
Abstract
Azospirillum spp. are plant-growth-promoting bacteria used worldwide as inoculants for a variety of crops. Among the beneficial mechanisms associated with Azospirillum inoculation, emphasis has been given to the biological nitrogen fixation process and to the synthesis of phytohormones. In Brazil, the application of inoculants containing A. brasilense strains Ab-V5 and Ab-V6 to cereals is exponentially growing and in this study we investigated the effects of maize inoculation with these two strains applied on seeds or by leaf spray at the V2.5 stage growth—a strategy to relieve incompatibility with pesticides used for seed treatment. We also investigate the effects of spraying the metabolites of these two strains at V2.5. Maize growth was promoted by the inoculation of bacteria and their metabolites. When applied via foliar spray, although A. brasilense survival on leaves was confirmed by confocal microscopy and cell recovery, few cells were detected after 24 h, indicating that the effects of bacterial leaf spray might also be related to their metabolites. The major molecules detected in the supernatants of both strains were indole-3-acetic acid, indole-3-ethanol, indole-3-lactic acid and salicylic acid. RT-PCR of genes related to oxidative stress (APX1, APX2, CAT1, SOD2, SOD4) and plant defense (pathogenesis-related PR1, prp2 and prp4) was evaluated on maize leaves and roots. Differences were observed according to the gene, plant tissue, strain and method of application, but, in general, inoculation with Azospirillum resulted in up-regulation of oxidative stress genes in leaves and down-regulation in roots; contrarily, in general, PR genes were down-regulated in leaves and up-regulated in roots. Emphasis should be given to the application of metabolites, especially of Ab-V5 + Ab-V6 that in general resulted in the highest up-regulation of oxidative-stress and PR genes both in leaves and in roots. We hypothesize that the benefits of inoculation of Azospirillum on seeds or by leaf spray, as well as of leaf spraying of Azospirillum metabolites, are strongly correlated with the synthesis of phytohormones and by eliciting genes related to plant-stress tolerance and defense against pathogens.
Similar content being viewed by others

Introduction
Inoculation with Azospirillum spp. has been the subject of several studies (Bashan and Holguin 1998) due to their remarkable capacity of promoting growth of important cereals, i.e. maize (Zea mays L.), wheat (Triticum aestivum L.) and rice (Oryza sativa L.), in addition to several grasses (e.g. Hungria et al. 2010, 2016; Cassán et al. 2015; Pereg et al. 2016). The benefits in plant growth have been attributed to a variety of single or combined mechanisms that act either accumulatively or in cascade (Bashan and de-Bashan 2010), including: enhanced uptake of nutrients and water (Ardakani et al. 2011); production and secretion of phytohormones and other signaling molecules such as auxins (Spaepen and Vanderleyden 2015), cytokinins (Tien et al. 1979), gibberellins (Bottini et al. 1989) and salicylic acid (Sahoo et al. 2014); biological nitrogen fixation (Marques et al. 2017); and phosphate solubilization (Rodriguez et al. 2004). However, although Azospirillum spp. seem remarkable in their apparent lack of specificity in promoting growth of practically every plant genus and species investigated so far (Pereg et al. 2016), there are also indications that species and strains may vary in determinants of niche-specific adaptation to the rhizosphere that affect plant–microbe interactions (Wisniewski-Dyé et al. 2012). Examples of determinants of adaptation include reactive oxygen species (ROS) as shown with A. lipoferum strain 4B in the rice rhizosphere (Drogue et al. 2014). ROS molecules encompass free radicals resulting from oxygen metabolism such as superoxide radicals (O2 −), hydroxyl radicals (OH−), hydrogen peroxide (H2O2) and singlet oxygen (1O2) (Bowler et al. 1992; Gill and Tuteja 2010). The most important ROS detoxification mechanism is represented by the activity of superoxide dismutase (SOD), ascorbate peroxidase (APX) and catalase (CAT) enzymes responsible for the scavenging of H2O2 by its conversion to water and O2 (Lamb and Dixon 1997; Asada 1999). In general, ROS detoxification systems vary with plant species, cultivar, and age, and also with the type and duration of abiotic and biotic stress (Hodges et al. 1996).
Another intriguing feature of Azospirillum spp. is that although the species comprise non-pathogenic bacteria, they are also able to induce plant-defense mechanisms that may help against further pathogen attacks (Cassán et al. 2014). This property is called ‘induced systemic resistance’ (ISR), in which the bacterium triggers a plant reaction by emitting signals—the pathogenesis-related proteins (PRs)—that spread systemically throughout the plant and enhance the defensive capacity of distant tissues against infection by pathogens (Van Loon and Bakker 2005). Once induced, plants may remain protected for prolonged periods (Van Loon 2007). For example, there are reports of Azospirillum helping protection against Colletotrichum acutatum (anthracnose) in strawberry (Fragaria ananassa Duch.) (Tortora et al. 2011), and resistance to Clavibacter michiganensis subsp. michiganensis (bacterial canker), Xanthomonas campestris pv. vesicatoria (Romero et al. 2003) and Rhizoctonia solani (damping-off disease) (Gupta et al. 1995) in tomato plants (Lycopersicon esculentum Mill).
Reports of plant-growth improvement by the exogenous application of synthetic growth regulators (e.g. auxins, gibberellins, cytokinins) have long been the subject of studies (e.g. Halmann 1990); more recently, emphasis has also been given to their effect in increasing tolerance of abiotic and biotic stresses (Robert-Seilaniantz et al. 2011). Similar effects on stresses have been reported with the application of jasmonic acid (Bari and Jones 2009; Wasternack 2007; Lorenzo and Solano 2005) and salicylic acid (Bari and Jones 2009), which might induce PR (pathogenesis-related) genes and, consequently, enhance resistance to several pathogens.
The commercial use of Azospirillum brasilense strains Ab-V5 and Ab-V6 on maize (Z. mays L.) and wheat (T. aestivum L.) crops in Brazil has grown exponentially since 2010 (Hungria et al. 2010; Hungria 2011). Our research group has started to investigate the effects of foliar-spray inoculation of Azospirillum, with the main practical purpose of avoiding the contact of the bacteria with harmful pesticides that are heavily applied to the seeds (Fukami et al. 2016). In this study we confirmed benefits to plant growth by seed and foliar applications of Azospirillum, but also verified responses to the application of their metabolites. We then investigated phytohormone production and the response of antioxidant systems with different methods of application of Azospirillum strains and their metabolites.
