
Abstract
To date, 12 members of the human ABCA subfamily are identified. They share a high degree of sequence conservation and have been mostly related with lipid trafficking in a wide range of body locations. Mutations in some of these genes have been described to cause severe hereditary diseases related with lipid transport, such as fatal surfactant deficiency or harlequin ichthyosis. In addition, most of them are hypothesized to participate in the subcellular sequestration of drugs, thereby being responsible for the resistance of several carcinoma cell lines against drug treatment. The objective of this review is to summarize the literature for this subfamily of ABC transporter proteins, excluding ABCA1 and ABCA4, which will be discussed in other chapters of this issue.
Similar content being viewed by others

In 1994, aiming at the identification of new members of the recently described ABC transporter family, Luciani et al. [32] designed a PCR approach based on the high degree of homology within this family of proteins. This resulted in the identification of two novel ABC transporter proteins, named ABC1 and ABC2, present on the human genome in very close locations on chromosome 9. This led to the establishment of a new subfamily for their classification, the ABCA subfamily.
This subfamily contains the largest ABC proteins known to date, some of them over 2,100 amino acids long and more than 200 kDa of predicted molecular weight. Excluding ABCA14–A17 proteins that have been only cloned in rodents [5, 12], the human ABCA subfamily is composed of 12 members and divided into two subgroups. One subgroup is formed by five genes, ABCA5, A6, A8, A9, and A10, that are located in a cluster on human chromosome 17 [4]. In addition to this special gene arrangement, the five members of this cluster present a series of structural differences not only as compared to the second subgroup of the A-family of ABC transporters, that is located dispersedly in the genome, but also in comparison to other known ABC transporters. The objective of this review is to summarize the literature for the members of the ABCA subfamily, excluding ABCA1 and ABCA4, which will be discussed in separate chapters of this issue. Information about gene and protein structures, function, tissue expression, and potential associations with human diseases will be specified for each ABCA transporter in the respective paragraphs. The main characteristics of all ABCA genes are presented in Table 1.
ABCA2
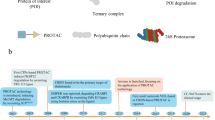
The human ABCA2 protein consists of 2,436 amino acids and has a molecular mass of approximately 250 kDa [22]. As mentioned above, ABCA2 is located on chromosome 9q close to ABCA1, a protein involved in cellular cholesterol efflux (see review by G. Chimini in this issue). Both genes share a high degree of homology, strongly suggesting a duplication event during evolution [32]. Interestingly, the ABCA2 gene presents an unusually compact structure, spanning only 21 kb, instead of the average 32–250 kb of other full-transporters [22]. The protein structure of ABCA2 is also similar to ABCA1: the protein contains two symmetrical halves and, between them, a long cytoplasmic regulatory domain interrupted by a stretch of highly hydrophobic residues (HH1; Fig. 1) that dips into the membrane [32, 48]. This highly hydrophobic segment represents a special structural feature of the members of the ABCA subfamily (Fig. 1).

Gene (a) and protein (b) structures of ABC transporters included in the A-subfamily. The two halves of the protein, each containing a transmembrane domain (TMD) and an ATP-binding cassette, are separated by a characteristic hydrophobic loop (HH1, arrow) that dips into the membrane. The NBDs and HH1 of all ABCA-subfamily members are encoded by 4–5 and 1–2 exons, respectively
Being predominantly expressed in the brain [48], ABCA2 has a unique expression profile within this subfamily. In the promoter region of this gene, target sites important in neural development, differentiation, and macrophage activation were identified [22]. ABCA2 is located in lysosomes where it is thought to play an important role in neuronal lipid transport. This hypothesis was tested in the rat in several studies by the group of Zhou and Inagaki [45, 55–57]. High expression of ABCA2, as a lysosome-associated protein, was found only in the cell bodies of oligodendrocytes [56] and solely in restricted regions of the newborn spinal cord. ABCA2 expression colocalized in time and space with myelination markers (i.e., O4 and myelin basic protein) [57] and appearance of thick myelin segments [45]. Recent works indicate a relationship between ABCA2 and Alzheimer’s disease (AD). Colocalization with beta-amyloid peptide and other AD markers was reported [11], and one synonymous SNP in human ABCA2 was significantly associated with early-onset development of this disease [33]. As AD is associated with lipid transport disorders and abnormal myelination, it is very likely that ABCA2 is implicated in controlling lipid trafficking from the neuronal cell body to the membrane [45].
As with other genes of the ABCA subfamily, a secondary role of ABCA2 in drug transport is a matter of intense research. The resistance of tumors to chemotherapy is considered to be partly due to overexpression of transmembrane transport proteins. Expression of ABCA2 was found to be significantly increased in tumor cell lines resistant to estramustine [29] and mitoxantrone [8], supporting the hypothesis that ABCA2 might store the drugs in lysosomes and possibly promote their export out of the cell. Consistently with the role of ABCA2 in the nervous system, ABCA2 expression has been reported to represent a useful marker in vestibular schwannoma, one of the most common benign intracranial tumors [50].
