9):405–409
International Helminth Genomes Consortium (2019) Comparative genomics of the major parasitic worms. Nat Genet 51(1):163–174
Islam MK, Miyoshi T, Yokomizo Y, Tsuji N (2004) The proteome expression patterns in adult Ascaris suum under exposure to aerobic/anaerobic environments analyzed by two-dimensional electrophoresis. Parasitol Res 93(2):96–101
James CE, Hudson AL, Davey MW (2009) Drug resistance mechanisms in helminths: is it survival of the fittest? Trends Parasitol 25(7):328–335
Janssen IJI, Krücken J, Demeler J, Basiaga M, Kornaś S, von Samson-Himmelstjerna G (2013) Genetic variants and increased expression of Parascaris equorum P-glycoprotein-11 in populations with decreased ivermectin susceptibility. PLoS One 8(4):e61635
Janssen IJI, Krücken J, Demeler J, von Samson-Himmelstjerna G (2015) Transgenically expressed Parascaris P-glycoprotein-11 can modulate ivermectin susceptibility in Caenorhabditis elegans. Int J Parasitol Drugs Drug Resist 5(2):44–47
Jimenez Castro PD, Howell SB, Schaefer JJ, Avramenko RW, Gilleard JS, Kaplan RM (2019) Multiple drug resistance in the canine hookworm Ancylostoma caninum: an emerging threat? Parasit Vectors 12(1):576
Jimenez Castro PD, Venkatesan A, Redman E, Chen R, Malatesta A, Huff H, Zuluaga Salazar DA, Avramenko R, Gilleard JS, Kaplan RM (2021) Multiple drug resistance in hookworms infecting greyhound dogs in the USA. Int J Parasitol Drugs Drug Resist 17:107–117
Jones WE (1985) Anthelmintic rotation. Equine Vet Data 6:97–100
Kaplan RM (2020) Biology, epidemiology, diagnosis, and management of anthelmintic resistance in gastrointestinal nematodes of livestock. Vet Clin North Am Food Anim Pract 36(1):17–30
Kaplan RM, Vidyashankar AN (2012) An inconvenient truth: global worming and anthelmintic resistance. Vet Parasitol 186(1–2):70–78
Kaufmann F, Das G, Sohnrey B, Gauly M (2011) Helminth infections in laying hens kept in organic free range systems in Germany. Livest Sci 141(2–3):182–187
Kelly TG, Madhavan VL, Peters JM, Kazacos KR, Silvera VM (2012) Spinal cord involvement in a child with raccoon roundworm (Baylisascaris procyonis) meningoencephalitis. Pediatr Radiol 42(3):369–373
Khan S, Nisar A, Yuan J, Luo X, Dou X, Liu F, Zhao X, Li J, Ahmad H, Mehmood SA, Feng X (2020) A whole genome re-sequencing based GWA analysis reveal candidate genes associated with ivermectin resistance in Haemonchus contortus. Genes 11(4):367
Kim SB, Goodfellow M (2002) Streptomyces avermitilis sp. nov., nom. rev., a taxonomic home for the avermectin-producing streptomycetes. Int J Syst Evol Micr 52(6):2011–2014
Kojima G, Usuki S, Mizokami K, Tanabe M, Machi J (2013) Intestinal anisakiasis as a rare cause of small bowel obstruction. Am J Emerg Med 31(9):1422.e1-1422.e2
Kotze AC, Prichard AK (2016) Anthelmintic resistance in Haemonchus contortus: history, mechanisms and diagnosis. Adv Parasitol 93:397–428
Kotze AC, Cowling K, Bagnall NH, Hines BM, Ruffell AP, Hunt PW, Coleman GT (2012) Relative level of thiabendazole resistance associated with the E198A and F200Y SNPs in larvae of a multi-drug resistant isolate of Haemonchus contortus. Int J Parasitol Drugs Drug Resist 2:92–97
Kotze AC, Hunt PW, Skuce P, von Samson-Himmelstjera G, Martin RJ, Sager H, Krücken J, Hodgkinson J, Lespine A, Jex AR, Gilleard JS, Beech RN, Wolstenholme AJ, Demeler J, Robertson A et al (2014) Recent advances in candidate-gene and whole-genome approaches to the discovery of anthelmintic resistance markers and the description of drug/receptor interactions. Int J Parasitol Drugs Drug Resist 4(3):164–184