Materials and methods
Bacterial strains and inoculation methods
Bacteria consisted of strains Ab-V5 (=CNPSo 2083) and Ab-V6 (=CNPSo 2084) of Azospirillum brasilense (from the “Culture Collection of Diazotrophic and Plant Growth-Promoting Bacteria of Embrapa Soja”, WFCC # 1213, WDCM # 1054). Both strains were derived from an Azospirillum selection program (Hungria et al. 2010) and are currently employed in commercial inoculants in Brazil (Hungria 2011).
The inoculants were initially prepared in DYGS medium (Rodrigues Neto et al. 1986) and, after growth for 48 h, cell concentrations were adjusted to 108 mL−1. For the production of metabolites, inoculants were produced under the same conditions and up to the same concentration and were centrifuged at 5000 rpm for 15 min. By plating the supernatants obtained on DYGS medium we confirmed that they were free of Azospirillum cells.
Three methods of inoculation were compared: (i) standard seed inoculation (SI)—considered as the control; (ii) inoculation by leaf spray (ILS) at the V2.5 stage of the maize growth cycle (Hickman and Shroyer 1994); and (iii) application with metabolites from A. brasilense strains Ab-V5 and Ab-V6 by leaf spray (MLS) at the V2.5 stage (about 7 days after transplanting) (Hickman and Shroyer 1994).
Seeds were inoculated 1 h before sowing by thoroughly coating them to provide a final concentration of 1.6 × 105 cells seed−1. For leaf-spray inoculation, an aerograph atomizer was employed to mimic the action of a spraying apparatus. The soil surface was covered with aluminum foil to prevent the inoculant reaching it. The final volume of liquid for leaf-spray inoculation was 1 mL (water + inoculant) per pot containing a single plant, and inoculants were diluted with sterile distilled water at 1:1000 (v:v) for spraying, to achieve an application rate of 1.6 × 105 cells plant−1. For leaf spray of metabolites, bacterial exudate corresponding to the same cell concentration as the seed inoculant used for leaf spray was used, with the application of 1 mL per plant corresponding to 1.6 × 105 cells plant−1. Foliar-spray inoculations of pots containing maize plants were performed 7 days after transplanting.
Greenhouse experiment
The experiment was performed under greenhouse conditions, using modified Leonard jars (Vincent 1970) containing sterilized substrate, consisting of a mixture of sand and pulverized coal (3:1, v/v) with application of sterile nutrient solution (Fahraeus 1957). Jars were arranged in a completely randomized design with nine treatments, a non-inoculated control, and six replicates. Each treatment received 60 kg N ha−1 (50% of the N application recommended for the crop). Inoculation treatments consisted of mineral-N fertilizer (50% N) and different methods of inoculation: SI (standard seed inoculation at sowing), ILS (inoculation by leaf spray, at the V2.5 stage of maize growth) and MLS (inoculation with metabolites by leaf spray of A. brasilense strains Ab-V5 and Ab-V6 at the at the V2.5 stage).
Hybrid maize seeds (DKB330 VT PRO2) were surface-sterilized with 70% ethanol and 3% sodium hypochlorite (Vincent 1970). They were pre-geminated for 48 h at 25 °C in Petri plates containing 1% (v/v) water agar. After germination, two seedlings were transplanted per jar and thinned to one plant after 3 days. Temperature at the greenhouse in controlled by means of air conditioners and average of day and night temperatures were of 28 ± 2.3/23 ± 1.9 °C (day/night); the experiment was performed at the summer growing season, where light intensity is the most adequate for maize growth. Sterile nutrient solution was applied as needed.
At 30 days after transplanting, leaf-chlorophyll contents (CC) were determined according to Kaschuk et al. (2010), based on the “Soil Plant Analysis Development” (SPAD) index, with readings taken from the lowermost third of the +3 leaf (Trani et al. 1983). Biometric parameters of plant height (cm; PH) and culm diameter (mm; CD) of plants were determined with the aid of a digital caliper. Plants were harvested, separating leaves and roots, with three biological replicates. Fresh weight was determined and 2 g of fresh material of each sample were dried at 60 °C for approximately 72 h, until constant weights were achieved; tissues were weighed to estimate the factor for conversion from fresh to dry weight of each replicate. The remaining sampled tissues were frozen in liquid nitrogen and stored at −80 °C until further analyses.
Data obtained were first evaluated for normality and variance homogeneity, followed by the analysis of variance (ANOVA). Tukey’s test was employed to compare means in cases where statistical significance had been detected by the ANOVA F test (p ≤ 0.05). Statistica software version 7.0 was employed.
Identification of phytohormones produced by A. brasilense by UHPLC-HRMS/MS
The identification of phytohormones produced by A. brasilense strains Ab-V5 and Ab-V6 was performed by ultrahigh-performance liquid chromatography-high-resolution mass spectrometry (UHPLC-HRMS/MS). Strains Ab-V5 and Ab-V6 were grown separately in DYGS medium (Rodrigues Neto et al. 1986) without tryptophan (TRP) or in DYGS supplemented with 500 µg mL−1 tryptophan (DYGS-TRP medium). Liquid bacterial inocula were incubated at 28 ± 2 °C with orbital shaking at 120 rpm for 14 days. The bacterial cultures were then filtered through nitrocellulose-membrane filters Millipore HA 0.45 µm to obtain the supernatants. The samples were filtered again in a microfiltration membrane, and 5-µL aliquots of each sample were analyzed. Hormones were identified by mass/charge ratio (m/z) values and by the retention times of the standard compounds indole-3-acetic acid (IAA), indole-3-butyric acid (IBA), indole-3-ethanol (TOL), indole-3-lactic acid (ILA), indole-3-pyruvic acid (IPyA), indole-3-propionic acid (IPA), kinetin (Kin), gibberellic acid (GA3), salicylic acid (SA) and jasmonic acid (JA); tri-methyl-indole-3-acetic acid (TmIAA) was used as internal standard.
RNA extraction, cDNA synthesis and quantitative RT-PCR
RNAs of leaves and roots were extracted with TRIzol® (Life Technologies/Thermo Fisher Scientific), and the concentration and purity were evaluated in a NanoDrop® ND1000 spectrophotometer (NanoDrop-Technologies, Inc.), while the integrity was evaluated by gel electrophoresis. Genomic DNA was removed with DNAseI (Invitrogen™) and the first strand of cDNA was synthesized using SuperscriptIII™ reverse transcriptase (Invitrogen™), according to the manufacturer’s protocol.
