
Abstract
Background
Hemp (Cannabis sativa) is a crop with a wide range of uses, from the production of fiber and seeds to the secondary metabolites for medicinal purposes. In addition, it is characterized by high biomass yield and the ability to accumulate heavy metals, which makes this plant convenient for phytoremediation purposes. In this study, the effect of applying exogenous biodegradable chelating agents, citric acid (CA) and nitrilotriacetic acid (NTA), to zinc-contaminated soil on zinc (Zn) uptake by two industrial hemp varieties ‘Felina 32’ and ‘Monoica’ was studied. The effect of CA and NTA on available Zn in soils was investigated using an ‘in pot’ experiment under controlled conditions. The effect of both tested compounds on soil microbial activity was simultaneously evaluated.
Results
After the application of NTA at a concentration of 5 mmol L−1, a > threefold increased accumulation of Zn in the above-ground parts was recorded in the ‘Felina 32’ variety. In the ‘Monoica’ variety, the levels of Zn in the above-ground parts were increased > twofold. NTA affected the soil microbiome negatively, causing decreased enzyme activity (in ‘Monoica’ planted soil) and induced respiration (in ‘Monoica’ and especially in ‘Felina 32’ planted soil). On the other hand, CA application did not lead to significantly increased Zn levels in any of the studied hemp varieties. Together with CA’s negative effects on some soil enzymes, CA enhanced urease activity, dehydrogenase and several respiration types for the ‘Felina 32’ variety and exerted less detrimental effect on the soil microbiome. No toxic effects from increased Zn uptake and accumulation in experimental plants were detected, accounting for the unchanged physiological stress markers (levels of photosynthetic pigments and proline in leaves, chlorophyll fluorescence parameters) and selected growth traits of the above-ground organs and root system.
Conclusions
From the studied varieties, ‘Felina 32’ seems to be more suitable for Zn-phytoextraction because of its higher tolerance to increased Zn levels, higher biomass production and Zn accumulation capacity. Our results indicate the potential of using the ‘Felina 32’ variety in NTA-assisted Zn phytoextraction from contaminated soils.
Graphical Abstract

Similar content being viewed by others

Introduction
Zinc (Zn) is a potentially toxic element (PTE), which is on the one hand essential for both plants and animals, but on the other hand, exerts various toxic effects in increased doses. For plants, Zn represents a micronutrient that is a cofactor in a number of metalloenzymes [1, 2], transcription factors [3], and is a structural component of ribosomes and some t-RNA synthetases [4]. Zinc is also important for photosynthesis [5] and maintaining the cell membrane integrity [6]. However, high Zn levels in the soil are phytotoxic, which is visually manifested by growth inhibition and a decrease in the leaf and root system dry matter, and leaf chlorosis and marginal necrosis. The level of reactive oxygen species (ROS) increases, and the content of chlorophyll b decreases. The intensity of photosynthesis, transpiration and stomatal conductance are reduced [7, 8].
A Global Zn concentration survey in United States’, Chinese and Indian uncontaminated soils showed values near to or slightly greater than that in the lithosphere, which implies a slight contamination worldwide [9]. The global world soil pollution dates back to the beginning of the industrial era and may be estimated from the world mine production records for Zn, which have been annually recorded from 1915. Zinc occurrence in the soil is mostly coupled with Cd, as Cd is a main Zn ore by-product [10]. The most severe Zn pollution is naturally localized to abandoned mine workings and surroundings [11], but associated industrial activities from Zn metal production result in the globally varied Zn metalloid spatial distribution in soils [12], including arable soil and pastures, which implies the need to assess regional soil pollution maps and predict the probability of exceeding a critical sanitation threshold for agricultural land use [13]. This task for general health risk estimation comes from global safe food security demands as some edible crops e.g., vegetable cultivated and marketed across the world, exerts significant Zn contamination reported mainly for third world countries [14,15,16]. The risk of emerging toxic Zn soil content may even be aggravated due to the lowest recycling efficiency of Zn (19%) compared to other heavy metals (HMs) [17].
One of the possibilities to reduce the excessive HMs content in polluted soils is applying phytoremediation technologies, specifically phytoextraction, i.e., using plants capable of absorbing HMs from the soil through the root system and accumulating them in above-ground organs that are subsequently harvested and removed from the contaminated area [18,19,20]. In the practical implementation of phytoremediation technologies, an important step is the choice of a suitable plant species for this purpose. Theoretically, HM hyperaccumulators are best suited for this purpose [21, 22] since they are able to grow in heavily contaminated soils and, at the same time, accumulate extremely high amounts of HMs in above-ground organs without phytotoxic effects. These species are characterized by increased uptake, faster translocation into leaves and a higher ability to detoxify HMs [23, 24]. However, the practical use of a number of hyperaccumulators in phytoremediation is hindered by the fact that in many cases they are small, slow-growing plants with low biomass production [25]. Besides wild plants, some field crops, such as sunflower (Helianthus annuus) [26], Jerusalem artichoke (Helianthus tuberosus) [27], barley (Hordeum vulgare) [28] or canola (Brassica napus) [29] have been identified as hyperaccumulators for some heavy metals in recent decades, which have a high potential for use in phytoremediation. However, many fast-growing non-hyperaccumulators with large above-ground biomass production have also been successfully tested as a promising phytoremediation crops, such as giant miscanthus (Miscanthus × giganteus) [30] or poplars (Populus sp.) [31].
Hemp (Cannabis sativa) is a crop that meets some characteristics of plants suitable for phytoremediation purposes, such as rapid growth, a deep root system, high production of above-ground biomass, and relatively low cultivation requirements [32]. In addition, several current works evaluate hemp as a plant capable of absorbing and accumulating certain amounts of HMs [33,34,35].
However, in addition to the lower HMs tolerance, the practical possibilities of using non-hyperaccumulators for phytoremediation purposes are complicated by the fact that HMs loading into xylem and its root-to-shoot translocation is restricted and therefore their subsequent accumulation in the shoot is limited in these plants [36].
The availability of HMs for uptake by plants depends on several factors, e.g., the content of organic matter in the soil, applied fertilizers and especially the pH value of the soil [37, 38]. Analyses show that many HMs, especially in alkaline soils, are bound in insoluble forms [39, 40]. Plants have evolved a number of mechanisms to increase the availability of micronutrients (including HMs) from the soil and facilitate their long-distance transport within the plant. Numerous endogenous chelators participate in the uptake, root-to-shoot translocation and detoxification of HMs. Non-proteinogenic amino acids (phytosiderophores, nicotianamine) and free proteinogenic amino acids (histidine, proline, alanine) are involved in the uptake and long-distance Zn translocation within the plant [41,42,43,44,45]. Carboxylic acids (malic, citric acid) also play an important role in the loading Zn into the xylem, its root-to-shoot translocation [45] and tolerance to increased levels of this metal in leaves [45,46,47].
One of the strategies leading to increasing the solubility and thus the HM bioavailability for phytoextraction is the exogenous application of chelating agents, which form complexes with numerous metal soil contaminants [48,49,50]. Similar to endogenous chelators, exogenously applied synthetic chelators can increase the uptake and root–shoot HM translocation via the transpiration stream [51], which is one of the basic prerequisites for successful phytoextraction.
A number of these compounds, especially aminopolycarboxylic acids, such as ethylenediaminetetraacetic acid (EDTA), ethylene glycol-bis-(2-aminoethylether)-tetraacetic acid (EGTA), diethylenetriaminepentaacetic acid (DTPA), hydroxyethylenediaminetetraacetic acid (HEDTA), etc., have been tested for phytoremediation purposes [52]. Compared to natural chelators, metal complexes with synthetic chelators are characterized by higher stability, in particular complexes with DTPA and especially EDTA [53], which is among the most effective synthetic chelators for phytoextraction purposes [54]. However, the wider use of some synthetic chelating agents such as EDTA may be associated with environmental risks e.g., low biodegradability, risk of groundwater contamination, phytotoxicity, negative impact on soil quality and soil microorganisms [52, 55, 56]. The biodegradability of the given substance therefore plays an important role in the choice of chelating agents. This criterion is met by some synthetic aminopolycarboxylic acids, while maintaining high chelation efficiency, for example nitrilotriacetic acid (NTA) and ethylenediaminedisuccinic acid (EDDS) [51, 57], natural low-molecular carboxylic acids (citric, tartaric, malic, gluconic acid) or the amino acids (proline, histidine and glycine) [52, 58,59,60].
As mentioned above, hemp represents a plant with phytoremediation potential, but so far, very few studies have tested the suitability of industrial hemp varieties for environmentally friendly chelating agents assisted phytoextraction [61, 62].
Our study aimed to verify whether applying selected biodegradable chelating agents can increase the uptake of Zn from the soil and its accumulation in the above-ground parts in two industrial hemp varieties approved for field cultivation in the Czech Republic. Another goal was to assess whether potentially increased levels of Zn accumulated in plants could have a phytotoxic effect and cause the inhibition of above-ground or root system growth and thus in consequence decrease biomass production. Last but not least, the subject of the study was also to assess the effect of tested chelating compound application on the soil’s microbial activity. For overall simplification, the experiments were performed on young, 4-week-old hemp plants in the vegetative phase of development. The novelty of this study is to demonstrate the enhanced phytoextraction by two industrial hemp varieties assisted by CA and NTA and reveal the underlying mechanism behind chelating agents-assisted phytoextraction.
Materials and methods
Soil
Common topsoil (classified as haplic chernozem according to WRB [63]) was obtained from the site in Brno Černovice (49° 16′ 37.3″ N, 16° 52′ 37,6″ E). For experimental purposes, the soil was dried at a room temperature and homogenized. The Zn content was adjusted by exogenous zinc sulphate application. 240 mL of an aqueous solution (9.29 g L−1 ZnSO4·7H2O) was applied to 1 kg of dry soil (particle size < 4 mm). Then, the artificially contaminated soil was regularly moistened and left at 20–25 °C for 10 days. To adjust the physical properties, the soil was subsequently mixed with silica sand in a ratio of 3:1 (soil: sand) and thoroughly homogenized.
The total Zn content in the soil extract was an average of 320 µg g−1 by aqua regia. The average content of available plant nutrients and cation exchange capacity in the two prepared soil samples were determined using Mehlich III extraction [64]. The average pH value was measured with a mixture of soil and 0.01 M CaCl2 (1:2.5, w/v) (Table 1.).
