
Abstract
Elevated levels of atmospheric CO2 lead to the increase of plant photosynthetic rates, carbon inputs into soil and root exudation. In this work, the effects of rising atmospheric CO2 levels on the metabolic active soil microbiome have been investigated at the Giessen free-air CO2 enrichment (Gi-FACE) experiment on a permanent grassland site near Giessen, Germany. The aim was to assess the effects of increased C supply into the soil, due to elevated CO2, on the active soil microbiome composition. RNA extraction and 16S rRNA (cDNA) metabarcoding sequencing were performed from bulk and rhizosphere soils, and the obtained data were processed for a compositional data analysis calculating diversity indices and differential abundance analyses. The structure of the metabolic active microbiome in the rhizospheric soil showed a clear separation between elevated and ambient CO2 (p = 0.002); increased atmospheric CO2 concentration exerted a significant influence on the microbiomes differentiation (p = 0.01). In contrast, elevated CO2 had no major influence on the structure of the bulk soil microbiome (p = 0.097). Differential abundance results demonstrated that 42 bacterial genera were stimulated under elevated CO2. The RNA-based metabarcoding approach used in this research showed that the ongoing atmospheric CO2 increase of climate change will significantly shift the microbiome structure in the rhizosphere.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The rise of atmospheric carbon dioxide (CO2) concentrations and global warming are well-documented processes. Total annual anthropogenic greenhouse gas emissions have continued to increase, comprising CO2, which represents around 75% of these emissions [1]. Elevated CO2 (eCO2) concentrations have several consequences on plants, such as increased growth in C3, C4, and CAM plants by 41%, 22%, and 15%, respectively [2, 3]; increased plant yield [4]; decreased evapotranspiration of both C3 [5] and C4 plants [6]; augmented photosynthetic capacity [3, 7, 8]; and increased below-ground biomass [9].
Considering that nearly up to 21% of all photosynthetically fixed carbon is transferred to the rhizosphere, roots and root exudates influence the composition and biomass of soil microbiome [10, 11]. Elevated atmospheric CO2 increases efflux amounts of total soluble sugars, amino acids, phenolic acids, and organic acids in the root exudates [12,13,14]. Similarly, the rates of organic carbon as energy sources enhance microbial degradation of soil organic matter (SOC), also known as priming effect [14]. Priming effect is defined as an accelerated decomposition of SOC due to an increased supply of labile C to the soil and changes in the microbial activity as a response [15]. The microbial succession is accompanied by the activation of various, previously dormant microorganisms that respond specifically to the added substrate [15, 16].
The effects of eCO2 levels on soil ecosystems have been studied in free-air CO2 enrichment (FACE) experiments, revealing significant effects of rising CO2 on soil organisms. However, with regard to microbial composition and function related to carbon and nitrogen cycling, mixed results have been obtained. At the BioCON field experiment, it was found that the structure of microbial communities was different between ambient CO2 (aCO2) and eCO2 [17]. Likewise, the abundance of genes involved in labile C degradation and C and N fixation, as RuBisCo, carbon monoxide dehydrogenase (CODH), propionyl-CoA/acetyl-CoA carboxylase (PCC/ACC), nifH and nirS genes were significantly increased under eCO2 [18]. Similarly, He et al. [19] and Xiong et al. [20] have reported a shift of soil microbial communities under eCO2 in a soybean and a maize agro-ecosystem, respectively. These changes included stimulation of key functional genes involved in carbon fixation and degradation, nitrogen fixation, denitrification, methane metabolism, and phosphorus cycling.
Oppositely, some FACE experiments have shown no effects of eCO2 on soil microbiome structure and activity, as Marhan et al. [21] who described that abundances of both total 16S rRNA genes and nitrate-reducing bacteria were not influenced by CO2 but by sampling date and depth. Dunbar et al. [22] described that neither bacterial nor fungal community structure nor composition were altered under eCO2. Pujol Pereira et al. [23] did not find any significant effects of eCO2 on bacterial abundance, soil C, and N concentrations. Butterly et al. [24] reported that changes in microbial community structure were not detected, although eCO2 reduced the abundance of C and N functional genes.
The Giessen free-air CO2 enrichment (Gi-FACE) experiment in Giessen, Germany, has been running since 1998. It is becoming a good predictor model to assess the effects of long-term increased CO2 concentrations on soil microbiome structure and function. Some studies carried out in this facility aimed to assess these changes. Regan et al. [25] reported that in the Gi-FACE extractable organic carbon, dissolved organic nitrogen, NH4+, NO3−, and abundances of genes involved in ammonia oxidation and denitrification depended more on soil depth and moisture gradient than on eCO2. Similarly, also de Menezes et al. [26] described that increases in atmospheric CO2 may cause only minor changes in Gi-FACE’s soil bacterial community composition and that functional responses of the soil community are due to factors like soil moisture rather than CO2 concentration. Brenzinger et al. [27] reported that the abundance and composition of microbial communities in the topsoil under eCO2 presented only small differences from soil under aCO2 (aCO2), concluding that + 20% CO2 had little to no effect on the overall microbial community involved in N-cycling in the Gi-FACE soil. More recently, Bei et al. [28] described that eCO2 had significant effects on the functional expression associated to both rhizosphere microbiomes and plant roots; and that abundances of Eukarya relative to Bacteria were significantly decreased in eCO2 as well.
The question of why some studies reported differences between eCO2 and aCO2 while some others did not is still open. Several abiotic and biotic factors could be the reason for the contradictive observations in the different experimental setups described above. However, all such previous studies conducted in the Gi-FACE used a DNA-based metagenomic approach, with the exception of Bei et al. [28], who utilized a metatranscriptomic approach. The disadvantage of using DNA is that, after a cell dies, amplifiable extracellular DNA can remain in soils for weeks to years and may bias DNA-based estimates of the diversity and structure of soil microbial communities [29, 30]. Moreover, Carini et al. [31] reported that DNA from dead cells or free DNA represented a large fraction of microbial DNA in many soils, comprising approximately 40.7% and 40.5% of amplifiable prokaryotic 16S rRNA genes and fungal ITS amplicons, respectively. Therefore, DNA-depending studies may overestimate the richness of the soil microbiome by up to 55% for prokaryotes and 52% for fungi [31] and in consequence may hide the active microorganisms that are involved in soil microbial processes.
A better approach for assessing differences between eCO2 and aCO2 is the use of RNA instead of DNA for 16S rRNA metabarcoding analysis. The ribosome numbers are correlated to the metabolic activity of bacteria [32], and different studies showed that, with this approach, the active organisms instead of the dormant ones were assessed [33,34,35]. Additionally, results of the metatranscriptomic methodological approach on the Gi-FACE soil microbiome reported by Bei et al. [28] demonstrated that RNA instead of DNA is a better predictor of microbiome composition and activity. For this reason, the aims of the present work were (i) to evaluate the effect of long-term eCO2 concentrations and increased C supply on active soil microbiome through an rRNA-based metabarcoding approach; (ii) to assess the differences between eCO2 and aCO2 conditions in rhizosphere and bulk soils; and (iii) to link these differences with environmental factors.
The following questions have been addressed:
-
1.
Is the community structure of active bacteria different between ambient and elevated CO2 in rhizosphere and/or bulk soil?
-
2.
Which other environmental parameters beside CO2 shape the community?
Material and Methods
Study Site Description
The Gi-FACE study is located at 50° 32′ N and 8° 41.3′ E near Giessen, Germany, at an elevation of 172 m above sea level. It consists of three pairs of rings with a diameter of 8 m; each pair consists of an ambient and an elevated CO2 treatment ring [36]. Since May 1998 until present, elevated CO2 rings have been continuously enriched by 20% above ambient CO2 concentrations during daylight hours. Ambient and elevated CO2 rings are separated by at least 20 m, and each pair is placed at the vertices of an equilateral triangle. The presence of a slight slope within the experimental site (between 0.5 and 3.5°) places the rings on a moisture gradient, such that pair 1 has the lowest mean moisture content (38.8% ± 10.2%) and pair 2 has the highest mean moisture content (46.1% ± 13.2%), whereas pair 3 is intermediate (40.7% ± 11%) [26, 36]. The average annual air temperature and precipitation are 9.4 °C and 580 mm, respectively.