Primers for the RT-qPCR targets were designed using primer3Plus (http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi/) (Table 1) to obtain amplicons of 110–150 bp. The endogenous control genes of maize used were UBCE and UBCP, corresponding to the ubiquitin-conjugating enzyme and the ubiquitin carrier protein, respectively (Manoli et al. 2012). These genes were used for data normalization of the cycle threshold (Ct) of RT-qPCR amplifications.
RT-qPCR reactions were performed in a 7500 RT-qPCR thermocycler (Applied Biosystems/Life Technologies). The reactions were performed in triplicate for each of the three biological replicates. The Platinum® SYBR® Green qPCR SuperMix-UDG (Invitrogen™) was used following the manufacturer’s instructions. Cycling conditions were as follows: 50 °C for 2 min, 95 °C for 10 min, 45 cycles at 95 °C for 2 min, 60 °C for 30 s and 72 °C for 30 s, in 45 cycles.
The data obtained were submitted to the Rest2009 software package (Pfaffl et al. 2002), providing a robust statistical analysis (p ≤ 0.05).
Confocal laser scanning microscopy of A. brasilense on maize leaves
Maize leaf colonization by A. brasilense strains Ab-V5 and Ab-V6 expressing the egfp (enconding for enhanced green fluorescent protein) and eyfp (encoding for enhanced yellow fluorescent protein) reporter genes were analyzed by Confocal Laser-Scanning Microscopy (CLSM). First, plasmids pMP4655 (egfp) and pMP4658 (eyfp) (Bloemberg et al. 2000) were transferred by conjugation to A. brasilense Ab-V5 and Ab-V6. To select the transconjugants of A. brasilense, plates with DYGS agar medium (Rodrigues Neto et al. 1986) were supplemented with nalidixic acid (final concentration 40 μg mL−1) and tetracycline (final concentration 20 μg mL−1). The Azospirillum strains exhibit intrinsic resistance to the antibiotic nalidixic acid, whereas Escherichia coli containing the transfer plasmid shows only tetracycline resistance. Transconjugants were obtained for both strains of Azospirillum.
Seeds of maize (hybrid DKB330 VT PRO2) were surface-sterilized (Vincent 1970). Pre-germinated seeds (2 days) were transplanted to test tubes containing 70 mL of sterilized nutrient solution (Fahraeus 1957), and were grown under controlled greenhouse conditions. Mean temperatures during the experiment were of 25/18 °C (day/night) and relative humidity of 70%. At the V2.5 stage of maize growth, plants were singly inoculated by leaf spray with either A. brasilense strain Ab-V5 or Ab-V6 harboring the reporter plasmids expressing egfp and eyfp genes, respectively. Inoculant concentrations applied to the leaves were estimated at 3 × 105 and 7 × 105 cells cm−2 of leaf, for strains Ab-V5 and Ab-V6, respectively. At 1 h, 1 and 2 days after leaf spraying, the leaves were examined for the presence of fluorescent bacteria using CLSM equipped with an Ar–Hg laser (Leica TCS SP2, Leica, Wetzlar, Germany); the filter sets for fluorescence microscopy consisted of a 458-nm band-pass excitation and a 520–560 nm emission. Microscopy analyses were performed on intact alive plant tissues. Simultaneously to the analysis by microscopy, the presence of the bacteria on the leaves surface was verified by evaluation of colony-forming units evaluated by the drop plate method (Miles et al. 1938) 1 h, 1 and 2 days after leaf spraying.
Results
Effects of inoculation of Azospirillum brasilense and their metabolites on plant-growth parameters
In the greenhouse experiment performed to evaluate effects of inoculation on plant growth, it is worth mentioning that all treatments received the same amount of N-fertilizer, corresponding to 50% (60 kg of N ha−1) of the dose recommended for the maize crop in Brazil. When different methods (via seed—SI at sowing or by leaf spray—ILS at the V2.5 stage) of inoculation of A. brasilense strains Ab-V5 and Ab-V6, in single or combined mixtures, or foliar-spray application of their metabolites (MLS), also at the V2.5 stage, were evaluated, statistically significant increases in chlorophyll content (CC) in relation to the non-inoculated control were observed in all treatments except for the SI with Ab-V5; the highest increases were observed in the treatments with MLS of Ab-V6 and MLS of Ab-V5 + Ab-V6, of 109 and 143%, respectively (Table 2). No statistical differences were observed for the parameters of plant high (PH) and culm diameter (CC). Shoot dry weight (SDW) was also improved by all inoculation treatments, except for MLS of Ab-V5. The best inoculation treatment of MLS of strains Ab-V5 and Ab-V6 increased SDW by 72%. In relation to this best treatment (T10), we should mention that the effect might be attributed mainly to the metabolites of Ab-V6, as the single metabolites of Ab-V6 (T9), but not of Ab-V5 (T8), resulted in increases in SDW (Table 2).
Identification of phytohormones produced by A. brasilense by UHPLC-HRMS/MS
UHPLC-HRMS/MS results obtained in the analysis of the supernatants from Azospirillum strains grown in DYGS or DYGS + TRP (tryptophan) media are presented in Table 3. For all samples, supplemented or not with tryptophan, the following main compounds were detected in the metabolites: indole-3-acetic acid (IAA), indole-3-ethanol (TOL), indole-3-lactic acid (ILA) and salicylic acid (SA). Other compounds have also been identified, but in relatively low amounts. In the supernatant of Ab-V5 grown without TRP, we detected gibberellic acid (GA3) and jasmonic acid (JA) and, when supplemented with TRP, we detected indole-3-propionic acid (IPA). In the supernatant of strain Ab-V6 supplied with TRP we detected GA3 (Table 3).