Plant material and cultivation conditions
Industrial hemp (Cannabis sativa L.) varieties ‘Felina 32’ and ‘Monoica’ (seeds were provided by Agritec Plant Research Ltd., CZ) were used for the experiments. Both varieties were chosen on the basis of previous experiments by Macečková et al. [65] for their increased ability to accumulate Zn in biomass. The seeds were sown directly into pots (10 × 10 × 11 cm) with a total soil volume of 450 g soil/pot. The plants were cultivated under controlled conditions. The temperature was 23/18 °C day/night, and illumination was provided with a 600 W high-pressure sodium lamp, PPFD 280 µmol m−2 s−1, photoperiod 20/4 h. The plants were watered regularly three times a week, with tap water in a volume of 50 mL per pot.
Three weeks after sowing, nitrilotriacetic acid (5 mmol L−1) and citric acid (10 mmol L−1) were applied in the form of an aqueous solution in a volume of 100 mL per pot. Concentrations of both substances were chosen on the basis of our previous experiments, as the highest applicable concentrations not causing phytotoxic effects in experimental plants. After another two weeks of cultivation, the plants were analyzed (Table 2).
Growth parameters
The following growth parameters were determined to evaluate the effect of experimental treatments on the growth of plants: total height (measured from the soil surface to the top of apical bud), dry weight of the root system and above-ground parts (the samples were dried at 80 °C to constant weight). The root system was scanned using an EPSON perfection V700 scanner and analyzed by Win RHIZO software (Regent Instruments Inc., CAN) to determine the total length, area and volume.
Determining zinc content in the root system and aerial parts by inductively coupled plasma mass spectrometry (ICP-MS)
Before the ICP-MS analysis, the plant samples were dried at 80 °C to constant weight and homogenized using a vibratory ball mill Fritsch PULVERISETTE 0 (Fritsch GmbH, GE). The samples were digested using a mixture of 4 mL of concentrated nitric acid and 1 mL of hydrogen peroxide in the MLS1200 Mega (Milestone, ITA) closed vessel digestion unit, according to the general power-controlled digestion program for biological samples [66]. The resulting digests were transferred into 25 mL volumetric flasks and diluted with ultrapure water to 25 mL. The total Zn content was determined by an inductively coupled plasma mass spectrometer (ICP-MS) (Agilent ORS–ICP-MS 7700 ×, Agilent Technologies, JPN). The ICP-MS method was previously fully validated, and the quality and reliability of the results was ensured by regular analysis of the independent QC sample. Chemicals and reagents used for analysis and ICP-MS method validation are described in the Additional file 1.
Calculation of translocation and bioconcentration factors
The translocation factor (TF) was calculated as the ratio between the Zn concentration in the shoot (µg g−1) and Zn concentration in the root system (µg g−1) [67]. Similarly, the bioconcentration factor (BCF) expresses the ratio between the Zn content in the roots (root BCF) or in the shoot (shoot BCF) and the Zn content in the soil, as described in Usman et al. [68].
Determining photosynthetic parameters and proline content
A handheld FP100 fluorometer Fluor Pen (Photon Systems Instruments /PSI/, CZ) with f-pulse 30%, F-pulse 70% and A-pulse 30% settings was used to measure fast chlorophyll fluorescence induction curves (OJIP). Data were processed by original PSI software and then graphically edited in MS Excel. Before the measurement, the experimental plants were kept in darkness for 15 min to ensure photosynthetic electron transport relaxation. For the measurement, fully expanded 5th node leaves were chosen in ‘Felina 32’ or 4th node in ‘Monoica’. Parameters Fv/Fm, Vj and Vi were automatically calculated by PSI original software. For photosynthetic pigments and proline analyzes, plant material samples were lyophilized and homogenized using a vibratory ball mill Fritsch PULVERISETTE 0 (Fritsch GmbH, GE). Homogenized samples were stored at – 20 °C prior to analysis. To determine pigment content, 50 mg of homogenized plant material was extracted three times with acetone, the extract was filtered using a Morton vacuum filter system and brought to a final volume of 25 mL. The content of chlorophyll a and b in the extract was determined spectrophotometrically (Spectronic 20 Genesys, Spectrum Chemical Mfg., USA). The absorbance of the extract was measured at 662 and 645 nm. The content of pigments was calculated according to Lichtenthaler and Buschmann [69]. The content of proline was analyzed from an amount of 50 mg of lyophilized plant material homogenized and extracted in 40% ethanol at 4 °C overnight. The samples were then centrifuged at 30,000×g for 5 min. The total proline content in the supernatant was determined spectrophotometrically after reacting with ninhydrin in 60% acetic acid and 20% ethanol, as described in Carillo and Gibon [70].
Determination of soil microbial properties
Soil samples were transported immediately to the laboratory, homogenized, sieved to fine earth (≤ 2 mm fraction) and subsampled for further analyses. Fresh fine samples were stored at 4 °C and analyzed for: (I) dehydrogenase activity (DHA) using 2,3,5-triphenyltetrazolium chloride (TTC) method [71]; (II) soil basal respiration (BR) and respiration induced by d-glucose (Glc-SIR), protocatechuic acid (Pro-SIR), D-trehalose (Tre-SIR), and d-mannose (Man-SIR) according to Campbell et al [72] using MicroResp® device (The James Hutton Institute, SCO). Freeze-dried samples were used for the following enzyme activity assays: β-glucosidase (GLU), n-acetyl-β-d-glucosaminidase (NAG), arylsulfatase (ARS), phosphatase (Phos), urease (Ure) according to (ISO 20130:2018 [73]).
Statistical evaluation
Data processing and statistical analyses were performed using freely available software R, version 3.6.1. [74]. Principal component analysis (PCA) and one-way variance analysis (ANOVA) type I (sequential) sum of squares at 5% significance level [75] were used to characterize the relationship between the variants and selected soil properties. Tukey’s HSD (honestly significant difference) test and “variant contrast” were used to calculate factor level means for each variant and detect a statistically significant difference among factor level means. The results were also graphically presented with Rohlf’s biplot for standardized PCA. Pearson correlation analysis was performed to measure the linear dependence between soil properties and this dependence was determined at 3 different levels of statistical significance: at p (p value) ≤ 0.05 (marked by *), p ≤ 0.01 (marked by **), p ≤ 0.001 (marked by ***). Pearson’s correlation coefficient was interpreted as follows: 0.0 < r < 0.3 (negligible correlation), 0.3 < r < 0.5 (low correlation), 0.5 < r < 0.7 (moderate correlation) and 0.7 < r < 0.9 (high correlation), 0.9 < r < 1.0 (very high correlation) [76]. The ANOVA table for all experiments is included in the Additional file 1 (Additional file 1: Table S5).
Results and discussion
Evaluation of the possible toxic effects of chelating agent application on the growth of experimental plants
Concentration ranges of 1–10 mmol L−1 for NTA and 5–15 mmol L−1 for CA were tested in preliminary experiments using non-contaminated common topsoil. Based on the results of these experiments, concentrations 5 mmol L−1 of NTA and 10 mmol L−1 of CA were chosen for subsequent studies as non-toxic for the tested plants, not causing significant (p ≤ 0.05) inhibition of above-ground parts or the root system growth (data not shown). This concentration of applied NTA appears to be optimal for soil application as it is well tolerated by a number of plant species and results in a significant increase in HMs uptake. Nanthavong and Sampanpanish [77] found up to 200 µg of NTA g−1 of soil as a non-toxic dose in pot experiments with Mimosa pudica, which approximately corresponds to the dose applied in our experiments. Hart et al. [78] tested the NTA concentration range of 1–10 mM in experiments on lead uptake by switchgrass (Panicum virgatum L.). An increase in NTA concentration from 1 to 5 mM was found to increase Pb accumulation in leaves by almost 800%, while a further increase in concentration from 5 to 10 mM no longer resulted in a significant increase in leaf Pb levels. In addition, a toxic effect was already observed at a concentration of 10 mM.
The potential impact of increased biologically available Zn levels on the growth of experimental plants was assessed based on the analysis of selected root system and above-ground parts’ growth parameters.
Growth parameters
The applied doses of NTA and CA did not result in either a significant (p ≤ 0.05) reduction or increase in aboveground biomass production and plant height in any of the tested varieties, as amended variants showed compared to an unamended control (Fig. 1a, b). This observation, done under the chosen degree of Zn at the level 320 µg g−1 soil contamination is important with respect to the reported standardized maximum permissible concentrations (MPC) for Zn in the Czech Republic and worldwide. The average Zn content in the tested soil was almost 3 times higher than the preventive value of 120 µg g−1 for Zn content in common soils in the Czech Republic set by Decree No. 153/2016 Coll. [79]. Similar MPC value limitations are also applied in other EU countries such as the Netherlands (MPC 160 mg kg−1) [80]. Moreover, this concentration (320 µg g−1) is well tolerated by hemp plants. In the pot experiments, ‘Felina 32’ variety tolerates up to 1000 µg g−1Zn with no significant (p ≤ 0.05) inhibition of shoot growth parameters and only a moderate inhibition of the root system (data not shown). Similarly, Shi and Cai [81] reported that hemp grows well with a Zn soil content below 400–800 µg g−1. Hemp together with flax and rapeseed was considered to be the most tolerant to elevated Zn levels in the soil, when eight various oil crops were tested.

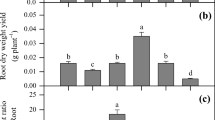
Influence of NTA or CA treatment on growth parameters of plants in experimental variants (Table 2): height of plants (a), dry weight of the aerial parts (b) and the root system (c), total length of the root system (d). Mean values in columns marked with same letters are not statistically significantly different (Tukey’s test, p ≤ 0.05), error bars = standard deviation
Overall, a higher production of above-ground biomass was recorded in the ‘Felina 32’ variety in all experimental variants compared to 039 ‘Monoica’. Since sufficient biomass production of above-ground parts is a prerequisite for successful HMs phytoextraction from soil, this is an important finding from the point of view of potential phytoremediation use. From the root system parameters, CA treatment had a significant (p ≤ 0.05) inhibitory effect on the total length of the root system in ‘Felina 32’ (Fig. 1d). However, the total root dry weight was similar in both varieties in all experimental variants regardless of the application of chelating agents (Fig. 1c), but CA treatment caused high variability in root dry weight values of single plants. Similarly, Li et al. [82] reported a decrease in total root length, volume and diameter in Sedum alfredii grown in Cd and Zn contaminated soil after applying 10 mM CA. In contrast, Muhammad et al. [83] proved an increase in root dry weight in Typha angustifolia after 2.5 and 5 mM CA application to Cd, Cu, Pb, and Cr contaminated soil.