The vegetation is an Arrhenatheretum elatioris Br.Bl. Filipendula ulmaria subcommunity, dominated by Arrhenatherum elatius, Galium album, and Geranium pratense. At least 12 grass species, 15 non-leguminous herbs and up to 5 legumes with small biomass contributions (< 5%) are present within a single plot [37]. The experimental field has not been ploughed for more than 100 years. It has received N fertilization in form of granular mineral calcium ammonium nitrate (40 kg N ha−1 year−1) once a year since 1995 and has been mown twice a year since 1993. The soil at the Gi-FACE site is classified as Fluvic Geysol; its texture is a sandy clay loam over a clay layer, with pH = 6.2 and average C and N contents of 4.5% and 0.45%, respectively, as measured in 2001 [36].
Soil Sampling and Physico-chemical Parameter Measurements
Soil sampling was performed utilizing sawed off 50 ml syringes (11 × 3 cm), and four samples were taken to a depth of ~ 10 cm within each ring in September 2015. Soil cores were gently shaken by hand to remove loosely attached soil (bulk soil), while the soil that remained attached to the roots was considered as rhizosphere soil. Bulk and rhizosphere soils were sieved (< 2 mm) and stored at − 80 °C for further analyses. Samples from each soil core were classified in four groups considering the CO2 conditions (ambient and elevated) and the soil habitat (bulk soil and rhizosphere soil).
Ammonium and nitrate concentrations were measured according to Kandeler et al. [38] and Bak et al. [39]. Water content, dry matter, and water holding capacity of soil samples were measured gravimetrically [40]. Carbon and nitrogen content of soil were measured by pyrolysis coupled to gas chromatography on a EA 1100 elemental analyzer (ThermoQuest, Milan, Italy) using a TCD detector by the Dumas method according to HBU (1996) [41] and VDLUFA (2012) method [42]. Injected CO2 and CO2 soil fluxes were determined from August to September 2015. Injected CO2 was measured at 60 cm above ground with an infrared gas analyzer (LI-COR 6252) [36]. CO2 soil fluxes were measured weekly using an automated closed dynamic chamber system (LI-COR 8100, LI-COR Inc., Lincoln, Nebraska, USA). Per ring, 4 PVC soil collars (20.3 cm diameter) were permanently installed as chamber bases in 2006 and held vegetation free since 2008. Fluxes were calculated from the increase in CO2 concentration in the chamber over the 1–3 min closure time as described by Keidel et al. [43].
Central tendency and dispersion measures were calculated for soil chemical data. CO2 injection and CO2 fluxes data were analyzed using growth curve analysis (GCA) [44], with R packages gazer version 0.1 [45] and lme4 version 1.1–23 [46], creating polynomial-transformed predictor variables, fitting them to a linear mixed model by maximum likelihood, and assessing differences between CO2 conditions with a t-test, using an alpha of < 0.05.
RNA Extraction and Reverse Transcription
RNA extraction was performed following a modified protocol of Mettel et al. [47]. For the extraction, 0.3–0.5 g of soil were weighed in reaction tubes containing 100 mg of sterile zirconia beads, added with 700 µl TPM buffer (50 mM Tris–HCl (pH 5), 1.7% [wt/vol] polyvinylpyrrolidone, 20 mM MgCl2), and vortexed for 30 s. Cells were then disrupted in a cell mill MM200 (Retsch, Haan, Germany) for 2 min at a frequency of 30 Hz. Soil and cell debris were precipitated by centrifugation in a microcentrifuge (Heraeus Fresco, Thermo Fisher Scientific Inc., Waltham) for 5 min at 17,000 g and 4 °C, and then the supernatant was transferred into a fresh reaction tube. To the resulting soil pellet 700 µL of buffer PBL (5 mM Tris–HCl (pH 5), 5 mM Na2EDTA, and 0.1% [wt/vol] sodium dodecyl sulfate) were added, and the disruption process was performed again as described above. Both supernatants from the lysis processes were pooled in one reaction tube.
The pooled supernatant was immediately extracted, initially with the addition of 500 µl of phenol/chloroform/isoamyl alcohol (25:24:1) and subsequent with chloroform/isoamyl alcohol (24:1). Afterwards, each time the sample was centrifuged for 5 min at 17,000 g and 4 °C. The resulting upper aqueous phase was transferred to a new reaction tube, 800 µl of PEG solution was added (30% [wt/vol] polyethylene glycol 6000 and 1.6 M NaCl), incubated in ice for 30 min and centrifuged for 30 min at 17,000 g and 4 °C. Subsequently, the DNA/RNA pellet was washed with 800 µl of ice-cold 75% ethanol, dried out and dissolved in 50 µl of nuclease free water.
After extraction, samples were treated for DNA digestion with RNase-Free DNase Set (QIAGEN GmbH — Germany) according to manufacturer instructions; DNase reaction was stopped with 10 µl of 50 mM EDTA. With the DNA-free RNA, a PCR was carried out, using the universal 16S rRNA gene primers 27F (5’-AGAGTTTGATCMTGGATCMTGGCTCAG-3’) and 1492R (5’- GGTTACCTTGTTACGACTT-3’) [48, 49] and checked on agarose gel electrophoresis to verify the absence of remaining DNA in the samples. Subsequently, reverse transcription was performed utilizing AccuScript High Fidelity 1st Strand cDNA Synthesis Kit (Agilent Technologies, Inc., Cedar Creek, TX, USA) following manufacturer instructions.
16S rRNA Ion Torren Sequencing and Metabarcoding Analysis
The 16S rRNA gene hypervariable regions (V4&V5) were PCR amplified using the set of primers 520F (5’-AYTGGGYDTAAAGNG-3’) [50] and 907R (5’-CCGTCAATTCMTTTRAGTTT-3’) [51] and sequenced by Ion Torrent technique following the protocol described by Kaplan et al. [52]. The obtained Ion Torrent sequencing output was analyzed using QIIME2 version 2020.6.0 [53], sequences were demultiplexed with the QIIME2 cutadapt command [54] using a barcode error rate of 0 and assigned to specific samples by corresponding barcodes. Later, quality control, denoising, sequences dereplication, and chimera filtering were performed using DADA2 software [55]; the first 15 nucleotides were trimmed, and sequences were truncated at a position of 320 nucleotides. Amplicon sequence variants (ASV) generated with DADA2 were taxonomically affiliated with a trained fitted classifier [56, 57] based on the SILVA 138 database [58, 59].
Alpha and beta diversity analyses were performed using R studio software 1.1.419, R packages Phyloseq 1.22.3 [60] and Vegan 2.4–6 [61]. Before diversity analyses, ASVs were collapsed by genera. For alpha diversity assessment, rarefaction was applied and diversity indices (Observed species, Simpson, Shannon, Fisher) were calculated and compared among CO2 conditions and soil habitats using the Wilcoxon test [62] with the Bonferroni correction method through 999 permutations. For non-constrained beta diversity analyses, data were transformed using centered log ratio (clr) method [63, 64], using R package Aldex2 1.18.0 [65]. Later, community dissimilarity distance matrices were created using the Aitchison distance [63, 64] and visualized using principal component analysis (PCA) [66]. Statistical differences among treatments, rings, and CO2 conditions were assessed by a permutational multivariate analysis of variance using Adonis method and employing 999 permutations [67]. Additionally, the degree of dispersion of the bacterial community composition from the four soil cores taken in each ring was assessed as described above. Redundancy analysis (RDA) was used to explore associations between microbial community structures and environmental parameters, and a permutation test of redundancy analysis using 999 permutations was applied for evaluating their statistical significance [68].
For the analysis of correlation between bacterial genera and environmental parameters, the genera belonging to the core microbiome of each of the soil sample groups were calculated, and their counts were transformed to relative abundance with package Microbiome version 1.8.0 [69]. Later, core microbiomes were calculated including genera with a total relative abundance of ≥ 0.01% and present in ≥ 85% of the corresponding group’s samples. A correlation test was performed using Aldex2 1.18.0 [65] and its “aldex.corr” function, utilizing Pearson's correlation coefficient, and p values were corrected using false discovery rate (FDR) method with an alpha of < 0.05.
Differential abundance of genera from rhizosphere soils was assessed by comparing the core microbiomes of each CO2 condition utilizing the R packages DESeq2 1.24.0 [70] and Aldex2 1.18.0 [65]. DESeq2 analysis was performed by estimating the size factor and the dispersion using the geometric mean of the core microbiome genera; later, values were fitted with a generalized linear model using negative binomial distribution and applying a Wald significance tests, the option “local” for fitting of dispersions to the mean intensity and an alpha threshold of < 0.05. Aldex2 analysis was done by performing a centered log ratio (clr) transformation using as denominator the geometric mean abundance of all features and 128 Monte Carlo instances; later, a Welch’s t-test with a Benjamini–Hochberg correction and threshold < 0.05 was performed.
