
Abstract
Background
Baseline information that is essential for determining the areas to target with larval control includes estimates of vector diversity and larval habitat preferences. Due to a lack of such information in Baringo County, Kenya, this study assessed species diversity and larval habitat preference of potential mosquito vectors of Rift Valley fever (RVF) and malaria.
Methods
Mosquito larvae were sampled from nine types of larval habitats and were identified morphologically. Species diversity was estimated by the Shannon’s diversity index while larval habitat preference by RVF and malaria vectors was determined by ANOVA.
Results
A total of 7724 immature mosquitoes comprising 17 species belonging to four genera, namely Anopheles, Culex, Aedes and Mansonia, were identified. Among the 17 species, three Anopheles species are responsible for malaria transmission: An. gambiae (s.l.), An. funestus (s.l.) and An. pharoensis. Rift Valley fever vectors included Mansonia spp. and Culex spp. The highest Shannon's diversity index was observed during the cold dry season (H = 2.487) and in the highland zone (H = 2.539) while the lowest diversity was recorded during the long rain season (H = 2.354) and in the riverine zone (H = 2.085). Ditches had the highest mean number of Anopheles larvae (16.6 larvae per sample) followed by swamp (12.4) and seasonal riverbed (10.7). Water pit and water pan had low mean numbers of Anopheles larvae (1.4 and 1.8, respectively) but relatively high mean numbers of culicines (16.9 and 13.7, respectively). Concrete tank was the least sampled type of habitat but had highest mean number of culicine larvae (333.7 l) followed distantly by water spring (38.9) and swamp (23.5). Overall, larval habitats were significantly different in terms of larval density (F(8,334) = 2.090, P = 0.036).
Conclusions
To our knowledge, the present study reports culicine larval species diversity in Baringo for the first time and the most preferred habitats were concrete tanks, water springs and swamps. Habitats preferred by Anopheles were mainly riverbed pools, ditches and swamps. Environmental management targeting the habitats most preferred by potential vectors can be part of integrated vector control in Baringo, especially during dry seasons.
Similar content being viewed by others


Background
More than 80% of the world’s population is at risk of one or more vector-borne diseases [1]. Mosquitoes are responsible for most vector-borne disease transmission and Africa bears a large burden [2]. Out of the 3000 known species of mosquitoes, about 100 are vectors of human diseases [3]. The common diseases transmitted by mosquito vectors include Rift Valley fever (RVF) and malaria. Rift Valley fever is a zoonotic disease transmitted by a Phlebovirus of the family Bunyaviradae [4]. Eleven epizootics have occurred in Kenya between 1951 and 2007 with an average inter-epizootic period of 3.6 years [5]. During the last RVF outbreak in Kenya in 2006/2007, the highest proportions of cases (31%) were from Garissa in the northeastern region followed by Baringo (24%) in the Rift Valley region. This was the first time RVF transmission was reported in Baringo [6].
Malaria is another vector-borne disease transmitted by mosquitoes. Global estimates of malaria cases and mortality were 212 million and 429,000, respectively, in 2015 with 90% of the cases occurring in sub-Saharan Africa [7]. This calls for the up-scaling of control strategies and inclusion of more innovative ways to supplement existing interventions. Malaria is a prevalent disease in Baringo, accounting for 11.8% of outpatients in health facilities [8]. The burden of malaria in Baringo is higher in the low-lying areas where transmission occurs throughout the year [9] but cases of malaria also occur in the highlands [10]. Malaria fatalities increase during outbreaks in areas where populations are immunologically vulnerable [11]. Such cases of explosive malaria were witnessed in October 2017 in Baringo East, a midland area that is rarely affected by malaria.
The current vector control strategies in Kenya, such as LLINs and IRS which target indoor resting mosquitoes, are insufficient. These strategies may not protect against outdoor resting mosquito vectors such as the culicine species that transmit RVF and secondary malaria vectors like An. pharoensis and An. coustani. Larval source management (LSM) should, therefore, be an additional strategy to supplement the existing interventions as part of an integrated vector management (IVM) policy [12]. This is possible because larval habitats in Baringo are mainly permanent artificial water bodies which are few in number, accessible and easily identifiable [13]. Information on the diversity and distribution of endemic vector species is essential and requires knowledge on the identity of mosquito species present in each locality for effective implementation of vector management [14,15,16]. The present study determined species diversity, seasonal occurrence and larval habitat preference by RVF and malaria vectors in Baringo.
Factors that affect mosquito species diversity include season, elevation and type of aquatic habitat [17]. Mosquito diversity parameters like species richness and abundance can be compared between different ecological zones and seasons. Areas with more diverse larval habitats are likely to have higher mosquito species diversity than areas with few larval sites [18]. Seasonal changes can affect larval habitat availability and productivity and thereby impact on species diversity. It is important to sample the same area continuously to cover different seasonal climatic conditions [17]. In sub-Saharan Africa, a significant decrease in the spatial distribution of larval habitats during the dry season [19] could affect species diversity. A survey of the Mara river basin found the highest number of mosquito larvae during the dry season at the dry stream beds compared to other habitat types [20].