In our experiments, no significant (p ≤ 0.05) differences between experimental variants were found even in the root:shoot value (Additional file 1: Table S3), which is considered a highly representative marker of environmental stress in plants [84].
For a more detailed insight into the effects of chelate mobilized Zn on the studied plants, selected HM stress biochemical markers were also evaluated, such as the content of photosynthetic pigments, selected parameters of chlorophyll fluorescence and levels of proline.
Photosynthetic characteristics, levels of photosynthetic pigments
One of the important processes in plants adversely affected by increased HMs levels is photosynthesis [85]. The appearance of the leaf chloroses is usually the most visible marker indicating a disruption in photosynthesis. It is associated with changes in the photosynthetic pigments content. Changes in chlorophyll content in high doses Zn-treated tomato plants were presented by Cherif et al. [86]. Their data show that chlorophyll a pool is more sensitive to excess zinc exposure than chlorophyll b or carotenoids. As a result, a significant reduction in the chlorophyll a/b ratio was observed. In correlation with the decreased chlorophyll a content, was also a decrease in the chlorophyll fluorescence parameters, which indicated Zn’s interaction with the photosystem II (PSII) reaction center of Zn-treated tomato plants. Nevertheless, in our experiments, no significant (p ≤ 0.05) differences were observed in the chlorophyll a and b levels between the two tested hemp varieties and the variants treated with different chelating agents (Fig. 2a, b). The only significant (p ≤ 0.05) difference reported was in the chlorophyll a/b ratio (Fig. 2c) between the ‘Felina 32’ variety treated with CA and ‘Monoica’ variety treated with NTA (ratio lower by almost 10%). Measuring chlorophyll fluorescence—OJIP test—used to assess possible changes in the photosynthetic apparatus generates the curves reporting on the function and capacity of the PSII, presented in Fig. 3. For better clarity, the points in curves represent the arithmetic mean with no statistical evaluation. Statistical analysis of selected parameters is presented in Table 3.

Influence of NTA or CA treatment on chlorophyll a (a) content, chlorophyll b (b) and chlorophyll a/b ratio (c) in plants (Table 2). Mean values in columns marked with same letters are not statistically significantly different (Tukey’s test, p ≤ 0.05), error bars = standard deviation

OJIP curves of ‘Felina 32’ (a) and 'Monoica’ (b) variety plants (Table 2). The presented curves reported average measured data values without the statistic for better curve shape clarity. Point O indicates the origin of the OJIP curve and corresponds to minimal fluorescence Fo, part O–J indicates changes in primary electron acceptor (QA) reduction, J–I indicates changes in reduction of secondary electron acceptor (QB, plastoquinone and cytochrome), point P corresponds with maximum fluorescence value Fm
The obtained OJIP curve shapes (Fig. 3) from all experimental variants (regardless of genotype or treatment) point to the normal function of the photosynthetic primary processes. However, the OJIP curves for the ‘Monoica’ variety show generally lower levels of maximal and variable chlorophyll fluorescence than ‘Felina 32’. A more accurate evaluation of the chlorophyll fluorescence parameters also revealed significant differences between both hemp varieties, including variants of the same treatment. The ‘Monoica’ variety treated with CA or NTA (as compared to ‘Felina 32’) showed significantly (p ≤ 0.05) lower values of Fv/Fm. The negative effect of increased Zn levels on photosynthesis was also observed by Huang et al. [87] in tomato plants. They reported that negative effect of Zn could probably be caused by Zn accumulation in leaves, which led to chloroplast function disruption. Reduction of Fv/Fm can be caused by the higher production of ROS under HMs stress, which impairs the chloroplast membrane integrity and disrupts the electron transport chain. On the contrary, no changes in the Fv/Fm parameter compared with the control were found in Zn-treated moss Fontinalis antipyretica (25–100 µM concentration [88]). Ciscato et al. [89] present only slight or no Zn effect (50 µM concentration) with decreasing Fv/Fm in their study with Phaseolus vulgaris. On the other hand, they observed an increase in the Vj part of the OJIP curve with partial recovery during their long-term experiment. In our study, the lower Zn tolerance of ‘Monoica’ could also be indicated by the detected higher average levels of proline in this variety (Fig. 4a, b), even though the total detected Zn amount was lower than in the ‘Felina 32’ variety (Fig. 4c, d). This variety showed no difference in the Fv/Fm parameter between chelating agent-treatments.

Influence of NTA or CA treatment on content of proline in root system (a) and above-ground parts (b) and total zinc content in root system (c) and above-ground parts (d) of plants in experimental variants (Table 2). Mean values in columns marked with same letters are not statistically significantly different (Tukey’s test, p ≤ 0.05), error bars = standard deviation
Generally, our observation suggests that the activity of the PSII was quite well preserved, at least under these experimental conditions. Because photosynthesis is the basic metabolic process of plant biomass production, the results reported the almost unchanged measured photosynthetic parameters, which agree with the biomass production data presented in Fig. 1. Based on the evaluation of analysed parameters, ‘Felina 32’ also seems to be more tolerant to Zn excess than ‘Monoica’.
Content of proline
An increase in the content of free proline (Pro) in plant tissues is a general phenomenon accompanying the effect of various abiotic stresses on the plant, such as osmotic stress, drought stress etc. However, no significant (p < 0.05) differences were detected in Pro content between the studied hemp genotypes and variants treated with both chelating agents (and the control), neither in shoot or root (Fig. 4a, b). In Scenedesmus sp. algae, the intracellular Pro level increased with increasing Zn concentration in the culture medium. Pro probably plays a role in detoxifying ROS, especially hydroxyl radicals, the production of which is induced under HM stress. Exogenous Pro treatment reduces Cu-induced oxidative stress, but only weakly Zn-induced stress [90].
In Solanum nigrum, an increase in proline content in the roots due to phytotoxic levels of Cu has been reported. On the contrary, no significant differences were observed in the stem [91]. On the other hand, a significant increase in the level of proline in leaves induced by HMs was recorded in a number of other plants, e.g. in beans under the influence of Hg, Cd, Cu or Pb [92], or in pepper under the influence of Pb and Cd [93].
When comparing average shoot Pro content values in variants FN and MN, as well as FC and MC, it suggests that (on average) Pro levels in the ‘Monoica’ variety increased as a response to the higher adverse effect of over-accessed Zn in this hemp variety compared to ‘Felina 32’ as presumed in the previous section. Nevertheless, high variability in Pro content values between measurements from single plants prevents assessment of a significant effect. Moreover, higher average Pro levels were recorded in the root system than the shoot (Fig. 4a, b). This may partly reflect differences in the varieties' HM stress sensitivity, however, as phytotoxic Zn levels in plants were apparently not reached, or were only slightly exceeded, further study would be needed to compare the sensitivity of the two varieties to HM stress. Insignificant (p < 0.05) differences among experimental variants indicated low level stress in chelating agent-treated plants. However, a strong negative correlation between Pro levels in the root system and some growth parameters—specifically total plant height (r = − 0.6, p ≤ 0.01), root system length (r = − 0.64, p ≤ 0.01) and root system weight (r = − 0.48, p ≤ 0.05) was demonstrated. A negative correlation was also revealed between the Pro content in the roots and shoots (r = − 0.5, p ≤ 0.05) and between Proline root and chlorophyll a/b ratio (r = − 0.52, p ≤ 0.05) (Fig. 6). This suggests that in hemp, similar to numerous other plant species, Pro may play a certain role in the response to HMs stress, although our results did not prove a correlation between Pro levels and Zn accumulation in roots or above-ground parts.
Accumulation of zinc in experimental plants
The average Zn content in the roots was increased to 655.7 µg g−1 in ‘Felina 32’ by NTA application and to 606 µg g−1 in ‘Monoica’ compared to the control, where it reached values of 431 µg g−1 in ‘Felina 32’ and 497 µg g−1 in ‘Monoica’ (Fig. 4c). However, due to the high variability in zinc content among the analyzed plants, this difference was not statistically significant (p ≤ 0.05).
In the control variant of both varieties, Zn levels detected in the root system were more than 10 times higher than the above-ground part (Fig. 4c, d). Such a pattern of Zn accumulation is typical for non-hyperaccumulator plants, of which hemp is one. In these plants the translocation of Zn2+ to shoot is limited [94]. Similarly, Shi and Cai [81] analyzed approximately 3–6 times higher Zn levels in the roots of hemp grown in contaminated soil than in above-ground parts. In contrast, Malik et al. [95] determined a translocation factor for Zn higher than 1 in wild hemp in Pakistan. Galic et al. [33] tested 4 varieties of industrial hemp for its ability to accumulate heavy metals. Most of the varieties accumulated higher amounts of heavy metals in the root system than in above-ground parts but some varieties had a TF higher than 1 for Zn, Hg, Mo and Cd. In our experiments, the shoot bioconcentration and translocation factors were lower than 1 for all tested variants (Additional file 1: Table S4).
NTA application significantly (p ≤ 0.05) increased Zn accumulation in the above-ground parts (Fig. 4d). In the ‘Felina 32’ variety, this increase in Zn levels was more than threefold and reached an average value of 133 µg g−1. In the ‘Monoica’ variety, more than twice as high with an average value of 87 µg g−1. In general, the ‘Felina 32’ variety accumulated a higher amount of Zn in the shoot after NTA application. On the contrary, in control plants not treated with chelating agents, the levels of Zn accumulated in the roots and shoots were similar in both varieties, with values for shoot Zn content 43 µg g−1 in ´Felina 32´ and 41 µg g−1 in ‘Monoica’ variety. The values of Zn content in the above-ground parts were similar to those found by De Vos et al. [94] in total above ground biomass of industrial hemp cultivated in HMs polluted soil with a total zinc content of 952 µg g−1. Similar to our experiments, Wang et al. [96] detected an approximately 300% increased uptake of Zn into the shoot in alfalfa (Medicago sativa) after NTA application at a concentration of 5 mmol L−1. Petrová et al. [61] recorded higher levels of accumulated Zn in hemp roots, stem and leaves than us, however, the plants were grown in hydroponic culture and a stronger EDTA chelation agent was applied to increase the bioavailability of HMs.