Functional capabilities based on the obtained 16S rRNA data were predicted using PICRUSt2 version (v2.3.0 beta) [71]. PICRUSt2 analysis was carried out using the default pipeline option. Afterward, EC number, KO functions, and MetaCyc non-constrained beta diversity and differential abundance analyses were performed as described above.
Quantitative PCR
The quantification of 16S rRNA gene to estimate total bacterial abundance was performed following the protocol described by Kaplan et al. [52], but instead of DNA, cDNA products described above were used for the quantification. Quantitative PCR (qPCR) was conducted on a Rotor-Gene Q (Qiagen, Hilden, Germany) by using Absolute qPCR SYBR Green Mix (Thermo Fisher Scientific). Statistical comparisons were done with Kruskal–Wallis and Wilcoxon tests with the Benjamini–Hochberg adjustment method using R Package stats version 3.6.3.
Results
Ion Torrent Sequencing
A total of 5,855,099 raw sequences were obtained. After demultiplexing, sequences were assigned to each sample, ranging sequence counts in each sample from 306,675 to 22,410. After quality control, denoising, sequence dereplication, and chimera filtering with DADA2 software, 2,674,159 sequences were removed, resulting in 3,180,940 non-chimeric sequences and 11,587 representative sequences which were grouped into ASVs (Amplicon sequence variations) at a 99% similarity. Later, sequences belonging to chloroplast and mitochondria were removed, resulting in 11,508 ASVs.
Soil Microbial Diversity
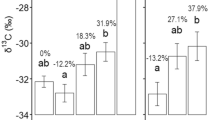
Diversity indexes were evaluated to assess differences in soil microbiome between eCO2 and aCO2 conditions. In the Gi-FACE, soil active bacterial diversity changed due to the influence of increased concentrations of CO2 (Fig. 1). These changes are better appreciated when comparing bulk and rhizosphere soil fractions from aCO2 and eCO2 rings separately. In regard to alpha diversity of rhizosphere and bulk soil fractions from aCO2 rings, significantly higher diversity values were observed in bulk compared to rhizosphere soils with Observed species (p value 0.00036), Shannon (p value 0.0086), and Fisher (p value 0.00036) indexes (Fig. 1). Nevertheless, this difference was not detected between bulk and rhizosphere soil fractions from eCO2 rings, indicating an evenness between the rhizosphere and bulk soils in eCO2 rings (Fig. 1). Likewise, eCO2 rhizosphere soil presented greater diversity values in comparison to its aCO2 counterpart, according to Observed species (p value 0.0193) and Fisher (p value 0.0193) indexes.

Alpha diversity metrics. aCO2, ambient CO2 conditions; eCO2, elevated CO2 conditions. * p smaller 0.01, ** p smaller 0.001, *** p smaller 0.0001
A distance matrix was created using the Aitchison distance and later ordinated using the principal component analysis (PCA) to further analyze the microbiome composition. Initially, the dispersion of the four soil cores taken within each ring and their distance to the centroids was assessed. They indicated a considerably different soil microbiome composition in each soil core, even when soil cores of the same rings were compared (S1). On the other hand, the assessment of differences among the evaluated habitats showed that the strongest effect on the bacterial microbiome differentiation in the soil was the ring factor, either for rhizosphere or bulk soils (p value 0.001).
Similarly, there were significant differences among the community composition of the four evaluated groups (aCO2 bulk soil, aCO2 rhizosphere soil, eCO2 bulk soil, eCO2 rhizosphere soil) (p value 0.001). In the same way, the PCA showed a clear differentiation between the microbiome composition of the rhizospheres from eCO2 and aCO2 rings (p value 0.002) (Fig. 2a). On the contrary, the separation of the microbial community composition between the bulk soils from aCO2 and eCO2 rings was not clear and statistically not significant (p value 0.327) (Fig. 2b).

Principal component analysis (PCA) calculated based on Aitchison community dissimilarity distance matrix of a rhizosphere soils from ambient and elevated CO2 rings and b bulk soils from ambient and elevated CO2 rings. A, ambient CO2 rings; E, elevated CO2 rings; aCO2, ambient CO2 conditions; eCO2, elevated CO2 conditions
Effect of Environmental Parameters on Microbial Community
A redundancy analysis (RDA) was carried out to assess the effect of environmental factors on the soil microbiome of the Gi-FACE. The results indicated that continuously higher environmental CO2 concentration was a factor that exerted a significant effect on the differentiation of the microbial communities of eCO2 rings (p value 0.021) (Table 1, Fig. 3a). Furthermore, CO2 soil fluxes on average were 35% higher in eCO2 rings in comparison to the aCO2 ones, and this difference was statistically significant throughout the assessed period of time (p value 0.031) (Fig. 3b). Moreover, increased soil fluxes of CO2 are associated with the differences that were observed in Gi-FACE soil microbiome (p value 0.001).

Time series data from August to September 2015 of a average environmental CO2 concentrations and b average soil CO2 fluxes of ambient (blue) and elevated (red) CO2 conditions; level of confidence interval of 0.95. Redundancy analysis (RDA) based on Aitchison community dissimilarity distance matrix of c rhizosphere soils from ambient (blue) and elevated (red) CO2 rings and d bulk soils from ambient (blue) and elevated (red) CO2 rings; black dots indicate soil bacterial genera
Likewise, the ammonium content in the whole soil, rhizosphere and bulk soil fractions had a significant influence on the community composition (Table 1), despite the fact that soil ammonium concentrations were not significantly different between eCO2 and aCO2 rings (p value 0.313) (S2). Similarly, the total carbon content had significant influence on the whole soil and bulk soil microbial community structure (Table 1), but likewise ammonium there were no significant differences in carbon content between aCO2 and eCO2 rings (p value 0.1304) (S2). On the contrary, the average carbon/nitrogen ratio in the whole soil of the eCO2 rings (11.1:1) was significantly higher in comparison with aCO2 rings (10.69:1) (p value 0.0069) and had a significant effect (p value 0.025) on the microbial community composition (Table 1).
Furthermore, when observing each habitat separately, the RDA indicated that in the rhizosphere soil, the CO2 atmospheric concentration had a significant effect on the microbiome differentiation between the aCO2 and eCO2 rings (p value 0.010) (Table 1, Fig. 3c). In contrast, eCO2 had no substantial influence on the composition of the microbial community’s structure of the bulk soils (p value 0.097) (Table 1, Fig. 3d).
Correlation analysis between environmental variables and rhizosphere soil core microbiome demonstrated that the abundance of several bacterial genera was either positively or negatively correlated with environmental CO2 concentrations and soil CO2 fluxes. Among the main bacterial families that were significantly positively correlated with environmental eCO2 and soil CO2 fluxes concentrations are Rhodanobacteraceae, “Labraceae,” Xanthomonadaceae, Rhodobacteraceae, Rhizobiaceae, Pseudomonadaceae, Phaselicystidaceae, Haliangiaceae, Bacillaceae, Streptomycetaceae, Xanthobacteraceae, Burkholderiaceae, Devosiaceae, Haliangiaceae, Comamonadaceae and Polyangiaceae (Table S3.1). On the contrary, families “Solibacteraceae,” Caulobacteraceae, Acetobacteraceae, Thermoactinomycetaceae, Beijerinckiaceae, and Blastocatellaceae were negatively correlated with environmental eCO2 and soil CO2 fluxes (Table S3.1). Moreover, bacterial orders Nitrospirales, Caulobacterales, “Rokubacteriales,” Vicinamibacterales, “Tistrellales,” and “Rokubacteriales” were significantly correlated with NH4+ content and soil water holding capacity in rhizosphere soils (Table S3.1).