Several studies have shown that different mosquito species prefer different larval habitats. Anopheles gambiae (s.l.), the principal vector of malaria, prefers slightly turbid, shallow, sunlit and transient water pools without aquatic plants [21, 22]. Anopheles pharoensis, a secondary vector of malaria, breeds in habitats with floating vegetation and with relatively shady conditions [22]. A recent study found no Anopheles larvae in an abandoned fishpond [23], a confirmation that Anopheles species do not prefer deep water bodies [24], probably due to lack of siphon used for breathing under water. However, it was not possible to identify clear characteristics of larval habitats for Anopheles species larvae in Tanzania [25]. Culicine mosquitoes have been shown to exploit a wide range of aquatic habitats with slight differences among individual species [18, 26]. All potential larval habitats can have one or more larval species of the genus Culex [18, 27]. Mansonia mosquitoes prefer habitats with aquatic plants such as Pistia spp. and polluted water [28,29,30]. The Mansonioides prefer habitats with well-developed macrophytes which provide mechanical support and favorable conditions for oviposition. Aedes spp. exploit a wide range of larval habitats with different temporal characteristics [31], probably due to adaptation to broad environmental components of physico-chemical factors. Specifically, Aedes aegypti mosquitoes prefer larval habitats of artificial water containers [32, 33].
Kenya is divided into four epidemiological regions based on malaria. A lot of research on larval vector surveys has been conducted in the endemic regions of the lake Victoria basin (western Kenya), central Kenya and coastal regions [16, 34,35,36,37] but few larval studies have been conducted in the semi-arid, seasonal transmission areas such as Baringo [13, 38]. Although some research has been performed in Baringo on mosquito vectors, there is limited information on species diversity and larval habitat preference. Furthermore, such previous studies did not cover the entire county because they were limited to areas around Lake Baringo and Lake 94. The aim of the present study was to investigate larval species diversity, seasonal occurrence and habitat preference of RVF and malaria vectors in a wider area of Baringo categorized into four ecological zones. Knowledge on seasonal mosquito species diversity and larval habitat preferences of the vectors will allow public health officials to more accurately carry out targeted larval source management in Baringo.
Methods
Description of the study area
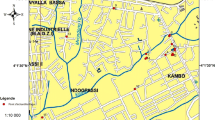
The study was conducted in Baringo County, Kenya. The area surveyed was between 0°32′28″–0°43′23″N, 35°36′7″–36°16′37″E at an altitude ranging between 870 and 2499 m above sea level (Fig. 1). The study area was divided into four ecological zones: lowland, midland, highland and riverine. Baringo is characterized by four lakes, Lake Baringo, Lake Bogoria, Lake 94 and Lake Kamnarok. The seasonal rivers in Baringo are often characterized by pockets of small pools of water along the riverbed, which provide suitable larval habitats for mosquitoes. Dams also exist, which form focal points where humans and livestock aggregate to access water, especially during the dry season.

Map of study area in Baringo County
Baringo County has two distinct weather patterns, namely dry and wet seasons. The dry season consist of months with distinctly low temperatures (June to August) and those with high temperatures (December to February). Mean monthly temperatures in the highlands range from 25 °C during the cold months to 30 °C during the hot months, while in the lowlands it ranges between 30 and 35 °C during cold and hot months, respectively. Baringo County experiences two rainy seasons: long rains (March to May) and short rains (September to November). The County experiences extreme spatial fluctuations in seasonal rainfall. It receives between 1000–1500 mm of rainfall annually in the highlands and 500–600 mm in the lowlands [39]. Monthly temperature and rainfall for Baringo during the study period (2014–2016) were obtained from IRI and CHIRPS [40].
Larval habitats, sampling and identification of mosquitoes
Twenty-four sites (six from each ecological zone) with potential mosquito larval habitats were identified and mapped with geo-positioning equipment (GPS) during a preliminary survey. Larval habitats were selected to represent the diverse larval habitats (Fig. 2) in the heterogeneous topography of Baringo ecological zones. The same larval habitats were sampled longitudinally once every month between June 2014 and June 2016. Larvae collected monthly were consolidated into seasons then analyzed to assess seasonal fluctuations in species diversity and density per dip. Ten to twenty dips were taken depending on size of habitat using the 350 ml standard dipper. Different points along the edge of large habitats such as lake shore were sampled hence more dips than those taken from smaller habitats such as water springs. However, the main objective was to find out the species found in different habitat types without considering the habitat size. The larvae were transferred into a sample container using a wide-bore pipette. Larvae were morphologically identified under a dissecting microscope to the lowest possible taxonomic unit [41, 42].

Larval habitats that were sampled regularly in Baringo County. a Lake Kamnarok covered with small floating plants. b Water pit with hoof prints. c Water spring. d Water pan without vegetation. e Water pan with algae and grass. f Riverbed pool. g Lake Baringo at Salabani. h Ditch
Statistical analyses
Analysis of species diversity
Average monthly temperature and rainfall during the sampling period were used to represent the seasonal climatic conditions. Information on species diversity in Baringo County for each ecological zone and season were estimated using Shannonʼs diversity index. Shannonʼs index was selected because it combines species richness and abundance and is also sensitive to rare and abundant species [43]. Pairwise comparisons of larval species diversity between ecological zones and seasons were made using the Shannonʼs diversity t-test as proposed by Hutcheson [44] based on the following equation:
where \(S_{{H^{\prime}_{1} - H^{\prime}_{2} }} = \sqrt {S_{{H^{\prime}_{1} }}^{2} + S_{{H^{\prime}_{2} }}^{2} }\)with each variance of \(H^{\prime}\) estimated by
where \(H^{\prime}\) is the Shannonʼs diversity index for each of the two samples, S is species richness (total number of species), S2 is the variance of each sample (\(H^{\prime}_{1}\) and \(H^{\prime}_{2}\)), n is the total abundance (number of individuals) and fi is the proportion each species makes towards total.