On the contrary, the effect of CA application was insignificant (p ≤ 0.05) in our experiments. In both varieties, the Zn content in the root system was similar to that of the control plants, while the content in the shoot was slightly increased. The low efficiency of CA in increasing the mobilization and uptake of Zn by plants in our experiments can be explained by the fact that CA is a weaker chelator than synthetic chelating agents, so it is usually less effective in solubilizing HMs in soil than most synthetic chelators [83, 97, 98]. As shown by other studies [99], increasing the applied concentration of CA, would make it possible to increase the mobility of Zn in the soil and its accumulation in plants, however, in our preliminary experiments, a concentration of CA higher than 10 mmol L−1 appeared to be phytotoxic for hemp. Moreover, the half-life of HMs mobilization in soil after CA application determined by Meers et al. [100] in the range of 1.5–5.7 days is relatively short, and is therefore possible that in our experiments it was not sufficient for a higher amount of Zn in plants to accumulate (especially if young, 14-day-old plants were used). Another possibility is that CA’s ability to mobilize Zn was reduced by the high content of carbonates in the soil used for experiments. Lesage et al. [101] noted that the high buffering capacity of calcareous soils results in CA application’s ineffectiveness in increasing HMs bioavailability in a soil. On the contrary, in the case of NTA application to soil, Wang et al. [96] reported only low competition between main soil cations like Ca and Mg with Zn for chelation.
As demonstrated earlier, the levels of accumulated Zn in the NTA or CA treated variant did not result in a significant (p ≤ 0.05) inhibitory effect on the growth of experimental plants, which may be due to the fact that phytotoxic levels of Zn in tissues were not reached. Such levels of Zn have not yet been published for hemp, but up to 657.5 µg g−1 in shoots and up to 5029 µg g−1 in roots have been demonstrated in plants growing on highly contaminated soil with Zn content of 800 µg g−1 [81]. The maximum bioaccumulation potential of hemp for Zn is therefore higher than recorded in the soil we used. In addition, the application of chelating agents mitigates the toxic effect of higher HMs concentrations on plant growth [102, 103]. The higher biomass production, the size of the root system and the overall higher ability to accumulate zinc in the above-ground biomass indicate a higher potential for ‘Felina 32’ in chelate assisted phytoremediation of Zn contaminated soils.
The influence of NTA and CA on soil microbial activity in zinc-polluted soil
Dehydrogenase activity was referred to as mitigated by higher content of HMs in soil [104, 105]. Thus, there was a presumption for decreased DHA due to the unimmobilized Zn ions in untreated soil controls FK, MK. However, comparable DHA in both variants FN, MN and respective controls FK, MK (Fig. 5a) indicated that NTA chelating agents did not alleviate Zn-derived toxicity on DHA produced by soil microbiota. However, treatment with CA was coupled with significantly (p ≤ 0.05) increased DHA in both ‘Felina 32’ and ‘Monoica’-planted soil variants (FC, MC) compared to the control (MK) and both FK and FN, Fig. 5a. This observed diverse effect for NTA and CA can be explained by the presumed NTA-derived increased solubility of Zn (compared to the CA-derived effect). Nevertheless, the consequential higher DHA inhibition was also accompanied with higher Zn phytoextraction efficacy in the NTA-treated variants (compared to untreated variants) and with significantly (p ≤ 0.05) increased Zn accumulation in shoots of both ‘Felina 32’ and ‘Monoica’ hemp plants (Fig. 4c, d).

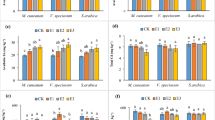
Enzyme activities in artificially Zn-contaminated soil treated with chelating agents. a Dehydrogenase (DHA), b n-acetyl-β-d-glucosaminidase (NAG), c β-glucosidase (GLU), d urease (Ure), e phosphatase (Phos), f arylsulfatase (ARS) in experimental variants (Table 2). Mean values (n = 27) in columns marked with same letters are not statistically significantly different (Tukey’s test, p ≤ 0.05), error bars = standard deviation
On the contrary, CA did not affect soil zinc availability similar to NTA, albeit CA being referred to as efficient in Zn removal from soil [106, 107] as well as in the promotion of [108]. The soil pH could play its role, as pKa2 of CA is higher (6.4) than NTA (3.1), resulting in more dissociated acidic groups on an NTA molecule and higher negative charge in soil experimental conditions, which would promote higher affinity to Zn ions. This assumption, which agrees with more efficient NTA-assisted phytoextraction, is relatively novel due to its difference from reported higher CA efficiency (compared to NTA) in Zn extraction from soil [109]. In contrast, DHA in the MC variant could be partially caused by enhanced microbial utilization of CA (as an easily degradable source of carbon and energy) [110]. Dehydrogenase values were related to indicators of plant fitness and stress (chlorophyll a/b ratio and Pro content in shoot). The ratio of chlorophyll a/b increased in hemp plants upon rising ROS levels, which may indicate enhanced phytoextraction activity (and increased Zn uptake), was coupled with decreased adverse effect on DHA in soil. Therefore, DHA significantly (p ≤ 0.05) positively correlated with the ratio of chlorophyll a/b (r = 0.5), and also negatively correlated with Proline_shoot (r = − 0.52), Fig. 6.

Pearson’s correlation of variables determined with artificially Zn-contaminated soil treated with chelating agents. Values of Pearson’s correlation matrix present correlation coefficient r, statistical significance of results is indicated by symbol(s): ˙p ≤ 0.1, *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001
n-acetyl-β-d-glucosaminidase (NAG) indicates soil fungal biomass turnover rate. Albeit not reported as markedly negatively affected by zinc in soil [111], a significant (p ≤ 0.05) decrease in NAG was detected in FN variant (compared to the control soil FK with ‘Felina 32’ hemp) and in the MC variant (as compared to both two variants seeded with ‘Monoica’), Fig. 5b. The results in ‘Felina 32’-planted variants were close to the DHA and ‘Monoica’ trends showing a NAG activity decrease in MC similar to GLU (β-glucosidase) determination results. The observed NAG decrease in the FN variant could again be associated with the adverse effect of increased mobility of Zn in soil (assumed from higher uptake of Zn by plants). Moreover, insignificantly (p ≤ 0.05) lowest quantitative indicators of plant biomass (except dry weight of roots, Fig. 1a–d) in the FN variant may presume that degradation activity in the respective NTA-treated soil was decreased due to the lowered hemp-derived stimulation of fungal community abundance and activity. Whereas DHA was enhanced putatively in the MC variant due to the access of citric acid, CA-derived mitigation of soil polysaccharide degradation in the MC variant could have been caused due to preferential utilization of a more easily degradable source of organic carbon (i.e. CA) before catabolising less labile intermediates of either chitin, or cellulose digestion. Therefore, MC variant with ‘Monoica’ hemp and CA had significantly (p ≤ 0.05) lower GLU than MK and MF (Fig. 7) from this same presumed reason as in cases with NAG. Nevertheless, GLU is one of the soil quality and carbon mineralization indicators, which was negatively affected by environmental metalloids (Zn) [112, 113]. However, GLU values in variants planted with the ‘Felina 32’ hemp variety were not affected by either NTA or CA treatment. Pearson’s analysis showed a statistically significant (p ≤ 0.05) moderate positive correlation of GLU with chlorophyll a and b (r were 0.57 and 0.56) and a weak negative correlation with Vj and Vi (r =− 0.49 both) (Fig. 6). GLU was synergic with chlorophyll a and b, as well as with Vj also on a PCA biplot (Fig. 7). Decreased chlorophyll fluorescence parameters in hemp leaves was coupled with decreased β-glucosidase activity and indicated chelating agents’ positive effect of on Zn soil mobility, resulting in improved plant uptake, similar to that referred to earlier [114]. Similarly, flattering Vj part of OJIP curves, which could indicate the worsened function of PSII on the level of the second electron acceptors [115], may lead to decreased (Vj) values as a response to increased phytoextraction rate in plants.

PCA Principal component analysis of variables determined with artificially Zn-contaminated soil treated with chelating agents
Phosphatase (Phos), urease (Ure), arylsulfatase (ARS) were also reported as negatively affected by Zn soil contamination, in this sequence (Phos > Ure > ARS) of their resistance to toxic impact of Zn2+ [113, 116, 117]. Urease (Ure) catalyzes the deamination pathway in nitrogen mineralization and was increased in the MC variant (compared to the control soil MK with ‘Monoica’ hemp), Fig. 5d, which corresponded to the results of DHA determination. As PCA biplot showed synergy between Ure and DHA, Phos, access of CA (in the form of labile carbon) in variant MC presumably caused higher microbial abundance (due to labile CA consumption) and together with slightly acidified soil this was followed by enhanced urease activity. Phosphatase (Phos) as an enzyme involved in phosphate solubilization and accessibility in soil [118], and was significantly (p ≤ 0.05) decreased in the FN variant (compared to FK and FC) but increased in the MN variant (compared to MK and MC), Fig. 5e. NTA presumably derived enhanced solubility and extractability of Zn which deteriorated Phos in the ‘Felina 32’ variant. This revealed adverse nitrilotriacetic acid effects on Phos assigned to the previously observed results of NTA-coupled inhibition of other enzymes. However, contrasting enhancement of phosphatase by NTA in the ‘Monoica’-planted soil was hardly explainable and would need further investigation to elucidate this issue. A possible explanation could be ascribed from the revealed significant (p ≤ 0.05) negative correlation of Phos with root system length (r = − 0.52, Fig. 6). It was assumed that less developed roots from the NTA-treated ‘Monoica’ variety decreased the efflux of phosphate into hemp plants in variant MN, resulting in the higher content of remaining available phosphate in the soil. More phosphate preserved in soil may promote higher phosphatase activity despite the inhibiting effect of NTA.