Krücken J, Fraundorfer K, Mugisha JC, Ramünke S, Sifft KC, Geus D, Habarugira F, Ndoli J, Sendegeya A, Mukampunga C, Bayingana C, Aebischer T, Demeler J, Gahutu JB, Mockenhaupt FP, von Samson-Himmelstjerna G (2017) Reduced efficacy of albendazole against Ascaris lumbricoides in Rwandan schoolchildren. Int J Parasitol Drugs Drug Resist 7(3):262–271
Lacey E (1988) The role of the cytoskeletal protein, tubulin, in the mode of action and mechanism of drug resistance to benzimidazoles. Int J Parasitol 18(7):885–936
Lacey E (1990) Mode of action of benzimidazoles. Parasitol Today 6(4):112–115
Lacey E, Gill JH (1994) Biochemistry of benzimidazole resistance. Acta Trop 56(2–3):245–262
Laing R, Maitland K, Lecová L, Skuce PJ, Tait A, Devaney E (2016) Analysis of putative resistance gene loci in UK field populations of Haemonchus contortus after 6 years of macrocyclic lactone use. Int J Parasitol 46(10):621–630
Lassen B, Peltola SM (2014) Anthelmintic resistance of intestinal nematodes to ivermectin and pyrantel in Estonian horses. J Helminthol 89(6):760–763
Laugier C, Sevin C, Ménard S, Maillard K (2012) Prevalence of Parascaris equorum infection in foals on French stud farms and first report of ivermectin-resistant P. equorum populations in France. Vet Parasitol 188(1–2)185–189
Lem MF, Vincent KP, Pone JW, Joseph T (2012) Prevalence and intensity of gastro-intestinal helminths in horses in the Sudano-Guinean climatic zone of Cameroon. Trop Parasitol 2(1):45–48
Lespine A, Ménez C, Bourguinat C, Prichard RK (2011) P-glycoproteins and other multidrug resistance transporters in the pharmacology of anthelmintics: Prospects for reversing transport-dependent anthelmintic resistance. Int J Parasitol Drugs Drug Resist 2:58–75
Li JC (1937) A six-chromosome Ascaris in Chinese horses. Science 86(2222):101–102
Li L, Lü L, Nadler SA, Gibson DI, Zhang LP, Chen HX, Zhao WT, Guo YN (2018) Molecular phylogeny and dating reveal a terrestrial origin in the early carboniferous for Ascaridoid nematodes. Syst Biol 67(5):888–900
Lightowlers MW, Rickard MD (1988) Excretory–secretory products of helminth parasites: effects on host immune responses. Parasitology 96:S123–S166
Lin TP (1954) The chromosomal cycle in Parascaris equorum (Ascaris megalocephala); oogenesis and diminution. Chromosoma 6(3):175–198
Lind EO, Christensson D (2009) Anthelmintic efficacy on Parascaris equorum in foals on Swedish studs. Acta Vet Scand 51(1):45
Lindgren K, Ljungvall Ö, Nilsson O, Ljungström BL, Lindahl C, Höglund J (2008) Parascaris equorum in foals and in their environment on a Swedish stud farm, with notes on treatment failure of ivermectin. Vet Parasitol 151(2–4):337–343
Linnaeus C (1758) Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Holmiæ. (Salvius)
Liu GH, Nadler SA, Liu SS, Podolska M, D’Amelio S, Shao R, Gasser RB, Zhu XQ (2016) Mitochondrial phylogenomics yields strongly supported hypotheses for Ascaridomorph nematodes. Sci Rep 6:39248
Lubega GW, Prichard RK (1990) Specific interaction of benzimidazole anthelmintics with tubulin: high-affinity binding and benzimidazole resistance in Haemonchus contortus. Mol Biochem Parasitol 38(2):221–232
Lubega GW, Klein RD, Geary TG, Prichard RK (1994) Haemonchus contortus: the role of two beta-tubulin gene subfamilies in the resistance to benzimidazole anthelmintics. Biochem Pharmacol 47(9):1705–1715
Lyons ET, Drudge JH, Tolliver SC (1990) Prevalence of some internal parasites found (1971–1989) in horses born on a farm in Central Kentucky. J Equine Vet Sci 10(2):99–107