Expression of genes related to defense mechanisms in maize
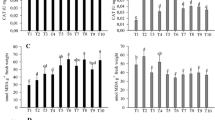
Effects of inoculation with Azospirillum or their metabolites on the expression of genes encoding for antioxidant and PR proteins were determined by RT-qPCR (Figs. 1, 2, 3). When compared to the non-inoculated control (T1), the gene of the cytosolic isoform APX1 in maize leaves was significantly up-regulated by inoculation in all treatments except for with the inoculation with strain Ab-V5 by leaf spray (T5) (Fig. 1a). The highest expression was achieved with treatment T10, with inoculation of metabolites of both strains, with an increase of 2.8-fold in comparison to the non-inoculated control. Contrarily, the expression of APX1 in roots was down-regulated in all treatments (Fig. 1a). The expression of the APX2 gene in leaves was up-regulated in all treatments, and statistically significant in six out of the nine inoculation treatments (Fig. 1b). Contrarily to APX1 gene-expression in roots, APX2 was significantly up-regulated with the metabolites of Ab-V5 (T8), and the metabolites of Ab-V5 + Ab-V6 (T10) (Fig. 1b). The same trend as for APX genes was observed with CAT1 (Fig. 1c). The highest expression in leaves was achieved by seed inoculation with Ab-V5 (T2, 5.5-fold) and spraying of the metabolites of the same strain (T8, 6.9-fold). CAT1 was down-regulated in roots, except for the seed inoculation with Ab-V5 + Ab-V6 (T4) and the metabolite-spray treatments (T8, T9, T10) (Fig. 1c). When SOD genes were investigated, up-regulation in leaves was achieved in all treatments, except for when both strains were leaf sprayed for SOD2 (Fig. 2a) and when Azospirillum cells were leaf sprayed for SOD4 (Fig. 2b). Contrarily, both genes were down-regulated in roots when living cells were applied to seeds or sprayed, whereas the application of metabolites on leaves resulted in up-regulation, with the highest expression of 2.5-fold for SOD2 and of 3.2-fold for SOD4 with the metabolites of Ab-V5 (Fig. 2a, b).

RT-qPCR analysis of the expression of a APX1, b APX2 and c CAT genes in maize leaves and roots when induced by Azospirillum brasilense strains Ab-V5 and Ab-V6 inoculated on seeds or by foliar spray, and also buy their metabolites applied by foliar spray. Data ± standard deviation from three biological replicates, each with three technical replicates. Data were normalized in relation to the endogenous control (UBCE and UBCP). The asterisks indicate statistically significant expression at the level α = 5%, determined by REST2009 software. Black bars leaf and dark gray bars root

RT-qPCR analysis of the expression of a SOD2 and b SOD4 genes, as described in the legend of Fig. 1

RT-qPCR analysis of the expression of several a PR1, b prp2 and c prp4 genes, as described in the legend of Fig. 1
Analyzing the PR group of genes (PR1, prp2 and prp4) (Fig. 3a–c), in general seed inoculation with single strains (Ab-V5 or Ab-V6) up-regulated gene expression in leaves, whereas seed co-inoculation, and foliar inoculation with single strains down-regulated the genes. Seed inoculation with Ab-V5 (T2) increased by 10.4-fold the expression of PR1 gene in leaves, whereas, with Ab-V6 (T3), up-regulation was of 5.1- and 5.5-fold for PR1 and prp2, respectively. In relation to the effects of metabolite sprays on gene expression in leaves, emphasis should be given to the Ab-V5 + Ab-V6 treatment (T10), always showing up-regulation, in particular of prp2 (6.4-fold). In relation to the gene expression in roots (Fig. 3a–c), in general all treatments resulted in up-regulation, but emphasis should be given to the co-inoculation of seeds on the expression of PR1 (20.2-fold) and prp4 (4.2-fold), respectively; down-regulation of prp2 with the metabolites of Ab-V6 of Ab-V5 + Ab-V6 was not statistically significant (Fig. 3b).
Colonization of maize leaves by A. brasilense
In order to check whether A. brasilense cells are able to colonize maize leaves, strains Ab-V5 and Ab-V6—harboring reporter plasmids expressing egfp and eyfp genes, respectively—were inoculated by leaf spray. After 1 h, 1 and 2 days of inoculation, the leaves were visualized by CLSM (Fig. 4). After 1 h of inoculation with both strains, (EGFP)-l and (EYFP)-labelled cells indicated that they were able to colonize leaves surface (Fig. 4a, d), and the same was observed after 1 day of inoculation (Fig. 4b, c). However, after 2 days of inoculation, we were unable to detect the strains on the leaf surfaces. Simultaneously, bacteria counts on leaves surface were performed after 1 h, 1 and 2 days of leaf spraying. Values obtained for colony-forming units (CFUs) were as follows: 2 × 105, 1 × 105 and 5 × 102 CFUs cm−2 of leaf for strain Ab-V5 and 6 × 105, 5 × 105 and 5 × 102 CFU cm−2 of leaf for strain Ab-V6 after 1 h, 1 and 2 days, respectively. The low bacterial counts at 2 days after inoculation might explain why the bacteria were not visualized by CLSM.

Confocal laser scanning microscopy analysis of maize leaf surface colonization by A. brasilense expressing EGFP when inoculated by leaf spray. a A. brasilense Ab-V5 after 1 h, b A. brasilense Ab-V5 after 1 day, c A. brasilense Ab-V6 after 1 h, d A. brasilense Ab-V6 after 1 day of inoculation
Discussion
When maize growth was evaluated under greenhouse conditions, the benefits of inoculation with A. brasilense Ab-V5 and/or Ab-V6 applied to seeds or by foliar application at the V2.5 stage of plant growth were confirmed. The benefits of inoculation with Azospirillum at sowing, via seeds or in-furrow, have been demonstrated under greenhouse and field conditions in cereals, with an emphasis on maize (Dobbelaere and Okon 2007; Hungria et al. 2010; Hungria 2011; Okon et al. 2015; Fukami et al. 2016), and increasing use of strains Ab-V5 and Ab-V6 has been exponential in Brazil since 2010 (Hungria 2011). Improvements in grain yields of maize and wheat by foliar application of Azospirillum have also been reported (Clemente et al. 2016; Fukami et al. 2016), but the physiological and genetic basis of such improvements have yet to be elucidated.
Intriguing were the positive responses observed in our study to foliar application of metabolites of Azospirillum—especially with Ab-V5 + Ab-V6—at the V2.5 stage. Previously, we reported benefits to the maize crop by the application of metabolites of rhizobia, suggesting that the effects could be attributed to lipo-chitooligosaccharides (LCOs) or Nod factors (Marks et al. 2013, 2015) synthesized by the bacteria. Positive effects with application of Nod factors in maize, cotton (Gossypium hirsutum) and beet (Beta vulgaris) were also reported by Smith et al. (2015). However, as far as we are aware, this is the first scientific report of effects of Azospirillum metabolites on cereal growth.
To achieve a better understanding of the effects of leaf spraying with Azospirillum cells, we investigated the bacterial colonization of leaves by microscopy. Strains Ab-V5 and Ab-V6 were detected on leaves surfaces up to 24 h after inoculation, but the numbers of surviving cells (CFU) were markedly reduced, and, after 48 h, cells were not detected by microscopy. It is possible that the number of recovered cells after 24 h was too low to be detected by CLSM, but the mortality in 24 h was of the order of 1000-fold. Furthermore, we must bear in mind that our experiment was performed under controlled optimized conditions, and that mortality under stressful field conditions—UV light, desiccation, high temperature—would certainly be far higher. Therefore, it is reasonable to suggest that the benefits observed in our study from foliar spraying of Azospirillum cells resulted from metabolites present in the inoculant rather than from the living cells.
