
Abstract
Eph receptors constitute the largest family of receptor tyrosine kinases, comprising 14 distinct members classified into two subgroups: EphAs and EphBs.. Despite their essential functions in normal physiological processes, accumulating evidence suggests that the involvement of the Eph family in cancer is characterized by a dual and often contradictory nature. Research indicates that Eph/ephrin bidirectional signaling influences cell–cell communication, subsequently regulating cell migration, adhesion, differentiation and proliferation. The contradictory functionalities may arise from the diversity of Eph signaling pathways and the heterogeneity of different cancer microenvironment. In this review, we aim to discuss the dual role of the Eph receptors in tumor development, attempting to elucidate the paradoxical functionality through an exploration of Eph receptor signaling pathways, angiogenesis, immune responses, and more. Our objective is to provide a comprehensive understanding of the molecular mechanisms underlying tumor development. Additionally, we will explore the evolving landscape of utilizing Eph receptors as potential targets for tumor therapy and diagnostic tools.
Similar content being viewed by others
Introduction
The human erythropoietin-producing hepatocellular (Eph) receptors consitute the largest family of receptor tyrosine kinases (RTKs), comprising 14 distinct members classified into two subgroups: EphAs and EphBs. The EphA subgroup includes nine receptors (EphA1-8 and EphA10), while the EphB subgroup consists five members (EphB1-4 and EphB6) [1]. Each member is comprised of an extracellular domain, a transmembrane region, and an intracellular tyrosine kinase domain. Eph receptors initiate signal transduction by interacting with their ligands, ephrins. These receptors are located on cell surfaces and transmit signals upon binding to the ligands [2]. EphA receptors predominantly bind to glycosylphosphatidylinositol (GPI) -linked ephrin-A ligands, whereas EphB receptors bind to ephrin-B ligands possessing transmembrane and intracellular domains [3]. Additionally, EphA4 and EphB2 receptors can also bind eprhins of a different class [4].
Eph/ephrin signaling displays at least two unique characteristics compared to other RTKs. Firstly, it exhibits bidirectional signaling, transmitting signals within both the receptor- and ligand-expressing cells upon binding to the membrane-bound ligand (forward and reverse signalings) [1]. Secondly, Eph receptors may function in a kinase activity-independent mechanism. The Eph/ephrin system is influenced not only by their own interactions but also by other RTKs [5].
Throughout various developmental processes, Eph/ephrin signaling plays diverse biological roles, including growth cone retraction in axon guidance, the formation of synaptic connections between neurons, cell sorting in embryo patterning, cell migration, platelet aggregation, and blood vessel remodeling [4, 6]. They regulate essential processes such as cell adhesion, migration, and cell growth. Despite its crucial functions in normal physiological processes, mounting evidence suggests that the Eph family's involvement in cancer is characterized by a dual and often contradictory role [7]. On one hand, some studies propose that the Eph family exhibits tumor-suppressive effects in certain tumor types, inhibiting tumor cell proliferation, invasion, and metastasis, thereby impeding tumor development. Conversely, an increasing body of evidence indicates that Eph receptors are associated with adverse prognosis, facilitating tumor growth and metastasis in other tumor types. Recent studies have focused on the functions of Eph/ephrin signaling in vasculogenesis and immune modulation [8]. However, the mechanism of Eph/ephrin involvement in cancer metastasis, invasion and angiogenesis remains to be fully understood [9]. The contradictory functionality may arise from the diversity of Eph signaling pathways and the heterogeneity of the different cancer microenvironments. In this review, our objective is to elucidate the dual role of Eph receptors in tumor development, attempting to explain the paradoxical functionality through an exploration of Eph receptor signaling pathways, angiogenesis, immune responses, and more. We aim to provide a deeper understanding of the molecular mechanisms underlying tumor development. Additionally, we will explore the evolving landscape of using Eph receptors as targets for tumor therapy and diagnostic tools.
The dual role of Eph family in cancer development
Tumorigenesis is a complex process characterized by distinct hallmarks extensively studied and expanded to encompass 10 organizing principles, including sustaining proliferative signaling, evading growth suppressors, resisting cell death, enabling replicative immortality, inducing angiogenesis, activating invasion and metastasis, genome instability, inflammation, reprogramming of energy metabolism, and evading immune destruction [10]. The tumor microenvironment also plays a critical role in increasing the complexity of tumorigenesis [10].
The Eph family of receptor tyrosine kinases has emerged as a crucial player in tumorigenesis, displaying a diverse range of roles in tumor development and progression. The function of Eph receptors and their ligands extends beyond a binary classification of either tumor suppression or tumor promotion (Table 1). Instead, they play diverse roles in different types of tumors, underscoring their complexity and context-dependent nature [8]. As depicted in Fig. 1, the Eph receptor expression exhibits a correlation with cancer patient survival. The Hazard Ratio (HR) indicates the variability of Eph functions across different cancer types (Fig. 1). To ensure that survival analyses involving multiple tumor groups were presented, the figures also displayed those with P-values exceeding 0.05.

The Hazard Ratio of survival events associated with Eph receptor expressions across different cancer types. The TISCH2 database (http://tisch.compgenomics.org/), containing scRNA-seq data from more than 50 cancer types, was utilized to analyze the association of Eph receptor expression and HR of survival. An HR > 1 indicates a high risk of death, while an HR < 1 indicates a low risk of death. A HR of patient survival according to the expression of EphA2 in distinct cancer types from datasets. B HR of patient survival according to the expression of EphB1 in distinct cancer types from datasets. C HR of patient survival according to the expression of EphB4 in distinct cancer types from datasets
Promoting tumor progression
Numerous studies have reported the overexpression of Eph receptors and ephrins in various types of cancer, including breast cancer, gastrointestinal cancer, melanoma and small-cell lung carcinoma (SCLC) [44]. These molecules exhibit increased levels in tumors, especially during the more aggressive stages of tumor progression [45]. For instance, EphA2, expressed at minimal levels in normal human breast epithelium [46], is significantly overexpressed in 60% to 80% of breast cancers [47]. Additionally, EphA2 upregulation has been observed in liver, prostate cancer, glioblastoma, esophageal squamous cell carcinoma, ovarian cancer, and melanoma, with the highest levels detected in the most invasive tumor cells [17]. Conversely, inhibiting EphA2 expression using small interfering RNA (siRNA) has been shown to impede the malignant progression of pancreatic, ovarian, and mesothelioma tumor cell lines, underscoring the pivotal role of EphA2 in tumor development [48]. In small cell lung cancer (SCLC) cell lines and tumor samples, transcripts encoding members of EphB receptors and ephrins are coexpressed. In human leukemia-lymphoma cell lines, both the EphB4 receptor and ephrinB2 ligand are commonly co-expressed, with EphB4 mRNA detected in 68/70 cell lines and 58/70 cell lines displaying positive ephrinB2 mRNA expression [49]. Although evidence suggests that the interaction between Eph receptors and other oncogenic pathways promotes tumor cell malignancy, this might occur in an ephrin-independent manner [5, 50, 51]. Of note, a paradoxical early response in the activation of cell surface receptors is the downregulation of these receptors, which involves ligand-stimulated endocytosis. However, the activity of RTK within intracellular lysosome following their internalization remains to be discussed [52].
Inhibiting tumor development
Numerous studies have provided compelling evidence supporting the role of Eph receptors in tumor suppression [50]. Specifically, EphB1 has been found to be down-regulated in gastric cancer, renal cell carcinomas and colorectal cancer [53, 54]. Additionally, there is a strong correlation between the loss of EphB1 protein and both metastasis and reduced survival rates in patients with serous ovarian cancer [55]. Ligand-dependent EphB1 signaling has been shown to effectively suppress glioma invasion and is positively correlated with patient survival [56]. In the context of breast cancer, treatment with ephrinA1-Fc has been reported to attenuate epidermal growth factor-mediated phosphorylation of Erk and inhibit the transformation of NIH3T3 cells expressing v-erbB2 [57]. Another member of the Eph receptors, EphA2, has been linked to increased susceptibility to chemical carcinogen-induced skin cancer, as well as elevated tumor cell proliferation and phosphorylation of Erk when deficient in mice [58]. In neuroblastoma patients, the expression of EphB6, ephrin-B2, and ephrin-B3 in tumors has been associated with a positive prognosis. Additionally, ectopic expression of EphB6 has shown the ability to suppress the malignant phenotype of neuroblastoma cell lines [59]. A study on squamous cell carcinoma revealed a significant down-regulation of EphB1 by a factor of 1.1-fold and a similar down-regulation of ephrinB2 by a factor of 1.0-fold [60]. Moreover, the EphB4 receptor has been reported to function as a tumour suppressor in a mouse xenograft model of breast cancer. This effect is elicited when the receptor is stimulated by its ligand, ephrinB2 [13].
The potential mechanisms of Eph function in tumor development
Although the role of Eph receptors in cancer development has been observed, the exact mechanisms underlying their oncogenic effects are still not fully understood [50]. Studies have shown that the overexpression of Eph receptors and ephrins is associated with angiogenesis and metastasis in various human cancers [17]. It has been hypothesized that these receptors may play a central role in angiogenesis or metastasis, rather than acting as primary oncogenes [45]. Ongoing research is dedicated to unraveling the complex cellular and molecular mechanisms involved in Eph-ephrin interactions and signaling. Our current focus is on understanding the potential mechanisms behind the dual roles of Eph receptors. Given their involvement in critical processes such as angiogenesis in tumor tissues, these molecules have the potential to serve as prognostic markers for tumors and show promise as targets for cancer therapy [17].
Conflicting effects of Eph signaling
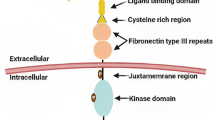
The Eph receptor family share a common structural feature known as the RTK domain, which contains the catalytic kinase domain responsible for phosphorylation. The extracellular domain of Eph receptors includes an N-terminal ligand-binding domain (LBD), a cysteine-rich domain (CRD) with an epidermal growth factor (EGF)-like motif, and two fibronectin-type III repests. This is followed by a single-pass transmembrane domain and an intracellular segment consisting of a juxtamembrane region, a tyrosine kinase domain, a sterile α motif (SAM), and a PDZ (postsynaptic densityprotein PSD95, Drosophila disc large tumor suppressor DlgA, and zonula occludens-1 protein ZO-1) -binding motif [61, 62]. Both EphA and EphB receptors share similar structural elements, including the extracellular ligand-binding domain, the transmembrane domain, and intracellular domains that initiate signaling cascades (Fig. 2A). Activation of EphA or EphB receptors can be induced preferentially by ephrin-A or ephrin-B ligands, respectively, or through cross-activation. In addition to ligand-induced phosphorylation signals, ligand-independent receptor clustering can generate oligomeric assemblies and modulate transmembrane signaling [63]. Eph receptors can also signal in a ligand-independent manner, particularly when they are overexpressed, as observed in cancer cells [64]. Eph receptors and ephrin ligands can engage in cis-interactions when both the ligand and receptor are expressed in the same cells. The diverse signaling mechanisms add complexity to the Eph-ephrin system, contributing to its diverse function in tumorigenesis.

The molecular structure of Ephrin-Eph and signaling pathway. A The molecular structures of ephrinA and ephrinB are different, while their receptors are similar. B, C The EphA and EphB receptor can bind to the ligands present on the same cell with the receptor, known as cis signaling, activating downstream signals. D, E The EphA and EphB receptor can bind to ligands located on neighboring cells, referred to as trans signaling, activating downstream signals. F Ephrin-Eph signaling engages in crosstalk with other RTKs such as EGFR and GFR, involving ERK, Ras and other downstream molecules
Forward and reverse signaling: tyrosine kinase-dependent pathways
Eph tyrosine kinases play a crucial role in a bidirectional signaling, involving forward signaling from ephrin-expressing cells to Eph receptor-expressing cells and reverse signaling from Eph receptor-expressing cells to ephrin-expressing cells [65]. Eph receptors mediate tyrosine kinase-dependent “forward” signaling, while ephrins, specifically ephrinB proteins, mediate ‘reverse’ signaling through phosphorylation by cytoplasmic tyrosine kinases [66] (Fig. 3A). Both types of signaling can lead to cell repulsion, depending on clustering intensity, downstream signaling strength, and Eph/ephrin internalization rates [67]. Eph-mediated forward and ephrin-mediated reverse signaling can be initiated through tyrosine phosphorylation of intracellular residues [68].

