
Abstract
Fusaric acid (FA) is one of the most harmful phytotoxins produced in various plant–pathogen interactions. Fusarium species produce FA as a secondary metabolite, which can infect many agronomic crops at all stages of development from seed to fruit, and FA production can further compromise plant survival because of its phytotoxic effects. FA exposure in plant species adversely affects plant growth, development and crop yield. FA exposure in plants leads to the generation of reactive oxygen species (ROS), which cause cellular damage and ultimately cell death. Therefore, FA-induced ROS accumulation in plants has been a topic of interest for many researchers to understand the plant–pathogen interactions and plant defence responses. In this study, we reviewed the FA-mediated oxidative stress and ROS-induced defence responses of antioxidants, as well as hormonal signalling in plants. The effects of FA phytotoxicity on lipid peroxidation, physiological changes and ultrastructural changes at cellular and subcellular levels were reported. Additionally, DNA damage, cell death and adverse effects on photosynthesis have been explained. Some possible approaches to overcome the harmful effects of FA in plants were also discussed. It is concluded that FA-induced ROS affect the enzymatic and non-enzymatic antioxidant system regulated by phytohormones. The effects of FA are also associated with other photosynthetic, ultrastructural and genotoxic modifications in plants.
Graphical abstract

Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fusarium toxins and their exposure
Phytotoxins are produced by several fungal species as secondary metabolites that can have severe toxic effects on plants and animals, including humans (Zain 2011). Every year, significant crop yields are lost because of the detrimental effects of phytotoxins, e.g. Fusarium toxins, on agricultural crops worldwide (Ismaiel and Papenbrock 2015; Pleadin et al. 2019; Rodríguez-Carrasco et al. 2013). These Fusarium toxins are low molecular weight compounds that are produced under many environmental conditions, mainly in cereals at all stages of development (Rodríguez-Carrasco et al. 2013). Therefore, these Fusarium toxins can contaminate many food sources and cause serious health problems when consumed (Alshannaq and Yu 2017; Iqbal et al. 2021a). The accumulation of these phytotoxins in the food chain is a major problem that is receiving considerable attention at the global level (Zain 2011; Rodríguez-Carrasco et al. 2013). Phytotoxin contamination in food and feed occurs as a result of malnutrition, poor handling techniques, inappropriate food storage and consumption of staple foods obtained from toxin-contaminated sites (Milićević et al. 2010). Fusarium toxins have been reported as carcinogens, hepatotoxins, nephrotoxins and neurotoxins (Arumugam et al. 2021). At the same time, the nature and damage caused by these phytotoxins vary with the type of phytotoxin, concentration, exposure time, route of consumption, health status and sex of the affected person (Zain 2011). Humans are mainly exposed to these toxins through the consumption of contaminated food and transfer of phytotoxins in animal products. In addition to oral ingestion of fungal contaminated food (Pinotti et al. 2016), skin contact (dermatophyte skin infection) and inhalation of spores (toxic pneumonitis) are also routes of exposure to Fusarium toxins (Kuruvilla and Dias 2012). Fusaric acid (FA), fumonisins, trichothecenes, zearalenone, deoxynivalenol, moniliformin, fusaproliferin, enniatins and beauvericin are classified as the most common and harmful phytotoxins (Arumugam et al. 2021; Ismaiel and Papenbrock 2015).
Basic overview of fusaric acid
FA is one of the most harmful and non-specific phytotoxins produced by various Fusarium species. The molecular formula and weight of FA are C10H13NO2 and 179.2157 g/mol, respectively (Singh et al. 2017). FA has been recognized as a phytotoxin involved in the development of plant diseases. During the separation of gibberellins in 1934, a Japanese scientist found a crystalline compound from the fungus Gibberella fujikuroi, which was later named FA (Brown et al. 2012; Sieber et al. 2013). FA is a derivative of picolinic acid (5-butylpicolinic acid; Fig. 1A) which is known as a metal chelating agent (Ismaiel and Papenbrock 2015). FA production is not restricted to Gibberella fujikuroi, but the entire Fusarium genus has the ability to produce it (Brown et al. 2012; Sieber et al. 2013). Interestingly, the 5-butyl side chain of FA is involved in enhancing the lipophilicity of FA which assists in the penetration through the cell membrane (Devnarain et al. 2017). In addition, FA has a carboxylic acid group, which acts as a proton donor and is responsible for its acidic properties in various reactions (Liu et al. 2016). As a result of the chelate-forming ability of FA, it binds with other divalent cations, causing the conjugation of iron, manganese, copper and zinc, which hinders their ability to perform biological functions (e.g. as cofactors of the antioxidant enzyme superoxide dismutase) (Arumugam et al. 2021).

FA-producing fungal species
Besides the FA-producing Fusarium fujikuroi, previously known as Gibberella fujikuroi, there are 50 to 100 phylogenetically closely related Fusarium species with the ability to produce FA such as F. verticillioides and F. proliferatum (Cen et al. 2020). Among the other closely related Fusarium species that are able to produce FA we can find F. oxysporum, F. mangiferae, F. subglutinans, F. circinatum and the remotely related F. solani, F. napiforme and F. crookwellense (Ismaiel and Papenbrock 2015). The high production of FA depends on the virulence of the fungal strain and the susceptibility of the host plants, such as banana, cucumber, potato, tobacco, tomato and wheat (Alshannaq and Yu 2017; Singh et al. 2017). Briefly, a virulent fungal strain can produce a higher amount of FA, resulting in more severe plant infection, whereas a susceptible plant with a weak immune system cannot resist fungal attack, resulting in more severe fungal infection (Singh et al. 2017).
FA biosynthesis
FA is a polyketide-derived secondary metabolite (Srivastava et al. 2020). The genes responsible for its biosynthesis are located next to each other in the genome. The genomic cluster for FA production involves genes encoding specific enzymes, transport proteins and transcription factors (Brown et al. 2015). Previous studies have reported that the FA biosynthesis gene cluster (FUB) comprises five genes in F. verticillioides (maize pathogen) and F. fujikuroi (rice pathogen) (Brown et al. 2012; Niehaus et al. 2014). However, transcription factors and transporter genes were not included in the cluster (Brown et al. 2015). Subsequently, the FUB region was analysed in several Fusarium species such as F. verticillioides, F. fujikuroi and F. oxysporum. A total of 12 genes (FUB1 to FUB12) are present in the FUB cluster (Brown et al. 2015). Furthermore, deletion analysis revealed that nine FUB genes and two transcription factors (Zn(II)2Cys6) are responsible for FA production (Brown et al. 2015). In one research study, the homologs of the FUB gene cluster were compared with the gene clusters of other Fusarium species, and it was found that certain homologs had non-FUB genes inserted at specific positions (Brown et al. 2012, 2015). The F. verticillioides and F. oxysporum infection of maize and cactus plants showed that FA production contributes to the phytotoxicity of Fusarium species but did not affect the virulence of the fungus (Brown et al. 2015). A proposed pathway for FA production in Fusarium sp. is shown in Fig. 1B. In this pathway, polyketide synthase (FUB1) converts three molecules of acetate to triketide, which further reacts with oxaloacetate to form FA. Nitrogen supply required for FA synthesis is derived from glutamine (Brown et al. 2015). The predicted functions of the FUB clusters were revealed by Brown and co-workers (2015) in F. verticillioides and F. oxysporum. As a result of the lack of distinct FA intermediates in fungal fub mutant cultures, the exact functions of the FUB3, FUB4, FUB5, FUB6 and FUB8 genes could not be assigned (Brown et al. 2015; Niehaus et al. 2014). Therefore, the exact role of the polyketide precursor in FA synthesis is still unknown. Similarly, FUB2 is assigned to encode a small protein, but its function is also currently unknown (Brown et al. 2015). However, FUB7 and FUB9 encode a sulfhydrylase and an oxidase, respectively, but their specific role in FA biosynthesis is unclear (Brown et al. 2015). Both FUB10 and FUB12 genes encode C6 transcription factors, while FUB11 acts as a transporter-encoding gene (Brown et al. 2015). FUB6, FUB8 and FUB10, which are responsible for FA production, may provide the basis for future research regarding the regulation of FA synthesis in Fusarium species. FB10 is a positive regulator of the other gene clusters while FB11 acts as a major facilitator superfamily (MFS) which plays a role in detoxification processes by removing excess FA (Studt et al. 2016). Moreover, FUB12 is also involved in the detoxification of FA to dehydrofusaric acid and then to fusarinolic acid by regulating the expression of cytochrome P450 (Studt et al. 2016). On the basis of the available literature, controlling the expression of FB6, FB8 and FB10 genes may lead to the development of novel strategies to limit FA production (Brown et al. 2015).