Changes on the Rhizosphere Microbial Community Composition
Differential abundance analyses demonstrated that several rhizosphere soil genera were affected. Both Aldex2 and DESeq2 demonstrated that 42 bacterial genera were stimulated under eCO2, among those are Haliangium, Phaselicystis, Rhizobacter, Pseudomonas, Rhizobium, Phyllobacterium, Mesorhizobium, Rhodanobacter, Labrys, unidentified genus of the class “Sericytochromatia,” Dokdonella, Massilia, Burkholderia, Bacillus, Novosphingobium, Acidibacter, and Streptomyces (Fig. 4a, Fig. 4b). These genera showed Log2 Fold changes ranging from 0.910 to 9.67. Furthermore, Aldex2 test showed that other 56 genera were significantly stimulated in the rhizosphere soil of eCO2 rings. These genera belonged mainly to bacterial families Nocardioidaceae, Beijerinckiaceae, Pyrinomonadaceae, “Koribacteraceae”, “Xiphinematobacteraceae,” Propionibacteriaceae, Dongiaceae, Geminicoccaceae, Solirubrobacteraceae, Blastocatellaceae, Caulobacteraceae, Nitrosomonadaceae, Xanthobacteraceae, Caulobacteraceae, Fibrobacteraceae, Acetobacteraceae, unidentified family of the phylum “Latescibacterota,” Myxococcaceae, “Solibacteraceae”, Rhizobiaceae, and Gemmatimonadaceae (S3).

Differential abundances of core microbiome bacterial genera of rhizosphere soil under elevated and ambient CO2. a DESeq2 test results with an alpha threshold < 0.05 and error expressed as standard error of log fold change. b Aldex2 results using centered log ratio (clr) transformation and the geometric mean abundance of all features; red points indicate significantly different genera after Welch’s t-test and Benjamini–Hochberg correction with an alpha threshold < 0.1
On the contrary, both differential abundance tests indicated that some genera presented a decreased of abundance under eCO2 conditions. Among these are unidentified genus of the family Vicinamibacteraceae, Pasteuria, Caulobacter, unidentified genus of the family “Entotheonellaceae,” Brevundimonas, Methylobacterium-Methylorubrum, Sumerlaea, Blastocatella, Phenylobacterium, Lacunisphaera, Roseomonas, and Opitutus. These genera had Log2 Fold changes from − 0.421 to − 9.31 in eCO2 ring (Fig. 4a, Fig. 4b, S3).
Functional Metagenomics Prediction
Beta diversity results of functional capabilities based on 16S rRNA data showed significant differences on functional metagenome’s composition of rhizosphere soils from aCO2 and eCO2 conditions. PICRUSt2 predicted functional metagenome were different regarding Enzyme Commission number (EC number) (p value 0.005), KEGG Orthology (KO) for molecular functions (p value 0.019), and MetaCyc Metabolic Pathways (p value 0.022) (S4). Moreover, similar to taxonomical results, predicted bulk soil’s functional metagenomics from aCO2 and eCO2 conditions did not show great differences regarding its beta diversity, neither on EC numbers (p value 0.197), KEGG Orthology (p value 0.179), or MetaCyc Metabolic Pathways (p value 0.317) (S4).
Besides, the analyses of significantly affected predicted enzymes indicated that several enzymes of degradation of carbon compounds were significantly stimulated in rhizospheric soils under eCO2 conditions. Enzymes involved in carbohydrates, lipids, amino acids, and polycyclic aromatic hydrocarbon degradation were significantly stimulated. Additionally, numerous predicted enzymes and pathways of synthesis of cellular components, membrane transporters, and quorum sensing were significantly higher under eCO2 conditions (S5). Also, according to KEGG Orthology for molecular functions, several enzymes involved in nitrogen fixation, nitric-oxide synthesis, and nitrite and nitrate reduction were predicted to be more abundant in eCO2 rhizosphere soil (S5).
Quantitative PCR
Active biomass estimation by 16S rRNA quantification demonstrated changes due to eCO2 concentrations. A 20% increase of 16S rRNA copy numbers per g dry weight soil in eCO2 rhizosphere (2.07 ± 0.50*108) in comparison to aCO2 rhizosphere (1.66 ± 0.44*108) was observed (p value 0.0001). Nevertheless, when comparing the 16S rRNA copy numbers per gram dry weight soil of bulk soils from aCO2 (2.35 ± 0.80*108) and eCO2 (2.35 ± 0.79*108) conditions, no significant differences were found (p value 0.9588) (Fig. 5). Moreover, significant differences were found between bulk and rhizosphere soils from aCO2 (p value 2.1 * 10–5) with in average 29% more copies per dry weigh in bulk soil compared to rhizosphere soil. Nonetheless, when comparing rhizosphere and bulk soils from eCO2 rings, this difference is lower and not significant (p value 0.1455), with the bulk soil having 12% more copies than the rhizosphere soil (Fig. 5).

Boxplot of 16S rRNA quantification of ambient CO2 rings bulk soil (a-bulk), elevated CO2 rings bulk soil (e-bulk), ambient CO2 rings rhizosphere soil (a-rhizo) and elevated CO2 rings rhizosphere soil (e-rhizo). Significance codes: 0.0001 ‘***’
Discussion
Changes in Microbiome Structure and Composition
Elevated CO2 concentrations affect the composition and biomass of soil microbial communities in the rhizosphere because of greater inputs of labile carbon (C) via root exudation may increase the microbial N demand. This causes an increased competition between plants and soil microorganisms for available N; therefore, N dynamics are likely to change under eCO2 [10, 11, 14, 72].
Our results showed that eCO2 had a strong effect in the Gi-FACE on the metabolic active microbiome of the rhizosphere soil, in contrast to the microbiome of the bulk soil which remained mostly unaffected. Alpha diversity indices indicate that a shift occurred under eCO2 conditions, producing an evenness in terms of alpha diversity between rhizosphere and bulk soil. Since significant differences were found between bulk and rhizosphere soil of aCO2 rings, this evenness represents an increase in alpha diversity of eCO2 rhizosphere soil (Fig. 1). Furthermore, beta diversity results revealed a different abundance and microbial community composition in the rhizosphere of eCO2 rings compared to aCO2 rings (Fig. 2).
These results differ from previous reports of the eCO2 effects on the Gi-FACE soil microbiome, which stated that only subtle or no effect occurred on microbial communities and that the differences were mostly due to soil conditions and the moisture gradient that occurs at this facility [25,26,27]. Similarly, to the aforementioned studies, our data confirmed that samples from ring-pair A1-E1 had lower water content in comparison to A2-E2 and A3-E3 samples (S2), and that water holding capacity (WHC) significantly influenced the soil microbiome (Table 1, Fig. 3). This was observable in the effect that the ring-pair factor had on the beta diversity of the Gi-FACE soil microbiome (p value 0.001) (Fig. 2).
Nonetheless, besides the moisture gradient, the observed differences caused by the eCO2 were likely detected due to the RNA-based metabarcoding approach used in our work, which is able to differentiate the metabolic active microorganisms from the inactive ones, avoiding the biases caused by DNA of dead cell or extracellular DNA, which can comprise approximately 41% of the amplifiable prokaryotic 16S rRNA genes in soil [31]. However, RNA metabarcoding has its limitations as well, mainly due to the fact that RNA conversion to cDNA requires the use of a reverse transcriptase which lacks proofreading activity, creating point mutations in some of the cDNA sequences [73]. Reverse transcriptase also regularly performs template switching, which can lead to the production of chimeric cDNA sequences and the creation of shortened isoform sequences from intramolecular template switching [74, 75]. Nevertheless, in our study these limitations were minimized by using a Moloney murine leukemia virus reverse transcriptase (MMLV-RT) derivative combined with a E. coli DNA polymerase III ε subunit which lowers the reverse transcription error rate by threefold, and later the resulting cDNA was amplified with a proofreading DNA polymerase which produced up to eightfold fewer errors [76].
The study of Bei et al.[28], which also addressed the active microbial community by using a metatranscriptomic approach, supports our results. For the summer of 2015, the same year that we took the samples for this study, they reported significant effects of eCO2 on the functional expression related to rhizosphere and plant roots associated microbiomes in the Gi-FACE. Also, similarly to our work, they described that the increase in bacterial abundance was related to significant enrichment of different taxonomical groups, including Acidobacteria, Actinobacteria, and Proteobacteria, and changes related to a significant decrease in Fungi and increase in Actinobacteria.
However, Bei et al. [28] found no significant eCO2 effect on the rhizosphere soil and root-associated microbiomes during the summer of 2017. These contrasting results for different years may result from climatic conditions in summer, since the summer 2015 was characterized by prolonged heat waves, while the mean temperature in summer 2017 was closer to the long-term average. The effect of eCO2 on the soil rhizosphere microbiome we found in our study may be affected by the above average temperatures of this particular year. Additionally, the prediction that heat waves will occur more frequently in the future [77] emphasizes the importance of our findings.