Working models of Eph receptors and their ligands in physiological functions. A Ephrin-Eph signaling contains reverse signal and forward signal, leading to the phosphorylation of downstream molecules. The forward signal influences the expression of additional downstream molecules, thereby impacting processes such as angiogenesis, cell proliferation, cell adhesion, and migration. B Ephrin-Eph signaling engages in cross talk with other RTKs, such as VEGFR, exerting regulatory effects that can either suppress or promote tumor development
Forward signaling of Eph receptors plays significant roles in various physiological and pathological processes, including embryonic development, neuronal guidance, immune responses, and cancer progression. Mutations that impair kinase function of Eph receptors, such as EphB2, EphA3, EphA5, are frequently found in cancer, suggesting a tumor suppressor role for Eph receptor forward signaling [69,70,71] (Table 2). Eph forward signaling inhibits proliferation, survival, migration and invasion in several cancer types through regulation of oncogenic signaling pathways, including ERK, RAS/MAPK, PI3K-AKT, Abl-Crk [11, 13, 50, 61]. Stimulation of EphA2 by ephrinA1 inhibits Akt activation and subsequently induces dephosphorylation of EphA2 on S897 [11]. EphrinA1-induced stimulation of EphA2 also leads to reduced ERK phosphorylation and suppresses growth of primary keratinocytes and prostate carcinoma cells [50]. EphB4, in breast cancer cells, activates an anti-oncogenic Abl-Crk pathway, inhibiting cell viability, proliferation, motility, and invasion, and downregulates the pro-invasive matrix metalloprotease, MMP-2 [13].
Eph forward signaling plays a crucial role in regulating the actin cytoskeleton, which is essential for cell migration and axon guidance. EphB forward signaling is pivotal for filopodial motility and synapse formation, activating PAK, a serine/threonine kinase that governs actin dynamics [67]. Eph receptors also contribute to epithelial homeostasis maintainance and can induce mesenchymal-to-epithelial transition (MET) in cancer cells. For instance, EphA2, EphA4 and several EphB forward signaling pathways, including EphB3 and EphB2, are involved in promoting MET by enhancing the expression of E-cadherin [61, 84]. Examining EphA2 in more detail, its phosphorylated tyrosine kinase activity inhibits the chemotactic migration of cancer cells. Conversely, ligand-independent overexpression of EphA2 promotes cancer cell migration through AKT-induced serine phosphorylation [11]. Importantly, EphrinA1 stimulation of EphA2 disrupts the interaction with AKT, leading to the dephosphorylation on S897 on EphA2. This, in turn, blocks the effects on migration and cell invasion [11]. Furthermore, the upregulation of EphA3 inhibits phosphorylation of PI3K/STAT3/AKT, enhancing apoptosis and reversing cellular resistance to cisplatin [85, 86].
Eph reverse signaling, initiated by ephrins within host cells, adds complexity to the Eph-ephrin system. Upon engagement with Eph receptors in neighboring cells, ephrins activate their cytoplasmic tails, recruiting and activating various intracellular signaling components, such as Src family kinases, ultimately influencing cellular responses like morphological changes, motility, and differentiation. The ephrinB cytoplasmic tail recruits and activates Src family kinases, resulting in tyrosine and serine phosphorylation and the recruitment of signaling effectors [67]. In vivo observations have highlighted the significance of ephrinB2-PDZ interaction for reverse signaling in lymphatic vessel development [87]. EphrinB2 reverse signaling triggers substantial morphological changes and increased motility [56], aligning with previous findings demonstrating that ephrin-B2 reverse signaling increases the motility of glioma cells [88]. During osteoclast differentiation, ephrinB2 expression is evident, and reverse signaling through ephrinB2 inhibits osteoclast differentiation by suppressing transcription factors Fos and NFATc1 [89].
In various cancer types, soluble Fc fusion proteins of ephrin ligands activate Eph forward signaling, resulting in decreased proliferation, survival, migration and invasion of various cancer cells in culture, also inhibiting tumor growth in several mouse models [13, 61]. The apparent paradox of Eph receptors being highly expressed but poorly activated by ephrins, as evidenced by their low level of tyrosine phosphorylation, suggests that Eph forward signaling may even be detrimental to tumor progression [90].
Eph cis- and trans- signaling: opposing functions
Eph receptors and ephrins engage in an intricate interplay involving both cis-interactions (within the same cell) (Fig. 2B and C) and trans-interactions (between neighboring cells) (Fig. 2D and E). Cis-interactions typically occur when both the ligand and receptor are expressed in the same cells [91]. Notably, cis-interactions do not activate Eph receptor tyrosine kinase activity. Instead, they diminish the receptor’s interaction with ephrin on adjacent cells, thereby dampening kinase activity [92]. The inhibition of cis-signal to trans-signal adds complexity to Eph receptor signaling and has been observed in various immune cell types, including NK cells, DC cells, mast cells, and T cells [93,94,95]. Plexin receptors and Notch receptors are also influenced by cis-binding [96]. The cell’s response to ligand-receptor interactions varies depending on expression levels and cell–cell contact. Tetramers of receptors and ligands are essential for both cis- and trans- signaling. The higher the number of cell surface receptors, the more likely they are to interact with ligands expressed on the same cells [96].
To explore the role of genes in tumorigenesis, researchers commonly employ gain-of-function or loss-of-function methodologies. In brief, introducing Eph receptors or ephrin ligand through transfection results in reduced kinase activity [97], leading to enhanced migration and invasion of cancer cells [41]. Conversely, using siRNA against Eph receptors leads to decreased migration and invasion [41]. For instance, Knockdown of EphA7 suppresses proliferation and metastasis in A549 human lung cancer cells [98]. Knockdown of EphB1 in medullobalstomas cells significantly reduces migration, which correlates with decreased β1-integrin expression and phosphorylated Src [99]. It is worth noting that Eph phosphorylative activity can suppress the progression of colorectal cancer [100]. Ligand-activated EphB1 signaling inhibits cancer cell migration and invasion through inducing EphB1 phosphorylation. Cis-interactions could be a strategy adopted by cancer cells to evade the tumor suppressive effects of Eph receptor signaling induced by trans-binding of ephrins [81].Upon tumor initiation, Eph receptor expression is upregulated through oncogenic signaling pathways [57]. However, the ephrin ligands often undergo down-regulation or are unable to bind to receptors due to the loss of cell adhesion [101]. Eph receptors require activation by ligands attached to the surface of neighboring cells, indicating that stable cell–cell adhesion may be necessary for Eph activation [101]. Another study suggests that impaired cell–cell adhesion in tumor cells hinders the activation of Eph receptors by ephrins located on adjacent cells [50].
In some cases, tumors exhibit EphA2 overexpression, often associated with the loss of corresponding ligands [102]. Eph receptors and co-expressed ligands engage in direct cis interactions, devoid of initiating intracellular signals. Nevertheless, this cis interaction hinders trans-interactions [92]. Similarly, coexpressed ephrin ligands with Eph receptor attenuate Eph receptor kinase activation [103]. Cis-interactions are ligand-independent, rendering Eph receptors less responsive to Ephrin ligands [6]. Trans-interactions, on the other hand, are ligand-dependent and rely on specific ephrins for Eph receptor functionality. Despite high expression in cancers, Eph recetors within cancer cells are often poorly tyrosine-phosphorylated [104].
The reversible process of epithelial-mesenchymal transtion (EMT) and mesenchymal-epithelial transtion (MET) further illustrates this complexity. In EMT cells, EphB1 cis-signaling counteracts trans-signaling due to reduced cell–cell contact [105]. However, in the MET state, where cell–cell adhesion is regained, EphB1 and ligand ephrinB2 interactions in trans promote cancer cell stemness, affecting genes like Nanog and Sox2. Cancer cells acquire stem cell characteristics and resistant to chemotherapy via context-dependent EphB1 signaling.
Ligand-dependent- and -independent signaling
Eph receptors have the ability to form clusters through direct Eph-Eph interactions, even in the absence of Ephrin ligands [62]. This clustering is facilitated by the ligand-binding domain (LBD) and cysteine-rich domain (CRD), both of which play a crucial role in promoting Eph-Eph interaction [62, 65] . Studies have indicated that the FN, CRD and LBD domains are pivotal in Eph signaling, allowing them to form tetrameric complexes independently of ephrins. It’s noteworthy that the FN domain may also be significant for EphB1 binding with ephrin ligands other than EphrinB2 [105].
Eph receptors and their ligands, ephrins, engage in a complex interplay of both ligand-dependent and ligand-independent signaling. Ligand-dependent signaling occurs when Eph receptors interact with their ligands (ephrins) on adjacent cells, initiating a series of events, including receptor clustering and subsequent phosphorylation. Conversely, ligand-independent signaling involves Eph receptors functioning independently of ephrin engagement. In cases where Eph receptors exhibit low levels of phosphorylation in cancer cells, their tumor-promoting activities are likely to be independent of Ephrin stimulation [61]. Factors such as increased phosphotyrosine phosphatases activity or the loss of E-cadherin in tumor cells can lead to under-phosphorylation of Eph receptors, despite their overexpression [50]. Loss of cell–cell contacts can also prevent ligand stimulation from neighouring cells, reducing Eph receptor phosphorylation [105]. E-cadherin plays a role in regulating the cell surface localization of EphA2 in breast cancer cells, and the loss of cell–cell contacts can impede interaction with ephrin ligands [101], making Eph receptor oncogenic activity appear to be ligand independent [50]. Upregulation of Eph receptors can facilitate Eph receptor-mediated signaling through dimerization and higher-order oligomerisation, independent of ephrin ligation, and involving intracellular SAM domain and extracellular LBD-LBD, CRD-CRD interface [62]. Ligand-independent Eph receptor signaling is distinct from, and sometimes even opposite to, ligand-induced RTK activity [62].
Ligand-independent Eph receptor signaling operates through various mechanisms. Besides tyrosine residues, Eph receptors can also undergo phosphorylation on serine/threonine residues, resulting in functional consequences [11]. For instance, ligand-independent serine phosphorylation of EphA2 stimulates migration, invasion, and cancer progression, particularly when ligand-induced forward signaling is decreased [11]. Phosphorylation of serine/threonine residues in Eph receptors can trigger ligand-independent signalings, as exemplified by S897-phosphorylated EphA2, which activates AKT and PI3K, promoting the migration and invasion of cancer cells. A mutation at the S897 site abolishes the ligand-independent promotion of cell motility [11, 106]. Ligand-independent EphB1 signaling promotes lung cancer cell invasion and migration through CDH2-induced EMT [41]. Interestingly, tyrosine phosphorylation of Eph receptors induced by ligands can lead to dephosphorylation of S897, and multiple studies have confirmed the pro-tumorigenic roles of ligand-independent Eph receptors [64].
Cross-talk with other signaling pathways
Eph receptors can also impact other signaling pathways by physically interacting with other RTKs [50]. This crosstalk between Eph receptors and other RTKs, such as ErbB2 (HER-2) and EGFR, leads to the phosphorylation of Eph receptor tyrosine kinase, subsequently increasing the activation Rho GTPases and Ras/ERK signaling pathways [5, 51] (Figs. 2F and 3B). This interplay between Eph receptors and other RTKs, possibly in an ephrin-independent manner, promotes pro-oncogenic effects, including enhanced cancer cell motility and proliferation [50].
In summary, when ligand ephrins are present, Eph receptor forward signaling and reverse signaling counteract pro-oncogenic effects. However, in the absence of ligand ephrins, Eph receptor ligand-independent signaling promotes the pro-oncogenic effects. The physical interactions between ephrin and Eph receptors within the same cell, as well as the interaction of Eph receptor with other RTKs, contribute to processes such as proliferation, differentiation, adhesion and migration [107].