Toxicity of FA and its mechanism
Toxicity of FA in animals, humans and other organisms
Consumption of contaminated plant-derived foods can result in the accumulation of mycotoxins in body cells of both humans and animals, where FA concentrations can reach dangerous levels up to 10−5 M (da Rocha et al. 2014; Sobral et al. 2018). Higher levels of FA are carcinogenic to the human immune system, reproductive organs, liver, kidney and brain (da Rocha et al. 2014; Mamur et al. 2020). For instance, FA toxicity in human hepatocellular carcinoma (HepG2) cell line induced DNA damage and post-translational modifications of p53 (a tumour suppressor protein) by increasing the activity histone deacetylases and inhibiting histone acetyltransferases followed by increased cell proliferation and apoptosis in the cells (Ghazi et al. 2017). In addition, FA induces skin and gastrointestinal disorders and alters gene function leading to abnormal embryo development (Mézes 2008). FA toxicity can inhibit the activity of dopamine β-hydroxylase, which is responsible for the proper functioning of the nervous system (Reddy et al. 1996). FA has been shown to be cytotoxic to human normal fibroblast as well as colon and breast cancer cells at concentrations of 500 µM (Fernandez-Pol et al. 1993). In another study, Mamur and co-workers (2020) analysed the toxic effects of FA on human lymphocytes and human cervical carcinoma cells, and showed that FA at a concentration of 400 µg/mL had both cytotoxic and genotoxic effects. FA also induced toxic effects on rat hepatoma cells, Chinese hamster ovary, mouse fibroblast and dog kidney fibroblast cells (Mamur et al. 2020; Vesonder et al. 1993). FA also induced toxic effects on zebrafish embryo development by chelating the active site of lysyl oxidase (an enzyme essential for respiratory function, ovulation and wound healing) with copper to inhibit its activity, resulting in teratogenic effects (Yin et al. 2015).
In addition to animals and humans, FA also affected bacterial growth (10−4 to 10−3 M), cell wall permeability in green algae (Spirogyra nitida) (5 × 10−3 M) and spore germination of the fungus Ustilago maydis at 1.5 × 10−4 M concentration (Srivastava et al. 2020). Therefore, more preventive measures are needed to reduce the toxic effects of FA in feed and food.
FA toxicity in plants
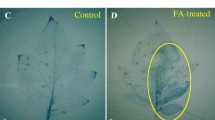
FA also causes plant diseases such as fusarium wilt, which reduces the yield of many agricultural crops (Singh et al. 2017). The toxicity of FA has been documented in many plant species such as tomato (Singh et al. 2017), potato (Sapko et al. 2011), cucumber (Wang et al. 2015), watermelon (Wu et al. 2007), banana (Fung et al. 2019), date palm (Bouizgarne et al. 2004), Arabidopsis (Bouizgarne et al. 2006a), maize (Spss and Oliveira 2009), cotton (Stipanovic et al. 2011), tobacco (Jiao et al. 2014), wheat (Li et al. 2021), barley (Liu et al. 2016), saffron (Samadi and Shahsavan Behboodi 2006), wax gourd (Wang et al. 2021), chihuahua flower (Antić et al. 2020), faba bean (Li et al. 2021), castor bean (Pavlovkin et al. 2004), cape gooseberry (Mendoza-Vargas et al. 2021), sword lily (Nosir et al. 2011) and hemp broomrape (Bouizgarne et al. 2006b). In plants, FA induces oxidative stress, chloroplast and mitochondrial dysfunction, lipid peroxidation, protein damage and DNA fragmentation (Jiao et al. 2014; Iqbal et al. 2021a) (Fig. 2). FA causes wilting, necrotic spots on leaves, reduction in roots and root hair growth (Singh et al. 2017), cell signalling perturbation, disruption of photosynthetic activity (Iqbal et al. 2021a), inhibition of respiration, membrane hyperpolarization and reduction in intracellular ATP levels (Bouizgarne et al. 2006a). FA has also been reported to inhibit cytochrome oxidase activity, alter membrane permeability, induce plasma membrane damage, chromatin condensation and electrolyte leakage (Kuźniak 2001; Singh and Upadhyay 2014).

General toxicity symptoms of fusaric acid (FA) in plants
In addition, FA can exert its phytotoxic effect on plant species in the dose range of 20–200 mg/kg (Manda and Srivastava 2021). Previous research results suggest that the FA concentration is also improved by the conversion of dehydrofusaric acid to FA (Manda and Srivastava 2021). Exposure of plants to FA results in chlorosis, chelation of copper and iron, changes in the cell wall permeability, disturbance of ionic balance as well as inhibition of enzymatic processes such as via the defectiveness of respiratory enzymes (Srivastava et al. 2020). In addition to FA, other phytotoxins have also caused similar wilt symptoms in plants, which require further investigation.
FA uptake and mechanism of action in plants
Many studies have reported the phytotoxicity and disease symptoms caused by FA in plants; however, some recent studies also investigated the mechanism of action of FA in plants (López–Díaz et al. 2018; Iqbal et al. 2023). FA induces wilt symptoms in tomato seedlings, when applied via the rooting medium (root uptake). The wilting characteristics in tomato cotyledons and lower leaves indicate the ability of FA to spread throughout the plant (López–Díaz et al. 2018). In the cucumber and banana plants, transport of FA from the roots to the upper parts (shoot, stem, leaf) was observed via both xylem and phloem pathways (Wang et al. 2013; Dong et al. 2014). The transport of FA within the plants causes membrane damage to the transport vessels which eventually leads to wilting (Dong et al. 2014; Wang et al. 2013). These findings are also consistent with another study where wilting symptoms were observed in watermelon seedlings (Wu et al. 2008a). However, Ruiz and co-workers (2015) suggested that the movement of FA within the plants was due to its chelating ability for metal ions which induces siderophore synthesis. Furthermore, the application of metal ions such as copper, zinc or iron improved the chelating ability of FA but at the same time inhibited the phytotoxicity of FA in tomato plants (López–Díaz et al. 2018). The inhibition of FA toxicity was also functional when FA and metal ions were applied to different plant parts such as leaves and roots, demonstrating that the chelating mechanism occurs in plants (López–Díaz et al. 2018). Furthermore, the presence of membrane-permeable TPEN (metal chelator) showed toxicity similar to that of FA (Fernandez-Pol et al. 1993). Although other possible mechanisms cannot be excluded, these results accurately elucidate the relationship between metal chelation and FA toxicity. Fusarium infection-induced disease symptoms in banana plants were further exacerbated by FA production, leading to acceleration of senescence (Dong et al. 2014). During this process, FA directly damaged the cell membranes and chloroplasts and then accumulated in the lower leaves to enhance senescence, demonstrating that FA can have both direct and indirect toxic effects in plants (Dong et al. 2014).
FA-induced oxidative and nitrosative stress in plants
FA can induce ROS production at or above 10−5 M concentration in different plant species (Singh and Upadhyay 2014). ROS include hydrogen peroxide (H2O2), superoxide radical (O2.−), hydroxyl ion (−OH) and singlet oxygen (1O2) (Singh and Upadhyay 2014). FA-mediated ROS accumulation is a major contributor to the development of disease symptoms in plants (Singh et al. 2017). It is well known that pathogenic attacks increase ROS production, which ultimately triggers the plant hypersensitive response (HR). HR is part of plant cell death that limits the spread of fungal infection by affecting many organelles such as mitochondria, chloroplast and nucleus (Singh et al. 2017; Singh and Upadhyay 2016; Karuppanapandian et al. 2011). Among ROS, H2O2 is a long-lasting molecule that plays an important role in plant immunity against pathogens, and acts as a signalling molecule under phytotoxin exposure (Zaynab et al. 2019). The control and regulation of ROS production as well as degradation was observed to be carried out by various antioxidant mechanisms (Ali et al. 2018). In addition, beneficial effects of ROS on cell functions such as growth and differentiation have also been reported at lower concentrations, but higher concentrations may be detrimental to cells by inducing cell death (Ma 2013). Under FA exposure, ROS accumulation affects many biochemical processes (Breusegem and Dat 2006). The balance between ROS production and ROS scavenging can be a determining factor for the infection of plant pathogens. Exposure to phytotoxins can perturb this redox balance and lead to ROS accumulation resulting in toxicity and disease (Arumugam et al. 2021; Mates 2000). At the same time, FA-induced ROS generation in different plant species has not been well studied and further research is needed to understand the exact mechanism of ROS production in the plant cell.