ABCA3
The ABCA3 gene was originally cloned in 1996 from a cDNA library constructed from a human medullary thyroid carcinoma cell line. It is a 1,704-amino acid protein (molecular weight approximately 150 kDa), mapping to human chromosome 16p13.3 and has a very similar structure as ABCA1 and ABCA2, including the HH1 hydrophobic residues (Fig. 1) [14, 27].
ABCA3 was found to be predominantly expressed in alveolar type II cells in the lung [53]. ABCA3-specific antibody staining of lung tissue revealed ring-like structures in the cytosol of the cells, suggesting that the protein is specifically concentrated in the membrane of lamellar bodies [53]. Lamellar bodies are densely packed lysosome-like structures storing pulmonary surfactant, a mixture of phospholipids and proteins, which reduces the surface tension at the air–liquid interface. The secretion of pulmonary surfactant represents a critical feature in the switch of the lung from an aqueous to an air environment at birth. After birth and throughout life, alveolar type II cells regulate the surfactant concentration in the alveolar space by secretion, reabsorption, and storage in lamellar bodies. As the lamellar structures obtained by ABCA3-specific staining are very similar to those generated by ABCA2 in the brain, ABCA3 is hypothesized to play an active role in the excretion of the lipid fraction of the pulmonary surfactant [35].
Because of this crucial function, ABCA3 expression levels have to be tightly regulated during fetal lung development. In rats, the highest ABCA3 transcript levels are detected 1–2 days before birth [34]. As glucocorticoids are known to induce fetal maturation, Yoshida et al. [54] tested the effect of intraperitoneal injection of dexamethasone (Dex) into pregnant rats. Significantly increased expression levels of ABCA3 in Dex-induced rats were demonstrated, and consistently, a glucocorticoid-response element (GRE) was identified within the 5′-flanking region of the gene. The promoter activity was induced twofold in the presence of Dex, an effect that was abolished after deletion of the GRE or introduction of a point mutation within the promoter sequence [54].
The clinical impact of defective ABCA3 function becomes apparent in the development of fatal surfactant deficiency in newborns [44]. More than two-thirds of the subjects suffering from this lethal disease carried nonsense or frameshift mutations in the ABCA3 gene. Electron micrograph images revealed abnormally small and dense lamellar bodies and peripheral inclusions in this organelle. More recently [10], it has been shown that mutations in ABCA3 are not always associated with fatal neonatal surfactant deficiency, but can be the cause of other nonlethal lung diseases.
In addition to thyroid carcinoma, ABCA3 has been detected in other tumoral cell lines, such as neuroblastoma or acute myeloid leukemia cells, showing resistance to drug treatment [18, 36, 52]. In these cells, ABCA3 is hypothesized to have a function in intralysosomal drug sequestration similar to ABCA2.
ABCA7
Among the multiple physiological pathways in which ABC transporters are implicated, lipid transport is one of the most studied due to its high impact on human health. In the year 2000, ABCA7, a new member of the A-subfamily, has been identified, and it extended the group of ABC transporters associated with lipid homeostasis. In pioneer cloning work [21], ABCA7 was characterized as a 2,146-amino acid protein sensitive to sterol concentrations. The expression levels of the protein were shown to change in different cholesterol import/export situations: in induced sterol uptake conditions, a notable increase in mRNA levels of this gene occurs, while cholesterol-depleting conditions results in down-regulation of its expression [21]. These studies propose that ABCA7 plays an important role in lipid trafficking at its distinct sites of expression.
However, several studies [17, 31] demonstrated that the role of ABCA7 in lipid transport is secondary in comparison to ABCA1, the major cellular lipid export protein. Experiments by Hayashi et al. [17] demonstrated that ABCA7 generates mostly small cholesterol-poor HDL particles, whereas ABCA1 forms predominantly big cholesterol-rich particles. Moreover, knockout of ABCA7 function in mouse macrophages did not seem to affect phospholipid or cholesterol efflux to apolipoprotein A-I, while inactivation of ABCA1 resulted in a complete absence of efflux [31].
ABCA7 has been suggested to play an important role in lipid transport within defined microenvironments. ABCA7 expression was detected in murine renal tubules in the apical brush border membrane [31]. In this context it should be noted that the kidney plays a major role in apolipoprotein A-I catabolism, and expression of other ABC transporters related with lipid homeostasis has been demonstrated in this organ [13]. In female, but not male, ABCA7 knockout mice, a significant reduction in visceral fat and serum cholesterol concentrations has been described [26]. Moreover, ABCA7 expression was detected during late keratinocyte differentiation in humans and was shown to significantly influence ceramide levels [25]. Taken together, these data indicate an important association of ABCA7 with lipid transport processes.