Role of Eph and ephrin in vascular system
Tumor angiogenesis plays a crucial role in promoting cancer cell growth and metastasis. The human vascular system comprises a complex network of blood vessels, including arteries, veins, and capillaries, acting as the primary conduit for information exchange and metabolic substance transport [108]. This intricate vascular network facilitates the absorption of essential nutrients and the removal of cellular and metabolic waste products [109]. During early embryonic development, the vascular network emerges through vasculogenesis and angiogenesis. In adulthood, under physiologic conditions, blood vessels generally remain quiescent, with limited new branch formation [110].
There are three main mechanisms of vessel formation. First, vasculogenesis involves the de novo generation new blood vessels from endothelial progenitor cells [110]. Initially thought to be restricted to embryonic development, recent studies have reported instances of vasculogenesis occurring in adult tissues [111]. Second, sprouting angiogenesis is the process of forming new blood vessels from preexisting ones and is observed during wound healing, ovulation, and embryo development [112]. This highly invasive process relies on the regulation of endothelial cell proliferation and migration by proangiogenic and anti-angiogenic molecules [113]. Third, intussusceptive angiogenesis is characterized by the formation of intraluminal tissue pillars within existing capillaries, small arteries, and veins, resulting in the splitting of vessels into two daughter vessels through the development of a central perforation and subsequent fusion of multiple capillaries [114].
The role of angiogenesis in cancer progression has gained considerable attention, as it sustains tumor growth, invasion, and metastasis. Tumors often stimulate the development of new blood vessels by secreting pro-angiogenic factors like vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF), while anti-angiogenic factors such as thrombospondin-1, angiostatin, and endostatin act as negative regulators of angiogenesis.
Eph receptor tyrosine kinases play pivotal roles in angiogenic processes, particularly in directing angiogenic sprouting and intussusceptive angiogenesis (Table 3). Several B-class ephrins and their receptors, including EphB2, EphB3, EphB4, ephrin-B1 and ephrinB2, are expressed in the murine embryonic vasculature [44]. Ephrin-B2, for example, is predominantly expressed in arteries and absent from veins [115], suggesting its role in defining boundaries between veins and arteries and influencing the location of capillary beds [44]. EphB2 and ephrinB2 are often located in the mesenchyme surrounding vessels, indicating their involvement in mesenchymal-endothelial interactions [44]. Genetic studies in mice have shown that targeted mutagenesis of ephrinB2 results in significant disruptions in normal vascular development during embryogenesis [116]. EphA2 activation induces a PI3-kinase-dependent signaling pathway that leads to the activation of Rac1 in endothelial cells in response to ephrin-A1. Analyzing single-cell sequencing data of colorectal cancer revealed endothelial cells as a prominent cluster, expressing the EphB1 gene significantly (Fig. 4). Moreover, similar analyses across 13 different tumors demonstrated substantial EphB1 expression in endothelial cells (Fig. 5A). EphA2 and EphB4, the most studied Eph molecules, were also found to be highly expressed in endothelial cells based on single-cell sequencing data (Fig. 5B and C). This signaling cascade orchestrated by Eph receptors contributes to vascular assembly and cell migration, further highlighting the crucial role in angiogenesis [117].

The expression of EphB1 in various cell types. The expression of EphB1 in various cell types was analyzed in publicly accessible single-cell RNA-seq (scRNA seq) dataset of colorectal cancer tissues (EMTAB8107). A UMAP plot with cell type annotations displayed at the top of the tab. B UMAP plot shows the expression level of EphB1 in different cell types. C Pie plot shows the cell number distribution of each cell type. D Violin plot reflects the distribution of EphB1 gene expression in various cell types

Average expression of Eph receptors. The TISCH2 database (http://tisch.compgenomics.org/), containing scRNA-seq data from more than 50 cancer types, was utilized to analyze the expression of Eph receptors. A The expression of EphA2 across datasets in different cell types. B The expression of EphB1 across datasets in different cell types. C The expression of EphB4 across datasets in different cell types
Role of Eph and ephrin in sprouting angiogenesis
Sprouting angiogenesis plays a fundamental role in the development of new blood vessels in cancer. This process involves the emergence of new capillaries or blood vessels branching out from existing ones, premarily driven by signals such as Vascular Endothelial Growth Factor (VEGF). In this intricate process, Eph receptors and their corresponding ephrin ligands play crucial roles as regulators, influencing various aspects of angiogenic sprouting, including endothelial cell migration, adhesion, proliferation, and vessel branching.
It’s important to note that Eph/ephrin signaling doesn’t directly induce cellular proliferation but rather exerts its effects through the involvement of other growth factors [118]. For instance, EphB/ephrinB signaling triggers Akt, PI3K, and MAPK signaling pathways, promoting endothelial cell proliferation and migration [29]. EphA2/ephrinA1 signaling regulates endothelail cell assembly and migration by PI3K-mediated activation of Rac1GTPase [119]. Inhibition of EphA receptors using soluble EphA-Fc or EphA3-Fc has been shown to effectively hinder tumor angiogenesis [120]. The role of Src as a key mediator in specific angiogenic processes has been highlighted [121]. It has been found that c-Src is an important target of EphB1. Ligand-stimulated EphB1 recruits c-Src and induces its active conformation through the phosphorylation of tyrosine Y418 [122].
Among the extensively studied Eph-ephrin pairs, EphB4 and ephrinB2 not only promotes the sprouting of new blood vessels but also play a crucial role in vessel maturation. EphB4 is primarily expressed in venous endothelial cells, while ephrinB2 is predominantly found in arteries. Their interaction facilitates endothelial cell migration, sprouting, and the assembly of new vessels. Additionally, interaction between endothelial ephrin-B2 and monocytes EphB4 plays a regulatory role in monocyte extravasation through the vascular endothelium [123, 124]. The analogous process of cancer cell extravasation driven by ephrinB2-EphB4 is yet to be determined [61].
Role of EphB and EphrinB2 in non-sprouting intussusceptive angiogenesis
Intussusceptive angiogenesis (IA) plays a significant role in both pre- and postnatal vascular growth,remodeling, tissue repair, and cancer progression [125]. Unlike sprouting angiogenesis, which relies on endothelial cell proliferation and migration, IA is a non-sprouting mechanism involving the division or splitting of pre-existing blood vessels. This distinctive process facilitates the expansion of the capillary bed and the development of intricate vascular networks while maintaining the architectural integrity of the tissue [125]. IA is a rapid process that does not require endothelial cell proliferation.
The mechanisms governing IA involve a complex interplay of hemodynamic forces, endothelial cell behavior, and extracellular matrix remodeling. Elevated fluid shear stress and mechanical forces within blood vessels are thought to initiate the formation of transvascular tissue pillars, leading to the division of the vessel lumen into multiple channels. Subsequently, these tissue pillars undergo growth and fusion, giving rise to new vessel segments and expanding the vascular network.
Several factors that are involved in sprouting angiogenesis, such as PDGF-B, MCP-1 and Eph/ephrins, are potential candidates for mediating IA [125]. Eph/ephrin signaling pathways, in particular, play a crucial role in regulating the positioning and segregation of arterial and venous endothelial cells during angiogenesis [116].
EphB4/ephrinB2 regulates IA in response to VEGF [126]. EphB4 in endothelial cells finely tunes the extent of endothelial proliferation induced by VEGF, thereby controlling the initial vascular enlargement without directly affecting VEGF-R2 activation [126]. Instead, it modulates downstream signaling through MAPK/ERK. Blocking ephrinB2/EphB4 signaling, without affecting TGF-β1/TGF-βR or angiopoietin/Tie2 pathways, can shift VEGF-induced angiogenesis from normal to aberrant. Conversly, activation of EphB4 signaling prevents the occurrence of aberrant angiogenesis induced by high VEGF doses.
The role of the Eph family in immune modulation and inflammation
Eph receptors play a multifaceted role in immune cell differentiation and activation (Table 3). These receptors are expressed widely in immune cells, such as monocytes, macrophages, DCs, B cells and T cells. They contribute to diverse functions within the immune system, encompassing immune cell activation, migration, adhesion, differentiation and proliferation, often modulated by interactions with the microenvironment [127].
Immune cell differentiation
Eph receptors play roles on hematopoietic cells during their differentiation into distinct immune cell fates [127]. The cell fate decisions are critically dependent on the interaction of the hematopoietic stem cells with their microenvironmental components [127]. The interplay between EphB4-positive human bone marrow hematopoietc progenitor cells and ephrin-B2-positive stromal cells significantly influences the differentiation of progenitor cells into mature erythroid cells [128]. Additionally, the interaction between EphB4 and ephrin-B2 governs the mobilization of hematopoietic stem and progenitor cells from the bone marrow to the circulation [129]. In cellular immunity, T cells play a crucial role and can be stimulated by ephrin-B1 and ephrin-B2, which regulate thymocyte development [130]. Deletion of ephrin-B1 and/or ephrin-B2 in thymocytes or thymic epithelial cells (TECs) results in decreased medullary areas and enlarged cysts [131].
Immune cell activation
Immune cell activation is a critical mechanism employed by the body to defend itself against infections and cancers. This intricate process involves antigen recognition, antigen presentation, activation signals, co-stimulation, and eventual immune cell activation. Upon activation, immune cells initiate a series of events, including the production of cytokines, chemokines, and antibodies, along with increased cell proliferation and recruitment to the site of infection, inflammation and cancer. While the immune system’s role in recognizing and eliminating cancer cells is pivotal, cancer cells can employ strategies to evade immune surveillance and suppression. The regulatory influence of Eph receptors extends to immune cell activation, influencing their capacity to detect and target tumor cells. This regulatory role can lead to either heightened or diminished anti-tumor immune responses, thereby adding complexity to the relationship between immune cell activation and cancer.
EphA3 is frequently overexpressed in various types of tumors, such as melanoma, lung carcinoma, and sarcoma, where it functions as a tumor-specific antigen [132]. Despite its oncogenic roles in gastric carcinoma and glioblastoma multiforme [133, 134], EphA3 may function as a tumor-suppressor in lung adenocarcinomas through suppressing ATK activation [86]. EphA3 is also the most highly mutated Eph receptor, and many EphA3 mutations identified in cancers impair kinase activity. This suggests that wild-type EphA3 may inhibit cancer formation or progression, and somatic cancer mutations disrupt these tumor-suppressive activities [86]. EphA3 mutations are particularly frequent in lung cancer and can act as “drivers” in lung cancer [135]. Dendritic cells (DCs) capture, process, and present antigens to other immune cells, such as T cells and B cells. DCs loaded with EphA2 peptide have been found to induce immune responses and reduce tumor burden [136, 137]. In T cells, EphB receptors, including EphB1, EphB2,EphB3 and EphB6, are expressed [127]. In vitro assays have shown that stimulation by ephrin B ligands can activate T cells, but high concentration of ephrinB1 and ephrinB2 can inhibit T-cell activation, suggesting that EphB-ephrinB signaling may also provide negative feedback during T cell activation [127, 138,139,140]. In a rat glioma model, ephrin-A1 can activate DCs [141]. EphB1 is expressed in plasmacytoid DCs, CD4+ and CD8+ T cells [127, 142]. EphB6, primarily expressed in thymocytes and a subpopulation of T cells, plays a crucial role in T cell activation and cancer cell death. The absence of EphB6 in mice reduces T cell activation and the recruitment of crucial signaling molecules like ZAP-70, LAT, SLP-76, PLCγ1, and P44/42 MAPK [143]. The compensatory functions within the ephrin family are evident, as the knockout of ephrin-B1 or ephrin-B2 in mice does not affect the activation and proliferation of T cells and native CD4 cells [130]. Notably, in human bone marrow-derived mesenchymal stromal cells, ephrinB2 binding to EphB4 on T cells inhibits T cell proliferation by inducing the production of immunosuppressive factors and reducing activating cytokines [140]. These findings highlight the regulatory roles of Eph receptors and ephrins in immune cell activation and their potential implications in anti-tumor immune responses.
