
Abstract
Coral calcification is critical for reef growth and highly dependent on environmental conditions. Yet, little is known about how corals calcify under sub-optimal conditions (e.g., turbid waters, high nutrients, sedimentation) or coral growth in understudied regions such as the Colombian Caribbean. We therefore assessed the calcification and linear extension rates of five coral species across an inshore-to-offshore gradient in the Colombian Caribbean. A suite of environmental variables (temperature, light intensity, visibility, pH, nutrients) measured during the rainy season (May – November 2022) demonstrated more sub-optimal conditions inshore compared to offshore. Across all species, calcification rates were 59% and 37% lower inshore compared to the offshore and midshore sites, respectively. Across all sites, massive corals calcified up to 92% more than branching species but were more susceptible to heat stress and sub-optimal inshore conditions. However, branching species had reduced survival due to extreme climatic events (i.e., bleaching, hurricanes). A comparison with published rates for the wider Caribbean revealed that massive species in the Colombian Caribbean grow up to 11 times more than those in the wider Caribbean while branching species generally have similar growth rates, but this finding may have been influenced by fragment size and/or heat stress. Our findings indicate that present-day environmental conditions, coupled with more frequent extreme climatic events, will favor massive over branching species in midshore areas of the Colombian Caribbean. This suggests a possible shift towards faster calcifying massive species in future coral communities, possibly exacerbating the ongoing regional decline in branching species over the last decades.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Growth rates of hermatypic corals, the primary builders of coral reefs, have a direct influence on reefs’ three-dimensional structure, their calcium carbonate production, and vertical accretion potential (e.g., Cornwall et al. 2021). In recent decades, however, coral reefs have become increasingly threatened by abiotic stressors, including rising temperatures, ocean acidification (OA), nutrient enrichment, and sedimentation, contributing to the global decline of coral reefs (Eddy et al. 2021). Many of these stressors negatively impact coral calcification rates (e.g., Chan and Connolly 2013; Kornder et al. 2018), which in turn contributes to declines in coral reef calcium carbonate production (Cornwall et al. 2021), particularly in the Caribbean where coral cover is low (e.g., Perry et al. 2013).
Predicted changes in the above-mentioned abiotic factors, both at global and local scales, are expected to have both positive and negative effects on coral growth (Kornder et al. 2018). Globally, increasing temperatures generally have a positive net effect on coral growth - until a threshold is reached, usually around + 3 °C above the annual average water temperature (Kornder et al. 2018), above which corals show reduced or suppressed calcification (Wírum et al. 2007; Carricart-Ganivet et al. 2012), can bleach or die (Baker et al. 2008). Additionally, even when corals are visually recovered from bleaching, calcification rates can remain suppressed for many months afterwards (e.g., Bove et al. 2020). In contrast, OA leads to reduced calcification rates in many, though not all, hermatypic corals, impacting calcium carbonate accretion and compromising the ability of corals to contribute to reef formation (Hoegh-Guldberg et al. 2007; Chan and Connolly 2013; Kornder et al. 2018). Additionally, OA leads to a lower seawater aragonite saturation state (Ωarag), ultimately shifting reef calcium carbonate budgets from net growth to net dissolution (Kornder et al. 2018; Cornwall et al. 2021).
Locally, factors such as light intensities, sedimentation, freshwater influxes, nutrients, and pollutants, influence coral calcification rates. For example, high sedimentation directly hinders coral growth by smothering and tissue abrasion (Fabricius 2005; Jones et al. 2016) and indirectly by reducing the light available for photosynthesis (Erftemeijer 2012). Similarly, high nutrient concentrations as well as other land-based pollutants generally lead to declines in coral growth (Nalley et al. 2021, 2023) but moderate nutrient exposure can also have a positive effect up to a threshold, which however is highly dependent on nutrient type, exposure duration, and life stage (D’Angelo and Wiedenmann 2014; Morris et al. 2019). Lastly, coral growth rates can vary greatly between biogeographical regions (Pratchett et al. 2015). Therefore, understanding the sensitivity of coral growth to changing global and local conditions is crucial for predicting future changes in coral calcification rates and reef accretion potential.
Since the 1970s, Caribbean coral reefs have experienced a significant decline in their growth and carbonate production because of coral cover loss and shifts in coral community composition (Gardner et al. 2003; Perry et al. 2013). However, data on coral growth are unevenly distributed among areas within the Caribbean, with sites like Florida and the U.S. Virgin Islands having more extensive datasets compared to underrepresented regions such as Colombia (Manzello et al. 2015; Groves et al. 2018; Rippe et al. 2018). This complicates the accurate estimation of Caribbean-wide coral reef carbonate production and highlights the need for a spatially more comprehensive approach to studying coral growth and addressing existing bias.
Coral reefs in the Colombian Caribbean span over 2800 km2, but most face severe threats from natural disturbances and numerous anthropogenic stressors (Spalding 2001; Rodríguez-Ramírez et al. 2010). Furthermore, the limited presence of hard substrate and the influence of large rivers (Magdalena, Atrato, Sinfi, and Rancheria) along its Caribbean coastline create conditions that are considered sub-optimal for coral growth and, thus, further constrain reef growth potential (Restrepo and Alvarado 2011). However, some coral reefs also exist on the mid shelf or near offshore islands where environmental conditions can differ substantially from the more extreme and sub-optimal conditions of the inshore reefs (Garzón-Ferreira and Díaz 2003). The presence of this strong inshore to offshore environmental gradient provides a unique opportunity to investigate how coral growth responds to various combinations of abiotic stressors, thus providing important insights into future coral growth in a changing ocean. However, research on coral growth in the Colombian Caribbean is still very scarce (e.g., Lizcano-Sandoval et al. 2019). To the best of our knowledge, this is the first study assessing surface area-normalized coral calcification (mg CaCO3 cm− 2 d− 1) in the Colombian Caribbean.
The goal of this study was to (1) determine coral growth rates (both calcification in mg d− 1 cm− 2 and linear/radial extension in cm yr− 1) of five coral species across a strong environmental gradient (inshore to offshore) in the Colombian Caribbean, and (2) conduct a comprehensive site characterization across three sites along this gradient, including an assessment of environmental conditions and benthic composition, to identify site-specific environmental drivers of coral growth. The calcification data obtained were then compared to those reported for the wider Caribbean. It was hypothesized that (1) environmental conditions are more optimal offshore (well-lit and oligotrophic) and more extreme inshore (turbid and nutrient-rich), leading to higher growth rates offshore than inshore; (2) certain key environmental parameters, such as temperature, light, and nutrients, have a more significant impact on coral growth rates than others; and (3) due to the sub-optimal conditions along most of the Colombian Caribbean coast, growth rates are generally lower there compared to the wider Caribbean.
Materials and methods
Study sites
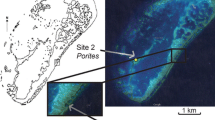
This study was conducted at three sites in the Colombian Caribbean (Fig. 1): (1) Providencia (13°21’04.3’’ N, 81°22’38.2’’ W), an offshore island belonging to an oceanic archipelago 700 km north of Colombia and 430 km east of Nicaragua; (2) Isla San Martín de Pajarales (10°10’35.9’’ N, 75°46’20.2’’ W), part of the coastal archipelago of Islas del Rosario, located about 25 km southwest off the coast of Cartagena de Indias, hereafter referred to as “Rosario” (midshore); and (3) Playa Inca Inca (11°13’01.3” N, 74°14’09.3” W), along the north coast of Colombia and close to the city of Santa Marta, hereafter referred to as “S. Marta” (inshore).
These sites were chosen because they are located along an inshore to offshore environmental gradient. Providencia (Fig. 1a) has a 32 km-long shallow bank reef with clear waters, an annual sea surface temperature oscillating between 27 and 30 °C (Diaz and Geister 1996), constant salinities of 34–36‰ (Garay et al. 1987) and a live coral cover estimated at 18% (Reef Check 2019). Terrestrial runoff is limited to the immediate vicinity of the two major islands of the archipelago (Providencia and San Andrés). The barrier reefs of Rosario (Fig. 1b) are part of the major mid-shelf coral reef system of the Colombian coast (Rodríguez-Ramírez et al. 2010). The distribution and structure of the coral communities are impacted by intense human activities, including blast fishing and unmanaged boat activity (Garzón-Ferreira and Díaz 2003). Additionally, the reefs are threatened by the ongoing dredging of the Canal del Dique which diverts freshwater from the river Magdalena towards the archipelago, carrying nutrients, sediments, and pollutants, thus causing the water to be highly turbid (Restrepo et al. 2016). During the rainy season, strong reductions in salinity (from 31.2 to 25.7) due to freshwater plumes are frequent daily. These reductions can persist for some time and gradually increase again throughout the day (Fig. S1). Both Providencia and Rosario are influenced by large currents, including the warm Panama-Colombia Countercurrent and the Caribbean Current (Andrade et al. 2003; Correa-Ramirez et al. 2019). However, Providencia is more frequently subjected to hurricanes than Rosario (Gómez et al. 2022). Lastly, the inshore reefs of S. Marta are restricted to sheltered bays along the rocky coastline (Fig. 1c) and are heavily influenced by riverine fluxes (Bayraktarov et al. 2014; Bayraktarov and Wild 2014). Additionally, during the major dry season (Dec-March), a quasi-permanent cold-water upwelling can lower sea surface temperature to 21 °C (normally 25–28 °C throughout the year), decreases pH, and increases salinity (∼ 37‰), turbidity, nutrients, and chlorophyll a levels (Gómez Gaspar and Acero P. 2020). Conversely, the major rainy season (Sept-Nov) brings warmer seawaters (up to 31 °C), lower salinity (as low as 24‰), and heavy sediment loads due to intense river runoffs (Rodríguez-Ramírez et al. 2010).

Map of study areas in Colombia and respective experimental coral locations (white circles). (a) Isla de Providencia, with the nursery site along the coral reef edge. The dotted lines represent the Caribbean current; (b) Isla Grande, the main island of the archipelago of Islas del Rosario, and the nursery site located within the CEINER (Centro de Investigación Educación y Recreación). The white solid line represents sediments carried by Canal del Dique and the yellow dotted line is the Panama Colombia Counter current (PCC); (c) The city of S. Marta and the nursery site in the “Acuario y Museo del Mar Fospina SAS”, situated in Playa Inca Inca. The white dotted lines represent the Caribbean Current and the yellow dotted line is the PCC. Map image created using ArcGIS [GIS Software]. Version 10.8. Ocean current data from Restrepo et al. 2012; Moreno-Madriñán et al. 2015 and Correa-Ramirez et al. 2020
Environmental monitoring
Environmental parameters, including seawater temperature, light intensity, visibility, NO2− + NO3−, NH4+, and PO43−, and pH on the total scale, were measured at the three sites to characterize the environmental conditions and their effects on growth rates. Continuous measurements of seawater temperature and light intensity were conducted simultaneously every 15 min, using three EnvLoggers (Electric Blue CRL) and HOBO® Pendant® Temp/Light loggers (MX2202, Onset Computer Corp.), respectively. Specifically, both logger types recorded data from 1 June to 18 November 2022 at Providencia, 21 May to 23 November 2022 at Rosario, and 17 June to 25 November 2022 at S. Marta. Although the HOBOs also recorded the temperature, only the EnvLoggers temperature data were used because of their higher accuracy (± 0.2 °C vs. ± 0.5 °C).