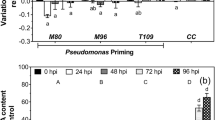
Under phytotoxin exposure, ROS interact with phytohormones to induce cell death (Overmyer et al. 2005). In addition, ROS production can cause numerous changes in the cellular components such as inactivation of enzymes, degradation of proteins and nucleic acids, and lipid peroxidation (Maksymiec and Krupa 2006). The contribution of ROS overproduction in plants to FA phytotoxicity has been a topic of interest for researchers (Malerba and Cerana 2021; Liu et al. 2020). Different concentrations of FA have been used in plants to induce ROS accumulation and to study their toxic effects on cellular functions (Sapko et al. 2011; Bouizgarne et al. 2006a). The results showed a time- and concentration-dependent phytotoxic effect of FA. For example, treatment of tomato plants with FA (0.1–1 mM) significantly increased ROS production after 24–72 h, and this oxidative burst damaged cellular functions and photosynthetic activity as well (Iqbal et al. 2021a). Similarly, FA doses of 50–300 µg/mL induced ROS generation (O2.− and H2O2) in tomato leaves after 72 h of treatment (Singh et al. 2017; Singh and Upadhyay 2014, 2016). Similarly, FA concentrations in the range of 10−8–10−3 M induced H2O2 production in cell suspension of potato plants after 48 h of exposure (Sapko et al. 2011). In comparison, various FA concentrations caused ROS accumulation (H2O2) and disrupted the cellular processes in many agronomic crops such as tobacco (Jiao et al. 2014), cucumber (Wang et al. 2020) and Arabidopsis (Bouizgarne et al. 2006a). In addition, high ROS (O2.−, H2O2) accumulation was also observed in saffron (Samadi and Shahsavan Behboodi 2006), tomato (Kuźniak 2001) and banana plants (Fung et al. 2019) under different FA doses, where the duration of FA exposure determined its effect on plant growth and development. Fung et al. (2019) reported rapid accumulation of both O2.− and H2O2 during FA-producing fungal infection at the early stage in banana plants. At the same time, higher ROS generation did not slow down the fungal infection, indicating that the accumulated ROS were not sufficient to prevent fungal pathogenicity (Fung et al. 2019). However, fungal tolerance to higher ROS production is still unknown. Interestingly, another study reported higher production of O2.− in culture medium (extracellular or filtrate) as compared to cellular extracts of tomato separated from culture medium (Kuźniak 2001). Nevertheless, H2O2 production showed the same levels in the culture medium and cellular extract upon FA treatment as compared to their respective controls (Kuźniak 2001). In addition, increased O2.− production was detected in the culture medium 12 h after the FA exposure and increased H2O2 production was measured 48 h after the treatment in cellular extracts (Kuźniak 2001). Therefore, these results clearly indicate that rapid or gradual induction of ROS production is directly proportional to the dose and exposure time of FA. The results also suggest that ROS accumulation may vary between plants and even within cells or extracellular spaces.
FA stress induced plasmolysis, cytoplasmic vacuole formation and cytoplasmic shrinkage in tobacco cell suspensions (Jiao et al. 2013). Interestingly, nitric oxide (NO) scavengers significantly reduced these FA-induced toxic effects in tobacco cells (Jiao et al. 2013). The PAL (phenylalanine ammonia-lyase) and Hsr203J (hypersensitivity-related protein) genes were investigated for their potential role in regulating plant–pathogen interactions and enhancing plant immunity against fungi or their phytotoxins (Jiao et al. 2013). In addition, the NO scavenger 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (CPTIO) also reduced the caspase-3-like activity and the expression of PAL and Hsr203J genes, indicating the crucial role of the NO signalling pathway in the regulation of plant cell death under FA exposure (Jiao et al. 2013). Therefore, the application of NO scavenger can also significantly reduce FA-induced phytotoxic effects and promote plant growth and development, confirming the significant role of NO under FA exposure.
FA-induced changes in enzymatic and non-enzymatic antioxidants
Plants contain both enzymatic and non-enzymatic antioxidants to attenuate ROS accumulation under phytotoxin FA exposure (Fung et al. 2019; Singh et al. 2017). These ROS scavengers maintain ROS homeostasis in the cells by removing ROS and play an important role in oxidative metabolism. ROS accumulation upon FA exposure induces both enzymatic and non-enzymatic antioxidants (Antić et al. 2020; Dong et al. 2014). These ROS-scavenging antioxidants include superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), peroxidase (POD), glutathione peroxidase (GPX), glutathione reductase (GR), glutathione S-transferase (GST), dehydroascorbate reductase (DHAR), glutathione (GSH), ascorbate (ASC) and phenolic compounds (Sharma et al. 2022; Hashem et al. 2021; Gupta et al. 2018; Sapko et al. 2011; Foyer and Noctor 2009). These antioxidants are responsible for cell protection and defence against toxic levels of overproduced ROS following fungal attacks (Gill and Tuteja 2010). SOD catalyses the dismutation of O2.− radicals to O2 and H2O2. Similarly, CAT and APX catalyse the conversion of H2O2 molecules into water and oxygen molecules. POD is responsible for the conversion of H2O2 and dangerous hydroperoxides (ROOH) to water and harmless ROH species. Similarly, GR can increase reduced GSH levels under oxidative stress and GST can enhance plant resistance to fungal pathogens by conjugating GSH with their toxins for the process of detoxification (Fung et al. 2019; Gullner et al. 2018; Liu et al. 2015). GPX can convert H2O2 to water and oxygen, and peroxide radicals to alcohol and oxygen molecules, while DHAR is involved in the conversion of dehydroascorbic acid to ascorbic acid (Loi et al. 2020). Nevertheless, further scientific research is needed to understand the role of these ROS-scavenging compounds under phytotoxin exposure, and their involvement in alleviating phytotoxin-induced oxidative damage in plants. The mechanisms of different ROS-scavenging antioxidants under FA exposure are summarized in Fig. 3.

Mechanism of ROS detoxification by enzymatic and non-enzymatic antioxidants under fusaric acid (FA) exposure. CAT catalase, SOD superoxide dismutase, POD peroxidase, GST glutathione S-transferase, GPX glutathione peroxidase, APX ascorbate peroxidase, GR glutathione reductase, DHAR dehydroascorbate reductase, GSH glutathione, ASC ascorbate
FA-induced oxidative stress affected antioxidant levels in several plant species. For example, treatment with 100 µM FA reduced APX and CAT activities in tobacco cell suspension culture after 24 h (Jiao et al. 2014). Similarly, Hashem et al. (2021) infected tomato plants with the FA-producing fungus F. oxysporum and then observed that the activities of the enzymes DHAR and GPX decreased, while the enzymatic activities of SOD and APX increased as a result of the fungal infection. Conversely, higher SOD (after 48 h) and CAT (after 12 and 24 h) activities were significantly increased after FA treatment (2 × 10−4 M) in cell suspensions of tomato plants (Kuźniak 2001). The same trend was observed for other antioxidants such as APX and POD. However, DHAR activity and ASC content were reduced under FA stress in a time-dependent manner (Kuźniak 2001). Similarly, different concentrations of FA treatments (0–400 mg/L) resulted in initially enhanced activities of SOD, POD and CAT in watermelon leaves, which then gradually decreased in a time-dependent manner in a 24-h-long experiment (Wu et al. 2008b). Similarly, banana seedlings infected with an FA-producing fungus showed significantly higher activities of GST, SOD, POD, CAT, GR and APX initially, which then gradually decreased in a 10-day-long experiment (Fung et al. 2019). Different concentrations of FA or FA-producing fungal species were used to determine the detoxification capacities of different enzymatic and non-enzymatic antioxidants in several plant species such as cucumber (Wang et al. 2020), potato (Sapko et al. 2011), wax gourd (Wang et al. 2021) and chihuahua flower (Antić et al. 2020). Interestingly, tomato leaves showed lower activities of CAT and APX due to the possible inhibitory effect of FA on these ROS scavengers, but SOD activity was increased with increasing time duration to detoxify ROS overproduction (Singh and Upadhyay 2014). Another study showed higher peroxidase (POX) activity due to its involvement in the cross-linking of cell wall proteins (Singh and Upadhyay 2016). FA treatment elevated the level of H2O2 which was detoxified by the POX enzyme. Furthermore, FA exposure decreased POD and CAT activities in wheat and faba bean (intercropping) plants, as the FA concentration increased from 0 to 200 mg/L (Li et al. 2021); however, the highest POD activity and the highest level of total phenolic compounds were found in banana plants during fungal infection due to FA production (Dong et al. 2014). These FA-induced changes in the antioxidant system could affect how plants respond by increasing or decreasing the amount of antioxidants produced, which can result in either plant survival or cell death. In addition, FA can have an inhibitory effect on antioxidant activities and higher levels can kill plant cells (Wu et al. 2008b; Singh and Upadhyay 2014). Therefore, a potent antioxidant system plays a critical role in detoxifying ROS produced in cellular compartments upon exposure to fungi or phytotoxin FA. The antioxidant system promotes plant growth by enhancing plant defence responses against phytotoxins such as FA. It is also noteworthy, that FA-induced changes in antioxidant activities are dependent on plant species, FA concentration, the examined plant organ and FA exposure time.