The reason why only the rhizosphere microbiome, in contrast to the bulk soil microbiome, was affected by eCO2 influx is most probably a consequence of the priming effect of the increased flux of roots exudates and consequently higher availability of carbon compounds. This increased supply of labile C causes an accelerated decomposition of soil organic C [15], which activates previously dormant microorganisms [15, 16].
Effect of eCO2Concentration on Microbial Community, C and N Cycles
Our results of the effect of environmental parameters on soil microbiome composition demonstrated that several rhizosphere bacterial families such as Rhodanobacteraceae, “Labraceae,” Xanthomonadaceae, Rhodobacteraceae, Rhizobiaceae, Pseudomonadaceae, Phaselicystidaceae, Haliangiaceae, Bacillaceae, Streptomycetaceae, Xanthobacteraceae, Burkholderiaceae, Devosiaceae, Haliangiaceae, Comamonadaceae, and Polyangiacea were positively correlated with eCO2 fumigation and soil CO2 fluxes. Within these families are found bacterial genera as Streptomyces, Burkholderia, Dokdonella, Bacillus, Pseudolabrys, Devosia, Mesorhizobium, Acidibacter, Rhizobacter, Rhodanobacter, Arenimonas, Amaricoccus, Phyllobacterium, Rhizobium, Pseudomonas, Phaselicystis, and Haliangium. Furthermore, the aforementioned genera had significant higher counts under eCO2 conditions according to DESeq2 and Aldex2 results. From other experiments, it was also reported that under eCO2 conditions the rhizosphere soil microbial communities had changed [78]. Increased atmospheric CO2 concentrations could also change the competitive ability of Rhizobium leguminosarum bv. trifolii, probably due to changes in root exudates [79]. In salt marsh systems containing the halophyte Suaeda japonica, it was reported that gene abundances and microbial community structures were both affected by eCO2, and rhizospheric microorganisms responded to eCO2 more strongly than those inhabiting the bulk soil [80]. Song et al. [81] described that community composition of soil microbiota associated with Phytolacca americana and Amaranthus cruentus roots were significantly affected by eCO2, and numbers of bacteria and fungi, as well as microbial C and N in the rhizosphere soils of both species, were higher at eCO2.
Greater carbon input due to eCO2 also explains the increase of 35% in soil CO2 fluxes and the 20% augmentation in 16S rRNA copy numbers from active bacterial biomass observed in rhizosphere soil under eCO2 in comparison to aCO2, which corresponded to an increased soil biological activity in Gi-FACE. Cheng et al. [82] described that eCO2 affected soil microbial respiration, producing an augmentation of microbial biomass and activities. Similarly, King et al. [83] showed that eCO2 increased soil respiration at four forest FACE experiments. Blagodatskaya et al. [84] demonstrated that augmented available organic C released by roots at eCO2 altered the ecological strategy of the soil microbial community, occurring a shift to a higher contribution of fast-growing species. The increased biological activity in eCO2 rhizosphere soil is supported by the predicted functional metagenome obtained with PICRUSt2, which shows significant increases in several enzymes involved in cellular components biosynthesis such as peptidoglycan, lipopolysaccharide, amino acids, bacterial motility proteins, and lipids synthesis (S5). These results differ from those obtained by Pujol Pereira et al. [23], who reported that on soybean [Glycine max (L.) Merr.], eCO2 decreased 16S rRNA gene abundance in rhizosphere soil by 31%. Also, Marhan et al. [21] described that abundances of total 16S rRNA were not influenced by CO2 but by sampling date and depth. Likewise, Brenzinger et al. [27] and Bei et al. [28] reported that at the Gi-FACE no differences between aCO2 and eCO2 rings were found regarding the 16S rRNA gene. However, Bei et al. [28] described that in summer of 2015 under eCO2 conditions the functional metagenome of rhizosphere soil presented an increase on amino acids and carbohydrates metabolisms, membrane transporters, and quorum sensing proteins: similar to our study’s PICRUSt2 results (S5).
In addition, several genera involved in the degradation of carbon (C) compounds were stimulated under eCO2 conditions; among these are Pseudomonas and Bacillus (Fig. 4, S2) that have been previously reported to degrade lignocellulose materials. Pseudomonas boreopolis produces a cellulase-free xylanase with a high activity of hemicellulose degradation [85]. Maki et al. [86] reported that Bacillus strain (55S5) and a Pseudomonas strain (AS1) displayed high potential for lignocellulose decomposition due to a variety of cellulase and xylanase activities. Trujillo-Cabrera et al. [87] described the isolation of cellulolytic bacteria from high humus content soils, as Bacillus thuringiensis and Pseudomonas gessardii. The augmentation of these taxa would agree with the predicted functional metagenome, which indicated an increment of several enzymes involved in lignocellulose materials degradation, as Chitinase (EC:3.2.1.14), Endo-1,3(4)-beta-glucanase (EC:3.2.1.6), Endo-1,4-beta-xylanase (EC:3.2.1.8), and Cellulase (EC:3.2.1.4) (S5). Similar results have been described by He et al. [17, 19], who reported that soils of a soybean agro-ecosystem and a glacial outwash sandplain showed increased abundance of encoding genes for enzymes involved in labile C degradation such as amylase, glucoamylase, pullulanase, fungal arabinofuranosidase, xylanase, endoglucanase, acetylglucosaminidase, and exochitinase. Likewise, Xiong et al. [20] described that alpha-amylase, cellobiase, endoglucanase, vanillin dehydrogenase, endochitinase, and phenoloxidase encoding genes were stimulated under eCO2 in soybean and maize fields. The above-mentioned increase of C compounds degradation could occur as a response of greater C availability due to an increase of root exudates under eCO2 conditions.
Moreover, these changes in the C influx could induce a reduction of available N in the soil ecosystem [24], which alters the N cycle and induces significant changes in soil biogeochemical characteristics in the rhizosphere, such as NO3−, available K+, soil microbial biomass carbon (SMBC), and available PO42− [78]. The aforementioned process could explain the higher carbon/nitrogen ratio found in our study in eCO2 rings in comparison with ambient ones and might also explain why some genera involved in different processes of the nitrogen cycle were stimulated under eCO2 conditions. Genera belonging to families Rhizobiaceae and Xanthobacteraceae as Rhizobium, Mesorhizobium, and Phyllobacterium have been extensively reported as nitrogen-fixing bacteria [88,89,90], and in our study presented Log2 fold increases ranging from 6.78 to 8.04. Also, PICRUSt2 results indicate that functional orthologs of the enzyme nitrogenase (EC:1.18.6.1) were significantly augmented in eCO2 rhizosphere soil (S5). This increase in the abundance of nitrogen-fixing bacteria could have occurred as response to N deficiency, which eventually became a limiting factor for biomass production under eCO2. Similar results were reported by Li et al. [91], who described a 24% increment of 15 N in mine tailing soils under eCO2 and a dominance of uncultured nitrogen-fixing bacteria.
Aldex2 correlation results demonstrated a significant negative correlation between NH4+ content and Nitrospira genus under eCO2 conditions. Although NH4+ values were not significantly different between aCO2 and eCO2, NH4+ content was on average 10% higher in aCO2 soils, which suggest that nitrification processes could have been affected due to elevated environmental CO2. Alterations in nitrification process in the Gi-FACE have been already described by Müller et al. [72], who reported that eCO2 reduced NH4+ oxidation to NO3− by 25%.
Furthermore, several denitrifying genera as Streptomyces, Rhodanobacter, Pseudomonas, Burkholderia, and Bacillus were significantly stimulated in eCO2 rhizospheric soil with Log2 fold changes between 5.37 and 8.065 [92,93,94,95,96]. Additionally, predicted functional metagenome indicate that several orthologs involved in the denitrification process, as nitric-oxide synthase (NAD(P)H) (EC:1.14.13.165), nitrate reductase (EC:1.7.99.4), nitrite reductase (NO-forming) (EC:1.7.2.1), nitrite reductase (cytochrome; ammonia-forming) (EC:1.7.2.2), periplasmic nitrate reductase NapA (EC:1.7.99.-), and nitric oxide reductase NorD protein, have significantly greater abundance under eCO2 conditions (S5).