Expression of ABCA7 has been found in myelolymphatic tissues such as thymus, spleen, bone marrow, and fetal tissues [21]. Further analysis revealed that ABCA7 mRNA is expressed in lymphocytes, granulocytes, and monocytes, suggesting a role of ABCA7 in hematopoiesis. In agreement with this hypothesis, Broccardo et al. [9] identified several regulation sites targeted by lymphomyeloid-specific transcription factors both in the human ABCA7 and in the mouse abca7 promoter regions. However, Sasaki et al. [43] reported highly specific ABCA7 expression in rat platelets, suggesting this to be responsible for the appearance of the protein in spleen and bone marrow in those previous works. Thus, the relationship of ABCA7 with blood cell lineages remains to be elucidated. Similar to ABCA1, one extracellular domain of ABCA7 has been associated with Sjogren’s syndrome [46], but the exact relationship with this disorder has not been assessed yet.
An alternative splicing event for ABCA7 has been described, resulting in the generation of two ABCA7 transcripts with different tissue distribution and subcellular localization (plasma membrane or endoplasmic reticulum) [19]. Whether this splicing event is of physiological relevance is yet unknown.
ABCA5, A6, A8, A9, and A10: the chromosome 17q24 cluster
According to their chromosome localization and phylogenetic analysis, the ABCA subfamily can be divided into two subgroups. Eight members of the subfamily are located dispersedly in the genome, while ABCA5, ABCA6, ABCA8, ABCA9, and ABCA10 are arranged in a chromosome cluster on mouse chromosome 11 [2] and human chromosome 17 [4].
In addition to their special chromosome arrangement, all genes included on the cluster are significantly shorter than the other members of the family (38–40 instead of usually 50–52 exons). Moreover, with the exception of ABCA5, the five members of the cluster show a high amino acid identity (up to 78%) [4], which is not found when these genes are compared with other members of the ABCA subfamily (around 30% in most cases). This led to the subdivision of this subfamily into two distinct subgroups.
Interestingly, for the five members of the cluster, the C-terminal nucleotide binding domain (NBD) contains the Walker A and B motifs, but lacks the signature C motif. This highly specific feature suggests that the five genes of the cluster have evolved from each other after gene duplication events or originate from a common ancestor gene. Excluding ABCA8, the members of this cluster present protein variants [23, 40, 51], pointing to an evolutionary development to acquire a broader functional flexibility, as described for other gene clusters with transport function [15]. The relative mRNA expression of the “ABCA6-like” transporters in human tissues is presented in Table 2.
Three members of the 17q24 cluster (ABCA6, ABCA9, and ABCA10) were demonstrated to be cholesterol-responsive and characterized in humans [23, 40, 51]. First to be cloned was ABCA6. Because of their high sequence homology and function analogy, these genes are referred to as “ABCA6-like transporters”.
ABCA5
The ABCA5 protein shows an amino acid sequence identity of maximal 42% to the proteins of the 17q24 cluster. However, the homology with other ABCA subfamily members is even lower, suggesting ABCA5 to claim a unique position within this subfamily [38]. The human ABCA5 protein consists of 1,642 amino acids, with a predicted molecular weight of approximately 183 kDa [38].
Sequencing of the human ABCA5 resulted in the identification of two alternative splicing sites, caused by base insertions at the end of exons 20 and 23; accordingly, two different protein transcripts were detected by Northern blot analysis [38]. Expression of a 6.2 kb ABCA5 transcript was found to be especially abundant in skeletal muscle, with lower levels in kidney, liver and placenta, while a >9 kb transcript was present in samples purified from human colon carcinoma cell lines. However, the biological meaning of these alternative splicing events is yet unknown. For example, the insertion of 16 bp in exon 21 results in a truncated polypeptide of 925 amino acids, comprising the first transmembrane module and the first NBD. Whether this structure functions as a half-transporter in conjunction with other ABC transporters or as a regulatory element remains to be elucidated.
Recently, rat Abca5 mRNA expression was located in Leydig cells, a major site of testosterone synthesis [39]. In previous work by Kubo et al. [28], murine ABCA5 protein was detected in lysosomes and late endosomes. ABCA5 −/− mice developed characteristic symptoms of lysosomal diseases in ABCA5 expressing tissues (mainly heart and thyroid gland) and died when reaching adulthood [28]. Therefore, ABCA5 is proposed to play an important role in intracellular trafficking. However, further functional assays are necessary to identify the substrates for ABCA5 and to assess a potential implication of this transporter in human diseases.
ABCA6
Human ABCA6 consists of 1,617 amino acids with a calculated molecular weight of approximately 160 kDa and a classical ABC full-transporter structure [23]. In the same work, an alternative ABCA6 mRNA variant was identified, caused by the skipping of exon 14 that introduces a preterminal stop codon. The transcript generated is composed of 637 amino acids and has the structure of a half-transporter. Whether this truncated protein homo- or heterodimerizes to form a complete transport complex or whether it has a different function (e.g., regulation of the expression, membrane signalling, etc.) remains to be elucidated.