The first hypothesis to explain increased plant growth by spraying cells or metabolites of A. brasilense Ab-V5 and Ab-V6 relies on phytohormone production. We have identified the main molecules in the supernatants of the Ab-V5 and Ab-V6 strains, induced and non-induced with tryptophan, as being indole-3-acetic acid (IAA), indole-3-ethanol (TOL), indole-3-latic acid (ILA) and salicylic acid (SA). Although the physiological functions of TOL and ILA remain unknown, it is possible that intermediates of IAA biosynthesis pathways are converted into these storage compounds whenever necessary (Cassán et al. 2014). In addition, in some combinations of strains and tryptophan we detected traces of gibberellic acid (GA3) and jasmonic acid (JA). The synthesis of phytohormones by Azospirillum has been broadly reported, and may differ between species and strains. The well studied A. brasilense strains Cd and Az39 produce IAA, zeatin, GA3, abscisic acid and ethylene (Perrig et al. 2007), strain UAP154 produces IAA and indole-butyric acid (IBA) (Martínez-Morales et al. 2003), strain 703Ebc produces IAA, TOL, ILA and indole-3-methanol (Crozier et al. 1988), and Sp13t produces IAA, ILA, GA3 and kinetin (Tien et al. 1979). Tien et al. (1979) also detected gibberellin-like molecules in the supernatants of A. brasilense Sp13t at low concentrations, of about 0.05 µg of GA3 mL−1. However, when applied at concentrations as low as 0.005 µg mL−1 to lettuce (Lactuca sativa), hypocotyls elongation was promoted and, in pearl millet (Pennisetum americanum L.), the number of lateral roots was increased. The benefits confirmed in our study of inoculation of seed with Azospirillum at sowing may be attributed to the effects of phytohormones in the rhizosphere, and we propose that these effects also occur from the application of cells and metabolites to the leaves.
Plants synthesize a variety of secondary metabolites that are involved in several physiological processes, and main functions of these compounds lie in providing stress tolerance and defense against pathogens (Sudha and Ravishankar 2002). Previous studies have reported that maize inoculation with Azospirillum results in significant changes in the secondary metabolic profiles of roots and shoots, suggesting the presence of finely-tuned interacting mechanisms (Walker et al. 2011). In addition, reactive oxygen species (ROS) in plants contribute to resisting biotic stresses such as pathogens and even symbiotic bacteria (before plant perceives benefit from the symbiosis) (Lamb and Dixon 1997; Santos et al. 2001), as well as to tolerating abiotic stresses (Ozyigit et al. 2016), such as saline conditions (Barakat 2011). However, ROS accumulation results in oxidative damage to cells such as lipid peroxidation with membrane destruction, protein inactivation or DNA mutation (Garcı́a-Limones et al. 2002). Oxidative stress is relieved in plants by antioxidant enzymes such as catalase, superoxide dismutase and ascorbate peroxidase (Wisniewski-Dyé et al. 2012; Ozyigit et al. 2016). The genes encoding the isoenzymes are found in different plant-cell compartments, such as the cytosolic SOD2, SOD4 (Jung et al. 2001), APX1 and APX2, which are inducible mainly under extreme light or heat-stress conditions (Davletova et al. 2005), and CAT1, found in peroxisomes, glyoxysomes and also in the cytosol (Scandalios et al. 1997; Jung et al. 2001). We evaluated the effects of Azospirillum and its metabolites on the expression of genes related to the synthesis of the H2O2-generating enzyme (SOD), the H2O2-scavenging enzymes (CAT and APX) in maize leaves and roots. In general, inoculation of seeds with A. brasilense and by foliar spraying resulted in down-regulation transcription of oxidative stress genes (APX1, APX2, SOD2, SOD4) in roots, but genes were always up-regulated by leaf spray of metabolites, except for APX1. The results suggest that oxidative stress in roots persisted longer with the application of living cells than with their metabolites. Seed inoculation up-regulated all genes in leaves, but when cells were sprayed on leaves, SOD4 with all strains and APX1 with Ab-V5 were down-regulated. Similarly to the roots, when the metabolites were sprayed on the leaves the genes—now including APX1—were up-regulated. The up-regulation of APX1 in leaves is particularly interesting, as APX genes might be essential for chloroplast protection during light stress (Pnueli et al. 2003; Mittler et al. 2004; Davletova et al. 2005).
Another defense mechanism of the plants is mediated by ISR (induced systemic resistance), resulting in plant resistance to some pathogenic bacteria, viruses and fungi (Lugtenberg and Kamilova 2009). ISR is triggered by non-pathogenic microorganisms and starts in primary infected parts, extending to other plant tissues (Dutta et al. 2008). Biochemical or physiological changes in plants include induced accumulation of pathogenesis-related (PR) proteins that have different functions like the proteins encoded by PR1 (a member of a multigene family) (Morris et al. 1998), PR-2 (a β-1-3-glucanase) (Kauffmann et al. 1987), PR4 (a chitinase family) (Nasser et al. 1988). Transcriptome studies of PR genes with Azospirillum sp. B510 applied as inoculum to rice (O. sativa L.) reported that one gene was up- and five were down-regulated (Drogue et al. 2014). In another study with Arabidopsis thaliana inoculated with A. brasilense Sp245, PR genes were also up-regulated (Spaepen et al. 2014). In our study, seed inoculation resulted in significant up-regulation of only one PR gene in roots, PR1, while foliar application in general resulted in up-regulation of PR1, prp2 and prp4 genes on roots. Up-regulation of PR1 and prp4 was also verified with metabolite spray. In relation to the gene expression in leaves, emphasis should be given to single-seed inoculation with both strains that up-regulated all PR genes. Interestingly, it has been shown that the use of more than one microorganism optimized ISR responses in pigeon pea (Cajanus cajan) (Dutta et al. 2008), similarly to our results with seed inoculation of Ab-V5 + Ab-V6 on roots. Bacillus subtilis also up-regulated PR1 and PR4, but not SOD2 genes in maize roots (Gond et al. 2015). It is also worth mentioning that ISR responses in different tissues from those where the microorganism is applied occurs, e.g. leaf spray with Pseudomonas fluorescens in rice induced ISR against the soil-borne plant pathogen Rhizoctonia solani (Vidhyasekaran and Muthamilan 1999).