Sulfate in soil form complexes with Zn, which decrease extractibility and prevent leaching of both (Zn2+, SO42−) [119]. Higher chelaton-derived Zn solubility as presumed (and mentioned above) was putatively coupled with the dissociation of complexes with sulfate and higher sulfate solubility in soil, resulting in increased leaching. Thus, higher organism demand for S was caused, which promoted arylsulfatase activity (ARS), Fig. 5f. Only the variant with MC had significantly lower ARS (p ≤ 0.05) than all other variants, which arylsulfatase activity was comparable to each other. The presumed mitigated CA-derived solubilization of complexed sulfate may prevent its leaching and, thus, induce lower ARS activity, which mediated organic S mineralization. ARS significantly (p ≤ 0.01) positively correlated with root system length (r = 0.65), Fig. 6, which could indicate that also more developed roots, providing higher efflux of sulfates from soil, may promote increased demand for ARS-derived organic S mineralization in soil.
The trends in enzyme activities for soil variants planted with ‘Felina 32’ and ‘Monoica’ hemp, were also corroborated by soil respiration determination results. Soil respiration was referred to as increased in heavy metal-contaminated soil under chelating agent treatment [119]. Similarly, significantly (p ≤ 0.05) increased basal respiration (BR) was found in both chelating agents-treated variants of ‘Felina 32’-planted soil (FN and FC) compared to control FK and in variant MC compared to ‘Monoica’ variants MK and MN), Fig. 8a. Nevertheless, contrasting increased respiration drivers are considered for different variants with higher BR. Chelating agents probably increased Zn availability and soil microbiota subsequently increased the energy investment in metabolism to resist HM stress. This imposed C release and promoted enhanced respiration in ‘Felina 32’ variant FN, as a similar microbial response was already referred [120]. On the other hand, CA amendment presented a source of labile and easily utilizable carbon source, which could further induce higher respiration due to its consumption in MC variant, even when increased Zn availability was not assumed. D-glucose-induced respiration (Glc-SIR) determination showed that maximal microbial potential for respiration (indicating an abundance of microbial aerobes in soil) was significantly (p ≤ 0.05) decreased in all chelating agent-treated variants (FN, FC and MN, MC) compared to Zn-contaminated untreated ones (FK, MK), Fig. 8b. This detrimental effect of chelating agents on soil microbial abundance was ascribed to increased Zn mobility (due to NTA and CA application), which casted higher selective pressure on soil microbial community. A similar decrease in microbial abundance but increase in activity due to the shift in composition of resistant microbiome groups was reported [121].

Respiration types in artificially Zn-contaminated soil treated with chelating agents. a Basal respiration (BR), b D-glucose-induced respiration (Glc-SIR), c D-mannose-induced respiration (Man-SIR), d protocatechuic acid-induced respiration (Pro-SIR), e D-trehalose-induced respiration (Tre-SIR) in the experimental variants
Therefore, it was ascribed that NTA amendment to soil (planted with ‘Monoica’ and especially with ‘Felina 32’) derived such efficient Zn solubilization and mobility, which burdened soil microbiome with strong toxic effect, which decreased both enzyme activities (DHA, NAG, Phos in ‘Felina 32’-planted soil) and substrate-induced respirations (d-mannose-, protocatechuic acid- and d-trehalose-induced) Man-SIR, Pro-SIR, Tre-SIR (Fig. 8b–e) in comparison to the other variants—controls (FK, MK) and MC. In contrast, CA application in the MC variant putatively promoted the least efficient solubilization of Zn (as it was assumed previously) and the weakest detrimental effect resulting in the highest Glc-SIR (in average) from all chelating agent-treated variants in ‘Monoica’ planted soil (Fig. 8b). Thus, the observed substrate-induced respirations in the MC variant (Man-SIR and Pro-SIR, Fig. 8b, c) were significantly (p ≤ 0.05) the highest, presumably not due to the strongest stress-induced enhancement of metabolism-transforming activity in response to Zn-toxicity, but due to the stimulation of growth and activity of functional groups of aerobes capable of rapidly degrading simple saccharides and intermediates of lignin degradation. This presumed coupling of all SIR types with the presumed promotion of functional groups of Man, Pro, Tre degraders could be ascribed from the significant (p ≤ 0.001) positive correlation between Man-SIR, Pro-SIR, Tre-SIR one another (0.40 ≤ r ≤ 0.79), Fig. 6.
Conclusion
In this study, greenhouse pot experiments were conducted to investigate the potential of industrial hemp plants assisted by CA and NTA to remediate Zn-polluted soils. The results of our study proved that the exogenous application of NTA chelating agent can significantly increase Zn uptake from contaminated soil by industrial hemp plants and its translocation to above-ground organs, and this, at the given level of Zn in the soil and at the concentrations of the chelating agent used, is without a significant impact on plant growth and their biomass production. In contrast, the soil microbiome was negatively affected, presumably due to NTA-derived increased availability of Zn, which lead to significantly decreased enzyme activities (in ‘Monoica’ planted soil) and induced respirations (in ‘Monoica’ and especially in ‘Felina 32’ planted soil). Conversely, the application of CA to increase the Zn bioavailability for hemp plants has been shown to be ineffective. Despite CA’s negative effects on some soil enzymes found in the ‘Felina 32’ variety, CA enhanced urease activity and carbon mineralization (dehydrogenase and several respiration types) and exerted less detrimental effect on the soil microbiome. Of the two tested varieties, the ‘Felina 32’ variety showed better growth parameters and higher Zn accumulation. Overall, enhancing phytoextraction of industrial hemp assisted with NTA is a promising method for remediating Zn-polluted soils, however, this needs to be further examined in the field.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- a.u.:
-
Arbitrary units
- ARS:
-
Arylsulfatase
- BCF:
-
Bioconcentration factor
- BR:
-
Soil basal respiration
- CA:
-
Citric acid
- CRM:
-
Certified reference material
- DHA:
-
Dehydrogenase activity
- DTPA:
-
Diethylenetriaminepentaacetic acid
- EDDS:
-
Ethylenediaminedisuccinic acid
- EDTA:
-
Ethylenediaminetetraacetic acid
- EGTA:
-
Ethylene glycol-bis-(2-aminoethylether)-tetraacetic acid
- Glc-SIR:
-
Respiration induced by d-glucose
- GLU:
-
β-Glucosidase
- HEDTA:
-
Hydroxyethylenediaminetetraacetic acid
- HM:
-
Heavy metal
- HSD:
-
Honestly significant difference
- ICP-MS:
-
Inductively coupled plasma mass spectrometer
- LOD:
-
Limit of detection
- LOQ:
-
Limit of quantitation
- Man-SIR:
-
Respiration induced by d-mannose
- MPC:
-
Maximum permissible concentrations
- NAG:
-
N-acetyl-β-d-glucosaminidase
- NTA:
-
Nitrilotriacetic acid
- OJIP:
-
Fast chlorophyll fluorescence induction curves
- PCA:
-
Principal component analysis
- Phos:
-
Phosphatase
- PPFD:
-
Photosynthetic photon flux density
- Pro:
-
Proline
- Pro-SIR:
-
Respiration induced by protocatechuic acid
- PSII:
-
Photosystem II
- PTE:
-
Potentially toxic element
- ROS:
-
Reactive oxygen species
- TF:
-
Translocation factor
- Tre-SIR:
-
Respiration induced by d-trehalose
- TTC:
-
2,3,5-Triphenyltetrazolium chloride
- Ure:
-
Urease
References
Kabir A, Swaraz A, Stangoulis J. Zinc-deficiency resistance and biofortification in plants. J Plant Nutr Soil Sci. 2014. https://doi.org/10.1002/jpln.201300326.
Castillo-González J, Ojeda-Barrios D, Hernández-Rodríguez A, González-Franco AC, Robles-Hernández L, López-Ochoa GR. Zinc metalloenzymes in plants. Interciencia. 2018;43(4):242–8.
Noguero M, Atif RM, Ochatt S, Thompson RD. The role of the DNA-binding One Zinc Finger (DOF) transcription factor family in plants. Plant Sci. 2013;209:32–45. https://doi.org/10.1016/J.PLANTSCI.2013.03.016.
Hänsch R, Mendel RR. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr Opin Plant Biol. 2009;12(3):259–66. https://doi.org/10.1016/j.pbi.2009.05.006.
Bailey S, Thompson E, Nixon PJ, Horton P, Mullineaux CW, Robinson C, Mann NH. A critical role for the Var2 FtsH homologue of Arabidopsis thaliana in the photosystem II repair cycle in vivo. J Biol Chem. 2002;277:2006–11. https://doi.org/10.1074/jbc.M105878200.
Rudani L, Vishal P, Kalavati P. The importance of zinc in plant growth—a review. Int Res J Nat Appl Sci. 2018;5:38–48.
Morina F, Jovanovic L, Mojovic M, et al. Zinc-induced oxidative stress in Verbascum thapsus is caused by an accumulation of reactive oxygen species and quinhydrone in the cell wall. Physiol Plant. 2010;140:209–24. https://doi.org/10.1111/j.1399-3054.2010.01399.x.
Andrejić G, Gajić G, Prica M, Dželetović Ž, Rakić T. Zinc accumulation, photosynthetic gas exchange, and chlorophyll a fluorescence in Zn-stressed Miscanthus × giganteus plants. Photosynthetica. 2018;56(4):1249–58. https://doi.org/10.1007/s11099-018-0827-3.
Han FX, Banin A, Su Y, Monts DL, Plodinec JM, Kingery WL, Triplett GE. Industrial age anthropogenic inputs of heavy metals into the pedosphere. Naturwissenschaften. 2002;89(11):497–504. https://doi.org/10.1007/s00114-002-0373-4.
Adriano DC. Trace elements in the terrestrial environment. New York: Springer; 1986. https://doi.org/10.1007/978-1-4757-1907-9.
Kebonye NM, Eze PN, John K, Agyeman PC, Němeček K, Borůvka L. An in-depth human health risk assessment of potentially toxic elements in highly polluted riverine soils, Příbram (Czech Republic). Environ Geochem Health. 2022;44(2):369–85. https://doi.org/10.1007/s10653-021-00877-3.
Ersoy A, Yunsel TY, Atici Ü. Geostatistical conditional simulation for the assessment of contaminated land by abandoned heavy metal mining. Environ Tox. 2008;23(1):96–109. https://doi.org/10.1002/tox.20314.
Van Meirvenne M, Meklit T. Geostatistical simulation for the assessment of regional soil pollution. Geogr Anal. 2010;42(2):121–35. https://doi.org/10.1111/j.1538-4632.2010.00786.x.
