
Abstract
The solvent-tolerant bacterium Pseudomonas putida S12 was engineered to efficiently utilize the C1 compounds methanol and formaldehyde as auxiliary substrate. The hps and phi genes of Bacillus brevis, encoding two key steps of the ribulose monophosphate (RuMP) pathway, were introduced to construct a pathway for the metabolism of the toxic methanol oxidation intermediate formaldehyde. This approach resulted in a remarkably increased biomass yield on the primary substrate glucose when cultured in C-limited chemostats fed with a mixture of glucose and formaldehyde. With increasing relative formaldehyde feed concentrations, the biomass yield increased from 35% (C-mol biomass/C-mol glucose) without formaldehyde to 91% at 60% relative formaldehyde concentration. The RuMP-pathway expressing strain was also capable of growing to higher relative formaldehyde concentrations than the control strain. The presence of an endogenous methanol oxidizing enzyme activity in P. putida S12 allowed the replacement of formaldehyde with the less toxic methanol, resulting in an 84% (C-mol/C-mol) biomass yield. Thus, by introducing two enzymes of the RuMP pathway, co-utilization of the cheap and renewable substrate methanol was achieved, making an important contribution to the efficient use of P. putida S12 as a bioconversion platform host.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The solvent-tolerant Pseudomonas putida S12 is used as a platform for the bioconversion of sugars into substituted aromatic compounds (Nijkamp et al. 2005; Nijkamp et al. 2007; Verhoef et al. 2007; Wierckx et al. 2005). These compounds are produced via central metabolite intermediates such as l-tyrosine and l-phenylalanine, the formation of which is directly linked to cellular growth. In such a process, a significant proportion of the available substrate is not used to form biomass and product, but utilized for the generation of free energy (ATP and proton gradient) and reducing equivalents (NAD(P)H). Since the sugar substrate represents an important cost factor in bioproduction processes (Schmid et al. 2001), the addition of a cheap auxiliary catabolic substrate to generate free energy and/or reducing equivalents may significantly improve the economy of the production process.
Previous studies have shown that co-utilization of thiosulfate or C1 compounds like formate and formaldehyde leads to an increased yield on the primary carbon source (Baerends et al. 2008; Bruinenberg et al. 1985; Harris et al. 2007; Masau et al. 2001). Another C1 compound that can be used as auxiliary substrate is methanol. Being more reduced than formate or formaldehyde, methanol can yield more reducing equivalents per C-mole. Since methanol can be derived from biomass via synthesis gas (Chmielniak and Sciazko 2003) it is a promising renewable auxiliary substrate for biotechnological processes.
The first step in methanol metabolism is the oxidation to formaldehyde, via dehydrogenases or oxidases (Anthony 1986; Sahm 1977). Formaldehyde is extremely toxic due to non-specific reactivity with proteins and nucleic acids. Therefore, rapid and efficient formaldehyde metabolization is a crucial step in the utilization of methanol. Several pathways for the metabolism of formaldehyde have been described. Methylotrophic yeasts may assimilate formaldehyde by the xylulose-5-phosphate cycle. In this pathway, formaldehyde is coupled to xylulose-5-phosphate and converted into dihydroxyacetone and glyceraldehyde-3-phosphate (GA3P) by a specialized transketolase-dihydroxyacetone synthase (Yurimoto et al. 2005a, b). Alternatively, formaldehyde is oxidatively dissimilated to form formate and eventually carbon dioxide and water (Yurimoto et al. 2005a, b).
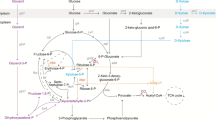
In bacteria, three formaldehyde metabolic pathways are known. Like yeasts, also bacteria may oxidatively dissimilate formaldehyde to formate and CO2. Alternatively, formaldehyde may be assimilated via the serine cycle. In this pathway formaldehyde is coupled to l-glycine to form l-serine (l-ser). l-ser is subsequently metabolized via a cyclic pathway that ensures replenishment of l-glycine and permits a drain on glycerate-3-phosphate to generate biomass (Chistoserdova et al. 2003). The third bacterial formaldehyde metabolic route is the RuMP pathway (Fig. 1) (Kato et al. 2006). In this pathway, formaldehyde is coupled to ribulose-5-phosphate (Ru5P) forming hexulose-6-phosphate (Hu6P). Hu6P is isomerized to fructose-6-phosphate (F6P), that can be further metabolized via the Embden–Meyerhof–Parnas (EMP) pathway, the Entner–Doudoroff (ED) pathway, or the pentose phosphate pathway (PPP). When F6P enters the oxidative part of the PPP (after isomerization to glucose-6-phosphate (G6P)), the RuMP pathway constitutes a cyclic oxidation pathway for formaldehyde: G6P yields Ru5P and CO2 while generating two NADPH. This cyclic oxidative dissimilation of formaldehyde has been described in, a.o., Methylobacillus flagellatus KT (Chistoserdova et al. 2000). The RuMP pathway may also constitute a formaldehyde assimilation pathway (Fig. 1). In this case one-third of the F6P formed enters either the EMP or ED pathway and is converted into GA3P and pyruvate. Pyruvate is used for the production of cell constituents whereas GA3P and two-thirds of the produced F6P are used to regenerate Ru5P by a combination of transketolase, transaldolase, and isomerization reactions (Jakobsen et al. 2006; Kato et al. 2006; Large and Bamforth 1988).