FA-mediated lipid peroxidation
As a fundamental component of biological membranes, lipids play a vital role in cellular function. FA-induced oxidative stress can damage lipids and disrupt the proper functioning of cells. ROS attack the carbon–carbon double bonds in polyunsaturated fatty acids, generating hydroperoxides and peroxyl radicals (Ayala et al. 2014). The disturbance of the plasma membrane caused by the lipid peroxyl radicals’ accelerated peroxidation alters the fluidity and permeability of the membrane, disrupts ion transport and interferes with cell signalling (Birben et al. 2012). In addition, 4-hydroxynonenal (HNE) and malondialdehyde (MDA) are reactive products of lipid peroxidation and are considered as indicators of oxidative stress (Arumugam et al. 2021).
Lipid peroxidation by FA exposure depends on the exposure time and the concentration of the phytotoxin. FA concentration between 50 and 200 mg/L resulted in lipid oxidation, and significantly increased the MDA content in wheat and faba bean seedlings (Li et al. 2021). Similarly, FA exposure at a concentration of 100 mg/L increased the MDA level in cape gooseberry plant after 12 h (Mendoza-Vargas et al. 2021). Further, other studies also showed that exposure of watermelon to FA (0–400 mg/L) increased MDA levels at 24 h. The MDA content significantly increased with the duration of FA exposure, reaching a maximum (5–11-fold) at 24 h post treatment as compared to control plants (Wu et al. 2008b). Interestingly, MDA is expressed as thiobarbituric acid-reacting substances (TBARS) (Singh et al. 2017). Therefore, numerous studies have been conducted on FA-induced oxidative stress leading to lipid peroxidation, and increased MDA and TBARS levels in the FA-treated plants. For example, FA (200 mg/L) significantly reduced the MDA levels in the roots of wax gourd during a 48-h-long experiment (Wang et al. 2021). In contrast, lipid peroxidation caused by FA treatment (100 µM) resulted in higher MDA levels (threefold) in tobacco cell suspension at 12 and 24 h; however, no rapid MDA generation was observed after treatment for 3 and 6 h (Jiao et al. 2014). In parallel, MDA levels in cucumber, banana, tomato, potato and chihuahua flowers were significantly increased as a function of the exposure time and increasing FA concentrations (Iqbal et al. 2021a; Wang et al. 2020; Fung et al. 2019; Dong et al. 2014; Sapko et al. 2011). In addition, higher MDA production was observed at the early stage of fungal infection after 2 h, which then gradually decreased until 6 h and finally started to increase again until day 7 after treatment with FA-producing fungi (Antić et al. 2020). Similarly, the toxic effects of FA also increased TBARS production, especially in tomato leaves, depending on the exposure time and concentration of FA (Singh and Upadhyay 2014, 2016). These observations suggest that FA toxicity can induce lipid peroxidation, which disrupts important cellular functions and reduces cellular stability. In addition, the disruption of biological functions of membranes, such as cell signalling, was found to increase the damage to cellular membranes in FA-treated plants as compared to control plants.
FA-induced effects on hormone signalling
Pathogen attack can trigger defence or resistance mechanisms in various plants. The major phytohormones, such as salicylic acid (SA), jasmonic acid (JA) and ethylene (ET), induce signal transduction via complex signalling pathways (Zhang et al. 2022; Ederli et al. 2021; Dhar et al. 2020). The defence responses mediated by SA- and JA-dependent signalling mechanisms are triggered against biotrophic and necrotrophic pathogens, respectively. However, the defence mechanisms against hemi-biotrophic and necrotrophic fungal species are much more complex (Palazzini et al. 2018). It is interesting to note that SA and JA compete with each other in the resistance responses to infections (Poór et al. 2017; Denancé et al. 2013). Higher ROS levels induced by phytotoxins can lead to increased SA production, resulting in HR-like cell death at the site of infection to prevent the spread of infection (Ding and Ding 2020; Poór et al. 2017). Already low levels of SA can initiate a systemic acquired response for additional defence mechanisms against pathogenic attack, leading to the induction of numerous antimicrobial pathogenesis-related proteins (Iqbal et al. 2021b). Besides SA, JA is also produced during pathogenic attacks or injury which plays an essential role in the synthesis of the antimicrobial plant defensin (PDF) and protease inhibitors, but it is also responsible for the positive regulation of plant cell death by altering oxidative metabolism (Coll et al. 2011). Interestingly, SA negatively affects ET signalling, and its biosynthesis under stress conditions (Poór et al. 2013).
The role of ET in FA-treated leaves of tomato plants was determined in a 72-h-long experiment (Singh and Upadhyay 2014). The results showed a significant increase in ET production upon FA exposure as compared to control plants. However, ET emission was the highest 6 h after the treatment, indicating the involvement of ET in the initiation of cell death under phytotoxin stress. Higher level of ET production was also related to enhanced ROS generation. Therefore, ET and ROS induce each other’s production, which eventually leads to cell death (Singh and Upadhyay 2014). Recently, Iqbal et al. (2021a) explained the crucial role of ET in defence progression in tomato leaves of two genotypes, Ailsa Craig (WT) and ET receptor Never ripe (Nr) mutant plants under FA exposure between of 0.1 and 1.0 mM after 24- and 72-h-long treatments. The results showed higher ET production after 72 h following FA treatment at higher concentrations, suggesting a time- and concentration-dependent effect of FA, and FA-induced ET production in tomato leaves. On the basis of these data, Nr leaves were more sensitive to FA phytotoxicity, as compared to WT plants (Iqbal et al. 2021a). In parallel, another study reported the effect of FA (200 mg/L) on the expression of six genes encoding ET responsiveness in Benincasa hispida seedlings and found higher induction of four transcription factors, ACC oxidase and ET insensitive-like 1 protein isoform X1, suggesting the involvement of ET in modulating and regulating FA-induced oxidative stress (Wang et al. 2021). Nevertheless, further research is required for a comprehensive understanding of hormonal signalling under phytotoxin exposure.
FA-elicited physiological effects
FA-induced phytotoxicity can affect the osmotic pressure, ion flux, micronutrient uptake, water potential, and it can result in the reduction of stomatal size, alterations in membrane permeability and cell death processes in plants (Wang et al. 2015; Dong et al. 2012; Bouizgarne et al. 2004). These stomatal abnormalities affect respiration and photosynthesis as well (Iqbal et al. 2021a). It has been reported that FA exposure mainly affects the electron transport chain, enzymatic activities and CO2 fixation (Singh et al. 2017). However, Wang and co-workers (2013, 2015, 2020) conducted experiments on cucumber leaves, and revealed also the toxic effects of FA on the photosynthetic process in a time- and FA concentration-dependent manner. Furthermore, FA stress leads to mitochondrial dysfunction due to higher levels of oxidative stress, leading to the initiation of cell death (Liu et al. 2020). For example, Dendrobium sonia 28 strain showed a higher mortality rate and growth reduction upon FA exposure (Dehgahi et al. 2016). Similarly, a high dose of FA exposure inhibited root growth and development of maize seedlings (Spss and Oliveira 2009). Likewise, the cell suspension cultures of Nicotiana tabacum treated with 100 µM of FA phytotoxin showed dilation of the endoplasmic reticulum and damage to cisternae (Jiao et al. 2013). In addition, FA-induced toxicity (1 mM) reduced root cell viability and adversely affected plasmodesmata and mitochondrial structure (Pavlovkin et al. 2004).
However, a number of other plant species have also been shown to be damaged by FA and the disease symptoms were associated with Fusarium infection. For example, tobacco cell suspensions showed significantly higher H2O2 production, lipid peroxidation and mitochondrial dysfunction under 100 µM of FA exposure after 24 h (Jiao et al. 2014). Similarly, FA treatment of Arabidopsis seedlings reduced root and root hair growth along with induction of membrane hyperpolarisation (Bouizgarne et al. 2006b). Similarly, FA-induced physicochemical changes such as reduced photosynthetic activity and plant biomass, turgor loss, as well as high proline content were observed in cape gooseberry plants at 100 mg/L FA concentration (Mendoza-Vargas et al. 2021). In addition, higher oxidative stress and antioxidant activity and reduced cell viability were observed in tomato cell suspensions at 2 × 10−4 M dose of FA. These FA-induced effects were enhanced with prolonged duration of FA exposure from 1 to 48 h (Kuźniak 2001). Furthermore, Hashem et al. (2021) documented the toxic effects of FA-producing Fusarium sp. on tomato plants showing wilting symptoms, reduction in chlorophyll content and plant growth. In addition, FA treatment (0.1–0.5 mM) in potato shoots significantly reduced the plant growth and length in a 4-week-long experiment (Arici and Meryem 2017). Surprisingly, in chihuahua plants (Tacitus bellus L.) infected with FA-producing Fusarium species, MDA levels were significantly reduced in a time-dependent manner together with high oxidative stress (Antić et al. 2020). In addition, wheat and faba bean plants showed growth inhibition under FA exposure when grown separately, but intercropping significantly reduced FA toxicity (Li et al. 2021). FA exposure at various doses can result in a variety of physiological and biochemical changes in different plant species, as well as impaired plant growth and development. However, these FA-mediated toxic effects depend on the plant species, the FA concentrations and the targeted plant organs in addition to the duration of exposure. In addition, plants resistant to FA-producing Fusarium species showed less toxicity symptoms and other physiological effects as compared to Fusarium-susceptible plants (Bouizgarne et al. 2004; Barna et al. 2011; Xie et al. 2011). These findings can also be applied to other phytotoxins to study their effects on plant growth and development, as well as metabolic processes (Table 1).