To assess cumulative heat stress (Liu et al. 2005), degree heating weeks (DHW) were calculated from the temperature logger data according to the methodology of the U.S. National Oceanic and Atmospheric Administration (NOAA) as an estimate of thermal stress that corals have experienced in the 12 weeks prior to a certain date, and which serves as a strong predictor of coral bleaching. The maximum monthly mean (MMM) temperature of the NOAA Coral Reef Watch Nicaragua Virtual Station was used to calculate DHW for Providencia while the MMM value of the Colombia Atlantic Virtual Station was used for both Rosario and S. Marta. To account for accumulated heat stress that occurred in the 12 weeks prior to the start of the logger measurements, NOAA’s DHW values from Coral Reef Watch daily global 5 km satellite data (v 3.1) were used for each station.
To convert from lux to photosynthetically active radiation (PAR), the HOBO light loggers were cross-calibrated using a miniPAR® logger (PME) over a period of ∼ 22 h using 1-minute intervals. In the field, both the temperature and the light loggers were attached with cable ties to the table nursery with the experimental coral fragments (see Experimental Design below), facing upward, and cleaned weekly with a toothbrush.
Visibility, as an indicator of turbidity, was evaluated weekly at each site using a 20 cm diameter Secchi Disk. Measurements were conducted at the same time of day (0900 − 11:00 h) when possible (except during rough seas or heavy rain) and always close (5–10 m) to the coral nurseries. Discrete water samples were also collected weekly from the nursery sites in 15 ml falcon tubes, filtered through a CHROMAFIL® Xtra Regenerated Cellulose filter (0.45 μm), and kept frozen until nutrient analyses (NO2− + NO3−, NH4+, and PO43−) were performed at the University of Amsterdam in January 2023 on a San + + System autoanalyzer (Skalar). Additional info about the environmental monitoring can be found in the Supplement. Lastly, seawater pH on the total scale was continuously recorded every 10 min over a period of three days at each site using an optical pH sensor (AquapHOx® Logger, PyroScience GmbH).
Site characterization
To characterize the benthic cover and coral diversity of each site along the environmental gradient, the composition of the benthos was determined according to the photoquadrat method (Preskitt et al. 2004). Six line transects of 10 m each were placed randomly parallel to the shore at depths between 1.5 and 4.5 m. Photos of the 50 × 50 cm quadrats (0.25 m2) were taken every 0.5 m alternating on the right and left side of the tape using a GoPro 10. The transects were placed in areas chosen to be representative of each site. In S. Marta and at Rosario, the transects were placed adjacent (25–50 m) to the nursery sites while in Providencia, due to the absence of a coral reef close to the coral nurseries, the transects were conducted on the closest reef (Three Cay, 13°27’30.15’’N, 81°22’05.65’’O). The digital analysis was performed with the software photoQuad (Trygonis and Sini 2012), and all hard corals were identified to species level or otherwise categorized as unknown. See the Supplement for more information.
Study species
Five coral species with different morphologies were selected for the study: branching Acropora cervicornis (Lamarck, 1816), branching Acropora palmata (Lamarck, 1816), digitate Porites porites (Pallas, 1766), massive Pseudodiploria strigosa (Dana, 1846), and encrusting Porites astreoides (Lamarck, 1816). Acropora cervicornis, A. palmata, and Ps. strigosa were selected because they are listed as “critically endangered” (IUCN 2022). Thus, it is of critical importance to assess the environmental drivers for changes in their growth. On the other hand, Po. astreoides and Po. porites were selected because they are among the most abundant corals in the selected sites (Reyes 2000; Díaz et al. 2000).
Experimental design
The study, conducted between September and November 2022, measured the growth rates of five coral species in the Colombian Caribbean using five parent colonies (genets) and three to five fragments (ramets) per genet. Although the growth experiment was conducted after the rainy season had already begun, all environmental data for the entire rainy season (May to November) was included, given its potential influence on the subsequent growth observed during the study period. Coral colonies were collected randomly by SCUBA divers or while snorkeling, between a depth of 1–5 m and 10–500 m apart from each other to maximize the chance of obtaining genetically different colonies (genets). A total of 290 ramets across all species and sites were obtained (Providencia, n = 80; Rosario, n = 122; S. Marta, n = 88; Table S1), and the size of each fragment varied according to the morphology of the polyp/coral itself (Table S2). Branching corals were fragmented using a chisel and a hammer, while massive corals were fragmented using a Diamond Band Saw (Gryphon Corporation®, CA). The ramets were then maintained in in-situ nurseries for 10–20 days to heal (Table 1).
Physiological measurements
Coral visual health condition
Coral health condition, including bleaching, was visually assessed using the Coral Health Chart (®CoralWatch) which indirectly measures the symbiont density through a six-point scale (Siebeck et al. 2006). A difference in two color-units corresponds to a significant change in symbiont density and chlorophyll a content, and thus a bleached condition (Siebeck et al. 2006).
Calcification rates
After the healing period, coral fragments were retrieved from the nurseries and transported to shore submerged in plastic baskets. The fragments were then weighed using the buoyant weight (BW) method described by Jokiel (1978) using an Ohaus Scout SKX balance (0.1 g accuracy) (OHAUS Corporation, Parsippany, NJ, USA). During weighing, water temperature and salinity were measured at every change in species using a Tetracon® 325 conductivity meter (WTW) to calculate the seawater density. Afterward, fragments of massive species were individually glued to cement pyramids with reef-safe epoxy (Oceans Wonders, USA) or a drop of cyanoacrylate super-glue (Super Pega, INFINITA) and re-weighed to determine the total wet weight. Pyramids were placed in table nurseries (Figure S2a) and branching species were tied with a thin nylon string of negligible weight to floating rope nurseries (Figure S2b).
To determine the dry weight of each coral fragment from the BW method, the following equation was applied:
where Wdry is the dry skeletal weight, Www is the wet weight of the coral in seawater, ρCaCO3 is the density of aragonite (2.93 g/cm3), and ρsw is the density of seawater. At the end of the experimental period, coral fragments were buoyantly weighed again. Before weighing, the pyramids and the nylon strings were cleared of any organisms (mostly algae) with the help of a toothbrush. Calcification rates (g) were calculated from measured surface areas and calculated dry weight according to the equation:
where Wf and Wi are the final and initial dry weights of each fragment respectively, t is number of days between weighing dates, and SAavg is the average between the final and the initial surface area of each fragment, respectively. See the Supplement for more information.
Surface area
To determine calcification rates (mg day− 1 cm− 2), dry weights were normalized to surface areas. The surface area of branching A. cervicornis and Po. porites were estimated through a geometric measurement called “Simple Geometry” (Naumann et al. 2009). The min and max diameter and total length were measured with a caliper (accuracy: ±1 mm). Since fragments of A. palmata were usually characterized by one or more jagged ends, the “foil method” (Marsh 1970), based on weighing a section of aluminum foil carefully shaped and trimmed on a coral fragment, was preferred. Surface areas of massive/encrusting Ps. strigosa, and Po. astreoides were obtained through digital image analysis using the ImageJ software (e.g., Ross et al. 2018). See the Supplement for more information.
Extension rates
To measure extension rates (cm yr− 1) of branching A. cervicornis, a small cable tie was placed at the furthest point from the tip (8–10 cm when possible) of each fragment. The linear extension was then measured as a unidirectional change in branch length. Cable ties did not cause damage to the coral tissue and allowed the fragments to be carefully handled during measurement. For massive species, the radial extension was calculated using ImageJ as the difference between the initial and the final geometric mean diameter (GMD), with GMD = √d1xd2 where d1 is the maximum diameter and d2 is the maximum perpendicular diameter. This approach has already been used in several studies and proved to be a highly accurate estimation of changes in surface area (Guest et al. 2013; Kikuzawa et al. 2018). Similarly, extension rates of Po. porites were calculated using ImageJ as the difference between the initial and the final length of each fragment. Extension rates were measured/calculated at each sampling period.
Statistical analyses
To characterize the coral assemblages at the three sites, a non-metric multidimensional scaling (nMDS) was performed using Bray-Curtis similarity matrix (Bray and Curtis 1957) based on fourth-root transformed data. To test for significant differences in the benthic composition between sites, a one-way PERMANOVA (9999 permutations) was performed. Ecological indices of species richness (S), Shannon’s Index (H’; Shannon and Weaver 1949), and Pielou evenness (J’; Pielou 1966) were then calculated using the DIVERSE function. To investigate the effects of the environmental variables on the benthic composition, a DistLM multivariate analysis (procedure BEST, criterion adjusted R2, and 9999 permutations) was performed (Legendre and Anderson 1999). Further, a Principal Coordinates analysis (PCO) was used to visualize the differences detected by DistLM using the same Bray-Curtis matrix. All benthos-related analyses were conducted using the software PRIMER 7 (Clarke and Gorley 2015).
Differences in environmental variables among sites were assessed using both multivariate and univariate tests in RStudio (version 4.3.0). First, a PERMANOVA (function adonis2, package vegan, version 2.6–6.1; Oksanen et al. 2024) using Euclidean distances with 9999 permutations was conducted on square root transformed and standardized data. Differences in multivariate dispersion among sites were assessed using the betadisper function from the vegan package, utilizing Euclidean distances calculated from square root transformed data. An ANOVA-like analysis was performed on the betadisper results to evaluate the statistical significance of these differences. Second, pairwise mean comparisons using the pairwise.adonis function (package pairwiseAdonis, version 0.4; Arbizu 2024) were performed to compare each site when environmental variables were pooled across sites, with p values adjusted for multiple comparisons using the Benjamini-Hochberg (BH) method. Finally, a series of univariate pairwise Wilcoxon tests were conducted to examine differences in individual environmental variables among sites, with BH correction applied to adjust for multiple comparisons.
To assess the overall fragment survival among the different sites, a logistic regression model (glm function, family binomial, link function logit) was fitted. The model evaluated the influence of site and species on fragment condition (dead or alive), incorporating an interaction term between site and species (Site*Species).
Differences in growth (calcification and extension) rates between species and sites were tested for with linear mixed models (LMMs, function lmer, package lme4; Bates et al. 2014), including genotype as a random factor nested within species. Extension rates were log-transformed to meet the assumption of normality. Both LMMs included calcification and extension rates as response variables, and site and species as predictors. Pairwise comparisons of estimated marginal means were performed using the emmeans package (Lenth et al. 2024), when significant differences between fixed effects and/or interaction were detected, with the Kenward-Roger degrees-of-freedom method and Tukey-adjusted p-values. To identify the main environmental drivers of coral growth and visualize the complex interactions between species and sites, a Principal Component Analysis (PCA) was utilized. Finally, to examine the relationship between calcification and extension, different types of relations were explored, including a linear regression, a polynomial regression, and a generalized additive model (GAM). The generalized additive model (GAM, package mcgv, Wood 2023) ultimately provided a better fit than a traditional linear model and was therefore selected as the GAM’s flexibility in capturing non-linear relationships allowed for a more accurate understanding of the factors influencing coral calcification rates. The residuals of all models were tested for normality and homogeneity of variance using diagnostic plots (i.e., Q–Q plots and fitted versus residuals plots, respectively). All GLM, GAM, and LMM analyses were conducted using RStudio (version 4.3.0).