Assimilation and dissimilation pathways for formaldehyde in P. putida S12pJNNhp(t). 1 Hexulose phosphate synthase, 2 hexulose phosphate isomerase, 3 hexose phosphate isomerase, 4 glucose-6-phosphate dehydrogenase, 5 6-phosphogluconate dehydratase, 6 2-dehydro-3-deoxy-phosphogluconate aldolase, 7 transketolase, 8 transaldolase, 9 pentose phosphate isomerase, 10 pentose phosphate epimerase; 11 6-phosphogluconate dehydrogenase, 12 formaldehyde dehydrogenase, 13 formate dehydrogenase. Ru ribulose, Hu hexulose, F fructose, G glucose, Xu xylulose, R ribulose, E erythrose, Su sedoheptulose, KDPG 2-dehydro-3-deoxy-6-phospho-d-gluconate, HCHO formaldehyde. Black arrows indicate the assimilatory RuMP pathway, dashed arrows indicate the dissimilatory RuMP pathway, gray arrows indicate the linear oxidation of formaldehyde to carbon dioxide
Previously, formaldehyde has been successfully applied as auxiliary substrate in yeast; however, its high toxicity hindered efficient co-utilization (Baerends et al. 2008). Expression of formaldehyde dehydrogenase (Fld) and a formate dehydrogenase (Fmd) from the methylotrophic yeast Hansenula polymorpha in Saccharomyces cerevisiae resulted in an increased biomass yield with formaldehyde as auxiliary substrate (Baerends et al. 2008). An increased tolerance towards formaldehyde was obtained by introducing the fld and fmd genes.
The aim of the present study was to develop and optimize a P. putida S12 strain capable of efficiently utilizing formaldehyde as auxiliary substrate. Since P. putida S12 possesses genes encoding formaldehyde and formate dehydrogenase, it was expected that P. putida S12 has a basic endogenous capacity to oxidize formaldehyde to formate and CO2. To further improve this capacity, additional formaldehyde metabolic pathways were constructed by expressing the two key enzymes of the RuMP pathway, 3-hexulose-6-phosphate synthase (Hps) and 6-phospho-3-hexulose isomerase (Phi). Biomass yields were determined in chemostat cultures for the engineered strain on different mixtures of glucose and formaldehyde. The RuMP pathway strain showed significantly improved performance over the control strain, achieving a biomass yield-on-glucose of 91% when using formaldehyde as auxiliary substrate. Replacing formaldehyde with the renewable auxiliary substrate methanol resulted in a biomass yield-on-glucose of 84%.
Materials and methods
Bacterial strains and culture conditions
The bacterial strains and plasmids used in this study are shown in Table 1. Luria broth (LB) (Sambrook and Russel 2001) or phosphate buffered mineral salts medium (MM; Hartmans et al. 1989) were used as indicated. MM was supplemented with 20 mM glucose (MMG) unless otherwise stated. Antibiotics were added as required at the following concentrations: ampicillin, 100 µg/ml; gentamicin, 10 µg/ml (MM), and 25 µg/ml (LB). Heterologous gene expression was induced by addition of 0.1 mM of salicylate unless otherwise stated.
Carbon-limited chemostat cultures were performed either in 1.0 l (working volume) fermentor using a BioFloIIc controller (New Brunswick Scientific) or in 0.7 l (working volume) fermentor using a BioFlo110 controller. MM was used with 10 mM glucose, 10 mg/l gentamicin and 0.1 mM sodium salicylate. Chemostats were inoculated with 50-ml overnight cultures on MMG. The dilution rate (D) was set at 0.05 h−1. After 36 h, the D was increased to 0.2 h−1 (glucose/formaldehyde chemostats) or 0.1 h−1 (glucose/methanol chemostats). The temperature was maintained at 30°C and the pH was maintained at 7.0 by automatic addition of 2 N NaOH. The stirring speed was controlled by dissolved oxygen (DO) concentration set at 15%. Either air or a mixture of air and oxygen was supplied, using a M + W Instruments D-5111 mass-flow controller (glucose/methanol chemostat), respectively, Brooks mass-flow controllers (5850 E series and 5850 TR series) and a 0154 control unit (Brooks). Dissolved oxygen tension was continuously monitored with an InPro model 6810 probe.
