
Abstract
Background
Understanding the ecology of larval malaria and lymphatic filariasis mosquitoes in a changing environment is important in developing effective control tools or programmes. This study characterized the breeding habitats of Anopheles mosquitoes in rural communities in different ecological zones in Ghana during the dry and rainy seasons.
Methods
The spatio-temporal distribution, species composition, and abundance of larval Anopheles mosquitoes in breeding habitats were studied in five locations in three ecological zones of Ghana. These were Anyakpor (coastal savannah area), Duase (forest area), and Libga, Pagaza, and Kpalsogu (Sahel savannah area). Larvae were collected using standard dippers and were raised in the insectary for identification.
Results
Out of a total of 7984 mosquito larvae collected, 2152 (27.26%) were anophelines and were more abundant in the rainy season (70.82%) than in the dry season (29.18%). The anophelines comprised 2128 (98.88%) An. gambiae s.l., 16 (0.74%) An. rufipes, and 8 (0.37%) An. pharoensis. In the coastal savannah and forest zones, dug-out wells were the most productive habitat during the dry (1.59 larvae/dip and 1.47 larvae/dip) and rainy seasons (11.28 larvae/dip and 2.05 larvae/dip). Swamps and furrows were the most productive habitats in the Sahel savannah zone during the dry (0.25 larvae/dip) and rainy (2.14 larvae/dip) seasons, respectively. Anopheles coluzzii was the most abundant sibling species in all the ecological zones. Anopheles melas and An. arabiensis were encountered only in the coastal savannah and the Sahel savannah areas, respectively. Larval habitat types influenced the presence of larvae as well as larval density (p < 0.001). The land-use type affected the presence of Anopheles larvae (p = 0.001), while vegetation cover influenced larval density (p < 0.05).
Conclusion
The most productive habitats were dug-out wells in the coastal savannah and forest zones, and furrows from irrigated canals in the Sahel savannah zone. Anopheles coluzzii was the predominant vector species in all the ecological zones. The abundance of Anopheles breeding habitats and larvae were influenced by anthropogenic activities. Encouraging people whose activities create the larval habitats to become involved in larval source management such as habitat manipulation to stop mosquito breeding will be important for malaria and lymphatic filariasis control.
Similar content being viewed by others


Background
Anopheles mosquitoes are important vectors that transmit diseases including malaria and lymphatic filariasis, among others [1]. The distribution and abundance of adult Anopheles mosquitoes are predicated on the presence and productivity of larval breeding habitats [2]. Species of the Anopheles gambiae complex prefer to breed in shallow water collections that are open to sunlight [3]. Their breeding habitats can have varying sizes of water bodies that are natural or man-made, permanent or temporary, freshwater or saline [2, 4]. Anopheles funestus, on the other hand, prefer to breed in shady permanent or semi-permanent water bodies, usually with floating or emergent vegetation such as that found in swamps, marshes, and edges of streams [5, 6].
The choice of oviposition sites of mosquitoes is influenced by myriad environmental factors, which include climatic components such as temperature, rainfall, vegetation, salinity and turbidity of the water, the size of the habitat, and the amount of sunlight [2]. The temperature of larval habitats can influence larval development, pupation rate and time, and larval survivorship [2, 7]. Variation in rainfall patterns or seasonal changes can also affect the availability of larval habitats and larval productivity [3].
Vector control is key to the elimination of vector-borne diseases such as malaria and lymphatic filariasis [8, 9]. Even though the most widely used vector control methods—long-lasting insecticide nets (LLINs) and indoor residual spraying (IRS)—have reduced the transmission of malaria in Africa [10, 11], these methods have not been successful in malaria eradication because of the emergence and rapid spread of insecticide resistance in mosquitoes [12,13,14]. Also, the use of LLINs and IRS which target indoor-biting and indoor-resting mosquitoes have driven behavioural changes in the Anopheles mosquito from indoor, late-night biting to early biting times when humans might be unprotected outside [15, 16]. Nevertheless, larval source management or source control could provide an additional valuable tool for the control of malaria vectors [17]. To assess the feasibility of larval control or larval source management, there is the need to assess the abundance of different types of habitats, measure the productivity in each habitat type [18], and also understand how these different habitat types are formed and how they interact with the society.
The presence and density of mosquito larvae, and consequently the number of competent adult malaria vectors, are regulated by a variety of ecosystem processes interacting at different levels and spatio-temporal scales. These include the presence of water and aquatic plants that protect larvae from predators and serve as detritus that support microbial communities, which in turn serve as food for mosquito larvae [19]. Changes in the structure of the ecosystem can have a considerable impact on mosquito populations and species distribution. Therefore, studies on the ecology of larval habitats should include a landscape context [2, 3, 20, 21]. Landscape features such as topography, land cover, and land use influence the formation, distribution, and microclimate conditions of larval habitats, which in turn influence the distribution of adult Anopheles vectors [2, 22,23,24,25]. Human activities can affect habitat distribution and stability through landscape changes such as deforestation, irrigation, and agricultural practices [2].
There are three main ecological zones in Ghana—the coastal savannah in southern Ghana, the forest in central Ghana, and the Sahel savannah in northern Ghana. These ecological zones affect the distribution of habitats and, importantly, species composition [26]. The coastal savannah and forest zones have a bimodal rainfall pattern, allowing for two peaks of malaria transmission, while the Sahel zone has a unimodal rainfall pattern, giving rise to seasonal malaria transmission.
The ecology of larval mosquitoes has implications for vector control, hence the need to understand the productivity and dynamics of larval habitats in the changing environment in order to model and predict the abundance of adult mosquitoes and ultimately develop effective control tools or programmes [2, 27]. The aim of this study, therefore, was to investigate the ecology of Anopheles mosquito larvae in different ecological zones in Ghana. The availability of larval habitats, their productivity, and distribution among different zones in Ghana was studied. The spatio-temporal distribution and species composition of larval malaria and lymphatic filariasis vectors were investigated.