Lyons ET, Tolliver SC, Drudge JH, Granstrom DE, Collins SS, Stamper S (1992) Critical and controlled tests of activity of moxidectin (CL 301,423) against natural infections of internal parasites of equids. Vet Parasitol 41(3–4):255–284
Lyons ET, Tolliver SC, Ionita M, Collins SS (2008) Evaluation of parasiticidal activity of fenbendazole, ivermectin, oxibendazole, and pyrantel pamoate in horse foals with emphasis on ascarids (Parascaris equorum) in field studies on five farms in Central Kentucky in 2007. Parasitol Res 103(2):287–291
Lyons ET, Tolliver SC, Kuzmina TA, Collins SS (2011) Further evaluation in field tests of the activity of three anthelmintics (fenbendazole, oxibendazole, and pyrantel pamoate) against the ascarid Parascaris equorum in horse foals on eight farms in Central Kentucky (2009–2010). Parasitol Res 109(4):1193–1197
Malekpour SH, Rakhshandehroo E, Yekaseresht A (2019) Molecular characterization of β-tubulin gene associated with benzimidazole resistance in larvae of field isolates of Parascaris (Nematoda: Ascarididae). J Parasi Dis 43(4):672–678
Martin F, Höglund J, Bergström TF, Lindsjö OK, Tydén E (2018) Resistance to pyrantel embonate and efficacy of fenbendazole in Parascaris univalens on Swedish stud farms. Vet Parasitol 264:69–73
Martin F, Dube F, Lindsjö OK, Eydal M, Höglund J, Bergström TF, Tydén E (2020) Transcriptional responses in Parascaris univalens after in vitro exposure to ivermectin, pyrantel citrate and thiabendazole. Parasit Vectors 13(1):342
Martin F, Eyda M, Höglund J, Tydén E (2021a) Constitutive and differential expression of transport protein genes in Parascaris univalens larvae and adult tissues after in vitro exposure to anthelmintic drugs. Vet Parasitol 298:109535
Martin F, Halvarsson P, Delhomme N, Höglund J, Tydén E (2021b) Exploring the β-tubulin gene family in a benzimidazole-resistant Parascaris univalens population. Int J Parasitol Drugs Drug Resist 17:84–91
Martin F, Svansson V, Eydal M, Oddsdóttir C, Ernback M, Persson I, Tydén E (2021c) First report of resistance to ivermectin in Parascaris univalens in Iceland. J Parasitol 107(1):16–22
Mateus TL, Cruzeiro ALF, Rocha H, Vieira-Pino MM (2015) Economic impact of hepatic rejections caused by Ascaris suum in swine during post mortem inspection at slaughterhouse. In: Safepork 2015 Proceedings Book: epidemiology and control of hazards in pork production chain, pp 269–272
Matthews JB (2014) Anthelmintic resistance in equine nematodes. Int J Parasitol Drugs Drug Resist 4(3):310–315
Molento MB, Antunes J, Bentes RN, Coles GC (2008) Anthelmintic resistant nematodes in Brazilian horses. Vet Rec 162(12):384–385
Morsy K, Fahmy S, Mohamed A, Ali S, El-Garhy M, Shazly M (2019) Optimizing and evaluating the antihelminthic activity of the biocompatible zinc oxide nanoparticles against the ascaridid nematode, Parascaris Equorum in Vitro. Acta Parasitol 64(4):873–886
Mulcahy G, O’Neill S, Fanning J, McCarthy E, Sekiya M (2005) Tissue migration by parasitic helminths – an immunoevasive strategy? Trends Parasitol 21(6):273–277
Muller F, Tobler H (2000) Chromatin diminution in the parasitic nematodes Ascaris suum and Parascaris univalens. Int J Parasitol 30(4):391–399
Mullis KB, Faloona FA (1987) Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol 155:335–350
Nadler SA (1987) Biochemical and immunological systematics of some Ascaridoid nematodes: genetic divergence between congeners. J Parasitol 73(4):811–816