For preparation of formaldehyde solutions, demineralised water was heated to 100°C. During cooling paraformaldehyde (Sigma-Aldrich) was added to obtain a 6 M stock solution. A 5 M NaOH solution was added to until the paraformaldehyde was completely dissolved. Before use, the pH of the formaldehyde solution was set to pH 7 with a 5 M HCl stock solution. The final formaldehyde concentration was measured by HPLC.
Construction of recombinant plasmids
The hps-phi genes were amplified from genomic DNA of B. brevis S1 (ATCC 12524) using forward (5′-GCCTCGAGGGAATACACACATTTGCTTGTAC-3′) and reverse (5′-CGTCTAGACTATCGAGATTGGCATGTC-3′) primers, designed on the published sequence of hps and phi (Takashita and Yasueda 2004). The original bicistronic gene organization was maintained as well as the native ribosome binding sites. Standard amplification methods were used with Accuprime Pfx polymerase (Invitrogen). The resulting PCR fragment was A-tailed for 30 min at 68°C using Supertaq (SphaeroQ). The fragment was subcloned in pGEM®-T Easy (Promega) yielding pGEM-HP. After digestion with NotI, the hps-phi fragment was isolated from agarose gel and ligated into NotI digested and dephosphorylated pJNN(t), yielding pJNNhp(t) with the hps-phi bicistron under the control of the nagAa promoter. After verifying the correct orientation by restriction analysis, pJNNhp(t) was transformed to P. putida S12 using a Gene Pulser electroporation device, yielding P. putida S12pJNNhp(t).
Plasmid DNA was isolated using QIAprep spin miniprep kit (QIAGEN). DNA fragments were isolated from 0.8% agarose gels with QIAEXII gel extraction kit (QIAGEN). Nucleotide sequencing reactions were performed by MWG Biotech AG. Standard molecular cloning techniques were performed according to Sambrook and Russell (2001).
Analytical methods
Cell densities were measured at 600 nm (OD600) with a Biowave Cell Density Meter (WPA Ltd). Cell dry weight (CDW) was calculated from OD600 values where 1 OD600 unit corresponds to 0.465 g/l CDW. For calculation of C-mol biomass, CDW was divided by 24 (Roels 1983).
Glucose concentrations were analyzed by HPLC (Waters) using an Aminex HDP-87 N column with 0.01 M Na2HPO4 as the eluent and a refractive index detector, or using an ion chromatography (Dionex ICS3000 system). For HPLC-RI, an Aminex HPX-87 N (Bio Rad) column (300 × 7.8 mm) was applied using 0.01 M Na2HPO4 as the eluent. For ion chromatography a CarboPac PA20 column was used with 10 mM NaOH as the eluent.
Organic acids were analyzed by HPLC-UV (Agilent 1100 system) or ion chromatography (Dionex ICS3000). For HPLC-UV, an Aminex HDP-87H (Bio Rad) column (300 × 7.8 mm) was used with 0.008 N H2SO4 as the eluent at a flow rate of 0.6 ml/min. The diode-array detector was set at 210 nm. For ion chromatography an IonPac ICE AS6 column (9 × 250 mm) was used with 0.4 mM heptafluorobutyric acid as the eluent.
Methanol, formate and formaldehyde were analyzed by ion chromatography (Dionex ICS3000) using an IonPac ICE AS1 column (9 × 250 mm) with 100 mM methyl sulphonic acid as the eluent. In addition, formaldehyde was analyzed by HPLC-UV (Agilent 1100) after derivatization with 2,4-dinitrophenylhydrazine (DNPH). Equal volumes of DNPH solution (0.5 mg/ml in 3% (w/v) phosphoric acid) and the sample were mixed and incubated for 10 min at 30°C prior to HPLC analysis. An Eclipse XDB-C8 column (4.6 × 15 mm) was used with a mixture of 45% acetonitrile and 55% water as the eluent at a flow rate of 1.2 mL/min. The diode-array detector set at 360 nm.
Protein expression was analyzed by SDS-PAGE. Samples were obtained by centrifuging 1 ml of culture at an OD600 of 1.5 (mid-exponential growth phase) at 16.000×g. The cell pellet was resuspended in 300 μl of loading buffer according to Sambrook and Russell (2001) and heated at 100°C for 10 min. After centrifugation at 16,000×g 20 μl of supernatant was applied on the gel for analysis. Routinely, 15% precast Tris–HCl gels (Bio Rad) placed in a Criterion™ electrophoresis cell (Bio Rad) were used. Electrophoresis was performed in running buffer containing 0.025 M Tris (pH 8.8), 0.192 M, glycine, and 3.5 mM SDS in demineralised water as described by Sambrook and Russell (2001) at 130 V. A broad-range unstained protein standard (Bio Rad) was used as a marker. Gels were stained using Coomassie Brilliant Blue R-250 staining solution (Bio Rad).