The peripheral B cell pool becomes populated with antigen-experienced memory cells, which is similar to the T pool. The activation and antibody production of these B cells require interactions with other immune cells. B cells are primarily responsible for orchestrating humoral immunity [144]. The expression levels of Eph receptors and ephrin ligands vary between activated and non-activated B cells [145]. This variation suggests that they may play a role in processes facilitating B cell activation. Research has shown that prior to activation, the upregulation of EphB2 leads to increased proliferation and antibody production in human B cells [127]. The differential expressions of Eph receptors and ephrin ligands in B cells not only regulate their development, activation, differentiation, and functionality but also potentially contribute to their specialize into various canonical B cell fates [127].
Inflammation and inflammatory pain
Eph receptors have gained increasing recognition for their significant roles in inflammatory diseases such as rheumatoid arthritis and bone cancer pain. They play a crucial role in regulating the migration and adhesion of leukocyte, contributing to the regulation of inflammatory signaling pathways. Eph receptors and ephrins also participate in tissue repair and remodeling following inflammation.
EphB1, for example, has been identified as beneficial to the activation of STAT3 transcripts and networks, suggesting a shift from proinflammatory activation towards an immune-modulatory or anti-inflammatory arm of the STAT3 pathway [146]. EphrinB1 and EphB1 exert significant influence on T cell function, playing crucial roles in inflammatory conditions like rheumatoid arthritis [147]. EphrinB1 can activate EphB1, leading to the production of TNF-α in peripheral blood lymphocytes and IL-6 in synovial cells [147]. Metastatic cancer-induced bone pain (CIBP) is a complex chronic condition [148]. Inflammation contributes to CIBP, and anti-inflammatory drugs such as nonsteroidal anti-inflammatory drugs (NSAIDs) are commonly used for treatment [149]. EphrinB-EphB receptor signaling, significantly upregulated in the dorsal root ganglion and spinal cord, has been implicated in the development of bone cancer pain [150]. Studies demonstrate that intrathecal injection of ephrinB2-Fc induces thermal hyperalgesia, mechanical allodynia, and activates spinal PKA and CREB. Inhibition of PKA can prevent and reverse thermal hyperalgesia and mechanical allodynia caused by ephrinB2-fc [151]. The involvement of EphB1 forward signaling in the spinal cord has been linked to the development of bone cancer pain and morphine tolerance. Agents designed to block the EphB1 receptor, such as EphB2-Fc, have demonstrated the potential to prevent and reverses bone cancer-induced pain [150, 152]. EphB1-Fc not only inhibits the activation of spinal MAPKs induced by inflammatory and neuropathic pain but also potentially hinders the formation of long-term potentiation (LTP) at synapses between dorsal root gnaglia and dorsal horn [153]. Furthermore, EphrinB-EphB receptor signaling activates astrocytes and microglial cells in the spinal cord by either activating or interacting with TLR4. This activation results in increased activity of proinflammatory cytokines like IL-1B and TNF-ɑ, contributing to bone cancer pain [154]. In another aspects, EphA4 receptor is involved in the generation and maintenance of CFA-induced chronic inflammatory pain, and blocking the spinal EphA4 receptor could potentially relieve persistent pain behaviors in mice [155].
Therapeutic implications
The intricate nature of Eph signaling presents a challenge for targeted cancer treatment. Various strategies exist to either enhance or inhibit the activities of individual Eph receptor or multiple family members. Despite the potential of these strategies, there are currently a lack of approved drugs specifically targeting the Eph family for cancer therapy.
Monoclonal antibody
Monoclonal antibody drugs represent a targeted therapy utilizing antibodies to specifically bind to certain proteins on the cell surface. This approach offers several advantages for the development of Eph-targeted therapeutics: (1) Monoclonal antibody drugs can be precisely designed to target specific proteins or cells, minimizing the risk of damage to healthy cells and tissues; (2) Monoclonal antibody drugs function as agonists for receptors by inducing receptor clustering, leading to activation. They can also act as antagonists by reducing receptor levels through processes like receptor endocytosis and degradation; (3) Monoclonal antibody drugs can inhibit receptor activity by blocking ligand binding, disrupting downstream signaling pathways; (4) Monoclonal antibody drugs can exert cytotoxic effects directly or indirectly through mechanisms such as antibody-dependent cell-mediated cytotoxicity (ADCC) or complement-dependent cytotoxicity (CDC) (Fig. 6); (5) Monoclonal antibody drugs can be loaded with various payloads, including radioactive isotype, conjugated drugs, or durg-containing nanoparticles [156]. For instance, anti-EphA2 mAbs have developed as agonistic antibodies, effectively inhibiting tumor growth and angiogenesis by inducing receptor phosphorylation and subsequent anticancer effects [157]. Humanized EphA2 mAb DS-8895a has demonstrated effecacy in inhibiting the growth of breast and gastric xenograft models through ADCC [158]. Additionally, EphA3, often overexpressed in hematologic malignancies, has been targeted by the humaneered derivative of the IIIA4 anti-EphA3 mAb (KB004) in clinical trials for hematological malignancies. KB004 exhibits a direct and antibody-mediated antileukemic effect on EphA3-positive human pre-B-ALL xenografts, specifically targeting leukemic cells, and its payload with an isotope is highly effective for antileukemia therapy [159].

Therapeutic strategies targeting Eph receptors. These strategies can inhibit tumor progression through various approaches and also impact pain behaviors
Among the EphBs, EphB4 has emerged as a prominent target for antibodies [156]. Disrupting the binding between ephrinB2 and EphB4 represents a novel strategy for anti-angiogenic therapy. Currently, two antibodies, including mAb47 and mAb131, have been developed to target the fibronectin domain of EphB4. Both mAbs have demonstrated the ability to reduce blood vessel density. Specifically, monoclonal antibody mAb47 has shown efficacy in suppressing tumor angiogenesis and inhibiting the growth of both EphB4-positive and EphB4-negative tumors, suggesting its impact on the tumor microenvironment [160, 161].
Targeting serine phosphorylation of Eph receptors
Targeting tyrosine phosphorylation of Eph receptors
Targeting tyrosine phosphorylation of Eph receptors involves strategies aimed at inhibiting or modulating the activity of these cell surface receptors, particularly RTKs. Targeting RTKs can be achieved through different approaches, such as small molecule inhibitors or monoclonal antibodies. Small molecule inhibitors typically bind to the ATP-binding site of the kinase domain, thereby preventing the phosphorylation of the receptor. An example is NVP-BHG712, which serves as an EphB4 kinase inhibitor. Treatment with NVP-BHG712 leads to a dose-dependent inhibition of RTK phosphorylation. It also shows activity against EphA2, VEGFR2, c-Raf, c-Src and c-Abl. By inhibiting VEGFR, NVP-BHG712 disrupts the growth and survival of cancer cells, leading to tumor regression. NVP-BHG712 exhibits excellent pharmacokinetic properties, inhibits EphB4 autophosphorylation after oral administration, and effectively suppresses VEGF-driven vessel formation (Fig. 6). The data suggests a close relationship between VEGFR and Eph receptors signaling during vessel formation, and EphB4 forward signaling is a significant mediator of VEGF-induced angiogenesis [162]. Despite its potential as an anti-cancer agent, further research is needed to fully understand its mechanisms of action and determine its optimal use in different cancer types and patient populations.
Interestingly, EphB1 receptor forward signaling is critical to the development and maintenance of pain, involving the activation of NR1 and NR2B receptors and subsequent Ca2+ -dependent signals. Inhibition of EphB1 prevents pain behaviors and NMDAR activation, even in the presence of active ephrinB2. Conversely, activation of EphB1 receptor induces pain behaviors and NMDAR activation, even with down-regulated ephrinB2 [150] (Fig. 6). The activated Eph receptors can also facilitate other protein–protein interactions via SAM and PDZ-binding motifs, contributing to signaling [163]. Investigating the roles of Eph receptors in neuropathic pain, researchers have identified tetracycline combinations that exhibit inhibitory activity against EphB1 tyrosine kinase through in silico screen in FDA-approved drugs. These combinations hold the potential for modulating the EphB1 pathway, which is relevant to neuropathic pain [164].
siRNA
Small interfering RNAs (siRNAs) approach serves as a valuable method to target the overexpression of mRNA. Inhibiting Eph receptors through siRNAs effectively disrupts Eph forward signaling, reverse signaling and even non-canonical signaling pathway [165]. Downregulation of EphA2 or EphB4 using small interfering RNAs (siRNAs) or antisense oligonucleotides has demonstrated significant impact on cancer cell malignancy in culture and has shown efficacy in inhibiting tumor growth in various mouse cancer models [61]. The knockdown of EphA2 in cancer cells in vitro has exhibited significant effects in glioma, NSCLC and breast cancer cells [166]. Furthermore, in vivo delivery of EphA2 siRNA using nanoliposomal in ovarian cancer has demonstrated effective targeting of cancer cell and remarkable anticancer effects [167, 168]. The knockdown of EphB4 in breast cancer cells has been shown to inhibit integrin-mediated cell adhesion, spreading and migration. However these effects are independent of ephrin stimulation [51].
Conclusions and perspectives
Ephs and Ephrins have been the focal points of extensive research over several decades, illuminating their classification, structure, bidirectional signaling mechanisms. The comprehensive exploration of Eph receptors have unraveled their diverse roles across various biological processes, from embryonic development to cancer progression, offering valuable insights into the intricate landscape of cellular signaling networks.
Eph receptors exhibit context-dependent and sometimes opposing effects, playing dual roles as both promoters and inhibitors of cancer progression. The delicate equilibrium among signaling pathways, angiogenesis, and immune responses is crucial for maintaining tissue homeostasis. Dysregulation of this balance is implicated in various pathological conditions, especially cancer. The discrepancies observed in the roles of Eph receptors in cancer underscore the molecular mechanisms involved in ligand-dependent and ligand-independent signaling, as well as their influence on angiogenesis and immune response modulation. Despite substantial research, several key issues remain to be addressed. Clarifying the crosstalk between Eph receptors and other signaling pathways are essential for understanding the molecular mechanisms. Further research holds the potential to uncover novel therapeutic targets and strategies, especially when considering the diverse context in which Eph receptors operate. This exploration provides valuable insights, revolutionizing the approach to cancer therapy.
Availability of data and materials
No datasets were generated or analysed during the current study.
References
Gucciardo E, Sugiyama N, Lehti K. Eph- and ephrin-dependent mechanisms in tumor and stem cell dynamics. Cell Mol Life Sci. 2014;71:3685–710. https://doi.org/10.1007/s00018-014-1633-0.
Unified nomenclature for Eph family receptors and their ligands, the ephrins. Eph Nomenclature Committee. Cell. 1997;90:403–4. https://doi.org/10.1016/s0092-8674(00)80500-0.
Nakada M, Hayashi Y, Hamada J. Role of Eph/ephrin tyrosine kinase in malignant glioma. Neuro Oncol. 2011;13:1163–70. https://doi.org/10.1093/neuonc/nor102.
Lisabeth EM, Falivelli G, Pasquale EB. Eph receptor signaling and ephrins. Cold Spring Harb Perspect Biol. 2013;5. https://doi.org/10.1101/cshperspect.a009159.
Brantley-Sieders DM, Zhuang G, Hicks D, Fang WB, Hwang Y, Cates JM, Coffman K, Jackson D, Bruckheimer E, Muraoka-Cook RS, Chen J. The receptor tyrosine kinase EphA2 promotes mammary adenocarcinoma tumorigenesis and metastatic progression in mice by amplifying ErbB2 signaling. J Clin Invest. 2008;118:64–78. https://doi.org/10.1172/JCI33154.
Arvanitis D, Davy A. Eph/ephrin signaling: networks. Genes Dev. 2008;22:416–29. https://doi.org/10.1101/gad.1630408.
Anderton M, van der Meulen E, Blumenthal MJ, Schafer G. The Role of the Eph Receptor Family in Tumorigenesis. Cancers (Basel). 2021;13. https://doi.org/10.3390/cancers13020206.
Wei W, Wang H, Ji S. Paradoxes of the EphB1 receptor in malignant brain tumors. Cancer Cell Int. 2017;17:21. https://doi.org/10.1186/s12935-017-0384-z.