Nadler SA, Hudspeth DS (2000) Phylogeny of the Ascaridoidea (Nematoda: Ascaridida) based on three genes and morphology: hypotheses of structural and sequence evolution. J Parasitol 86(2):380–393
Nadler SA, Carreno RA, Mejía-Madrid H, Ullberg J, Pagan C, Houston R, Hugot JP (2007) Molecular phylogeny of clade III nematodes reveals multiple origins of tissue parasitism. Parasitology 134(Pt 10):1421–1442
Näreaho A, Vainio K, Oksanen A (2011) Impaired efficacy of ivermectin against Parascaris equorum, and both ivermectin and pyrantel against strongyle infections in trotter foals in Finland. Vet Parasitol 182(2–4):372–377
Nejsum P, Parker ED Jr, Frydenberg J, Roepstorff A, Boes J, Haque R, Astrup I, Prag J, Sørensen UBS (2005) Ascariasis is a zoonosis in Denmark. J Clin Microbiol 43(3):1142–1148
Nicholls JM, Clayton HM, Pirie HM, Duncan JL (1978) A pathological study of the lungs of foals infected experimentally with Parascaris equorum. J Comp Pathol 88(2):261–274
Niedermaier J, Moritz KB (2000) Organization and dynamics of satellite and telomere DNAs in Ascaris: implications for formation and programmed breakdown of compound chromosomes. Chromosoma 109(7):439–452
Nielsen MK, Wang J, Davis R, Bellaw JL, Lyons ET, Lear TL, Goday C (2014) Parascaris univalens – a victim of large-scale misidentification? Parasitol Res 113(12):4485–4490
Nielsen MK, Branan MA, Wiedenheft AM, Digianantonio R, Garber LP, Kopral CA, Phillippi-Taylor AM, Traub-Dargatz JL (2018) Parasite control strategies used by equine owners in the United States: a national survey. Vet Parasitol 250:45–51
Nielsen MK, Gee EK, Hansen A, Waghorn T, Bell J, Leathwick DM (2021) Monitoring equine ascarid and cyathostomin parasites: Evaluating health parameters under different treatment regimens. Equine Vet J 53(5):902–910
Nielsen MK, Mittel L, Grice A, Erskine M, Graves E, Vaala W, Tully RC, French DD, Bowman R, Kaplan RM (2019) AAEP parasite control guidelines. American Association of Equine Practitioners
Nielsen MK (2016) Evidence-based considerations for control of Parascaris spp. infections in horses. Equine Vet Ed 28(4):224–231
Nobel Prize (2015) The Nobel Prize in Physiology or Medicine 2015. The Nobel Prize. https://www.nobelprize.org/prizes/medicine/2015/press-release/. Accessed July-December 2021.
Özben M, von Samson-Himmelstjerna G, Freiin von Streit, MKB, Wilkes, EJA, Hughes, KJ, Krücken, J (2022) Absence of polymorphisms in codons 167, 198 and 200 of all seven β-tubulin isotypes of benzimidazole susceptible and resistant Parascaris spp. specimens from Australia. Pathogens 11(5):490
Palma A, Matamoros G, Escobar D, Sánchez AL, Fontecha G (2020) Absence of mutations associated with resistance to benzimidazole in the beta-tubulin gene of Ascaris suum. Rev De Soc Bras Med Trop 53:e20190155
Pape M, Posedi J, Failing K, Schnieder T, von Samson-Himmelstjerna G (2003) Analysis of the beta-tubulin codon 200 genotype distribution in a benzimidazole-susceptible and -resistant cyathostome population. Parasitology 127(Pt 1):53–59
Peng Z, Shen D, Zhang D, Li X, Wang X, Zhai Q, Hou Z, Li H (2019) Genetic characteristics and phylogenetic relationship of Parascaris spp. from Equus zebra, E. caballus, and E. asinus. Vet Parasitol 271:76–79
Prichard R (1994) Anthelmintic resistance. Vet Parasitol 54(1–3):259–268
Rakhshandehroo E, Asadpour M, Jafari A, Malekpour SH (2016) The effectiveness of Cinnamomum zeylanicum, Punica granatum flower and Capsicum annuum extracts against Parascaris equorum infective larvae. Acta Vet Eurasia 42:132–137