The human ABCA6 gene is ubiquitously expressed in the organism, with high levels found in liver, lung, heart, brain, and ovaries. It has been described to be up-regulated during macrophage differentiation and to be responsive to cholesterol concentrations [23]. Therefore, it is likely that ABCA6 plays an important role in macrophage lipid transport. Whether this function is performed as a single gene or in conjunction with other ABC transporters implicated in lipid transport, such as ABCA1, ABCA7 or other ABCA6-like transporters, is still unclear. ABCA6 may also play a role in drug transport, as significant expression was found in lymphoma cell lines with weak response to drug treatment [20].
ABCA8
The human ABCA8 protein consists of 1,581 amino acids and has a classical full-transporter structure, with 14 predicted membrane spanning segments [47]. It has been demonstrated to exert an ATPase-dependent drug transport function, with a substrate specificity very close to the multidrug resistance-associated protein 2 (MRP2; ABCC2), a member of the ABCC subfamily implicated in substance trafficking through the blood–brain barrier [16] (see review by D. Keppler and A. Nies in this issue). Expression of ABCA8 mRNA was found in various organs including heart, liver, and muscle. Further studies are necessary to assess the exact function of ABCA8 in these tissues.
ABCA9
To date, this ABC transporter has been cloned and characterized in humans in only one published work [40]. However, the data presented do support the classification of ABCA9 within the subgroup of ABCA6-like transporters: short length (1,624 amino acids), identification of splicing variants, and a potential role in lipid homeostasis. As previously demonstrated for the ABCA6 transporter [23], ABCA9 mRNA levels are up-regulated during monocyte differentiation into macrophages and respond to stimulation with cholesterol [40].
Expression of this gene occurs in a wide range of tissues, with high levels found in lung, central nervous system, reproductive tissues, and fetal tissues. Expression of the closely related ABCA6 gene in fetal tissues was not detectable or of a lower range, suggesting a specialization of ABCA9 for covering similar functions during organ development.
Piehler et al. [40] described a canonical ABCA9 protein and three splicing variants, two of them generated by intron retention and one by exon skipping. All alternative splicing events result in the introduction of preterminal stop codons. The biological roles of these alternative transcripts are still unknown. However, expression levels of the ABCA9Δ+55 transcript in human macrophages were regulated coordinately with the canonical ABCA9 transcript, supporting the hypothesis that it has a biological function related with that of the canonical protein.
ABCA10
ABCA10 represents the most recently cloned gene of the 17q24 cluster [51]. This transporter shares all structural characteristics with the other members of the cluster, and is also categorized in the subgroup of “ABCA6-like transporters”.
This 1,543-amino acid / 176 kDa protein has a full-transporter structure, with the characteristic absence of the signature sequence in the C-terminal NBD. It shares the highest nucleotide sequence homology with ABCA6, A8, and A9 [51], suggesting that these four genes originate from the same ancestor. ABCA10 and most of the currently identified ABCA transporters contain PEST sequences within their N-terminal transmembrane domains [42]. These sequences regulate the calpain-dependent degradation of the protein, and therefore control the membrane availability of the transporter, as shown before for ABCA1 [49].
Similar to the other “ABCA6-like” genes, the existence of splicing variants and the responsiveness to sterol concentrations have also been described for ABCA10 [51]. This gene is expressed predominantly in the intestinal tract, where other ABC transporters (ABCG5 and ABCG8, see review by S. E. Hazard and S. B. Patel in this issue) play an important role in lipid transport processes [7]. The exact function of ABCA10 is supposed to be related with those processes, but this remains to be verified.
In summary, all “ABCA6-like” transporters present highly homologous sequences and are regulated by cholesterol levels. However, their expression pattern and tissue distribution reveal significant differences. The marked similarities and localization on a gene cluster support the hypothesis that they originate from a duplication process during evolution. Whether this occurred as a specialization process for developing similar functions at different locations or whether these genes exert synergistic activities are challenging hypotheses to be tested in future studies. In addition, it will be of high interest to investigate the potential biological significance of the splicing variants present in all “ABCA6-like” transporters.
ABCA12
The human ABCA12 gene is located on chromosome 2q34 and encodes for two splicing variants of 2,595 and 2,516 amino acids; both of them are predicted to have two NBDs and two transmembrane domains [3].
Expression of ABCA12 was predominantly found on the skin and in the stomach. The ABCA12 gene is located on a chromosomal region previously linked to skin diseases, diabetes mellitus, and congenital cataracts [3]. In agreement with the assumption that ABCA12 plays a role in skin lipid homeostasis, high ABCA12 expression was found in healthy keratinocytes. Mutations in the ABCA12 gene were detected in patients suffering from lamellar ichthyosis type 2, a rare hereditary disease characterized by skin desquamation over the whole body [30, 37]. Harlequin ichthyosis (HI), the most severe form of this disease, was described to be causally linked to mutations in ABCA12 [24]. This often-lethal disease is caused by a disorder in the keratinization process resulting in the coverage of the whole body with a thick “armor”-like scale layer. Prior to identification of ABCA12 mutations in this disease, prenatal diagnosis for HI was only possible in fetal biopsies. Therefore, the recent work by Akiyama et al. [1], suggesting the feasibility of HI diagnosis during pregnancy, could represent a major diagnostic and analytical advance in this severe disease. In this study, five distinct mutations in the ABCA12 gene were identified in HI patients. The ABCA12 protein was localized in the lamellar granules of keratinocytes [1], where it may play a major role in the regulation of lipid trafficking. Moreover, the defect in cultured keratinocytes could be corrected after gene transfer [1], implying the possibility of a future gene therapy for HI.