Kandouz M. The Eph/Ephrin family in cancer metastasis: communication at the service of invasion. Cancer Metastasis Rev. 2012;31:353–73. https://doi.org/10.1007/s10555-012-9352-1.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–74. https://doi.org/10.1016/j.cell.2011.02.013.
Miao H, Li DQ, Mukherjee A, Guo H, Petty A, Cutter J, Basilion JP, Sedor J, Wu J, Danielpour D, et al. EphA2 mediates ligand-dependent inhibition and ligand-independent promotion of cell migration and invasion via a reciprocal regulatory loop with Akt. Cancer Cell. 2009;16:9–20. https://doi.org/10.1016/j.ccr.2009.04.009.
Lee HH, Wang YN, Yang WH, Xia W, Wei Y, Chan LC, Wang YH, Jiang Z, Xu S, Yao J, et al. Human ribonuclease 1 serves as a secretory ligand of ephrin A4 receptor and induces breast tumor initiation. Nat Commun. 2021;12:2788. https://doi.org/10.1038/s41467-021-23075-2.
Noren NK, Foos G, Hauser CA, Pasquale EB. The EphB4 receptor suppresses breast cancer cell tumorigenicity through an Abl-Crk pathway. Nat Cell Biol. 2006;8:815–25. https://doi.org/10.1038/ncb1438.
Liang Z, Wang X, Dong K, Li X, Qin C, Zhou H. Expression Pattern and Prognostic Value of EPHA/EFNA in Breast Cancer by Bioinformatics Analysis: Revealing Its Importance in Chemotherapy. Biomed Res Int. 2021;2021:5575704. https://doi.org/10.1155/2021/5575704.
Nikolova Z, Djonov V, Zuercher G, Andres AC, Ziemiecki A. Cell-type specific and estrogen dependent expression of the receptor tyrosine kinase EphB4 and its ligand ephrin-B2 during mammary gland morphogenesis. J Cell Sci. 1998;111(Pt 18):2741–51. https://doi.org/10.1242/jcs.111.18.2741.
Chen G, Wang Y, Zhou M, Shi H, Yu Z, Zhu Y, Yu F. EphA1 receptor silencing by small interfering RNA has antiangiogenic and antitumor efficacy in hepatocellular carcinoma. Oncol Rep. 2010;23:563–70.
Surawska H, Ma PC, Salgia R. The role of ephrins and Eph receptors in cancer. Cytokine Growth Factor Rev. 2004;15:419–33. https://doi.org/10.1016/j.cytogfr.2004.09.002.
Wang X, Zhang M, Ping F, Liu H, Sun J, Wang Y, Shen A, Ding J, Geng M. Identification and Therapeutic Intervention of Coactivated Anaplastic Lymphoma Kinase, Fibroblast Growth Factor Receptor 2, and Ephrin Type-A Receptor 5 Kinases in Hepatocellular Carcinoma. Hepatology. 2019;69:573–86. https://doi.org/10.1002/hep.29792.
Feng YX, Zhao JS, Li JJ, Wang T, Cheng SQ, Yuan Y, Wang F, Wang XF, Xie D. Liver cancer: EphrinA2 promotes tumorigenicity through Rac1/Akt/NF-kappaB signaling pathway. Hepatology. 2010;51:535–44. https://doi.org/10.1002/hep.23313.
Husain A, Chiu YT, Sze KM, Ho DW, Tsui YM, Suarez EMS, Zhang VX, Chan LK, Lee E, Lee JM, et al. Ephrin-A3/EphA2 axis regulates cellular metabolic plasticity to enhance cancer stemness in hypoxic hepatocellular carcinoma. J Hepatol. 2022;77:383–96. https://doi.org/10.1016/j.jhep.2022.02.018.
Yuan W, Zhao H, Zhou A, Wang S. Interference of EFNA4 suppresses cell proliferation, invasion and angiogenesis in hepatocellular carcinoma by downregulating PYGO2. Cancer Biol Ther. 2022;23:1–12. https://doi.org/10.1080/15384047.2022.2149039.
Sawai Y, Tamura S, Fukui K, Ito N, Imanaka K, Saeki A, Sakuda S, Kiso S, Matsuzawa Y. Expression of ephrin-B1 in hepatocellular carcinoma: possible involvement in neovascularization. J Hepatol. 2003;39:991–6. https://doi.org/10.1016/s0168-8278(03)00498-7.
Jamshidi-Parsian A, Griffin RJ, Kore RA, Todorova VK, Makhoul I. Tumor-endothelial cell interaction in an experimental model of human hepatocellular carcinoma. Exp Cell Res. 2018;372:16–24. https://doi.org/10.1016/j.yexcr.2018.09.001.
Li S, Zhu Y, Ma C, Qiu Z, Zhang X, Kang Z, Wu Z, Wang H, Xu X, Zhang H, et al. Downregulation of EphA5 by promoter methylation in human prostate cancer. BMC Cancer. 2015;15:18. https://doi.org/10.1186/s12885-015-1025-3.
Li S, Ma Y, Xie C, Wu Z, Kang Z, Fang Z, Su B, Guan M. EphA6 promotes angiogenesis and prostate cancer metastasis and is associated with human prostate cancer progression. Oncotarget. 2015;6:22587–97. https://doi.org/10.18632/oncotarget.4088.
Mertens-Walker I, Fernandini BC, Maharaj MS, Rockstroh A, Nelson CC, Herington AC, Stephenson SA. The tumour-promoting receptor tyrosine kinase, EphB4, regulates expression of integrin-β8 in prostate cancer cells. BMC Cancer. 2015;15:164. https://doi.org/10.1186/s12885-015-1164-6.
Iwasaki K, Ninomiya R, Shin T, Nomura T, Kajiwara T, Hijiya N, Moriyama M, Mimata H, Hamada F. Chronic hypoxia-induced slug promotes invasive behavior of prostate cancer cells by activating expression of ephrin-B1. Cancer Sci. 2018;109:3159–70. https://doi.org/10.1111/cas.13754.
Tang L, Wang L, Yu T, Xu T, Zhang L, Shu Y. Knockdown of EphB3 inhibits cell proliferation partly through the AKT signaling pathway and represses epithelial-mesenchymal transition in esophageal squamous cell carcinoma. Transl Cancer Res. 2022;11:85–98. https://doi.org/10.21037/tcr-21-1567.
Steinle JJ, Meininger CJ, Forough R, Wu G, Wu MH, Granger HJ. Eph B4 receptor signaling mediates endothelial cell migration and proliferation via the phosphatidylinositol 3-kinase pathway. J Biol Chem. 2002;277:43830–5. https://doi.org/10.1074/jbc.M207221200.
Chu LY, Huang BL, Huang XC, Peng YH, Xie JJ, Xu YW. EFNA1 in gastrointestinal cancer: Expression, regulation and clinical significance. World J Gastrointest Oncol. 2022;14:973–88. https://doi.org/10.4251/wjgo.v14.i5.973.
Easty DJ, Hill SP, Hsu MY, Fallowfield ME, Florenes VA, Herlyn M, Bennett DC. Up-regulation of ephrin-A1 during melanoma progression. Int J Cancer. 1999;84:494–501. https://doi.org/10.1002/(sici)1097-0215(19991022)84:5%3c494::aid-ijc8%3e3.0.co;2-o.
Wang YC, Dai Y, Xu GL, Yu W, Quan RL, Zhao YJ. Association Between EphA1 and Tumor Microenvironment in Gastric Carcinoma and its Clinical Significance. Med Sci Monit. 2020;26:e923409. https://doi.org/10.12659/msm.923409.
Peng Q, Chen L, Wu W, Wang J, Zheng X, Chen Z, Jiang Q, Han J, Wei L, Wang L, et al. EPH receptor A2 governs a feedback loop that activates Wnt/β-catenin signaling in gastric cancer. Cell Death Dis. 2018;9:1146. https://doi.org/10.1038/s41419-018-1164-y.
Oki M, Yamamoto H, Taniguchi H, Adachi Y, Imai K, Shinomura Y. Overexpression of the receptor tyrosine kinase EphA4 in human gastric cancers. World J Gastroenterol. 2008;14:5650–6. https://doi.org/10.3748/wjg.14.5650.
Lee PC, Chen ST, Kuo TC, Lin TC, Lin MC, Huang J, Hung JS, Hsu CL, Juan HF, Lee PH, Huang MC. C1GALT1 is associated with poor survival and promotes soluble Ephrin A1-mediated cell migration through activation of EPHA2 in gastric cancer. Oncogene. 2020;39:2724–40. https://doi.org/10.1038/s41388-020-1178-7.
Tanaka M, Kamata R, Takigahira M, Yanagihara K, Sakai R. Phosphorylation of ephrin-B1 regulates dissemination of gastric scirrhous carcinoma. Am J Pathol. 2007;171:68–78. https://doi.org/10.2353/ajpath.2007.070033.
Zhou S, Wang L, Guo S, Zhang Z, Wang J. EphA4 protein promotes invasion in clear cell renal cell carcinomas. Int J Clin Exp Pathol. 2017;10:11737–42.
Kou CJ, Kandpal RP. Differential Expression Patterns of Eph Receptors and Ephrin Ligands in Human Cancers. Biomed Res Int. 2018;2018:7390104. https://doi.org/10.1155/2018/7390104.
Giaginis C, Tsoukalas N, Bournakis E, Alexandrou P, Kavantzas N, Patsouris E, Theocharis S. Ephrin (Eph) receptor A1, A4, A5 and A7 expression in human non-small cell lung carcinoma: associations with clinicopathological parameters, tumor proliferative capacity and patients’ survival. BMC Clin Pathol. 2014;14:8. https://doi.org/10.1186/1472-6890-14-8.
Li L, He Y, Liu D, Li L, Chen F, Ran J, Yang L, Zhang L. Prognostic values of EphB1/B2 and p-EphB1/B2 expression in non-small cell lung cancer. Int J Clin Exp Pathol. 2017;10:10092–101.
Wang L, Peng Q, Sai B, Zheng L, Xu J, Yin N, Feng X, Xiang J. Ligand-independent EphB1 signaling mediates TGF-β-activated CDH2 and promotes lung cancer cell invasion and migration. J Cancer. 2020;11:4123–31. https://doi.org/10.7150/jca.44576.
Kumar SR, Singh J, Xia G, Krasnoperov V, Hassanieh L, Ley EJ, Scehnet J, Kumar NG, Hawes D, Press MF, et al. Receptor tyrosine kinase EphB4 is a survival factor in breast cancer. Am J Pathol. 2006;169:279–93. https://doi.org/10.2353/ajpath.2006.050889.
Efazat G, Novak M, Kaminskyy VO, De Petris L, Kanter L, Juntti T, Bergman P, Zhivotovsky B, Lewensohn R, Hååg P, Viktorsson K. Ephrin B3 interacts with multiple EphA receptors and drives migration and invasion in non-small cell lung cancer. Oncotarget. 2016;7:60332–47. https://doi.org/10.18632/oncotarget.11219.
Nakamoto M, Bergemann AD. Diverse roles for the Eph family of receptor tyrosine kinases in carcinogenesis. Microsc Res Tech. 2002;59:58–67. https://doi.org/10.1002/jemt.10177.
Dodelet VC, Pasquale EB. Eph receptors and ephrin ligands: embryogenesis to tumorigenesis. Oncogene. 2000;19:5614–9. https://doi.org/10.1038/sj.onc.1203856.
Kouros-Mehr H, Werb Z. Candidate regulators of mammary branching morphogenesis identified by genome-wide transcript analysis. Dev Dyn. 2006;235:3404–12. https://doi.org/10.1002/dvdy.20978.
Ogawa K, Pasqualini R, Lindberg RA, Kain R, Freeman AL, Pasquale EB. The ephrin-A1 ligand and its receptor, EphA2, are expressed during tumor neovascularization. Oncogene. 2000;19:6043–52. https://doi.org/10.1038/sj.onc.1204004.
Duxbury MS, Ito H, Zinner MJ, Ashley SW, Whang EE. EphA2: a determinant of malignant cellular behavior and a potential therapeutic target in pancreatic adenocarcinoma. Oncogene. 2004;23:1448–56. https://doi.org/10.1038/sj.onc.1207247.