Chabukdhara M, Munjal A, Nema AK, Gupta SK, Kaushal RK. Heavy metal contamination in vegetables grown around peri-urban and urban-industrial clusters in Ghaziabad. India Hum Ecol Risk Assess. 2016;22(3):736–52. https://doi.org/10.1080/10807039.2015.1105723.
Madu DI, Anthonius AE, Vivian A, Nonso CE, Ikechukwu EE. Assessment of heavy metal contents, and probable health risks of some staple vegetables in enugu metropolis. Eur J Nutr Food Saf. 2021;13:1–14. https://doi.org/10.9734/ejnfs/2021/v13i130341.
Ngweme GN, Konde JNN, Laffite A, Kiyombo GM, Mulaji CK, Poté JW. Contamination levels of toxic metals in marketed vegetable (Amaranthus viridis) at Kinshasa, Democratic Republic of the Congo. J Food Sci Nutr. 2021;7:87. https://doi.org/10.24966/FSN-1076/100087.
US Department of the Interior and US Geological Survey. Minerals yearbook: metals and minerals. Washington: US Gov Printing Office. 1994–2000.
Shahid M, Austruy A, Echevarria G, Arshad M, Sanaullah M, Aslam M, Nadeem M, Nasim W, Dumat C. EDTA-enhanced phytoremediation of heavy metals: a review. Soil Sediment Contam Int J. 2014;23(4):389–416. https://doi.org/10.1080/15320383.2014.831029.
Quarshie SDG, Xiao XF, Zhang LL. Enhanced phytoremediation of soil heavy metal pollution and commercial utilization of harvested plant biomass: a review. Water Air Soil Pollut. 2021;232(11):28. https://doi.org/10.1007/s11270-021-05430-7.
Deng X, Yuan XQ, Chen LH, Chen YX, Rong XM, Zeng QR, Yang Y. Field-scale remediation of cadmium-contaminated farmland soil by Cichorium intybus L.: Planting density, repeated harvests, and safe use of its Cd-enriched biomass for protein feed. Ind Crop Prod. 2022;188:9. https://doi.org/10.1016/j.indcrop.2022.115604.
Adomako MO, Yu FH. Effects of resource availability on the growth, Cd accumulation, and photosynthetic efficiency of three hyperaccumulator plant species. J Environ Manage. 2023;345:8. https://doi.org/10.1016/j.jenvman.2023.118762.
Sut-Lohmann M, Grimm M, Kästner F, Raab T, Heinrich M, Fischer T. Brassica juncea as a feasible hyperaccumulator of chosen potentially toxic metals under extreme environmental conditions. Int J Environ Res. 2023;17(3):10. https://doi.org/10.1007/s41742-023-00528-8.
Rascio N, Navari-Izzo F. Heavy metal hyperaccumulating plants: how and why do they do it? And what makes them so interesting? Plant Sci. 2011;180:169–81. https://doi.org/10.1016/j.plantsci.2010.08.016.
Yuan Y, Imtiaz M, Rizwan M, Dai ZH, Hossain MM, Zhang YH, Huang HL, Tu SX. The role and its transcriptome mechanisms of cell wall polysaccharides in vanadium detoxication of rice. J Hazard Mater. 2022. https://doi.org/10.1016/j.jhazmat.2021.127966.
Khan AG, Kuek C, Chaudhry TM, Khoo CS, Hayes WJ. Role of plants, mycorrhizae and phytochelators in heavy metal contaminated land remediation. Chemosphere. 2000;41:197–207. https://doi.org/10.1016/S0045-6535(99)00412-9.
Dhiman SS, Zhao X, Li J, Kim D, Kalia VC, Kim IW, Kim JY, Lee JK. Metal accumulation by sunflower (Helianthus annuus L.) and the efficacy of its biomass in enzymatic saccharification. PLoS ONE. 2017;12(4): e0175845. https://doi.org/10.1371/journal.pone.0175845.
Antonkiewicz J, Kołodziej B, Bielińska EJ, Witkowicz R, Tabor S. Using Jerusalem artichoke to extract heavy metals from municipal sewage sludge amended soil. Pol J Environ Stud. 2018;27(2):513–27. https://doi.org/10.15244/pjoes/75200.
Hadad M, Nassar D, Shtaya M. Heavy metals accumulation in soil and uptake by barley (Hordeum vulgare) irrigated with contaminated water. Sci Rep. 2023;13:4121. https://doi.org/10.1038/s41598-022-18014-0.
Dhiman SS, Selvaraj C, Li J, Singh R, Zhao X, Kim D, Kim JY, Kang YC, Lee JK. Phytoremediation of metal-contaminated soils by the hyperaccumulator canola (Brassica napus L.) and the use of its biomass for ethanol production. Fuel. 2016;183:107–14. https://doi.org/10.1016/j.fuel.2016.06.025.
Grzegórska A, Czaplicka N, Antonkiewicz J, Rybarczyk P, Baran A, Dobrzyński K, Zabrocki D, Rogala A. Remediation of soils on municipal rendering plant territories using Miscanthus x giganteus. Environ Sci Pollut Res. 2023;30(9):22305–18. https://doi.org/10.1007/s11356-022-23724-z.
Ancona V, Barra Caracciolo A, Campanale C, De Caprariis B, Grenni P, Uricchio VF, Borello D. Gasification treatment of poplar biomass produced in a contaminated area restored using plant assisted bioremediation. J Environ Manage. 2019;1:137–41. https://doi.org/10.1016/j.jenvman.2019.03.038.
Citterio S, Santagostino A, Fumagalli P, Prato N, Ranalli P, Sgorbati S. Heavy metal tolerance and accumulation of Cd, Cr and Ni by Cannabis sativa L. Plant Soil. 2003;256:243–52. https://doi.org/10.1023/A:1026113905129.
Galic M, Percin A, Zgorelec Z, Kisic I. Evaluation of heavy metals accumulation potential of hemp (Cannabis sativa l.). J Cent Eur Agric. 2019;20(2):700–11. https://doi.org/10.5513/jcea01/20.2.2201.
Placido DF, Lee CC. Potential of industrial hemp for phytoremediation of heavy metals. Plants. 2022;11:595. https://doi.org/10.3390/plants11050595.
Golia EE, Bethanis J, Ntinopoulos N, Kaffe GG, Komnou AA, Vasilou C. Investigating the potential of heavy metal accumulation from hemp: the use of industrial hemp (Cannabis sativa l.) for phytoremediation of heavily and moderated polluted soils. Sustain Chem Pharm. 2023;31:20. https://doi.org/10.1016/j.scp.2022.100961.
Weber M, Harada E, Vess C, Roepenack-Lahaye EV, Clemens S. Comparative microarray analysis of Arabidopsis thaliana and Arabidopsis halleri roots identifies nicotianamine synthase, a ZIP transporter and other genes as potential metal hyperaccumulation factors. Plant J. 2004;37:269–81. https://doi.org/10.1046/j.1365-313X.2003.01960.x.
Čásová K, Černý J, Száková J, Balík J, Tlustoš P. Cadmium balance in soils under different fertilization managements including sewage sludge application. Plant Soil Environ. 2009;55(8):353–61. https://doi.org/10.17221/142/2009-PSE.
Roberts TL. Cadmium and phosphorous fertilizers: the issues and the science. Procedia Eng. 2014;83:52–9. https://doi.org/10.1016/j.proeng.2014.09.012.
Sadeghzadeh B. A review of zinc nutrition and plant breeding. J Soil Sci Plant Nutr. 2013;13(4):907–27. https://doi.org/10.4067/S0718-95162013005000072.
Valipour M, Shahbazi K, Khanmirzaei A. Chemical immobilization of lead, cadmium, copper, and nickel in contaminated soils by phosphate amendments. Clean-Soil Air Water. 2016;44(5):572–8. https://doi.org/10.1002/clen.201300827.
Suzuki M, Tsukamoto T, Inoue H, et al. Deoxymugineic acid increases Zn translocation in Zn-deficient rice plants. Plant Mol Biol. 2008;66:609–17. https://doi.org/10.1007/s11103-008-9292-x.
Banakar R, Alvarez Fernandez A, Díaz-Benito P, Abadia J, Capell T, Christou P. Phytosiderophores determine thresholds for iron and zinc accumulation in biofortified rice endosperm while inhibiting the accumulation of cadmium. J Exp Bot. 2017;68(17):4983–95. https://doi.org/10.1093/jxb/erx304.
Hazama K, Nagata S, Fujimori T, Yanagisawa S, Yoneyama T. Concentrations of metals and potential metal-binding compounds and speciation of Cd, Zn and Cu in phloem and xylem saps from castor bean plants (Ricinus communis) treated with four levels of cadmium. Physiol Plant. 2015;154:243–55. https://doi.org/10.1111/ppl.12309.
Khodamoradi K, Khoshgoftarmanesh AH, Dalir N, Afyuni M, Schulin R. How do glycine and histidine in nutrient solution affect zinc uptake and root-to-shoot translocation by wheat and triticale? Crop Pasture Sci. 2015;66(11):1105–10. https://doi.org/10.1071/CP14227.
Monsant AC, Kappen P, Wang Y, et al. In vivo speciation of zinc in Noccaea caerulescens in response to nitrogen form and zinc exposure. Plant Soil. 2011;348:167. https://doi.org/10.1007/s11104-011-0887-7.
Sagardoy R, Morales F, Rellán-Álvarez R, Abadía A, Abadía J, López-Millán AF. Carboxylate metabolism in sugar beet plants grown with excess Zn. J Plant Physiol. 2011;168(7):730–3. https://doi.org/10.1016/j.jplph.2010.10.012.
Barrameda-Medina Y, Montesinos-Pereira D, Romero L, Ruiz JM, Blasco B. Comparative study of the toxic effect of Zn in Lactuca sativa and Brassica oleracea plants: I. Growth, distribution, and accumulation of Zn, and metabolism of carboxylates. Environ Exp Bot. 2014;107:98–104. https://doi.org/10.1016/j.envexpbot.2014.05.012.
Sarkar D, Syam SA, Sumathi KMS, Rupali D. Chelant-aided enhancement of lead mobilization in residential soils. Environ Pollut. 2008;156:1139–48. https://doi.org/10.1016/j.envpol.2008.04.004.
Dipu S, Kumar AA, Thanga SG. Effect of chelating agents in phytoremediation of heavy metals. Remediat J. 2012;22:133–46. https://doi.org/10.1002/rem.21304.