ABCA13
Human ABCA13 is the largest ABC transporter protein described to date, with a length of 5,058 amino acids and a predicted molecular weight of >450 kDa [41]. A characteristic feature of ABCA13 is the extraordinarily long N-terminal cytoplasmic tail, which is encoded by 30 exons and spans more than 3,500 amino acids. The ABCA13 gene maps to chromosome 7p12.3; this region is genetically linked to Schwachman–Diamond syndrome, a genetic disorder affecting the pancreas, and a locus involved in T-cell tumor invasion and cancer metastasis. Consistently, elevated expression of the ABCA13 gene was found in leukemia, prostate tumor, and CNS tumor cell lines, where it could play a role in transport of xenobiotics and subsequent drug resistance.
However, the expression levels of ABCA13 mRNA in healthy tissues are very low, and the transcript was only detectable in a small number of tissues, including the trachea, testis, and bone marrow [41]. The murine ABCA13 promoter region was found to contain transcription factor binding sites associated with myeloid and lymphoid cell types [6]. Ubiquitous mRNA expression in blood derived cells was demonstrated [6], suggesting a role associated with hematopoiesis. Nevertheless, future functional studies are needed to identify the exact function and substrates of this novel transporter.
Other members of the ABCA subfamily
Recently, four other members were added to the A-subfamily of ABC transporters, but they have been only cloned from mouse and rat [5, 12]; to date no orthologues are described in the human genome. ABCA14, ABCA15, and ABCA16 are arranged in a cluster on mouse chromosome 7, while ABCA17 is located separately. ABCA14–17 are full-transporter proteins exclusively expressed in the testis [12]. It has been proposed that these proteins have functions related with lipid transport during testis development and/or spermatogenesis [12]. Indeed, ABCA17 was localized by in situ hybridization to germ cells in the seminiferous tubule [5]. In HEK293 cell lines transfected with ABCA17, its subcellular expression was restricted to the endoplasmic reticulum. The function of ABCA17 has been associated with lipid export to the sperm [12].
In summary, the members of the ABCA subfamily of transporters play an important role in lipid transport processes. Most of them were found to be expressed in lysosome-like structures and to be related with lipid trafficking. Consistently, mutations in ABC transporters of the A-subfamily have been identified to be the cause of severe disorders associated with lipid transport. Members of the human chromosome 17q24 cluster seem to have evolved from similar evolutionary processes and have developed similar functions in different locations within the body. Potential synergistic relationships between the members of the ABCA subfamily or with other ABC transporters remain to be elucidated.
References
Akiyama M, Sugiyama-Nakagiri Y, Sakai K, McMillan JR, Goto M, Arita K, Tsuji-Abe Y, Tabata N, Matsuoka K, Sasaki R, Sawamura D, Shimizu H (2005) Mutations in lipid transporter ABCA12 in harlequin ichthyosis and functional recovery by corrective gene transfer. J Clin Invest 115:1777–1784
Annilo T, Chen ZQ, Shulenin S, Dean M (2003) Evolutionary analysis of a cluster of ATP-binding cassette (ABC) genes. Mamm Genome 14:7–20
Annilo T, Shulenin S, Chen ZQ, Arnould I, Prades C, Lemoine C, Maintoux-Larois C, Devaud C, Dean M, Denefle P, Rosier M (2002) Identification and characterization of a novel ABCA subfamily member, ABCA12, located in the lamellar ichthyosis region on 2q34. Cytogenet Genome Res 98:169–176
Arnould I, Schriml LM, Prades C, Lachtermacher-Triunfol M, Schneider T, Maintoux-Larois C, Lemoine C, Debono D, Devaud C, Naudin L, Bauche S, Annat M, Annilo T, Allikmets R, Gold B, Denefle P, Rosier M, Dean M (2005) Identifying and characterizing a five-gene cluster of ATP-binding cassette transporters mapping to human chromosome 17q24: a new subgroup within the ABCA subfamily. GeneScreen 1:157–164