Rakhshandehroo E, Asadpour M, Malekpour SH, Jafari A (2017) The anthelmintic effects of five plant extracts on the viability of Parascaris equorum larvae. Equine Vet Ed 29(4):219–224
Rashwan N, Scott M, Prichard R (2017) Rapid genotyping of β-tubulin polymorphisms in Trichuris trichiura and Ascaris lumbricoides. PLoS Negl Trop Dis 11(1):e0005205
Read AF, Skorping A (1995) The evolution of tissue migration by parasitic nematode larvae. Parasitology 111(Pt 3):359–371
Reinemeyer CR (2009) Diagnosis and control of anthelmintic-resistant Parascaris equorum. Parasit Vectors 2(Suppl 2):S8
Reinemeyer CR, Nielsen MK (2017) Control of helminth parasites in juvenile horses. Equine Vet Ed 29(4):225–232
Relf VE, Lester HE, Morgan ER, Hodgkinson JE, Matthews JB (2014) Anthelmintic efficacy on UK Thoroughbred stud farms. Int J Parasitol 44(8):507–514
Rendle D, Austin C, Bowen M, Cameron I, Furtado T, Hodgkinson J, McGorum B, Matthews J (2019) Equine de-worming: a consensus on current best practice. UK-Vet Equine 3:1–14
Rezansoff AM, Lain R, Gilleard JS (2016) Evidence from two independent backcross experiments supports genetic linkage of microsatellite Hcms8a20, but not other candidate loci, to a major ivermectin resistance locus in Haemonchus contortus. Int J Parasitol 46(10):653–661
Robert M, Hu W, Nielsen MK, Stowe CJ (2015) Attitudes towards implementation of surveillance-based parasite control on Kentucky thoroughbred farms – current strategies, awareness and willingness-to-pay. Equine Vet J 47(6):694–700
Robertson SJ, Pennington AJ, Evans AM, Martin RJ (1994) The action of pyrantel as an agonist and an open channel blocker at acetylcholine receptors in isolated Ascaris suum muscle vesicles. Eur J Pharmacol 272(2–3):273–282
Rose H, Rinaldi L, Bosco A, Mavrot F, de Waal T, Skuce P, Charlier J, Torgerson PR, Hertzberg H, Hendrickx G, Vercruysse J, Morgan ER (2015) Widespread anthelmintic resistance in European farmed ruminants: a systematic review. Vet Rec 176(21):546
Saffra NA, Perlman JE, Desai RU, Kazacos KR, Coyle CM, Machado FS, Kedhar SR, Engelbert M, Tanowitz HB (2010) Baylisascaris procyonis induced diffuse unilateral subacute neuroretinitis in New York City. J Neuroparasitology 1:N100401
Santschi EM, Slone DE, Embertson RM, Clayton MK, Markel MD (2000) Colic surgery in 206 juvenile thoroughbreds: survival and racing results. Equine Vet J Suppl 32:32–36
Saunders GI, Wasmuth JD, Beech R, Laing R, Hunt M, Naghra H, Cotton JA, Berriman M, Britton C, Gilleard JS (2013) Characterization and comparative analysis of the complete Haemonchus contortus β-tubulin gene family and implications for benzimidazole resistance in strongylid nematodes. 43(6):465–475
Scare JA, Dini P, Norris JK, Steuer AE, Scoggin K, Gravatte HS, Howe DK, Sulsarewicz P, Nielsen MK (2020) Ascarids exposed: a method for in vitro drug exposure and gene expression analysis of anthelmintic naïve Parascaris spp. Parasitology 147(6):659–666
Scare JA, Steuer AE, Shaffer CL, Slusarewicz P, Mousley A, Nielsen MK (2019) Long live the worms: methods for maintaining and assessing the viability of intestinal stages of Parascaris spp. in vitro. Parasitology 146(5):685–693
Schougaard H, Nielsen MK (2007) Apparent ivermectin resistance of Parascaris equorum in Danish foals. Vet Rec 160(13):439–440
Sharma N, Hunt PW, Hine BC, Ruhnke I (2019) The impacts of Ascaridia galli on performance, health, and immune responses of laying hens: new insights into an old problem. Poul Sci 98(12):6517–6526