Methods
Study sites
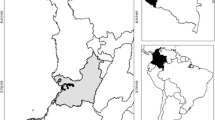
The study was conducted in five locations in three ecological landscapes of Ghana—the coastal savannah, the forest, and the Sahel savannah zones (Fig. 1). Anyakpor (5°46′51.96″N, 0°35′12.84″E) was the site in the coastal savannah area. It is a rural coastal community about 5 km west of Ada Foah in southern Ghana, and has a dry equatorial climate with temperatures ranging from 23 °C to 28 °C throughout the year and maximum temperatures reaching 33 °C. Its rainfall pattern is bimodal, with a long rainy season from April to June and a short rainy season from September to November. It has coastal savannah type vegetation. The community is divided into two main parts—the fishing and farming communities. The farmers are involved in vegetable farming on raised beds. Because it is a low-lying area with a high water table, farmers in Anyakpor in coastal Ghana dig wells and ponds to get underground water for irrigation purposes. The nature of the soil of the communities along the coast makes it difficult to hold water; as a result, there are little to no breeding habitats within the settlement areas of the community throughout the year. An. gambiae s.l. is the dominant malaria vector in the coastal savannah zone, followed by An. funestus [28].

Map of study area
Duase (6°32′3.05″N, 1°14′42.22″W) was the site located in the forest zone. It is a rural community close to Konongo. It has a wet semi-equatorial climate characterized by two distinct rainy seasons, with a long rainy season from May to July and a short rainy season from September to November. The mean annual temperature and relative humidity are 26 °C and 77%, respectively. The vegetation is a semi-deciduous forest composed of open and closed forests. Duase is drained by one main river which stretches throughout the community, forming large and small water collections at various points which serve as suitable breeding habitats for mosquitoes. The river is diverted at several points to create ponds and wells for farm animals as well as for irrigation purposes. An. gambiae is the major malaria vector in the forest area.
Kpalsogu (9°33′45.2″N, 1°01′54.6″W), Pagaza (9°22′33.34″N, 0°42′29.67″W), and Libga (9°35′32.26″N, 0°50′48.8″W) were the selected sites in the Sahel savannah region of northern Ghana. They have a unimodal rainfall pattern from May to November. The mean annual temperature, which is 28 °C, appears to be favourable for Anopheles larval development, but temperature can reach a maximum of 42 °C. There are dug-out dams and other water impoundments which collect water during the rainy season for irrigation in the dry season. In the rainy season, these dams overflow, creating many swamps which are suitable breeding habitats for Anopheles mosquitoes. Water from the dams which is diverted through canals to farms also serves as breeding sites for mosquitoes. In the harsh dry season, most of these dug-out dams dry up, forming small, temporary open sunlit water collections which are suitable breeding habitats for An. gambiae s.s. [29]. An. gambiae s.l. and An. funestus are the major malaria vectors in the Sahel savanna zone. Secondary vectors found in the Sahel savannah include An. rufipes, An. nili, and An. pharoensis [29], Kpalsogu has been an active IRS site since 2008; in Libga, IRS was stopped after 2014, and Pagaza has never been under IRS intervention. This study was undertaken during the dry (February–March) and rainy (May in the forest zone and coastal savannah zone, and August–September in the Sahel savannah zone) seasons of 2019. Each habitat was sampled once during each season.
Larval habitat characterization
All larval habitats in each site were classified as natural or man-made. Natural habitats included swamps, streams, and natural ponds, while man-made habitats included drainage ditches, footprints, and hoofprints. Land-use type was classified based on the natural vegetation and activities taking place on the land where the larval habitat was found. These included forest for sites with high canopy cover, farmland for cultivated areas, pasture for grazing areas, shrubland for bushy areas with short trees, roads, and swamps, and compounds or home for places with human settlement. The length and width of each habitat were measured and recorded in metres. The percentage of vegetation covering the surface of the water was visually estimated [30]. The vegetation cover was categorized as follows: zero if vegetation was not present in the habitat, ≤ 24% surface coverage, and 25–49, 50–74, and 75–100% surface coverage [31].
Larval sampling and densities
Larval sampling was conducted for all potential breeding sites by the standard dipping method using the WHO 350 ml standard dipper. The size of each habitat was categorized as ≤ 1, > 1–2, > 2–5, > 5–10, > 10–100, or > 100 m, and a maximum of 2, 4, 6, 10, 50, and 150 dips were taken, respectively (i.e., depending on the size of the habitat), as described by Gouagna and Mereta [28, 36]. For habitats pf much smaller size such as hoofprints and footprints, a ladle was used to collect the samples. Larvae collected were classified as early instars (L1 and L2) or late instars (L3 and L4). The number of larvae and pupae were recorded, and the larval density was estimated as the ratio of the number of larvae collected per dip [32,33,34,35].
Mosquito species identification
Anopheline larvae specimens were transported to the insectary of the Department of Medical Microbiology, University of Ghana, where they were bred into adults. The larvae were fed on TetraMin® fish meal and maintained at 27 ± 2 °C. Emerging adult mosquitoes were morphologically identified under a stereomicroscope using the taxonomic keys by Gillies and Coetzee [36]. Anopheles gambiae s.l. were further identified to sibling species and molecular form using rDNA polymerase chain reaction (PCR) [37] and PCR–restriction fragment length polymorphism (RFLP) [38] analysis, respectively.
Data analysis
Descriptive analysis was performed to compare the abundance of the various habitat types and larval densities in the different study sites (ecological zones) and seasons. Larval densities were calculated by dividing the total number of larvae collected by the total number of dips taken. The total number of dips for smaller habitats such as footprints and hoofprints was considered to be one dip. A test for normality of the larval density distribution using the Shapiro–Wilk test showed a non-normal distribution. The density of Anopheles mosquito larvae was compared among the various breeding habitats and study sites. The Mann–Whitney U test and the Kruskal–Wallis test were used to test the associations between continuous and categorical variables. The chi-square and Fisher's exact tests were used to test the association between two categorical variables. Logistic regression was used to assess the association between the habitat characteristics with categorical data and the presence of Anopheles larvae. Nested generalized linear mixed models with sites nested within ecological zones were used to model the effect of habitat characteristics on larval densities. All statistical analyses were conducted in STATA version 15 software (StataCorp. 2017. Stata Statistical Software: Release 15. College Station, TX: StataCorp LLC).