Simpsonʼs index was included to measure species evenness. Berger-Parkerʼs index was also used to indicate the proportion of the most abundant species in each ecological zone and climatic season. Species rarefaction curves were used to estimate sampling sufficiency and expected occurrence of species for smaller groups. Cumulative species abundance (ln S), Shannonʼs index (H) and log evenness (ln E) (SHE) profiles were used to estimate ecological heterogeneity.
Analysis of larval habitat preference
Larval density per dip was determined by dividing total number of larvae by number of dips to get the mean. The mean was then standardized by multiplying by highest number of dips since different numbers of dips were taken (10–20) based on size of larval habitat. A test of the data for normality using the Shapiro-Wilk test showed a non-normal distribution. Thus, data were log-transformed [log10 (n+1) because the data had many zeroes] to reduce skewness and improve normality. After the transformation, data were re-tested and found to have a normal distribution. One-way analysis of variance (ANOVA) was then used to compare mean larvae in each habitat so as to determine habitat preference by malaria and RVF vectors. When significant differences were observed in ANOVA, Tukeyʼs post-hoc test was used for pairwise comparisons of the means [18].
Results
Larval species diversity in ecological zones and seasons in Baringo
A total of 7724 immature mosquitoes comprising of 17 species belonging to 4 genera (Anopheles, Culex, Aedes and Mansonia) were identified from various larval habitats in the four ecological zones. The 17 species included five Anopheles species, three Aedes species, eight Culex species and one Mansonia species. The Mansonia species were collected at pupa stage and left to emerge before identification was done. Among the 17 species identified, three Anopheles species were malaria vectors: An. gambiae (s.l.) (8.1%), An. funestus (s.l.) (0.1%) and An. pharoensis (15.4%).
The Shannonʼs diversity index (Table 1) was highest in the highland zone (H = 2.539) followed by the lowland zone (H = 2.536), midland zone (H = 2.327) and riverine zone (H = 2.085). Anopheles pharoensis was the dominant species in the riverine zone and accounted for 18.9% of the total number of larvae collected in the riverine zone (Berger-Parkerʼs index = 0.189). A pairwise comparison of species diversity between zones by Shannon diversity t-test showed that only highland and riverine zones were significantly different in species diversity (t(15.876) = − 2.534, P = 0.049). The Simpsonʼs index was highest (1-D = 0.915) in the highland zone, indicating a high level of species evenness, and lowest (1-D = 0.860) in the riverine zone, an indication that species were not evenly represented.
The rarefaction curve for the riverine zone (P in Fig. 3) showed that the common six species were obtained after 8 samples while for the highland zone (N in Fig. 3), the common ten species were obtained after 17 samples. Similarly, no new species were collected from the midland and lowland zones after 14 and 16 samples, respectively.

Rarefaction curves of the number of species against number of samples for ecological zones. Abbreviations: M, highland; N, lowland; O, midland; P, riverine
The combined plots of cumulative species abundance (ln S), Shannonʼs index (H) and log evenness (ln E) (SHE) profiles showed that the four ecological zones were not obviously heterogeneous as the lines representing each measurement did not change much in direction (Fig. 4a). Similarly, species abundance, diversity and evenness were not different between seasons (Fig. 4b).

a SHE profiles for zones. b SHE profiles for seasons. Key: ln S, number of species (taxa); ln E, evenness; H, Shannonʼs index
Monthly data analysis showed that the average number of larvae for all species combined per dip was high in April (2.0 larvae per dip), January (1.99), March (1.93) and May (1.81). A low density of larvae was observed in December and September, 0.83 and 1.07 larvae per dip, respectively. Overall, larval density was high in the long rain season followed by the cold dry season (Fig. 5).

Monthly average larvae per dip and overall mean per season for all species
Although the trend for combined species showed high density during the long rain season (March to May), individual species showed different trends (Fig. 6). Anopheles pharoensis density was high in April during the long rain season and lowest in November (short rain season) when An. gambiae (s.l.) density was highest. On the other hand, Culex quinquefasciatus peaked in May during the long rain season but was lowest in December during the dry season.

Monthly average larvae per dip for individual species
Anopheles gambiae (s.l.) was notably low in abundance during the long rain season (n = 61) and the highest abundance was recorded during the short rain season (n = 263) while An. pharoensis was lowest in abundance (n = 127) during the short rain season. However, statistically there was no significant difference between seasons in An. gambiae (s.l.) abundance (F(3,406) = 2.115, P = 0.098) but An. pharoensis was significantly different in abundance between seasons (F(3,406) = 4.544, P = 0.004). On the other hand, Mansonia species which are the main vectors of RVF in Baringo constituted 0.42% while Culex species which are secondary vectors of RVF constituted 59.97%. The genus Culex was represented by 8 species dominated by Cx. quinquefasciatus.
The highest Shannonʼs diversity index (Table 2) of species was observed during the cold dry season (H = 2.487) whereas the lowest diversity was recorded during the long rain season (H = 2.354). Culex quinquefasciatus was the dominant species during three of the four seasons except the long rain season when Aedes aegypti was the dominant species constituting 42.9% of the total larvae (Berger-Parkerʼs index = 0.429). Culex quinquefasciatus constituted 40, 28.1 and 28.3% during the short rain, cold dry and dry season, respectively. However, these variations in abundance were not statistically different (F(3,406) = 0.036, P = 0.991).
Pairwise comparison between the four seasons by Shannonʼs diversity t-test showed that they were all significantly different from each other in species diversity. When the four seasons were merged into the two groups referred to as dry and wet seasons, still there was a significant difference (t(7570.1) = 7.57, P < 0.0001) with the dry season having a higher species diversity than the wet season. The Simpsonʼs index was highest (1-D = 0.912) during the cold dry season indicating a high level of species evenness. The long rain season had the lowest Simpsonʼs index for species evenness (1-D = 0.896).