Results
Site characterization
The composition of the benthos differed significantly across all three sites (PERMANOVA, df = 2, SS = 19181, MS = 9590.6, p < 0.001) (Figure S4a). In Providencia, the benthic community was primarily comprised of algae (31% ± 5.3, mean ± SE, n = 6), CCA (25% ± 7, n = 6), and hard substrate (23.5% ± 1.4, n = 6). In contrast, Rosario’s benthic community was dominated by sand (27% ± 4, n = 6), followed by hard substrate (25.1% ± 5, n = 6) and sponges (16% ± 1.1, n = 6). S. Marta exhibited a dominance of turf algae, comprising nearly 50% ± 5.3 (n = 6), followed by sand (31% ± 6.2, n = 6) and coral rubble (8.7% ± 2.3, n = 6). The highest percentage of live coral cover was observed in Providencia (10.3% ± 1.7, mean ± SE, n = 6), followed by Rosario and S. Marta (6% ± 1.2 and 5.3% ± 1.5, n = 6, respectively). In total, 20 hermatypic coral species were identified across all sites (Table S2), with an unequal distribution among the sites (Figure S4b). The dominant coral species in Providencia and Rosario was O. faveolata (33.2% ± 1.5 and 39.7% ± 1, n = 6, respectively), while Ps. strigosa was most abundant in S. Marta (27.7% ± 0.6, n = 6). Providencia exhibited the highest species richness and Shannon’s diversity, indicating a higher coral diversity and richness compared to Rosario and S. Marta (Table 2). In contrast, Rosario displayed the highest evenness, followed by Providencia and S. Marta, indicating a less uniform distribution of individuals among species in the coastal site (Table 2). See the Supplement for more details.
Environmental conditions
Temperature and heat stress
All environmental parameters measured in this study differed significantly between sites (PERMANOVA, F = 48.24, p < 0.001), including seawater temperature (Table 3). Between June and November 2022, Rosario experienced the warmest average daily temperature of 30.0 °C ± 0.03 SE (n = 176), followed by Providencia with 29.3 °C ± 0.04 SE (n = 171), and S. Marta with 29.0 °C ± 0.09 SE (n = 162) (Fig. 2). Seawater temperatures in S. Marta were influenced by seasonal upwelling during the first part of the study, resulting in a temperature drop as low as 26 °C in July 2022 (Fig. 2c). In Providencia, temperatures showed a consistent increase throughout the rainy season, except for a moderate drop in October 2022 after the passage of Hurricane Julia (Fig. 2a). Conversely, no substantial fluctuations or trends were observed at Rosario, except for a drop of ∼ 2 °C in November 2022, which coincided with the intensification of the rainy season (Fig. 2b). Notably, S. Marta exhibited the highest maximum daily temperature range of 4.1 °C in October 2022, followed by Rosario (2.3 °C) in July 2022 and Providencia (1.3 °C) in September 2022.
Moderate to severe heat stress occurred at all sites during the study period (Fig. 2). Coral fragments in Rosario were found to be at the highest risk of bleaching, as they experienced water temperatures above the local MMM for 184 days from 21 May to 23 November 2022, resulting in a maximum of 18.0 DHW in early August 2022 (Fig. 2b). Notably, heat stress had already reached > 3 DHW by June 2022, when the fragments were placed in their nursery to grow. In Providencia, the water temperatures exceeded the local MMM for 162 days from 1 June to 18 November 2022, resulting in a maximum of 16.4 DHW by the end of November (Fig. 2a). Lastly, coral fragments in S. Marta experienced 10.7 DHW by 25 November 2022, with water temperatures measured above the local MMM for 109 days from 17 June to 25 November 2022 (Fig. 2c).
Photosynthetically active radiation (PAR) and visibility
Two of the three light loggers malfunctioned, resulting in some data loss at Providencia and Rosario. Consequently, light loggers recorded only between 05 September − 18 November 2022 in Providencia (Fig. 2d) and from 01 June to 15 July 2022 at Rosario (Fig. 2e). Thus, light intensity (PAR) could not be compared across all three sites. Nevertheless, the available light data indicated that coral fragments in Providencia experienced significantly higher average light intensities (+ 40.8%) compared to S. Marta during the same period (Providencia: 252.3 ± 15.6 µmol m− 2 s− 1 vs. S. Marta: 148.9 ± 11, mean ± SE; n = 75, Table 3). Similarly, light intensities at Rosario differed significantly from S. Marta between 17 June and 15 July 2022 (Rosario: 338.5 ± 18.7 µmol m− 2 s− 1 vs. S. Marta: 310 ± 20.1, n = 29, Table 3).
Like temperature, light intensity followed a seasonal pattern in S. Marta, being the highest at the beginning of the rainy season and lower at its end. Specifically, the maximum light intensity and the maximum average daily intensity of 1,795 µmol m− 2 s− 1 and 540 µmol m− 2 s− 1, respectively, were recorded in July 2022. In contrast, the lowest average daily light intensity of 5 µmol m− 2 s− 1 was recorded in September 2022 (Fig. 2f). In Providencia, although initial data are missing, light intensities gradually decreased as the rainy season intensified (Fig. 2d). At Rosario, no clear trend was observed due to data loss (Fig. 2e).
Visibility (Secchi depth) exhibited significant differences between all sites (Table 3). Overall, estimated turbidity was, on average, 74 ± 6% and 38 ± 3% higher in S. Marta and Rosario, respectively, compared to Providencia. Providencia consistently had the clearest waters, with an average visibility of 15.4 m ± 1.2 SE (n = 27) which, however, declined gradually during the rainy season and dropped sharply after the passage of hurricane Julia (Fig. 2g). In contrast, Rosario had an average visibility of 9.5 m ± 0.7 SE (n = 25), with occasional spikes up to 15 m (Fig. 2h). S. Marta exhibited the most turbid waters, with an average visibility of 4 m ± 0.3 SE (n = 26) throughout the study period (Fig. 2i).
Seawater pH
Measurements of seawater pHT across a three-day period (Fig. 2) showed that the pHT in Providencia varied greatly (0.27 pH units) between day and night, whereas no clear diel pattern was observed in either Rosario or S. Marta. Specifically, average pHT levels measured in Providencia were significantly higher than those of Rosario and S. Marta (+ 0.08 and + 0.03 pH units, respectively), while the pHT of S. Marta was significantly higher (+ 0.05 pH units) than that of Rosario (Table 3).
Nutrients
Overall, nutrients measured once per week varied significantly among sites (Table 3). Both S. Marta and Providencia showed fluctuating trends for all nutrients whereas Rosario did not exhibit any particular trend except for occasional NH4+ spikes reaching 5 µMol L− 1 (Fig. 2o-q). Rosario had significantly lower NO2− + NO3− concentrations (-66.9%) than Providencia (Rosario: 0.82 ± 0.12 µMol L− 1 vs. Providencia: 2.48 ± 0.27, mean ± SE, n = 21, Table 3) and 64% lower concentrations than S. Marta (2.31 ± 0.32 µMol L− 1, mean ± SE, n = 17, Table 3). There was no significant difference between Providencia and S. Marta (Table 3). NH4+ levels differed significantly between all sites (Table 3). NH4+ concentrations at Rosario were 72.9% and 62.9% lower compared to Providencia and S. Marta, respectively (Rosario: 1.84 ± 0.48 µMol L− 1 vs. Providencia: 6.78 ± 0.76 µMol L− 1, mean ± SE, n = 21; S. Marta: 4.96 ± 1.06 µMol L− 1, mean ± SE, n = 17, Table 3). Lastly, PO43− concentrations in Providencia were 83.8% and 53.2% significantly higher compared to Rosario and S. Marta, respectively (Providencia: 0.62 ± 0.08 µMol L− 1 vs. Rosario: 0.10 ± 0.03, mean ± SE, n = 21; S. Marta: 0.29 ± 0.04, mean ± SE, n = 17). Additionally, PO43− remained consistently at < 1 µMol L− 1 at Rosario and S. Marta, while fluctuating between 0.07 and 1.64 µMol L− 1 in Providencia (Fig. 2q).

Environmental variables measured throughout the rainy season (May – November 2022) in the Colombian Caribbean. (a-c) Seawater temperature and heat stress exposure. Daily average, minimum, maximum temperature, and degree heating weeks (DHW) in (a) Providencia (offshore) from 1 June to 18 November 2022, (b) Rosario (midshore) from 21 May to 23 November 2022 and (c) S. Marta (inshore) from 17 June to 25 November 2022. The black arrow in panel (a) represents the passing of Hurricane Julia in Providencia. The blue dashed lines represent the local maximum monthly mean (MMM) temperature. Red solid lines are weekly cumulative heat stress (= degree heating weeks). (d-f) Photosynthetically Active Radiation (PAR) (µmol m− 2 s− 1) and (g-i) visibility measured using a Secchi disk (m). Daily light intensities in (d) Providencia from 5 September to 18 November 2022, (e) Rosario from 1 June to 15 July 2022, and (f) S. Marta from 17 June to 25 November 2022. Visibility measured once a week in (g) Providencia, (h) Rosario and (i) S. Marta. (l-n) Seawater pH recorded on the total scale at 10 min intervals over a period of three days at (l) Providencia, (m) Rosario, and (n) S. Marta. (o-q) Weekly nutrient concentrations measured in Providencia, Rosario, and S. Marta. (o) NO2− + NO3− (p) NH4+ and (q) PO43−. Note that the Y-axis scaling differs between nutrient types
Physiological measurements
Survival and visual coral health
By the end of the study, overall fragment survival differed significantly between Providencia and Rosario and between Rosario and S. Marta (Table S6), and species-specific patterns related to coral morphology were observed, affecting especially corals in S. Marta. However, their interaction (Site * Species) did not result significant. The highest mortality was observed in Providencia (60%), followed by S. Marta (57%) and Rosario (15%) (Table S7). However, the particularly high mortality in Providencia and S. Marta was due to extreme weather events (Hurricane Julia and a storm, respectively) that caused the detachment and the collapse of the rope nurseries where some branching species were growing. Specifically, 78% of the dead branching fragments in Providencia (35% A. cervicornis and 65% A. palmata) and 93% in S. Marta (100% Po. porites) can be attributed to these events. Excluding the mortality attributed to these extreme weather events, the survival of coral fragments differed significantly only between Rosario and S. Marta and only between some of the species (Table S6). Bleaching primarily affected massive corals in S. Marta (86% of all bleached fragments, Table S7). Once bleached, the fragments quickly died and were covered by turf algae, thus preventing recovery. No bleaching was observed in both Providencia and Rosario (Table S6).
Calcification rates
Calcification rates were significantly affected by the interactive effect of species and site, as well as genotype (Table S8). Pooled across all species, calcification rates at Rosario and S. Marta were significantly lower by 34.9% and 58.8%, respectively, compared to Providencia (Table S8). In addition, substantial variations in calcification rates were observed among the different coral morphologies. In general, massive/encrusting species calcified approximately five times more than branching species at all sites (Fig. 3a). When pooled across all sites, the calcification rate of A. cervicornis was 0.83 mg day− 1 cm− 2 ± 0.1 SE (n = 33), significantly lower compared to Po. astreoides and Ps. strigosa by 94.9% and 91.9%, respectively (Table S9). Similarly, A. palmata calcified 0.61 mg day− 1 cm− 2 ± 0.1 SE (n = 41), which was significantly slower than Po. astreoides and Ps. strigosa by 96.2% and 93.9%, respectively (Table S9). On the other hand, Po. astreoides calcified 16.4 mg day− 1 cm− 2 ± 0.7 SE (n = 22), significantly more than Po. porites by 89.6% and Ps. strigosa by 37.8%. Lastly, Po. porites calcified 1.7 mg day− 1 cm− 2 ± 0.2 SE (n = 25). Although limited due to widespread bleaching/mortality at some sites, pairwise comparisons revealed that Ps. strigosa was the only species with significantly different calcification rates between sites (Table S9). Specifically, Ps. strigosa calcified 51.8% and 45.4% more at Rosario and Providencia, compared to S. Marta.