Results
Distribution and abundance of larval habitats in different ecological zones and seasons
A total of 383 breeding habitats comprising 11 different habitat types were encountered and recorded during the study (Fig. 2). Most of the habitats were man-made (69.71%, 267/383), and the others (30.29%, 116/383) were natural. The most abundant habitat type was man-made ponds (27.15%, 104/383), which were present throughout the rainy and dry seasons mostly on farmlands. This was followed by natural ponds (12.01%, 46/383), swamps (11.49%, 44/383), dug-out wells (10.97%, 42/383), and concrete wells (9.92%, 38/383). Other habitat types included tyre tracks (7.83%, 30/383), puddles (6.27%, 24/383), and drainage ditches (6.01%, 23/383). The less abundant habitat types were furrows (4.18%, 16/383), hoofprints (2.87%, 11/383), and footprints (1.31%, 5/383) (Table 1).

Habitat types found during the study period. a Dug-out well, b concrete well, c hoofprints, d man-made pond, e swamp, f furrow, g footprint, h natural pond, i drainage ditch, j tyre track, k puddle
The distribution of mosquito larval habitat types and their abundance varied among the study sites (χ2 = 498.2658; df = 40; p = 0.0001) as well as ecological zones (χ2 = 369.5865; df = 20; p = 0.0001) (Additional file 1: Table S1). In Anyakpor in the coastal savanna zone, only four habitats types were encountered: man-made ponds (57%, 97/168), dug-out wells (20.83%, 35/168), concrete wells (20.83%, 35/168), and a natural pond (0.60%, 1/168). In Duase in the forest area, eight habitat types were found: natural ponds (29.03%, 18/62), puddles (14.52%, 9/62), tyre tracks (12.90%, 8/62), man-made ponds (11.29%, 7/62), drainage ditches (11.29%, 7/62), dug-out wells (11.29%, 7/62), swamps (6.455, 4/62), and concrete wells (3.23%, 2/62). Eight habitat types were also encountered in Kpalsogu in the Sahel savannah area. These included swamps (31.50%, 30/96), tyre tracks (21.88%, 21/96), hoofprints (11.46, 11/96), natural ponds (11.46, 11/96), puddles (10.42%, 10/96), furrows (7.29%, 7/96), footprints (4.17%, 4/96), and drainage ditches (2.08%, 2/96). In Libga in the Sahel savannah area, only four habitat types were found: furrows (28.13%, 9/32), drainage ditches (28.13%, 9/32), natural ponds (21.88%, 7/32), and a concrete well (3.13%, 1/32). In Pagaza, also in the Sahel savannah area, the six habitats encountered were natural ponds (36%, 9/25), puddles (20%, 5/25), drainage ditches (20%, 5/25), swamps (16%, 4/25), tyre tracks (4%, 1/25), and footprints (4%, 1/25).
Larval habitats were significantly more abundant in the rainy season (63.45%, 243/383) than in the dry season (36.55%, 140/383) (χ2 = 91.3295; df = 10; p = 0.001) (Additional file 1: Table S1). Dug-out wells were the most abundant habitat type during the dry season, while man-made ponds were the most abundant during the rainy season. The presence of a larval habitat type was associated with the land-use type (χ2 = 587.4192; df = 60; p = 0.0001). Larval habitats were mostly found on farmlands (58.75%), followed by pastures (16.19%) and on roads (13.05%). The rest were present in forested areas (4.70%), around homes or compounds (4.18%), by streams and rivers (1.83%), and in swamps (1.31%). Most of the habitats had vegetation cover of less than 24% (χ2 = 215.1340; df = 40; p = 0.0001) (Additional file 1: Table S1).
Larval habitat types, and the presence and densities of Anopheles larvae
The presence of Anopheles larvae was dependent on the type of habitat present (χ2 = 41. 3651; df = 10; p < 0.0001). Even though there was a significant increase in the number of habitats during the rainy season (χ2 = 91.3295; df = 10; p < 0.0001) (Additional file 1: Table S1) compared to the dry season, the proportion of habitats where Anopheles larvae were present did not differ significantly between the two seasons (χ2 = 0.0051; df = 1; p = 0.943) (Additional file 2: Table S2). In the dry season, 29.29% (41/140) of the habitats had Anopheles larvae present, while in the rainy season, 29.63% (72/243) of the habitats were positive for Anopheles larvae. In all, dug-out wells were the most frequent habit of Anopheles larvae during the dry season (39.02%, 16/41), followed by man-made ponds (21.95%, 9/41), whereas during the rainy season, man-made ponds were the most commonly inhabited (30.56%, 22/41), followed by swamps (16.67% 12/72) (Table 2).
The type of habitat present was associated with An. gambiae s.l. larval density (χ2 = 29.593; df = 10; p < 0.001) (Additional file 2: Table S2). Even though the mean An. gambiae s.l. larval density in the rainy season (1.49 larvae/dip) was slightly higher than that in the dry season (1.15 larvae/dip), the difference was not significant (z = −0.232; p = 0.8166). In Anyakpor, the most productive habitat types were dug-out wells during both the dry (1.6 larvae/dip) and rainy (11.28 larvae/dip) seasons (Table 3). The preferred breeding habitats in Kpalsogu were natural ponds during the dry season (0.89 larvae/dip) and swamps (2.57 larvae/dip) during the rainy season. In Libga, drainage ditches were the only habitat type present in the dry season, with mean larval density of 0.3 larvae/dip, while furrows were the only productive habitats in the rainy season (1.83 larvae/dip). There were no productive habitats in Pagaza during the dry season, but in the rainy season the most productive habitat type was puddles (1.44 larvae/dip). In Duase, the most productive habitat type was dug-out wells in both the dry season (1.47 larvae/dip) and the rainy season (2.05 larvae/dip) (Table 3).