FA-induced DNA fragmentation and genotoxicity
To maintain cellular homeostasis, plant cell death under toxin exposure or fungal infection eliminates old, damaged or diseased plant cells in a highly coordinated manner. In addition, cell death plays an essential role in the coordination of plant growth and development, as well as in the induction of cellular responses to various stresses (Valandro et al. 2020). Plants induce cell death at the infection site to prevent further spread of pathogen infection and toxin production (Iqbal et al. 2022; Xing et al. 2013). Saffron root tip cells were exposed to 25–200 µM FA for 60 h, and it was observed that FA induced higher H2O2 accumulation, chromatin condensation and DNA breakage followed by cell death (Samadi and Shahsavan Behboodi 2006). The maximum DNA fragmentation (50–60%) was detected at 25–100 µM FA over the different time periods, but less DNA fragmentation was observed at 200 µM of FA exposure. Furthermore, DNA fragmentation was suppressed by the activation of various inhibitors such as serine protease and caspase (Samadi and Shahsavan Behboodi 2006). FA exposure also resulted in the condensation of chromatin material (spheres budding from the nucleus) and the release of cytochrome c from mitochondria into cytosol (Samadi and Shahsavan Behboodi 2006). FA toxicity was also reported in tobacco cell suspension exposed to 100 µM FA for 24 h, showing high production of NO leading to cell death, dilated endoplasmic reticulum, chromatin condensation and DNA fragmentation (Dehgahi et al. 2016). Similarly, FA exposure (50–300 µg/mL) caused lipid peroxidation and formation of necrotic lesions in tomato leaves, followed by DNA degradation. FA infiltration into tomato leaves resulted in DNA fragmentation on agarose gel, whereas DNA fragmentation was absent in the case of control leaf tissues (Singh and Upadhyay 2014). Therefore, it can be concluded that FA toxicity may have detrimental effects on the control of defence-related genes engaged in hormonal signalling pathways (e.g. JA, ET and SA) regulated by ROS and NO, as well as cause DNA damage or fragmentation by activating the proteases in the cytosol upon FA exposure, which may contribute to cell death.
Approaches to control FA toxicity
Application of metal ions
FA toxicity can affect various physiological, biochemical and metabolic processes at the molecular level, which can hinder plant growth and development resulting in crop yield loss. Several strategies have been used to moderate or reduce FA toxicity in plants (Wang et al. 2020). For example, the use of exogenous zinc (Zn) and copper (Cu) increased the tolerance of cucumber plants to FA toxicity (Wang et al. 2020). The results showed that both Zn and Cu significantly reduced the deleterious effects of FA exposure, and altered the antioxidant activity. Application of these metals reduced wilting symptoms, cell membrane damage of roots and leaves, and promoted the plants’ growth and photosynthetic activity (Wang et al. 2020). In addition, Zn and Cu alleviated FA toxicity by 17% and 20%, respectively, by reducing the H2O2 production and lipid peroxidation (Wang et al. 2020). Furthermore, both metal treatments enhanced antioxidant enzymatic activities (SOD, CAT and POD) by preventing FA-induced oxidative burst (Wang et al. 2020). Cu and Zn have a significant influence on FA tolerance in cucumber plants by modifying the uptake and transport of FA in plants (Wang et al. 2020).
Biological detoxification
Several bacterial and fungal species have been tested to detoxify or degrade FA. Interestingly, some antagonistic fluorescent strains of Pseudomonas sp. showed resistance to FA at a concentration of 500 ppm (Fakhouri et al. 2003). In addition, a non-pathogenic fungal strain of Colletotrichum sp. was found to be more efficient in detoxifying FA (200 ppm) after 4 days of treatment. This fungal strain degraded FA to 4-butyl-2-carboxypyrimidine, which had no toxic effects on tomato seedlings and fungal spore germination. However, Colletotrichum sp. was unable to degrade the high concentration of FA (400 ppm) and showed no growth even after 10 days of treatment (Fakhouri et al. 2003). Maina and co-workers (2008) used three bacterial strains, namely Pseudomonas putida 53, Pseudomonas fluorescens T58 and Bacillus sphaericus B43, to reduce the toxic effects of FA on tomato plants. These bacterial strains activated induced systemic resistance (ISR) to reduce the damage caused by F. oxysporum f. sp. lycopersici, which produces FA in tomato plants. The results showed that the H2O2 accumulation, chlorophyll degradation and ion leakage were significantly reduced in bacteria-treated plants as compared to control plants. In addition, SOD and GPX activities were reduced following bacterial treatment. However, ROS such as O2.− showed higher levels in bacteria-treated plants in a time-dependent manner, eventually leading to membrane damage (Maina et al. 2008). These results clearly showed that ISR caused by biocontrol agents may be relevant to prevent FA-related damage in plants and serve as a basis for further studies to elucidate the unidentified mechanisms of action of biocontrol agents.
In addition, another fungal strain, Mucor rouxii, has shown the potential to degrade toxic FA to 8-hydroxyfusaric acid, a less toxic compound in Gossypium hirsutum and G. barbadense (Crutcher et al. 2017). The hydroxylation of the butyl chain of FA resulted in detoxification, and showed less phytotoxic effects in cotton plants as compared to the parent compound (FA) (Crutcher et al. 2017). Interestingly, controlling the genes responsible for this hydroxylation could be helpful in developing biocontrol agents to reduce FA toxicity (Crutcher et al. 2017). Similarly, the Burkholderia ambifaria T16 strain, which was isolated from the rhizosphere of barley, was shown to detoxify FA in barley seedlings and to rely solely on FA for its carbon, nitrogen and energy sources. Inoculation of barley seedlings with this bacterial species increased cell viability, cell density, indole-3-acetic acid production and biofilm formation (Simonetti et al. 2018). Furthermore, B. ambifaria T16 treatment also improved siderophore production, ACC deaminase activity and resistance to phytopathogenic Fusarium sp. (Simonetti et al. 2018). These results confirm the important role of B. ambifaria T16 in FA detoxification, and open up new avenues for the development of gene-modified FA detoxification technologies. A recent study also reported the conversion of FA into less toxic compounds, when the FA-producing fungus F. verticillioides was co-cultured with another fungus, Gliocladium roseum (Kuang et al. 2022). Using nuclear magnetic resonance (NMR) and mass spectrophotometry, Kuang et al. (2022) detected four different FA products in the co-culture inoculation and identified them as 4-(5-butylpicolinamido)butanoic acid (45BBA), 5-butylpyridine-2-carboxylic acid methyl ester (5B2CAM), bis(5-butyl-2-pyridinecarboxylate-N1,O2)-copper (B52P) and methyl 4-(5-butylpicolinamido)butanoate (M45BBA). These FA derivatives were tested on Botrytis cinerea and Aspergillus niger and showed reduced toxicity as compared to the parent compound FA. The 45BBA derivative of FA showed the lowest toxicity among tested FA derivatives (Kuang et al. 2022). Therefore, these results indicate the efficient antagonistic interaction between these fungal species leading to the biotransformation or detoxification of FA (Brauer et al. 2019).