In summary, our results demonstrate that in the Gi-FACE, the rhizosphere soil microbiome was significantly affected due to the influence of increased CO2 concentrations alongside other environmental factors. The increment of carbon input due to eCO2 possibly augmented labile carbon degradation in rhizosphere soil reflected by the increment of bacteria biomass and CO2 soil emissions. The aforementioned processes could cause a nitrogen constraint, observed in the increment of the C:N ratio, and decreased of NH4+, which likely triggered a shift in the rhizosphere soil microbiome with an increment of nitrogen fixing and denitrifying taxa. The observed increase of denitrifier genera might explain the increased N2O fluxes under eCO2 conditions, previously described in the Giessen FACE [27, 97, 98]. Similarly, our data support the results described by Moser et al. [98], who reported that under eCO2 conditions, N2O emissions were 1.79-fold higher and that the linear interpolations showed a 2.09-fold increase in N2O emissions mostly because of the oxidation of organic N and incomplete reduction of NO2−, emitting N2O instead of N2 (Fig. 6).

Model diagram of the effect of elevated CO2 on the rhizosphere microbiome of C and N cycles bacterial taxa of the Giessen FACE experiment
Our findings suggest that alterations in carbon cycle affects nitrogen cycle dynamics in grassland soils, due to changes on the microorganisms involved on the different processes of these cycles. Nonetheless, further analyses would be necessary to assess the Gi-FACE microbiome metatranscriptome of carbon and nitrogen cycles, how they are affected by eCO2, and how this effect depends on ambient temperature regimes like summer heat waves.
Data Availability
The authors declare that the data supporting the findings of this study are available within the article and its Supplementary Information. cDNA sequence data are available in the GenBank database under the accession number PRJNA656997.
Code Availability
Not applicable.
References
IPCC (2014) Climate change 2014: Synthesis report. Contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change. IPCC, Geneva, Switzerland
Idso K (1994) Plant responses to atmospheric CO2 enrichment in the face of environmental constraints: a review of the past 10 years’ research. Agric For Meteorol 69:153–203. https://doi.org/10.1016/0168-1923(94)90025-6
He P, Bader KP, Radunz A, Schmid GH (1995) Consequences of high CO2-concentrations in air on growth and gas-exchange rates in Tobacco mutants. Z Naturforsch 50c:781–788
Kimball BA (1983) Carbon dioxide and agricultural yield: An assemblage and analysis of 430 prior observations. Agron J 75:779. https://doi.org/10.2134/agronj1983.00021962007500050014x
Owensby CE, Ham JM, Knapp AK et al (1997) Water vapour fluxes and their impact under elevated CO2 in a C4-tallgrass prairie. Glob Chang Biol 3:189–195. https://doi.org/10.1046/j.1365-2486.1997.00084.x
Kimball BA (2016) Crop responses to elevated CO2 and interactions with H2O, N, and temperature. Curr Opin Plant Biol 31:36–43. https://doi.org/10.1016/j.pbi.2016.03.006
Habash DZ, Paul MJ, Parry MAJ et al (1995) Increased capacity for photosynthesis in wheat grown at elevated CO2 - the relationship between electron transport and carbon metabolism. Planta 197:482–489. https://doi.org/10.1007/BF00196670
Johnson RM, Pregitzer KS (2007) Concentration of sugars, phenolic acids, and amino acids in forest soils exposed to elevated atmospheric CO2 and O3. Soil Biol Biochem 39:3159–3166. https://doi.org/10.1016/j.soilbio.2007.07.010
Jongen M, Jones MB, Hebeisen T et al (1995) The effects of elevated CO2 concentrations on the root growth of Lolium perenne and Trifolium repens grown in a FACE* system. Glob Chang Biol 1:361–371
Walker TS, Bais HP, Grotewold E, Vivanco JM (2003) Root exudation and rhizosphere Biology root exudation and rhizosphere biology. Plant Physiol 132:44–51. https://doi.org/10.1104/pp.102.019661.Although
Li K, Guo XW, Xie HG et al (2013) Influence of root exudates and residues on soil microecological environment. Pakistan J Bot 45:1773–1779
Phillips RP, Meier IC, Bernhardt ES et al (2012) Roots and fungi accelerate carbon and nitrogen cycling in forests exposed to elevated CO2. Ecol Lett 15:1042–1049. https://doi.org/10.1111/j.1461-0248.2012.01827.x
Jia X, Wang W, Chen Z et al (2014) Concentrations of secondary metabolites in tissues and root exudates of wheat seedlings changed under elevated atmospheric CO2 and cadmium-contaminated soils. Environ Exp Bot 107:134–143. https://doi.org/10.1016/j.envexpbot.2014.06.005
Dong J, Hunt J, Delhaize E et al (2021) Impacts of elevated CO2 on plant resistance to nutrient deficiency and toxic ions via root exudates: a review. Sci Total Environ 754:142434. https://doi.org/10.1016/j.scitotenv.2020.142434
Blagodatskaya E, Kuzyakov Y (2008) Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: critical review. Biol Fertil Soils 45:115–131. https://doi.org/10.1007/s00374-008-0334-y
Di Lonardo DP, De Boer W, Klein Gunnewiek PJA et al (2017) Priming of soil organic matter: chemical structure of added compounds is more important than the energy content. Soil Biol Biochem 108:41–54. https://doi.org/10.1016/j.soilbio.2017.01.017
He Z, Xu M, Deng Y et al (2010) Metagenomic analysis reveals a marked divergence in the structure of belowground microbial communities at elevated CO2. Ecol Lett 13:564–575. https://doi.org/10.1111/j.1461-0248.2010.01453.x
Xu M, He Z, Deng Y et al (2013) Elevated CO2 influences microbial carbon and nitrogen cycling. BMC Microbiol 13:124. https://doi.org/10.1186/1471-2180-13-124
He Z, Xiong J, Kent AD et al (2014) Distinct responses of soil microbial communities to elevated CO2 and O3 in a soybean agro-ecosystem. ISME J 8:714–726. https://doi.org/10.1038/ismej.2013.177
Xiong J, He Z, Shi S et al (2015) Elevated CO2 shifts the functional structure and metabolic potentials of soil microbial communities in a C4 agroecosystem. Sci Rep 5:1–9. https://doi.org/10.1038/srep09316
Marhan S, Philippot L, Bru D et al (2011) Abundance and activity of nitrate reducers in an arable soil are more affected by temporal variation and soil depth than by elevated atmospheric [CO2]. FEMS Microbiol Ecol 76:209–219. https://doi.org/10.1111/j.1574-6941.2011.01048.x
Dunbar J, Gallegos-Graves LV, Steven B et al (2014) Surface soil fungal and bacterial communities in aspen stands are resilient to eleven years of elevated CO2 and O3. Soil Biol Biochem 76:227–234. https://doi.org/10.1016/j.soilbio.2014.05.027
Pujol Pereira EI, Chung H, Scow K, Six J (2013) Microbial communities and soil structure are affected by reduced precipitation, but not by elevated carbon dioxide. Soil Sci Soc Am J 77:482. https://doi.org/10.2136/sssaj2012.0218
Butterly CR, Phillips LA, Wiltshire JL et al (2016) Long-term effects of elevated CO2 on carbon and nitrogen functional capacity of microbial communities in three contrasting soils. Soil Biol Biochem 97:157–167. https://doi.org/10.1016/j.soilbio.2016.03.010
Regan K, Kammann C, Hartung K et al (2011) Can differences in microbial abundances help explain enhanced N2O emissions in a permanent grassland under elevated atmospheric CO2? Glob Chang Biol 17:3176–3186. https://doi.org/10.1111/j.1365-2486.2011.02470.x
de Menezes AB, Müller C, Clipson N, Doyle E (2016) The soil microbiome at the Gi-FACE experiment responds to a moisture gradient but not to CO2 enrichment. Microbiology 162:1572–1582. https://doi.org/10.1099/mic.0.000341
Brenzinger K, Kujala K, Horn MA et al (2017) Soil conditions rather than long-term exposure to elevated CO2 affect soil microbial communities associated with N-cycling. Front Microbiol 8:1–14. https://doi.org/10.3389/fmicb.2017.01976