Dong W, Wang R, Li H, Yang X, Li J, Wang H, Jiang C, Wang Z. Effects of chelating agents addition on ryegrass extraction of cadmium and lead in artificially contaminated soil. Water. 1929;2023:15. https://doi.org/10.3390/w15101929.
Santos FS, Hernández-Allica J, Becerril MJ, Amaral Sobrinho N, Mazur N, Garbisu C. Chelate-induced phytoextraction of metal polluted soils with Brachiaria decumbens. Chemosphere. 2006;65(1):43–50. https://doi.org/10.1016/j.chemosphere.2006.03.012.
Bian X, Cui J, Tang B, Yang L. Chelant-induced phytoextraction of heavy metals from contaminated soils: a review. Pol J Environ Stud. 2018;27(6):2417–24. https://doi.org/10.15244/pjoes/81207.
You Y, Dou J, Xue Y, Jin N, Yang K. Chelating agents in assisting phytoremediation of uranium-contaminated soils: a review. Sustainability. 2022;14:6379. https://doi.org/10.3390/su14106379.
Dermont G, Bergeron M, Mercier G, Richer-Laflèche M. Soil washing for metal removal: a review of physical/chemical technologies and field applications. J Hazard Mater. 2008;152(1):1–31. https://doi.org/10.1016/j.jhazmat.2007.10.043.
Zhou J, Dang Z, Chen N, Xu S, Xie Z. Enhanced phytoextraction of heavy metal contaminated soil by chelating agents and auxin indole-3-acetic acid. Huanjing Kexue/Environ Sci. 2007;28(9):2085–8.
Wang H, Wu C, Liu J, et al. Changes in soil microbial communities induced by warming and N deposition accelerate the CO2 emissions of coarse woody debris. J For Res. 2023;2023(34):1051–63. https://doi.org/10.1007/s11676-022-01544-8.
Xu X, Thomson NR. An evaluation of the green chelant EDDS to enhance the stability of hydrogen peroxide in the presence of aquifer solids. Chemosphere. 2007;69(5):755–62. https://doi.org/10.1016/j.chemosphere.2007.05.008.
Evangelou MWH, Ebel M, Schaeffer A. Evaluation of the effect of small organic acids on phytoextraction of Cu and Pb from soil with tobacco Nicotiana tabacum. Chemosphere. 2006;63(6):996–1004. https://doi.org/10.1016/j.chemosphere.2005.08.042.
Karczewska A, Milko K. Effects of chelating agents on Cu, Pb and Zn solubility in polluted soils and tailings produced by copper industry. Ecol Chem Eng. 2010;A17:95–403.
Kaushik P. Use of natural organic and synthetic chelating agents for efficient phytoremediation. Int J Enhanc Res Sci Technol Eng. 2015;4(6):99–101.
Petrová Š, Benešová D, Soudek P, Vaněk T. Enhancement of metal(loid)s phytoextraction by Cannabis sativa L. J Food Agric Environ. 2012;10:631–41.
Ferrarini A, Fracasso A, Spini G, Fornasier F, Taskin E, Fontanella MC, Beone GM, Amaducci S, Puglisi E. Bioaugmented phytoremediation of metal-contaminated soils and sediments by hemp and giant reed. Front Microbiol. 2021;12:645893. https://doi.org/10.3389/fmicb.2021.645893.
FAO. World reference base for soil resources. Rome: Food and Agriculture Organization of the United Nations; 2014.
Mehlich A. Mehlich 3 soil test extractant: a modification of Mehlich 2 extractant. Commun Soil Sci Plant Anal. 1984;15:1409–16. https://doi.org/10.1080/00103628409367568.
Macečková B, Bjelková M, Větrovcová M, Motyka O, Seidlerová J, Khest F. Comparative assessment for phytoextraction capability using different varieties of non-hyperaccumulator plant species Cannabis sativa L. In: Recent Advances in Neglected and Under-utilized Species Research. October 18–20, 2015, Nitra, Slovak Republic. pp. 25–8.
Kuba M, Gallo J, Pluhacek T, Hobza M, Milde D. Content of distinct metals in periprosthetic tissues and pseudosynovial joint fluid in patients with total joint arthroplasty. J Biomed Mater Res Part B Appl Biomater. 2019;107:454–62.
Han Y, Zhang L, Gu J, Zhao J, Fu J. Citric acid and EDTA on the growth, photosynthetic properties and heavy metal accumulation of Iris halophila Pall. cultivated in Pb mine tailings. Int Biodeterior Biodegrad. 2018;128:15–21. https://doi.org/10.1016/j.ibiod.2016.05.011.
Usman K, Al-Ghouti MA, Abu-Dieyeh MH. The assessment of cadmium, chromium, copper, and nickel tolerance and bioaccumulation by shrub plant Tetraena qataranse. Sci Rep. 2019;9(1):5658. https://doi.org/10.1038/s41598-019-42029-9.
Lichtenthaler HK, Buschmann C. Chlorophylls and carotenoids: measurement and characterization by UV-VIS spectroscopy. Curr Protoc Food Anal Chem. 2001;1:F4.3.1-F4.3.8. https://doi.org/10.1002/0471142913.faf0403s01.
Carillo P, Gibon Y. PROTOCOL: extraction and determination of proline. PrometheusWiki: Clayton South, Australia. 2011. https://prometheusprotocols.net/function/tissue-chemistry/primary-metabolites/extraction-and-determination-of-proline/. Accessed on 15 Apr 2021
Doi R, Ranamukhaarachchi SL. Soil dehydrogenase in a land degradation-rehabilitation gradient: observations from a savanna site with a wet/dry seasonal cycle. Rev Biol Trop. 2009;57:223–34. https://doi.org/10.15517/rbt.v57i1-2.11317.
Campbell CD, Chapman SJ, Cameron CM, Davidson MS, Potts JM. A rapid microtiter plate method to measure carbon dioxide evolved from carbon substrate amendments so as to determine the physiological profiles of soil microbial communities by using whole soil. Appl Environ Microbiol. 2003;69(6):3593–9. https://doi.org/10.1128/AEM.69.6.3593-3599.2003.
ISO 20130. Soil quality—measurement of enzyme activity patterns in soil samples using colorimetric substrates in micro-well plates. Geneva: International Organization for Standardization; 2018.
R Core Team R. A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2020.
Zar JH. Biostatistical analysis. Englewood Cliffs: Prentice-Hall Inc.; 1984.
Hinkle DE, Wiersma W, Jurs SG. Applied statistics for the behavioral sciences. Boston: Houghton Mifflin; 2003.
Nanthavong K, Sampanpanish P. Effect of NTA and EDTA on arsenic uptake from contaminated soil by Mimosa pudica. Mod Applied Sci. 2015;9:280–91. https://doi.org/10.5539/mas.v9n9p280.
Hart G, Koether M, McElroy T, Greipsson S. Evaluation of chelating agents used in phytoextraction by switchgrass of lead contaminated soil. Plants. 2022;11:1012. https://doi.org/10.3390/plants11081012.
Decree no. 153/2016 Coll. on details on protection of agricultural soil quality and on amendment to Decree no. 13/1994 Coll., specifying some detail on agricultural land fund protection.
Crommentuijn T, Polder MD, van de Plassche EJ. Maximum permissible concentrations and negligible concentrations for metals, taking background concentrations into account RIVM Report 601501001. Bilthoven, Netherlands. 1997. https://doi.org/10.1006/jema.2000.0354.
Shi G, Cai Q. Zinc tolerance and accumulation in eight oil crops. J Plant Nutr. 2010;33:982–97. https://doi.org/10.1080/01904161003728669.
Li Y, Wang Y, Khan MA, Luo W, Xiang Z, Xu W, Zhong B, Ma J, Ye Z, Zhu Y, Duan L, Liu D. Effect of plant extracts and citric acid on phytoremediation of metal-contaminated soil. Ecotoxicol Environ Saf. 2021;15(211):111902. https://doi.org/10.1016/j.ecoenv.2021.111902.
Muhammad D, Chen F, Zhao J, Zhang G, Wu F. Comparison of EDTA- and citric acid-enhanced phytoextraction of heavy metals in artificially metal contaminated soil by Typha angustifolia. Int J Phytoremed. 2009;11(6):558–74. https://doi.org/10.1080/15226510902717580.
Chiu KK, Ye ZH, Wong MH. Growth of Vetiveria zizanioides and Phragmites australis on Pb/Zn and Cu mine tailings amended with manure compost and sewage sludge: a greenhouse study. Bioresour Technol. 2006;97(1):158–70. https://doi.org/10.1016/j.biortech.2005.01.038.
Souri Z, Cardoso AA, Da-Silva CJ, De Oliveira LM, Dari B, Sihi D, Karimi N. Heavy metals and photosynthesis. In: Ahmad P, Ahanger MA, Alyemeni MN, Alam P, editors. Recent developments: photosynthesis, productivity, and environmental stress. Hoboken: Wiley; 2019. p. 107–34. https://doi.org/10.1002/9781119501800.ch7.
Cherif J, Derbel N, Nakkah M, von Bergmann H, Jemal F, Lakhdar ZB. Analysis of in vivo chlorophyll fluorescence spectra to monitor physiological state of tomato plants growing under zinc stress. J Photochem Photobiol B. 2010;101(3):332–9. https://doi.org/10.1016/j.jphotobiol.2010.08.005.
Huang XH, Zhu F, Yan WD, Chen XY, Wang GJ, Wang RJ. Effects of Pb and Zn toxicity on chlorophyll fluorescence and biomass production of Koelreuteria paniculata and Zelkova schneideriana young plants. Photosynthetica. 2019;57(2):688–97. https://doi.org/10.32615/ps.2019.050.
Rau S, Miersch J, Neumann D, Weber E, Krauss GJ. Biochemical responses of the aquatic moss Fontinalis antipyretica to Cd, Cu, Pb and Zn determined by chlorophyll fluorescence and protein levels. Environ Exp Bot. 2007;59:299–306. https://doi.org/10.1016/j.envexpbot.2006.03.001.
Ciscato M, Vangronsveld J, Valcke R. Effects of heavy metals on the fast chlorophyll fluorescence induction kinetics of photosystem II: a comparative study. Zeitschrift für Naturforschung C. 1999;54(9–10):735–9. https://doi.org/10.1515/znc-1999-9-1019.