Ban N, Sasaki M, Sakai H, Ueda K, Inagaki N (2005) Cloning of ABCA17, a novel rodent sperm-specific ABC (ATP-binding cassette) transporter that regulates intracellular lipid metabolism. Biochem J 389:577–585
Barros SA, Tennant RW, Cannon RE (2003) Molecular structure and characterization of a novel murine ABC transporter, Abca13. Gene 307:191–200
Berge KE, Tian H, Graf GA, Yu L, Grishin NV, Schultz J, Kwiterovich P, Shan B, Barnes R, Hobbs HH (2000) Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 290:1771–1775
Boonstra R, Timmer-Bosscha H, Echten-Arends J, van der Kolk DM, van den BA, de Jong B, Tew KD, Poppema S, de Vries EG (2004) Mitoxantrone resistance in a small cell lung cancer cell line is associated with ABCA2 upregulation. Br J Cancer 90:2411–2417
Broccardo C, Osorio J, Luciani MF, Schriml LM, Prades C, Shulenin S, Arnould I, Naudin L, Lafargue C, Rosier M, Jordan B, Mattei MG, Dean M, Denefle P, Chimini G (2001) Comparative analysis of the promoter structure and genomic organization of the human and mouse ABCA7 gene encoding a novel ABCA transporter. Cytogenet Cell Genet 92:264–270
Bullard JE, Wert SE, Whitsett JA, Dean M, Nogee LM (2005) ABCA3 mutations associated with pediatric interstitial lung disease. Am J Respir Crit Care Med 172(8):1026–1031
Chen ZJ, Vulevic B, Ile KE, Soulika A, Davis W Jr, Reiner PB, Connop BP, Nathwani P, Trojanowski JQ, Tew KD (2004) Association of ABCA2 expression with determinants of Alzheimer’s disease. FASEB J 18:1129–1131
Chen ZQ, Annilo T, Shulenin S, Dean M (2004) Three ATP-binding cassette transporter genes, Abca14, Abca15, and Abca16, form a cluster on mouse chromosome 7F3. Mamm Genome 15:335–343
Christiansen-Weber TA, Voland JR, Wu Y, Ngo K, Roland BL, Nguyen S, Peterson PA, Fung-Leung WP (2000) Functional loss of ABCA1 in mice causes severe placental malformation, aberrant lipid distribution, and kidney glomerulonephritis as well as high-density lipoprotein cholesterol deficiency. Am J Pathol 157:1017–1029
Connors TD, Van Raay TJ, Petry LR, Klinger KW, Landes GM, Burn TC (1997) The cloning of a human ABC gene (ABC3) mapping to chromosome 16p13.3. Genomics 39:231–234
Gellner K, Eiselt R, Hustert E, Arnold H, Koch I, Haberl M, Deglmann CJ, Burk O, Buntefuss D, Escher S, Bishop C, Koebe HG, Brinkmann U, Klenk HP, Kleine K, Meyer UA, Wojnowski L (2001) Genomic organization of the human CYP3A locus: identification of a new, inducible CYP3A gene. Pharmacogenetics 11:111–121
Graff CL, Pollack GM (2004) Drug transport at the blood–brain barrier and the choroid plexus. Curr Drug Metab 5:95–108
Hayashi M, Abe-Dohmae S, Okazaki M, Ueda K, Yokoyama S (2005) Heterogeneity of high density lipoprotein generated by ABCA1 and ABCA7. J Lipid Res 46:1703–1711
Hirschmann-Jax C, Foster AE, Wulf GG, Nuchtern JG, Jax TW, Gobel U, Goodell MA, Brenner MK (2004) A distinct “side population” of cells with high drug efflux capacity in human tumor cells. Proc Natl Acad Sci U S A 101:14228–14233
Ikeda Y, Abe-Dohmae S, Munehira Y, Aoki R, Kawamoto S, Furuya A, Shitara K, Amachi T, Kioka N, Matsuo M, Yokoyama S, Ueda K (2003) Posttranscriptional regulation of human ABCA7 and its function for the apoA-I-dependent lipid release. Biochem Biophys Res Commun 311:313–318
Islam TC, Asplund AC, Lindvall JM, Nygren L, Liden J, Kimby E, Christensson B, Smith CI, Sander B (2003) High level of cannabinoid receptor 1, absence of regulator of G protein signalling 13 and differential expression of Cyclin D1 in mantle cell lymphoma. Leukemia 17:1880–1890
Kaminski WE, Orso E, Diederich W, Klucken J, Drobnik W, Schmitz G (2000) Identification of a novel human sterol-sensitive ATP-binding cassette transporter (ABCA7). Biochem Biophys Res Commun 273:532–538
Kaminski WE, Piehler A, Pullmann K, Porsch-Ozcurumez M, Duong C, Bared GM, Buchler C, Schmitz G (2001) Complete coding sequence, promoter region, and genomic structure of the human ABCA2 gene and evidence for sterol-dependent regulation in macrophages. Biochem Biophys Res Commun 281:249–258
Kaminski WE, Wenzel JJ, Piehler A, Langmann T, Schmitz G (2001) ABCA6, a novel a subclass ABC transporter. Biochem Biophys Res Commun 285:1295–1301