Abundance and distribution of Anopheles larvae in the different ecological zones
A total of 7894 mosquito larvae were collected during this study. Of this number, 2152 (27.26%) were anophelines, while 5742 (72.74%) were culicines. Of the anopheline species, 2128 (98.88%) were An. gambiae s.l., 16 (0.74%) were An. rufipes, and 8 (0.37%) were An. pharoensis. During the rainy season, 1500 (70.49%) An. gambiae s.l. were collected, while 628 (29.51%) were collected in the dry season.
Anyakpor in the coastal savannah area had the highest abundance of An. gambiae s.l. larvae (1286), with 343 (26.67%) occurring in the dry season and 943 (73.33%) in the rainy season (Table 4). In Duase, situated in the forest area, 30.17% of An. gambiae s.l. larvae were collected during the dry season, while 69.83% were collected in the rainy season. Kpalsogu and Libga in the Sahel savannah recorded 39.00 and 35.79% of An. gambiae s.l. larvae in the dry season, while the rainy season contributed 61.00 and 64.21%, respectively. In Pagaza, also in the Sahel savannah, An. gambiae s.l. larvae were found only in the rainy season (Table 4), with none during the dry season.
The distribution of An. gambiae sibling species varied across the ecological zones (χ2 = 45.9887 df = 8; p = 0.0001). An. coluzzii was the most abundant species (53.44%) in all ecological zones, followed by An. gambiae s.s. (25.98%) and An. arabiensis (6.27%), which were found only in the Sahel savannah: Kpalsogu (19.30%), Libga (17.86%), and Pagaza (35.71%). Anopheles melas were the least abundant species (4.19%) and were present only in Anyakpor in the coastal savannah area (Table 5). All the species were more abundant in the rainy season (χ2 = 21.2510; df = 2; p = 0.0001) than in the dry season. An. gambiae s.s. and An. coluzzii were found in all the habitat types encountered in this study. An. arabiensis were predominantly found in swamps (52.38%) and furrows (28.57%), whereas An. melas were found in dug-out wells (55.56%) and man-made ponds (44.44%). While An. rufipes were found only in Kpalsogu and Libga in the Sahel savannah area, An. pharoensis were found in Anyakpor in the coastal savannah area and Libga in the Sahel savannah area An. pharoensis were found only in man-made ponds (75.00%) and furrows (25.00%). An. rufipes were found in swamps (56.25%), footprints (25.00%), furrows (12.50%), and puddles (6.25%).
Habitat characteristics, occurrence, and densities of Anopheles larvae
The type of habitat influenced the presence of Anopheles larvae (χ2 = 41.3651; df = 10; p = 0.0001) and their density (p < 0.01). A significant majority (84.07% (95/113) of Anopheles-positive habitats were less than 10 m2 in size (χ2 = 11.9217; df = 2; p = 0.0001). Land-use type influenced both the presence of Anopheles larvae (χ2 = 26.5920; df = 6; p = 0.0001) (Additional file 2: Table S2) and their larval density (χ2 = 16.117; df = 6; p = 0.013) (Additional file 2: Table S2). Fifty-five percent of all larval habitats found around homes or compounds contained Anopheles larvae. The odds of finding Anopheles larvae in any habitat was twice as high if the vegetation cover was less than 24% (OR 2.24 [1.02, 4.93]; p = 0.045) (Table 6). As the vegetation cover increased, the density of Anopheles larvae decreased (B = −0.016 [−0.28, 0.003]; p = 0.015) (Additional file 3: Table S3). The present study, again, showed that Anopheles larvae share a habitat preference with culicine larvae. The likelihood of encountering Anopheles larvae in a breeding habitat was more than three times as high when culicines were present (OR 3.13 [1.75,5.59]; p < 0.01) (Table 6).
Discussion
Understanding the ecology of anopheline larvae in a changing environment is crucial for the development and successful implementation of targeted control measures [36, 37] to supplement current adult vector control tools. In this study, the distribution of Anopheles breeding habitats in rural communities in the different ecological zones of Ghana was characterized. The study revealed differences in the abundance and distribution of Anopheles breeding habitats in the different ecological zones. Although man-made ponds were the most abundant habitat type, dug-out wells were the most productive for Anopheles mosquito larvae. Anopheles larvae also preferred to breed in small habitats, while increasing vegetation cover reduced Anopheles larval densities.
The common habitat types were man-made ponds, natural ponds, drainage ditches, and swamps. Other habitats such as tyre tracks and puddles are usually formed during the rainy season when rainwater collects on untarred roads [35]. Such habitats are temporal. The habitats encountered were mostly associated with anthropogenic activities. This explains why communities that practice irrigation farming, Anyakpor and Kpalsogu, had the highest number of habitats.
Duase and Kpalsogu had the most diverse breeding habitat types, which included 8 of the 11 habitat types encountered. The persistence of breeding habitats during both dry and rainy seasons in the forest and coastal savannah areas accounts for perennial malaria found within these sites, while seasonal variations are observed in the Sahel savannah areas [39, 40].
In all, the most abundant habitat type was man-made ponds. Natural ponds were the only habitat type found in all the study sites. Hoofprints were found only in the Sahel savannah zone where livestock are left to graze on swampy pastures. Most of the habitats were man-made and found on farmlands. This emphasizes the importance of human activities and, for that matter, land-use in the creation of Anopheles breeding habitats and the impact they have on malaria transmission.
The fact that most of the breeding habitats are found on farmland can be attributed to the practice of irrigation. Irrigation provides ideal breeding habitats for Anopheles vectors, and this study corroborates that of Appawu et al. [41], who found that irrigated fields generated large numbers of mosquitoes. It is important to note that agrochemicals used on these farms end up polluting the water sources which serve as breeding sites on these farms, thereby leading to the development and spread of insecticide resistance by exposing mosquito larvae to high or sublethal doses of the agrochemicals [42,43,44,45].