Enzyme assays
For preparation of cell extracts, 50-ml cultures were harvested at late logarithmic growth phase by centrifugation at 3,200×g for 15 min at 4°C. Cell pellets were resuspended in 3 ml of 50 mM potassium phosphate buffer (pH 7.5). All samples were kept on ice during the preparation of extracts. Cells were disrupted using a Branson sonifier with a micro tip at a pulse mode, output set at 4 and effective output set to 60%. After three rounds of sonication (45 s of pulsing and 15 s pause), cell debris was removed by centrifugation at 3,200×g for 15 min at 4°C. Supernatants were desalted using a PD10 column (GE healthcare) and used for enzyme assays. Protein concentration was measured using Bradford reagent (Sigma-Aldrich).
Hps- and Phi enzyme activities were determined in a single combined assay as described previously (Arfman et al. 1990; Kato 1990; Orita et al. 2006). The final product formed by these two enzymes from formaldehyde and Ru5P is F6P that is converted into G6P by glucose-6-phosphate isomerase. The formation of G6P was coupled to NADPH formation via glucose-6-phosphate dehydrogenase that converts G6P into glucono-1,5-lactone-6-phosphate. The reaction mixture (0.95 ml) consisted of 50 mM potassium phosphate (pH 7.5), 5 mM MgCl2, 5 mM ribose-5-phosphate (Ri5P; Sigma-Aldrich), 2.5 mM NADP (Sigma-Aldrich), 10 U phosphoribose isomerase (PRI; Sigma-Aldrich), 10 U phosphoglucose isomerase (Roche), 10 U glucose-6-phosphate dehydrogenase (Sigma-Aldrich). The mixture was preincubated for 5 min at 30°C to achieve equilibrium between Ri5P and Ru5P (catalyzed by PRI). The reaction was started by adding formaldehyde to a concentration of 5 mM. For calculation of the combined Hps- and Phi activity, a molar extinction coefficient of 6.22 mM−1 cm−1 at 340 nm was used for NADPH. One unit is defined as the overall Hps and Phi activity catalyzing the formation of 1 μmol/min of NADPH at 30°C.
Results
Cloning and functional expression of Hps and Phi in P. putida S12
The thermotolerant methylotrophic bacterium B. brevis S1 was used as the source of the hps and phi genes encoding the first two enzymes of the RuMP pathway. Previously, these enzymes have been successfully expressed in the non-methylotrophic mesophile Escherichia coli (Yurimoto et al. 2002). Therefore, a similar approach was followed for P. putida S12. Heterologous protein expression in the constructed RuMP pathway strain, P. putida S12pJNNhp(t), was confirmed by SDS-PAGE (not shown). As the individual activities are difficult to assess, the functionality of the expressed enzymes was confirmed by measuring their combined activity according to Arfman et al. (1990). The overall activity of the two conversions performed by these enzymes was 6,800 U/g protein in cell extract of mid-log phase cells from P. putida S12pJNNhp(t). As expected, no activity was observed in the empty vector control strain (P. putida S12pJNNmcs(t)).
Formaldehyde as auxiliary substrate: C-limited chemostat cultures at a dilution rate of 0.2 h−1
P. putida S12pJNNhp(t) and an empty vector control strain were cultured in a C-limited chemostat with glucose as the primary carbon source and formaldehyde as auxiliary substrate. The chemostat feed was supplied at a dilution rate of 0.2 h−1 and contained a fixed concentration of carbon (60 mM) consisting of a mixture of glucose and formaldehyde the ratio of which was altered during the experiment. The experiment was started with 10 mM glucose (60 mM carbon) and no formaldehyde. After reaching steady state (i.e., after approximately 25 h) 6 mM of glucose-carbon (i.e., 1 mM of glucose) was replaced with 6 mM of formaldehyde-carbon (i.e., 6 mM of formaldehyde). More glucose was replaced with formaldehyde in this step-wise fashion until wash-out of the chemostat cultures was observed.
Both the Hps/Phi-expressing P. putida S12pJNNhp(t) and the empty vector control strain reached steady state when up to 50% of the total carbon feed consisted of formaldehyde. No accumulation of glucose or its corresponding acid metabolites gluconate and 2-ketogluconate was observed, confirming that the cultures were C-limited. Also, no formaldehyde or formate accumulated in the cultures of either strain, indicating the presence of endogenous formaldehyde and formate dehydrogenases in P. putida S12 by which these compounds were metabolized to completion.
Figure 2a shows the biomass yield-per-glucose (C-mol biomass/C-mol glucose) for both the empty vector control and the RuMP pathway strain. Both strains showed a yield increase with increasing formaldehyde levels (Fig. 2b). However, the performance of the RuMP pathway strain was consistently superior to the performance of the control strain. The yield increase of the RuMP pathway strain relative to the yield increase of the control strain (Fig. 2c) consistently shows that the introduction of the RuMP pathway genes approximately doubled the endogenous yield increase. This effect is independent from the relative formaldehyde concentration within the range tested.