ISR responses to a variety of plant pathogens usually have been associated with the signaling compounds jasmonate and ethylene (Glick 2012; Ahemad and Kibret 2014), the levels of which are increased in tissue independent of SA (Van Loon 2007); this mechanism has also been reported in the association of Azospirillum sp. B510 with rice (Yasuda et al. 2009). Indeed, several studies have demonstrated that exogenous applications of SA (Bari and Jones 2009) and JA (Agrawal et al. 2000; Lorenzo and Solano 2005; Wasternack 2007; Bari and Jones 2009) induce PR genes and consequently increase the resistance to several pathogens. In addition, exogenous applications of JA also increase the activities of CAT and SOD enzymes in soybean [Glycine max (L.) Merr.] plants stressed by cadmium (Noriega et al. 2012). The ISR might be related also to the reported effects of A. brasilense against soil-borne plant pathogens such as Rhizoctonia spp. (Russo et al. 2008) and Fusarium oxysporum f. sp. matthiolae (Somers et al. 2005).
It is worth considering that the exogenous application of synthetic growth regulators (e.g. IAA, GA, kin) has been broadly adopted by foliar spraying due to plant-growth promotion (Halmann 1990), but the commercial products are usually very expensive. However, in our study, the foliar spray of Azospirillum metabolites in general improved not only plant growth, but also up-regulated plant genes related to defense mechanisms, and might represent an alternative biological plant regulator.
In conclusion, we reported that, regardless of the method of inoculation—on seeds or by foliar application—the A. brasilense strains Ab-V5 and Ab-V6 promoted plant growth. Intriguingly, the foliar application of their metabolites also improved growth. The benefits of cell and metabolite application can be attributed both to the synthesis of phytohormones and to the induction of plant defense-related genes. Clearly, the application of biological low-cost inoculants containing Azospirillum cells or their metabolites, promoting plant growth and eliciting plant resistance to biotic and abiotic stresses, have important agronomic implications.
References
Agrawal GK, Jwa NS, Rakwal R (2000) A novel rice (Oryza sativa L.) acidic PR1 gene highly responsive to cut, phytohormones, and protein phosphatase inhibitors. Biochem Biophys Res Commun 274:157–165
Ahemad M, Kibret M (2014) Mechanisms and applications of plant growth promoting rhizobacteria: current perspective. J King Saud Univ Sci 26:1–20
Ardakani MR, Mazaheri D, Mafakheri S, Moghaddam A (2011) Absorption efficiency of N, P, K through triple inoculation of wheat (Triticum aestivum L.) by Azospirillum brasilense, Streptomyces sp., Glomus intraradices and manure application. Physiol Mol Biol Plants 17(2):181–192
Asada K (1999) The water–water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annu Rev Plant Physiol Plant Mol Biol 50:601–639
Barakat NAM (2011) Oxidative stress markers and antioxidant potential of wheat treated with phytohormones under salinity stress. J Stress Physiol Biochem 7(4):250–267
Bari R, Jones JDG (2009) Role of plant hormones in plant defence responses. Plant Mol Biol 69(4):473–488
Bashan Y, de-Bashan LE (2010) How the plant growth-promoting bacterium Azospirillum promotes plant growth—a critical assessment. Adv Agron 108:77–136
Bashan Y, Holguin G (1998) Proposal fot the division of plant growth-promoting rhizobacteria into two classifications: biocontrol-PGPB (plant growth-promoting bacteria) and PGPB. Soil Biol Biochem 30(8/9):1225–1228
Bloemberg GV, Wijfjes AHM, Lamers GEM, Stuurman N, Lugtenberg BJJ (2000) Simultaneous imaging of Pseudomonas fluorescens WCS365 populations expressing three different autofluorescent proteins in the rhizosphere: new perspectives for studying microbial communities. Mol Plant Microbe Interact 13(11):1170–1176
Bottini R, Fulchieri M, Pearce D, Pharis RP (1989) Identification of gibberellins A1, A3, and iso-A3 in cultures of Azospirillum lipoferum. Plant Physiol 90:45–47
Bowler C, Montagu MV, Inzé D (1992) Superoxide dismutase and stress tolerance. Annu Rev Plant Physiol Plant Mol Biol 43:83–116
Cassán F, Vanderleyden J, Spaepen S (2014) Physiological and agronomical aspects of phytohormone production by model plant-growth-promoting rhizobacteria (PGPR) belonging to the genus Azospirillum. J Plant Growth Regul 33:440–459
Cassán FD, Okon Y, Creus CM (2015) Handbook for Azospirillum. Springer, Basel
Clemente JA, Condé AB, Andrade AT, Cardoso CR, Flor IM, Martins AD, Lima WT, Oliveira CB (2016) Azospirillum brasilense and nitrogen fertilization affecting wheat productivity. Afr J Agric Res 11:2179–2184
Crozier A, Arruda P, Jasmin JM, Monteiro AM, Sandberg G (1988) Analysis of indole-3-acetic acid and related indoles in culture medium from Azospirillum lipoferum and Azospirillum brasilense. Appl Environ Microbiol 54(11):2833–2837
Davletova S, Rizhsky L, Liang H, Shengqiang Z, Olivier DJ, Coutu J, Shulaev V, Schlauch K, Mittler R (2005) Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsis. Plant Cell 17:268–281
Dobbelaere S, Okon Y (2007) The plant growth-promoting effect and plant responses. In: Elmerich C, Newton WE (eds) Associative and endophytic nitrogen-fixing bacteria and cyanobacterial associations. Springer, Dordrecht
Drogue B, Sanguin H, Chamam A, Mozar M, Llauro C, Panaud O, Prigent-Combaret C, Picault N, Wisniewski-Dyé F (2014) Plant root transcriptome profiling reveals a strain-dependent response during Azospirillum-rice cooperation. Front Plant Sci 5:1–14
Dutta S, Mishra AK, Kumar BSD (2008) Induction of systemic resistance against fusarial wilt in pigeon pea through interaction of plant growth promoting rhizobacteria and rhizobia. Soil Biol Biochem 40:452–461
Fahraeus G (1957) The infection of clover root hairs by nodule bacteria studied by a simple glass slide technique. J Gen Microbiol 16:374–381
Fukami J, Nogueira MA, Araujo RS, Hungria M (2016) Accessing inoculation methods of maize and wheat with Azospirillum brasilense. AMB Express 6:3