Future prospects
In addition to computational and experimental techniques, recent developments in “omics” approaches such as proteomics, transcriptomics, metagenomics, lipidomics and metabolomics have provided us new opportunities to search for novel bioactive fungicides for the agricultural sector in order to reduce crop losses and increase crop productivity (Brauer et al. 2019). For example, epigenetic approaches could use RNA interference such as bidirectional trafficking of plant–pathogen miRNAs to control pathogens and toxin production (Ratajczak and Ratajczak 2016). Similarly, metabolomics helps to study microbial interactions with plants and identify novel bioactive biomarkers with high efficacy and pathogen specificity to control pathogenic fungi (Johanningsmeier et al. 2016). Similarly, the integrated approach of metagenomics could play an essential role in the production of antifungal drugs, using an integrated method of metabolite evaluation and molecular library screening to select a drug candidate through the application of computational models (Brauer et al. 2019). However, more research is needed to find new antifungal molecules that can solve the problem of antifungal drug resistance in plants. The development of new chemicals against fungal species is a challenging task because of the time and resources required, as well as the need for accurate identification and characterisation of candidate compounds. The discovery of new antifungal molecules is far less frequent than the emergence of fungal species resistant to antifungal drugs. In addition, modification of the genome of economically important crops, either by breeding or transgenesis could be useful to control Fusarium diseases and their potential phytotoxins, including FA. The nature of plant tolerance or resistance to fungal infection, which can reduce crop yield and produce fungal resistant crops under biotic stress conditions, therefore needs to be understood in great detail. In addition, to advance the scientific understanding of FA phytotoxicity and its hidden host plant infection mechanism, the present study will be useful in understanding the long-term control of the phytotoxin. Further research on the synergistic effects of two or more phytotoxins is also needed to understand their phytotoxic effects in cross-kingdoms. To promote biocontrol of fungal diseases caused by FA, additional knowledge is needed to screen biocontrol strains that secrete siderophores and other metal chelating agents, such as iron supplementation to limit FA production and the growth of fungal pathogens.
Conclusion
Exposure to FA increases oxidative stress via the enhanced accumulation of ROS, and affects both enzymatic and non-enzymatic antioxidant processes. FA-mediated ROS production also causes changes in hormonal signalling, photosynthetic apparatus, cellular structure and DNA damage. These FA-induced changes include mitochondrial dysfunction, lipid peroxidation, chlorosis and ultimately impaired plant growth and development, resulting in loss of crop production. However, various strategies such as the use of metals, such as Zn and Cu, co-culture inoculation with biocontrol agents and control of the FA biosynthesis genes in fungal species are being used to control FA production and reduce its effects in various crops.
References
Ali M, Cheng Z, Ahmad H, Hayat S (2018) Reactive oxygen species (ROS) as defences against a broad range of plant fungal infections and case study on ROS employed by crops against Verticillium dahliae wilts. J Plant Interact 13:353–363
Alshannaq A, Yu JH (2017) Occurrence, toxicity, and analysis of major mycotoxins in food. Int J Environ Res Public Health 14:632
Antić TC, Janošević D, Maksimović VM, Živić M, Budimir S, Glamočlija J, Mitrović AL (2020) Biochemical and histological characterization of succulent plant Tacitus bellus response to Fusarium verticillioides infection in vitro. J Plant Physiol 244:153086
Arici ŞE, Meryem SARI (2017) Growth and antioxidant responses of potato (Solanum tuberosum, cv Agria) shoots cultured in vitro under different fusaric acid and boron concentrations. Plant Prot Bull 57:73–87
Arumugam T, Ghazi T, Abdul NS, Chuturgoon AA (2021) A review on the oxidative effects of the fusariotoxins: fumonisin B1 and fusaric acid. Toxicology. Elsevier, Amsterdam, pp 181–190
Ayala A, Munoz MF, Arguelles S (2014) Lipid peroxidation: production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid Med Cell Longev 2014:360438
Barna B, Jansen C, Kogel KH (2011) Sensitivity of barley leaves and roots to fusaric acid, but not to H2O2, is associated with susceptibility to Fusarium infections. J Phytopathol 159:720–725
Birben E, Sahiner UM, Sackesen C, Erzurum S, Kalayci O (2012) Oxidative stress and antioxidant defence. World Allergy Organ J 5:9–19
Bouizgarne B, Brault M, Pennarun AM, Rona JP, Ouhdouch Y, El Hadrami I, Bouteau F (2004) Electrophysiological responses to fusaric acid of root hairs from seedlings of date palm-susceptible and-resistant to Fusarium oxysporum f. sp. albedinis. J Phytopathol 152:321–324
Bouizgarne B, El-Maarouf-Bouteau H, Frankart C, Reboutier D, Madiona K, Pennarun AM, Monestiez M, Trouverie J, Amiar Z, Briand J, Brault M (2006a) Early physiological responses of Arabidopsis thaliana cells to fusaric acid: toxic and signalling effects. New Phytol 169:209–218
Bouizgarne B, El-Maarouf-Bouteau H, Madiona K, Biligui B, Monestiez M, Pennarun AM, Amiar Z, Rona JP, Ouhdouch Y, El Hadrami I, Bouteau F (2006b) A putative role for fusaric acid in biocontrol of the parasitic angiosperm Orobanche ramosa. Mol Plant Microbe Interact 19:550–556
Brauer VS, Rezende CP, Pessoni AM, De Paula RG, Rangappa KS, Nayaka SC, Gupta VK, Almeida F (2019) Antifungal agents in agriculture: friends and foes of public health. Biomolecules 9:521
Breusegem FV, Dat JF (2006) Reactive oxygen species in plant cell death. Plant Physiol 141:384–390
Brown DW, Butchko RA, Busman M, Proctor RH (2012) Identification of gene clusters associated with fusaric acid, fusarin, and perithecial pigment production in Fusarium verticillioides. Fungal Genet Biol 49:521–532
Brown DW, Lee SH, Kim LH, Ryu JG, Lee S, Seo Y, Kim YH, Busman M, Yun SH, Proctor RH, Lee T (2015) Identification of a 12-gene fusaric acid biosynthetic gene cluster in Fusarium species through comparative and functional genomics. Mol Plant Microbe Interact 28:319–332
Cen YK, Lin JG, Wang YL, Wang JY, Liu ZQ, Zheng YG (2020) The gibberellin producer Fusarium fujikuroi: methods and technologies in the current toolkit. Front Bioeng Biotechnol 8:232
Coll NS, Epple P, Dangl JL (2011) Programmed cell death in the plant immune system. Cell Death Differ 18:1247–1256
Crutcher FK, Puckhaber LS, Bell AA, Liu J, Duke SE, Stipanovic RD, Nichols RL (2017) Detoxification of fusaric acid by the soil microbe Mucor rouxii. J Agric Food Chem 65:4989–4992
Curir P, Guglieri L, Dolci M, Capponi A, Aurino G (2000) Fusaric acid production by Fusarium oxysporum f. sp. lilii and its role in the lily basal rot disease. Eur J Plant Pathol 106:849–856
da Rocha MEB, Freire FDCO, Maia FEF, Guedes MIF, Rondina D (2014) Mycotoxins and their effects on human and animal health. Food Control 36:159–165
Dehgahi R, Zakaria L, Mohamad A, Joniyas A, Subramaniam S (2016) Effects of fusaric acid treatment on the protocorm-like bodies of Dendrobium sonia-28. Protoplasma 253:1373–1383
Denancé N, Sánchez-Vallet A, Goffner D, Molina A (2013) Disease resistance on growth: the role of plant hormones in balancing immune responses and fitness costs. Front Plant Sci 4:155
Devnarain N, Tiloke C, Nagiah S, Chuturgoon AA (2017) Fusaric acid induces oxidative stress and apoptosis in human cancerous oesophageal SNO cells. Toxicon 126:4–11
Dhar N, Chen JY, Subbarao KV, Klosterman SJ (2020) Hormone signaling and its interplay with development and defence responses in Verticillium-plant interactions. Front Plant Sci 11:584997
Ding P, Ding Y (2020) Stories of salicylic acid: a plant defence hormone. Trends Plant Sci 25:549–565