Slocombe JOD, de Gannes RVG, Lake MC (2007) Macrocyclic lactone resistant Parascaris equorum on stud farms in Canada and effectiveness of fenbendazole and pyrantel pamoate. Vet Parasitol 145(3–4):371–376
Sohn WM, Na BK, Kim TH, Park TJ (2015) Anisakiasis: Report of 15 gastric cases caused by Anisakis Type I larvae and a brief review of Korean Anisakiasis cases. Korean J Parasitol 54(4):465–470
Sotilli J, Robinson MW, Kimber MJ, Cucher M, Ancarola ME, Nejsum P, Marcilla A, Eichenberger RM, Tritten L (2020) The protein and microRNA cargo of extracellular vesicles from parasitic helminths – current status and research priorities. Int J Parasitol 50(9):635–645
Southwood LL, Ragle CA, Snyder S, Hendrickson DA (1996) Surgical treatment of ascarid impactions in horses and foals. Proc Am Assoc Equine Practnrs 42:258–261
Stewart TB, Hale OM (1988) Losses to internal parasites in swine production. J Anim Sci 66(6):1548–1554
Stoneham S, Coles G (2006) Ivermectin resistance in Parascaris equorum. The Vet Rec 158(16):572
Studzińska MB, Sallé G, Roczeń-Karczmarz M, Szczepaniak K, Demkowska-Kutrzepa M, Tomczuk K (2020) A survey of ivermectin resistance in Parascaris species infected foals in south-eastern Poland. Acta Vet Scand 62(1):28
Sutherland IA, Leathwick DM (2011) Anthelmintic resistance in nematode parasites of cattle: a global issue? Trends Parasitol 27(4):176–181
Takiguchi Y, Mishima H, Okuda M, Terao M, Aoki A, Fukuda R (1980) Milbemycins, a new family of macrolide antibiotcs: fermentation, isolation, and physico-chemical properties. J Antibiot 33(10):1120–1127
Tatz AJ, Segev G, Steinman A, Berlin D, Milgram J, Kelmer G (2012) Surgical treatment for acute small intestinal obstruction caused by Parascaris equorum infection in 15 horses (2002–2011). Equine Vet J Suppl 43:111–114
Tchou S (1937) Une nouvelle race chinoise d’Ascaris megalocephala (type trivalens). Comptes Rendus De L’académie Des Sciences 204:1676–1677
Thapa S, Hinrichsen LK, Brenninkmeyer C, Gunnarsson S, Heerkens JL, Verwer C, Niebuhr K, Willet A, Grilli G, Thamsborg SM, Sørensen JT (2015) Prevalence and magnitude of helminth infections in organic laying hens (Gallus gallus domesticus) across Europe. Vet Parasitol 214(1–2):118–125
Thompson RC, Roberts MG (2001) Does pet helminth prophylaxis increase the rate of selection for drug resistance? Trends Parasitol 17(12):576–578
Tinkler SH (2020) Preventive chemotherapy and anthelmintic resistance of soil-transmitted helminths - can we learn nothing from veterinary medicine? One Health 9:100106
Townsend LB, Wise DS (1990) The synthesis and chemistry of certain anthelmintic benzimidazoles. Parasitol Today 6(4):107–112
Trailovic SM, Rajkovic M, Marjanovic DS, Neveu C, Charvet CL (2021) Action of carvacrol on Parascaris sp. and antagonistic effect on nicotinic acetylcholine receptors. Pharmaceuticals 14(6):505
Tydén E, Engström A, Morrison DA, Höglund J (2013a) Sequencing of the β-tubulin genes in the ascarid nematodes Parascaris equorum and Ascaridia galli. Mol Biochem 190(1):38–43
Tydén E, Morrison DA, Engström A, Nielsen MK, Edal M, Höglund J (2013b) Population genetics of Parascaris equorum based on DNA fingerprinting. Infect Genet Evol 13:236–241
Tydén E, Dahlberg J, Karlberg O, Höglund J (2014) Deep amplicon sequencing of preselected isolates of Parascaris equorum in β-tubulin codons associated with benzimidazole resistance in other nematodes. Parasit Vectors 7:410
Tydén E, Skarin M, Andersson-Franko M, Sjöblom M, Höglund J (2016) Differential expression of β-tubulin isotypes in different life stages of Parascaris spp after exposure to thiabendazole. Mol Biochem Parasitol 205(1–2):22–28