Bei Q, Moser G, Wu X et al (2019) Metatranscriptomics reveals climate change effects on the rhizosphere microbiomes in European grassland. Soil Biol Biochem 138:1–10. https://doi.org/10.1016/j.soilbio.2019.107604
Morrissey EM, McHugh TA, Preteska L et al (2015) Dynamics of extracellular DNA decomposition and bacterial community composition in soil. Soil Biol Biochem 86:42–49. https://doi.org/10.1016/j.soilbio.2015.03.020
Dlott G, Maul JE, Buyer J, Yarwood S (2015) Microbial rRNA: RDNA gene ratios may be unexpectedly low due to extracellular DNA preservation in soils. J Microbiol Methods 115:112–120. https://doi.org/10.1016/j.mimet.2015.05.027
Carini P, Marsden PJ, Leff JW et al (2016) Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. Nat Microbiol 2:1–6. https://doi.org/10.1038/nmicrobiol.2016.242
Felske A, Engelen B, Nübel U, Backhaus H (1996) Direct ribosome isolation from soil to extract bacterial rRNA for community analysis. Appl Environ Microbiol 62:4162–4167
Duineveld BM, Kowalchuk GA, Keijzer A et al (2001) Analysis of bacterial communities in the rhizosphere of chrysanthemum via denaturing gradient gel electrophoresis of PCR-amplified 16S rRNA as well as DNA fragments coding for 16S rRNA. Appl Environ Microbiol 67:172–178. https://doi.org/10.1128/AEM.67.1.172-178.2001
Hoshino YT, Matsumoto N (2007) DNA- versus RNA-based denaturing gradient gel electrophoresis profiles of a bacterial community during replenishment after soil fumigation. Soil Biol Biochem 39:434–444. https://doi.org/10.1016/j.soilbio.2006.08.013
Hunt DE, Lin Y, Church MJ et al (2013) Relationship between abundance and specific activity of bacterioplankton in open ocean surface waters. Appl Environ Microbiol 79:177–184. https://doi.org/10.1128/AEM.02155-12
Jäger HJ, Schmidt SW, Kammann C et al (2003) The University of Giessen free-air carbon dioxide enrichment study: description of the experimental site and of a new enrichment system. J Appl Bot Bot 77:117–127
Andresen LC, Yuan N, Seibert R et al (2018) Biomass responses in a temperate European grassland through 17 years of elevated CO2. Glob Chang Biol 24:3875–3885. https://doi.org/10.1111/gcb.13705
Kandeler E, Gerber H (1988) Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol Fertil Soils 6:68–72. https://doi.org/10.1007/BF00257924
Bak F, Scheff G, Jansen KH (1991) A rapid and sensitive ion chromatographic technique for the determination of sulfate and sulfate reduction rates in freshwater lake sediments. FEMS Microbiol Lett 85:23–30. https://doi.org/10.1111/j.1574-6968.1991.tb04694.x
Forster JC (1995) Methods in Applied Soil Microbiology and Biochemistry. In: Alef K, Nannipieri P (eds) Methods in applied soil microbiology and biochemistry. Academic Press, San Diego, pp 105–106
HBU (1996) Handbuch der Bodenuntersuchung (HBU), Bodenbeschaffenheit - Bestimmung von organischem Kohlenstoff und Gesamtkohlenstoff nach trockener Verbrennung (Elementaranalyse). In: DIN ISO 10. GmbH, Berlin, p 3.4.1.31.1a
VDLUFA (2012) Methodenbuch- Band I, Die Untersuchung von Böden, 6. VDLUFA - Verlag, Darmstadt
Keidel L, Kammann C, Grünhage L et al (2015) Positive feedback of elevated CO2 on soil respiration in late autumn and winter. Biogeosciences 12:1257–1269. https://doi.org/10.5194/bg-12-1257-2015
Mirman D (2014) Growth curve analysis and visualization using R, 1st ed. Chapman and Hall/CRC in R series, Boca Raton
Geller J, Winn MB, Mahr T, Mirman D (2020) GazeR: a package for processing gaze position and pupil size data. Behav Res Methods 52:2232–2255. https://doi.org/10.3758/s13428-020-01374-8
Bates D, Mächler M, Bolker BM, Walker SC (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–41. https://doi.org/10.18637/jss.v067.i01
Mettel C, Kim Y, Shrestha PM, Liesack W (2010) Extraction of mRNA from soil. Appl Environ Microbiol 76:5995–6000. https://doi.org/10.1128/AEM.03047-09
Lane DJ (1991) 16S/23S rRNA sequencing. In Stackebrandt E, Goodfellow M (eds) Nucleic acid techniques in bacterial systematics. John Wiley and Sons, New York, pp 115–175
Weisburg WG, Barns SM, Pelletier DA, Lane DJ (1991) 16S Ribosomal DNA amplification for phylogenetic study. J Bacteriol 173:697–703
Claesson MJ, O’Sullivan O, Wang Q et al (2009) Comparative analysis of pyrosequencing and a phylogenetic microarray for exploring microbial community structures in the human distal intestine. PLoS ONE 4:e6669. https://doi.org/10.1371/journal.pone.0006669
Engelbrektson A, Kunin V, Wrighton KC et al (2010) Experimental factors affecting PCR-based estimates of microbial species richness and evenness. ISME J 4:642–647. https://doi.org/10.1038/ismej.2009.153
Kaplan H, Ratering S, Felix-Henningsen P, Schnell S (2019) Stability of in situ immobilization of trace metals with different amendments revealed by microbial 13C-labelled wheat root decomposition and efflux-mediated metal resistance of soil bacteria. Sci Total Environ 659:1082–1089. https://doi.org/10.1016/j.scitotenv.2018.12.441
Bolyen E, Rideout JR, Dillon MR et al (2019) Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol 37:852–857. https://doi.org/10.1038/s41587-019-0209-9
Martin M (2011) Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J 17:10
Callahan BJ, McMurdie PJ, Rosen MJ et al (2016) DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods 13:581–583. https://doi.org/10.1038/nmeth.3869
Pedregosa F, Varoquaux G, Gramfort A et al (2011) Scikit-learn: machine learning in python. J Mach Learn Res 12:2825–2830. https://doi.org/10.1145/2786984.2786995
Bokulich NA, Kaehler BD, Rideout JR et al (2018) Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 6:1–17. https://doi.org/10.1186/s40168-018-0470-z
Quast C, Pruesse E, Yilmaz P et al (2013) The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res 41:590–596. https://doi.org/10.1093/nar/gks1219
Glöckner FO, Yilmaz P, Quast C et al (2017) 25 years of serving the community with ribosomal RNA gene reference databases and tools. J Biotechnol 261:169–176. https://doi.org/10.1016/j.jbiotec.2017.06.1198
McMurdie PJ, Holmes S (2013) Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 8:e61217. https://doi.org/10.1371/journal.pone.0061217
Oksanen J, Blanchet FG, Friendly M et al (2018) vegan: Community ecology package. R package version 2.5-7. https://CRAN.R-project.org/package=vegan
Wilcoxon F (1945) Individual comparisons of grouped data by ranking methods. Biometrics Bull 1:80–83. https://doi.org/10.1093/jee/39.2.269
Aitchison J (1982) The statistical analysis of compositional data. J ofthe R Stat Soc Ser B 44:139–177. https://doi.org/10.1007/978-94-009-4109-0
Aitchison J (1986) Book review, XII. Chapman and Hall, London
Fernandes AD, Macklaim JM, Linn TG et al (2013) ANOVA-like differential expression (ALDEx) analysis for mixed population RNA-Seq. PLoS ONE 8:e67019. https://doi.org/10.1371/journal.pone.0067019
Jolliffe IT, Cadima J (2016) Principal component analysis: a review and recent developments subject areas. PhilTransRSocA 374:1–16
Anderson MJ (2001) A new method for non parametric multivariate analysis of variance. Austral Ecol 26:32–46. https://doi.org/10.1111/j.1442-9993.2001.01070.pp.x
Legendre P, Oksanen J, ter Braak CJF (2011) Testing the significance of canonical axes in redundancy analysis. Methods Ecol Evol 2:269–277. https://doi.org/10.1111/j.2041-210X.2010.00078.x
Lahti L, Shetty S (2019) microbiome R package. http://microbiome.github.io
Love MI, Huber W, Anders S (2014) Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 15:550. https://doi.org/10.1186/s13059-014-0550-8
Douglas GM, Maffei VJ, Zaneveld J et al (2019) PICRUSt2 for prediction of metagenome frunctions. Nat Biotechnol 38:669–673. https://doi.org/10.1038/s41587-020-0550-z