Kelsell DP, Norgett EE, Unsworth H, Teh MT, Cullup T, Mein CA, Dopping-Hepenstal PJ, Dale BA, Tadini G, Fleckman P, Stephens KG, Sybert VP, Mallory SB, North BV, Witt DR, Sprecher E, Taylor AE, Ilchyshyn A, Kennedy CT, Goodyear H, Moss C, Paige D, Harper JI, Young BD, Leigh IM, Eady RA, O’Toole EA (2005) Mutations in ABCA12 underlie the severe congenital skin disease harlequin ichthyosis. Am J Hum Genet 76:794–803
Kielar D, Kaminski WE, Liebisch G, Piehler A, Wenzel JJ, Mohle C, Heimerl S, Langmann T, Friedrich SO, Bottcher A, Barlage S, Drobnik W, Schmitz G (2003) Adenosine triphosphate binding cassette (ABC) transporters are expressed and regulated during terminal keratinocyte differentiation: a potential role for ABCA7 in epidermal lipid reorganization. J Invest Dermatol 121:465–474
Kim WS, Fitzgerald ML, Kang K, Okuhira K, Bell SA, Manning JJ, Koehn SL, Lu N, Moore KJ, Freeman MW (2005) Abca7 null mice retain normal macrophage phosphatidylcholine and cholesterol efflux activity despite alterations in adipose mass and serum cholesterol levels. J Biol Chem 280:3989–3995
Klugbauer N, Hofmann F (1996) Primary structure of a novel ABC transporter with a chromosomal localization on the band encoding the multidrug resistance-associated protein. FEBS Lett 391:61–65
Kubo Y, Sekiya S, Ohigashi M, Takenaka C, Tamura K, Nada S, Nishi T, Yamamoto A, Yamaguchi A (2005) ABCA5 resides in lysosomes, and ABCA5 knockout mice develop lysosomal disease-like symptoms. Mol Cell Biol 25:4138–4149
Laing NM, Belinsky MG, Kruh GD, Bell DW, Boyd JT, Barone L, Testa JR, Tew KD (1998) Amplification of the ATP-binding cassette 2 transporter gene is functionally linked with enhanced efflux of estramustine in ovarian carcinoma cells. Cancer Res 58:1332–1337
Lefevre C, Audebert S, Jobard F, Bouadjar B, Lakhdar H, Boughdene-Stambouli O, Blanchet-Bardon C, Heilig R, Foglio M, Weissenbach J, Lathrop M, Prud’homme JF, Fischer J (2003) Mutations in the transporter ABCA12 are associated with lamellar ichthyosis type 2. Hum Mol Genet 12:2369–2378
Linsel-Nitschke P, Jehle AW, Shan J, Cao G, Bacic D, Lan D, Wang N, Tall AR (2005) Potential role of ABCA7 in cellular lipid efflux to apoA-I. J Lipid Res 46:86–92
Luciani MF, Denizot F, Savary S, Mattei MG, Chimini G (1994) Cloning of two novel ABC transporters mapping on human chromosome 9. Genomics 21:150–159
Mace S, Cousin E, Ricard S, Genin E, Spanakis E, Lafargue-Soubigou C, Genin B, Fournel R, Roche S, Haussy G, Massey F, Soubigou S, Brefort G, Benoit P, Brice A, Campion D, Hollis M, Pradier L, Benavides J, Deleuze JF (2005) ABCA2 is a strong genetic risk factor for early-onset Alzheimer’s disease. Neurobiol Dis 18:119–125
Mulugeta S, Gray JM, Notarfrancesco KL, Gonzales LW, Koval M, Feinstein SI, Ballard PL, Fisher AB, Shuman H (2002) Identification of LBM180, a lamellar body limiting membrane protein of alveolar type II cells, as the ABC transporter protein ABCA3. J Biol Chem 277:22147–22155
Nagata K, Yamamoto A, Ban N, Tanaka AR, Matsuo M, Kioka N, Inagaki N, Ueda K (2004) Human ABCA3, a product of a responsible gene for abca3 for fatal surfactant deficiency in newborns, exhibits unique ATP hydrolysis activity and generates intracellular multilamellar vesicles. Biochem Biophys Res Commun 324:262–268
Norwood K, Wang RY, Hirschmann-Jax C, Andreeff M, Brenner MK, Goodell MA, Wulf GG (2004) An in vivo propagated human acute myeloid leukemia expressing ABCA3. Leuk Res 28:295–299
Parmentier L, Lakhdar H, Blanchet-Bardon C, Marchand S, Dubertret L, Weissenbach J (1996) Mapping of a second locus for lamellar ichthyosis to chromosome 2q33-35. Hum Mol Genet 5:555–559
Petry F, Kotthaus A, Hirsch-Ernst KI (2003) Cloning of human and rat ABCA5/Abca5 and detection of a human splice variant. Biochem Biophys Res Commun 300:343–350
Petry F, Ritz V, Meineke C, Middel P, Kietzmann T, Schmitz-Salue C, Hirsch-Ernst KI (2005) Subcellular localisation of rAbca5, a rat ATP-binding cassette transporter expressed in Leydig cells, and characterisation of its splice variant apparently encoding a half-transporter. Biochem J 393(Pt 1):79–87