Steube KG, Meyer C, Habig S, Uphoff CC, Drexler HG. Expression of receptor tyrosine kinase HTK (hepatoma transmembrane kinase) and HTK ligand by human leukemia-lymphoma cell lines. Leuk Lymphoma. 1999;33:371–6. https://doi.org/10.3109/10428199909058438.
Vaught D, Brantley-Sieders DM, Chen J. Eph receptors in breast cancer: roles in tumor promotion and tumor suppression. Breast Cancer Res. 2008;10:217. https://doi.org/10.1186/bcr2207.
Larsen AB, Pedersen MW, Stockhausen MT, Grandal MV, van Deurs B, Poulsen HS. Activation of the EGFR gene target EphA2 inhibits epidermal growth factor-induced cancer cell motility. Mol Cancer Res. 2007;5:283–93. https://doi.org/10.1158/1541-7786.MCR-06-0321.
Lemmon MA, Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell. 2010;141:1117–34. https://doi.org/10.1016/j.cell.2010.06.011.
Sheng Z, Wang J, Dong Y, Ma H, Zhou H, Sugimura H, Lu G, Zhou X. EphB1 is underexpressed in poorly differentiated colorectal cancers. Pathobiology. 2008;75:274–80. https://doi.org/10.1159/000151707.
Zhou S, Wang L, Li G, Zhang Z, Wang J. Decreased expression of receptor tyrosine kinase of EphB1 protein in renal cell carcinomas. Int J Clin Exp Pathol. 2014;7:4254–60.
Wang H, Wen J, Wang H, Guo Q, Shi S, Shi Q, Zhou X, Liu Q, Lu G, Wang J. Loss of expression of EphB1 protein in serous carcinoma of ovary associated with metastasis and poor survival. Int J Clin Exp Pathol. 2014;7:313–21.
Teng L, Nakada M, Furuyama N, Sabit H, Furuta T, Hayashi Y, Takino T, Dong Y, Sato H, Sai Y, et al. Ligand-dependent EphB1 signaling suppresses glioma invasion and correlates with patient survival. Neuro Oncol. 2013;15:1710–20. https://doi.org/10.1093/neuonc/not128.
Macrae M, Neve RM, Rodriguez-Viciana P, Haqq C, Yeh J, Chen C, Gray JW, McCormick F. A conditional feedback loop regulates Ras activity through EphA2. Cancer Cell. 2005;8:111–8. https://doi.org/10.1016/j.ccr.2005.07.005.
Guo H, Miao H, Gerber L, Singh J, Denning MF, Gilliam AC, Wang B. Disruption of EphA2 receptor tyrosine kinase leads to increased susceptibility to carcinogenesis in mouse skin. Cancer Res. 2006;66:7050–8. https://doi.org/10.1158/0008-5472.Can-06-0004.
Tang XX, Zhao H, Robinson ME, Cnaan A, London W, Cohn SL, Cheung NK, Brodeur GM, Evans AE, Ikegaki N. Prognostic significance of EPHB6, EFNB2, and EFNB3 expressions in neuroblastoma. Med Pediatr Oncol. 2000;35:656–8. https://doi.org/10.1002/1096-911x(20001201)35:6%3c656::aid-mpo37%3e3.0.co;2-a.
Hafner C, Schmitz G, Meyer S, Bataille F, Hau P, Langmann T, Dietmaier W, Landthaler M, Vogt T. Differential gene expression of Eph receptors and ephrins in benign human tissues and cancers. Clin Chem. 2004;50:490–9. https://doi.org/10.1373/clinchem.2003.026849.
Pasquale EB. Eph receptors and ephrins in cancer: bidirectional signalling and beyond. Nat Rev Cancer. 2010;10:165–80. https://doi.org/10.1038/nrc2806.
Liang LY, Patel O, Janes PW, Murphy JM, Lucet IS. Eph receptor signalling: from catalytic to non-catalytic functions. Oncogene. 2019;38:6567–84. https://doi.org/10.1038/s41388-019-0931-2.
Nikolov DB, Xu K, Himanen JP. Homotypic receptor-receptor interactions regulating Eph signaling. Cell Adh Migr. 2014;8:360–5. https://doi.org/10.4161/19336918.2014.971684.
Taylor H, Campbell J, Nobes CD. Ephs and ephrins. Curr Biol. 2017;27:R90–5. https://doi.org/10.1016/j.cub.2017.01.003.
Himanen JP. Ectodomain structures of Eph receptors. Semin Cell Dev Biol. 2012;23:35–42. https://doi.org/10.1016/j.semcdb.2011.10.025.
Wu Z, Ashlin TG, Xu Q, Wilkinson DG. Role of forward and reverse signaling in Eph receptor and ephrin mediated cell segregation. Exp Cell Res. 2019;381:57–65. https://doi.org/10.1016/j.yexcr.2019.04.040.
Klein R. Bidirectional modulation of synaptic functions by Eph/ephrin signaling. Nat Neurosci. 2009;12:15–20. https://doi.org/10.1038/nn.2231.
Lim YS, McLaughlin T, Sung TC, Santiago A, Lee KF, O’Leary DD. p75(NTR) mediates ephrin-A reverse signaling required for axon repulsion and mapping. Neuron. 2008;59:746–58. https://doi.org/10.1016/j.neuron.2008.07.032.
Alazzouzi H, Davalos V, Kokko A, Domingo E, Woerner SM, Wilson AJ, Konrad L, Laiho P, Espin E, Armengol M, et al. Mechanisms of inactivation of the receptor tyrosine kinase EPHB2 in colorectal tumors. Cancer Res. 2005;65:10170–3. https://doi.org/10.1158/0008-5472.CAN-05-2580.
Huusko P, Ponciano-Jackson D, Wolf M, Kiefer JA, Azorsa DO, Tuzmen S, Weaver D, Robbins C, Moses T, Allinen M, et al. Nonsense-mediated decay microarray analysis identifies mutations of EPHB2 in human prostate cancer. Nat Genet. 2004;36:979–83. https://doi.org/10.1038/ng1408.
Davies H, Hunter C, Smith R, Stephens P, Greenman C, Bignell G, Teague J, Butler A, Edkins S, Stevens C, et al. Somatic mutations of the protein kinase gene family in human lung cancer. Cancer Res. 2005;65:7591–5. https://doi.org/10.1158/0008-5472.CAN-05-1855.
Faoro L, Singleton PA, Cervantes GM, Lennon FE, Choong NW, Kanteti R, Ferguson BD, Husain AN, Tretiakova MS, Ramnath N, et al. EphA2 mutation in lung squamous cell carcinoma promotes increased cell survival, cell invasion, focal adhesions, and mammalian target of rapamycin activation. J Biol Chem. 2010;285:18575–85. https://doi.org/10.1074/jbc.M109.075085.
London M, Gallo E. Critical role of EphA3 in cancer and current state of EphA3 drug therapeutics. Mol Biol Rep. 2020;47:5523–33. https://doi.org/10.1007/s11033-020-05571-8.
Light TP, Gomez-Soler M, Wang Z, Karl K, Zapata-Mercado E, Gehring MP, Lechtenberg BC, Pogorelov TV, Hristova K, Pasquale EB. A cancer mutation promotes EphA4 oligomerization and signaling by altering the conformation of the SAM domain. J Biol Chem. 2021;297:100876. https://doi.org/10.1016/j.jbc.2021.100876.
Chen Z, Chen J, Ren D, Zhang J, Yang Y, Zhang H, Mao B, Ma H. EPHA5 mutations predict survival after immunotherapy in lung adenocarcinoma. Aging (Albany NY). 2020;13:598–618. https://doi.org/10.18632/aging.202169.
Andersson E, Eldfors S, Edgren H, Ellonen P, Väkevä L, Ranki A, Mustjoki S. Novel TBL1XR1, EPHA7 and SLFN12 mutations in a Sezary syndrome patient discovered by whole exome sequencing. Exp Dermatol. 2014;23:366–8. https://doi.org/10.1111/exd.12405.
Yoo S, Shin J, Park S. EphA8-ephrinA5 signaling and clathrin-mediated endocytosis is regulated by Tiam-1, a Rac-specific guanine nucleotide exchange factor. Mol Cells. 2010;29:603–9. https://doi.org/10.1007/s10059-010-0075-2.
Truitt L, Freywald A. Dancing with the dead: Eph receptors and their kinase-null partners. Biochem Cell Biol. 2011;89:115–29. https://doi.org/10.1139/o10-145.
Schulz J, Mah N, Neuenschwander M, Kischka T, Ratei R, Schlag PM, Castaños-Vélez E, Fichtner I, Tunn PU, Denkert C, et al. Loss-of-function uORF mutations in human malignancies. Sci Rep. 2018;8:2395. https://doi.org/10.1038/s41598-018-19201-8.
Zhang H, Cui Z, Pan T, Hu H, He R, Yi M, Sun W, Gao R, Wang H, Ma X, et al. RNF186/EPHB2 Axis Is Essential in Regulating TNF Signaling for Colorectal Tumorigenesis in Colorectal Epithelial Cells. J Immunol. 2022;209:1796–805. https://doi.org/10.4049/jimmunol.2200229.
Li G, Ji XD, Gao H, Zhao JS, Xu JF, Sun ZJ, Deng YZ, Shi S, Feng YX, Zhu YQ, et al. EphB3 suppresses non-small-cell lung cancer metastasis via a PP2A/RACK1/Akt signalling complex. Nat Commun. 2012;3:667. https://doi.org/10.1038/ncomms1675.
Ferguson BD, Carol Tan YH, Kanteti RS, Liu R, Gayed MJ, Vokes EE, Ferguson MK, John Iafrate A, Gill PS, Salgia R. Novel EPHB4 Receptor Tyrosine Kinase Mutations and Kinomic Pathway Analysis in Lung Cancer. Sci Rep. 2015;5:10641. https://doi.org/10.1038/srep10641.
Yoon S, Choi JH, Kim SJ, Lee EJ, Shah M, Choi S, Woo HG. EPHB6 mutation induces cell adhesion-mediated paclitaxel resistance via EPHA2 and CDH11 expression. Exp Mol Med. 2019;51:1–12. https://doi.org/10.1038/s12276-019-0261-z.
Cortina C, Palomo-Ponce S, Iglesias M, Fernandez-Masip JL, Vivancos A, Whissell G, Huma M, Peiro N, Gallego L, Jonkheer S, et al. EphB-ephrin-B interactions suppress colorectal cancer progression by compartmentalizing tumor cells. Nat Genet. 2007;39:1376–83. https://doi.org/10.1038/ng.2007.11.
Peng J, Wang Q, Liu H, Ye M, Wu X, Guo L. EPHA3 regulates the multidrug resistance of small cell lung cancer via the PI3K/BMX/STAT3 signaling pathway. Tumour Biol. 2016;37:11959–71. https://doi.org/10.1007/s13277-016-5048-4.
Zhuang G, Song W, Amato K, Hwang Y, Lee K, Boothby M, Ye F, Guo Y, Shyr Y, Lin L, et al. Effects of cancer-associated EPHA3 mutations on lung cancer. J Natl Cancer Inst. 2012;104:1182–97. https://doi.org/10.1093/jnci/djs297.
Makinen T, Adams RH, Bailey J, Lu Q, Ziemiecki A, Alitalo K, Klein R, Wilkinson GA. PDZ interaction site in ephrinB2 is required for the remodeling of lymphatic vasculature. Genes Dev. 2005;19:397–410. https://doi.org/10.1101/gad.330105.
Nakada M, Anderson EM, Demuth T, Nakada S, Reavie LB, Drake KL, Hoelzinger DB, Berens ME. The phosphorylation of ephrin-B2 ligand promotes glioma cell migration and invasion. Int J Cancer. 2010;126:1155–65. https://doi.org/10.1002/ijc.24849.
Chen J, Song W, Amato K. Eph receptor tyrosine kinases in cancer stem cells. Cytokine Growth Factor Rev. 2015;26:1–6. https://doi.org/10.1016/j.cytogfr.2014.05.001.