The rarefaction curve for the short rain season (D in Fig. 7) showed that the common eight species were obtained after 13 samples, while for cold dry season (A in Fig. 7), the common nine species were obtained after 16 samples. Similarly, no new species were collected during dry season and long rain season after 14 and 15 samples, respectively.

Rarefaction curves of the number of species against number of samples for seasons. Abbreviations: A, Cold dry season; B, Dry season, C, Long rain season, D, Short rain season
Seasonality of larval habitats
Nine categories of larval habitats were sampled, and the most commonly inhabited were lake margins, ditches, swamps, seasonal river beds, water pits, water pans and water springs. The least preferred habitats were the dam, which was consistently sampled, and concrete tanks, which were sampled only when they contained water (Fig. 8a, b). Lake margins receded during the dry season making sampling unfeasible. This happened at the Salabani sampling site on the shores of Lake Baringo and at the Sirata sampling site at the swampy Lake 94 (Fig. 8c–e). Water pans and some water pits also dried completely during the dry season. Swamps, ditches, water springs and riverbed pools persisted throughout the sampling period although the volume of water decreased in water springs and river-bed pools during the dry season.

Larval sites that dried during the dry seasons in Baringo County. a, b, Concrete tank: a external view, b internal view during cold dry season. c Dried edge of Lake Baringo at Salabani. d Dried edge of Lake 94. e Wet area inside Lake 94
A total of 29 larval habitats classified into 9 categories were sampled for several months when they contained water. Overall, 411 samples were taken cumulatively from all habitats during the study duration (2014–2016) and the most sampled habitat type was seasonal river bed at 5 sites totaling 81 samples (Table 3). This was closely followed by Lake Margin at 4 sites and water spring at 3 sites. The habitats that were least in number were dams and swamps, one each at two sites.
Larval habitat preference by culicines and Anopheles species
The ditch had the highest mean of Anopheles larvae (16.6 larvae per sample) followed by swamp (12.4 per sample) and seasonal river bed (10.7 per sample). The ditch was the only habitat which had higher mean of Anopheles larvae than culicine larvae. In the swamp, culicine larvae mean was almost double that of Anopheles larvae. Concrete tank was the least sampled type of habitat but had the highest mean number of culicine larvae (333.7 larvae per sample) with a low mean number of Anopheles larvae (2.6 larvae per sample). Water pit and water pan had the lowest mean number of Anopheles larvae, 1.4 and 1.8 larvae per sample, respectively (Table 3).
Overall, larval habitats were significantly different in terms of larval density (F(8,334) = 2.090, P = 0.036). Multiple comparisons by post-hoc test showed that the concrete tank with a mean of 333.7 per sample was significantly different from dam edge, lake margin, river-bed, water pan and water pit. However, concrete tank was not statistically different from ditch, swamp and water spring which had relatively high mean number per sample. When concrete tank was excluded from the analysis because of its outstandingly high mean number, there was no significant difference between all habitats (F(7,328) = 0.866, P = 0.534).
A separate analysis involving only Anopheles species showed that habitats were significantly different in terms of larval mean per sample (F(8,401) = 9.595, P < 0.0001). Multiple comparisons by post-hoc test showed that the ditch which had the highest mean number of Anopheles larvae was significantly different (P < 0.05) from all other habitats except the swamp (P = 0.233), dam edge (P = 0.728) and concrete tank (P = 0.162). There was no significant difference between concrete tank and all other habitats in terms of Anopheles larval mean.
When analysis was performed for culicines only, there was also a significant difference between all sampled habitats (F(8,401) = 4.903, P < 0.0001) and when concrete tank was excluded (F(7,395) = 5.670, P < 0.0001). Concrete tank, dam edge and water pan were not significantly different from all other larval habitats in terms of the mean number of culicine larvae. On the other hand, water spring which had second highest mean number of culicine larvae was significantly different from ditch, lake margin and river-bed (P < 0.05). Although the swamp had a larval mean ranking 3rd highest, it was only different from ditch and lake margin.
Occurrence of malaria and RVF vectors among the surveyed larval habitats
Out of the five Anopheles species identified, three are malaria vectors: An. funestus (s.l.), An. gambiae (s.l.) and An. pharoensis. Anopheles funestus (s.l.) was only collected from the ditch in the lowland zone while An. gambiae (s.l.) and An. pharoensis were collected from all types of larval habitats in all ecological zones. For An. gambiae (s.l.), riverbed pool had the highest mean number of larvae per sample (2.6) followed by ditch and swamp with means of 2.5 and 2.1 per sample, respectively. Anopheles pharoensis on the other hand had the highest mean of 6.9 larvae per sample in the ditch followed by swamp and riverbed pools with means of 4.0 and 3.5 per sample, respectively. Ditches, riverbed pools and swamp were the most three preferred larval habitats by malaria vectors followed by lake margin (Table 4).
Further analysis to determine An. gambiae (s.l.) habitat preference showed a significant difference between habitats (F(8,401) = 3.891, P < 0.0001). The post-hoc test showed that the water pit which had the least mean number of An. gambiae (s.l.) larvae per sample was significantly different from ditch (P = 0.001) and river-bed (P = 0.011) which had relatively high means. Water pan and water pit were the least preferred habitats by An. gambiae (s.l.). Analysis to determine habitat preference by An. pharoensis showed an overall significant difference between habitats (F(8,401) = 5.949, P = 0.0001). The post-hoc test showed that the ditch which had the highest mean number of An. pharoensis was significantly different from river-bed and lake margin although they also had relatively high means (P < 0.005). There was no significant difference between the ditch and the swamp which had the second highest mean number of An. pharoensis larvae (P = 0.674).