Urban Jr JF, Nielsen MK, Gazzola D, Xie Y, Beshah E, Hu Y, Li H, Rus F, Flanagan K, Draper A, Vakalapudi S, Li RW, Ostroff GR, Aroian RV (2021) An inactivated bacterium (paraprobiotic) expressing Bacillus thuringiensis Cry5B as a therapeutic for Ascaris and Parascaris spp. infections in large animals. One Health 12:100241
van Beneden E (1883) Recherches sur la Maturation de L’Oeuf, la Fécondation et la Division Cellulaire. Clemm, Gand & Leipzig. https://archive.org/details/recherchessurla00benegoog
van Loon JPAM, Visser EMS, de Mik-van MM, Kerbert P, Huppes T, Menke ES (2020) Colic surgery in horses: a retrospective study into short- and long-term survival rate, complications and rehabilitation toward sporting activity. J Equine Vet Sci 90:103012
van Wyk JA, Malan FS, Gerber HM, Alves RM (1987) Two field strains of Haemonchus contortus resistant to rafoxanide. Onderstepoort J Vet Res 54(2):143–146
Vercruysse J, Harrids EA, Kaboret Y, Pangui LJ, Gibson DI (1986) Gastrointestinal helminths of donkeys in Burkina Faso. Z Parasitenkd 72:821–825
Vercruysse J, Albonico M, Behnke JM, Kotze AC, Prichard RK, McCarthy JS, Montresor A, Levecke B (2011) Is anthelmintic resistance a concern for the control of human soil-transmitted helminths? Int J Parasitol Drugs Drug Resist 1(1):14–27
Veronesi F, Moretta I, Moretti A, Fioretti DP, Genchi C (2009) Field effectiveness of pyrantel and failure of Parascaris equorum egg count reduction following ivermectin treatment in Italian horse farms. Vet Parasitol 161(1–2):138–141
Veronesi F, Fioretti DP, Genchi C (2010) Are macrocyclic lactones useful drugs for the treatment of Parascaris equorum infections in foals? Vet Parasitol 172(1–2):164–167
Volk N, Tormey L (2017) Not just globetrotters: zoonotic transmission of Ascaris suum in Vermont. Am J Gastroenterol 112:S1373
von Samson-Himmelstjerna G (2012) Anthelmintic resistance in equine parasites – detection, potential clinical relevance and implications for control. Vet Parasitol 185(1):2–8
von Samson-Himmelstjerna G, Buschbaum S, Wirtherle N, Pape M, Schnieder T (2003) TaqMan minor groove binder real-time PCR analysis of β-tubulin codon 200 polymorphism in small strongyles (Cyathostomin) indicates that the TAC allele is only moderately selected in benzimidazole-resistant populations. Parasitology 127(Pt 5):489–496
von Samson-Himmelstjerna G, Blackhall WJ, McCarthy JS, Skuce PJ (2007a) Single nucleotide polymorphism (SNP) markers for benzimidazole resistance in veterinary nematodes. Parasitology 134(Pt 8):1077–1086
von Samson-Himmelstjerna G, Fritzen B, Demeler J, Schurmann S, Rohn K, Schnieder T, Epe C (2007b) Cases of reduced cyathostomin egg-reappearance period and failure of Parascaris equorum egg count reduction following ivermectin treatment as well as survey on pyrantel efficacy on German horse farms. Vet Parasitol 144(1–2):74–80
von Samson-Himmelstjerna G, Janssen IJI, Ramünke S, Goday C, Borges FA, Koudela B, Niedźwiedź A, Tomczuk K, Studzińska B, Kornas S, Krücken J (2021a) Very low intraspecific sequence variation in selected nuclear and mitochondrial Parascaris univalens genes. Infect Genet Evol 95:105035
von Samson-Himmelstjerna G, Thompson RCA, Krücken J, Grant W, Bowman DD, Schnyder M, Deplazes P (2021b) Spread of anthelmintic resistance in intestinal helminths of dogs and cats is currently less pronounced than in ruminants and horses – yet it is of major concern. Int J Parasitol Drugs Drug Resist 17:36–45