The variation in the presence of Anopheles larvae may be due to the differences in the physical, chemical, and biological properties as well as the quality of the water present in the various habitats [46]. These properties directly influence the choice of oviposition sites by gravid females and also influence the development and survivorship of larvae [47,48,49,50]. The presence of Anopheles larvae in man-made ponds and dug-out wells further establishes the influence of human activities on the presence and distribution of Anopheles vectors [31, 37, 50, 53,54,55].
The presence of Anopheles larvae in habitats close to human settlements, where it was easy to find the next blood meal source, corroborates the findings of studies from Kenya [51, 52], which also suggest that because An. gambiae s.l. are closely associated with humans, they will make use of the closest habitat for oviposition when they become gravid [51, 52]. Choosing habitats close to human settlements where An. gambiae may have taken a blood meal is also an evolutionary strategy to conserve energy [46, 53].
This study also revealed that Anopheles larvae were predominantly present in breeding habitats with vegetation cover of less than 24%. Similar findings have been observed in Kenya [54,55,56]. Low vegetation cover allows the habitat to be more exposed to sunlight, a preference for ovipositing mosquitoes [5]. Also, adequate exposure to sunlight warms the water to suitable temperatures, as temperature is also a key factor influencing larval development and survival [2, 5, 7, 57]. Inadequate exposure to sunlight caused by high vegetation cover affects the photosynthetic efficiency of algae biomass which serves as food for mosquito larvae [58]. It was evident in this study that as the percentage of vegetation cover increased, the density of Anopheles larvae decreased. This is in line with studies conducted in Ethiopia [35, 59]. As vegetation cover increases, the amount of sunlight reaching the habitat is decreased.
In Anyakpor in the coastal zone, the predominant species was An. coluzzii (60.82%), similar to what was reported by Kudom [60] and Fossog et al. [61], whose model showed An. coluzzii to dominate the coastal line of Africa. The species was mostly found in man-made ponds, dug-out wells, and concrete wells. Compared to An. gambiae s.s., both inland and coastal An. coluzzii are known to have a higher tolerance to salinity [61, 62]. An. melas in Anyakpor were found only in dug-out wells and man-made ponds, because these habitats are fed by salty underground water which An. melas prefer to breed in, unlike concrete wells which are fed by rainwater. An. coluzzii and An. gambiae s.s. were the only species found in Duase, in the forest zone, with An. coluzzii being the dominant species, which is contrary to the findings of other studies [39, 63, 64]. This could be as a result of the deforestation which is caused by rapid urbanization. Rapid deforestation affects climatic conditions, and this might be causing Duase to become drier, thereby allowing An. coluzzii to thrive there. An. coluzzii was also the dominant species in Kpalsogu, followed by An. gambiae s.s. and An. arabiensis. On the other hand, An. gambiae s.s. was the dominant species in Libga. In Pagaza, An. coluzzii and An. arabiensis were co-dominant. An. arabiensis were found only in the Sahel savannah area because of its dry sub-arid environments [65]. An. arabiensis prefer to be zoophilic even though they can also be anthropophilic [66], and this can explain why they were most commonly found in swamps, since swamps in the Sahel savannah areas mostly serve grazing cattle. As shown by this study, An. coluzzii and An. gambiae s.s. are known to live in sympatry in most parts of Ghana [60]. However, An. coluzzii predominates in the coastal and Sahel savannah regions of Ghana [63, 64, 67]. The dominance of An. coluzzii in the coastal regions of Ghana has been attributed to permanent habitats created on irrigation farms [63, 64]. An. coluzzii is usually seen breeding in large permanent habitats, but in this study they were also found in small temporary habitats such as hoofprints. A study by Edillo et al. [68] showed that An. coluzzii preferred different breeding habitat types from An. gambiae s.s. and An. arabiensis, but this study showed otherwise. An. pharoensis and An. rufipes are secondary malaria vectors in Ghana [15], and even though they are less abundant, they can gradually replace the current malaria vectors and become dominant in the very near future.
This study might have missed some of the larval habitats and some species of mosquitoes, since it was difficult to access all habitats in all study sites. Some habitats might have been in dense thicket and bushes that were not reached for sampling. This study provides baseline insight into the development of integrated larval source management suitable for the larval habitats in the various study sites. The results revealed that irrigated farms contribute to higher populations of malaria mosquitoes, as this system of agriculture creates many habitats suitable for Anopheles mosquito larvae. As a result, communities that practice irrigation farming have a higher abundance of Anopheles larvae. This study, again, revealed that human activities contribute greatly to the presence and abundance of Anopheles mosquito larvae. This implies that changes in agricultural methods, including methods of irrigation and environmental management such as surface water drainage, landfilling, and land reclamation, would be extremely beneficial in controlling malaria transmission. Considering that An. gambiae s.l. prefers to breed in small habitats, habitat modification would be a suitable method of larval source reduction. Other integrated larval source management approaches such as water and environmental management and biological control methods are also feasible. In communities that rely on the same water collections for domestic purposes, and in situations where the water collections cannot be drained, microbial bio-larvicides may be used.
Conclusion
In this study, the presence and availability of Anopheles breeding habitats varied among the study site, and, ecological zones. The abundance of breeding habitats was influenced by rainfall, as more habitats were created during the rainy season and this, in turn, increased the abundance of Anopheles mosquitoes in all the ecological zones. Man-made ponds were the most abundant breeding habitat in the coastal savannah zone, while natural ponds were the most abundant in both the forest and Sahel savannah zones. Dug-out wells were the most productive habitats in the coastal savannah and forest zones, while furrows were the most productive in the Sahel savannah zone. Anopheles coluzzii was the predominant species in all the study sites. Anopheles melas was found only in the coastal savannah, whereas An. arabiensis was found only the Sahel savanna zone. The abundance of Anopheles breeding habitats and larvae was influenced by anthropogenic activities. Encouraging people whose activities create the larval habits to become involved in larval source management such as habitat manipulation to stop mosquito breeding will be important for malaria and lymphatic filariasis control.
Availability of data and materials
All datasets generated and/or analysed during the current study are available on request.