Extension rates
Linear and radial extension rates differed significantly between species and sites but were not affected by their interaction or genotype (Table S8). When pooled across all species, extension rates at Rosario were 23.7% higher than Providencia and significantly higher (+ 83.2%) compared to S. Marta. In Providencia, extension rates were significantly higher by 78% compared to S. Marta (Fig. 3b). When pooled across sites, A. cervicornis grew the most in terms of linear extension (12.4 cm yr − 1 ± 1.1 SE, n = 33). Porites porites grew 3.31 cm yr − 1 ± 0.6 SE (n = 25). Encrusting Po. astreoides and massive Ps. strigosa grew 1.13 cm yr − 1 ± 0.2 SE (n = 22) and 1.43 cm yr − 1 ± 0.2 SE (n = 52), respectively. Pairwise mean comparisons showed that A. cervicornis grew significantly more than Po. astreoides, Ps. strigosa and Po. porites by 91.1%, 88.7%, and 73.4% respectively, and that Ps. strigosa grew significantly more than Po. astreoides by 21.4%. Porites porites also grew significantly more than Ps. strigosa by 66.7%. (Table S9). As for the interaction of species and sites, pairwise mean comparisons showed that only Ps. strigosa at Rosario had significantly higher extension rates by 50.8% compared to S. Marta (Table S9). The PCA revealed clear clustering of sites and species, with Ps. strigosa being particularly distinct across the three sites (Fig. 3c). Variation in growth along the first principal component (PC1) was primarily driven by temperature, NO2− + NO3−, and NH4+ levels, while the second principal component (PC2) explained variation mainly due to minimum temperature, visibility, and minimum pH (Fig. 3c, Table S10). Lastly, the generalized additive model (GAM) showed a significant correlation between mean extension rates and calcification rates (F = 39.84, p < 0.0001), explaining 76% of the variability in daily calcification rates (Table S11).

Calcification (mg cm-2 d-1) (a) and linear/radial extension rates (cm yr-1) (b) measured in Providencia (September - November 2022), Rosario and S. Marta (October – November 2022). ACERV = A. cervicornis, APALM = A. palmata, PASTR = Po. astreoides, PPOR = Po. porites and PSTRIG = Ps. strigosa. The boxes represent the interquartile range (IQR), with the median indicated by the bold horizontal line inside each box. The whiskers extend to the minimum and maximum values within 1.5 times the IQR, with individual data points beyond the whiskers shown as dots. Boxes identified by two different letters indicate a significant interaction between species and sites (Pairwise mean comparison, p < 0.05; also reported in Table S9), while there is no significant difference if there is no letter. (c) Spatial variation of coral growth metrics (calcification and extension) at the three sites (P = Providencia, R = Rosario, S = S. Marta), illustrated by the first two components (PC1 and PC2) of a principal component analysis on square root-transformed data. Vectors represent the relative contribution of site-specific environmental variables to the total variation (Table S10)
Discussion
This study investigated the effects of a strong environmental gradient (inshore to offshore) on the survival and coral growth rates in five species in the Colombian Caribbean. By comparing abiotic data and coral growth rates across this gradient, it was possible to assess the relative importance of environmental vs. species-specific drivers. We found that complex species and site interactions governed coral growth rates, though extreme climatic events prevented full comparison of all species across all sites. To the best of our knowledge, this is the first study to investigate both calcification and linear extension rates of multiple coral species in the understudied Colombian Caribbean, which represents an ideal location for investigating coral growth across a strong environmental gradient due to the presence of inshore, midshore and offshore reefs and a range of environmental conditions along this gradient. Finally, considering the projected increase in coral reefs facing sub-optimal conditions due to climate change and intensifying local stressors (urbanization, coastal development, etc.), insights from inshore/midshore reef studies gain growing global relevance for the future, highlighting the significance of this work.
Site-specific environmental conditions
The study coincided with an unusually intense rainy season, which significantly impacted environmental conditions at the three study sites and amplified the inshore to offshore environmental gradient. Rainfalls surpassed the three-decade average by 20% (IDEAM 2023) due to a La Niña phase, which is known to lead to prolonged rainfalls (Poveda et al. 2011) and a higher frequency of river plumes in the Colombian Caribbean (Hoyos et al. 2013). The impact of the 2020–2023 La Niña was evident in strong salinity fluctuations and much higher nutrient levels in inshore S. Marta (+ 85% nitrate, + 48% phosphate) and midshore Rosario (+ 85% nitrate, + 57% phosphate) compared to past La Niña-associated rainy seasons (Vivas-Aguas et al. 2012, 2014; Bayraktarov et al. 2014). In offshore Providencia, historical nutrient data are scarce during the La Niña-associated rainy seasons (2008, 2011, 2012) and limited to one measurement per season, making the comparison difficult due to high discrepancies between years (Gavio et al. 2010). These findings confirm that the observed site-specific differences in nutrients suggest a more pronounced La Niña effect on the two coastal sites compared to offshore Providencia.
As expected, visibility conformed to the inshore-offshore gradient, being lowest in S. Marta (inshore) and highest in Providencia (offshore), with Rosario being an intermediate point (midshore) (Fig. 2g-i). The low visibility at S. Marta can be attributed to elevated sedimentation and nutrient input due to riverine runoffs (intensified by the impact of La Niña), compounded by restricted water flow due to its specific topography (western orientation) which leads to accumulation of sediments (Bayraktarov et al. 2014). The generally higher visibility recorded in offshore Providencia temporarily decreased by 55% after the passing of Hurricane Julia in October 2022. Nonetheless, Providencia still experienced significantly higher light intensity (+ 41%) compared to Santa Marta during September-November 2022 (Fig. 2d, f; Table 3). As for Rosario, the measured visibility of ∼ 11 m agrees with recent work (Bula-Meyer 2002; Henao-Castro 2013), and reflects a historical decline compared to the 1970s due to higher frequency and intensity of the Canal del Dique’s plumes (Lonin et al. 2004).
Providencia was the warmest and thermally most stressed site, followed by Rosario and S. Marta (Fig. 2a-c), which is likely the result of the interaction between warmer-than-normal seawater temperatures and the site-specific water currents and bathymetry. Both Providencia and Rosario are influenced by the warm Panama-Colombia Countercurrent (PCC) which probably contributed to the high temperatures and the higher cumulative heat stress (16.4 DHW and 15.1 DHW, respectively) compared to S. Marta (10.7 DHW). In contrast to Providencia and S. Marta, whose sea floors drop off steeply to the atoll/continental slope, Rosario lies in a bay on a much shallower portion of the continental shelf with an average depth of ∼ 35 m. This likely led to the site experiencing severe heat stress (15.1 DHW), with temperatures comparable to past rainy seasons (Severiche et al. 2017) due to limited cool water influxes (Chollett and Mumby 2013). In contrast, the unique combination of the coastal morphology, the bathymetry, and the quasi-permanent upwelling of S. Marta likely contributed to consistently lower thermal stress (10.7 DHW) compared to the other sites (Rueda-Roa et al. 2018; Gómez Gaspar and Acero P. 2020).
Impact of climatic disturbances on coral survival and health
Overall, corals at Providencia showed no bleaching but 60% of all fragments died due to the collapse of the rope nursery after the passing of hurricane Julia in October 2022. Similarly, 57% of all corals at S. Marta died but this was due to an intense tropical storm in November 2022. Conversely, Rosario had no bleaching, and the limited mortality (15%) was mainly due to grazing by juvenile parrotfish (M. Bravo, pers. Obs.). Besides storms, the observed spatial variation in coral survival and their physiological responses to heat stress is likely due to the interacting effects of temperature, turbidity, water flow, sedimentation, and nutrient concentrations.
Massive and encrusting species in S. Marta exhibited high bleaching susceptibility, with up to 28% of fragments being bleached and 53% dead by the end of November 2022, whereas branching species only showed little bleaching (4%) (Table S7). The high bleaching in S. Marta was unexpected for several reasons. First, the average water temperature during the experimental period remained below the local bleaching threshold (29.31 °C ± 0.13 SE vs. 29.69 °C, NOAA) and less cumulative heat stress (which accumulated throughout the rainy season, Fig. 2c) occurred compared to the other sites (10.7 DHW vs. 16.4 DHW at Providencia and 15.1 DHW at Rosario, Fig. 2a, b). Second, the high daily temperature variability (up to 4 °C), known to generally promote coral heat resistance (Safaie et al. 2018), seemingly did not have the expected beneficial effect. Third, massive and encrusting species are generally considered more bleaching resilient than branching species (e.g., Loya et al. 2001; Smith et al. 2013), yet massive Ps. strigosa and encrusting Po. astreoides showed 60% and 100% more bleaching than branching A. palmata. These findings, however, agree with similar responses observed in the Tropical Eastern Pacific in Panama (Manzello 2010) and branching Acropora spp. from both the Caribbean (Manzello et al. 2007; Williams et al. 2017) and the Indo-Pacific (Khen et al. 2023). In contrast, both Po. astreoides and Ps. strigosa also bleached severely in Colombia’s Parque Nacional Natural Tayrona (PNNT, 8–34 km north of S. Marta) during the 2010 and 2011 bleaching events where heat stress reached 9.9 DHW (Bayraktarov et al. 2013; Vega-Sequeda et al. 2017). Potential reasons for the severe bleaching in S. Marta include - but are not limited to - the lack of vertical mixing due to the absence of trade winds during the rainy season (Bayraktarov and Wild 2014) coupled with intense coastal runoffs and nutrient input, both of which have been shown to increase bleaching susceptibility (Wiedenmann et al. 2013; Vega Thurber et al. 2014).
Conversely, despite the higher heat stress experienced by the corals in Providencia (16.4 DHW) and Rosario (15.1 DHW), no bleaching was recorded at these two sites (Table S7). In Providencia, the passing of hurricane Julia caused a decrease of 0.5 °C in daily seawater temperatures and a reduction in light intensity (Fig. 2a, d), likely mitigating (at least partially) the accumulated thermal stress (Wilkinson and Souter 2008; Carrigan and Puotinen 2014). Contrarily to S. Marta, the higher bleaching resistance observed at Rosario may be attributed to the ameliorating effect of turbidity (Sully and van Woesik 2020). Although both sites are turbid, this difference between the two coastal sites may be explained by the different sediment composition and the distance from the source origin: while Rosario is located ∼ 25 km off the Canal del Dique’s mouth, S. Marta is within an enclosed bay, and runoffs are directly adjacent to coral nurseries. Additionally, compared to the coarser sediment at S. Marta (Vargas et al. 2016), the fine Dique silts (Restrepo et al. 2016) typically remain suspended in the surface layer, without directly affecting the coral tissues, creating a periodically stratified water column recognized to be crucial for the survival of some local reefs (e.g., Varadero; López-Londoño et al. 2023). This stratification, coupled with adequate water flow (Jones et al. 2019), can reduce thermal stress by limiting light penetration (Morgan et al. 2017). Finally, turbidity might promote coral heterotrophic plasticity aided by a higher sediment-feeding rate (Anthony and Fabricius 2000) which has been shown to increase bleaching resistance (e.g., Grottoli et al. 2006).
Variation in growth rates among genets and ramets and environmental drivers of growth trends
Complex species and site interactions influenced coral growth rates, though extreme climatic events prevented full comparison of all species across all sites due to bleaching- and hurricane-related mortality. Specifically, only Ps. strigosa could be fully assessed across all sites, while the comparison between the other species was limited to two sites each: A. cervicornis (Providencia and Rosario), A. palmata (Rosario and S. Marta), and Po. astreoides (Providencia and S. Marta). Porites porites only survived at Rosario.