a Biomass yield on glucose in chemostat cultures of strains S12pJNNhp(t) (white bars) and S12pJNN(t) (gray bars). Y xs represents the biomass yield (C-mol biomass per C-mol glucose). b Biomass yield increase (on glucose) as function of relative formaldehyde concentration in chemostat cultures of strains S12pJNNhp(t) (triangles) and S12pJNN(t) (squares). The biomass yield on glucose at 0% relative formaldehyde concentration was considered as the baseline yield. The baseline yield was assumed to be independent on relative formaldehyde concentration. c Yield increase factor of strain S12pJNNhp(t) over strain S12pJNN(t) as function of relative formaldehyde concentration in chemostat cultures. Data are averages of duplicate experiments; error bars denote the maximum deviation from the average of two independent experiments
Formaldehyde as auxiliary substrate: C-limited chemostat cultures at low dilution rate
Both the empty vector control strain and the RuMP pathway strain washed out in chemostats at D = 0.2 h−1 when the fraction of formaldehyde was increased to 60% of total carbon. Wash-out of chemostat cultures coincided with accumulation of formaldehyde and formic acid. This result suggests that both the endogenous and the engineered formaldehyde metabolic pathway activities were too low to cope with the formaldehyde feed applied. To investigate this hypothesis, chemostat cultures were performed at a lower dilution rate and, consequently, a lower formaldehyde feed rate that is more in agreement with previous studies on continuous culture experiments with methylotrophic bacteria on formaldehyde (Arfman et al. 1992; De Boer et al. 1990; Mitsui et al. 2005).
The experiment was started at a dilution rate of 0.2 h−1 and a 7:3 (C-mol) glucose-to-formaldehyde ratio (60 mM total C). When steady state was reached, the dilution rate was set to 0.03 h−1. The formaldehyde feed concentration was kept constant whereas the glucose feed concentration was lowered to 2 mM (12 mM C) to achieve a 4:6 glucose-to-formaldehyde ratio (C-mol; 30 mM C total).
Figure 3 shows that the empty vector control strain did not reach steady state under these conditions. Formaldehyde and formate accumulated and wash-out was observed. In contrast, P. putida S12pJNNhp(t) reached steady state with a biomass yield on glucose of 91% (C-mol biomass/C-mol glucose). Since the RuMP pathway strain should theoretically be able to utilize formaldehyde as a sole C-source (see Fig. 1), the glucose feed was stopped. However, formaldehyde and formic acid accumulated and subsequent wash-out of biomass occurred, indicating that the assimilatory pathway activity for formaldehyde was insufficient to sustain growth at the conditions tested.

C-limited chemostat cultures of P. putida S12pJNNhp(t) (triangles) and P. putida S12pJNN(t) (squares) at D = 0.03 h−1, grown on mineral salts medium with 30 mM of total carbon (60% of all carbon originates from formaldehyde). The data presented are from a single representative experiment
Methanol as auxiliary substrate: C-limited chemostat cultures at low dilution rate
In the chemostat cultures performed in this study, formaldehyde (and formate) accumulated above a critical formaldehyde feed concentration, probably reflecting the upper limit of the formaldehyde metabolic capacity of the culture. Accumulation of only trace amounts of formaldehyde was always accompanied by wash-out of the culture, underlining the acute and low-threshold nature of formaldehyde toxicity. Thus, for the purpose of better process control, formaldehyde should be replaced with a less toxic auxiliary substrate such as methanol. Utilization of methanol as (co-)substrate, however, requires the ability to oxidize methanol to formaldehyde.
Unexpectedly, formation of NADPH was observed in Hps/Phi enzyme assays with P. putida S12 cell extracts when methanol was added as a substrate instead of formaldehyde. The overall activity amounted to 145 U/g of protein in cell extract. Although the activity with methanol was lower by a factor 50 compared to formaldehyde, this result indicated that methanol is oxidized endogenously by P. putida S12.
The C-limited chemostat cultures at D = 0.03 h−1 were repeated, replacing formaldehyde with methanol. The chemostats were started at D = 0.1 h−1 and a mixed feed of glucose and methanol was applied according to the following regime between the different steady states: glucose/methanol: 10 mM/15 mM; 5 mM/15 mM, and 5/30 mM. After the final steady state, the dilution rate was set at 0.03 h−1 and the total feed carbon concentration was decreased to 30 mM with 60% originating from methanol.
P. putida S12pJNNhp(t) reached steady state under these conditions, in contrast to the empty vector control strain (Fig. 4). Approximately 25% of the methanol feed was not metabolized, but no formaldehyde or formate accumulation was observed. The co-utilization of methanol resulted in a significant improvement of the biomass yield to 84% biomass-per-glucose (C-mol). After stopping the glucose feed, the culture washed out as observed in the glucose/formaldehyde experiments while culture methanol levels increased.