Liu W, Yu C, Li J, Fang J. The Roles of EphB2 in Cancer. Front Cell Dev Biol. 2022;10:788587. https://doi.org/10.3389/fcell.2022.788587.
Kao TJ, Kania A. Ephrin-mediated cis-attenuation of Eph receptor signaling is essential for spinal motor axon guidance. Neuron. 2011;71:76–91. https://doi.org/10.1016/j.neuron.2011.05.031.
Yin Y, Yamashita Y, Noda H, Okafuji T, Go MJ, Tanaka H. EphA receptor tyrosine kinases interact with co-expressed ephrin-A ligands in cis. Neurosci Res. 2004;48:285–96. https://doi.org/10.1016/j.neures.2003.11.009.
Doucey MA, Scarpellino L, Zimmer J, Guillaume P, Luescher IF, Bron C, Held W. Cis association of Ly49A with MHC class I restricts natural killer cell inhibition. Nat Immunol. 2004;5:328–36. https://doi.org/10.1038/ni1043.
Masuda A, Nakamura A, Maeda T, Sakamoto Y, Takai T. Cis binding between inhibitory receptors and MHC class I can regulate mast cell activation. J Exp Med. 2007;204:907–20. https://doi.org/10.1084/jem.20060631.
Endo S, Sakamoto Y, Kobayashi E, Nakamura A, Takai T. Regulation of cytotoxic T lymphocyte triggering by PIR-B on dendritic cells. Proc Natl Acad Sci U S A. 2008;105:14515–20. https://doi.org/10.1073/pnas.0804571105.
Held W, Mariuzza RA. Cis-trans interactions of cell surface receptors: biological roles and structural basis. Cell Mol Life Sci. 2011;68:3469–78. https://doi.org/10.1007/s00018-011-0798-z.
Carvalho RF, Beutler M, Marler KJ, Knoll B, Becker-Barroso E, Heintzmann R, Ng T, Drescher U. Silencing of EphA3 through a cis interaction with ephrinA5. Nat Neurosci. 2006;9:322–30. https://doi.org/10.1038/nn1655.
Li R, Sun Y, Jiang A, Wu Y, Li C, Jin M, Yan H, Jin H. Knockdown of ephrin receptor A7 suppresses the proliferation and metastasis of A549 human lung cancer cells. Mol Med Rep. 2016;13:3190–6. https://doi.org/10.3892/mmr.2016.4904.
Bhatia S, Baig NA, Timofeeva O, Pasquale EB, Hirsch K, MacDonald TJ, Dritschilo A, Lee YC, Henkemeyer M, Rood B, et al. Knockdown of EphB1 receptor decreases medulloblastoma cell growth and migration and increases cellular radiosensitization. Oncotarget. 2015;6:8929–46. https://doi.org/10.18632/oncotarget.3369.
Kim Y, Ahmed S, Miller WT. Colorectal cancer-associated mutations impair EphB1 kinase function. J Biol Chem. 2023;105115. https://doi.org/10.1016/j.jbc.2023.105115.
Zantek ND, Azimi M, Fedor-Chaiken M, Wang B, Brackenbury R, Kinch MS. E-cadherin regulates the function of the EphA2 receptor tyrosine kinase. Cell Growth Differ. 1999;10:629–38.
Herath NI, Boyd AW. The role of Eph receptors and ephrin ligands in colorectal cancer. Int J Cancer. 2010;126:2003–11. https://doi.org/10.1002/ijc.25147.
Falivelli G, Lisabeth EM, Rubio de la Torre E, Perez-Tenorio G, Tosato G, Salvucci O, Pasquale EB. Attenuation of eph receptor kinase activation in cancer cells by coexpressed ephrin ligands. PLoS ONE. 2013;8:e81445. https://doi.org/10.1371/journal.pone.0081445.
Pasquale EB. Eph receptor signalling casts a wide net on cell behaviour. Nat Rev Mol Cell Biol. 2005;6:462–75. https://doi.org/10.1038/nrm1662.
Wang L, Peng Q, Xie Y, Yin N, Xu J, Chen A, Yi J, Shi W, Tang J, Xiang J. Cell-cell contact-driven EphB1 cis- and trans- signalings regulate cancer stem cells enrichment after chemotherapy. Cell Death Dis. 2022;13:980. https://doi.org/10.1038/s41419-022-05385-5.
Li JY, Xiao T, Yi HM, Yi H, Feng J, Zhu JF, Huang W, Lu SS, Zhou YH, Li XH, Xiao ZQ. S897 phosphorylation of EphA2 is indispensable for EphA2-dependent nasopharyngeal carcinoma cell invasion, metastasis and stem properties. Cancer Lett. 2019;444:162–74. https://doi.org/10.1016/j.canlet.2018.12.011.
Cecchini A, Cornelison DDW. Eph/Ephrin-Based Protein Complexes: The Importance of cis Interactions in Guiding Cellular Processes. Front Mol Biosci. 2021;8:809364. https://doi.org/10.3389/fmolb.2021.809364.
Pugsley MK, Tabrizchi R. The vascular system. An overview of structure and function. J Pharmacol Toxicol Methods. 2000;44:333–40. https://doi.org/10.1016/s1056-8719(00)00125-8.
Ma Q, Reiter RJ, Chen Y. Role of melatonin in controlling angiogenesis under physiological and pathological conditions. Angiogenesis. 2020;23:91–104. https://doi.org/10.1007/s10456-019-09689-7.
Viallard C, Larrivee B. Tumor angiogenesis and vascular normalization: alternative therapeutic targets. Angiogenesis. 2017;20:409–26. https://doi.org/10.1007/s10456-017-9562-9.
Ratajska A, Jankowska-Steifer E, Czarnowska E, Olkowski R, Gula G, Niderla-Bielinska J, Flaht-Zabost A, Jasinska A. Vasculogenesis and Its Cellular Therapeutic Applications. Cells Tissues Organs. 2017;203:141–52. https://doi.org/10.1159/000448551.
Bikfalvi A. History and conceptual developments in vascular biology and angiogenesis research: a personal view. Angiogenesis. 2017;20:463–78. https://doi.org/10.1007/s10456-017-9569-2.
Vandekeere S, Dewerchin M, Carmeliet P. Angiogenesis Revisited: An Overlooked Role of Endothelial Cell Metabolism in Vessel Sprouting. Microcirculation. 2015;22:509–17. https://doi.org/10.1111/micc.12229.
Burri PH, Djonov V. Intussusceptive angiogenesis–the alternative to capillary sprouting. Mol Aspects Med. 2002;23:S1-27. https://doi.org/10.1016/s0098-2997(02)00096-1.
Adams RH, Wilkinson GA, Weiss C, Diella F, Gale NW, Deutsch U, Risau W, Klein R. Roles of ephrinB ligands and EphB receptors in cardiovascular development: demarcation of arterial/venous domains, vascular morphogenesis, and sprouting angiogenesis. Genes Dev. 1999;13:295–306. https://doi.org/10.1101/gad.13.3.295.
Wang HU, Chen ZF, Anderson DJ. Molecular distinction and angiogenic interaction between embryonic arteries and veins revealed by ephrin-B2 and its receptor Eph-B4. Cell. 1998;93:741–53. https://doi.org/10.1016/s0092-8674(00)81436-1.
Brantley-Sieders DM, Caughron J, Hicks D, Pozzi A, Ruiz JC, Chen J. EphA2 receptor tyrosine kinase regulates endothelial cell migration and vascular assembly through phosphoinositide 3-kinase-mediated Rac1 GTPase activation. J Cell Sci. 2004;117:2037–49. https://doi.org/10.1242/jcs.01061.
Cheng N, Brantley DM, Chen J. The ephrins and Eph receptors in angiogenesis. Cytokine Growth Factor Rev. 2002;13:75–85. https://doi.org/10.1016/s1359-6101(01)00031-4.
Miao H, Wei BR, Peehl DM, Li Q, Alexandrou T, Schelling JR, Rhim JS, Sedor JR, Burnett E, Wang B. Activation of EphA receptor tyrosine kinase inhibits the Ras/MAPK pathway. Nat Cell Biol. 2001;3:527–30. https://doi.org/10.1038/35074604.
Brantley DM, Cheng N, Thompson EJ, Lin Q, Brekken RA, Thorpe PE, Muraoka RS, Cerretti DP, Pozzi A, Jackson D, et al. Soluble Eph A receptors inhibit tumor angiogenesis and progression in vivo. Oncogene. 2002;21:7011–26. https://doi.org/10.1038/sj.onc.1205679.
Eliceiri BP, Paul R, Schwartzberg PL, Hood JD, Leng J, Cheresh DA. Selective requirement for Src kinases during VEGF-induced angiogenesis and vascular permeability. Mol Cell. 1999;4:915–24. https://doi.org/10.1016/s1097-2765(00)80221-x.
Vindis CC, Cerretti DP, Daniel TO, Huynh-Do U. EphB1 recruits c-Src and p52Shc to activate MAPK/ERK and promote chemotaxis. J Cell Biol. 2003;162:661–71. https://doi.org/10.1083/jcb.200302073.
Pfaff D, Heroult M, Riedel M, Reiss Y, Kirmse R, Ludwig T, Korff T, Hecker M, Augustin HG. Involvement of endothelial ephrin-B2 in adhesion and transmigration of EphB-receptor-expressing monocytes. J Cell Sci. 2008;121:3842–50. https://doi.org/10.1242/jcs.030627.
Korff T, Braun J, Pfaff D, Augustin HG, Hecker M. Role of ephrinB2 expression in endothelial cells during arteriogenesis: impact on smooth muscle cell migration and monocyte recruitment. Blood. 2008;112:73–81. https://doi.org/10.1182/blood-2007-12-128835.
Burri PH, Hlushchuk R, Djonov V. Intussusceptive angiogenesis: its emergence, its characteristics, and its significance. Dev Dyn. 2004;231:474–88. https://doi.org/10.1002/dvdy.20184.
Groppa E, Brkic S, Uccelli A, Wirth G, Korpisalo-Pirinen P, Filippova M, Dasen B, Sacchi V, Muraro MG, Trani M, et al. EphrinB2/EphB4 signaling regulates non-sprouting angiogenesis by VEGF. EMBO Rep. 2018;19. https://doi.org/10.15252/embr.201745054.
Darling TK, Lamb TJ. Emerging Roles for Eph Receptors and Ephrin Ligands in Immunity. Front Immunol. 2019;10:1473. https://doi.org/10.3389/fimmu.2019.01473.
Suenobu S, Takakura N, Inada T, Yamada Y, Yuasa H, Zhang XQ, Sakano S, Oike Y, Suda T. A role of EphB4 receptor and its ligand, ephrin-B2, in erythropoiesis. Biochem Biophys Res Commun. 2002;293:1124–31. https://doi.org/10.1016/S0006-291X(02)00330-3.
Kwak H, Salvucci O, Weigert R, Martinez-Torrecuadrada JL, Henkemeyer M, Poulos MG, Butler JM, Tosato G. Sinusoidal ephrin receptor EPHB4 controls hematopoietic progenitor cell mobilization from bone marrow. J Clin Invest. 2016;126:4554–68. https://doi.org/10.1172/JCI87848.
Jin W, Qi S, Luo H. The effect of conditional EFNB1 deletion in the T cell compartment on T cell development and function. BMC Immunol. 2011;12:68. https://doi.org/10.1186/1471-2172-12-68.
Cejalvo T, Munoz JJ, Tobajas E, Alfaro D, García-Ceca J, Zapata A. Conditioned deletion of ephrinB1 and/or ephrinB2 in either thymocytes or thymic epithelial cells alters the organization of thymic medulla and favors the appearance of thymic epithelial cysts. Histochem Cell Biol. 2015;143:517–29. https://doi.org/10.1007/s00418-014-1296-9.
Chiari R, Hames G, Stroobant V, Texier C, Maillere B, Boon T, Coulie PG. Identification of a tumor-specific shared antigen derived from an Eph receptor and presented to CD4 T cells on HLA class II molecules. Cancer Res. 2000;60:4855–63.