References
World Health Organization. A global brief on vector-borne diseases [Internet]. World Heal. Organ. 2014. pp 54. http://apps.who.int/iris/bitstream/10665/111008/1/WHO_DCO_WHD_2014.1_eng.pdf
Rejmánková E, Grieco J, Achee N, Roberts DR. Ecology of Larval Habitats. In: Manguin S, editor. Anopheles mosquitoes—new insights into Malar vectors. InTech Open; 2013. p. 397–446.
Ondiba IM, Oyieke FA, Athinya DK, Nyamongo IK, Estambale BB. Larval species diversity, seasonal occurrence and larval habitat preference of mosquitoes transmitting Rift Valley fever and malaria in Baringo County, Kenya. Parasit Vectors. 2019;12:1–14. https://doi.org/10.1186/s13071-019-3557-x.
MacHault V, Gadiaga L, Vignolles C, Jarjaval F, Bouzid S, Sokhna C, et al. Highly focused anopheline breeding sites and malaria transmission in Dakar. Malar J. 2009;8:1–21.
Williams J, Pinto J. Training manual on malaria entomology for entomology and vector control technicians (basic level). USAID. Washington, DC. 2012;78.
Emidi B, Kisinza WN, Mmbando BP, Malima R, Mosha FW. Effect of physicochemical parameters on Anopheles and Culex mosquito larvae abundance in different breeding sites in a rural setting of Muheza, Tanzania. Parasit Vectors. 2017;10:1–12.
Afrane YA, Lawson BW, Githeko AK, Yan G. Effects of microclimatic changes caused by land use and land cover on duration of gonotrophic cycles of Anopheles gambiae (Diptera: Culicidae) in western Kenya highlands. J Med Entomol. 2005;45:974–80.
Dery DB, Asante KP, Zandoh C, Febir LG, Brown C, Adjei G, et al. Baseline malaria vector transmission dynamics in communities in Ahafo mining area in Ghana. Malar J. 2015;14:1–8.
WHO. A Framework for Malaria Elimination [Internet]. WHO Press. World Heal. Organ. 2017. http://apps.who.int/iris/bitstream/handle/10665/254761/9789241511988-eng.pdf?sequence=1
Govella NJ, Ferguson H. Why use of interventions targeting outdoor biting mosquitoes will be necessary to achieve malaria elimination. Front Physiol. 2012;3:1–6.
Derua YA, Kweka EJ, Kisinza WN, Githeko AK, Mosha FW. Bacterial larvicides used for malaria vector control in sub—Saharan Africa : review of their effectiveness and operational feasibility. Parasit Vectors. 2019;12:1–18. https://doi.org/10.1186/s13071-019-3683-5.
Fillinger U, Knols BGJ, Becker N. Efficacy and efficiency of new Bacillus thuringiensis var. israelensis and Bacillus sphaericus formulations against Afrotropical anophelines in Western Kenya. Trop Med Int Heal. 2003;8:37–47.
Bhatt S, Weiss DJ, Cameron E, Bisanzio D, Mappin B, Dalrymple U, et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature. 2015;526:207–11. https://doi.org/10.1038/nature15535.
Tusting LS, Thwing J, Sinclair D, Fillinger U, Gimnig J, Bonner KE, et al. Mosquito larval source management for controlling malaria. Cochrane Database Syst Rev. 2013. https://doi.org/10.1002/14651858.CD008923.pub2.
Afrane YA, Bonizzoni M, Yan G. Secondary malaria vectors of sub-saharan africa: threat to malaria elimination on the continent? In: Rodriguez-Morales AJ, editor. Current topics in malaria. Intech Open Science; 2016. p. 473–90.
Sougoufara S, Doucouré S, Sembéne PMB, Harry M, Sokhna C. Challenges for malaria vector control in sub-Saharan Africa : Resistance and behavioral adaptations in Anopheles populations. J Vector Borne Dis. 2017;54:4–15.
Ndenga BA, Simbauni JA, Mbugi JP, Githeko AK, Fillinger U. Productivity of malaria vectors from different habitat types in the Western Kenya highlands. PLoS ONE. 2011;6:1–11.
Becker N. Microbial control of mosquitoes : management of the upper Rhine mosquito population as a model programme. Parasitol Today. 1997;13:485–7.
Rejmankova E, Grieco J, Archee N, Masuoka P, Pope K, Roberts D, et al. Freshwater community interactions and malaria. In: Collinge SK, Ray C, editors., et al., Disease ecology. Oxford: Oxford University Press; 2006. p. 241.
Gimnig JE, Ombok M, Kamau L, Hawley WA. Characteristics of larval anopheline (Diptera: Culicidae) habitats in Western Kenya. J Med Entomol. 2001;38:282–8.
Kibret S, Lautze J, Mccartney M, Nhamo L, Yan G. Malaria around large dams in Africa: Effect of environmental and transmission endemicity factors. Malar J. 2019;18:1–12. https://doi.org/10.1186/s12936-019-2933-5.
Minakawa N, Munga S, Atieli F, Mushinzimana E, Zhou G, Githeko AK, et al. Spatial distribution of anopheline larval habitats in Western Kenyan highlands: Effects of land cover types and topography. Am J Trop Med Hyg. 2005;73:157–65.
Mushinzimana E, Munga S, Minakawa N, Li L, Feng CC, Bian L, et al. Landscape determinants and remote sensing of anopheline mosquito larval habitats in the western Kenya highlands. Malar J. 2006;5:1–11.
Afrane YA, Zhou G, Lawson BW, Githeko AK, Yan G. Life-table analysis of Anopheles arabiensis in western Kenya highlands: Effects of land covers on larval and adult survivorship. Am J Trop Med Hyg. 2007;77:660–6.
Mutuku FM, Bayoh MN, Hightower AW, Vulule JM, Gimnig JE, Mueke JM, et al. A supervised land cover classification of a western Kenya lowland endemic for human malaria: Associations of land cover with larval Anopheles habitats. Int J Health Geogr. 2009;8:1–13.
Dadzie S, Boakye D, Asoala V, Koram K, Kiszewski A, Appawu M. A community-wide study of malaria reduction: Evaluating efficacy and user-acceptance of a low-cost repellent in Northern Ghana. Am J Trop Med Hyg. 2013;88:309–14.