C-limited chemostat cultures of P. putida S12pJNNhp(t) (triangles) and P. putida S12pJNN(t) (squares) at D = 0.03 h−1, grown on mineral salts medium with 30 mM of total carbon (60% of all carbon originates from methanol). The data presented are from a single representative experiment
Discussion
P. putida S12 was shown to have an innate capability to co-utilize formaldehyde and glucose, in relative concentrations of up to 50% in C-limited chemostats. This ability is indicative of the presence of an endogenous formaldehyde metabolic pathway in P. putida S12. Database searches revealed the presence of several genes coding for formaldehyde and formate dehydrogenases in the P. putida S12 genome (manuscript in preparation). Therefore, this pathway probably consists of a linear route that oxidizes formaldehyde, via formate, to CO2. The observed yield increase of the control strain may be attributed to the reducing equivalents generated during formaldehyde oxidation.
Although P. putida S12 disposes of an endogenous formaldehyde detoxification pathway, the introduction of an additional, heterologous formaldehyde metabolic pathway clearly had a positive effect in chemostat cultures grown on a mixture of glucose and formaldehyde. This was reflected by a consistently improved biomass yield on glucose and the ability to grow at elevated relative concentrations of formaldehyde.
The presence of an additional formaldehyde metabolic route may allow for growth at higher relative formaldehyde concentrations, since accumulation of the highly toxic formaldehyde is prevented more efficiently. This unlikely explains, however, the higher biomass yield gain for the strain expressing the RuMP pathway enzymes. Possibly, the improved yield gain relates to a better cofactor balance: when constituting a cyclic oxidation pathway, the RuMP cycle generates 2 mol of NADPH per mol formaldehyde (Fig. 1), whereas the linear oxidation pathway yields 2 mol of NADH. It should be noted, however, that the introduction of the hps and phi genes will also constitute an assimilatory RuMP pathway (Fig. 1). Thus, (co-)assimilation of formaldehyde may occur, in which case formaldehyde not only serves as a catabolic auxiliary substrate, but also as an assimilatory substrate. It was shown that the presence of the RuMP pathway results in a yield increase that is twice the yield increase attained by linear formaldehyde oxidation. Considering that the same amount of formaldehyde was metabolized in both strains, it may be concluded that the RuMP pathway strain utilizes formaldehyde twice as efficient as the empty vector control strain. Since the linear and the cyclic oxidation pathway yield an equal amount of reducing equivalents (Fig. 1; although of different nature), co-assimilation of formaldehyde is the most likely explanation for the observed additional yield increase found with the RuMP pathway strain. Thus, the biomass yield expressed as yield-per-glucose likely is an overestimation of the actual biomass yield-per-assimilated carbon for the RuMP-pathway strain grown on formaldehyde-glucose mixtures.
The exact relative contributions of linear oxidation, cyclic oxidation, cofactor balance, and assimilation of formaldehyde to biomass yield cannot be determined as it is not possible to selectively shut down either of the pathways. Attempts to knock-out formaldehyde dehydrogenase-encoding genes in order to abolish linear oxidation in P. putida S12 have consistently failed (unpublished). No viable double cross-over recombinants could be obtained, suggesting that the linear oxidation pathway apparently is crucial for the detoxification of formaldehyde. Also in non-methylotrophs, formaldehyde is an endogenous metabolite that is produced by oxidative demethylation of DNA (Aas et al. 2003; Falnes et al. 2002) and during methionine, histidine, and choline metabolism (Arnstein 1954). Moreover, the contributions of the cyclic oxidation pathway and the assimilatory pathway cannot be separated since the expression of Hps and Phi constitutes both pathways at a time.
Despite the likely ability of P. putida S12pJNNhp to utilize formaldehyde as sole C-source, chemostat cultures washed out when deprived of glucose while maintaining the formaldehyde feed. The activity of the first two enzymes of the RuMP pathway as measured in mid-log phase batch cultures should suffice to assimilate the formaldehyde administered to the low-D (0.03 h−1) cultures. The net Hps/Phi activity amounted to approximately 6,800 U/g protein. Assuming a protein content of 50%, this activity should allow 1 g of cells (dry weight) to cope with a formaldehyde feed of about 200 mmol h−1. However, in the low-D cultures 270 mg of CDW was not able to fully metabolize formaldehyde that was administered at a rate of 0.54 mmol h−1. This observation suggests that the metabolic flux through the endogenous part of the formaldehyde assimilatory cycle may be the bottleneck in preventing formaldehyde accumulation, and subsequent wash-out of cells, when deprived of other C-sources. Alternatively, the activity of the RuMP-pathway enzymes in batch cultures may not be representative for chemostat cultures.