Day BW, Stringer BW, Al-Ejeh F, Ting MJ, Wilson J, Ensbey KS, Jamieson PR, Bruce ZC, Lim YC, Offenhauser C, et al. EphA3 maintains tumorigenicity and is a therapeutic target in glioblastoma multiforme. Cancer Cell. 2013;23:238–48. https://doi.org/10.1016/j.ccr.2013.01.007.
Xi HQ, Wu XS, Wei B, Chen L. Aberrant expression of EphA3 in gastric carcinoma: correlation with tumor angiogenesis and survival. J Gastroenterol. 2012;47:785–94. https://doi.org/10.1007/s00535-012-0549-4.
Ding L, Getz G, Wheeler DA, Mardis ER, McLellan MD, Cibulskis K, Sougnez C, Greulich H, Muzny DM, Morgan MB, et al. Somatic mutations affect key pathways in lung adenocarcinoma. Nature. 2008;455:1069–75. https://doi.org/10.1038/nature07423.
Hatano M, Kuwashima N, Tatsumi T, Dusak JE, Nishimura F, Reilly KM, Storkus WJ, Okada H. Vaccination with EphA2-derived T cell-epitopes promotes immunity against both EphA2-expressing and EphA2-negative tumors. J Transl Med. 2004;2:40. https://doi.org/10.1186/1479-5876-2-40.
Yamaguchi S, Tatsumi T, Takehara T, Sakamori R, Uemura A, Mizushima T, Ohkawa K, Storkus WJ, Hayashi N. Immunotherapy of murine colon cancer using receptor tyrosine kinase EphA2-derived peptide-pulsed dendritic cell vaccines. Cancer. 2007;110:1469–77. https://doi.org/10.1002/cncr.22958.
Yu G, Mao J, Wu Y, Luo H, Wu J. Ephrin-B1 is critical in T-cell development. J Biol Chem. 2006;281:10222–9. https://doi.org/10.1074/jbc.M510320200.
Kawano H, Katayama Y, Minagawa K, Shimoyama M, Henkemeyer M, Matsui T. A novel feedback mechanism by Ephrin-B1/B2 in T-cell activation involves a concentration-dependent switch from costimulation to inhibition. Eur J Immunol. 2012;42:1562–72. https://doi.org/10.1002/eji.201142175.
Nguyen TM, Arthur A, Hayball JD, Gronthos S. EphB and Ephrin-B interactions mediate human mesenchymal stem cell suppression of activated T-cells. Stem Cells Dev. 2013;22:2751–64. https://doi.org/10.1089/scd.2012.0676.
Li M, Wang B, Wu Z, Shi X, Zhang J, Han S. Treatment of Dutch rat models of glioma using EphrinA1-PE38/GM-CSF chitosan nanoparticles by in situ activation of dendritic cells. Tumour Biol. 2015;36:7961–6. https://doi.org/10.1007/s13277-015-3486-z.
Rissoan MC, Duhen T, Bridon JM, Bendriss-Vermare N, Péronne C, de Saint Vis B, Brière F, Bates EE. Subtractive hybridization reveals the expression of immunoglobulin-like transcript 7, Eph-B1, granzyme B, and 3 novel transcripts in human plasmacytoid dendritic cells. Blood. 2002;100:3295–303. https://doi.org/10.1182/blood-2002-02-0638.
Luo H, Yu G, Tremblay J, Wu J. EphB6-null mutation results in compromised T cell function. J Clin Invest. 2004;114:1762–73. https://doi.org/10.1172/JCI21846.
Weiskopf D, Weinberger B, Grubeck-Loebenstein B. The aging of the immune system. Transpl Int. 2009;22:1041–50. https://doi.org/10.1111/j.1432-2277.2009.00927.x.
Alonso CL, Trinidad EM, de Garcillan B, Ballesteros M, Castellanos M, Cotillo I, Muñoz JJ, Zapata AG. Expression profile of Eph receptors and ephrin ligands in healthy human B lymphocytes and chronic lymphocytic leukemia B-cells. Leuk Res. 2009;33:395–406. https://doi.org/10.1016/j.leukres.2008.08.010.
Tyzack GE, Hall CE, Sibley CR, Cymes T, Forostyak S, Carlino G, Meyer IF, Schiavo G, Zhang SC, Gibbons GM, et al. A neuroprotective astrocyte state is induced by neuronal signal EphB1 but fails in ALS models. Nat Commun. 2017;8:1164. https://doi.org/10.1038/s41467-017-01283-z.
Kitamura T, Kabuyama Y, Kamataki A, Homma MK, Kobayashi H, Aota S, Kikuchi S, Homma Y. Enhancement of lymphocyte migration and cytokine production by ephrinB1 system in rheumatoid arthritis. Am J Physiol Cell Physiol. 2008;294:C189-196. https://doi.org/10.1152/ajpcell.00314.2007.
Zajaczkowska R, Kocot-Kepska M, Leppert W, Wordliczek J. Bone Pain in Cancer Patients: Mechanisms and Current Treatment. Int J Mol Sci. 2019;20. https://doi.org/10.3390/ijms20236047.
Colosia A, Njue A, Bajwa Z, Dragon E, Robinson RL, Sheffield KM, Thakkar S, Richiemer SH. The Burden of Metastatic Cancer-Induced Bone Pain: A Narrative Review. J Pain Res. 2022;15:3399–412. https://doi.org/10.2147/JPR.S371337.
Liu S, Liu WT, Liu YP, Dong HL, Henkemeyer M, Xiong LZ, Song XJ. Blocking EphB1 receptor forward signaling in spinal cord relieves bone cancer pain and rescues analgesic effect of morphine treatment in rodents. Cancer Res. 2011;71:4392–402. https://doi.org/10.1158/0008-5472.CAN-10-3870.
Zhou XL, Wang Y, Zhang CJ, Yu LN, Cao JL, Yan M. PKA is required for the modulation of spinal nociceptive information related to ephrinB-EphB signaling in mice. Neuroscience. 2015;284:546–54. https://doi.org/10.1016/j.neuroscience.2014.10.025.
Dong Y, Mao-Ying QL, Chen JW, Yang CJ, Wang YQ, Tan ZM. Involvement of EphB1 receptor/ephrinB1 ligand in bone cancer pain. Neurosci Lett. 2011;496:163–7. https://doi.org/10.1016/j.neulet.2011.04.008.
Ruan JP, Zhang HX, Lu XF, Liu YP, Cao JL. EphrinBs/EphBs signaling is involved in modulation of spinal nociceptive processing through a mitogen-activated protein kinases-dependent mechanism. Anesthesiology. 2010;112:1234–49. https://doi.org/10.1097/ALN.0b013e3181d3e0df.
Liu S, Liu YP, Song WB, Song XJ. EphrinB-EphB receptor signaling contributes to bone cancer pain via Toll-like receptor and proinflammatory cytokines in rat spinal cord. Pain. 2013;154:2823–35. https://doi.org/10.1016/j.pain.2013.08.017.
Wang Y, Wen C, Xie G, Jiang L. Blockade of Spinal EphA4 Reduces Chronic Inflammatory Pain in Mice. Neurol Res. 2021;43:528–34. https://doi.org/10.1080/01616412.2021.1884798.
Janes PW, Vail ME, Gan HK, Scott AM. Antibody Targeting of Eph Receptors in Cancer. Pharmaceuticals (Basel). 2020;13. https://doi.org/10.3390/ph13050088.
Landen CN Jr, Lu C, Han LY, Coffman KT, Bruckheimer E, Halder J, Mangala LS, Merritt WM, Lin YG, Gao C, et al. Efficacy and antivascular effects of EphA2 reduction with an agonistic antibody in ovarian cancer. J Natl Cancer Inst. 2006;98:1558–70. https://doi.org/10.1093/jnci/djj414.
Hasegawa J, Sue M, Yamato M, Ichikawa J, Ishida S, Shibutani T, Kitamura M, Wada T, Agatsuma T. Novel anti-EPHA2 antibody, DS-8895a for cancer treatment. Cancer Biol Ther. 2016;17:1158–67. https://doi.org/10.1080/15384047.2016.1235663.
Charmsaz S, Al-Ejeh F, Yeadon TM, Miller KJ, Smith FM, Stringer BW, Moore AS, Lee FT, Cooper LT, Stylianou C, et al. EphA3 as a target for antibody immunotherapy in acute lymphoblastic leukemia. Leukemia. 2017;31:1779–87. https://doi.org/10.1038/leu.2016.371.
Du E, Li X, He S, Li X, He S. The critical role of the interplays of EphrinB2/EphB4 and VEGF in the induction of angiogenesis. Mol Biol Rep. 2020;47:4681–90. https://doi.org/10.1007/s11033-020-05470-y.
Krasnoperov V, Kumar SR, Ley E, Li X, Scehnet J, Liu R, Zozulya S, Gill PS. Novel EphB4 monoclonal antibodies modulate angiogenesis and inhibit tumor growth. Am J Pathol. 2010;176:2029–38. https://doi.org/10.2353/ajpath.2010.090755.
Martiny-Baron G, Holzer P, Billy E, Schnell C, Brueggen J, Ferretti M, Schmiedeberg N, Wood JM, Furet P, Imbach P. The small molecule specific EphB4 kinase inhibitor NVP-BHG712 inhibits VEGF driven angiogenesis. Angiogenesis. 2010;13:259–67. https://doi.org/10.1007/s10456-010-9183-z.
Chrencik JE, Brooun A, Recht MI, Kraus ML, Koolpe M, Kolatkar AR, Bruce RH, Martiny-Baron G, Widmer H, Pasquale EB, Kuhn P. Structure and thermodynamic characterization of the EphB4/Ephrin-B2 antagonist peptide complex reveals the determinants for receptor specificity. Structure. 2006;14:321–30. https://doi.org/10.1016/j.str.2005.11.011.
Ahmed MS, Wang P, Nguyen NUN, Nakada Y, Menendez-Montes I, Ismail M, Bachoo R, Henkemeyer M, Sadek HA, Kandil ES. Identification of tetracycline combinations as EphB1 tyrosine kinase inhibitors for treatment of neuropathic pain. Proc Natl Acad Sci U S A. 2021;118. https://doi.org/10.1073/pnas.2016265118.
Barquilla A, Pasquale EB. Eph receptors and ephrins: therapeutic opportunities. Annu Rev Pharmacol Toxicol. 2015;55:465–87. https://doi.org/10.1146/annurev-pharmtox-011112-140226.
Amato KR, Wang S, Hastings AK, Youngblood VM, Santapuram PR, Chen H, Cates JM, Colvin DC, Ye F, Brantley-Sieders DM, et al. Genetic and pharmacologic inhibition of EPHA2 promotes apoptosis in NSCLC. J Clin Invest. 2014;124:2037–49. https://doi.org/10.1172/JCI72522.
Landen CN Jr, Chavez-Reyes A, Bucana C, Schmandt R, Deavers MT, Lopez-Berestein G, Sood AK. Therapeutic EphA2 gene targeting in vivo using neutral liposomal small interfering RNA delivery. Cancer Res. 2005;65:6910–8. https://doi.org/10.1158/0008-5472.CAN-05-0530.
Xiao T, Xiao Y, Wang W, Tang YY, Xiao Z, Su M. Targeting EphA2 in cancer. J Hematol Oncol. 2020;13:114. https://doi.org/10.1186/s13045-020-00944-9.
Acknowledgements
Not applicable.
Funding
This study was supported by National Natural Science Foundation, China (grant number 81773147, 81972198); Natural Science Foundation, Changsha (grant number: kq2208299); Strategic Priority Research Program of Central South University (ZLXD2017004); Health Special Fund Research Project of Hunan province (B 2021–04);the Fundamental Research Funds for the Central Universities of Central South University(2023ZZTS0864).
Author information
Authors and Affiliations
Contributions
X.T.G. , J.Q.T. and Y.Y.Y finished writing the manuscript, J.J.X. and , J.Q.T. participated in the revision of manuscripts and graphics. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Guo, X., Yang, Y., Tang, J. et al. Ephs in cancer progression: complexity and context-dependent nature in signaling, angiogenesis and immunity. Cell Commun Signal 22, 299 (2024). https://doi.org/10.1186/s12964-024-01580-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12964-024-01580-3