Coral morphology strongly influences growth rates (e.g., González-Barrios et al. 2021). Massive and encrusting corals calcified consistently faster (up to 12 times more) compared to branching species which, however, grew more in terms of linear extension (up to 6 times more) (Fig. 3). While this trend agrees with well-established extension patterns associated with morphology in the Tropical Eastern Pacific in Panama (Manzello 2010), Mexico (Medellín-Maldonado et al. 2016), and in lab experiments (Mallon et al. 2022), branching species typically calcify faster than massive species (Pratchett et al. 2015). Although unusual, our results agree with those of Foster et al. (2014), who found higher growth rates in massive compared to branching species. This pattern was attributed to the sub-lethal effect of temperature stress on the more sensitive branching A. pulchra and Pocillopora damicornis compared to the massive Goniastrea aspera used in their study. Similarly, in the Gulf of Chiriquí, Panama, massive pavonid corals maintained their extension rates despite thermal stress and acidification, though they exhibited less tolerance to repeated bleaching compared to branching pocilloporid corals (Manzello 2010). At Davies Reef in the central Great Barrier Reef (GBR), branching A. muricata experienced a substantial decline in linear extension rates over time, whereas massive Porites corals showed stable growth rates despite rising sea surface temperatures (Anderson et al. 2018). Similarly, the severe heat stress experienced by offshore and midshore corals in our study might have contributed to the observed differences in growth rates. These observations highlight the complex interplay between coral morphology and environmental stressors, suggesting that while branching corals, such as acroporids or pocilloporids, appear to be highly sensitive to thermal stress and possibly cause a reduction in their growth rates, the same conditions could be less detrimental to massive species.
Alternatively, it is possible that fragment size influenced - or at least contributed to - the observed differences in growth rates between massive and branching corals because fragments of massive coral species in this study were substantially smaller than those of branching species (Table S2). Calcification rates of (small) coral fragments can be significantly higher than those of full-size colonies or larger fragments (Forsman et al. 2006; Page et al. 2018; García-Medrano et al. 2023). Despite this, some studies have shown that growth rates of some species, such as massive Porites lobata, are highly variable depending on local environmental conditions (Lough and Barnes 2000; Elizalde-Rendón et al. 2010). In yet other studies, smaller fragments grew more slowly than larger fragments (Lirman et al. 2010, 2014; Forsman et al. 2015; Lizcano-Sandoval et al. 2018; Ishida-Castañeda et al. 2020; Rapuano et al. 2023). In conclusion, it is therefore possible that fragment size contributed to differences in growth rates between massive and branching species, but it is unlikely that it was the sole driver.
Species calcified differently across the environmentally different sites. Porites astreoides calcified significantly faster than all the other species (Table S9), highlighting its weedy life history strategy and recognized opportunistic ability to outcompete other coral species (Green et al. 2008), resulting in a relatively higher abundance of this coral compared to other species (e.g., 37% at Rosario). Acropora cervicornis had similar calcification rates in Providencia and Rosario (Fig. 3a), but remarkably lower (-44%), although not significant, extension rates in Providencia (Fig. 3b), suggesting a potential physiological tradeoff of higher skeletal density at the cost of reduced extension. This tradeoff, consistent with the findings of Lohr and Patterson (2017) for nursery-raised A. cervicornis, has been proposed as an adaptive response to periodic disturbances, such as the higher frequency of hurricanes in Providencia compared to the rest of the Colombian Caribbean (Highsmith et al. 1980). Generally considered a fast-growing species (Darling et al. 2012), the lower calcification rates of A. palmata observed at both Rosario and S. Marta compared to the wider Caribbean (Table S12) suggest an overall suppressed growth in coastal and midshore reef sites. This reduction is congruent with the species’ overall decline at Rosario (between 80 and 90%, (Garzón-Ferreira and Díaz 2003) and S. Marta (up to 30%, García-Urueña and Garzón-Machado 2020) over the last decades.
This study has certain limitations related to measurements of environmental conditions such as the non-continuous (but weekly) nutrient measurements, lack of PAR data at Rosario due to logger malfunctioning, and the limited carbonate chemistry data (pH was measured across only three days throughout the rainy season, see Supplementary Methods). Furthermore, while we measured several key environmental parameters, others that were not measured (e.g., flow, sedimentation), or not included by the models (e.g., light) may also play an important role. For example, the significantly higher light levels at Providencia compared to S. Marta (+ 41%) might have promoted growth by enhancing photosynthesis (Ross et al. 2022), enhancing coral growth offshore (Carricart-Ganivet and Merino-Ibarra 2001; Lough et al. 2016). In contrast, the reduced growth rates of Ps. strigosa found at S. Marta suggest a more negative growth response to the generally more extreme and sub-optimal inshore environmental conditions as this species could be compared across all three sites. Additionally, reduced calcification rates at S. Marta were likely also a consequence of the widespread bleaching (Mendes and Woodley 2002; Baird and Marshall 2002; Leuzinger et al. 2012) and potentially coastal acidification due to increased freshwater and nutrient input (Kornder et al. 2018; Osborne et al. 2022).
Comparative analysis of growth trends across the Caribbean
The comparison of coral growth trends between the Colombian and wider Caribbean (Tables S12, S13) revealed that the literature is strongly biased towards extension rates (up to 3x more studies) because only few studies measured both extension and calcification rates. This is probably because measuring linear extension is comparatively easier, especially for certain species (e.g., A. cervicornis with cable ties and caliper).
Analysis of 31 studies showed that branching species in the Colombian Caribbean generally exhibit lower calcification rates (up to 77% lower for A. palmata and 31% for A. cervicornis) compared to other Caribbean sites, whereas the opposite was the case for massive or encrusting species. For example, A. palmata calcified consistently less at Rosario and S. Marta compared to Florida (Kuffner et al. 2020) and the U.S. Virgin Islands (Gladfelter et al. 1978), while A. cervicornis had calcification rates comparable to those in Florida and across the Mesoamerican reefs (Enochs et al. 2018; González-Barrios and Álvarez-Filip 2018). Notably, Po. astreoides and Ps. strigosa calcified up to eight times more in the Colombian Caribbean than various Caribbean sites (Groves et al. 2018; Martinez et al. 2019; Manzello et al. 2021), while their extension rates exhibited variable trends (Tables S12, S13). The observed differences in growth rates are likely influenced by regional environmental drivers such as sea surface temperature, sedimentation, precipitations, Ωarag and nutrient inputs, reflecting latitudinal variations (Gledhill et al. 2008; Kwiatkowski et al. 2013). The Atlantic Multi-decadal Oscillation (AMO) has also been identified as an important factor impacting coral growth (Lizcano-Sandoval et al. 2019). Future research assessing growth rates across the Caribbean should therefore account for variations in water physicochemical parameters (e.g., temperature, pH, nutrients) and other environmental factors (e.g., currents, disturbance events, and human impacts) for a comprehensive understanding of species-specific responses across different biogeographical regions.
Conclusion
This research provides important new insights into growth patterns and bleaching susceptibility of five coral species in response to the inshore to offshore gradient in the understudied Colombian Caribbean. It was evident that a strong inshore to offshore environmental gradient, mainly dictated by variables we measured such as turbidity and nutrients, and others not included in this study such as sediment composition, played a key role in the observed spatial variation in growth and bleaching responses. Additionally, this gradient was further amplified by an intense La Niña-associated rainy season. However, this study has certain limitations. First, it was mostly limited to the rainy season, thus capturing only a partial seasonal pattern of the environmental conditions at each site, since the dry season - especially in the coastal sites - has very different conditions (e.g., stronger upwelling). Second, due to extreme climatic events, the fragments of several species (especially branching ones) did not survive, reducing the species available for growth comparison across all sites. Third, only limited pH data could be obtained during the experiment due to logistical reasons, yet we acknowledge seawater carbonate chemistry as an important driver of coral growth.
Nonetheless, our findings highlight coral growth rates in the Colombian Caribbean are overall lower midshore/inshore than offshore due to more sub-optimal environmental conditions (e.g., riverine influxes, sedimentation, reduced water flow), which is why these sites should be considered extreme and marginal coral environments (sensu Schoepf et al. 2023). As local and global stressors continue to intensify over the coming decades, this may also lead to a future shift towards faster calcifying massive species (Fig. 3) across nearshore reef habitats that may exacerbate the ongoing regional decline in branching species over the last decades. The extreme environmental conditions at S. Marta, in particular, will likely continue to pose a severe threat to coral (and reef) growth in coastal nearshore areas (Cooper et al. 2008; De’ath et al. 2009). This highlights the need to reduce local stressors to maintain coral growth and increase reef resilience to global stressors (e.g., Donovan et al. 2021). Finally, investigating both parameters related to coral growth - calcification and extension - is crucial to understand growth-related trade-offs under multiple stressors and to increase our understanding of regional disparities in coral growth and future trajectories in this era of climate change.
Data availability
The data and code associated with this study will be made available after publication in the online open access data repository Figshare.