Replacement of formaldehyde with methanol in C-limited chemostat cultures at a low dilution rate also resulted in improved biomass yield. Methanol was not completely utilized, however, and the biomass yield on glucose was slightly lower than for the corresponding formaldehyde experiments. The accumulation of methanol but not formaldehyde/formate suggests that the endogenous methanol dehydrogenase activity is the bottleneck for methanol utilization. This was also indicated by the 50-fold lower activity with methanol as the substrate in the Hps/Phi enzyme assay. From the P. putida S12 genome sequence (manuscript in preparation) no close homologues to established methanol dehydrogenases could be identified. Therefore, it is likely that the low methanol dehydrogenase activity of P. putida S12 results from a side activity of a broad-specificity alcohol dehydrogenase.
In conclusion, the enhanced ability of P. putida S12pJNNhp(t) to utilize C1 compounds as auxiliary substrate can be used to substantially improve raw feedstock utilization efficiency. Since both primary and auxiliary substrates can be obtained from biomass, process economy can be improved without compromising sustainability. Although somewhat less efficient than formaldehyde, methanol is the preferred auxiliary substrate as it is less toxic, easier to handle, cheap and renewable. Currently, possibilities are explored to improve the utilization of methanol by increasing the endogenous methanol dehydrogenase activity or by co-expressing a heterologous methanol dehydrogenase.
References
Aas PA, Otterlei M, Falnes PO, Vagbo CB, Skorpen F, Akbari M, Sundheim O, Bjoras M, Slupphaug G, Seeberg E, Krokan HE (2003) Human and bacterial oxidative demethylases repair alkylation damage in both RNA and DNA. Nature 421:859–863
Anthony C (1986) Bacterial oxidation of methane and methanol. Adv Microb Physiol 27:113–210
Arfman N, Bystrykh L, Govorukhina NI, Dijkhuizen L (1990) 3-Hexulose-6-phosphate synthase from thermotolerant methylotroph Bacillus C1. Methods Enzymol 188:391–397
Arfman N, de Vries KJ, Moezelaar HR, Attwood MM, Robinson GK, van Geel M, Dijkhuizen L (1992) Environmental regulation of alcohol metabolism in thermotolerant methylotrophic Bacillus strains. Arch Microbiol 157:272–278
Arnstein HRV (1954) The metabolism of glycine. In: Anson ML, Baily K, Edsall JT (eds) Advances in protein chemistry, vol IX. Academic, New York, pp 2–82
Baerends RJ, de Hulster E, Geertman JM, Daran JM, van Maris AJ, Veenhuis M, van der Klei IJ, Pronk JT (2008) Engineering and analysis of a Saccharomyces cerevisiae strain that uses formaldehyde as an auxiliary substrate. Appl Environ Microbiol 74:3182–3188
Bruinenberg PM, Jonker R, Dijken JP, Scheffers WA (1985) Utilization of formate as an additional energy source by glucose-limited cchemostate cultures of Candida utilis CBS 621 and Saccheromyces cerevisiae CBS 8066. Arch Microbiol 142:302–306
Chistoserdova L, Chen SW, Lapidus A, Lidstrom ME (2003) Methylotrophy in Methylobacterium extorquens AM1 from a genomic point of view. J Bacteriol 185:2980–2987
Chistoserdova L, Gomelsky L, Vorholt JA, Gomelsky M, Tsygankov YD, Lidstrom ME (2000) Analysis of two formaldehyde oxidation pathways in Methylobacillus flagellatus KT, a ribulose monophosphate cycle methylotroph. Microbiology 146:233–238
Chmielniak T, Sciazko M (2003) Co-gasification of biomass and coal for methanol synthesis. Applied energy 74:393–403
De Boer LE, van der Vlag GJJ, Dijkhuizen L (1990) Regulation of methanol metabolism in the facultative methylothroph Nocardia sp. 239 during growth on mixed substrates in batch- and continuous cultures. Arch Microbiol 153:337–343
Falnes PO, Johansen RF, Seeberg E (2002) AlkB-mediated oxidative demethylation reverses DNA damage in Escherichia coli. Nature 419:178–182
Harris DM, van der Krogt ZA, van Gulik WM, van Dijken JP, Pronk JT (2007) Formate as an auxiliary substrate for glucose-limited cultivation of Penicillium chrysogenum: impact on penicillin G production and biomass yield. Appl Environ Microbiol 73:5020–5025
Hartmans S, Smits JP, van der Werf MJ, Volkering F, de Bont JAM (1989) Metabolism of styrene oxide and 2-phenylethanol in the styrene-degrading Xanthobacter Strain 124X. Appl Environ Microbiol 55:2850–2855
Hartmans S, van der Werf MJ, de Bont JA (1990) Bacterial degradation of styrene involving a novel flavin adenine dinucleotide-dependent styrene monooxygenase. Appl Environ Microbiol 56:1347–1351