Mansonia species, which are the only known vectors of RVF in Baringo, were collected from swamps, water pits, river-beds and water springs in small numbers. Culex quinquefasciatus and Cx. pipiens, which have only been implicated in RVF virus transmission, were collected from all habitats. Generally, Cx. quinquefasciatus was the most abundant larval species constituting 51.4% of potential arboviral larval species collected from all habitats. Of the three Aedes species collected, only Ae. aegypti and Ae. africanus are known vectors of yellow fever virus which is also an arbovirus belonging to the same group as RVF virus. Aedes aegypti and Ae. africanus were both collected from water pits, water pans and water springs in small numbers but a very large number of Ae. Aegypti was collected from concrete tanks. The concrete tank contributed 98.9% of Ae. aegypti larvae collected from all habitats and had a high mean of 155.9 larvae per sample indicating a high preference of water containers by this species (Table 5).
Statistical analysis was performed only for Cx. quinquefasciatus because it was the most abundant arboviral vector. Although there was a significant difference in habitat preference by Cx. quinquefasciatus (F(8,401) = 2.132, P = 0.032), it was only ditch and river bed that were different from swamp in terms of larval density. Swamp had the highest mean number of Cx. quinquefasciatus larvae while river-bed and ditch had low means (Table 5). All other larval habitats were not significantly different from each other in terms of larval mean for Cx. quinquefasciatus.
Discussion
This study reveals a more complex larval species composition compared to previous studies in Baringo where only four anopheline species and one Aedes species had been identified [13, 33, 38]. Culex larvae have been collected from Baringo by other researchers but identification up to species level has not been undertaken [38]. The present study recorded larvae of 17 species, including Anopheles and culicines. The most favorable seasons for most mosquito species were cold dry and dry seasons as depicted by the high species diversity index. Therefore, the two seasons are appropriate for implementation of larval source management (LSM). Application of larvicides during these two seasons would be effective since there would be no wash off. Highland and lowland zones had high species diversity and should also be targeted for LSM.
The three Aedes species larvae collected in the present study, namely Ae. aegypti, Ae. taylori and Ae. africanus, are all vectors of arboviruses [45] but only Ae. africanus had previously been reported in high altitude woodlands in Baringo during the yellow fever outbreak of 1992–1993 [33]. The information on larval vector species can be instrumental for integrated control strategies in view of the fact that control of immature stages would be more appropriate since they are confined in small aquatic habitats where they cannot escape as opposed to adults which are highly mobile [13, 46].
Mansonia spp. are the main vectors of RVF in Baringo [47] but larval stages are not easy to find most likely because of their habit of attaching to aquatic plants [28, 29]. However, adult mosquitoes of Mansonia species have been collected from Baringo in the previous studies [47,48,49,50]. A few pupae of Mansonia species were collected during this study and were identified to genus level after emergence into adults. Mansonia species were found in habitats with vegetation. Removal of such vegetation can be an effective control method to prevent development of Mansonia [29] and hence reduce their population. Species of Culex were found in all habitats in the four ecological zones. This finding is consistent with previous studies in which one or more Culex species were found in all types of habitat [18, 27]. Whereas Cx. quinquefasciatus did not show a preference for any particular habitat across the four ecological zones, Cx. univittatus was only collected from the lake margin and concrete tank.
The Aedes species incriminated in the transmission of RVF virus (Ae. mcintoshi and Ae. ochraceus) develop and emerge from flood waters after unusually heavy and persistent rainfall [4]. However, these Aedes primary vectors of RVF have not been reported in Baringo, even during the previous active epizootics. Aedes aegypti specifically breed in containers but the present study focused more on sampling large and relatively permanent larval habitats. Nevertheless, sampling during the few months when rain was heavy yielded a large number of Ae. aegypti larvae from two concrete tanks in the lowland zone. The abundance of Ae. aegypti during the long rain season and its confinement to containers makes it easier to control at larval stage unlike adults which rest outdoors in diverse places [35]. A previous survey of domestic and peridomestic water receptacles in Baringo found no Aedes larvae except in one isolated cistern in Marigat town [33]. Similar results were reported from a study in Malaysia which revealed that indoor containers were more preferred larval habitats for Ae. aegypti [32]. Aedes aegypti is the primary vector of dengue, chikungunya and yellow fever viruses [33, 51,52,53]. Its presence in the lowlands is indicative of potential risks of spread of arboviruses in the event of an outbreak.
The presence of An. gambiae (s.l.), An. funestus (s.l.), An. coustani and An. pharoensis, previously reported in Baringo [13, 38] and confirmed in the present study, shows that they are the most predominant Anopheles species in the region. According to the study of Mala et al. [13], An. arabiensis is the most abundant species of An. gambiae complex in Baringo. Anopheles gambiae (s.l.) abundance was not significantly different among seasons. This is consistent with findings in western Kenya where no difference was found in Anopheles larval abundance between seasons [16]. Therefore, it is advisable to control malaria vector larvae in all seasons by targeting all habitats [38].