References
Anderson KD, Cantin NE, Heron SF, Lough JM, Pratchett MS (2018) Temporal and taxonomic contrasts in coral growth at Davies Reef, central Great Barrier Reef, Australia. Coral Reefs 37:409–421. https://doi.org/10.1007/s00338-018-1666-1
Andrade CA, Barton ED, Mooers CNK (2003) Evidence for an eastward flow along the Central and South American Caribbean Coast. J Geophys Res 108:2002JC001549. https://doi.org/10.1029/2002JC001549
Anthony KR, Fabricius KE (2000) Shifting roles of heterotrophy and autotrophy in coral energetics under varying turbidity. J Exp Mar Biol Ecol 252:221–253. https://doi.org/10.1016/s0022-0981(00)00237-9
Arbizu PM (2024) pairwiseAdonis: Pairwise multilevel comparison using adonis. R package version 0.4
Baird A, Marshall P (2002) Mortality, growth and reproduction in scleractinian coral following bleaching on the great barrier reef. Mar Ecol Prog Ser 237:133–141. https://doi.org/10.3354/meps237133
Baker AC, Glynn PW, Riegl B (2008) Climate change and coral reef bleaching: an ecological assessment of long-term impacts, recovery trends and future outlook. Estuar Coast Shelf Sci 80:435–471. https://doi.org/10.1016/j.ecss.2008.09.003
Bates D, Mächler M, Bolker B, Walker S (2014) Fitting Linear mixed-effects models using lme4. ArXiv e-prints. https://doi.org/10.18637/jss.v067.i01
Bayraktarov E, Wild C (2014) Spatiotemporal variability of sedimentary organic matter supply and recycling processes in coral reefs of Tayrona National Natural Park, Colombian caribbean. Biogeosciences 11:2977–2990. https://doi.org/10.5194/bg-11-2977-2014
Bayraktarov E, Pizarro V, Eidens C, Wilke T, Wild C (2013) Bleaching susceptibility and Recovery of Colombian Caribbean Corals in response to Water current exposure and Seasonal Upwelling. PLoS ONE 8:e80536. https://doi.org/10.1371/journal.pone.0080536
Bayraktarov E, Pizarro V, Wild C (2014) Spatial and temporal variability of water quality in the coral reefs of Tayrona National Natural Park, Colombian caribbean. Environ Monit Assess 186:3641–3659. https://doi.org/10.1007/s10661-014-3647-3
Bove CB, Umbanhowar J, Castillo KD (2020) Meta-analysis reveals reduced coral calcification under projected ocean warming but not under acidification across the Caribbean Sea. Front Mar Sci 7:127. https://doi.org/10.3389/fmars.2020.00127
Bray JR, Curtis JT (1957) An ordination of the Upland Forest Communities of Southern Wisconsin. Ecol Monogr 27:325–349. https://doi.org/10.2307/1942268
Bula-Meyer G (2002) Perturbación biológica Extrema en Los sistemas coralinos del caribe. Revista De La Acad Colombiana De Ciencias Exactas Físicas Y Naturales 26:105–111
Carricart-Ganivet JP, Merino-Ibarra M (2001) Growth responses of the reef-building coral Montastrea annularis along a gradient of continental influence in the southern Gulf of Mexico. Bull Mar Sci 68:133–146
Carricart-Ganivet JP, Cabanillas-Terán N, Cruz-Ortega I, Blanchon P (2012) Sensitivity of calcification to thermal stress varies among Genera of massive reef-building corals. PLoS ONE 7:e32859. https://doi.org/10.1371/journal.pone.0032859
Carrigan AD, Puotinen M (2014) Tropical cyclone cooling combats region-wide coral bleaching. Glob Change Biol 20:1604–1613. https://doi.org/10.1111/gcb.12541
Chan NCS, Connolly SR (2013) Sensitivity of coral calcification to ocean acidification: a meta-analysis. Glob Change Biol 19:282–290. https://doi.org/10.1111/gcb.12011
Chollett I, Mumby PJ (2013) Reefs of last resort: locating and assessing thermal refugia in the wider Caribbean. Biol Conserv 167:179–186. https://doi.org/10.1016/j.biocon.2013.08.010
Clarke K, Gorley R (2015) PRIMER version 7: User manual/tutorial
Cooper TF, De’ath G, Fabricius KE, Lough JM (2008) Declining coral calcification in massive Porites in two nearshore regions of the northern Great Barrier Reef. Glob Change Biol 14:529–538. https://doi.org/10.1111/j.1365-2486.2007.01520.x
Cornwall CE, Comeau S, Kornder NA, Perry CT, Van Hooidonk R, DeCarlo TM, Pratchett MS, Anderson KD, Browne N, Carpenter R, Diaz-Pulido G, D’Olivo JP, Doo SS, Figueiredo J, Fortunato SAV, Kennedy E, Lantz CA, McCulloch MT, González-Rivero M, Schoepf V, Smithers SG, Lowe RJ (2021) Global declines in coral reef calcium carbonate production under ocean acidification and warming. Proc Natl Acad Sci USA 118:e2015265118. https://doi.org/10.1073/pnas.2015265118
Correa-Ramirez M, Rodriguez-Santana Á, Ricaurte-Villota C, Paramo J (2019) Water masses and mixing processes in the Southern Caribbean upwelling system off Colombia. Ocean Sci Discuss. https://doi.org/10.5194/os-2019-17
Correa-Ramirez M, Rodriguez-Santana Á, Ricaurte-Villota C, Paramo J (2020) The Southern Caribbean upwelling system off Colombia: Water masses and mixing processes. Deep-Sea Res I: Oceanog Res Pap 155:103145. https://doi.org/10.1016/j.dsr.2019.103145
D’Angelo C, Wiedenmann J (2014) Impacts of nutrient enrichment on coral reefs: new perspectives and implications for coastal management and reef survival. COSUST 7:82–93. https://doi.org/10.1016/j.cosust.2013.11.029
Darling ES, Alvarez-Filip L, Oliver TA, McClanahan TR, Côté IM (2012) Evaluating life-history strategies of reef corals from species traits. Ecol Lett 15:1378–1386. https://doi.org/10.1111/j.1461-0248.2012.01861.x
De’ath G, Lough JM, Fabricius KE (2009) Declining coral calcification on the great barrier reef. Science 323:116–119. https://doi.org/10.1126/science.1165283
Diaz J, Geister J (1996) A field guide to the atolls and reefs of the Archipelago of San Andres and Providencia (Colombia)
Díaz JM, Díaz-Pulido G, Sánchez JA (2000) Distribution and structure of the southernmost Caribbean coral reefs: golfo de Urabá, Colombia. Scientia Mar 64:327–336. https://doi.org/10.3989/scimar.2000.64n3327
Donovan MK, Burkepile DE, Kratochwill C, Shlesinger T, Sully S, Oliver TA, Hodgson G, Freiwald J, Van Woesik R (2021) Local conditions magnify coral loss after marine heatwaves. Science 372:977–980. https://doi.org/10.1126/science.abd9464
Eddy T, Lam V, Reygondeau G, Cisneros-Montemayor A, Greer K, Palomares M, Bruno J, Ota Y, Cheung W (2021) Global decline in capacity of coral reefs to provide ecosystem services. One Earth. https://doi.org/10.1016/j.oneear.2021.08.016
Elizalde-Rendón EM, Horta-Puga G, González-Diaz P, Carricart-Ganivet JP (2010) Growth characteristics of the reef-building coral Porites astreoides under different environmental conditions in the Western Atlantic. Coral Reefs 29:607–614. https://doi.org/10.1007/s00338-010-0604-7
Enochs IC, Manzello DP, Jones PJ, Aguilar C, Cohen K, Valentino L, Schopmeyer S, Kolodziej G, Jankulak M, Lirman D (2018) The influence of diel carbonate chemistry fluctuations on the calcification rate of Acropora cervicornis under present day and future acidification conditions. J Exp Mar Bio Ecol 506:135–143. https://doi.org/10.1016/j.jembe.2018.06.007
Erftemeijer P (2012) Environmental impacts of dredging and other sediment disturbances on corals: A review
Fabricius K (2005) Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Mar Pollut Bull 50:125–146. https://doi.org/10.1016/j.marpolbul.2004.11.028
Forsman ZH, Rinkevich B, Hunter CL (2006) Investigating fragment size for culturing reef-building corals (Porites lobata and P. compressa) in ex situ nurseries. Aquaculture 261:89–97. https://doi.org/10.1016/j.aquaculture.2006.06.040
Forsman ZH, Page CA, Toonen RJ, Vaughan D (2015) Growing coral larger and faster: micro-colony-fusion as a strategy for accelerating coral cover. PeerJ 3:e1313. https://doi.org/10.7717/peerj.1313
Foster T, Short JA, Falter JL, Ross C, McCulloch MT (2014) Reduced calcification in western Australian corals during anomalously high summer water temperatures. J Exp Mar Bio Ecol 461:133–143. https://doi.org/10.1016/j.jembe.2014.07.014
Garay J, Castillo F, Andrade C, Aguilera J, Niño L, De La Pava M, López W, Márquez G (1987) Estudio oceanográfico Del área insular y oceánica del caribe colombiano: Archipiélago de San Andrés Y Providencia Y cayos Vecinos, vol 9. Boletín Científico CIOH, pp 1–33
García-Medrano D, López-Pérez A, Guendulain-García S, Valencia-Méndez O, Granja-Fernández R, González-Mendoza T, Torres-Hernández P (2023) Gardening Pocillopora spp. fragments and their potential for rebuilding reef systems in the southern Mexican Pacific. Restor Ecol 31:e14006. https://doi.org/10.1111/rec.14006
García-Urueña R, Garzón-Machado MA (2020) Current status of Acropora palmata and Acropora cervicornis in the Colombian Caribbean: demography, coral cover and condition assessment. Hydrobiologia 847:2141–2153. https://doi.org/10.1007/s10750-020-04238-6
Gardner TA, Côté IM, Gill JA, Grant A, Watkinson AR (2003) Long-term region-wide declines in Caribbean corals. Science 301:958–960. https://doi.org/10.1126/science.1086050
Garzón-Ferreira J, Díaz JM (2003) The Caribbean coral reefs of Colombia. In: Cortés J (ed) Latin American Coral reefs. Elsevier Science, Amsterdam, pp 275–301
Gavio B, Palmer-Cantillo S, Mancera JE (2010) Historical analysis (2000–2005) of the coastal water quality in San Andrés Island, SeaFlower Biosphere Reserve, Caribbean Colombia. Mar Pollut Bull 60:1018–1030. https://doi.org/10.1016/j.marpolbul.2010.01.025
Gladfelter EH, Monahan RK, Gladfelter WB (1978) Growth Rates of Five Reef-Building Corals in the Northeastern Caribbean
Gledhill DK, Wanninkhof R, Millero FJ, Eakin M (2008) Ocean acidification of the Greater Caribbean Region 1996–2006. J Geophys Res Doi. https://doi.org/10.1029/2007JC004629
Gómez C, Millán S, Acosta-Chaparro A, Abril-Howard A, Navas-Camacho R (2022) Preliminary assessment of the impact of Hurricane Iota on coral reefs in Providencia and Santa Catalina Islands: Evaluación preliminar del impacto del huracán iota en arrecifes coralinos de las islas de Providencia y Santa Catalina. Boletín De Investigaciones Marinas Y Costeras 51:193–200. https://doi.org/10.25268/bimc.invemar.2022.51.1.1164
Gómez Gaspar A, Acero PA (2020) Comparación De las surgencias de la Guajira colombiana y del oriente venezolano. Bol Investig Mar Costeras 49:131–172. https://doi.org/10.25268/bimc.invemar.2020.49.2.943
González-Barrios FJ, Álvarez-Filip L (2018) A framework for measuring coral species-specific contribution to reef functioning in the Caribbean. Ecol Indic 95:877–886. https://doi.org/10.1016/j.ecolind.2018.08.038
González-Barrios FJ, Cabral-Tena RA, Alvarez-Filip L (2021) Recovery disparity between coral cover and the physical functionality of reefs with impaired coral assemblages. Glob Change Biol 27:640–651. https://doi.org/10.1111/gcb.15431
Green DH, Edmunds PJ, Carpenter RC (2008) Increasing relative abundance of Porites astreoides on Caribbean reefs mediated by an overall decline in coral cover. Mar Ecol Prog Ser 359:1–10. https://doi.org/10.3354/meps07454
Grottoli AG, Rodrigues LJ, Palardy JE (2006) Heterotrophic plasticity and resilience in bleached corals. Nature 440:1186–1189. https://doi.org/10.1038/nature04565
Groves SH, Holstein DM, Enochs IC, Kolodzeij G, Manzello DP, Brandt ME, Smith TB (2018) Growth rates of Porites astreoides and Orbicella franksi in mesophotic habitats surrounding St. Thomas, US Virgin Islands. Coral Reefs 37:345–354. https://doi.org/10.1007/s00338-018-1660-7