Kipyab PC, Khaemba BM, Mwangangi JM, Mbogo CM. The physicochemical and environmental factors affecting the distribution of Anopheles merus along the Kenyan coast. Parasites Vectors Vectors. 2015;8:1–8.
Appawu MA, Baffoe-Wilmort A, Afari EA, Dunyo S, Koram KA, Nkrumah FK. Malaria vector studies in two ecological zones in southern Ghana. African Entomol. 2001;9:59–65.
Coleman S, Dadzie SK, Seyoum A, Yihdego Y, Mumba P, Dengela D, et al. A reduction in malaria transmission intensity in Northern Ghana after 7 years of indoor residual spraying. Malar J. 2017;16:324.
Imbahale SS, Paaijmans KP, Mukabana WR, Lammeren R Van, Githeko AK, Takken W, et al. A longitudinal study on Anopheles mosquito larval abundance in distinct geographical and environmental settings in western Kenya. Malar J 2011;10:1–13. http://www.malariajournal.com/content/10/1/81
Gouagna LC, Rakotondranary M, Boyer S, Lempérière G, Dehecq J-SS, Fontenille D. Abiotic and biotic factors associated with the presence of Anopheles arabiensis immatures and their abundance in naturally occurring and man-made aquatic habitats. Parasites Vectors. 2012;5:1–12.
Silver JB. Mosquito ecology-field sampling methods. 3rd ed. Springer; 2008.
Imbahale SS, Paaijmans KP, Mukabana WR, Van LR, Githeko AK, Takken W. A longitudinal study on Anopheles mosquito larval abundance in distinct geographical and environmental settings in western Kenya. Malar J. 2011;10(1):1–13.
Kweka EJ, Munga S, Himeidan Y, Githeko AK, Yan G. Assessment of mosquito larval productivity among different land use types for targeted malaria vector control in the western Kenya highlands. Parasites Vectors. 2015;8:1–8. https://doi.org/10.1186/s13071-015-0968-1.
Getachew D, Balkew M, Tekie H. Anopheles larval species composition and characterization of breeding habitats in two localities in the Ghibe River Basin, southwestern Ethiopia. Malar J. 2020;19:1–13. https://doi.org/10.1186/s12936-020-3145-8.
Gillies MT, Coetzee M. A supplement to the Anophelinae of Africa South of the Sahara. Publ South African Inst Med Res 1987;55:1–146. http://ww.w.sandflycatalog.org/files/pdfs/190531.pdf
Scott JA, Brogdon WG, Collins FH. Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction. Am J Trop Med Hyg. 1993;49:520–9.
Fanello C, Santolamazza F, Della Torre A. Simultaneous identification of species and molecular forms of the Anopheles gambiae complex by PCR-RFLP. Med Vet Entomol. 2002;16:461–4.
Dery DB, Brown C, Asante KP, Adams M, Dosoo D, Amenga-Etego S, et al. Patterns and seasonality of malaria transmission in the forest-savannah transitional zones of Ghana. Malar J. 2010;9:1–8.
PMI-Ghana. President’s Malaria Initiative - Ghana [Internet]. Malar. Oper. Plan FY 2018. 2018. http://www.pmi.gov/docs/default-source/default-document-library/malaria-operational-plans/fy14/ghana_mop_fy14.pdf?sfvrsn=20
Appawu M, Owusu-Agyei S, Dadzie S, Asoala V, Anto F, Koram K, et al. Malaria transmission dynamics at a site in northern Ghana proposed for testing malaria vaccines. Trop Med Int Heal. 2004;9:164–70.
Baffour-Awuah S, Annan AA, Maiga-Ascofare O, Dieudonné SD, Adjei-Kusi P, Owusu-Dabo E, et al. Insecticide resistance in malaria vectors in Kumasi, Ghana. Parasit Vectors. 2016;9:633. https://doi.org/10.1186/s13071-016-1923-5.
Kudom AA, Anane LN, Afoakwah R, Adokoh CK. Relating high insecticide residues in larval breeding habitats in urban residential areas to the selection of pyrethroid resistance in Anopheles gambiae s.l. (Diptera: Culicidae) in Akim Oda, Ghana. J Med Entomol 2018;55:490–5. https://academic.oup.com/jme/article/55/2/490/4756456
Reid MC, McKenzie FE. The contribution of agricultural insecticide use to increasing insecticide resistance in African malaria vectors. Malar J BioMed Central. 2016;15:1–8.
Soko W, Chimbari MJ, Mukaratirwa S. Insecticide resistance in malaria-transmitting mosquitoes in Zimbabwe: a review. Infect Dis poverty. 2015;4:46. https://doi.org/10.1186/s40249-015-0076-7.
Gu W, Utzinger J, Novak RJ. Habitat-based larval interventions: A new perspective for malaria control. Am J Trop Med Hyg. 2008;78:2–6.
Himeidan YE, Temu EA, El Rayah EA, Munga S, Kweka EJ. Chemical cues for malaria vectors oviposition site selection: challenges and opportunities. J Insects. 2013;2013:1–9.
Mereta ST, Yewhalaw D, Boets P, Ahmed A, Duchateau L, Speybroeck N, et al. Physico-chemical and biological characterization of anopheline mosquito larval habitats ( Diptera : Culicidae ): implications for malaria control. Parasit Vectors. 2013;6:1–16.
Nikookar SH, Fazeli-Dinan M, Azari-Hamidian S, Mousavinasab SN, Aarabi M, Ziapour SP, et al. Correlation between mosquito larval density and their habitat physicochemical characteristics in Mazandaran Province, northern Iran. PLoS Negl Trop Dis. 2017;11:1–19.
Djamouko-djonkam L, Mounchili-Ndam S, Kala-chouakeu N, Nana-ndjangwo SM, Kopya E, Sonhafouo-chiana N, et al. Spatial distribution of Anopheles gambiae sensu lato larvae in the urban environment of Yaoundé, Cameroon. Infect Dis Poverty. 2019;8:1–15.