The present study shows that Anopheles species were distributed in all ecological zones (980–2200 m above sea level) except larvae of An. funestus (s.l.) which were found only in the lowland. This is consistent with studies conducted elsewhere in which it was found that malaria vectors are found in all levels of elevation [54]. The small and open, sunlit water pools preferred by An. gambiae (s.l.) [42, 55] were common in seasonal river beds in midland zone where this species was most abundant. This is similar to findings of studies conducted in Eritrea and Ethiopia where high larval productivity was recorded at stream bed pools [12]. This implies that riverbed pools could sustain malaria vectors responsible for transmission during the dry season in Baringo so they should be targeted for larval source management since they are easily identifiable. Findings from other studies link An. gambiae complex to artificial, environmentally-disturbed habitats and small shallow habitats without emergent vegetation [36, 37]. However, a study conducted in an urban environment in Tanzania demonstrated that it was not clear to define larval habitats for An. gambiae (s.l.) as high densities were found in polluted water [25]. In the present study, An. gambiae (s.l.) and An. pharoensis were found co-existing in the same larval habitats such as riverbed pools, ditches and lake margins. The riverbed pools and ditches were small in size and discrete hence can easily be treated to destroy larvae. On the other hand, An. funestus (s.l.) prefers deeper and more persistent habitats with vegetation [36]. The three species [An. gambiae (s.l.), An. pharoensis and An. funestus (s.l.)] appear to prefer different types of habitats which are all present in Baringo County, a factor that could be enhancing malaria transmission throughout the year.
Concrete tanks, water springs and swamps should be targeted for potential RVF vectors, while ditches and riverbed pools should be targeted for potential malaria vectors. Larval source management would reduce mosquito vector populations and supplement the current vector control strategies which target only indoor adult stages. Thus, larval source management is a feasible strategy than can be implemented in Baringo to control both indoor and outdoor resting mosquitoes such as An. pharoensis and culicine species.
Limitations of the study
Anopheles gambiae (s.l.) larvae were not identified to the species level by molecular techniques and Mansonia species were not identified to the species level. This study focused on identifying larval sites, seasonal effects and species diversity and therefore sizes of larval habitats were not quantified. Future studies, aimed specifically at productivity of larval habitats, should take into account measurements of such sites and perform molecular identification of species complexes.
Conclusions
The present study reports a higher diversity of culicine and Anopheles larvae than previous studies in Baringo. Occurrence of Mansonia species, Aedes species and several species of Culex indicates the potential for a rapid spread of arboviral diseases such as RVF and yellow fever which have been reported in Baringo in previous years. The most important larval habitats were riverbed pools, ditches, swamps and lake margins which should be targeted during larval control operations. The presence of malaria vectors in all seasons implies that transmission of malaria occurs throughout the year unlike previous assumptions that malaria transmission is seasonal in semi-arid areas. Knowledge on vector species diversity, availability and types of preferred larval sites can inform comprehensive control strategies such as inclusion of environmental management as a component of integrated vector management. These results have implications for control strategies and suggest a greater need for increased surveillance and research in the region due to ongoing climate change.
Availability of data and materials
The study is part of a larger project on “Early warning systems for improved human health and resilience to climate-sensitive vector-borne diseases in Kenya”. The data can be requested from the project’s scientific committee through the following e-mail address: climatechange@jooust.ac.ke.
Abbreviations
- ANOVA:
-
analysis of variance
- IRS:
-
indoor residual spraying
- IVM:
-
integrated vector management
- LLINs:
-
long-lasting insecticide-treated nets
- LSM:
-
larval source management
- RVF:
-
Rift Valley fever
References
WHO. Global vector control reponse 2017 to 2030. Geneva: World Health Organization; 2017. p. 2017.
Hotez PJ, Kamath A. Neglected tropical diseases in sub-saharan Africa: review of their prevalence, distribution, and disease burden. PLoS Negl Trop Dis. 2009;3:e412.
WHO. Vector control methods for use by individuals and communities. Geneva: World Health Organization; 1997. p. 10–7.
CDC. Rift Valley fever transmission. Atlanta: Centers for Disease Control and Prevention; 2013.
Murithi RM, Munyua P, Ithondeka PM, Macharia JM, Hightower A, Luman ET, et al. Rift Valley fever in Kenya: history of epizootics and identification of vulnerable districts. Epidemiol Infect. 2011;139:372–80.
Nguku PM, Sharif S, Mutonga D, Amwayi S, Omolo J, Mohammed O, et al. An investigation of a major outbreak of Rift Valley fever in Kenya: 2006–2007. Am J Trop Med Hyg. 2010;83(Suppl. 2):5–13.
WHO. World malaria report 2016. Geneva: World Health Organization; 2016.
KIRA-Kenya Inter-Agency Rapid Assessment. Baringo County baseline analysis. Nairobi; 2014.http://www.humanitarianresponse.info/system/file/document/files/Baringo. Accessed 18 Jan 2016.
Omondi CJ, Onguru D, Kamau L, Nanyingi M, Ong’amo G, Estambale B. Perennial transmission of malaria in the low altitude areas of Baringo County, Kenya. Malar J. 2017;16:257.
Kipruto EK, Ochieng AO, Anyona DN, Mbalanya M, Mutua EN, Onguru D, et al. Effects of climatic variability on malaria trends in Baringo County, Kenya. Malar J. 2017;16:220.
Kiszewski AE, Teklehaimanot A. A review of the clinical and epidemiologic burdens of epidemic malaria. Am J Med Trop Hyg. 2004;71(Suppl. 2):128–35.
Shililu J, Mbogo C, Ghebremeskel T, Githure J, Novak R. Mosquito larval habitats in a semiarid ecosystem in Eritrea: impact of larval habitat management on Anopheles arabiensis population. Am J Trop Med Hyg. 2007;76:103–10.