Piehler A, Kaminski WE, Wenzel JJ, Langmann T, Schmitz G (2002) Molecular structure of a novel cholesterol-responsive A subclass ABC transporter, ABCA9. Biochem Biophys Res Commun 295:408–416
Prades C, Arnould I, Annilo T, Shulenin S, Chen ZQ, Orosco L, Triunfol M, Devaud C, Maintoux-Larois C, Lafargue C, Lemoine C, Denefle P, Rosier M, Dean M (2002) The human ATP binding cassette gene ABCA13, located on chromosome 7p12.3, encodes a 5058 amino acid protein with an extracellular domain encoded in part by a 4.8-kb conserved exon. Cytogenet Genome Res 98:160–168
Rechsteiner M, Rogers SW (1996) PEST sequences and regulation by proteolysis. Trends Biochem Sci 21:267–271
Sasaki M, Shoji A, Kubo Y, Nada S, Yamaguchi A (2003) Cloning of rat ABCA7 and its preferential expression in platelets. Biochem Biophys Res Commun 304:777–782
Shulenin S, Nogee LM, Annilo T, Wert SE, Whitsett JA, Dean M (2004) ABCA3 gene mutations in newborns with fatal surfactant deficiency. N Engl J Med 350:1296–1303
Tanaka Y, Yamada K, Zhou CJ, Ban N, Shioda S, Inagaki N (2003) Temporal and spatial profiles of ABCA2-expressing oligodendrocytes in the developing rat brain. J Comp Neurol 455:353–367
Toda Y, Aoki R, Ikeda Y, Azuma Y, Kioka N, Matsuo M, Sakamoto M, Mori S, Fukumoto M, Ueda K (2005) Detection of ABCA7-positive cells in salivary glands from patients with Sjogren’s syndrome. Pathol Int 55:639–643
Tsuruoka S, Ishibashi K, Yamamoto H, Wakaumi M, Suzuki M, Schwartz GJ, Imai M, Fujimura A (2002) Functional analysis of ABCA8, a new drug transporter. Biochem Biophys Res Commun 298:41–45
Vulevic B, Chen Z, Boyd JT, Davis W Jr, Walsh ES, Belinsky MG, Tew KD (2001) Cloning and characterization of human adenosine 5′-triphosphate-binding cassette, sub-family A, transporter 2 (ABCA2). Cancer Res 61:3339–3347
Wang N, Chen W, Linsel-Nitschke P, Martinez LO, Agerholm-Larsen B, Silver DL, Tall AR (2003) A PEST sequence in ABCA1 regulates degradation by calpain protease and stabilization of ABCA1 by apoA-I. J Clin Invest 111:99–107
Wang Y, Yamada K, Tanaka Y, Ishikawa K, Inagaki N (2005) Expression of ABCA2 protein in human vestibular schwannoma and peripheral nerve. J Neurol Sci 232:59–63
Wenzel JJ, Kaminski WE, Piehler A, Heimerl S, Langmann T, Schmitz G (2003) ABCA10, a novel cholesterol-regulated ABCA6-like ABC transporter. Biochem Biophys Res Commun 306:1089–1098
Wulf GG, Modlich S, Inagaki N, Reinhardt D, Schroers R, Griesinger F, Trumper L (2004) ABC transporter ABCA3 is expressed in acute myeloid leukemia blast cells and participates in vesicular transport. Haematologica 89:1395–1397
Yamano G, Funahashi H, Kawanami O, Zhao LX, Ban N, Uchida Y, Morohoshi T, Ogawa J, Shioda S, Inagaki N (2001) ABCA3 is a lamellar body membrane protein in human lung alveolar type II cells. FEBS Lett 508:221–225
Yoshida I, Ban N, Inagaki N (2004) Expression of ABCA3, a causative gene for fatal surfactant deficiency, is up-regulated by glucocorticoids in lung alveolar type II cells. Biochem Biophys Res Commun 323:547–555
Zhao LX, Zhou CJ, Tanaka A, Nakata M, Hirabayashi T, Amachi T, Shioda S, Ueda K, Inagaki N (2000) Cloning, characterization and tissue distribution of the rat ATP-binding cassette (ABC) transporter ABC2/ABCA2. Biochem J 350(Pt 3):865–872
Zhou C, Zhao L, Inagaki N, Guan J, Nakajo S, Hirabayashi T, Kikuyama S, Shioda S (2001) Atp-binding cassette transporter ABC2/ABCA2 in the rat brain: a novel mammalian lysosome-associated membrane protein and a specific marker for oligodendrocytes but not for myelin sheaths. J Neurosci 21:849–857
Zhou CJ, Inagaki N, Pleasure SJ, Zhao LX, Kikuyama S, Shioda S (2002) ATP-binding cassette transporter ABCA2 (ABC2) expression in the developing spinal cord and PNS during myelination. J Comp Neurol 451:334–345
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Albrecht, C., Viturro, E. The ABCA subfamily—gene and protein structures, functions and associated hereditary diseases. Pflugers Arch - Eur J Physiol 453, 581–589 (2007). https://doi.org/10.1007/s00424-006-0047-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00424-006-0047-8