Guest J, Baria MV, Gomez E, Heyward A, Edwards A (2013) Closing the circle: is it feasible to rehabilitate reefs with sexually propagated corals? Coral reefs. https://doi.org/10.1007/s00338-013-1114-1
Henao-Castro A (2013) Efectos De Los aportes Del Canal Del Dique Sobre El reclutamiento de especies de coral en Los arrecifes del Archipiélago Nuestra Señora Del Rosario, área marina protegida. https://doi.org/10.13140/RG.2.2.12191.97448
Highsmith RC, Riggs AC, D’Antonio CM (1980) Survival of hurricane-generated coral fragments and a disturbance model of reef calcification/growth rates. Oecologia 46:322–329. https://doi.org/10.1007/BF00346259
Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ, Caldeira K, Knowlton N, Eakin CM, Iglesias-Prieto R, Muthiga N, Bradbury RH, Dubi A, Hatziolos ME (2007) Coral Reefs under Rapid Climate Change and Ocean Acidification. Science 318:1737–1742. https://doi.org/10.1126/science.1152509
Hoyos N, Escobar J, Restrepo JC, Arango AM, Ortiz JC (2013) Impact of the 2010–2011 La Niña phenomenon in Colombia, South America: the human toll of an extreme weather event. Appl Geogr 39:16–25. https://doi.org/10.1016/j.apgeog.2012.11.018
IDEAM 2023 Plan Nacional de Respuesta Segunda Temporada Lluvias 2022 con condiciones Fenómeno La Niña (Sep – Dic 2022), Boletín Climatológico Mensual - CLIMATOLÓGICO MENSUAL - IDEAM. http://www.ideam.gov.co/web/tiempo-y-clima/climatologico-mensual. Accessed 9 Mar 2024
Ishida-Castañeda J, Pizarro V, López-Victoria M, Zapata FA (2020) Coral reef restoration in the Eastern Tropical Pacific: feasibility of the coral nursery approach. Restor Ecol 28:22–28. https://doi.org/10.1111/rec.13047
IUCN (2022) The IUCN Red List of Threatened Species. In: The IUCN Red List of Threatened Species. https://www.iucnredlist.org/en. Accessed 8 Mar 2024
Jokiel P (1978) Coral growth: buoyant weight technique. pp 529–541
Jones R, Bessell-Browne P, Fisher R, Klonowski W, Slivkoff M (2016) Assessing the impacts of sediments from dredging on corals. Mar Pollut Bull 102:9–29. https://doi.org/10.1016/j.marpolbul.2015.10.049
Jones R, Fisher R, Bessell-Browne P (2019) Sediment deposition and coral smothering. PLoS ONE 14:e0216248. https://doi.org/10.1371/journal.pone.0216248
Khen A, Wall CB, Smith JE (2023) Standardization of in situ coral bleaching measurements highlights the variability in responses across genera, morphologies, and regions. PeerJ 11:e16100. https://doi.org/10.7717/peerj.16100
Kikuzawa YP, Toh TC, Ng CSL, Sam SQ, Taira D, Afiq-Rosli L, Chou LM (2018) Quantifying growth in maricultured corals using photogrammetry. Aquac Res 49:2249–2255. https://doi.org/10.1111/are.13683
Kornder NA, Riegl BM, Figueiredo J (2018) Thresholds and drivers of coral calcification responses to climate change. Glob Change Biol 24:5084–5095. https://doi.org/10.1111/gcb.14431
Kuffner I, Stathakopoulos A, Toth L, Bartlett L (2020) Reestablishing a stepping-stone population of the threatened elkhorn coral Acropora palmata to aid regional recovery. Endang Species Res 43:461–473. https://doi.org/10.3354/esr01083
Kwiatkowski L, Cox PM, Economou T, Halloran PR, Mumby PJ, Booth BBB, Carilli J, Guzman HM (2013) Caribbean coral growth influenced by anthropogenic aerosol emissions. Nat Geosci 6:362–366. https://doi.org/10.1038/ngeo1780
Legendre P, Anderson MJ (1999) Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecol Monogr 69:1–24 10.1890/0012-9615(1999)069[0001:DBRATM]2.0.CO;2
Lenth RV, Bolker B, Buerkner P, Giné-Vázquez I, Herve M, Jung M, Love J, Miguez F, Riebl H, Singmann H (2024) emmeans: Estimated Marginal Means, aka Least-Squares Means
Leuzinger S, Willis BL, Anthony KRN (2012) Energy allocation in a reef coral under varying resource availability. Mar Biol 159:177–186. https://doi.org/10.1007/s00227-011-1797-1
Lirman D, Thyberg T, Herlan J, Hill C, Young-Lahiff C, Schopmeyer S, Huntington B, Santos R, Drury C (2010) Propagation of the threatened staghorn coral Acropora cervicornis: methods to minimize the impacts of fragment collection and maximize production. Coral Reefs 29:729–735. https://doi.org/10.1007/s00338-010-0621-6
Lirman D, Schopmeyer S, Galvan V, Drury C, Baker AC, Baums IB (2014) Growth dynamics of the threatened caribbean Staghorn Coral Acropora cervicornis: influence of Host Genotype, Symbiont identity, colony size, and Environmental setting. PLoS ONE 9:e107253. https://doi.org/10.1371/journal.pone.0107253
Liu G, Strong A, Skirving W, Arzayus F (2005) Overview of NOAA coral reef watch program’s near-real time satellite global coral bleaching monitoring activities. Proc 10th Int Coral Reef Symp 1:1783–1793
Lizcano-Sandoval LD, Londoño-Cruz E, Zapata FA (2018) Growth and survival of Pocillopora damicornis (Scleractinia: Pocilloporidae) coral fragments and their potential for coral reef restoration in the Tropical Eastern Pacific. Mar Biol Res 14:887–897. https://doi.org/10.1080/17451000.2018.1528011
Lizcano-Sandoval LD, Marulanda-Gómez Á, López-Victoria M, Rodriguez-Ramirez A (2019) Climate Change and Atlantic Multidecadal Oscillation as drivers of recent declines in Coral Growth Rates in the Southwestern Caribbean. Front Mar Sci 6:38. https://doi.org/10.3389/fmars.2019.00038
Lohr KE, Patterson JT (2017) Intraspecific variation in phenotype among nursery-reared staghorn coral Acropora cervicornis (Lamarck, 1816). J Exp Mar Biol Ecol 486:87–92. https://doi.org/10.1016/j.jembe.2016.10.005
Lonin S, Parra C, Andrade C, Thomas Y-F (2004) Patrones de la pluma turbia del canal del Dique en la bahía de Cartagena. In: Boletín Científico CIOH. pp 77–89
López-Londoño T, Gómez-Campo K, Galindo-Martínez CT, González-Guerrero LA, Roitman S, Pollock FJ, Pizarro V, López-Victoria M, Medina M, Iglesias-Prieto R (2023) Sobrevivencia y respuestas fisiológicas de corales expuestos a elevada turbidez en El Arrecife Varadero, Caribe Colombiano. Boletín De Investigaciones Marinas Y Costeras 52:135–158. https://doi.org/10.25268/bimc.invemar.2023.52.1.1192
Lough JM, Barnes DJ (2000) Environmental controls on growth of the massive coral Porites. J Exp Mar Bio Ecol 245:225–243. https://doi.org/10.1016/S0022-0981(99)00168-9
Lough JM, Cantin NE, Benthuysen JA, Cooper TF (2016) Environmental drivers of growth in massive Porites corals over 16 degrees of latitude along Australia’s northwest shelf: environmental drivers of massive coral growth. Limnol Oceanogr 61:684–700. https://doi.org/10.1002/lno.10244
Loya Y, Sakai K, Yamazato K, Nakano Y, Sambali H, van Woesik R (2001) Coral bleaching: the winners and the losers. Ecol Lett 4:122–131. https://doi.org/10.1046/j.1461-0248.2001.00203.x
Mallon J, Cyronak T, Hall ER, Banaszak AT, Exton DA, Bass AM (2022) Light-driven dynamics between calcification and production in functionally diverse coral reef calcifiers. Limnol Oceanogr 67:434–449. https://doi.org/10.1002/lno.12002
Manzello DP (2010) Coral growth with thermal stress and ocean acidification: lessons from the eastern tropical Pacific. Coral Reefs 29:749–758. https://doi.org/10.1007/s00338-010-0623-4
Manzello DP, Berkelmans R, Hendee JC (2007) Coral bleaching indices and thresholds for the Florida Reef Tract, Bahamas, and St. Croix, US Virgin Islands. Mar Pollut Bull 54:1923–1931. https://doi.org/10.1016/j.marpolbul.2007.08.009
Manzello DP, Enochs IC, Kolodziej G, Carlton R (2015) Coral growth patterns of Montastraea cavernosa and Porites astreoides in the Florida Keys: the importance of thermal stress and inimical waters. J Exp Mar Bio Ecol 471:198–207. https://doi.org/10.1016/j.jembe.2015.06.010
Manzello DP, Kolodziej G, Kirkland A, Besemer N, Enochs IC (2021) Increasing coral calcification in Orbicella faveolata and pseudodiploria strigosa at Flower Garden Banks, Gulf of Mexico. Coral Reefs 40:1097–1111. https://doi.org/10.1007/s00338-021-02108-8
Marsh JA (1970) Primary Productivity of reef-building calcareous red Algae. Ecology 51:255–263. https://doi.org/10.2307/1933661
Martinez A, Crook ED, Barshis DJ, Potts DC, Rebolledo-Vieyra M, Hernandez L, Paytan A (2019) Species-specific calcification response of Caribbean corals after 2-year transplantation to a low aragonite saturation submarine spring. Proc R Soc B 286:20190572. https://doi.org/10.1098/rspb.2019.0572
Medellín-Maldonado F, Cabral-Tena RA, López-Pérez A, Calderón-Aguilera LE, Norzagaray-López CO, Chapa-Balcorta C, Zepeta-Vilchis RC (2016) Calcification of the main reef-building coral species on the Pacific coast of southern Mexico. Cienc Mar 42:209–225. https://doi.org/10.7773/cm.v42i3.2650
Mendes J, Woodley J (2002) Effect of the 1995–1996 bleaching event on polyp tissue depth, growth, reproduction and skeletal band formation in Montastraea annularis. Mar Ecol Prog Ser 235:93–102. https://doi.org/10.3354/meps235093
Moreno-Madriñán MJ, Rickman DL, Ogashawara I, Irwin DE, Ye J, Al-Hamdan MZ (2015) Using remote sensing to monitor the influence of river discharge on watershed outlets and adjacent coral reefs: Magdalena River and Rosario Islands, Colombia. Int J Appl Earth Obs Geoinf 38:204–215. https://doi.org/10.1016/j.jag.2015.01.008
Morgan KM, Perry CT, Johnson JA, Smithers SG (2017) Nearshore turbid-zone corals exhibit high bleaching tolerance on the great barrier reef following the 2016 Ocean Warming Event. Front mar sci. https://doi.org/10.3389/fmars.2017.00224
Morris L, Voolstra C, Quigley K, Bourne D, Bay L (2019) Nutrient availability and metabolism affect the Stability of Coral–Symbiodiniaceae Symbioses. Trends Microbiol 27:678–689. https://doi.org/10.1016/j.tim.2019.03.004
Nalley EM, Tuttle LJ, Barkman AL, Conklin EE, Wulstein DM, Richmond RH, Donahue MJ (2021) Water quality thresholds for coastal contaminant impacts on corals: a systematic review and meta-analysis. Sci Total Environ 794:148632. https://doi.org/10.1016/j.scitotenv.2021.148632
Nalley EM, Tuttle LJ, Conklin EE, Barkman AL, Wulstein DM, Schmidbauer MC, Donahue MJ (2023) A systematic review and meta-analysis of the direct effects of nutrients on corals. Sci Total Environ 856:159093. https://doi.org/10.1016/j.scitotenv.2022.159093
Naumann MS, Niggl W, Laforsch C, Glaser C, Wild C (2009) Coral surface area quantification–evaluation of established techniques by comparison with computer tomography. Coral Reefs 28:109–117. https://doi.org/10.1007/s00338-008-0459-3
Oksanen J, Simpson GL, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB, Solymos P, Stevens MHH, Szoecs E, Wagner H, Barbour M, Bedward M, Bolker B, Borcard D, Carvalho G, Chirico M, Caceres MD, Durand S, Evangelista HBA, FitzJohn R, Friendly M, Furneaux B, Hannigan G, Hill MO, Lahti L, McGlinn D, Ouellette M-H, Cunha ER, Smith T, Stier A, Braak CJFT, Weedon J (2024) vegan: Community Ecology Package
Osborne E, Hu X, Hall ER, Yates K, Vreeland-Dawson J, Shamberger K, Barbero L, Martin Hernandez-Ayon J, Gomez FA, Hicks T, Xu Y-Y, McCutcheon MR, Acquafredda M, Chapa-Balcorta C, Norzagaray O, Pierrot D, Munoz-Caravaca A, Dobson KL, Williams N, Rabalais N, Dash P (2022) Ocean acidification in the Gulf of Mexico: drivers, impacts, and unknowns. Prog Oceanogr 209:102882. https://doi.org/10.1016/j.pocean.2022.102882