Mala AO, Irungu LW, Shililu JI, Muturi EJ, Mbogo C, Njagi JK, Githure JI. Dry season ecology of Anopheles gambiae complex mosquitoes at larval habitats in two traditionally semi arid villages in Baringo, Kenya. Parasit Vectors. 2011;4:25.
Vanlalruia K, Senthilkumar N, Gurusubramanian G. Diversity and abundance of mosquito species in relation to their larval habitats in Mizoram, North Eastern Himalayan region. Acta Trop. 2014;137:1–18.
Bond JG, Casas-Martínez M, Quiroz-Martínez H, Novelo-Gutiérrez R, Marina CF, Ulloa A, et al. Diversity of mosquitoes and the aquatic insects associated with their oviposition sites along the Pacific coast of Mexico. Parasit Vectors. 2014;7:41.
Kweka EJ, Zhou G, Munga S, Lee M-C, Atieli HE, Atieli M, et al. Anopheline larval habitats seasonality and species distribution: a prerequisite for effective targeted larval habitats control programmes. PLoS ONE. 2012;7:e52084.
Mendoza F, Ibáñez-Bernal S, Cabreno-Sañudo FJ. A standardized sampling method to estimmate mosquito richness and abundance for research and public health surveillance programmes. Bull Entomol Res. 2008;98:323–32.
Muturi JE, Shililu JI, Weidong GU, Jacob BG, Githure JI, Novak RJ. Larval habitat dynamics and diversity of Culex mosquitoes in rice agro ecosystem in Mwea, Kenya. Am J Trop Med Hyg. 2007;76:95–102.
Govoetchan R, Gnanguenon V, Ogouwalé E, Oké-Agbo F, Azondékon R, Sovi A, et al. Dry season refugia for anopheline larvae and mapping of the seasonal distribution in mosquito larval habitats in Kandi, northeastern Benin. Parasit Vectors. 2014;7:137.
Dida GO, Anyona DN, Abuom PO, Akoko D, Adoka SO, Matano A-S. Spatial distrribution and habitat characterization of mosquito species during the dry season along the Mara River and its tributaries, in Kenya and Tanzania. Infect Dis Poverty. 2018;7:2.
Benedict MQ, Sandve SR, Wilkins EE, Roberts JM. Relationship of larval desiccation to Anopheles gambiae Giles and An. arabiensis Patton survival. J Vector Ecol. 2010;35:116–23.
Teklu BM, Tekie H, McCartney M, Kibret S. The effect of physical water quality and water level changes on the occurrence and density of Anopheles mosquito larvae around the shoreline of the Koka reservoir, central Ethiopia. Hydrol Earth Syst Sci. 2010;14:2595–603.
Amadi JA, Ong’amo GO, Olago DO, Oriaso SO, Nyamongo IK, Estambale BBA. Mapping potential of Anopheles gambiae s.l. larval distribution using remotely sensed climatic and environmental variables in Baringo, Kenya. Med Vet Entomol. 2018. https://doi.org/10.1111/mve.12312.
Sinka M, Bangs M, Manguin S, Coetzee M, Mbogo C, Hemingway J, et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis. Parasit Vectors. 2010;3:117.
Sattler MA, Mtasiwa D, Kiama M, Premji Z, Tanner M, Killeen GF, et al. Habitat characterization and spatial distribution of Anopheles sp. mosquito larvae in Dar es Salaam (Tanzania) during an extended dry period. Malar J. 2005;4:4.
Mwangangi JM, Muturi EJ, Mbogo CM. Seasonal mosquito larval abundance and composition in Kibwezi, lower eastern Kenya. J Vector Borne Dis. 2009;46:65–71.
Fillinger U, Lindsay SW. Larval source management for malaria control in Africa: myths and reality. Malar J. 2011;10:353.
Rajendran G, Krishnamoorthy K, Sabesan S, Panicker KN. Attachment behaviour of immatures of Mansonia annulifera (Theobald), 1901. Indian J Med Res. 1989;89:192–7.
Chandra G, Ghosh A, Biswas D, Chatterjee SN. Host plant preference of Mansonia mosquitoes. J Aquat Plant Manage. 2006;44:42–144.
Vijayakumari KN, Murali D. Oviposition and larval attachment by Mansonia mosquitoes on host plants. Millennium Zool. 2010;11:23–7.
Dom NC, Ahmad AH, Ismail R. Habitat characterization of Aedes sp. breeding in urban hotspot area. Procd Soc Behav. 2013;85:100–9.
Cheong WH. Preferred Aedes aegypti larval habitats in urban areas. Bull World Health Organ. 1967;36:586–9.
Reiter P, Cordellier R, Ouma JO, Cropp CB, Savage HM, Sanders EJ, et al. First recorded outbreak of yellow fever in Kenya, 1992–1993. ii. Entomologic investigations. Am J Trop Med Hyg. 1998;59:650–6.
Omukunda E, Githeko A, Ndong’a MS, Mushinzimana E, Yan G. Effect of swamp cultivation on distribution of anopheline larval habitats in Western Kenya. J Vector Borne Dis. 2012;49:61–71.
Ndenga BA, Mutuku FM, Ngugi HN, Mbakaya JO, Aswni P, Musunzaji PS, et al. Characteristics of Aedes aegypti adult mosquitoes in rural and urban areas of western and coastal Kenya. PLoS ONE. 2017;12:e0189971.
Gimnig J, Ombok M, Kamau L, Hawley W. Characteristics of larval anopheline (Diptera: Culicidae) habitats in Western Kenya. J Med Entomol. 2001;38:282–8.