GarcÍa-Limones C, Hervás A, Navas-Cortés JA, Jiménez-Díaz RM (2002) Induction of an antioxidant enzyme system and other oxidative stress markers associated with compatible and incompatible interactions between chickpea (Cicer arietinum L.) and Fusarium oxysporum f. sp. ciceris. Physiol Mol Plant Pathol 61:325–337
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48(12):909–930
Glick BR (2012) Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012:1–15
Gond SK, Bergen MS, Torres MS, White JF Jr (2015) Endophytic Bacillus spp. produce antifungal lipopeptides and induce host defence gene expression in maize. Microbiol Res 172:79–87
Gupta S, Arora DK, Srivastava AK (1995) Growth promotion of tomato plants by Rhizobacteria and imposition of energy stress on Rhizoctonia solani. Soil Biol Biochem 27(8):1051–1058
Halmann M (1990) Synthetic plant growth regulators. Adv Agron 43:47–105
Hickman JS, Shroyer JP (1994) Corn production handbook. Publication C, Manhattan
Hodges DM, Andrews CJ, Johnson DA, Hamilton RI (1996) Antioxidant compound response to chilling stress in differentially sensitive inbred maize line. Physiol Plant 98:685–692
Hungria M (2011) Inoculação com Azospirillum brasilense: inovação em rendimento a baixo custo. Circular Técnica 325. Embrapa Soja, Londrina
Hungria M, Campo RJ, Souza EM, Pedrosa FO (2010) Inoculation with selected strains of Azospirillum brasilense and A. lipoferum improves yields of maize and wheat in Brazil. Plant Soil 331:413–425
Hungria M, Nogueira MA, Araujo RS (2016) Inoculation of Brachiaria spp. with the plant growth-promoting bacterium Azospirillum brasilense: an environment-friendly component in the reclamation of degraded pastures in the tropics. Agric Ecosyst Environ 221:125–131
Jung S, Kernodle SP, Scandalios JG (2001) Differential antioxidant responses to norflurazon-induced oxidative stress in maize. Redox Rep 6(5):311–317
Kaschuk G, Hungria M, Leffelaar PA, Giller KE, Kuyper TW (2010) Differences in photosynthetic behaviour and leaf senescence of soybean (Glycine max [L.] Merrill) dependent on N2 fixation or nitrate supply. Plant Biol 12(1):60–69
Kauffmann S, Legrand M, Geoffroy P, Fritig B (1987) Biological function of ‘pathogenesis-related’ proteins: four PR proteins of tobacco have 1,3-β-glucanase activity. EMBO J 6(11):3209–3212
Lamb C, Dixon RA (1997) The oxidative burst in plant disease resistance. Annu Rev Plant Physiol Plant Mol Biol 48:251–275
Lorenzo O, Solano R (2005) Molecular players regulating the jasmonate signalling network. Curr Opin Plant Biol 8(5):532–540
Lugtenberg B, Kamilova F (2009) Plant-growth-promoting rhizobacteria. Annu Rev Microbiol 63:541–556
Manoli A, Sturaro A, Trevisan S, Quaggiotti S, Nonis A (2012) Evaluation of candidate reference genes for qPCR in maize. J Plant Physiol 169:807–815
Marks BB, Megías M, Nogueira MA, Hungria M (2013) Biotechnological potential of rhizobial metabolites to enhance the performance of Bradyrhizobium spp. and Azospirillum brasilense inoculants with soybean and maize. AMB Express 3:21
Marks BB, Megías M, Ollero FJ, Nogueira MA, Araujo RS, Hungria M (2015) Maize growth promotion by inoculation with Azospirillum brasilense and metabolites of Rhizobium tropici CIAT 899 enriched on lipo-chitooligosaccharides (LCOs). AMB Express 5:71
Marques ACR, Oliveira LB, Nicoloso FT, Jacques JS, Giacomini SJ, Quadros FLF (2017) Biological nitrogen fixation in C4 grasses of different growth strategies of South America natural grasslands. Appl Soil Ecol 113:54–62
Martínez-Morales LJ, Soto-Urzúa L, Baca BE, Sánchez-Ahédo JA (2003) Indole-3-butyric acid (IBA) production in culture medium by wild strain Azospirillum brasilense. FEMS Microbiol Lett 228(2):167–173
Miles AA, Misra SS, Irwin JO (1938) The estimation of the bactericidal power of the blood. Epidemiol Infect 38(6):732–749
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F (2004) Reactive oxygen gene network of plants. Trends Plant Sci 9(10):490–498
Morris SW, Vernooij B, Titatarn S, Starrett M, Thomas S, Wiltse CC, Frederiksen RA, Bhandhufalck A, Hulbert S, Uknes S (1998) Induced resistance responses in maize. Mol Plant Microbe Interact 11(7):643–658
Nasser W, Tapia M, Kauffmann S, Montasser-Kouhsari S, Burkard G (1988) Identification and characterization of maize pathogenesis-related proteins. Four maize PR proteins are chitinases. Plant Mol Biol 11(4):529–538
Noriega G, Cruz DS, Batlle A, Tomaro M, Balestrasse K (2012) Heme oxygenase is involved in the protection exerted by jasmonic acid against cadmium stress in soybean roots. J Plant Growth Regul 31:79–89
Okon Y, Labandera-Gonzales C, Lage M, Lage P (2015) Agronomic applications of Azospirillum and other PGPR. In: de Brujin FJ (ed) Biological nitrogen fixation. Wiley, Hoboken
Ozyigit II, Filiz E, Vatansever R, Kurtoglu KY, Koc I, Öztürk MX, Anjum NA (2016) Identification and comparative analysis of H2O2-scavenging enzymes (ascorbate peroxidase and glutathione peroxidase) in selected plants employing bioinformatics approaches. Front Plant Sci 7:1–23
Pereg L, de-Bashan LE, Bashan Y (2016) Assessment of affinity and specificity of Azospirillum for plants. Plant Soil 399:389–414
Perrig D, Boiero ML, Masciarelli OA, Penna C, Ruiz OA, Cassán FD, Luna MV (2007) Plant-growth-promoting compounds produced by two agronomically important strains of Azospirillum brasilense, and implications for inoculant formulation. Appl Microbiol Biotechnol 75:1143–1150
Pfaffl MW, Horgan GW, Dempfle L (2002) Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 30(9):e36
Pnueli L, Liang H, Rozenberg M, Mittler R (2003) Growth suppression, altered stomatal responses, and augmented induction of heat shock proteins in cytosolic ascorbate peroxidase (Apx1)-deficient Arabidopsis plants. Plant J 34:187–203
Robert-Seilaniantz A, Grant M, Jones JDG (2011) Hormone crosstalk in plant disease and defense: more than just jasmonate–salicylate antagonism. Annu Rev Phytopathol 49:317–343