Husken LE, Beeftink R, de Bont JA, Wery J (2001) High-rate 3-methylcatechol production in Pseudomonas putida strains by means of a novel expression system. Appl Microbiol Biotechnol 55:571–577
Jakobsen OM, Benichou A, Flickinger MC, Valla S, Ellingsen TE, Brautaset T (2006) Upregulated transcription of plasmid and chromosomal ribulose monophosphate pathway genes is critical for methanol assimilation rate and methanol tolerance in the methylotrophic bacterium Bacillus methanolicus. J Bacteriol 188:3063–3072
Kato N (1990) 3-Hexulose-6-phosphate synthase from Mycobacterium gastri MB19. Methods Enzymol 188:397–401
Kato N, Yurimoto H, Thauer RK (2006) The physiological role of the ribulose monophosphate pathway in bacteria and archaea. Biosci Biotechnol Biochem 70:10–21
Large J, Bamforth CW (1988) Methylotrophy and biotechnology. Longman, London
Masau RJ, Oh JK, Suzuki I (2001) Mechanism of oxidation of inorganic sulfur compounds by thiosulfate-grown Thiobacillus thiooxidans. Can J Microbiol 47:348–358
Mitsui R, Omori M, Kitazawa H, Tanaka M (2005) Formaldehyde-limited cultivation of a newly isolated methylotrophic bacterium, Methylobacterium sp. MF1: enzymatic analysis related to C1 metabolism. J Biosci Bioeng 99:18–22
Nijkamp K, van Luijk N, de Bont JAM, Wery J (2005) The solvent-tolerant Pseudomonas putida S12 as host for the production of cinnamic acid from glucose. Appl Microbiol Biotechnol 69:170–177
Nijkamp K, Westerhof RG, Ballerstedt H, de Bont JAM, Wery J (2007) Optimization of the solvent-tolerant Pseudomonas putida S12 as host for the production of p-coumarate from glucose. Appl Microbiol Biotechnol 74:617–624
Orita I, Sato T, Yurimoto H, Kato N, Atomi H, Imanaka T, Sakai Y (2006) The ribulose monophosphate pathway substitutes for the missing pentose phosphate pathway in the archaeon Thermococcus kodakaraensis. J Bacteriol 188:4698–4704
Roels JA (1983) Energetics and kinetics in biotechnology. Elsevier, Amsterdam
Sahm H (1977) Metabolism of methanol by yeast. Adv Biochem Eng 6:77–103
Sambrook J, Russel DW (2001) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, New York
Schmid A, Dordick JS, Hauer B, Kiener A, Wubbolts M, Witholt B (2001) Industrial biocatalysis today and tomorrow. Nature 409:258–268
Takashita R and Yasueda H (2004) Coryneform bacterium transformed to utilize methanol as carbon source. JP2004261150
Verhoef S, Ruijssenaars HJ, de Bont JAM, Wery J (2007) Bioproduction of p-hydroxybenzoate from renewable feedstock by solvent-tolerant Pseudomonas putida S12. J Biotechnol 132:49–56
Wierckx NJ, Ballerstedt H, de Bont JA, Wery J (2005) Engineering of solvent-tolerant Pseudomonas putida S12 for bioproduction of phenol from glucose. Appl Environ Microbiol 71:8221–8227
Yurimoto H, Hirai R, Matsuno N, Yasueda H, Kato N, Sakai Y (2005a) HxlR, a member of the DUF24 protein family, is a DNA-binding protein that acts as a positive regulator of the formaldehyde-inducible hxlAB operon in Bacillus subtilis. Mol Microbiol 57:511–519
Yurimoto H, Hirai R, Yasueda H, Mitsui R, Sakai Y, Kato N (2002) The ribulose monophosphate pathway operon encoding formaldehyde fixation in a thermotolerant methylotroph, Bacillus brevis S1. FEMS Microbiol Lett 214:189–193
Yurimoto H, Kato N, Sakai Y (2005) Assimilation, dissimilation, and detoxification of formaldehyde, a central metabolic intermediate of methylotrophic metabolism. Chem Rec 5:367–375
Acknowledgments
The authors thank Ton van Maris for his expert help during the research of this paper, and Nick Wierckx for critical reading of this manuscript. This project is financially supported by the Netherlands Ministry of Economic Affairs and the B-Basic partner organizations (www.b-basic.nl) through B-Basic, a public–private NWO-ACTS program (ACTS = Advanced Chemical Technologies for Sustainability). This project was (co)financed by the Kluyver Center for Genomics of Industrial Fermentation, which is part of the Netherlands Genomics Initiative/Netherlands Organization for Scientific Research.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Koopman, F.W., de Winde, J.H. & Ruijssenaars, H.J. C1 compounds as auxiliary substrate for engineered Pseudomonas putida S12. Appl Microbiol Biotechnol 83, 705–713 (2009). https://doi.org/10.1007/s00253-009-1922-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-009-1922-y