Carlson JC, Byrd BD, Omlin FX. Field assessments in western Kenya link malaria vectors to environmentally disturbed habitats during the dry season. BMC Public Health. 2004;4:33.
Arum SO, Kokwaro ED, Mulwa F. Diversity and colonization of permanent and semi-permanent breeding habitats by Anopheles mosquitoes in a semi-arid ecosystem of Baringo district in Kenya, in Second RUFORUM Biennial Meeting. Uganda: Entebbe; 2010.
Kenya Information Guide; 2015. Baringo County, Kenya. http://www.kenya-information-guide.com/baringo-county.html. Accessed 25 July 2018.
Funk CC, Peterson PJ, Landsfeld MF, Pedreros DH, Verdin JP, Rowland JD, et al. A quasi-global precipitation time series for drought monitoring. US Geological Survey Data Series 832. Reston, Virginia: U.S. Department of the Interior; 2014.
Highton RB. Taxonomic keys for the identification of the Afrotropical mosquitoes. Highton April: Revised and prepared by R.B; 1983.
Gillies MT, Coetzee M. A supplement to the Anophelinae of Africa south of the Sahara (Afrotropical region). Johannesburg: Publication of the South African Institute of Medical Research; 1987.
Morris EK, Caruso T, Buscot F, Fischer M, Hancock C, Maier TS, et al. Choosing and using diversity indices: insights for ecological applications from the German Biodiversity Exploratories. Ecol Evol. 2014;4:3514–24.
Hutcheson KA. Test for comparing diversity based on the Shannon formula. J Theor Biol. 1970;29:151–4.
Gillett JD. Common African mosquitoes and their medical importance. London: W.H. Med. Books Ltd; 1972.
Killeen G, Fillinger U, Knols BGJ. Advantages of larval control for African malaria vectors: low mobility and behavioural responsiveness of immature mosquito stages allow high effective coverage. Malar J. 2002;1:8.
Lutomiah J, Bast J, Clark J, Richardson J, Yalwala S, Oullo D, et al. Abundance, diversity, and distribution of mosquito vectors in selected ecological regions of Kenya: public health implications. J Vector Ecol. 2013;38:134–42.
Sang R, Kioko E, Lutomiah J, Warigia M, Ochieng C, O’Guinn M, et al. Rift Valley fever virus epidemic in Kenya, 2006/2007: the entomologic investigations. Am J Trop Med Hyg. 2010;83:28–37.
Omondi D, Masiga D, Ajamma YU, Fielding BC, Njoroge L, Villinger J. Unraveling host-vector-arbovirus interactions by two-gene high resolution melting mosquito bloodmeal analysis in a Kenyan wildlife-livestock interface. PLoS ONE. 2015;10:e0134375.
Ondiba IM, Oyieke FA, Nyamongo IK, Estambale BBA. Diversity, distribution and abundance of potential rift valley fever vectors in Baringo County. Kenya. Intern J Mosq Res. 2017;4:42–8.
Ibanez-Bernal S, Briseno B, Mutebi JP, Argot E, Rodriguez G, Martinez-Campos C, Paz R, et al. First record in America of Aedes albopictus naturally infected with dengue virus during the 1995 outbreak at Reynosa, Mexico. Med Vet Entomol. 1997;11:305–9.
O’Meara GF, Evans LF, Gettman AD, Cuda JP. Spread of Aedes albopictus and decline of Ae. aegypti (Diptera: Culicidae) in Florida. J Med Entomol. 1995;32:554–62.
Burt FJ, Rolph MS, Rulli NE, Mahalingam S, Heise MT. Chikungunya: a re-emerging virus. Lancet. 2012;379:662–71.
Minakawa N, Sonye G, Mogi M, Githeko A, Yan G. The effects of climatic factors on the distribution and abundance of malaria vectors in Kenya. J Med Entomol. 2002;39:833–41.
Gillies MT, Meillon D. The Anophelinae of Africa south of the Sahara (Ethiopian zoogeographic region). Johannesburg: Publications of the South African Institute for Medical Research; 1968.
Acknowledgements
We thank the field project staff, D. Anyona, E. Agure, J. Chepkosgei and M. Mbaranya, for field logistics and data collection and A. Ochieng for study site map preparation. We also thank M. Rotich and R. Bor for assisting in species identification. The study received financial assistance from the WHO’s Special Programme for Research and Training in Tropical Diseases (TDR) through a grant agreement with the International Development Research Centre of Canada (106905-00). This in turn funded Project no. B20278 through a collaboration between the University of Nairobi and Jaramogi Oginga Odinga University of Science and Technology. We thank all the above organizations and institutions for funding the project and making the study possible.
Funding
This study was funded by the WHO’s Special Programme for Research and Training in Tropical Diseases (TDR) through a grant agreement with the International Development Research Centre of Canada (106905-00).
Author information
Authors and Affiliations
Contributions
IMO designed the study, mapped larval sites and collected data in the field, identified specimens taxonomically, carried out data entry and analysis, and wrote the first draft of the manuscript. FAO participated in the study design, enhanced the literature survey, and edited the article. DKA participated in data collection and species identification. BBAE and IKN conceived the idea for the study and participated in the study design. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Ondiba, I.M., Oyieke, F.A., Athinya, D.K. et al. Larval species diversity, seasonal occurrence and larval habitat preference of mosquitoes transmitting Rift Valley fever and malaria in Baringo County, Kenya. Parasites Vectors 12, 295 (2019). https://doi.org/10.1186/s13071-019-3557-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-019-3557-x