Ding Z, Yang L, Wang G, Guo L, Liu L, Wang J, Huang J (2018) Fusaric acid is a virulence factor of Fusarium oxysporum f. sp. cubense on banana plantlets. Trop Plant Pathol 43:297–305
Dong X, Ling N, Wang M, Shen Q, Guo S (2012) Fusaric acid is a crucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Plant Physiol Biochem 60:171–179
Dong X, Xiong Y, Ling N, Shen Q, Guo S (2014) Fusaric acid accelerates the senescence of leaf in banana when infected by Fusarium. World J Microbiol Biotechnol 30:1399–1408
Ederli L, Salerno G, Quaglia M (2021) In the tripartite combination Botrytis cinerea–Arabidopsis–Eurydema oleracea, the fungal pathogen alters the plant–insect interaction via jasmonic acid signalling activation and inducible plant-emitted volatiles. J Plant Res 134:523–533
Fakhouri W, Walker F, Armbruster W, Buchenauer H (2003) Detoxification of fusaric acid by a nonpathogenic Colletotrichum sp. Physiol Mol Plant Pathol 63:263–269
Fernandez-Pol JA, Klos DJ, Hamilton PD (1993) Cytotoxic activityof fusaric acid on human adenocarcinoma cells in tissue culture. Anticancer Res 13:57–64
Foyer CH, Noctor G (2009) Redox regulation in photosynthetic organisms: signaling, acclimation, and practical implications. Antioxid Redox Signal 11:861–905
Fung SM, Razali Z, Somasundram C (2019) Reactive oxygen species scavenging enzyme activities in berangan banana plant infected by Fusarium oxysporum f. sp. cubense. Chiang Mai J Sci 46:1084–1095
Ghazi T, Nagiah S, Tiloke C, Sheik Abdul N, Chuturgoon AA (2017) Fusaric acid induces DNA damage and post-translational modifications of p53 in human hepatocellular carcinoma (HepG2) cells. J Cell Biochem 118(11):3866–3874
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Gullner G, Komives T, Király L, Schröder P (2018) Glutathione S-transferase enzymes in plant-pathogen interactions. Front Plant Sci 9:1836
Gupta DK, Palma JM, Corpas FJ (eds) (2018) Antioxidants and antioxidant enzymes in higher plants. Springer, Berlin
Hashem A, Akhter A, Alqarawi AA, Singh G, Almutairi KF, Abd Allah EF (2021) Mycorrhizal fungi induced activation of tomato defence system mitigates Fusarium wilt stress. Saudi J Biol Sci 28:5442–5450
Iqbal N, Czékus Z, Ördög A, Poór P (2021a) Ethylene-dependent effects of fusaric acid on the photosynthetic activity of tomato plants. Photosynthetica 59:337–348
Iqbal N, Czékus Z, Poór P, Ördög A (2021b) Plant defence mechanisms against mycotoxin fumonisin B1. Chem Biol Interact 343:109494
Iqbal N, Czékus Z, Angeli C, Bartók T, Poór P, Ördög A (2022) Fumonisin B1-induced oxidative burst perturbed photosynthetic activity and affected antioxidant enzymatic response in tomato plants in ethylene-dependent manner. J Plant Growth Regul 42(3):1865–1878
Iqbal N, Czékus Z, Poór P, Ördög A (2023) Ethylene-dependent regulation of oxidative stress in the leaves of fusaric acid-treated tomato plants. Plant Physiol Biochem 196:841–849
Ismaiel AA, Papenbrock J (2015) Mycotoxins: producing fungi and mechanisms of phytotoxicity. Agriculture 5:492–537
Jiao J, Zhou B, Zhu X, Gao Z, Liang Y (2013) Fusaric acid induction of programmed cell death modulated through nitric oxide signalling in tobacco suspension cells. Planta 238:727–737
Jiao J, Sun L, Zhou B, Gao Z, Hao Y, Zhu X, Liang Y (2014) Hydrogen peroxide production and mitochondrial dysfunction contribute to the fusaric acid-induced programmed cell death in tobacco cells. J Plant Physiol 171:1197–1203
Johanningsmeier SD, Harris GK, Klevorn CM (2016) Metabolomic technologies for improving the quality of food: practice and promise. Annu Rev Food Sci Technol 7:413–438
Karuppanapandian T, Moon JC, Kim C, Manoharan K, Kim W (2011) Reactive oxygen species in plants: their generation, signal transduction, and scavenging mechanisms. Aust J Crop Sci 5:709–725
Kuang Y, Scherlach K, Hertweck C, Yang S, Sampietro DA, Karlovsky P (2022) Fusaric acid detoxification: a strategy of Gliocladium roseum involved in its antagonism against Fusarium verticillioides. Mycotoxin Res 38(1):13–25
Kuruvilla TS, Dias M (2012) Fusarium solani: a causative agent of skin and nail infections. Indian J Dermatol 57:308
Kuźniak E (2001) Effects of fusaric acid on reactive oxygen species and antioxidants in tomato cell cultures. J Phytopathol 149:575–582
Li Y, Lv J, Zhao Q, Chen L, Dong Y, Dong K (2021) Wheat/faba bean intercropping improves physiological and structural resistance of faba bean to fusaric acid stress. Plant Pathol 70:827–840
Liu X, Zheng X, Song Z, Liu X, Kumar-Kopparapu N, Wang X, Zheng Y (2015) Preparation of enzymatic pretreated corn gluten meal hydrolysate and in vivo evaluation of its antioxidant activity. J Funct Foods 18:1147–1157
Liu S, Dai H, Orfali RS, Lin W, Liu Z, Proksch P (2016) New fusaric acid derivatives from the endophytic fungus Fusarium oxysporum and their phytotoxicity to barley leaves. J Agric Food Chem 64:3127–3132
Liu S, Li J, Zhang Y, Liu N, Viljoen A, Mostert D, Zuo C, Hu C, Bi F, Gao H, Li C (2020) Fusaric acid instigates the invasion of banana by Fusarium oxysporum f. sp. cubense TR 4. New Phytol 225:913–929
Loi M, De Leonardis S, Mulè G, Logrieco AF, Paciolla C (2020) A novel and potentially multifaceted dehydroascorbate reductase increasing the antioxidant systems is induced by beauvericin in tomato. Antioxidants 9:435
López-Díaz C, Rahjoo V, Sulyok M, Ghionna V, Martín-Vicente A, Capilla J, Di Pietro A, López-Berges MS (2018) Fusaric acid contributes to virulence of Fusarium oxysporum on plant and mammalian hosts. Mol Plant Pathol 19:440–453
Ma Q (2013) Role of nrf2 in oxidative stress and toxicity. Annu Rev Pharmacol Toxicol 53:401–426
Maina M, Hauschild R, Sikora R (2008) Protection of tomato plants against fusaric acid by resistance induction. J Appl Biosci 1:18–31
Maksymiec W, Krupa Z (2006) The effects of short term exposition to Cd, excess Cu ions and jasmonate on oxidative stress appearing in Arabidopsis thaliana. Environ Exp Bot 57:187–194
Malerba M, Cerana R (2021) Plant cell cultures as a tool to study programmed cell death. Int J Mol Sci 22:2166
Mamur S, Ünal F, Yılmaz S, Erikel E, Yüzbaşıoğlu D (2020) Evaluation of the cytotoxic and genotoxic effects of mycotoxin fusaric acid. Drug Chem Toxicol 43:149–157
Manda RR, Srivastava VAAS (2021) Biochemistry of wilted tomato plants. Int Res J Chem 33:14–22
Mates J (2000) Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology 153:83–104
Mendoza-Vargas LA, Villamarín-Romero WP, Cotrino-Tierradentro AS, Ramírez-Gil JG, Chávez-Arias CC, Restrepo-Díaz H, Gomez-Caro S (2021) Physiological response of cape gooseberry plants to Fusarium oxysporum f. sp. physali, fusaric acid, and water deficit in a hydrophonic system. Front Plant Sci. https://doi.org/10.3389/fpls.2021.702842
Mézes M (2008) Mycotoxins and other contaminants in rabbit feeds. In: Proc 9th World Rabbit Congr, Verona, Italy, pp 491–506
Milićević DR, Škrinjar M, Baltić T (2010) Real and perceived risks for mycotoxin contamination in foods and feeds: challenges for food safety control. Toxins 2:572–592
Niehaus EM, von Bargen KW, Espino JJ, Pfannmuller A, Humpf HU, Tudzynski B (2014) Characterization of the fusaric acid gene cluster in Fusarium fujikuroi. Appl Microbiol Biotechnol 98:1749–1762
Nosir W, McDonald J, Woodward S (2011) Impact of biological control agents on fusaric acid secreted from Fusarium oxysporum f. sp. gladioli (Massey) Snyder and Hansen in Gladiolus grandiflorus corms. J Ind Microbiol Biotechnol 38:21–27
Overmyer K, Brosche M, Pellinen R, Kuittinen T, Tuominen H, Ahlfors R, Keinanen M, Saarma M, Scheel D, Kangasjarvi J (2005) Ozone-induced programmed cell death in the Arabidopsis radical-induced cell death 1 mutant. Plant Physiol 137:1092–1104
Palazzini J, Roncallo P, Cantoro R, Chiotta M, Yerkovich N, Palacios S, Echenique V, Torres A, Ramirez M, Karlovsky P, Chulze S (2018) Biocontrol of Fusarium graminearum sensu stricto, reduction of deoxynivalenol accumulation and phytohormone induction by two selected antagonists. Toxins 10:88
Pavlovkin J, Mistrik I, Prokop M (2004) Some aspects of the phytotoxic action of fusaric acid on primary Ricinus roots. Plant Soil Environ 50:397–401