Tripathi BN, Gaur JP. Relationship between copper and zinc induced oxidative stress and proline accumulation in Scenedesmus sp. Planta. 2004;219:397–404. https://doi.org/10.1007/s00425-004-1237-2.
Fidalgo F, Azenha M, Silva FA, de Sousa A, Santiago A, Ferraz P, Teixeira J. Copper-induced stress in Solanum nigrum L. and antioxidant defense system responses. Food Energy Secur. 2013;2:70–80. https://doi.org/10.1002/fes3.20.
Zengin FK, Munzuroglu O. Effects of some heavy metals on content of chlorophyll, proline and some antioxidant chemicals in bean (Phaseolus vulgaris L.) seedlings. Acta Biol Crac Ser Bot. 2005;47:157–64.
Kaya C, Akram NA, Sürücü A, Ashraf M. Alleviating effect of nitric oxide on oxidative stress and antioxidant defence system in pepper (Capsicum annuum L.) plants exposed to cadmium and lead toxicity applied separately or in combination. Sci Hortic. 2019;255:52–60. https://doi.org/10.1016/j.scienta.2019.05.029.
De Vos B, Souza MF, Michels E, Meers E. Industrial hemp (Cannabis sativa L.) in a phytoattenuation strategy: remediation potential of a Cd, Pb and Zn contaminated soil and valorization potential of the fibers for textile production. Ind Crops Prod. 2022;178:114592. https://doi.org/10.1016/j.indcrop.2022.114592.
Malik RN, Husain SZ, Nazir I. Heavy metal contamination and accumulation in soil and wild plant species from industrial area of Islamabad, Pakistan. Pak J Bot. 2010;42(1):291–301.
Wang X, De Souza MF, Li H, Qiu J, Ok YS, Meers E. Biodegradation and effects of EDDS and NTA on Zn in soil solutions during phytoextraction by alfalfa in soils with three Zn levels. Chemosphere. 2022;292:133519. https://doi.org/10.1016/j.chemosphere.2022.133519.
Chaturvedi N, Dhal NK, Patra HK. EDTA and citric acid-mediated phytoextraction of heavy metals from iron ore tailings using Andrographis paniculata: a comparative study. Int J Min Reclam Environ. 2015;29(1):33–46. https://doi.org/10.1080/17480930.2014.955328.
Zhang X, Zhong B, Shafi M, Guo J, Liu C, Guo H, Peng D, Wang Y, Liu D. Effect of EDTA and citric acid on absorption of heavy metals and growth of Moso bamboo. Environ Sci Pollut Res Int. 2018;25(19):18846–52. https://doi.org/10.1007/s11356-018-2040-0.
Quartacci MF, Baker AJM, Navari-Izzo F. Nitrilotriacetate- and citric acid-assisted phytoextraction of cadmium by Indian mustard (Brassica juncea (L.) Czernj, Brassicaceae). Chemosphere. 2005;59(9):1249–55. https://doi.org/10.1016/j.chemosphere.2004.11.053.
Meers E, Lesage E, Lamsal S, Hopgood M, Vervaeke P, Tack FMG, Verloo MG. Enhanced phytoextraction: I. effect of EDTA and citric acid on heavy metal mobility in a calcareous soil. Int J Phytorem. 2005;7:129–42. https://doi.org/10.1080/16226510590950423.
Lesage E, Meers E, Vervaeke P, Lamsal S, Hopgood M, Tack FMG, Verloo MG. Enhanced phytoextraction: II. Effect of EDTA and citric acid on heavy metal uptake by Helianthus annuus from a calcareous soil. Int J Phytoremed. 2005;7(2):143–52. https://doi.org/10.1080/16226510590950432.
Badamasi H, Dagari MS, Sale I. Impacts of citric acid on the phytoextraction of zinc (Zn) using sorghum (Sorghum bicolor (l.) Moench) plants. Malays J Anal Sci. 2020;24(4):587–98.
Saleem MH, Ali S, Kamran M, Iqbal N, Azeem M, Tariq Javed M, Ali Q, Zulqurnain Haider M, Irshad S, Rizwan M. Ethylenediaminetetraacetic acid (EDTA) mitigates the toxic effect of excessive copper concentrations on growth, gaseous exchange and chloroplast ultrastructure of Corchorus capsularis L. and improves copper accumulation capabilities. Plants. 2020;9(6):1–20. https://doi.org/10.3390/plants9060756.
Mansour H, Saber M, Awad F, Zaghloul A. Dehydrogenase activity and zinc equivalent parameters as indicators for potential toxic elements remediation in polluted soil ecosystem. Bioremediat J. 2019;23(4):326–33. https://doi.org/10.1080/10889868.2019.1670132.
Gallego S, Esbri JM, Campos JA, Peco JD, Martin-Laurent F, Higueras P. Microbial diversity and activity assessment in a 100-year-old lead mine. J Hazard Mat. 2021;410:12. https://doi.org/10.1016/j.jhazmat.2020.124618.
Khodadoust AP, Reddy KR, Maturi K. Effect of different extraction agents on metal and organic contaminant removal from a field soil. J Hazard Mater. 2005;117(1):15–24. https://doi.org/10.1016/j.jhazmat.2004.05.021.
Asadzadeh F, Maleki-Kaklar M, Soiltanalinejad N, Shabani F. Central composite design optimization of zinc removal from contaminated soil, using citric acid as biodegradable chelant. Sci Rep. 2018;8:2633. https://doi.org/10.1038/s41598-018-20942-9.
Yu XZ, Wang DQ, Zhang XH. Chelator-induced phytoextraction of zinc and copper by rice seedlings. Ecotoxicology. 2014;23(4):749–56. https://doi.org/10.1007/s10646-014-1188-8.
Xiao JH, Zhou J, Wang ZH, Li SY, Zhang WC, Fang CI, Guo YG, Xiao DX, Lou XY, Liu JS. Extraction of copper and zinc from naturally contaminated copper mine soils: Chemical fractionation analysis and risk assessment. J Cent South Univ. 2018;25(6):1274–84. https://doi.org/10.1007/s11771-018-3824-6.
Thinh NV, Osanai Y, Adachi T, Vuong BTS, Kitano I, Chung NT, Thai PK. Removal of lead and other toxic metals in heavily contaminated soil using biodegradable chelators: Glda, citric acid and ascorbic acid. Chemosphere. 2021;263:12. https://doi.org/10.1016/j.chemosphere.2020.127912.
Anza M, Salazar O, Epelde L, Alkorta I, Garbisu C. The application of nanoscale zero-valent iron promotes soil remediation while negatively affecting soil microbial biomass and activity. Front Environ Sci. 2019. https://doi.org/10.3389/fenvs.2019.00019.
Kucharski J, Wieczorek K, Wyszkowska J. Changes in the enzymatic activity in sandy loam soil exposed to zinc pressure. J Elemntol. 2011;16(4):577–89. https://doi.org/10.5601/jelem.2011.16.4.07.
Borowik A, Wyszkowska J, Kucharski J, Bacmaga M, Boros-Lajszner E, Tomkiel M. Sensitivity of soil enzymes to excessive zinc concentrations. J Elementol. 2014;19(3):637–47. https://doi.org/10.5601/jelem.2014.19.2.456.
Ali A, Guo D, Zhang Y, Sun X, Jiang S, Guo Z, Huang H, Liang W, Li R, Zhang Z. Using bamboo biochar with compost for the stabilization and phytotoxicity reduction of heavy metals in mine-contaminated soils of China. Sci Rep. 2017;7(1):2690. https://doi.org/10.1038/s41598-017-03045-9.
Paunov M, Koleva L, Vassilev A, Vangronsveld J, Goltsev V. Effects of different metals on photosynthesis: cadmium and zinc affect chlorophyll fluorescence in durum wheat. Int J Mol Sci. 2018;19(3):787. https://doi.org/10.3390/ijms19030787.
Kucharski J, Wyszkowska J, Wieczorek K. Changes in the enzymatic activity in sandy loam soil exposed. J Elem. 2011;16(4/2011):577–89. https://doi.org/10.5601/jelem.2011.16.4.07.
Wasilkowski D, Nowak A, Plaza G, Mrozik A. Effects of pulp and na-bentonite amendments on the mobility of trace elements, soil enzymes activity and microbial parameters under ex situ aided phytostabilization. PLoS ONE. 2017;12(1):23. https://doi.org/10.1371/journal.pone.0169688.
Jha V, Bombaywala S, Purohit H, Dafale NA. Differential colonization and functioning of microbial community in response to phosphate levels. J Environ Manage. 2022;321:14. https://doi.org/10.1016/j.jenvman.2022.115856.
Phillips CJC, Chiy PC. Effects of applying sodium and sulphur fertilisers on the concentrations of elements in water leached from permanent pasture. J Sci Food Agricul. 2002;82(8):806–15. https://doi.org/10.1002/jsfa.1114.
Xu M, Cui Y, Beiyuan J, Wang X, Duan C, Fang L. Heavy metal pollution increases soil microbial carbon limitation: evidence from ecological enzyme stoichiometry. Soil Ecol Let. 2021;3(3):230–41. https://doi.org/10.1007/s42832-021-0094-2.
Xu Y, Seshadri B, Bolan N, Sarkar B, Ok YS, Zhang W, Rumpel C, Sparks D, Farrell M, Hall T, Dong Z. Microbial functional diversity and carbon use feedback in soils as affected by heavy metals. Environ Int. 2019;125:478–88. https://doi.org/10.1016/j.envint.2019.01.071.
Acknowledgements
The authors gratefully acknowledge Ing. Marie Bjelková, Ph.D. from Agritec Plant Research, Ltd. in Šumperk for providing plant material.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
PK and PS: conceptualization. PK, PS, TH, KŠL, TP and PT: investigation and methodology. PK, JH, KŠL and MB: writing—original draft preparation. PK, JH, KŠL, TV, MB, TP, PT and MR: writing—review and editing. TH, JH and MR: data curation. MB and TV: supervision. All authors contributed to the article and approved the submitted version.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Determination of total zinc content by inductively coupled plasma mass spectrometry (ICP-MS).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kalousek, P., Holátko, J., Schreiber, P. et al. The effect of chelating agents on the Zn-phytoextraction potential of hemp and soil microbial activity. Chem. Biol. Technol. Agric. 11, 23 (2024). https://doi.org/10.1186/s40538-024-00544-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40538-024-00544-6