Mutuku FM, Bayoh MN, Gimnig JE, Vulule JM, Kamau L, Walker ED, et al. Pupal habitat productivity of Anopheles gambiae complex mosquitoes in a rural village in western Kenya. Am Soc Trop Med Hyg. 2006;74:54–61.
Minakawa N, Seda P, Yan G. Influence of host and larval habitat distribution on the abundance of African malaria vectors in Western Kenya. Am J Trop Med Hyg. 2002;67:32–8.
Kweka EJ, Zhou G, Munga S, Lee MC, Atieli HE, Nyindo M, et al. Anopheline larval habitats seasonality and species distribution: a prerequisite for effective targeted larval habitats control programmes. PLoS ONE. 2012;7(12):e52084.
Muturi EJ, Mwangangi J, Shililu J, Muriu S, Jacob B, Kabiru E, et al. Mosquito species succession and physicochemical factors affecting their abundance in rice fields in Mwea. Kenya J Med Entomol. 2007;44:336–44.
Yasuoka J, Levins R. Impact of deforestation and agricultural development on anopheline ecology and malaria epidemiology. Am J Trop Med Hyg. 2007;76:450–60.
Mwangangi JM, Shililu J, Muturi EJ, Muriu S, Jacob B, Kabiru EW, et al. Anopheles larval abundance and diversity in three rice agro-village complexes Mwea irrigation scheme, central Kenya. Malar J. 2010;9:1–10.
WHO. Malaria entomology and vector control. Geneva: WHO Press; 2013. p. 190.
Mwingira VS, Mayala BK, Senkoro KP, Rumisha SF, Shayo EH, Mlozi MR, et al. Mosquito larval productivity in rice-fields infested with Azolla in Mvomero District, Tanzania. Tanzan J Health Res. 2009;11:17–22.
Kenea O, Balkew M, Gebre-Michael T. Environmental factors associated with larval habitats of anopheline mosquitoes (diptera: Culicidae) in irrigation and major drainage areas in the middle course of the rift valley, central ethiopia. J Vector Borne Dis. 2011;48:85–92.
Kudom AA. Larval ecology of Anopheles coluzzii in Cape Coast, Ghana: Water quality, nature of habitat and implication for larval control. Malar J BioMed Central. 2015;14:1–13.
Fossog BT, Ayala D, Acevedo P, Kengne P, Ngomo Abeso Mebuy I, Makanga B, et al. Habitat segregation and ecological character displacement in cryptic African malaria mosquitoes. Evol Appl. 2015;8:326–45.
Nwaefuna EK, Bagshaw II, Gbogbo F, Osae M. Oviposition and development of Anopheles coluzzii Coetzee and Wilkerson in salt water. MMalaria Res Treat. 2019;2019:1–7.
Yawson AE, McCall PJ, Wilson MD, Donnelly MJ. Species abundance and insecticide resistance of Anopheles gambiae in selected areas of Ghana and Burkina Faso. Med Vet Entomol. 2004;18:372–7.
de Souza D, Kelly-Hope L, Lawson B, Wilson M, Boakye D. Environmental factors associated with the distribution of Anopheles gambiae s.s in Ghana; an important vector of lymphatic filariasis and malaria. PLoS ONE. 2010;5(3):e9927.
Tay SCK, Danuor SK, Morse A, Caminade C, Badu K, Abruquah H. Entomological survey of malaria vectors within the kumasi metropolitan area—a study of three communities: Emena Atonsu and Akropong. Int J Parasitol. 2012;1:144–54.
Dida GO, Anyona DN, Abuom PO, Akoko D, Adoka SO, Matano AS, et al. Spatial distribution and habitat characterization of mosquito species during the dry season along the Mara River and its tributaries, in Kenya and Tanzania. Infect Dis Poverty. 2018;7:1–16.
Chabi J, Baidoo PK, Datsomor AK, Okyere D, Ablorde A, Iddrisu A, et al. Insecticide susceptibility of natural populations of Anopheles coluzzii and Anopheles gambiae (sensu stricto) from Okyereko irrigation site, Ghana, West Africa. Parasites Vectors. 2016;9:1–8. https://doi.org/10.1186/s13071-016-1462-0.
Edillo FE, Tripét F, Touré YT, Lanzaro GC, Dolo G, Taylor CE. Water quality and immatures of the M and S forms of Anopheles. Malar J. 2006;5:1–10.
Acknowledgements
The authors are grateful to the people of Anyakpor, Duase, Kpalsogu, Libga, and Pagaza for permitting us to sample mosquitoes in their communities and farms. We greatly appreciate Shittu Bunkunmi Dhikrullahi, Abdul Rahim Mohammed, and Isaac Sun Sraku for their technical assistance during field surveys and the molecular work in the lab. The authors also thank all the community field assistants for helping in the data collection in the various communities.
Funding
This study was supported by a grant from the National Institutes of Health (NIH: R01 A1123074). The funders had no role or influence on the design of this study, the collection, analysis, or interpretation of the data, or the writing of this manuscript.
Author information
Authors and Affiliations
Contributions
IAH, SKA, AOF, and YAA conceived and designed the study. IAH and YAA were responsible for designing and coordinating the entomological surveys. IAH was responsible for data collection. YAA, SKA, and AOF supervised the data collection. IAH and BAM were responsible for the data analysis and drafted the first manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethical approval for this study with protocol identification number CHS-Et/M2-5.5/2019–2020 was obtained from the Ethics and Protocol Review Committee of the College of Health Sciences, University of Ghana.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Univariate analysis on the distribution of Anopheles breeding habitats.
Additional file 2: Table S2.
Univariate analysis of habitat characteristics, and the presence and larval density of Anopheles larvae.
Additional file 3: Table S3.
GLMM of habitat characteristics and Anopheles larval density.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hinne, I.A., Attah, S.K., Mensah, B.A. et al. Larval habitat diversity and Anopheles mosquito species distribution in different ecological zones in Ghana. Parasites Vectors 14, 193 (2021). https://doi.org/10.1186/s13071-021-04701-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-021-04701-w