Pinotti L, Ottoboni M, Giromini C, Dell’Orto V, Cheli F (2016) Mycotoxin contamination in the EU feed supply chain: a focus on cereal byproducts. Toxins 8:45
Pleadin J, Frece J, Markov K (2019) Mycotoxins in food and feed. Adv Food Nutr Res 89:297–345
Poór P, Kovács J, Szopkó D, Tari I (2013) Ethylene signaling in salt stress-and salicylic acid-induced programmed cell death in tomato suspension cells. Protoplasma 250:273–284
Poór P, Takács Z, Bela K, Czékus Z, Szalai G, Tari I (2017) Prolonged dark period modulates the oxidative burst and enzymatic antioxidant systems in the leaves of salicylic acid-treated tomato. J Plant Physiol 213:216–226
Ratajczak MZ, Ratajczak J (2016) Horizontal transfer of RNA and proteins between cells by extracellular microvesicles: 14 years later. Clin Transl Med 5:1487
Reddy RV, Larson CA, Brimer GE, Frappier BL, Reddy CS (1996) Developmental toxic effects of fusaric acid in CD1 mice. Bull Environ Contam Toxicol 57:354–360
Rodríguez-Carrasco Y, Ruiz MJ, Font G, Berrada H (2013) Exposure estimates to Fusarium mycotoxins through cereals intake. Chemosphere 93:2297–2303
Ruiz JA, Bernar EM, Jung K (2015) Production of siderophores increases resistance to fusaric acid in Pseudomonas protegens Pf-5. PLoS ONE 10:e0117040
Samadi L, Shahsavan Behboodi B (2006) Fusaric acid induces apoptosis in saffron root-tip cells: roles of caspase-like activity, cytochrome c, and H2O2. Planta 225:223–234
Sánchez-Rangel D, Hernández-Domínguez EE, Pérez-Torres CA, Ortiz-Castro R, Villafán E, Rodríguez-Haas B, Ibarra-Laclette E (2018) Environmental pH modulates transcriptomic responses in the fungus Fusarium sp. associated with KSHB Euwallacea sp. near fornicatus. BMC Genom 19:1–21
Sapko OA, Utarbaeva AS, Makulbek S (2011) Effect of fusaric acid on prooxidant and antioxidant properties of the potato cell suspension culture. Russ J Plant Physiol 58:828–835
Sharma SK, Singh D, Pandey H, Jatav RB, Singh V, Pandey D (2022) An overview of roles of enzymatic and nonenzymatic antioxidants in plant. In: Aftab T, Hakeem KR (eds) Antioxidant defence in plants. Springer, Singapore, pp 1–13
Sieber CMK, von Bargen KW, Studt L, Niehaus E-M, Espino JJ (2013) Deciphering the cryptic genome: genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites. PLoS Pathol 9:e1003475
Simonetti E, Roberts IN, Montecchia MS, Gutierrez-Boem FH, Gomez FM, Ruiz JA (2018) A novel Burkholderia ambifaria strain able to degrade the mycotoxin fusaric acid and to inhibit Fusarium spp. growth. Microbiol Res 206:50–59
Singh VK, Upadhyay RS (2014) Fusaric acid induced cell death and changes in oxidative metabolism of Solanum lycopersicum L. Bot Stud 55:1–11
Singh VK, Upadhyay R (2016) Induction of defence responses by fusaric acid (Fusarium toxin) in tomato plant. In: Proceedings of the 6th International Conference on Agriculture, Environment and Biological Sciences, Kuala Lumpur, Malaysia, pp 21–22
Singh VK, Singh HB, Upadhyay RS (2017) Role of fusaric acid in the development of ‘Fusarium wilt’symptoms in tomato: physiological, biochemical and proteomic perspectives. Plant Physiol Biochem 118:320–332
Sobral MMC, Faria MA, Cunha SC, Ferreira IM (2018) Toxicological interactions between mycotoxins from ubiquitous fungi: impact on hepatic and intestinal human epithelial cells. Chemosphere 202:538–548
Spss D, Oliveira RC (2009) Effects of fusaric acid on Zea mays L. seedlings. Phyton 78:155
Srivastava S, Singh VP, Rana M (2020) Fusaric acid: a potent vivotoxin. Eur J Mol Clin Med 7:2020
Stipanovic RD, Puckhaber LS, Liu J, Bell AA (2011) Phytotoxicity of fusaric acid and analogs to cotton. Toxicon 57:176–178
Studt L, Janevska S, Niehaus EM, Burkhardt I, Arndt B, Sieber CM, Humpf HU, Dickschat JS, Tudzynski B (2016) Two separate key enzymes and two pathway-specific transcription factors are involved in fusaric acid biosynthesis in Fusarium fujikuroi. Environ Microbiol 18(3):936–956
Valandro F, Menguer PK, Cabreira-Cagliari C, Margis-Pinheiro M, Cagliari A (2020) Programmed cell death (PCD) control in plants: new insights from the Arabidopsis thaliana deathosome. Plant Sci 299:110603
Vesonder RF, Gasdorf H, Peterson RE (1993) Comparison of the cytotoxicities of Fusarium metabolites and Alternaria metabolite AAL-toxin to cultured mammalian cell lines. Arch Environ Contam Toxicol 24:473–477
Wang M, Xiong Y, Ling N, Feng X, Zhong Z, Shen Q, Guo S (2013) Detection of the dynamic response of cucumber leaves to fusaric acid using thermal imaging. Plant Physiol Biochem 66:68–76
Wang M, Ling N, Dong X, Liu X, Shen Q, Guo S (2014) Effect of fusaric acid on the leaf physiology of cucumber seedlings. Eur J Plant Pathol 138:103–112
Wang M, Sun Y, Sun G, Liu X, Zhai L, Shen Q, Guo S (2015) Water balance altered in cucumber plants infected with Fusarium oxysporum f. sp. cucumerinum. Sci Rep 5:1–7
Wang R, Huang J, Liang A, Wang Y, Mur LAJ, Wang M, Guo S (2020) Zinc and copper enhance cucumber tolerance to fusaric acid by mediating its distribution and toxicity and modifying the antioxidant system. Int J Mol Sci 21:3370
Wang M, Liu W, Yan J, Sun P, Chen F, Jiang B, Xie D, Lin YE, Peng Q, He X (2021) A transcriptomic analysis of gene expression in chieh-qua in response to fusaric acid stress. Horticulturae 7:88
Wu HS, Yin XM, Zhu YY, Guo SW, Wu CL, Lu YL, Shen QR (2007) Nitrogen metabolism disorder in watermelon leaf caused by fusaric acid. Physiol Mol Plant Pathol 71:69–77
Wu HS, Bao W, Liu DY, Ling N, Ying RR, Raza W, Shen QR (2008a) Effect of fusaric acid on biomass and photosynthesis of watermelon seedlings leaves. Caryologia 61:258–268
Wu HS, Yin XM, Liu DY, Ling N, Bao W, Ying RR, Zhu YY, Guo SW, Shen QR (2008b) Effect of fungal fusaric acid on the root and leaf physiology of watermelon (Citrullus lanatus) seedlings. Plant Soil 308:255–266
Xie D, Ma L, Šamaj J, Xu C (2011) Immunohistochemical analysis of cell wall hydroxyproline-rich glycoproteins in the roots of resistant and susceptible wax gourd cultivars in response to Fusarium oxysporum f. sp. Benincasae infection and fusaric acid treatment. Plant Cell Rep 30:1555–1569
Xing F, Li Z, Sun A, Xing D (2013) Reactive oxygen species promote chloroplast dysfunction and salicylic acid accumulation in fumonisin B1-induced cell death. FEBS Lett 587:2164–2172
Yin ES, Rakhmankulova M, Kucera K, de Sena Filho JG, Portero CE, Narváez-Trujillo A, Holley SA, Strobel SA (2015) Fusaric acid induces a notochord malformation in zebrafish via copper chelation. Biometals 28:783–789
Zain ME (2011) Impact of mycotoxins on humans and animals. J Saudi Chem Soc 15:129–144
Zaynab M, Fatima M, Sharif Y, Zafar MH, Ali H, Khan KA (2019) Role of primary metabolites in plant defence against pathogens. Microb Pathog 137:103728
Zhang X, Dai YS, Wang YX, Su ZZ, Yu LJ, Zhang ZF, Chen QF (2022) Overexpression of the Arabidopsis MACPF protein AtMACP2 promotes pathogen resistance by activating SA signaling. Int J Mol Sci 23:8784
Funding
Open access funding provided by University of Szeged. This work was supported by the grant from the National Research, Development, and Innovation Office of Hungary – NKFIH (Grant no. NKFIH FK 124871 and 138867) and by the ÚNKP-22–4 and ÚNKP-22–5 New National Excellence Program of the Ministry of Human Capacities and the University of Szeged Open Access Fund (6429). Péter Poór and Attila Ördög were supported by the János Bolyai Research Scholarship of the Hungarian Academy of Sciences.
Author information
Authors and Affiliations
Contributions
Writing—original draft preparation, NI; writing—review and editing, NI, ZC, AÖ and PP.
Corresponding author
Ethics declarations
Conflict of interest
No conflict of interest is declared.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Iqbal, N., Czékus, Z., Ördög, A. et al. Fusaric acid-evoked oxidative stress affects plant defence system by inducing biochemical changes at subcellular level. Plant Cell Rep 43, 2 (2024). https://doi.org/10.1007/s00299-023-03084-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00299-023-03084-9