Page CA, Muller EM, Vaughan DE (2018) Microfragmenting for the successful restoration of slow growing massive corals. Ecol Eng 123:86–94. https://doi.org/10.1016/j.ecoleng.2018.08.017
Perry CT, Murphy GN, Kench PS, Smithers SG, Edinger EN, Steneck RS, Mumby PJ (2013) Caribbean-wide decline in carbonate production threatens coral reef growth. Nat Commun 4:1402. https://doi.org/10.1038/ncomms2409
Pielou EC (1966) The measurement of diversity in different types of biological collections. J Theor Biol 13:131–144. https://doi.org/10.1016/0022-5193(66)90013-0
Poveda G, Álvarez DM, Rueda ÓA (2011) Hydro-climatic variability over the Andes of Colombia associated with ENSO: a review of climatic processes and their impact on one of the Earth’s most important biodiversity hotspots. Clim Dyn 36:2233–2249. https://doi.org/10.1007/s00382-010-0931-y
Pratchett M, King K, Hoogenboom M, Widman E, Baird A, Pandolfi J, Edmunds P, Lough J Spatial, temporal and taxonomic variation in coral growth - implications for the structure and function of coral reef ecosystems., Pratchett MS, Anderson KD, Hoogenboom MO, Widman E, Baird AH, Pandolfi JM, Edmunds PJ (2015) & J.M. Lough Oceanography and Marine Biology: An Annual Review, 2015, 53:215–295. Oceanography and marine biology: an annual review 53:215–295
Preskitt LB, Vroom PS, Smith CM (2004) A Rapid Ecological Assessment (REA) quantitative Survey Method for Benthic Algae using photoquadrats with Scuba. Pac Sci 58:201–209. https://doi.org/10.1353/psc.2004.0021
Rapuano H, Shlesinger T, Roth L, Bronstein O, Loya Y (2023) Coming of age: annual onset of coral reproduction is determined by age rather than size. iScience 26:106533. https://doi.org/10.1016/j.isci.2023.106533
Reef Check (2019) Colombia’s Corales de Paz Completes First Survey of Providencia. In: Reef Check. https://www.reefcheck.org/colombia-s-corales-de-paz-completes-first-survey-of-providencia/. Accessed 7 Mar 2024
Restrepo JD, Alvarado EM (2011) Assessing Major Environmental issues in the Caribbean and Pacific coasts of Colombia, South America. Treatise on Estuarine and Coastal Science. Elsevier, pp 289–314
Restrepo JC, Ortíz JC, Maza M, Otero L, Alvarado M, Aguirre J (2012) Estimating fluvial discharge in the caribbean seaboard of Colombia: Magnitude, variability and extreme events. In: Coast Eng Proc. p 44
Restrepo JD, Park E, Aquino S, Latrubesse EM (2016) Coral reefs chronically exposed to river sediment plumes in the southwestern Caribbean: Rosario Islands, Colombia. Sci Total Environ 553:316–329. https://doi.org/10.1016/j.scitotenv.2016.02.140
Reyes J (2000) Lista De Los Corales (Cnidaria: Anthozoa: Scleractinia)De Colombia. Biota Colombiana 1:164–176
Rippe JP, Baumann JH, De Leener DN, Aichelman HE, Friedlander EB, Davies SW, Castillo KD (2018) Corals sustain growth but not skeletal density across the Florida Keys Reef Tract despite ongoing warming. Glob Change Biol 24:5205–5217. https://doi.org/10.1111/gcb.14422
Rodríguez-Ramírez A, Reyes-Nivia MC, Zea S, Navas-Camacho R, Garzón-Ferreira J, Bejarano S, Orozco C (2010) Recent dynamics and condition of coral reefs in the Colombian caribbean. Rev Biol Trop 58(Suppl 1):107–131. https://doi.org/10.15517/rbt.v58i1.20027
Ross CL, Schoepf V, DeCarlo TM, McCulloch MT (2018) Mechanisms and seasonal drivers of calcification in the temperate coral Turbinaria reniformis at its latitudinal limits. Proc R Soc B 285:20180215. https://doi.org/10.1098/rspb.2018.0215
Ross CL, Warnes A, Comeau S, Cornwall CE, Cuttler MVW, Naugle M, McCulloch MT, Schoepf V (2022) Coral calcification mechanisms in a warming ocean and the interactive effects of temperature and light. Commun Earth Environ 3:72. https://doi.org/10.1038/s43247-022-00396-8
Rueda-Roa D, Ezer T, Muller-Karger F (2018) Description and mechanisms of the Mid-year Upwelling in the Southern Caribbean Sea from Remote Sensing and Local Data. JMSE 6:36. https://doi.org/10.3390/jmse6020036
Safaie A, Silbiger NJ, McClanahan TR, Pawlak G, Barshis DJ, Hench JL, Rogers JS, Williams GJ, Davis KA (2018) High frequency temperature variability reduces the risk of coral bleaching. Nat Commun 9:1671. https://doi.org/10.1038/s41467-018-04074-2
Schoepf V, Baumann JH, Barshis DJ, Browne NK, Camp EF, Comeau S, Cornwall CE, Guzmán HM, Riegl B, Rodolfo-Metalpa R, Sommer B (2023) Corals at the edge of environmental limits: a new conceptual framework to re-define marginal and extreme coral communities. Sci Total Environ 884:163688. https://doi.org/10.1016/j.scitotenv.2023.163688
Severiche CA, Baldiris I, Acosta JC, Bedoya EA, Castro I, Pacheco H (2017) Multivariate Analysis of Water Quality in Rosario Islands National Park (Colombia)
Shannon CE, Weaver W (1949) The mathematical theory of communication. University of Illinois Press, Champaign, IL, US
Siebeck UE, Marshall NJ, Klüter A, Hoegh-Guldberg O (2006) Monitoring coral bleaching using a colour reference card. Coral Reefs 25:453–460. https://doi.org/10.1007/s00338-006-0123-8
Smith TB, Brandt ME, Calnan JM, Nemeth RS, Blondeau J, Kadison E, Taylor M, Rothenberger P (2013) Convergent mortality responses of Caribbean coral species to seawater warming. Ecosphere 4:art87. https://doi.org/10.1890/ES13-00107.1
Spalding M (2001) World atlas of coral reefs. University of California Press, Berkeley
Sully S, van Woesik R (2020) Turbid reefs moderate coral bleaching under climate-related temperature stress. Glob Chang Biol 26:1367–1373. https://doi.org/10.1111/gcb.14948
Trygonis V, Sini M (2012) photoQuad: a dedicated seabed image processing software, and a comparative error analysis of four photoquadrat methods. J Exp Mar Bio Ecol 424–425:99–108. https://doi.org/10.1016/j.jembe.2012.04.018
Vargas TVH, Uribe PE, Castellanos AOM, Ríos RCA (2016) Coastal landforms caused by deposition and erosion along the shoreline between Punta Brava and Punta Betín, Santa Marta, 1102 Colombian Caribbean. Rev Acad Colomb Cienc Ex Fis Nat 40:664. https://doi.org/10.18257/raccefyn.387
Vega Sequeda JC, Zea S, Bernal G (2017) Effect of extreme oceanic events in the coral formations of Islas Del Rosario, Colombian caribbean. Cic Ocea 32:25–38. https://doi.org/10.37543/oceanides.v32i1.194
Vega Thurber RL, Burkepile DE, Fuchs C, Shantz AA, McMinds R, Zaneveld JR (2014) Chronic nutrient enrichment increases prevalence and severity of coral disease and bleaching. Glob Change Biol 20:544–554. https://doi.org/10.1111/gcb.12450
Vivas-Aguas LJ, Sánchez J, Betancourt J, Quintero M, Moreno Y, Santana C, Cuadrado I, Ibarra K, Ríos M Obando P, Sánchez D (2014). Diagnóstico y Evaluación de la Calidad de las Aguas Marinas y Costeras del Caribe y Pacifico colombianos. Serie de Publicaciones Periódicas del Invemar No. 4 (2014). Red de vigilancia para la conservación y protección de las aguas marine y costeras de Colombia (REDCAM). Informe técnico 2013. INVEMAR. Santa Marta, p 314
Vivas-Aguas LJ, Espinosa L, Sánchez J, Cadavid B, Bautista P, Quintero M, Betancourt J, Parra JP, Parra LG, Cuadraro I, Ibarra K (2012). Diagnóstico y Evaluación de la Calidad Ambiental Marina en el Caribe y Pacífico colombiano. Red de vigilancia para la conservación y protección de las aguas marine y costeras de Colombia - REDCAM. Informe técnico 2012. INVEMAR. Santa Marta, p 310
Wiedenmann J, D’Angelo C, Smith EG, Hunt AN, Legiret F-E, Postle AD, Achterberg EP (2013) Nutrient enrichment can increase the susceptibility of reef corals to bleaching. Nat Clim Change 3:160–164. https://doi.org/10.1038/nclimate1661
Wilkinson C, Souter D (2008) Status of Caribbean coral reefs after bleaching and hurricanes in 2005
Williams DE, Miller MW, Bright AJ, Pausch RE, Valdivia A (2017) Thermal stress exposure, bleaching response, and mortality in the threatened coral Acropora palmata. Mar Pollut Bull 124:189–197. https://doi.org/10.1016/j.marpolbul.2017.07.001
Wírum FP, Carricart-Ganivet JP, Benson L, Golicher D (2007) Simulation and observations of annual density banding in skeletons of Montastraea (Cnidaria: Scleractinia) growing under thermal stress associated with ocean warming. Limnol Oceanogr 52:2317–2323. https://doi.org/10.4319/lo.2007.52.5.2317
Wood S (2023) mgcv: Mixed GAM Computation Vehicle with Automatic Smoothness Estimation
Acknowledgements
We thank: L Cotrino, A Sanchez Diosa, LF Gaitan, M Sandoval Rivera, A Hernandez, and JC Otero Peréz for assistance during fieldwork, the staff of both the “Oceanario Islas del Rosario” and “Acuario y Museo del Mar Fospina SAS” for logistic support, Pieter Slot for analyzing the nutrient samples, and Pyroscience GmbH for the generous loan of an AquapHOx pH logger via the ICYMARE 2022 BREMERHAVEN award to MB. We acknowledge the kind and crucial collaboration with the team Corales de Paz during field operations in Providencia and as constant support throughout the study.
Funding
This research was partly funded by the Amsterdam University Fund (AUF; www.auf.nl) and the Institute for Biodiversity and Ecosystems Dynamics at the University of Amsterdam (UvA). VS was supported by a MacGillavry Fellowship from UvA and a Vidi grant from the Dutch Research Council (VI.Vidi.203.069).
Author information
Authors and Affiliations
Contributions
MB and VS designed the study and provided materials. MB conducted the fieldwork, collected the data, performed the data analysis, and led the writing of the manuscript. VS edited and reviewed drafts and provided supervision. All authors approved the final manuscript.
Corresponding author
Ethics declarations
Ethical approval
No approval of research ethics committees was required to accomplish the goals of this study because experimental work was conducted with unregulated invertebrate species (scleractinian corals).
Field study permissions
Field research permissions were granted by the organizations undertaking individual projects within the umbrella project “Un Millon de Corales por Colombia” (1MC), complying with all relevant regulations. In Providencia, permission was granted by “Corales de Paz”, in Santa Marta by “Acuario y Museo del Mar Fospina SAS”, in Rosario Islands by “Oceanario Islas del Rosario”. The project is led by the Ministry of Environment and Sustainable Development with the support of important allies such as National Natural Parks, Conservation International Colombia (CI), Corales de Paz, the Institute of Marine and Coastal Research (INVEMAR), the Malpelo Foundation, Regional Autonomous Corporations (Coralina, Cardique, Corpamag, Carsucre, and Corpourabá), schools and diving centers, local NGOs, artisanal fishermen’s organizations, and academia.
Competing interests
The authors declare no conflicts of interest that are relevant to the content of this article.
Additional information
Communicated by Ch. Wild.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bravo, M., Schoepf, V. Growth rates of five coral species across a strong environmental gradient in the Colombian Caribbean. Mar Biol 171, 187 (2024). https://doi.org/10.1007/s00227-024-04511-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-024-04511-5