Wang J (2021) Genomics of the parasitic nematode Ascaris and its relatives. Genes 12(4):493
Wang J, Davis RE (2014) Programmed DNA elimination in multicellular organisms. Curr Op Genet Dev 27:26–34
Wang J, Gao S, Mostovoy Y, Kang Y, Zagoskin M, Sun Y, Zhang B, White LK, Easton A, Nutman TB, Kwok PY, Hu S, Nielsen MK, Davis DE (2017) Comparative genome analysis of programmed DNA elimination in nematodes. Genome Res 27(12):2001–2014
Wells MM (1924) Ascaris megalocephala. General Biological Supply House, Chicago
Whittaker JH, Carlson SA, Jones DE, Brewer MT (2017) Molecular mechanisms for anthelmintic resistance in strongyle nematode parasites of veterinary importance. J Vet Pharmacol Ther 40(2):105–115
Wilkes EJA, McConaghy FF, Thompson RL, Dawson K, Sangster NC, Hughes KJ (2017) Efficacy of a morantel–abamectin combination for the treatment of resistant ascarids in foals. Aust Vet J 95(3):85–88
Wolstenholme AJ (2012) Glutamate-gated chloride channels. J Biol Chem 287(48):40232–40238
Xinou E, Lefkoupoulos A, Gelagoti M, Drevelegas A, Diakou A, Milonas I, Dimitriadis AS (2003) CT and MR imaging findings in cerebral toxocaral disease. ANJR Am J Neuroradiol 24(4):714–718
Xu M, Molento M, Blackhall W, Ribeiro P, Beech R, Prichard R (1998) Ivermectin resistance in nematodes may be cause by alteration of P-glycoprotein homolog. Mol Biochem Parasitol 91(2):327–335
Xu MJ, Fu JH, Nisbet AJ, Huang SY, Zhou DH, Lin RQ, Song HQ, Zhu XQ (2013) Comparative profiling of microRNAs in male and female adults of Ascaris suum. Parasitol Res 112(3):1189–1195
Yazwinksi TA, Tucker CA, Wray E, Jones L, Clark FD (2013) Observations of benzimidazole efficacies against Ascaridia dissimilus, Ascaridia galli, and Heterakis gallinarum in naturally infected poultry. J Appl Poult Res 22(1):75–79
Yoshihara S, Nakagawa M, Suda H, Ikeda K, Hanashiro K (1983) White spots of the liver in pigs experimentally infected with Ascaris suum. Natl Inst Anim Health Q 23(4):127–137
Zakeri A, Whitehead BJ, Stensballe A, de Korne C, Williams AR, Everts B, Nejsum P (2021) Parasite worm antigens instruct macrophages to release immunoregulatory extracellular vesicles. J Extracell Vesicles 10(10):e12131
Zibaei M, Sadjjadi SM, Jahadi-Hosseini SH (2014) Toxocara cati larvae in the eye of a child: a case report. Asian Pac J Trop Biomed 4(Suppl 1):S53–S55
Zuccherato LW, Furtado LF, Medeiros CdS, Pinheiro CdS, Rabelo ÉM (2018) PCR-RFLP screening of polymorphisms associated with benzimidazole resistance in Necator americanus and Ascaris lumbricoides from different geographical regions in Brazil. PLoS Negl Trop Dis 12(9):e0006766
Author information
Authors and Affiliations
Contributions
Jennifer L. Cain conceptualized the work, performed the literature search and data analysis, and drafted and revised the work. Martin K. Nielsen revised the work and acted as supervisor.
Corresponding author
Ethics declarations
Ethics approval
N/A.
Consent to participate
N/A.
Consent for publication
N/A.
Conflict of interest
Dr. Martin Nielsen holds stock in and Jennifer Cain is an employee of Parasight System, Inc., a company that is manufacturing an automated parasite egg counting technique.
Additional information
Section Editor: Abdul Jabbar
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Cain, J.L., Nielsen, M.K. The equine ascarids: resuscitating historic model organisms for modern purposes. Parasitol Res 121, 2775–2791 (2022). https://doi.org/10.1007/s00436-022-07627-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-022-07627-z