Rodrigues Neto J, Malavolta VA Jr, Victor O (1986) Meio simples para o isolamento e cultivo de Xanthomonas campestris pv. citri tipo B. Summa Phytopathol 12:32
Rodriguez H, Gonzalez T, Goire I, Bashan Y (2004) Gluconic acid production and phosphate solubilization by the plant growth-promoting bacterium Azospirillum spp. Naturwissenschaften 91:552–555
Romero AM, Correa OS, Moccia S, Rivas JG (2003) Effect of Azospirillum-mediated plant growth promotion on the development of bacterial diseases on fresh-market and cherry tomato. J Appl Microbiol 95:832–838
Russo A, Vettori L, Felici C, Fiaschi G, Morini S, Toffanin A (2008) Enhanced micropropagation response and biocontrol effect of Azospirillum brasilense Sp245 on Prunus cerasifera L. clone Mr.S 2/5 plants. J Biotechnol 134:312–319
Santos R, Hérouart D, Sigaud S, Touati D, Puppo A (2001) Oxidative burst in alfalfa-Sinorhizobium meliloti symbiotic interaction. Mol Plant Microbe Interact 14:86–89
Sahoo RK, Ansari MW, Pradhan M, Dangar TK, Mohanty S, Tuteja N (2014) Phenotypic and molecular characterization of native Azospirillum strains from rice fields to improve crop productivity. Protoplasma 251(4):943–953
Scandalios JG, Guan L, Polidoros AN (1997) Catalases in plants: gene structure, properties, regulation, and expression. In: Scandalios JG (ed) Oxidative stress and the molecular biology of antioxidant defenses. Cold Spring Harbor Laboratory, New York
Smith S, Habib A, Kang Y, Legget M, Diaz-Zorita M (2015) LCO applications provide improved responses with legumes and nonlegumes. In: de Bruijn FJ (ed) Biological nitrogen fixation. Wiley, Hoboken
Somers E, Ptacek D, Gysegom P, Srinivasan M, Vanderleyden J (2005) Azospirillum brasilense produces the auxin-like phenylacetic acid by using the key enzyme for indole-3-acetic acid biosynthesis. Appl Environ Microbiol 71(4):1803–1810
Spaepen S, Vanderleyden J (2015) Auxin signaling in Azospirillum brasilense: a proteome analysis. In: de Brujin FJ (ed) Biological nitrogen fixation. Wiley, Hoboken
Spaepen S, Bossuyt S, Engelen K, Marchal K, Vanderleyden J (2014) Phenotypical and molecular responses of Arabidopsis thaliana roots as a result of inoculation with the auxin-producing bacterium Azospirillum brasilense. New Phytol 201(3):850–861
Sudha G, Ravishankar GA (2002) Involment and interaction of various signaling compounds on the plant metabolic events during defense response, resistance to stress factors, formation of secondary metabolites and their molecular aspects. Plant Cell Tissue Organ Cult 71:181–212
Tien TM, Gaskins MH, Hubbell DH (1979) Plant growth substances produced by Azospirillum brasilense and their effect on the growth of Pearl Millet (Pennisetum americanum L.). Appl Environ Microbiol 37(5):1016–1024
Tortora ML, Díaz-Ricci JC, Pedraza RO (2011) Protection of strawberry plants (Fragaria ananassa Duch.) against anthracnose disease induced by Azospirillum brasilense. Plant Soil 356:279–290
Trani PE, Hiroce R, Bataglia OC (1983) Análise foliar: amostragem e interpretação. Fundação Cargill, Campinas
van Loon LC (2007) Plant responses to plant growth-promoting rhizobacteria. Eur J Plant Pathol 119(3):243–254
van Loon LC, Bakker PAHM (2005) Induced systemic resistance as a mechanism of disease suppression by rhizobacteria. In: Siddiqui ZA (ed) PGPR: biocontrol and biofertilization. Springer, Netherlands
Vidhyasekaran P, Muthamilan M (1999) Evaluation of a powder formulation of Pseudomonas fluorescens Pf1 for control of rice sheath blight. Biocontrol Sci Technol 9:67–74
Vincent JM (1970) A manual for the practical study of root-nodule bacteria. Blackwell, Oxford
Walker V, Bertrand C, Bellvert F, Moënne-Loccoz Y, Bally R, Comte G (2011) Host plant secondary metabolite profiling shows a complex, strain-dependent response of maize to plant growth-promoting rhizobacteria of the genus Azospirillum. New Phytol 189:494–506
Wasternack C (2007) Jasmonates: an update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann Bot 100(4):681–697
Wisniewski-Dyé F, Lozano L, Acosta-Cruz E, Borland S, Drogue B, Prigent-Combaret C, Rouy Z, Barbe V, Herrera AM, González V, Mavingui P (2012) Genome sequence of Azospirillum brasilense CBG497 and comparative analyses of Azospirillum core and accessory genomes provide insight into niche adaptation. Genes 3:576–602
Yasuda M, Isawa T, Shinozaki S, Minamisawa K, Nakashita H (2009) Effects of colonization of a bacterial endophyte, Azospirillum sp. B510, on disease resistance in rice. Biosci Biotechnol Biochem 73(12):2595–2599
Authors’ contributions
JF, FJO, MM and MH initiated and designed the study. FJO and MH contributed with reagents/materials. JF performed the experiments. JF, FJO, MM and MH analyzed the data and wrote the paper. All authors read and approved the final manuscript.
Acknowledgements
Authors thank Dr. Allan R.J. Eaglesham for English suggestions, J. Fukami acknowledges a Ph.D. fellowship and a sandwich fellowship to Spain from CAPES. M. Hungria is also a fellow from CNPq.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All data and materials cited on the manuscript are freely available for the scientific community.
Consent for publication
All authors gave the consented for publication.
Ethics approval and consent of participation
The study has not involved any human or animal participation or data.
Funding information
Financed by Embrapa (02.13.08.003.00.00), INCT-Plant-Growth Promoting Microorganisms for Agricultural Sustainability and Environmental Responsibility (465133/2014-2; Fundação Araucária) and Ministerio de Economía y Competitividad (Mineco, AGL2016-77163-R).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Fukami, J., Ollero, F.J., Megías, M. et al. Phytohormones and induction of plant-stress tolerance and defense genes by seed and foliar inoculation with Azospirillum brasilense cells and metabolites promote maize growth. AMB Expr 7, 153 (2017). https://doi.org/10.1186/s13568-017-0453-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13568-017-0453-7