Müller C, Rütting T, Abbasi MK et al (2009) Effect of elevated CO2 on soil N dynamics in a temperate grassland soil. Soil Biol Biochem 41:1996–2001. https://doi.org/10.1016/j.soilbio.2009.07.003
Houseley J, Tollervey D (2010) Apparent non-canonical trans-splicing is generated by reverse transcriptase in vitro. PLoS ONE 5:e12271. https://doi.org/10.1371/journal.pone.0012271
Cocquet J, Chong A, Zhang G, Veitia RA (2006) Reverse transcriptase template switching and false alternative transcripts. Genomics 88:127–131. https://doi.org/10.1016/j.ygeno.2005.12.013
Laroche O, Wood SA, Tremblay LA et al (2017) Metabarcoding monitoring analysis: the pros and cons of using co-extracted environmental DNA and RNA data to assess offshore oil production impacts on benthic communities. PeerJ 5:e3347. https://doi.org/10.7717/peerj.3347
Arezi B, Hogrefe HH (2007) Escherichia coli DNA polymerase III ε subunit increases Moloney murine leukemia virus reverse transcriptase fidelity and accuracy of RT-PCR procedures. Anal Biochem 360:84–91. https://doi.org/10.1016/j.ab.2006.10.009
Russo S, Sillmann J, Fischer EM (2015) Top ten European heatwaves since 1950 and their occurrence in the coming decades. Environ Res Lett 10:124003. https://doi.org/10.1088/1748-9326/10/12/124003
Yu Z, Li Y, Wang G et al (2016) Effectiveness of elevated CO2 mediating bacterial communities in the soybean rhizosphere depends on genotypes. Agric Ecosyst Environ 231:229–232. https://doi.org/10.1016/j.agee.2016.06.043
Montealegre CM, Van Kessel C, Blumenthal JM et al (2000) Elevated atmospheric CO2 alters microbial population structure in a pasture ecosystem. Glob Chang Biol 6:475–482. https://doi.org/10.1046/j.1365-2486.2000.00326.x
Lee SH, Kang H (2016) Elevated CO2 causes a change in microbial communities of rhizosphere and bulk soil of salt marsh system. Appl Soil Ecol 108:307–314. https://doi.org/10.1016/j.apsoil.2016.09.009
Song N, Zhang X, Wang F et al (2012) Elevated CO2 increases Cs uptake and alters microbial communities and biomass in the rhizosphere of Phytolacca americana Linn (pokeweed) and Amaranthus cruentus L. (purple amaranth) grown on soils spiked with various levels of Cs. J Environ Radioact 112:29–37. https://doi.org/10.1016/j.jenvrad.2012.03.002
Cheng L, Booker FL, Burkey KO et al (2011) Soil microbial responses to elevated CO2 and O3 in a nitrogen-aggrading agroecosystem. PLoS ONE 6:e21377. https://doi.org/10.1371/journal.pone.0021377
King JS, Hanson PJ, Bernhardt ES et al (2004) A multiyear synthesis of soil respiration responses to elevated atmospheric CO2 from four forest FACE experiments. Glob Chang Biol 10:1027–1042. https://doi.org/10.1111/j.1365-2486.2004.00789.x
Blagodatskaya E, Blagodatsky S, Dorodnikov M, Kuzyakov Y (2010) Elevated atmospheric CO2 increases microbial growth rates in soil: results of three CO2 enrichment experiments. Glob Chang Biol 16:836–848. https://doi.org/10.1111/j.1365-2486.2009.02006.x
Guo H, Hong C, Zheng B et al (2018) Biotechnology for biofuels improving enzymatic digestibility of wheat straw pretreated by a cellulase - free xylanase - secreting Pseudomonas boreopolis G22 with simultaneous production of bioflocculants. Biotechnol Biofuels 11:1–10. https://doi.org/10.1186/s13068-018-1255-0
Maki ML, Idrees A, Tin Leung K, Qin W (2012) Newly isolated and characterized bacteria with great application potential for decomposition of lignocellulosic. J Mol Microbiol Biotechnol 22:156–166. https://doi.org/10.1159/000341107
Trujillo-Cabrera Y, Ponce-Mendoza A, Rivera-Orduña FN, Wang ET (2013) Diverse cellulolytic bacteria isolated from the high humus, alkaline-saline chinampa soils. 63:779–792. https://doi.org/10.1007/s13213-012-0533-5
Zahran HH (2001) Rhizobia from wild legumes: diversity, taxonomy, ecology, nitrogen fixation and biotechnology. J Biotechnol 91:143–153. https://doi.org/10.1016/S0168-1656(01)00342-X
Teixeira H, Rodríguez-Echeverría S (2016) Identification of symbiotic nitrogen-fixing bacteria from three African leguminous trees in Gorongosa National Park. Syst Appl Microbiol 39:350–358. https://doi.org/10.1016/j.syapm.2016.05.004
Masson-Boivin C, Sachs JL (2018) Symbiotic nitrogen fixation by rhizobia — the roots of a success story. Curr Opin Plant Biol 44:7–15. https://doi.org/10.1016/j.pbi.2017.12.001
Li Y, Wu Z, Dong X et al (2019) Variance in bacterial communities, potential bacterial carbon sequestration and nitrogen fixation between light and dark conditions under elevated CO2 in mine tailings. Sci Total Environ 652:234–242. https://doi.org/10.1016/j.scitotenv.2018.10.253
Kester RA, De Boer W, Laanbroek HJ (1997) Production of NO and N2O by pure cultures of nitrifying and denitrifying bacteria during changes in aeration. Appl Environ Microbiol 63:3872–3877
Arai H, Kodama T, Igarashi Y (1999) Effect of nitrogen oxides on expression of the nir and nor genes for denitrification in Pseudomonas aeruginosa. FEMS Microbiol Lett 170:19–24. https://doi.org/10.1016/S0378-1097(98)00517-5
Verbaendert I, Boon N, De Vos P, Heylen K (2011) Denitrification is a common feature among members of the genus Bacillus. Syst Appl Microbiol 34:385–391. https://doi.org/10.1016/j.syapm.2011.02.003
Yang XP, Wang SM, Zhang DW, Zhou LX (2011) Isolation and nitrogen removal characteristics of an aerobic heterotrophic nitrifying-denitrifying bacterium, Bacillus subtilis A1. Bioresour Technol 102:854–862. https://doi.org/10.1016/j.biortech.2010.09.007
Feng L, Jia R, Zeng Z et al (2018) Simultaneous nitrification–denitrification and microbial community profile in an oxygen-limiting intermittent aeration SBBR with biodegradable carriers. Biodegradation 29:473–486. https://doi.org/10.1007/s10532-018-9845-x
Kammann C, Müller C, Grünhage L, Jäger HJ (2008) Elevated CO2 stimulates N2O emissions in permanent grassland. Soil Biol Biochem 40:2194–2205. https://doi.org/10.1016/j.soilbio.2008.04.012
Moser G, Gorenflo A, Brenzinger K et al (2018) Explaining the doubling of N2O emissions under elevated CO2 in the Giessen FACE via in-field 15N tracing. Glob Changs Biol 24:3897–3910. https://doi.org/10.1111/gcb.14136
Acknowledgements
We thank Professor Bernd Honermeier group for their support to perform soil carbon and nitrogen analyses, and we are grateful to our colleagues Rita Geissler-Plaum and Bellinda Schneider for their excellent technical support. For the long-term funding of the Environmental Monitoring and Climate Impact Research Station Linden that enabled the long-term Gi-FACE experiment, we acknowledge the Hessian Agency for Nature Conservation, Environment and Geology (HLNUG).
Funding
Open Access funding enabled and organized by Projekt DEAL. This study was partially funded by the Environmental Monitoring and Climate Impact Research Station Linden that enabled the long-term Gi-FACE experiment and the Hessian Agency for Nature Conservation, Environment and Geology (HLNUG).
Author information
Authors and Affiliations
Contributions
DR conducted experiments, data curation, data analysis, and writing of the manuscript. SR contributed with methodology, review and editing. MC contributed with methodology, review and editing. CM conducted experiments and data curation. GM contributed with data curation, review and editing. MD conducted experiments, data curation, review and editing. Ch M contributed with methodology, review and editing. SS contributed with methodology, review and editing.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rosado-Porto, D., Ratering, S., Cardinale, M. et al. Elevated Atmospheric CO2 Modifies Mostly the Metabolic Active Rhizosphere Soil Microbiome in the Giessen FACE Experiment. Microb Ecol 83, 619–634 (2022). https://doi.org/10.1007/s00248-021-01791-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-021-01791-y